

 下载:
下载:

-
益智(Alpinia oxyphylla Miq.)为姜科(Zingiberaceae)山姜属(Alpinia)多年生丛生型草本植物,主产于广东、广西、海南和福建等地,其中海南岛因其独特的水热组合气候条件成为益智的道地性产区[1 − 2]。益智性温味辛,具有温中止泻、固精缩尿等多种功效[3]。现代研究发现益智代谢产物化学成分结构多样,药理活性丰富[4],其中倍半萜和二苯基庚烷是其中两类重要成分;益智果实中重要的倍半萜类成分诺卡酮的生物合成途径和机制已经得到解析[5 − 6];益智果实中二苯基庚烷类化合物的代表姜黄素不仅结构类型多样,而且生物活性显著。姜黄素是姜科植物的特征性成分之一,在姜黄属、山姜属以及姜属植物中被广泛报道[7]。目前关于益智中二苯基庚烷类化合物的生物合成途径和机制尚不明确,为此对益智中二苯基庚烷类化合物的合成路径有探索和发掘的意义。文献[8]报道姜黄中的两种的III型聚酮合酶,即二酮辅酶A(后文辅酶A用 CoA 表示)合成酶(DCS)和姜黄素合酶(CURS),可将两种带 CoA的底物聚合,经过多步反应生成姜黄素,为此本团队通过对益智中的III型聚酮合酶进行聚类分析,选取 AoPKS5 候选基因进行克隆及对其进行表达分析,这不仅有助于阐明益智中二苯基庚烷类化合物的生物合成机制,还可为药用活性成分的代谢工程改造提供分子依据。
植物Ⅲ型聚酮合酶(PKSⅢ)最早在植物中克隆并鉴定出,因此 PKSⅢ 常被称为植物特有的聚酮合酶[9]。文献[8]表明,目前已在不同植物物种中克隆并功能鉴定了二十余种具有不同催化特性的Ⅲ型 PKS,如查尔酮合酶(CHS),二苯乙烯合酶(STS),联苄合酶(BBS),二酮 CoA 合成酶(DCS),姜黄素合酶(CURS)等。Kreuzaler F等[10 − 11]在欧芹(Petroselinum crispum)细胞悬浮培养中发现一种查尔酮酶能将对香豆酰 CoA 和丙二酰 CoA 生成柚皮素(5, 7, 4' - 三羟基黄酮)。Katsuyama Y等[12 − 13]从姜黄中首次克隆出二酮 CoA 合成酶(DCS)和姜黄素合成酶(CURS),其中,二酮 CoA 合成酶通过阿魏酰 CoA 和丙二酰 CoA 的缩合,催化生成阿魏酰二酮 CoA;姜黄素合成酶在体外催化肉桂酰二酮-乙酰半胱胺和阿魏酰 CoA 生成姜黄素 。Wang J等[14]从蛇足石杉(Huperzia serrata)中克隆了一种新的III型 PKS,命名为 HsPKS3,将香豆酰 CoA、丙二酰 CoA 和重组酶 HsPKS3 孵育,-产生了双去甲氧基姜黄素的二苯基庚类支架,同时形成了羟基苯甲丙酮的C6−C3支架,HsPKS3 还可以催化肉桂酰 CoA 类生成多种二苯基庚烷,且 HsPKS3 接受含氮的大体积n-甲基苯基 CoA 作为起始物,产生喹诺酮类生物碱[14]。Sakamoto S等[15]从白花丹中克隆了两个III型聚酮合酶的 cDNA( PKS1 、PKS2),PKS1 通过接受乙酰 CoA 和丙二酰 CoA,催化从3到6个乙酸单位形成5个α-吡喃酮,而 PKS2 通过接受对香豆酰 CoA 和丙二酰 CoA 催化柚皮素和去甲杨梅酮的形成。Choi GS 等[16]使用两种底物(邻氨基苯甲酸酯和丙二酰 CoA )在大肠杆菌中通过两种酶(III型聚酮合酶的一类吖啶酮合成酶(ACS)和邻氨基苯甲酰 CoA 连接酶)共同作用产生两种吖啶酮:1,3-二羟基-9(10H)吖啶酮(DHA)和1,3-二羟基-10-甲基吖啶酮(NMA)。Zhou SG等[17]从铁皮石斛中克隆了一个新的III型聚酮合酶(DoBS1),DoBS1 蛋白能够在体外以4-香豆酰 CoA 和3-丙二酰 CoA 为底物合成二氢白藜芦醇(DHR)。Chen Y等[18]从华石斛中克隆并鉴定了一个III型PKS( DsBBS),归类为联苯合成酶(bibenzyl synthase, BBS),DsBBS酶活性分析表明,重组 DsBBS-HisTag 蛋白能以4-香豆酰 CoA 和丙二酰 CoA 为底物在体外合成白藜芦醇。Wang XH等[19]从白木香(Aquilaria sinensis)中鉴定并解析了一个新的III型聚酮合酶(PECPS),该酶在体外能以4-羟基苯丙酰 CoA 与丙二酰 CoA 生成四氢双去甲氧基姜黄素,而用苯甲酰 CoA、丙二酰 CoA 和4-羟基苯丙酰 CoA 共孵育方法构建C6-C5-C6支架后再与 PECPS 共反应后,除产生四氢双去甲氧基姜黄素外还产生了5-(4-羟苯基)-1-苯戊烷-1,3-二酮。Manoilenko S等[20]选取了来自非洲菊和拟南芥的两个III型PKS,使用了20个芳香和脂肪族 CoA 酯作为底物,基于UHPLC-ESI-HRMS的反应中间体和产物分析显示非洲菊和拟南芥的这两个III型PKS酶具有广泛的底物特异性。Takemoto K等[21]从旅顺桤木中克隆出三个III型PKS( AsPKSIII1、AsPKSIII2、AsPKSIII3),并通过添加底物对香豆酸、阿魏酸、咖啡酸、芥子酸、二氢对香豆酸或肉桂酸,同时将4CL和ACC 基因与各自的AsPKSIII 基因转化,结果显示:①AsPKSIII1 合成了对应于对香豆酸、阿魏酸和咖啡酸的二酮 CoA,这提示AsPKSIII1参与了二苯基类庚烷生物合成的第一步反应;②AsPKSIII2 和AsPKSIII3 合成了对应于对香豆酸、咖啡酸、二氢对香豆酸和肉桂酸的三酮酮吡咯酮和查尔酮。本研究团队首次从南药益智基因组中系统筛选出Ⅲ型聚酮合酶(PKSⅢ)基因家族 AoPKSs,并结合聚类分析与转录组数据筛选获得候选基因 AoPKS5,还通过生物信息学与分子实验相结合的方法,对AoPKS5的序列结构与功能特征进行系统解析,结果显示:①序列分析与结构预测结果显示AoPKS5 含有典型的 Cys-His-Asn 催化三联体和保守的 GFGTG 结构域;②RT-qPCR 结果表明AoPKS5 在益智的叶、花、茎、块茎和果实中呈现组织特异性表达,这提示AoPKS5可能参与二苯基庚烷类次生代谢物的合成与调控。本研究团队进一步构建了 pNC-ET28-AoPKS5 重组表达载体和优化IPTG浓度与诱导时间,实现了AoPKS5蛋白在大肠杆菌中的高效原核表达,并经 Ni-NTA 亲和层析纯化获得高纯度重组蛋白,这为后续酶学功能验证及代谢工程应用奠定了分子基础。
-
实验材料为种植于中国热带农业科学院热带生物技术研究所(110°19'E,19°59'N)的多年生益智植株。主要试剂为总 RNA 提取试剂盒:FOREGENE Plant Total RNA Isolation Kit Plus;琼脂糖:IOWEST REGULAR AGAROSE G-10;cDNA 反转录试剂盒:TIANGEN FastKing 一步法除基因组 cDNA 第一链合成预混试剂;2×Taq AS D:iScience 2×Taq-AS PCR Mix(+Dye);高保真酶:TaKaRa PrimeSTAR HS(Premix);Marker:TIANGEN D2000分子量标准;胶回收试剂盒:Magen HiPure Gel Pure DNA Mini Kit;载体 pMD-19T: TaKaRa pMDTM19-T Vector Cloning Kit;DH5α感受态细胞:江苏愚公生命科技有限公司 E.coli DH5α;质粒提取试剂盒:FOREGENE General Plasmid Mini Kit;荧光定量试剂盒:TIANGEN FastReal 快速荧光定量PCR预混试剂(SYBR Green);NC克隆试剂盒:海南你行生物 Nimble Cloning 试剂盒;IPTG:TIANGEN IPTG(Sterile)50 mg·mL-1;BL21(DE3):TIANGEN E. coli BL21(DE3)感受态;裂解酶:EMD Millipore Bugbuster Protein Extraction Reagent;Ni-NTA:MACHEREY-NAGEL GmbH&Co.KG Protino Ni-NTA Column 1 mL。主要仪器为PCR 扩增仪:BIO-RAD;实时荧光定量 PCR 仪:Roche LightCycler96;琼脂糖凝胶电泳仪:北京 LIUYI 琼脂糖水平电泳仪;蛋白电泳仪:北京 LIUYI SDS-PAGE 电泳仪;凝胶成像系统:Analytik Jena。
-
使用TBTools对益智基因组[22]分析,使用Blast比对将益智的蛋白序列文件和PKSs基因家族参考序列蛋白序列文件进行两两比对,得到益智PKSs基因家族蛋白文件。使用eggNOG-MAPPER网站(
http://eggnog-mapper.embl.de/ )、Pfam网站(http://pfam.xfam.org/ )和NCBI网站(https://www.ncbi.nlm.nih.gov/Structure/bwrpsb/bwrpsb.cgi )对益智PKSs基因家族蛋白在线预测,进一步筛选符合PKSs蛋白结构的序列。 -
通过NCBI网站(
https://www.ncbi.nlm.nih.gov/ )下载65个物种的共计80条PKSs和益智PKSs蛋白序列通过MEGA X64软件中的邻接法(NJ)构建系统树,使用MEGA X64 软件中的最大似然法(ML)构建系统树,从中筛选出与目标蛋白亲缘关系较近的序列用于进一步比对。使用DNAMAN将AoPKS5氨基酸序列与姜黄(Curcuma longa, DCS, CURS)、大米七香蕉(Musa acuminata AAA Group, DCS, CURS)、生姜(Zingiber officinale, DCS, CURS)、莪术(Curcuma zedoaria, CURS)、非洲菊(Gerbera hybrid cultivar, 2-PS)、向日葵(Helianthus annuus, 2-PS)、紫花丹(Plumbago indica, HKS)、喜马拉雅大黄(Rheum australe, ALS)、药用大黄(Rheum officinale, ALS)、掌叶大黄(Rheum palmatum, ALS)以及印度獐牙菜(Swertia chirayita , BPS)共计15个PKSIII序列进行序列比对。 -
采取益智果实、花、茎和叶4个部位样品送转录组检测(相关结果未发表)。使用 TBTools 将从基因组筛选出的益智PKSs基因家族通过转录组分析制作转录组表达量热图。
-
选取生长茂盛的益智叶片,迅速置于液氮中速冻,速冻后,用研杵在研钵中迅速将其研磨成粉末。称取约50 mg研磨样品用于后续实验步骤。用RNA提取试剂盒提取益智总RNA,使用琼脂糖凝胶电泳检测。挑选条带清晰、无降解的RNA样品,使用cDNA试剂盒进行反转录反应,合成cDNA产物。将所得cDNA冻存于−20℃冰箱中,备用于后续各项分子实验。
-
利用TBtools软件对AoPKSs基因进行转录组水平的表达分析,同时评估AoPKS5蛋白的分子量、等电点等基本理化性质。借助NetPhos-3.1(
https://services.healthtech.dtu.dk/services/NetPhos-3.1/ )、TMHMM-2.0(https://services.healthtech.dtu.dk/services/TMHMM-2.0/ )、SignalP 5.0(https://services.healthtech.dtu.dk/services/SignalP-5.0/ )、SOPMA(https://npsa-prabi.ibcp.fr/cgi-bin/npsa_automat.pl?page=npsa_sopma.html ) 及 AlphaFold 3(https://alphafoldserver.com/ )在线工具分别对AoPKS5蛋白的磷酸化修饰位点、跨膜结构、信号肽以及二级结构和三级结构进行预测及分析。 -
使用IDT公司(https:hidtdna.com)在线设计AoPKS5基因特异性引物,得到正向引物 AoPKS5F为5′-ATGGAAGTGAACGGCTACC-3′,反向引物AoPKS5R为5′-CTAGTTCAGACTGCAACTATGGA-3′。以益智叶片的cDNA为模板进行PCR扩增。用琼脂糖凝胶电泳检测无误后切取对应条带并利用胶回收试剂盒回收纯化DNA,与 pMD-19T 载体进行连接,转化至DH5α感受态细胞,筛选阳性克隆后提取质粒,并将提取产物送至测序公司进行序列验证。
-
使用IDT公司在线设计AoPKS5基因荧光定量PCR特异性引物:正向引物 q-AoPKS5F 5′-CGTCGTAGATCAGAACGCTTAT-3′,反向引物 q-AoPKS5R 5′-GCGCCTAAACTTGGCTTTG-3′。同时选取内参基因作为标准化依据,其引物序列分别为 q-F:5′-TCTAATTGTGTGCCTGGTAGTG-3′和q-R:5′-GGCTGGGTTTCGGCTTATTA-3′。荧光信号采集由实时定量PCR系统完成,用于检测AoPKS5在益智不同组织中的表达水平。实验设置3组独立的生物学重复。采用2−ΔΔCt法分析,使用单因素方差分析法(One-way ANOVA)结合Tukey's多重比较检验,评价目标基因的组织两两之间的特异性表达差异。
-
以1.3步骤获得的质粒为模板,进行PCR扩增反应。扩增后获得的DNA条带经电泳检测,切取与目标大小相符的单一条带并进行胶回收。以回收产物为模板与载体pNC-ET28相连。将连接产物转化至DH5α 感受态细胞中,筛选阳性克隆并进行质粒提取与测序验证。验证无误的重组质粒pNC-ET28-AoPKS5随后转化至大肠杆菌感受态BL21(DE3)中,继续筛选阳性菌株并进行测序确认。阳性菌株接种液以1∶100比例扩增至25 mL,并平均分装至5个50 mL离心管中,于37℃、180 r·min−1 振荡培养,直至OD600 =0.6~0.8。分别向各组添加不同终浓度的IPTG(0、0.1、0.2 和 0.5 mmol·L−1),在16℃、220 r·min−1条件下诱导表达18 h,以筛选出最佳诱导剂浓度。在筛选得到的最佳IPTG浓度条件下,继续接种5 mL菌液分别培养14、16、18、20和22 h,采集菌液后制备蛋白样品,使用SDS-PAGE进行蛋白表达检测,确定最适诱导时间与诱导剂浓度组合。根据筛选结果,采用最优条件诱导5 mL菌液,取其中1 mL于4℃下12 000 r·min−1离心10 min,弃去上清液后加入150 µL细胞裂解液,于4℃条件下裂解过夜。分别提取可溶性蛋白(上清液)与包涵体部分,制备蛋白样品并通过SDS-PAGE检测蛋白表达情况。最终根据优化条件大规模诱导表达,接种至500 mL菌液中进行蛋白表达。培养结束后,菌液于4℃、8 000 r·min−1离心15 min收集菌体。接着进行细胞破碎与菌体溶解处理,后续包括蛋白重折叠和透析步骤。最后使用Ni-NTA亲和层析对目标蛋白进行纯化,纯化效果通过SDS-PAGE检测验证。
-
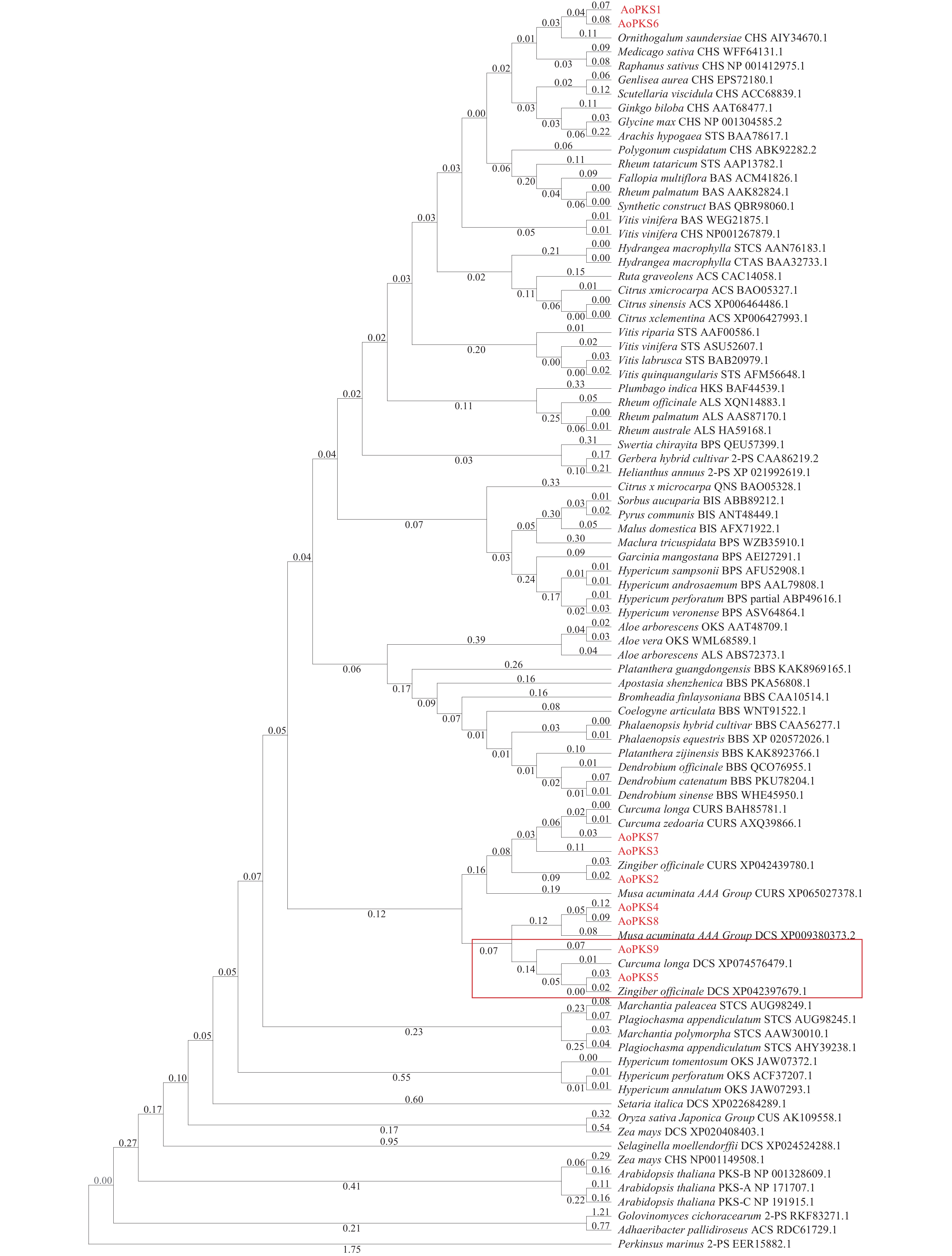
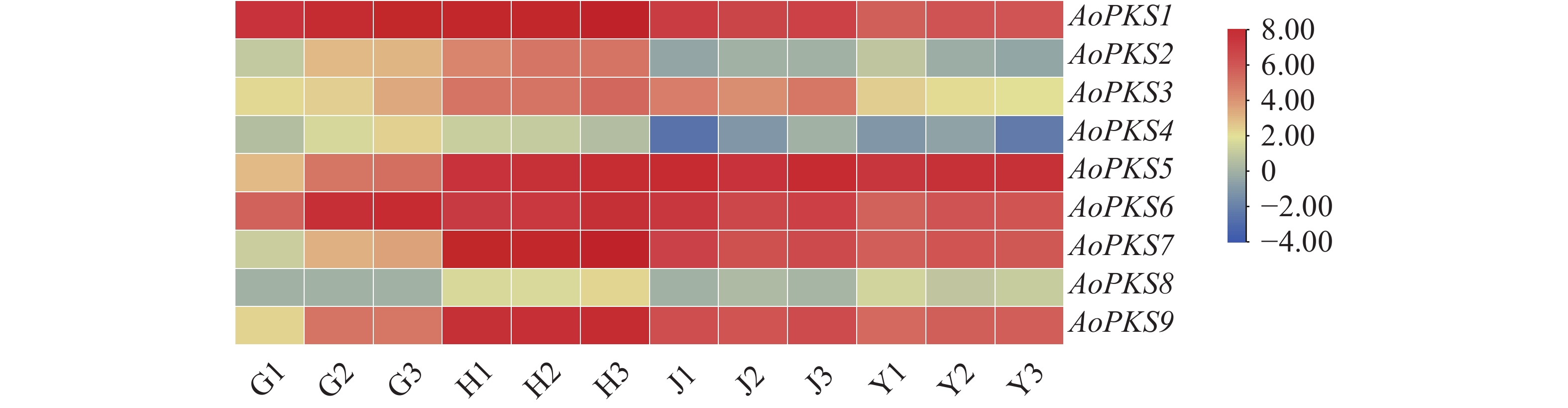
在益智基因组[22]中共筛选出9条PKSs基因蛋白序列,与65个物种共计80条PKSs通过MAGA 11建立系统树通过邻接法(NJ)(图1)和最大似然法(ML)(图2)进行聚类分析,再在转录组文件中对9条PKSs蛋白序列FPKM值取log2对数,使用TBTOOLS对得到的9条PKSs蛋白序列FPKM值绘制表达量热图(图3)。进化树将亲缘关系相近的基因聚在一起,推测聚在同一枝的基因有相同或者相似的功能。NJ 法建立的系统树中AoPKS5和AoPKS9与同是姜科植物的已验证功能的生姜和姜黄的DCS聚在一起,推测四者之间具有相同或者相似的功能。结合转录组表达量热图,结果显示,基因AoPKS5在果、茎、叶3个组织中表达量高于AoPKS9,再根据ML法建立的系统树:枝上的数字表示遗传差异,数字越小表示二者序列相似性高,进化上亲缘关系越近,故选取AoPKS5作为研究对象。序列比对结果表明AoPKS5和14种PKSIII的蛋白同源性分别为95.89%、65.63%、70.77%、59.49%、95.63%、62.76%、65.38%、59.95%、56.33%、51.26%、53.45%、53.45%、53.71%和58.42%;通过多重序列比对(图4)显示这些III型聚酮合酶都拥有查尔酮酶超基因家族共同的催化三联体 Cys-His-Asn[23],同时它们都拥有植物III型聚酮合酶的序列标签 GFGPG[24]。
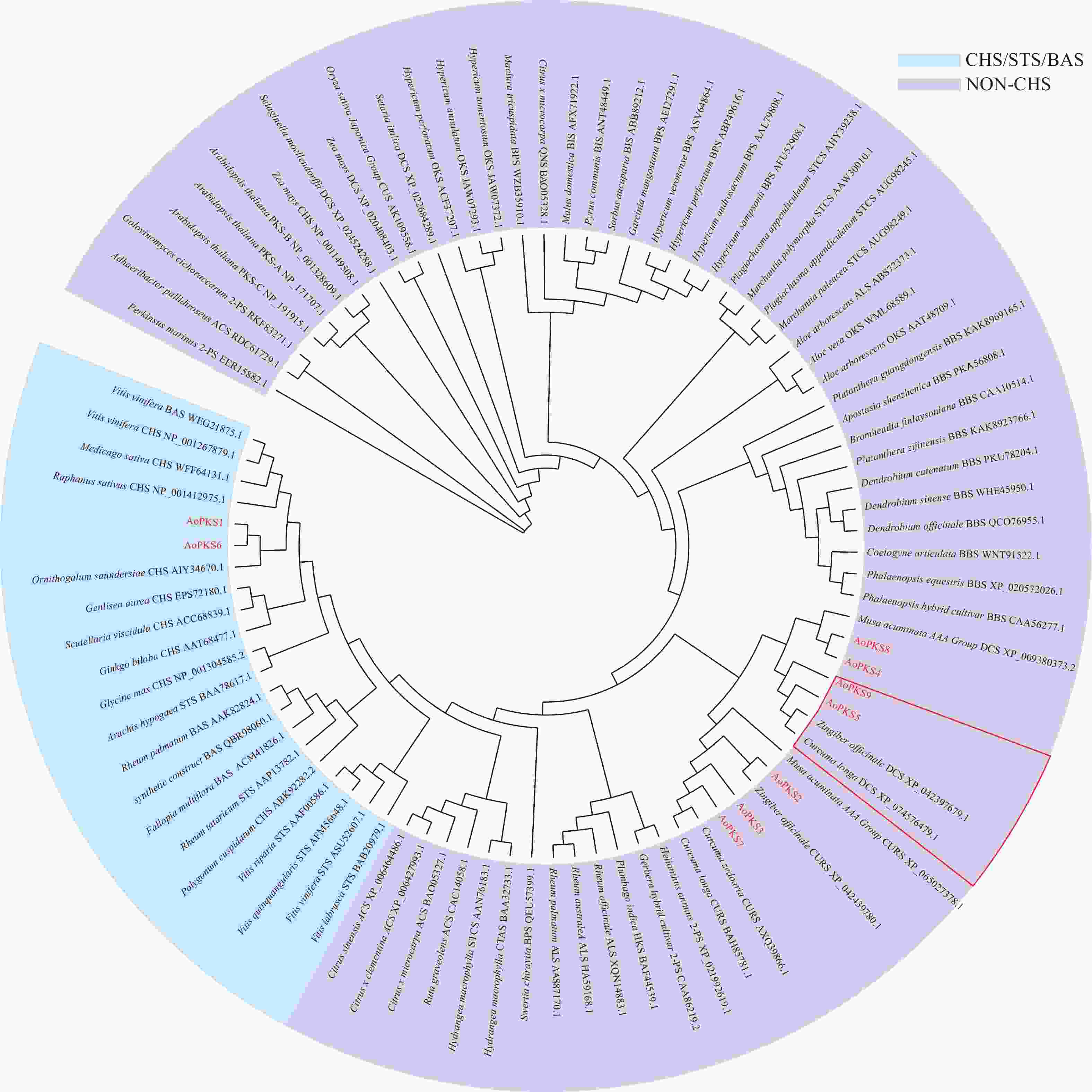
图 1 不同物种III型聚酮合酶氨基酸序列系统树构建(NJ)
Figure 1. Construction of phylogenetic tree of PKSIII amino acid sequence in different species(NJ)
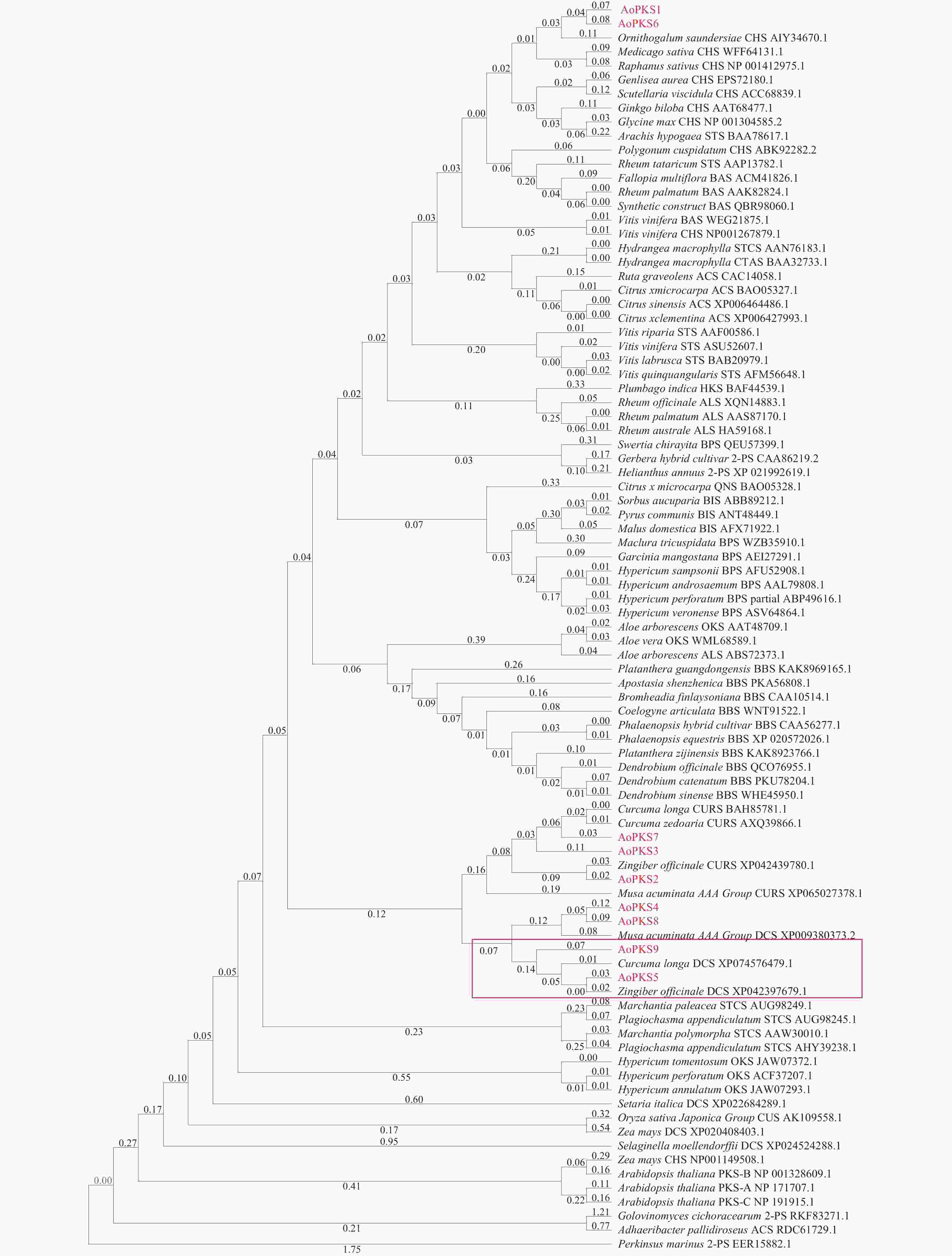
图 2 不同物种III型聚酮合酶氨基酸序列系统树构建(ML)
Figure 2. Construction of phylogenetic tree of PKSIII amino acid sequence in different species(ML)
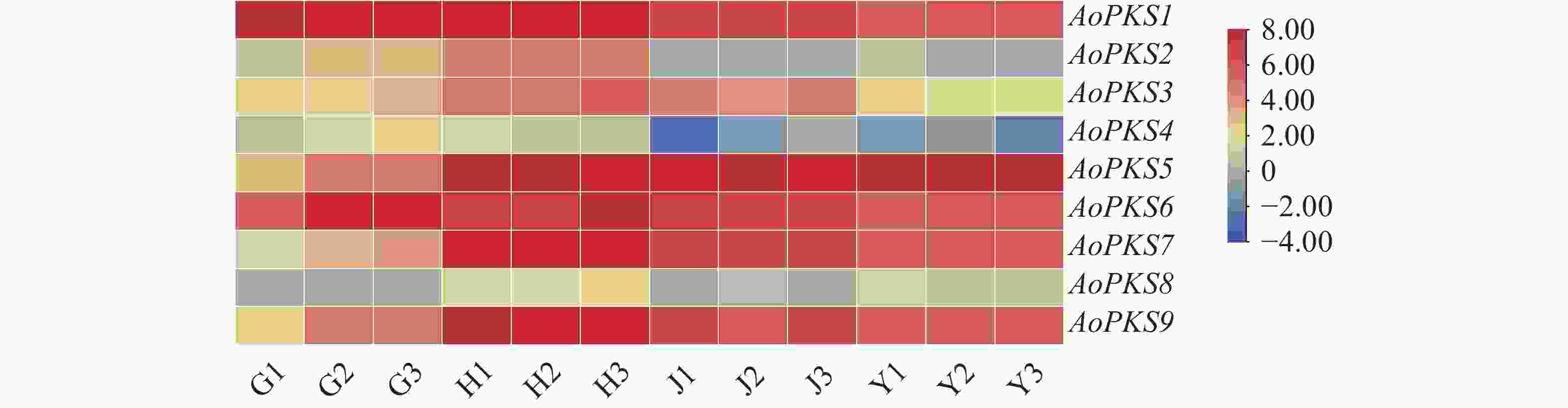
图 3 益智不同组织差异表达 AoPKSs 基因
Figure 3. AoPKSs gene was differently expressed in different tissues of Alpinia oxyphylla

图 4 AoPKS5 与14种植物III型聚酮合酶多序列比对
Figure 4. Multisequence alignment of AoPKS5 with PKSIII from 14 species of plants
-
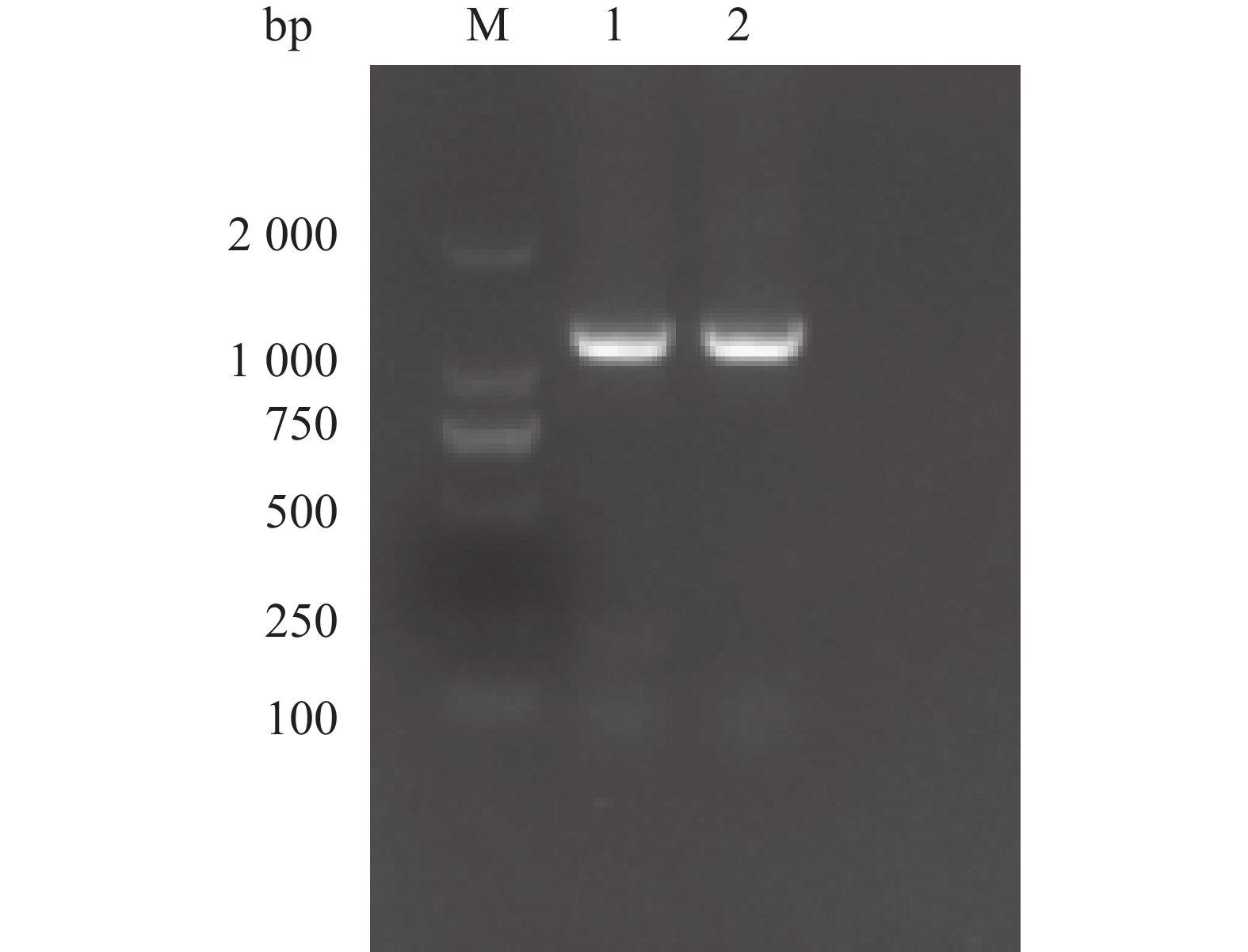
以从益智中提取的总RNA反转录获得的cDNA为模板,利用特异性设计的正反向引物对AoPKS5基因进行了PCR扩增。扩增产物经1%琼脂糖凝胶电泳分析后,在约
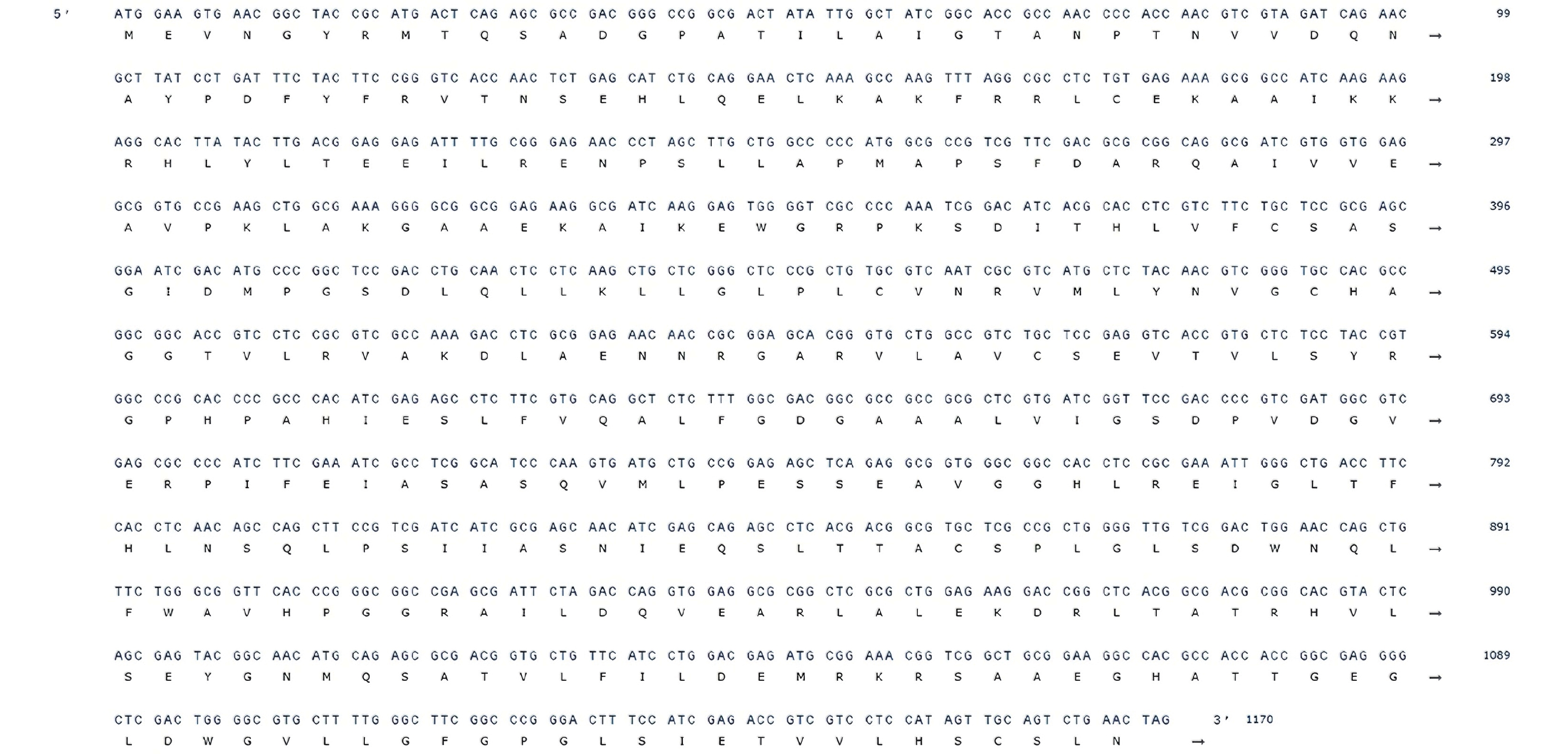
1200 bp处观察到清晰条带,其大小与预期的目标基因相符(图5)。随后对电泳条带对应的DNA 片段进行胶回收纯化,并将其与载体进行连接,转化至感受态细胞中。经筛选阳性克隆并进行测序验证完全正确后,确认AoPKS5的开放阅读框(ORF)长度为1 170 bp,编码389个氨基酸残基(图6)。
图 5 AoPKS5 基因克隆
Figure 5. Cloning of AoPKS5 gene
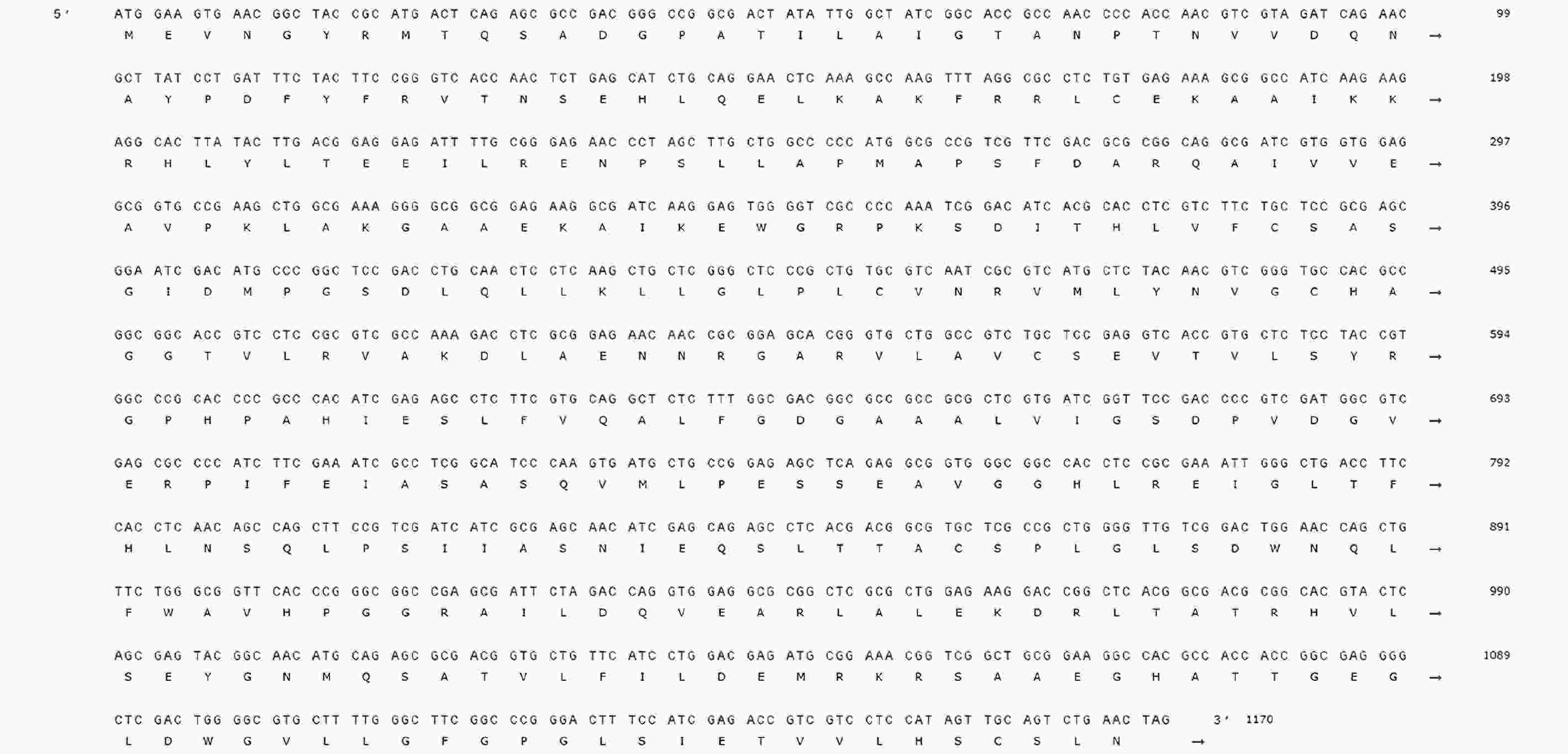
图 6 AoPKS5 基因序列
Figure 6. AoPKS5 gene sequence
-
AoPKS5 蛋白的分子式为C1863H
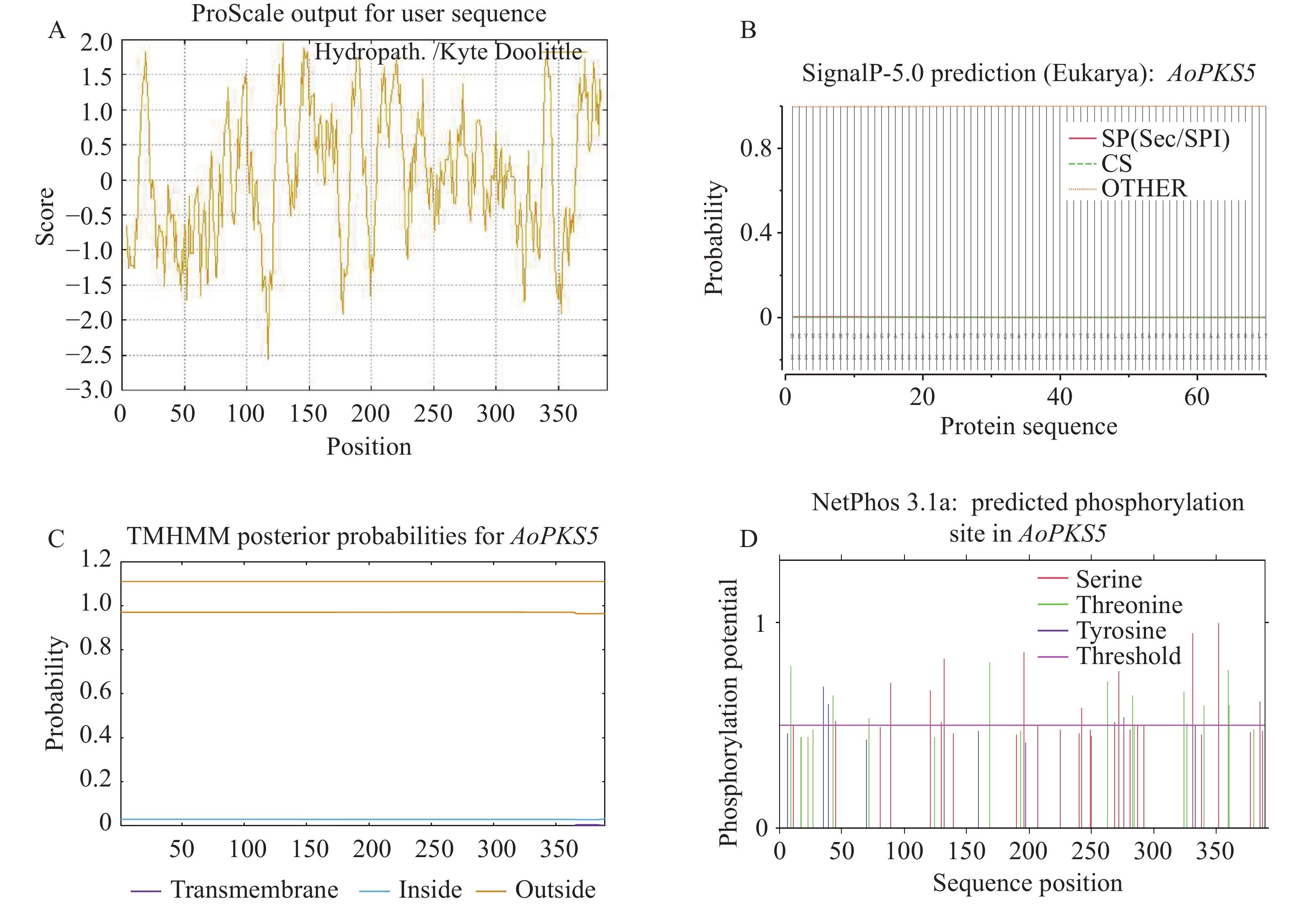
2981 N521O554S15,相对分子质量约为42 023.22 Da,理论等电点(pI)为5.96。不稳定指数为41.33,表明该蛋白属于不稳定类型;脂肪指数为100.33,亲水性指数为0.042,综合判断该蛋白具有疏水性特征,属于不稳定的疏水性蛋白(图7-A)。进一步通过TMHMM分析发现AoPKS5蛋白不含跨膜结构域,推测AoPKS5蛋白为胞外蛋白(图7-B)。使用SignalP 5.0工具进行信号肽预测,结果显示AoPKS5蛋白不具有信号肽结构,表明AoPKS5蛋白为非分泌型蛋白(图7-C)。此外,借助 NetPhos-3.1对AoPKS5蛋白潜在磷酸化位点进行预测分析,发现 AoPKS5 蛋白包含13个丝氨酸(Ser)、11个苏氨酸(Thr)以及2个酪氨酸(Tyr)残基为潜在磷酸化修饰位(7-D)。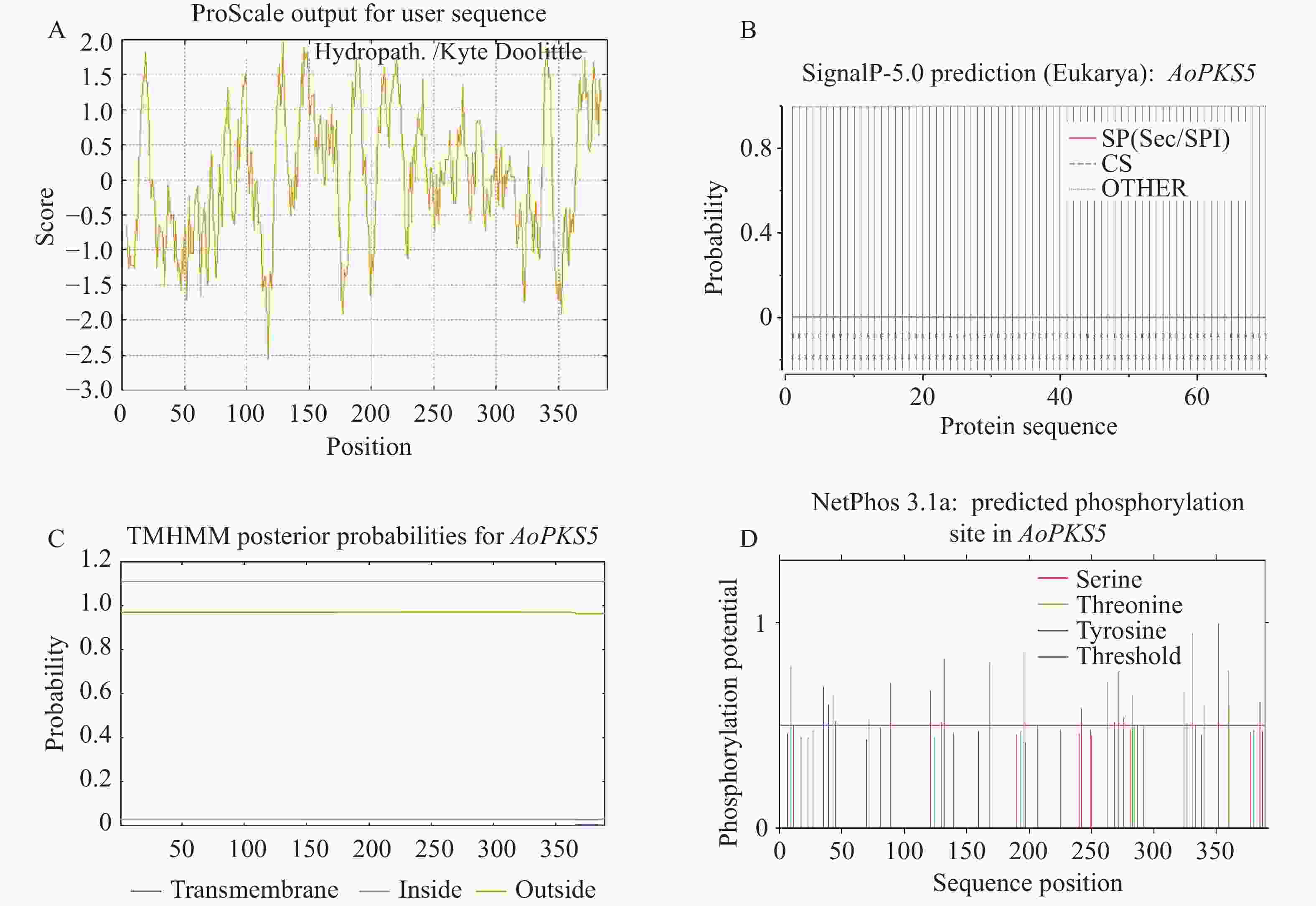
图 7 AoPKS5 蛋白预测分析
Figure 7. AoPKS5 protein prediction analysis
-
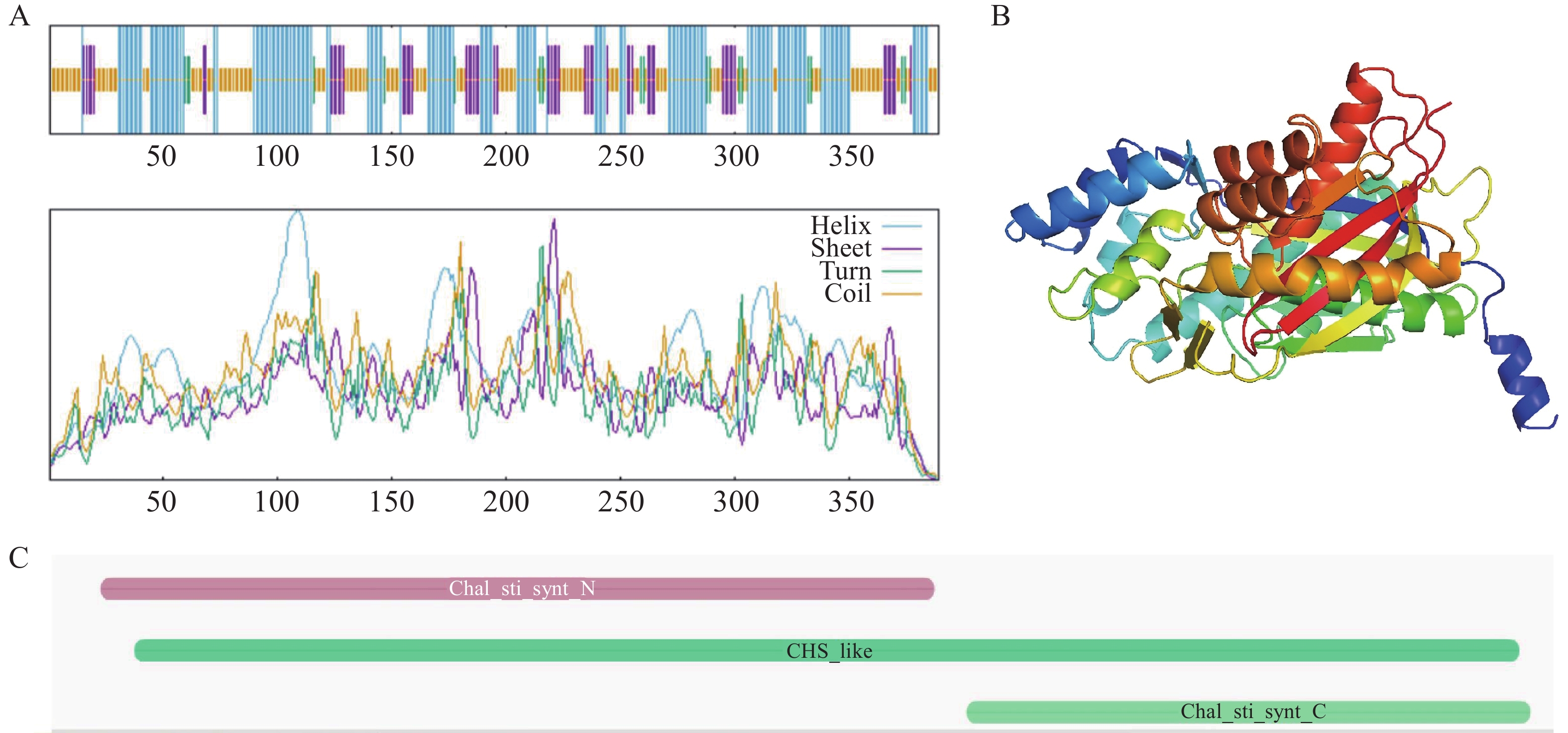
采用SOPMA在线工具对AoPKS5蛋白的二级结构进行预测,结果显示AoPKS5蛋白以α-螺旋为主要构象,占比为41.65%;其次为无规则卷曲,占39.33%;β-折叠结构占14.40%,而 β-转角部分占4.63%(图8-A)。通过AlphaFold 3平台对AoPKS5蛋白三维结构进行建模分析(图8-B)。在保守结构域方面,利用InterPro数据库进行功能结构注释,发现AoPKS5 蛋白包含1个典型的CHS-like结构域,其范围为第15至383位氨基酸残基;N端6~227位氨基酸残基构成Chal_sti_synt_N具有硫代酶作用的PF00195结构域,C端237~386位氨基酸有硫代酶作用的PF02797结构域(图8-C)。
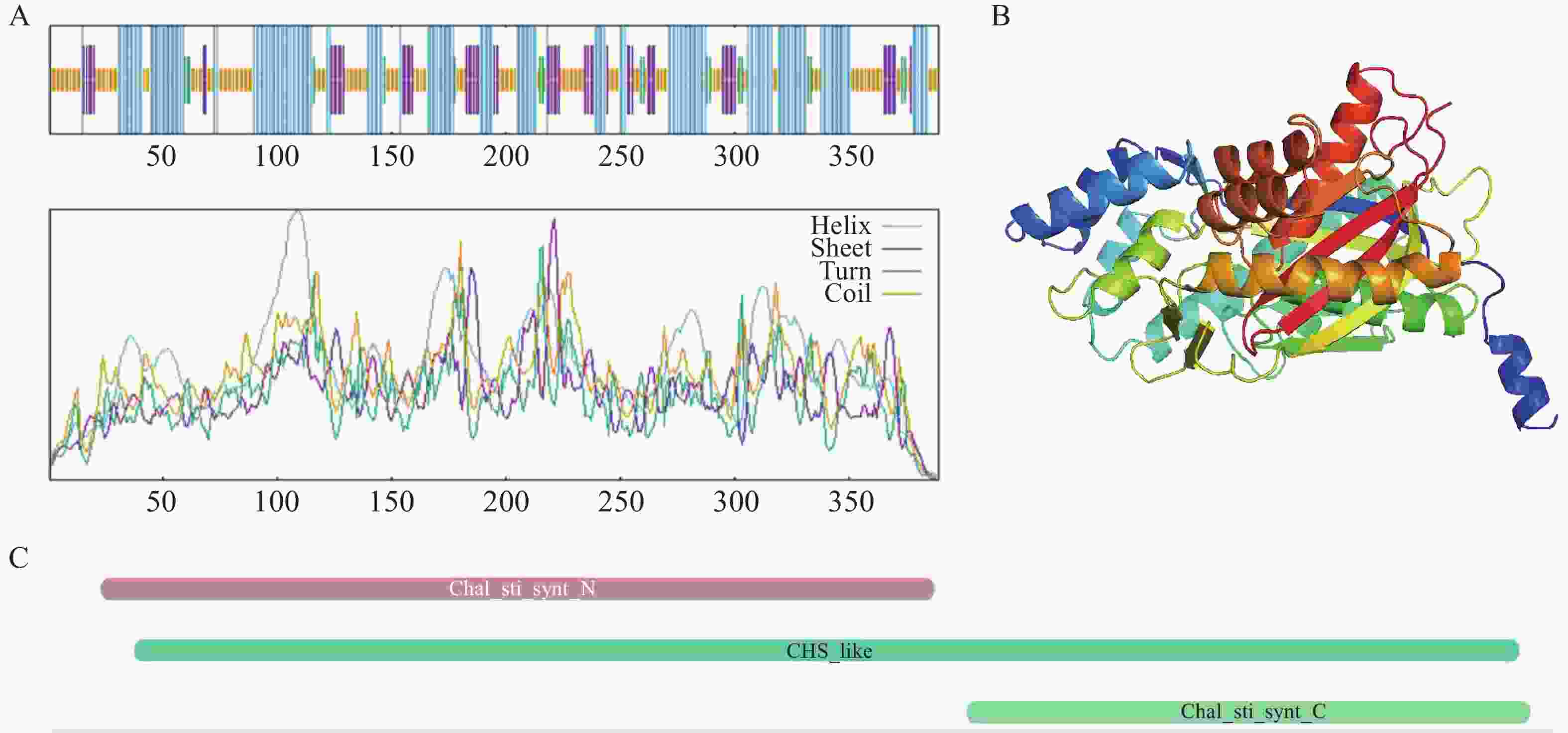
图 8 AoPKS5 蛋白结构预测与结构域
Figure 8. Structure prediction and domain of AoPKS5 protein
-
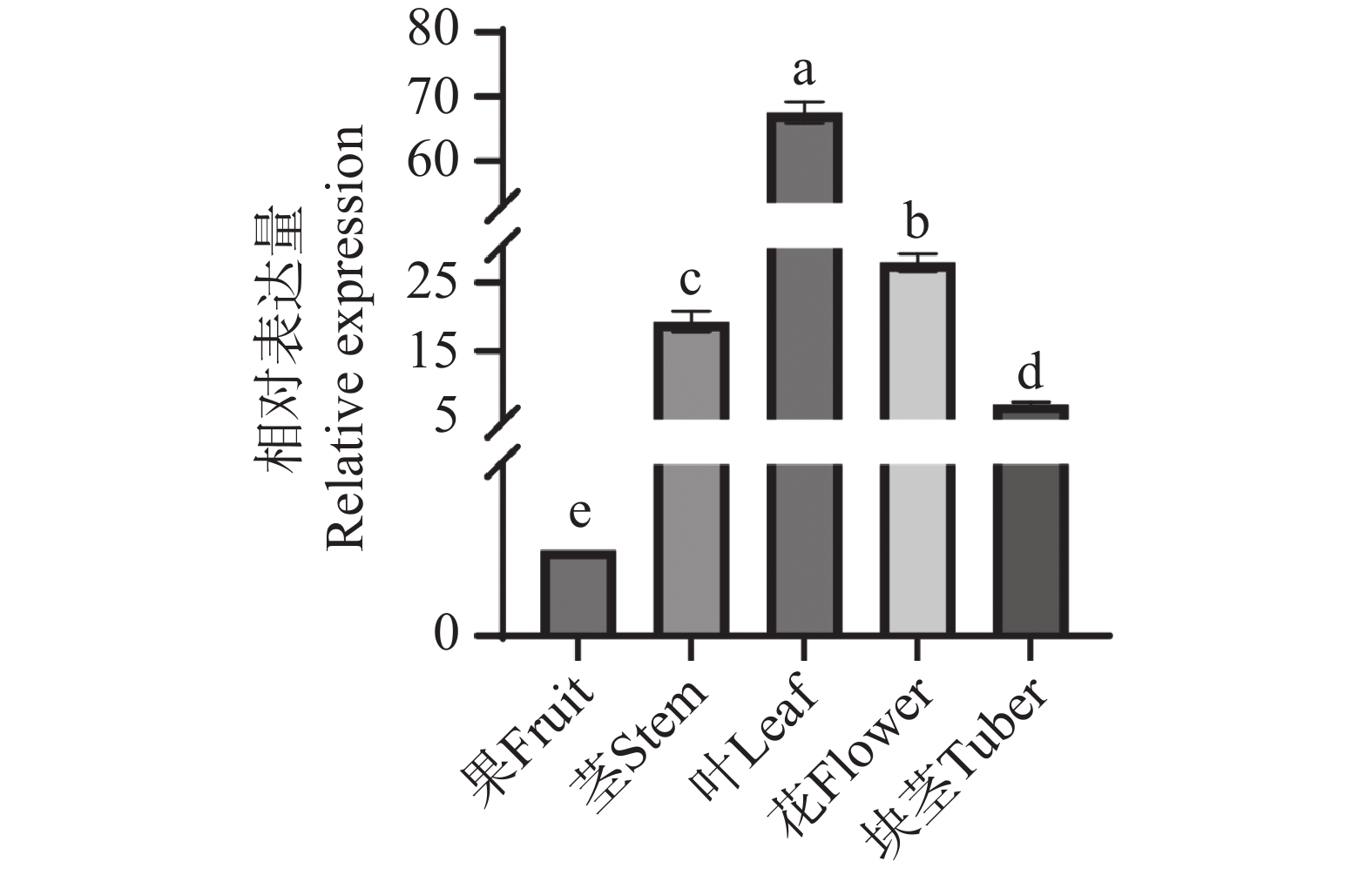
采用荧光定量PCR技术检测AoPKS5基因在益智的茎、叶、花、果以及块茎5个部位的相对表达量,结果显示5个部位均有AoPKS5的表达,且在不同组织中的表达呈现显著的组织特异性。结果显示,叶中的表达量最高,其次是花、茎、块茎和果。以益智果实中的基因表达量为参考,对5个组织部位进行单因素方差分析(One-way ANOVA),结合 Tukey's 多重比较检验,采用显著性差异分析abcd字母标记法,表达量均值最高的部位设置为 a,以此类推,10组数据中P值均小于0.05,可得知5个部位两两之间均存在显著水平差异(图9)。结合转录组表达量数据分析可知荧光定量PCR数据与转录组表达数据呈现相似的变化规律。
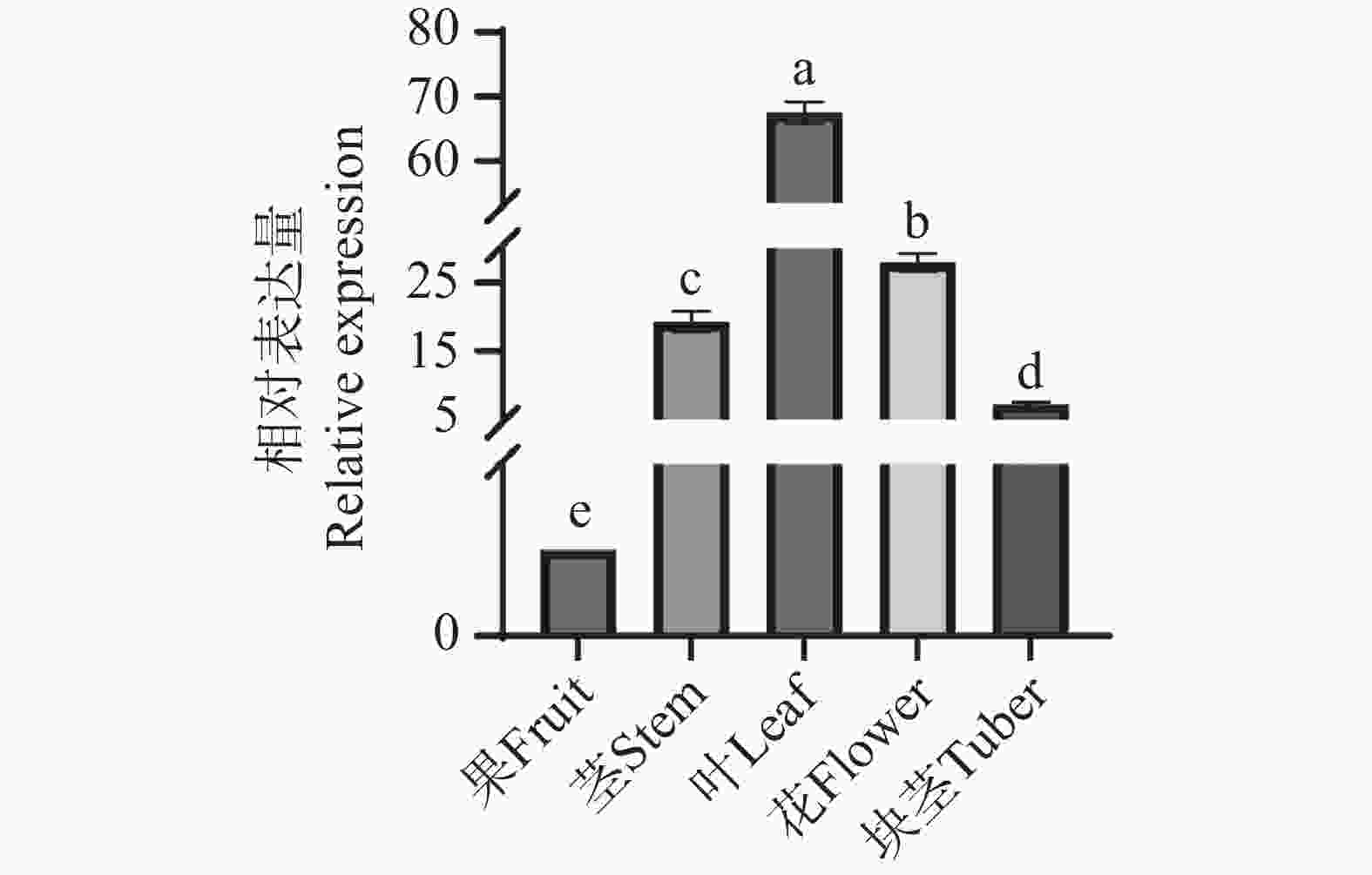
图 9 益智不同组织 AoPKS5 基因表达量情况
Figure 9. Expression levels of the AoPKS5 gene in different tissues in A. oxyphylla
-
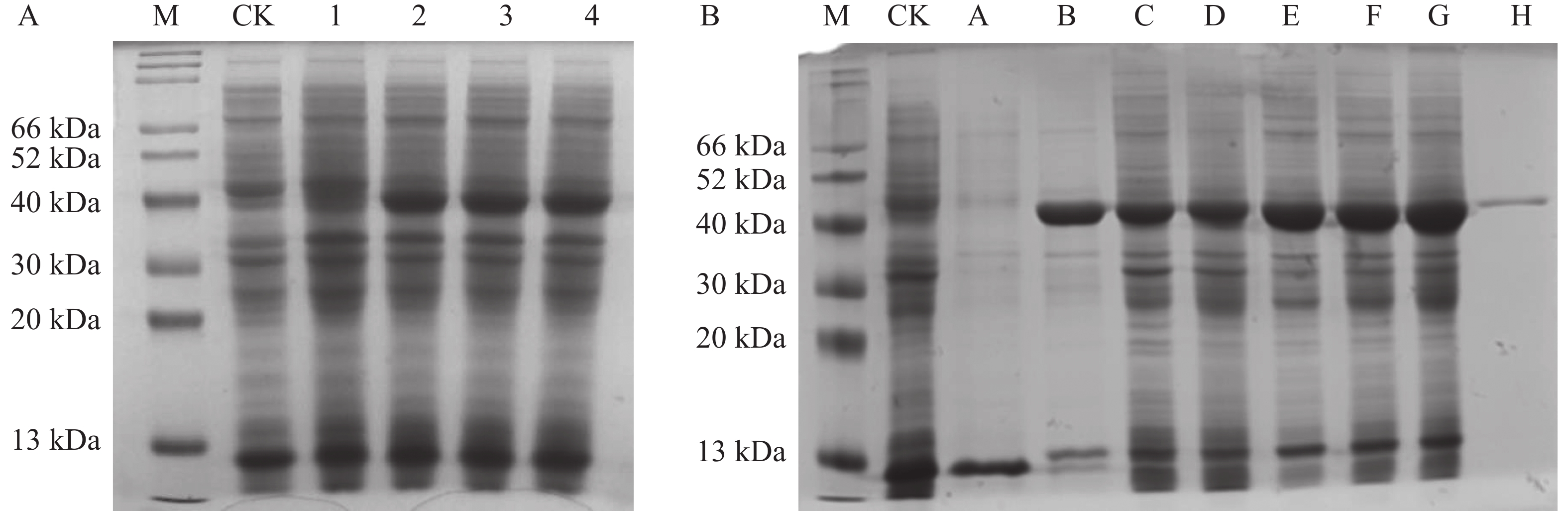
将重组质粒pNC-ET28-AoPKS5转化至感受态E. coli BL21(DE3)中以实现目的蛋白的表达。通过IPTG诱导后,利用SDS-PAGE凝胶电泳对表达产物进行检测。结果显示,相较于空载对照及未诱导组,诱导表达组在约40 kDa位置出现了明显的蛋白条带。蛋白在IPTG 添加后表达,当IPTG为0.1 mmol·L−1时出现了目的蛋白分子量条带,且后续表达量不再随着IPTG浓度的增加而增加(图10-A)。此外,诱导时间对蛋白表达也具有显著影响,蛋白产量随着诱导时间的延长而上升。表达的重组蛋白主要富集于沉淀部分,表明其以包涵体形式存在,因此采用包涵体处理策略进行后续操作。进一步使用Ni-NTA亲和层析对目标蛋白进行纯化,SDS-PAGE分析结果显示,纯化后的蛋白条带清晰,无杂蛋白干扰,且与理论分子质量一致(图10-B)。
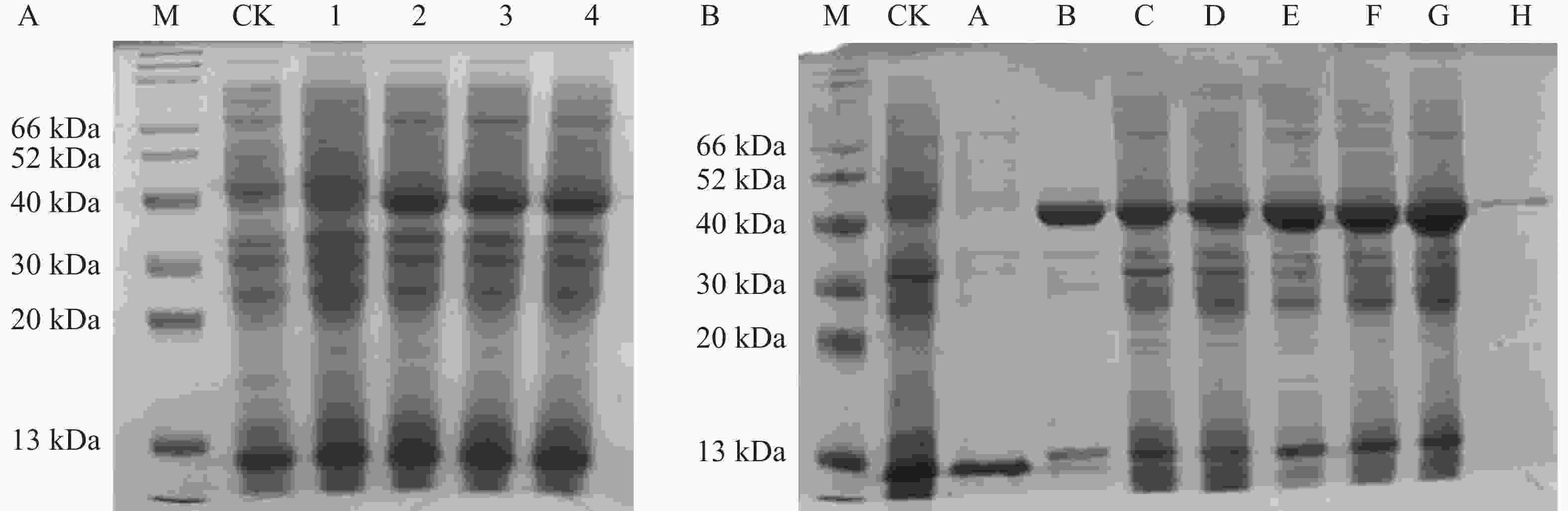
图 10 AoPKS5 蛋白 SDS-PAGE 蛋白凝胶电泳
Figure 10. SDS-PAGE protein gel electrophoresis of AoPKS5 protein
-
本研究首次克隆并初步分析了南药益智中III型聚酮合酶基因AoPKS5的结构。通过序列比对与系统发育分析,AoPKS5与已鉴定的姜科植物二酮辅酶A合成酶(姜黄Curcuma longa DCS; 生姜Zingiber officinale DCS)高度同源,提示其在二苯基庚烷骨架合成方面具有类似催化活性[12 − 13]。AoPKS5蛋白保留了典型的Cys–His–Asn催化三联体和GFGPG序列标签[23 − 24],二者为Ⅲ型PKS功能实现的关键结构模块,与2-PS、DCS、CURS等功能已明确的植物Ⅲ型PKS保持高度一致,这些保守位点在类型Ⅲ PKS 的催化机制与底物整合中发挥关键作用,决定了起始底物结合、延链次数与环化类型等催化特性。因此,从分子水平推断AoPKS5具备形成二苯基类骨架的催化能力是合理的[8]。
在原核表达体系优化中,本研究确定了AoPKS5蛋白高效包涵体表达的最优条件,并纯化获得高纯度重组蛋白。虽然包涵体形式产量相较于可溶性蛋白高,但酶学活性的恢复与重折叠步骤的效率密切相关,后续可通过优化表达条件(进一步降低诱导温度、减少IPTG浓度或缓慢诱导等)以提高可溶性表达,还可通过优化折叠条件,如添加共表达分子伴侣(如 GroEL-GroES、DnaK-DnaJ-GrpE 或触发因子 TF)以辅助折叠提升可溶性蛋白的得率[25],以便进行酶动力学和底物特异性研究。
尽管本研究系统阐述了AoPKS5蛋白的分子特征和表达特性,但仍有若干问题有待深入探索。一方面是AoPKS5蛋白的底物范围及产物谱尚未通过体外酶学实验加以验证,但是关于底物特异性与产物多样性的分子基础已有较为清晰的研究进展。PKSIII的底物范围、延链次数和环化方式主要受活性位点口袋的空间构象与若干关键残基控制[26](例如环口袋大小、GFGPG回路及附近的氨基酸侧链位置),因此,通过比对AoPKS5与已鉴定的 DCS/CURS、HsPKS3 等蛋白的关键位点,可初步预测 AoPKS5的底物偏好并为定点突变提供候选位点(但具体残基及其作用需结合三维结构建模与功能验证来确认)。文献[20]亦显示某些Ⅲ型PKS对起始底物具有较宽的底物兼容性,这可能导致体外/体内产物谱的多样性,;因此在设计体外实验时应同时检测多种苯丙酰-CoA 类起始底物(如对香豆酰-CoA、阿魏酰-CoA、肉桂酰-CoA 等)与 malonyl-CoA/丙二酰单元的不同组合,以捕捉可能的多种产物。另一方面是AoPKS5在植物体内的调控机理(如转录因子结合位点、启动子活性)需要进一步挖掘。此外,还可借助基因编辑或过表达策略在模式植物(如本氏烟草)中重构二苯基庚烷生物合成途径,这将有助于评估 AoPKS5 蛋白的生理功能及其在代谢工程中的应用潜力。
-
本研究首次克隆南药益智中AoPKS5基因并对其系统分析,结果显示 AoPKS5 基因编码蛋白为389个氨基酸的Ⅲ型聚酮合酶,并带有 Cys-His-Asn 催化三联体和 GFGPG 序列标签。系统树分析表明,AoPKS5蛋白与姜黄和生姜中的二酮辅酶A合成酶(DCS)蛋白高度同源,推测其在二苯基庚烷类化合物的合成中可能具有相同或相似的催化功能。RT-qPCR 结果显示,AoPKS5基因在益智不同组织中的表达呈显著差异,表达量从高到低依次为叶、花、茎、块茎和果。通过NC克隆的方法,成功构建pNC-ET28-AoPKS5 蛋白表达载体并优化表达条件,获得纯化后的重组AoPKS5蛋白,为AoPKS5蛋白酶学活性验证及益智中功能性次生代谢产物的生物合成机制研究奠定了基础,也为后续代谢工程与天然产物开发提供了重要参考。
Analysis of gene cloning and expression of type III polyketide synthase AoPKS5 in Alpinia oxyphylla
-
摘要: 本研究为探究南药益智(Alpinia oxyphylla)中Ⅲ型聚酮合酶(PKSⅢ)的分子特征及其在二苯基庚烷类化合物合成中的作用,从益智基因组中筛选 PKSⅢ 基因家族,选取候选基因 AoPKS5 进行结构与表达分析。基于基因组与转录组数据进行系统筛选和表达分析,结合系统发育树探讨其进化关系;利用生物信息学预测 AoPKS5 蛋白的理化性质与催化位点;通过RT-qPCR检测其组织表达,并构建 pNC-ET28-AoPKS5 载体在大肠杆菌BL21(DE3)中实现原核表达。结果显示,AoPKS5 全长
1170 bp,编码389 个氨基酸,理论分子量42 kDa,等电点5.96,无信号肽与跨膜区,含典型 Cys–His–Asn 催化三联体和 GFGPG 保守序列。系统发育分析表明其与姜科植物 DCS 蛋白同源性较高;RT-qPCR结果显示在叶中表达最高。研究成功克隆并解析了 AoPKS5 基因,建立其原核表达体系,为阐明 Ⅲ 型 PKS 在益智二苯基庚烷类天然产物生物合成中的作用奠定基础。Abstract: An analysis was made of the molecular characteristics of type III polyketide synthases (PKSIII) in Alpinia oxyphylla and their potential role in the biosynthesis of diphenylheptanoid compounds. The PKSIII gene family was identified from the A. oxyphylla genome, and the candidate gene AoPKS5 was selected for structural and expression analysis. Based on genomic and transcriptomic data, PKSIII genes were systematically screened and analyzed for expression levels, and phylogenetic analysis was conducted to explore their evolutionary relationships. Bioinformatics tools were used to predict the physicochemical properties and catalytic sites of AoPKS5. Tissue-specific expression was analyzed by using RT-qPCR, and the recombinant vector pNC-ET28-AoPKS5 was constructed for heterologous expression in Escherichia coli BL21 (DE3). The results showed that AoPKS5 contained a1170 bp ORF encoding 389 amino acids, with a theoretical molecular weight of 42 kDa and an isoelectric point of 5.96, lacking both signal peptide and transmembrane domains, but possessing the conserved Cys–His–Asn catalytic triad and GFGPG motif. Phylogenetic analysis indicated high homology with DCS proteins from Curcuma longa and Zingiber officinale. RT-qPCR revealed that AoPKS5 was most highly expressed in leaves. The AoPKS5 gene was successfully cloned and characterized and its prokaryotic expression system established, providing a theoretical and technical foundation for elucidating the molecular mechanisms of PKSIII involvement in diphenylheptanoid biosynthesis in A. oxyphylla. -
图 2 不同物种III型聚酮合酶氨基酸序列系统树构建(ML)
注:图中数字表示遗传差异,数字越小表示二者序列相似性高,进化上亲缘关系越近。
Fig. 2 Construction of phylogenetic tree of PKSIII amino acid sequence in different species(ML)
Note: The numbers in the figure represent genetic differences; the smaller the number, the higher the sequence similarity between the two, indicating a closer evolutionary relationship.
图 3 益智不同组织差异表达 AoPKSs 基因
G:果;H:花;J:茎;Y:叶;1,2,3,代表三次重复。
Fig. 3 AoPKSs gene was differently expressed in different tissues of Alpinia oxyphylla
G: Fruit; H: Flower; J: Stem; Y: Leaf; 1, 2, 3 represents three repetitions.
图 4 AoPKS5 与14种植物III型聚酮合酶多序列比对
Fig. 4 Multisequence alignment of AoPKS5 with PKSIII from 14 species of plants
图 5 AoPKS5 基因克隆
M:D2000 分子量标准(100-2000 bp);1,2表示两个重复
Fig. 5 Cloning of AoPKS5 gene
M: D2000 Molecular Weight Standard(100-2000 bp); 1, 2 represents two repetitions
图 7 AoPKS5 蛋白预测分析
A:蛋白亲疏水性;B:蛋白跨膜结构;C:信号肽;D:磷酸化位点。
Fig. 7 AoPKS5 protein prediction analysis
A: Protein hydrophobicity; B: Protein transmembrane structure; C: Signal peptide; D: Phosphorylation site.
图 8 AoPKS5 蛋白结构预测与结构域
注:A: AoPKS5 蛋白的二级结构(蓝色为α-螺旋,紫色为β-折叠,绿色为β转角,黄色为无规则卷曲);B: AoPKS5 蛋白的三级结构;C: AoPKS5 蛋白的结构域
Fig. 8 Structure prediction and domain of AoPKS5 protein
Note: Secondary structure of AoPKS5 protein(blue indicates alpha helix, purple indicates beta sheet, green indicates beta turn, yellow indicates random coil); B: Tertiary structure of AoPKS5 protein; C: Domain of AoPKS5 protein
图 9 益智不同组织 AoPKS5 基因表达量情况
注:每组数据生物学重复3次,统计方法为单因素方差分析和Tukey's 多重比较检验,使用显著性abcd 字母标注法,每个单字母均表示与其他组织有显著差异。
Fig. 9 Expression levels of the AoPKS5 gene in different tissues in A. oxyphylla
Note: Each data set was biologically replicated three times, and the statistical methods used were one-way analysis of variance(ANOVA)and Tukey's multiple comparison test, with significance indicated using the abcd notation. Each single letter represents significant difference from other tissues.
图 10 AoPKS5 蛋白 SDS-PAGE 蛋白凝胶电泳
A图中,M代表Marker;CK代表空载;1、2、3和4分别代表IPTG浓度为0 、0.1 、0.2 和0.5 mmol·L-1。B图中,M代表Marker;CK代表空载;A代表裂解液裂解蛋白后的上清样品;B代表裂解液裂解蛋白后的沉淀样品;C、D、E、F 和G分别代表在0.1 mmol·L-1 IPTG浓度下诱导14 、16 、18h、20 和22 h;H代表经 NI-NTA 纯化后的蛋白。
Fig. 10 SDS-PAGE protein gel electrophoresis of AoPKS5 protein
A: M represents Marker; CK represents no load, and 1, 2, 3 and 4 represent IPTG concentrations of 0, 0.1 , 0.2 and 0.5 mmol·L-1, respectively. B: M represents Marker; CK represents no load; A represents the supernatant after the lysate lysed the protein; B represents the precipitated sample after the lysate lysed the protein; C, D, E, F and G represent the induction at the IPTG concentration of 0.1 mmol·L-1 for 14, 16, 18, 20 and 22 h, respectively; H represents the protein purified by NI-NTA.
-
[1] 中国科学院中国植物志编辑委员会. 中国植物志 第16卷 第2分册[M]. 北京: 科学出版社, 2016: 67-67. [2] 国家药典委员会. 中华人民共和国药典[M]. 北京: 中国医药科技出版社: 2020: 303-304. [3] 吴珊珊, 龚晓猛, 张美, 等. 缩泉丸中益智仁盐炙前后对肾阳虚大鼠肾脏功能的改善作用[J]. 中国实验方剂学杂志, 2016, 22(5), 1−4. https://doi.org/10.13422/j.cnki.syfjx.2016050001 doi: 10.13422/j.cnki.syfjx.2016050001 [4] 李海波, 周汨, 董洁, 等. 益智化学成分及药理作用研究进展[J]. 世界科学技术-中医药现代化, 2024, 26(7), 1870−1887. https://doi.org/10.11842/wst.20230519004 doi: 10.11842/wst.20230519004 [5] BUSAYO F K, YANG J L, DING X P, et al. Identification of volatile compounds and their bioactivities from unpolar fraction of Alpinia oxyphylla Miq. and mining key genes of nootkatone biosynthesis [J]. Natural Product Research, 2024, 38 (10), 1793−1798. https://doi.org/10.1080/14786419.2023.2220063 doi: 10.1080/14786419.2023.2220063 [6] DENG X M, YE Z L, DUAN J Y, et al. Complete pathway elucidation and heterologous reconstitution of (+)-nootkatone biosynthesis from Alpinia oxyphylla [J]. New Phytologist, 2024, 241 (2), 779−792. https://doi.org/10.1111/nph.19375 doi: 10.1111/nph.19375 [7] 罗京超, 冯毅凡, 吉星. 天然线性二苯基庚烷类化合物的研究进展[J]. 中草药, 2008, 39(12), 1912−1916. https://doi.org/10.3321/j.issn:0253-2670.2008.12.054 doi: 10.3321/j.issn:0253-2670.2008.12.054 [8] MORITA H, WONG C P, ABE I. How structural subtleties lead to molecular diversity for the type III polyketide synthases [J]. Journal of Biological Chemistry, 2019, 294 (41), 15121−15136. https://doi.org/10.1074/jbc.REV119.006129 doi: 10.1074/jbc.REV119.006129 [9] FERRER J L, JEZ J M, BOWMAN M E, et al. Structure of chalcone synthase and the molecular basis of plant polyketide biosynthesis [J]. Nature Structural Biology, 1999, 6 (8), 775−784. https://doi.org/10.1038/11553 doi: 10.1038/11553 [10] KREUZALER F, HAHLBROCK K. Enzymatic synthesis of aromatic compounds in higher plants: formation of naringenin (5, 7, 4'-trihydroxyflavanone) from p-coumaroyl coenzyme A and malonyl coenzyme A [J]. Federation of European Biochemical Societies, 1972, 28 (1), 69−72. https://doi.org/10.1016/0014-5793(72)80679-3 doi: 10.1016/0014-5793(72)80679-3 [11] KREUZALER F, RAGG H, FAUTZ E, et al. UV-induction of chalcone synthase mRNA in cell suspension cultures of Petroselinum hortense [J]. Proceedings of the National Academy of Sciences of the United States of America, 1983, 80 (9), 2591−2593. https://doi.org/10.1073/pnas.80.9.2591 doi: 10.1073/pnas.80.9.2591 [12] KATSUYAMA Y, MIYAZONO K I, TANOKURA M, et al. Structural and biochemical elucidation of mechanism for decarboxylative condensation of β-keto acid by curcumin synthase [J]. Journal of Biological Chemistry, 2011, 286 (8), 6659−6668. https://doi.org/10.1074/jbc.M110.196279 doi: 10.1074/jbc.M110.196279 [13] KATSUYAMA Y, KITA T, FUNA N, et al. Curcuminoid biosynthesis by two type III polyketide synthases in the herb Curcuma longa [J]. Journal of Biological Chemistry, 2009, 284 (17), 11160−11170. https://doi.org/10.1074/jbc.M900070200 doi: 10.1074/jbc.M900070200 [14] WANG J, WANG X H, LIU X, et al. Synthesis of unnatural 2-substituted quinolones and 1, 3-diketones by a member of type III polyketide synthases from Huperzia serrata [J]. Organic Letters, 2016, 18 (15), 3550−3553. https://doi.org/10.1021/acs.orglett.6b01501 doi: 10.1021/acs.orglett.6b01501 [15] SAKAMOTO S, MORITA Y, YUSAKUL G, et al. Molecular cloning and characterization of type III polyketide synthase from Plumbago zeylanica [J]. Journal of Asian Natural Products Research, 2021, 23 (5), 478−490. https://doi.org/10.1080/10286020.2020.1754399 doi: 10.1080/10286020.2020.1754399 [16] CHOI G S, CHOO H J, KIM B G, et al. Synthesis of acridone derivatives via heterologous expression of a plant type III polyketide synthase in Escherichia coli [J]. Microbial Cell Factories, 2020, 19 (1), 73. https://doi.org/10.1186/s12934-020-01331-2 doi: 10.1186/s12934-020-01331-2 [17] ZHOU S G, ZHONG K, YAN F X, et al. Biological characteristics of a novel bibenzyl synthase (DoBS1) gene from Dendrobium officinale catalyzing dihydroresveratrol synthesis [J]. Molecules, 2024, 29 (22), 5320. https://doi.org/10.3390/molecules29225320 doi: 10.3390/molecules29225320 [18] CHEN Y, WANG Y, LIANG C J, et al. Characterization of the key bibenzyl synthase in Dendrobium sinense [J]. International Journal of Molecular Sciences, 2022, 23 (12), 6780. https://doi.org/10.3390/ijms23126780 doi: 10.3390/ijms23126780 [19] WANG X H, GAO B W, NAKASHIMA Y, et al. Identification of a diarylpentanoid-producing polyketide synthase revealing an unusual biosynthetic pathway of 2-(2-phenylethyl)chromones in agarwood [J]. Nature Communications, 2022, 13 (1), 348. https://doi.org/10.1038/s41467-022-27971-z doi: 10.1038/s41467-022-27971-z [20] MANOILENKO S, DIPPE M, FUCHS T, et al. Enzymatic one-step synthesis of natural 2-pyrones and new-to-nature derivatives from coenzyme A esters [J]. Journal of Biotechnology, 2024, 388, 72−82. https://doi.org/10.1016/j.jbiotec.2024.04.006 doi: 10.1016/j.jbiotec.2024.04.006 [21] TAKEMOTO K, MIKOTA Y, MORIUCHI R, et al. Cloning of three Alnus sieboldiana type III polyketide synthases and formation of polyketides in recombinant Escherichia coli using cinnamic acid analogs as substrates [J]. Heliyon, 2024, 10 (6), e27698. https://doi.org/10.1016/j.heliyon.2024.e27698 doi: 10.1016/j.heliyon.2024.e27698 [22] PAN K, DAI S P, TIAN J P, et al. Chromosome-level genome and multi-omics analyses provide insights into the geo-herbalism properties of Alpinia oxyphylla [J]. Frontiers in Plant Science, 2023, 14, 1161257. https://doi.org/10.3389/fpls.2023.1161257 doi: 10.3389/fpls.2023.1161257 [23] 李成磊, 张晓伟, 吴琦, 等. 甜荞查尔酮合酶基因Chs的克隆及序列分析[J]. 广西植物, 2011, 31(3), 383−387. https://doi.org/10.3969/j.issn.1000-3142.2011.05.019 doi: 10.3969/j.issn.1000-3142.2011.05.019 [24] ABE I, MORITA H. Structure and function of the chalcone synthase superfamily of plant type III polyketide synthases [J]. Natural Product Reports, 2010, 27 (6), 809−838. https://doi.org/10.1039/b909988n doi: 10.1039/b909988n [25] GOYAL M, CHAUDHURI T K. GroEL-GroES assisted folding of multiple recombinant proteins simultaneously over-expressed in Escherichia coli [J]. The International Journal of Biochemistry & Cell Biology, 2015, 64, 277−286. https://doi.org/10.1016/j.biocel.2015.04.018 doi: 10.1016/j.biocel.2015.04.018 [26] STEWART C, Jr, WOODS K, MACIAS G, et al. Molecular architectures of benzoic acid-specific type III polyketide synthases[J]. Acta Crystallographica Section D: Structural Biology, 2017, 73(Pt 12): 1007-1019. https://doi.org/10.1107/S2059798317016618 doi: 10.1107/S2059798317016618 -

 点击查看大图
点击查看大图
计量
- 文章访问数: 415
- HTML全文浏览量: 350
- 被引次数: 0
