

-
木薯(Manihot esculenta)原产于南美洲,是热带地区重要的粮食和经济作物。目前,中国的福建、广东、海南、广西及云贵等省区都有栽培。在木薯产业发展过程中,生物胁迫和非生物胁迫严重影响了其产量。由菜豆黄单胞菌木薯萎蔫致病变种(Xanthomonas phaseoli pv. manihotis, Xpm)引起的木薯细菌性枯萎病(cassava bacteria blight, CBB),是一种致死性叶片和维管束疾病,可导致木薯产量损失高达75%,严重影响木薯种植的经济可行性和粮食安全[1-2]。
APETALA2/ethylene responsive factor(AP2/ERF)超家族,是植物最大的转录因子超家族之一,参与植物的生长发育和多种生物及非生物胁迫的信号传导过程,在植物应对生物与非生物胁迫机制中发挥重要作用[3]。ERF家族属于AP2/ERF超家族,是一种含有AP2保守结构域(由57~70个高度保守的氨基酸残基组成,其N端含有1个YRG保守元件,C端含有1个RAYD保守元件),以及与其靶基因上游启动子区域中的GCC-box(AGCCGCC)特异性结合的DNA结合域,并能够进一步激活或者抑制乙烯/茉莉酸(ET/JA,ethylene/jasmonic acid)偶联的防御机制的转录因子[4]。GCC-box常位于许多编码致病相关蛋白基因的启动子区内,通过与ERF转录因子结合从而调控基因表达,例如编码β−1,3−葡聚糖酶、几丁质酶和渗透蛋白的基因。当植物受到病原菌侵染,ERF转录因子与GCC-box结合,直接或者间接激活下游病程相关(pathogenesis related,PR)基因,使抗病相关基因在植物响应生物胁迫的过程中发挥作用[5]。大多数植物ERF基因家族分类均根据ERF转录因子的结构域的异同进行分类,例如拟南芥(Arabidopsis thaliana)、水稻(Oryza sativa)和光皮桦(Betula luminifera)[6-8]。前人已在木薯基因组v6.1中挖掘了木薯ERF基因家族成员,并根据拟南芥ERF基因家族分类规则将其分为10类,其中Ⅰ~Ⅳ类是与植物生长发育密切相关,且在应对温度、湿度等非生物胁迫方面起作用的CBF/DREB亚族,Ⅴ~Ⅹ类是在非生物和生物胁迫均发挥作用的ERF亚族[9-11]。在木薯ERF基因家族的Ⅴ~Ⅹ类中,Ⅶ亚族在木薯组织特异性表达中丰度较高,且其表达量明显受到Xpm的诱导[9]。已有研究表明,木薯ERF转录因子参与调控木薯生长发育和胁迫应答。木薯MeERF5基因能快速响应干旱、盐和低温胁迫[12];过表达MeERF127基因的木薯植株可能通过调控下游靶基因的表达,从而提高了木薯的抗旱性和耐盐性[13];木薯MeERF109基因结合MeCBF1启动子上游区域并抑制其表达,进而促进淀粉积累并负调控低温耐受[14]。木薯MeERF72基因可以负调控木薯块根薯肉淀粉的合成[15]。木薯MeERF72过表达可以与MeCPK1相互作用,从而正向调节对Xpm的抗病性[16]。然而,已报道的基因大多数是关于木薯生长发育与非生物胁迫的研究,关于木薯ERF与木薯抵抗细菌性枯萎病抗性的研究仍较少,因此研究可能在木薯抗病中发挥关键作用的木薯ERF Ⅶ亚族基因具有重要意义。
本研究利用生物信息学的手段,对ERF Ⅶ亚族基因进行系统进化、基因结构、顺式作用元件及表达模式分析,并对其编码蛋白进行保守基序和互作关系分析,进一步结合实时荧光定量PCR进行验证分析结果,旨在为解析木薯响应Xpm侵染的分子机制提供基因资源和理论基础。
-
供试的木薯品种:‘TM60444’,种植于海南大学儋州试验基地,由本实验室种植和保存。菌株:由本实验室在广西分离和保存的Xpm11菌株。主要试剂:多糖多酚植物总RNA提取试剂盒(TIANGEN,DP441,北京),FastKing gDNA Dispelling RT SuperMix FastKing一步法除基因组cDNA第一链合成预混试剂(TIANGEN,KR118,北京),TB GREEN Ⅱ(TAKARA,RR820A,北京),DNA Marker(TAKARA,3427A,北京),引物由北京擎科生物科技股份有限公司合成。
-
利用软件Hmmer v3.0和本地blast,以拟南芥ERF基因为探针[17-18],在下载于Phytozome网站(
https://phytozome-next.jgi.doe.gov/ )的木薯基因组v8.1中筛选候选的木薯ERF基因家族成员。运用SMART网站(http://smart.embl.de/ )和CDD网站(https://www.ncbi.nlm.nih.gov/Structure/cdd/cdd.shtml )鉴定候选基因的结构域,保留含有AP2结构域的基因为木薯ERF基因家族成员。MEGA v7.0软件和Evolview网站(https://www.evolgenius.info/evolview/#/ )分别用于构建和美化系统进化树。MEME网站(https://meme-suite.org/meme/ )用于分析木薯ERF基因家族成员的保守基序并用TBtools可视化。利用TBtools软件分析并构建基因的染色体定位图和基因结构图。提取木薯ERF基因家族Ⅶ亚族基因上游2 000 bp的启动子序列,并利用PlantCARE网站(http://bioinformatics.psb.ugent.be/webtools/plantcare/html/ )对其进行预测分析,筛选与植物生长发育、光响应、激素响应和环境胁迫响应相关元件,并运用TBtools进行可视化。PlantTFDB网站用于预测木薯ERF基因家族Ⅶ亚族转录因子的靶基因。以拟南芥为参考,在STRING数据库(https://cn.string-db.org/ )完成蛋白互作网络的构建。不同组织表达的转录组数据(PRJNA324539、GSE82279)下载于GEO数据库(https://www.ncbi.nlm.nih.gov/geo/ ),病原菌侵染后的转录组数据来源于实验室先前已保存的数据,后用TBtools可视化分析。利用WGCNA算法构建基因共表达网络。 -
RNA提取试剂盒提取叶片总RNA,并以1%的琼脂糖凝胶电泳检测提取的总RNA质量。以该RNA为模板,利用FastKing gDNA Dispelling RT SuperMix FastKing一步法除基因组cDNA第一链合成预混试剂(TIANGEN,KR118)进行反转录并获得cDNA。之后用木薯通用内参基因(Manes.15G054700,F: TCCACTGGGTAGATTCGCTGTTA ,R: TAGCTCCACTTGGGTCCTTCTT)检测cDNA质量,保存该cDNA方便进行后续实验。使用NCBI网站设计木薯ERF基因家族Ⅶ亚族基因的qPCR引物,以病原菌处理前和处理后不同时间点的cDNA为模板,在Rotor-Gene Q(QIAGEN)仪器上进行实时荧光定量PCR分析。qPCR体系为:TB GREEN Ⅱ酶10 μL,上下游引物各1 μL,木薯cDNA模板2 μL,ddH2O 6 μL,每个时间点设计3个技术性重复。
-
以拟南芥ERF蛋白序列为探针,在木薯基因组v8.1中鉴定木薯ERF基因家族成员。根据保守结构域,共筛选获得161个木薯ERF蛋白,根据其在染色体上的位置依次命名为MeERF1至MeERF161,具体信息见表1。
表 1 木薯ERF转录因子家族信息
Table 1. Information on cassava ERF transcription factor family
重命名
Rename基因编号
Gen ID蛋白长度/aa
Protein length相对分子质量
Mw等电点
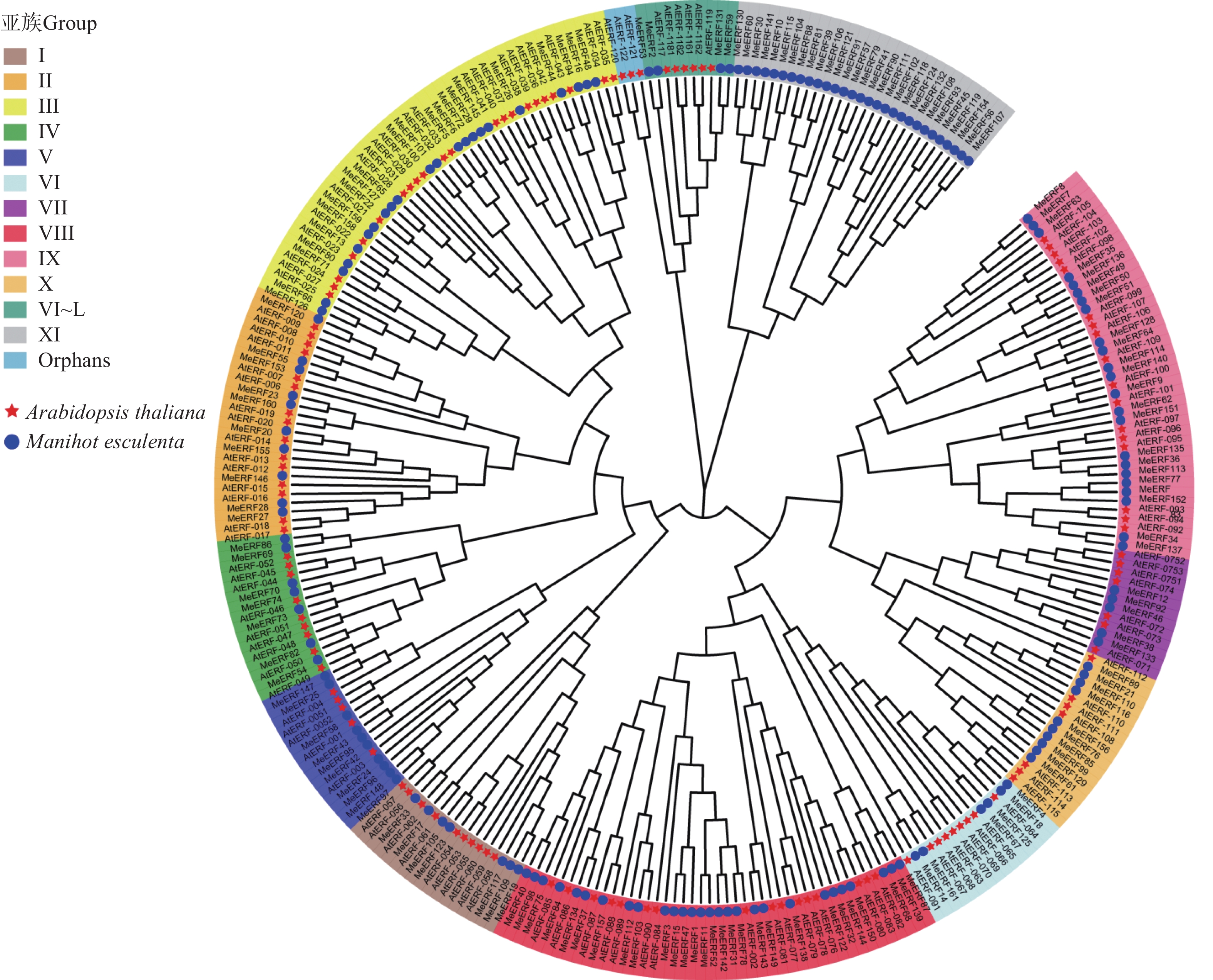
Theoretical pIMeERF1 Manes.01G000062.1 338 35665.73 5.18 MeERF2 Manes.01G012200.6 308 35086.46 5.21 MeERF3 Manes.01G022500.1 220 24351.21 9.49 MeERF4 Manes.01G046200.1 337 37280.05 4.93 MeERF5 Manes.01G067400.1 243 26965.04 5.82 MeERF6 Manes.01G067500.1 233 26178.39 5.27 MeERF7 Manes.01G085100.1 260 28968.13 7.67 MeERF8 Manes.01G085200.1 315 35275.61 6.56 MeERF9 Manes.01G085400.1 289 30917.88 6.36 MeERF10 Manes.01G085800.1 318 36809.66 9.41 MeERF11 Manes.01G101000.1 402 45128.87 4.59 MeERF12 Manes.01G222600.1 309 34577.41 6.06 MeERF13 Manes.01G245200.1 187 20860.34 5.54 MeERF14 Manes.01G249600.1 310 35421.18 4.97 MeERF15 Manes.01G262100.2 283 31426.76 9.07 MeERF16 Manes.01G271300.1 251 27577.35 5.45 MeERF17 Manes.02G010700.1 308 34347.42 8.80 MeERF18 Manes.02G013200.1 335 37300.15 5.26 MeERF19 Manes.02G154400.1 355 39070.65 6.21 MeERF20 Manes.02G161700.1 232 24812.79 6.11 MeERF21 Manes.02G182000.1 458 48601.51 6.39 MeERF22 Manes.02G208000.2 176 19697.98 5.43 MeERF23 Manes.02G208500.1 174 19131.18 5.74 MeERF24 Manes.03G011000.1 201 22830.86 8.69 MeERF25 Manes.03G015400.1 189 21225.84 6.97 MeERF26 Manes.03G025200.1 279 30468.31 5.30 MeERF27 Manes.03G044100.1 206 23073.70 4.94 MeERF28 Manes.03G056100.1 209 23281.35 4.81 MeERF29 Manes.03G058500.1 269 29083.08 4.84 MeERF30 Manes.03G090396.1 374 43206.81 9.13 MeERF31 Manes.03G098000.1 288 32246.12 8.14 MeERF32 Manes.03G106300.1 228 24636.33 9.47 MeERF33 Manes.03G147500.1 332 37682.17 7.25 MeERF34 Manes.03G165100.1 219 25037.03 5.61 MeERF35 Manes.03G165200.1 150 17202.76 6.12 MeERF36 Manes.03G165300.1 140 15483.99 6.31 MeERF37 Manes.03G192400.1 331 37005.58 5.26 MeERF38 Manes.03G199300.1 259 28942.74 6.54 MeERF39 Manes.04G027000.8 504 55441.53 7.13 MeERF40 Manes.04G094600.1 265 29036.53 4.79 MeERF41 Manes.04G100700.1 330 37395.26 8.61 MeERF42 Manes.04G105000.1 179 20194.42 7.13 MeERF43 Manes.04G114000.1 178 20154.76 8.94 MeERF44 Manes.04G131900.1 231 25233.03 4.75 MeERF45 Manes.04G144500.1 573 63762.94 6.18 MeERF46 Manes.04G150100.4 381 42281.90 4.75 MeERF47 Manes.05G040000.1 302 33130.72 8.92 MeERF48 Manes.05G052000.1 244 27100.90 5.57 MeERF49 Manes.05G054200.1 222 24764.44 4.80 MeERF50 Manes.05G054300.1 222 24887.60 4.97 MeERF51 Manes.05G054400.1 230 25481.27 5.35 MeERF52 Manes.05G106400.1 368 41238.68 4.77 MeERF53 Manes.05G120878.1 319 36175.14 4.59 MeERF54 Manes.05G153700.1 233 25772.96 5.32 MeERF55 Manes.05G175800.1 152 17157.18 9.12 MeERF56 Manes.05G184000.1 661 72872.84 6.60 MeERF57 Manes.06G032500.1 353 40399.73 6.33 MeERF58 Manes.06G058200.1 211 23455.48 6.97 MeERF59 Manes.06G068900.1 350 38842.80 4.82 MeERF60 Manes.06G071700.1 369 41012.18 9.07 MeERF61 Manes.06G127000.1 247 27672.40 9.24 MeERF62 Manes.06G131100.1 248 27664.05 7.07 MeERF63 Manes.06G131200.1 285 31753.29 9.21 MeERF64 Manes.06G131300.1 212 23612.22 5.67 MeERF65 Manes.06G143102.1 231 25542.74 5.49 MeERF66 Manes.06G143200.1 230 24770.37 5.39 MeERF67 Manes.06G156400.1 372 41907.57 4.70 MeERF68 Manes.07G114500.1 236 25810.98 9.26 MeERF69 Manes.07G131900.1 344 37015.33 6.38 MeERF70 Manes.07G135300.2 388 42958.18 4.94 MeERF71 Manes.08G021600.1 199 21599.19 5.42 MeERF72 Manes.08G022000.1 202 22777.86 6.66 MeERF73 Manes.08G048500.1 294 33611.76 6.02 MeERF74 Manes.08G096900.1 421 47903.26 6.08 MeERF75 Manes.08G099800.1 274 29891.50 4.68 MeERF76 Manes.08G101300.1 250 28280.07 5.81 MeERF77 Manes.08G137700.1 222 25471.20 4.91 MeERF78 Manes.08G145400.1 277 31562.01 5.52 MeERF79 Manes.09G020700.1 427 47250.60 8.53 MeERF80 Manes.09G058852.1 204 21585.89 5.64 MeERF81 Manes.09G080100.1 458 51437.98 6.18 MeERF82 Manes.09G146100.1 200 21356.54 5.12 MeERF83 Manes.09G149000.1 222 25191.10 5.15 MeERF84 Manes.09G173200.1 232 25325.83 4.51 MeERF85 Manes.09G188100.1 231 26127.79 6.30 MeERF86 Manes.10G010300.1 332 35806.27 6.20 MeERF87 Manes.10G031600.1 240 26187.29 8.84 MeERF88 Manes.10G041100.3 478 53902.31 7.10 MeERF89 Manes.10G056300.1 263 29211.72 8.22 MeERF90 Manes.10G063200.1 422 47166.90 5.12 MeERF91 Manes.10G119500.4 513 57285.11 5.95 MeERF92 Manes.11G013700.7 385 42590.45 4.86 MeERF93 Manes.11G020300.1 560 62316.43 6.53 MeERF94 Manes.11G035600.1 236 25593.38 5.11 MeERF95 Manes.11G055000.1 196 21564.58 9.62 MeERF96 Manes.11G064100.1 178 20254.65 6.31 MeERF97 Manes.11G064200.1 178 20096.35 6.17 MeERF98 Manes.11G074500.1 278 30326.10 4.78 MeERF99 Manes.11G076500.1 280 31229.21 6.25 MeERF100 Manes.11G158533.1 217 23760.89 8.46 MeERF101 Manes.11G158600.1 217 23760.89 8.46 MeERF102 Manes.12G002400.1 706 77507.62 6.04 MeERF86 Manes.10G010300.1 332 35806.27 6.2 MeERF87 Manes.10G031600.1 240 26187.29 8.84 MeERF88 Manes.10G041100.3 478 53902.31 7.10 MeERF89 Manes.10G056300.1 263 29211.72 8.22 MeERF90 Manes.10G063200.1 422 47166.90 5.12 MeERF91 Manes.10G119500.4 513 57285.11 5.95 MeERF92 Manes.11G013700.7 385 42590.45 4.86 MeERF93 Manes.11G020300.1 560 62316.43 6.53 MeERF94 Manes.11G035600.1 236 25593.38 5.11 MeERF95 Manes.11G055000.1 196 21564.58 9.62 MeERF96 Manes.11G064100.1 178 20254.65 6.31 MeERF97 Manes.11G064200.1 178 20096.35 6.17 MeERF98 Manes.11G074500.1 278 30326.10 4.78 MeERF99 Manes.11G076500.1 280 31229.21 6.25 MeERF100 Manes.11G158533.1 217 23760.89 8.46 MeERF101 Manes.11G158600.1 217 23760.89 8.46 MeERF102 Manes.12G002400.1 706 77507.62 6.04 MeERF103 Manes.12G022600.3 414 45296.72 6.21 MeERF104 Manes.12G047500.5 232 26223.60 9.94 MeERF105 Manes.12G087500.1 471 52295.81 6.02 MeERF106 Manes.12G106400.1 510 56821.22 6.96 MeERF107 Manes.12G117800.1 677 75540.73 6.42 MeERF108 Manes.12G125500.1 496 54660.26 5.81 MeERF109 Manes.12G136800.1 361 40158.47 6.08 MeERF110 Manes.12G146000.1 349 38119.39 5.89 MeERF111 Manes.13G001800.1 701 76935.96 6.21 MeERF112 Manes.13G023700.1 403 43709.99 6.43 MeERF113 Manes.13G033000.1 117 13330.77 5.39 MeERF114 Manes.13G033100.1 146 16311.89 8.96 MeERF115 Manes.13G049600.3 235 26376.60 9.73 MeERF116 Manes.13G081100.1 316 34021.08 8.95 MeERF117 Manes.13G093300.1 358 39861.37 5.90 MeERF118 Manes.13G100600.1 503 55506.27 5.86 MeERF119 Manes.13G108800.1 632 70853.51 6.27 MeERF120 Manes.13G120400.1 191 21567.66 8.70 MeERF121 Manes.13G120700.1 484 53738.25 8.68 MeERF122 Manes.13G143500.1 215 23535.44 9.30 MeERF123 Manes.13G148300.1 474 52704.53 6.28 MeERF124 Manes.14G000900.2 544 59966.77 6.12 MeERF125 Manes.14G019032.1 345 38561.03 5.07 MeERF126 Manes.14G029500.2 230 24322.77 4.78 MeERF127 Manes.14G029600.1 219 24140.19 5.11 MeERF128 Manes.14G038900.1 236 26378.58 6.00 MeERF129 Manes.14G044700.1 243 26918.67 9.80 MeERF130 Manes.14G098000.1 366 40348.38 9.24 MeERF131 Manes.14G101900.1 358 40001.20 5.08 MeERF132 Manes.14G154900.1 562 61094.18 6.95 MeERF133 Manes.15G009900.1 269 29932.67 6.03 MeERF134 Manes.15G014600.1 315 34961.60 5.28 MeERF135 Manes.15G039500.1 132 14731.29 6.37 MeERF136 Manes.15G039600.1 162 18420.07 7.97 MeERF137 Manes.15G039700.1 210 23781.70 5.77 MeERF138 Manes.15G066800.1 236 25840.91 8.91 MeERF139 Manes.15G066900.1 233 25825.94 9.42 MeERF140 Manes.15G092200.1 142 15906.60 6.31 MeERF141 Manes.15G102800.1 371 42235.59 8.78 MeERF142 Manes.15G108400.1 294 32691.73 8.79 MeERF143 Manes.16G030400.1 162 17410.50 8.07 MeERF144 Manes.16G030600.1 252 27628.79 9.24 MeERF145 Manes.16G075800.1 264 28999.09 5.13 MeERF146 Manes.16G090900.1 159 17690.76 9.20 MeERF147 Manes.16G121500.1 192 21619.25 6.59 MeERF148 Manes.16G126900.1 203 22932.96 8.99 MeERF149 Manes.17G048200.1 161 17432.51 9.62 MeERF150 Manes.17G048400.1 248 27075.57 9.47 MeERF151 Manes.17G051600.1 186 20613.92 9.29 MeERF152 Manes.17G051700.1 280 31615.20 6.34 MeERF153 Manes.18G040400.1 152 17222.06 8.55 MeERF154 Manes.18G050000.1 683 75301.40 6.23 MeERF155 Manes.18G075600.1 222 23871.83 5.45 MeERF156 Manes.18G093800.1 410 43467.85 5.91 MeERF157 Manes.18G097200.1 150 17004.09 6.73 MeERF158 Manes.18G118800.1 192 21220.90 4.85 MeERF159 Manes.18G118900.3 193 21114.49 5.41 MeERF160 Manes.18G119500.1 178 19721.91 7.01 MeERF161 Manes.18G125700.1 267 30424.46 6.27 为了分析木薯和拟南芥ERF基因家族的系统进化关系,利用234个拟南芥ERF和161个木薯ERF蛋白构建种间系统发育进化树(图1)。从图1可知,木薯ERF基因家族分为13个亚族,包括Ⅰ~Ⅹ和Ⅵ~L亚族,一组只含有拟南芥ERF的Orphans亚族,以及仅含有29个MeERFs的Ⅺ亚族。木薯ERF基因家族分类与拟南芥ERF基因家族分类大部分一致,说明大部分木薯ERF蛋白在长期进化中并未遗失。除了仅含有MeERFs的Ⅺ亚族,第Ⅷ亚族和第Ⅵ~L亚族分别含有28个和4个MeERFs,分别是含有木薯ERF基因数量最多和最少的亚族。
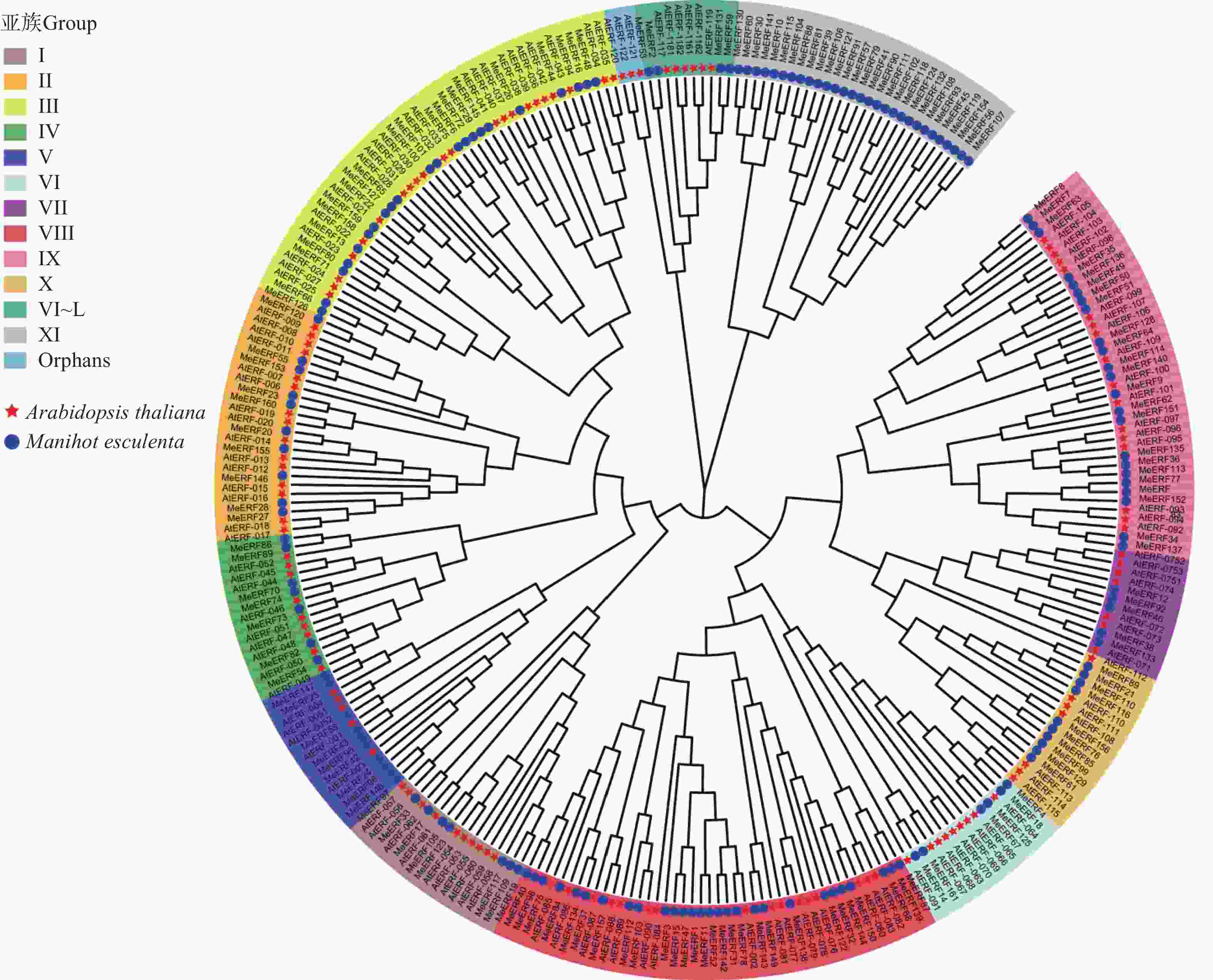
图 1 ERF基因家族系统发育关系
Figure 1. Phylogenetic relationships of ERF gene family
-
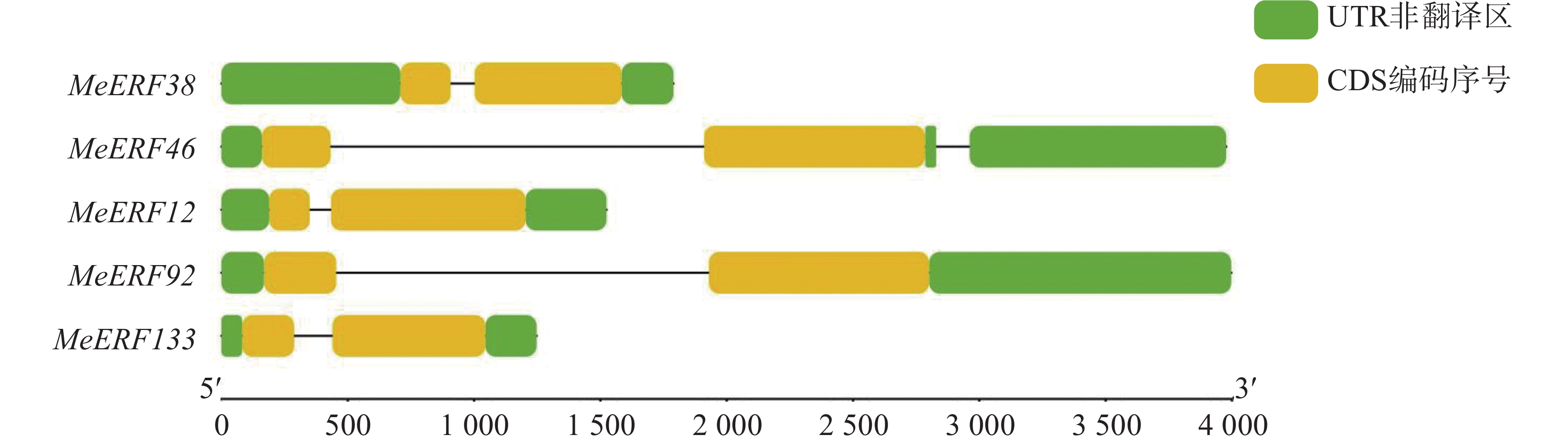
运用TBtools软件分析木薯ERF基因家族Ⅶ亚族成员的基因结构。从图2可知,木薯ERF基因家族Ⅶ亚族所有成员均含有2个外显子,非编码区的数量为2~3个。该结果暗示,木薯ERF基因家族Ⅶ亚族成员的功能和亚细胞定位的差异可能受基因结构的影响[19](表2,图2)。
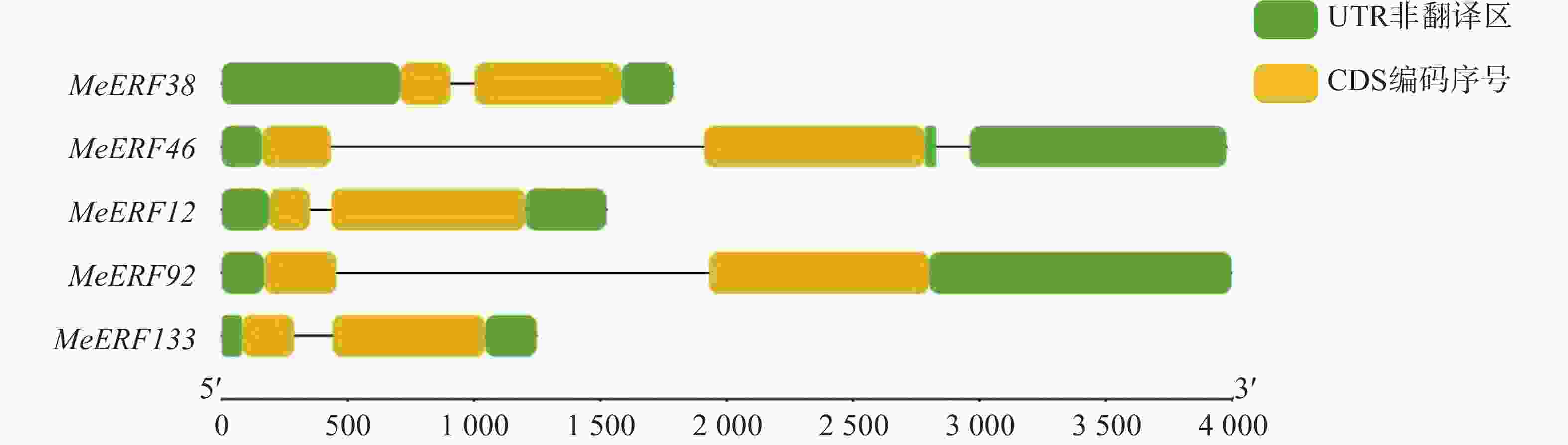
图 2 木薯ERF基因家族Ⅶ亚族基因结构
Figure 2. Gene structure of Ⅶ subfamily genes in cassava ERF gene family
表 2 木薯ERF基因家族Ⅶ亚族保守基序
Table 2. Conserved motifs of cassava ERF Ⅶ subfamily genes
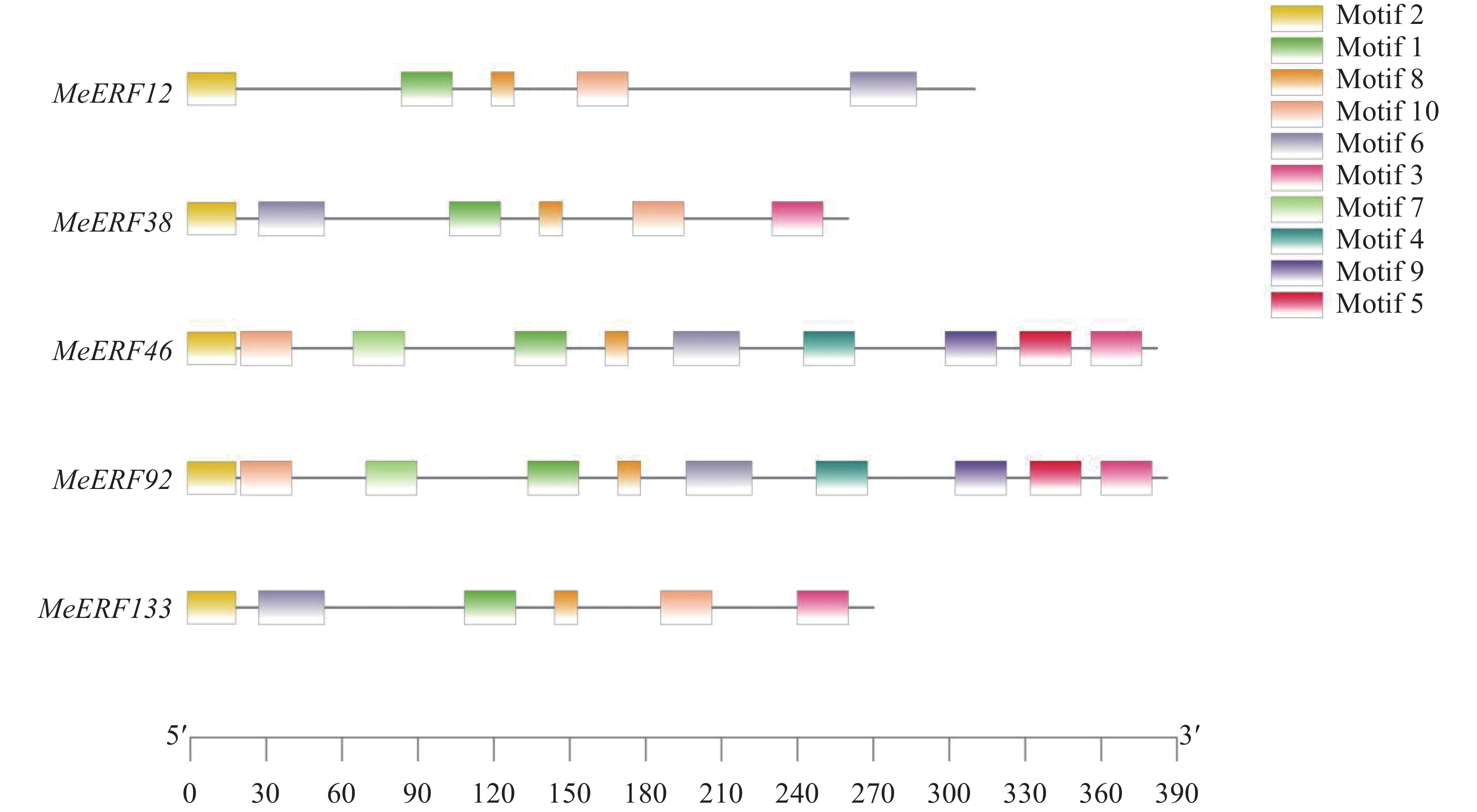
保守基序 Motif 基序氨基酸序列 Sequence Motif1 IYRGIRQRPWGKWAAEIRDPRKGVRVWLGTFNTAEEAARAYDEEARRIRG Motif2 MCGGAIISDFIPVTRARR Motif3 QLGGSSQDGGEPVDLWSFDDLPNHVGGQR Motif4 LMDSFPANGDGANKSIPPCDNVPMFFNSDQGSNSFECSDFGWGEQASKTP Motif5 NQMNFQMPYLEGSWE Motif6 LDTYNKFLGLDYLBNABPDYFSTQGSL Motif7 FQEFKDESDVDEEDDVLFDVKPFAFSATASPPPRNRSPSRGSTAVKSVEF Motif8 KKAKVNFPDE Motif9 EIDESLFMEDANPKKKIKADSENAVPIEENNGKSPSEELLAF Motif10 RLTADFLAPPLKKPIGVGY 运用MEME网站并以保守基序数目10为条件,对木薯ERF基因家族Ⅶ亚族成员进行保守基序分析。分析结果显示,所有MeERFs均具有包括了AP2结构域的motif1,MeERF46和MeERF92含有10个种类的motifs,其余3个MeERFs仅含有motif1、motif2、motif6、motif8和motif10(图3,表2)。木薯ERF基因家族Ⅶ亚族成员具有类型和数量相似的保守基序,推测该亚族成员的生物学功能相似。
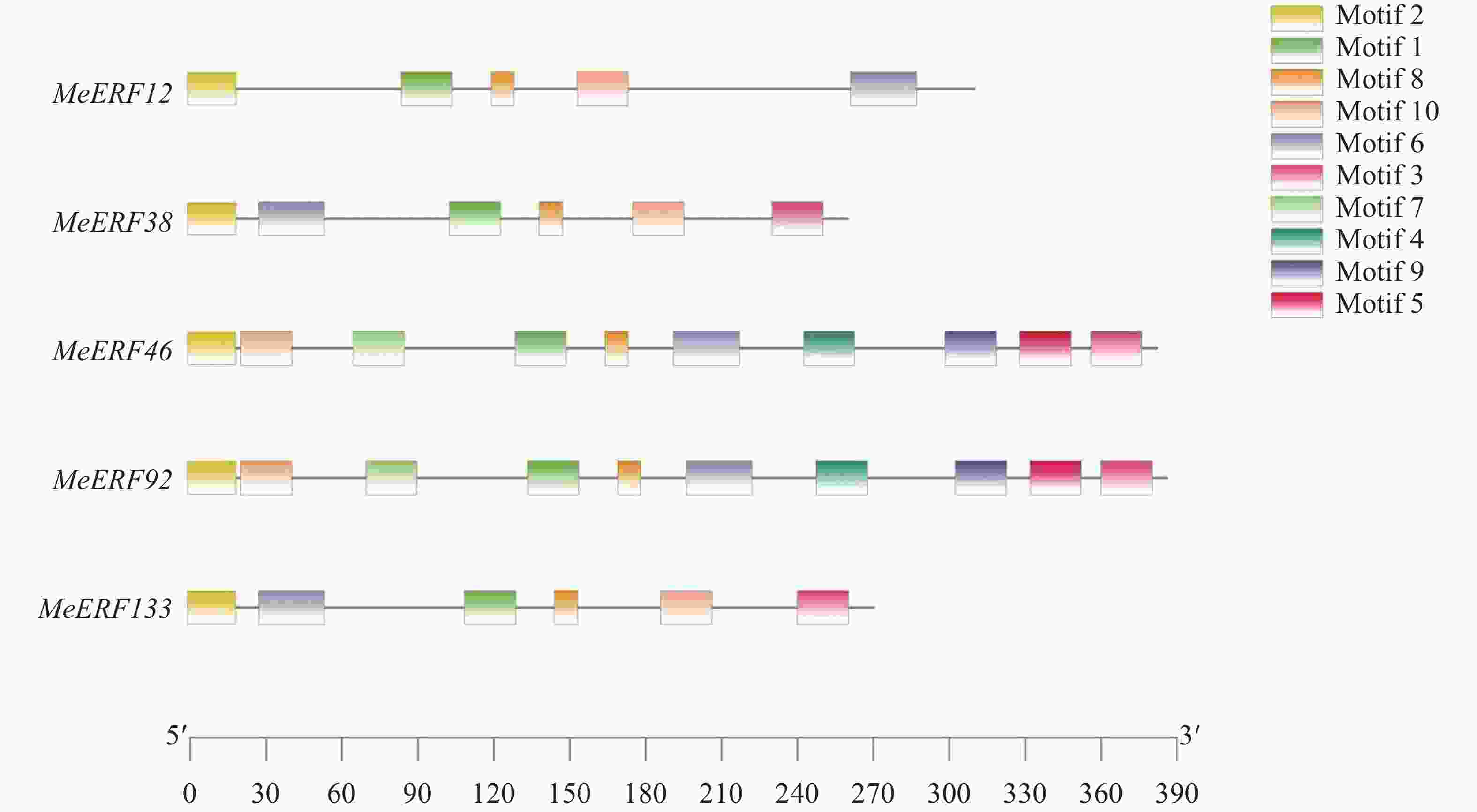
图 3 木薯ERF基因家族Ⅶ亚族保守基序分析
Figure 3. Conserved motif analysis of Ⅶ subfamily of cassava ERF gene family
-
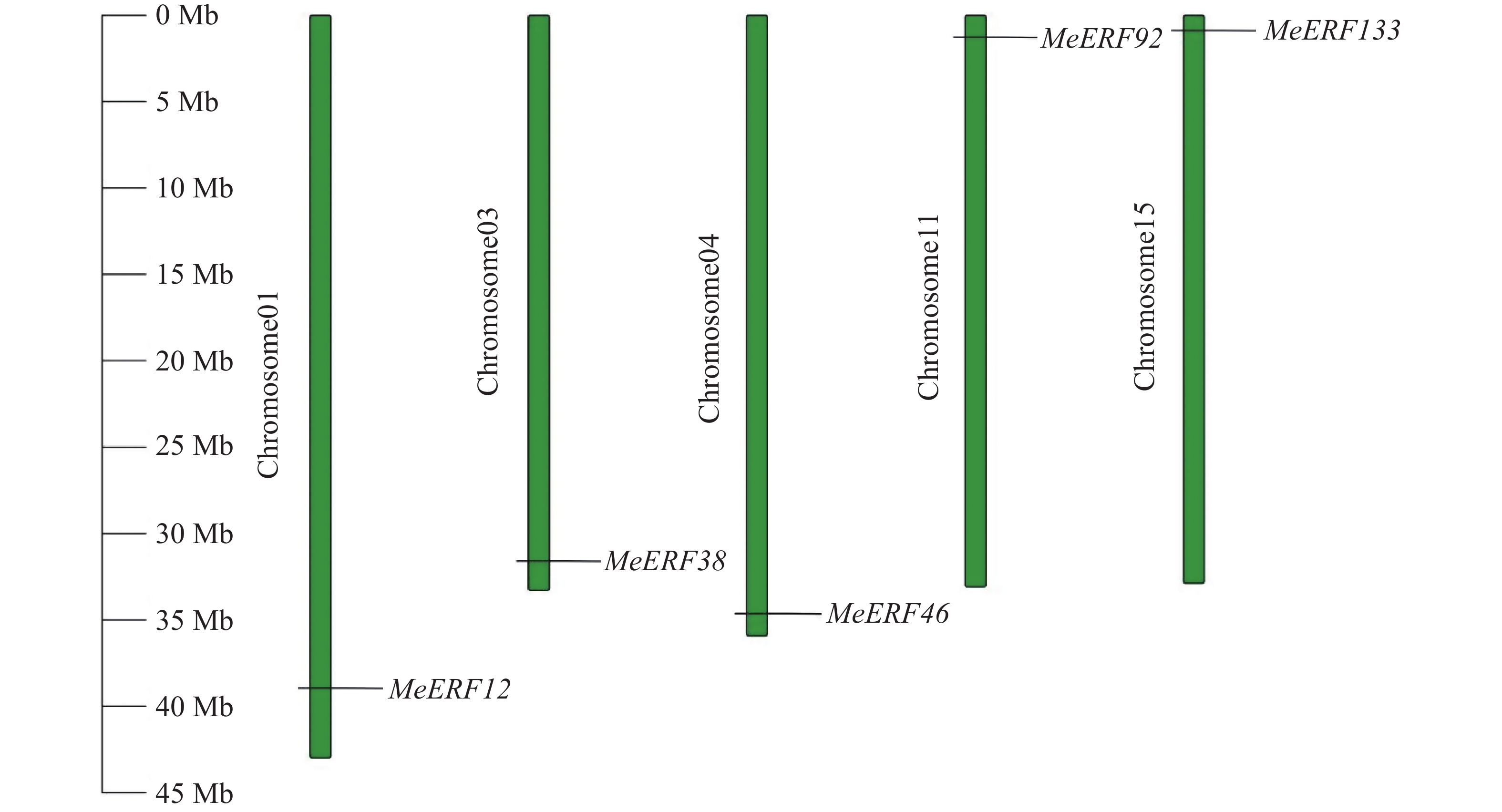
对木薯ERF基因家族Ⅶ亚族的5个基因进行染色体位置分布分析,结果如图4所示。木薯ERF基因家族Ⅶ亚族的成员均分布在染色体两端。其中,60%的基因(MeERF12、MeERF38和MeERF46)分布在染色体下端,其余基因(MeERF92和MeERF133)分布在染色体上端。5个基因分别定位于5条不同的染色体,MeERF12、MeERF38和MeERF46分别定位于1号、3号和4号染色体,MeERF92和MeERF133分别定位于11号和15号染色体。
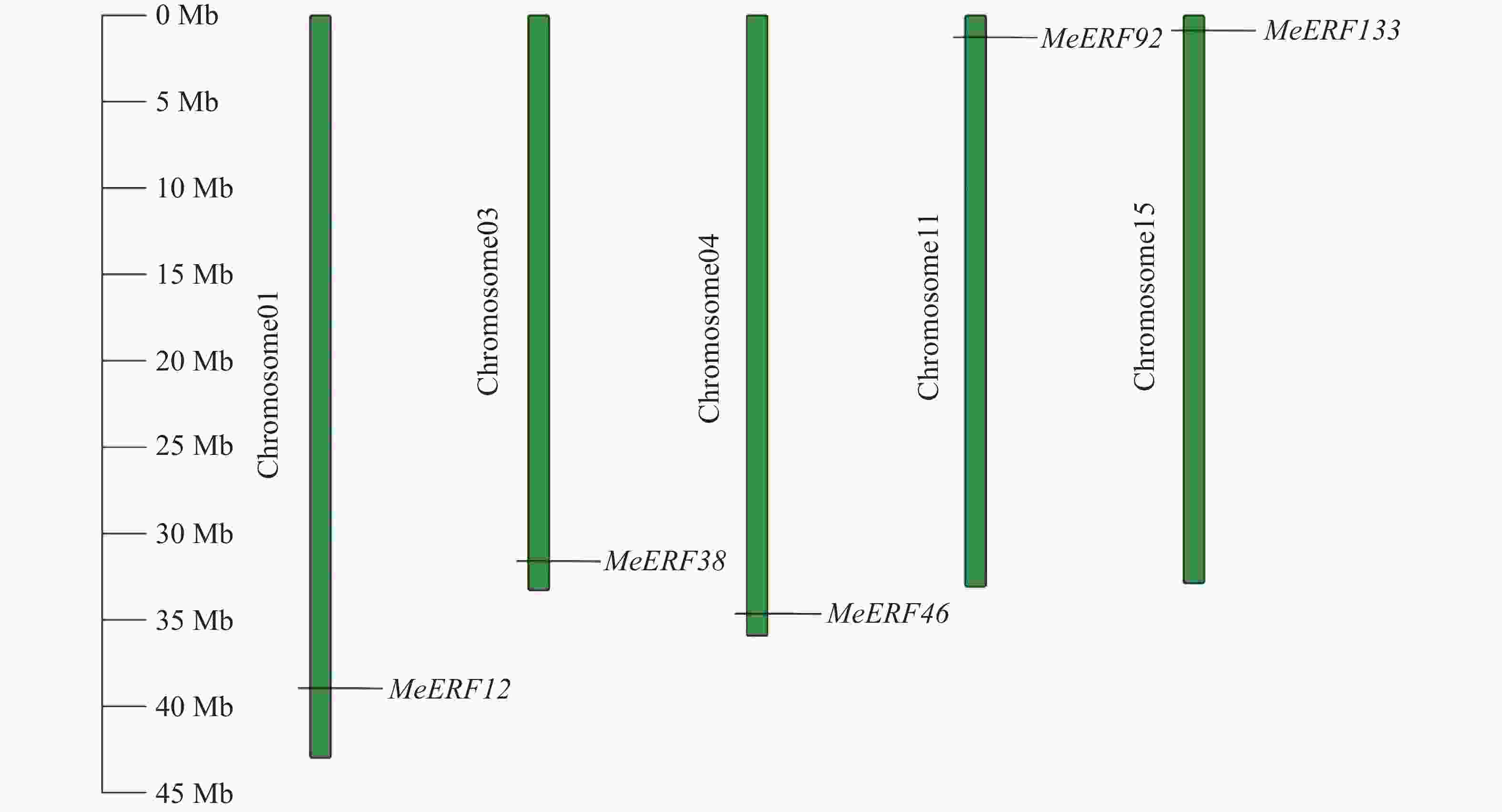
图 4 木薯ERF基因家族Ⅶ亚族成员的染色体定位
Figure 4. Chromosome localization of cassava ERF Ⅶ subfamily
-
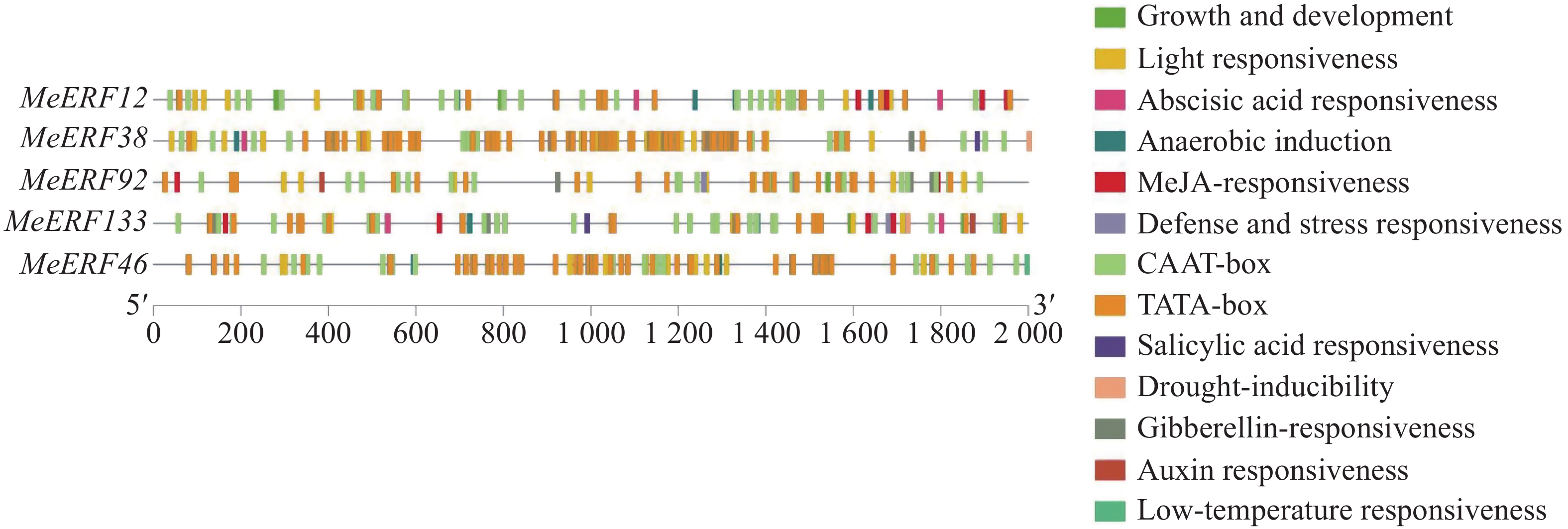
提取木薯ERF基因家族Ⅶ亚族基因上游2 000 bp的启动子序列进行顺式作用元件分析。由图5可知,木薯ERF基因家族Ⅶ亚族基因均包含具有启动子活性的TATA-box和CAAT-box元件,并且主要含有13种顺式作用元件,分别参与植物的生长发育、光响应、激素响应和环境胁迫响应等过程。木薯ERF基因家族Ⅶ亚族所有成员都含有光响应元件,是数量最多的作用元件。4个木薯ERFs含有生长发育相关的反应元件。此外,激素和胁迫响应的元件类型和数目也较多,包括5种激素相关作用元件和3种胁迫相关作用元件。水杨酸、赤霉素和脱落酸作为激素,其相关作用元件与胁迫响应有关。在5个基因中,MeERF133包含所有激素相关作用元件,且含有的脱落酸响应元件数量最多。MeERF38和MeERF133含有响应干旱胁迫的顺式作用元件,仅有MeERF46含有响应低温胁迫的顺式作用元件。顺式作用元件分析的结果表明,该亚族成员的表达可能受到光照、多种激素和环境胁迫等因素的诱导。
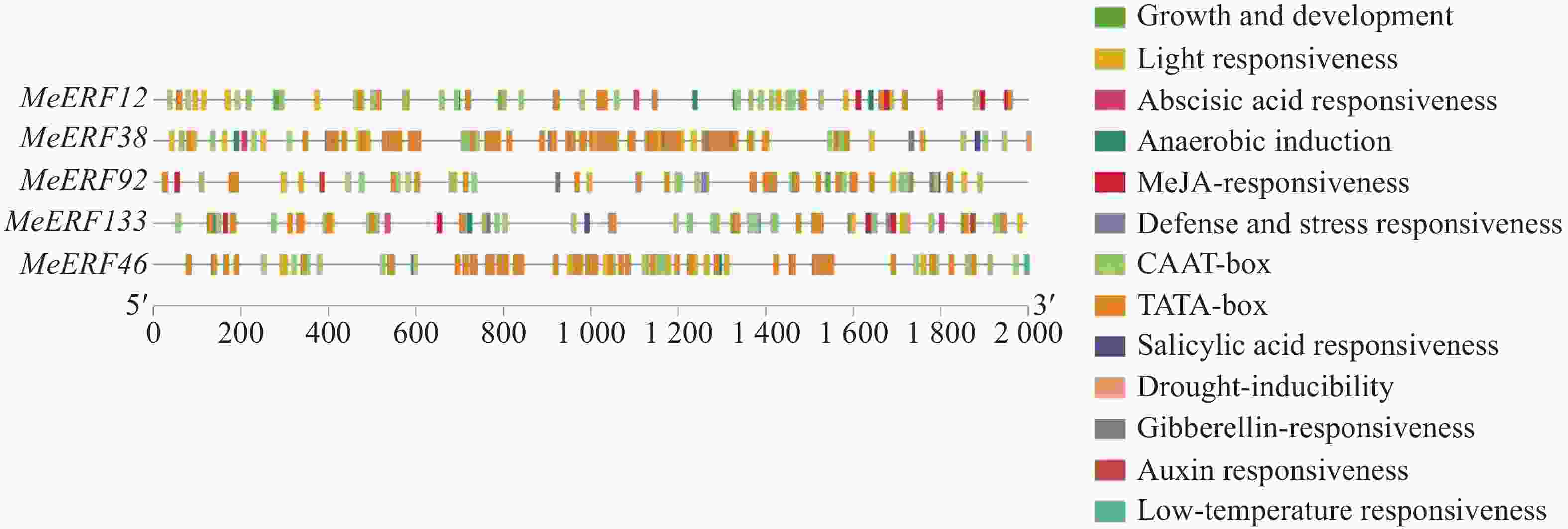
图 5 木薯ERF基因家族Ⅶ亚族启动子顺式作用元件分析
Figure 5. Analysis of cis acting elements of the Ⅶ subfamily promoter of cassava ERF gene family
-
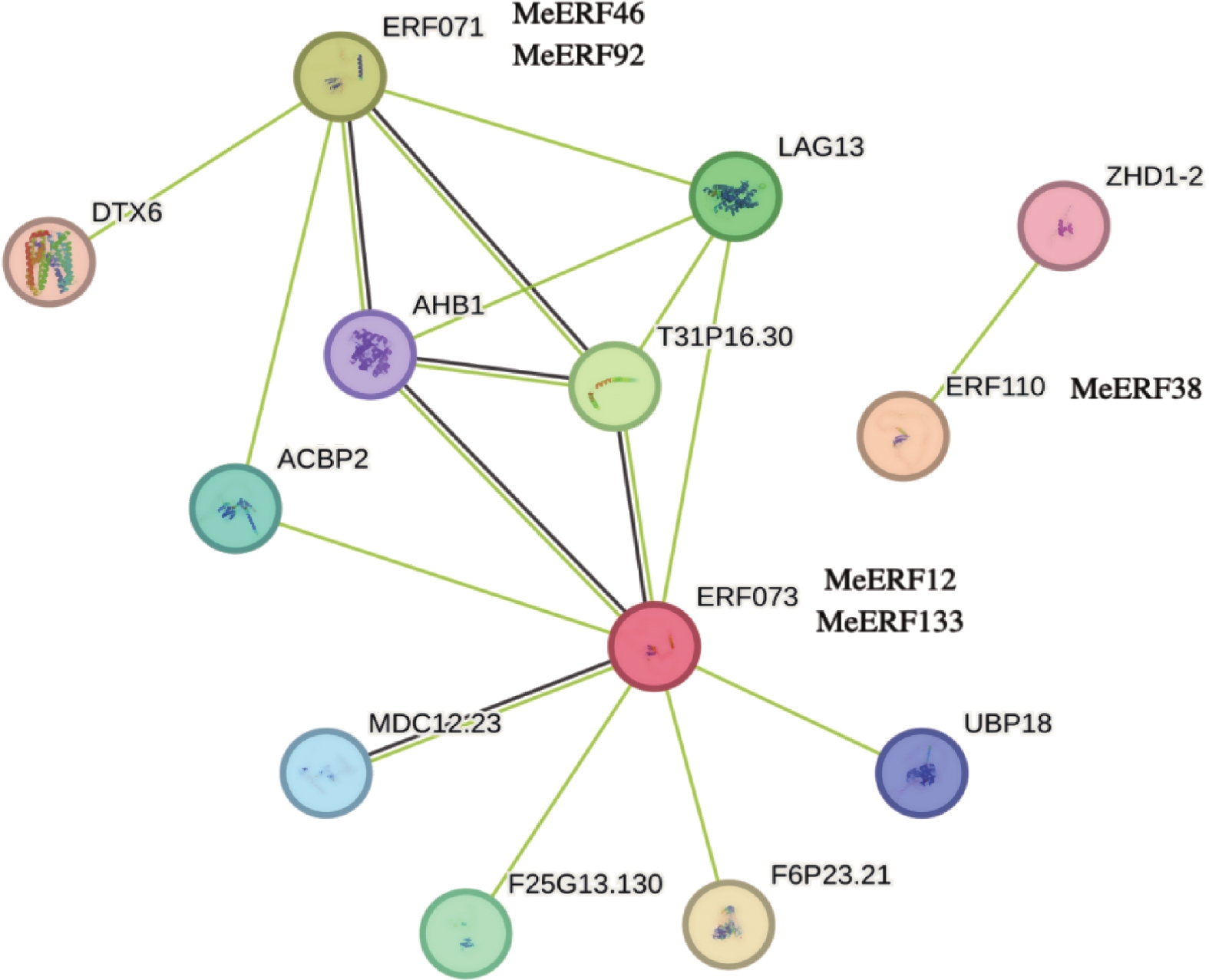
以拟南芥数据库为参考,通过STRING网站构建蛋白互作网络。如图6所示,MeERF46和MeERF92的同源蛋白是ERF071,MeERF12和MeERF133的同源蛋白为ERF073,MeERF38是ERF110的同源蛋白。由蛋白互作网络图可知,ERF073拥有最多的互作蛋白,而ERF110只有一个互作蛋白(Zinc-finger homeodomain protein 2, ZHD1-2),推测MeERF12和MeERF133可能与更多的蛋白有互作关系(图6)。ERF071和ERF073蛋白与重组拟南芥非共生血红蛋白1(Arabidopsis thaliana Non-symbiotic hemoglobin 1, AHB1)、跨膜蛋白(Transmembrane protein, T31P16.30)、LAG1长寿保证同源物3(LAG1 longevity assurance homolog 3, LAG13)和酰基辅酶A结合蛋白2(Acyl-CoA-binding domain-containing protein 2, ACBP2)4种蛋白都有直接的互作关系。该结果暗示,4种MeERF(MeERF46、MeERF92、MeERF12、MeERF133)可能是这4类蛋白的互作蛋白。
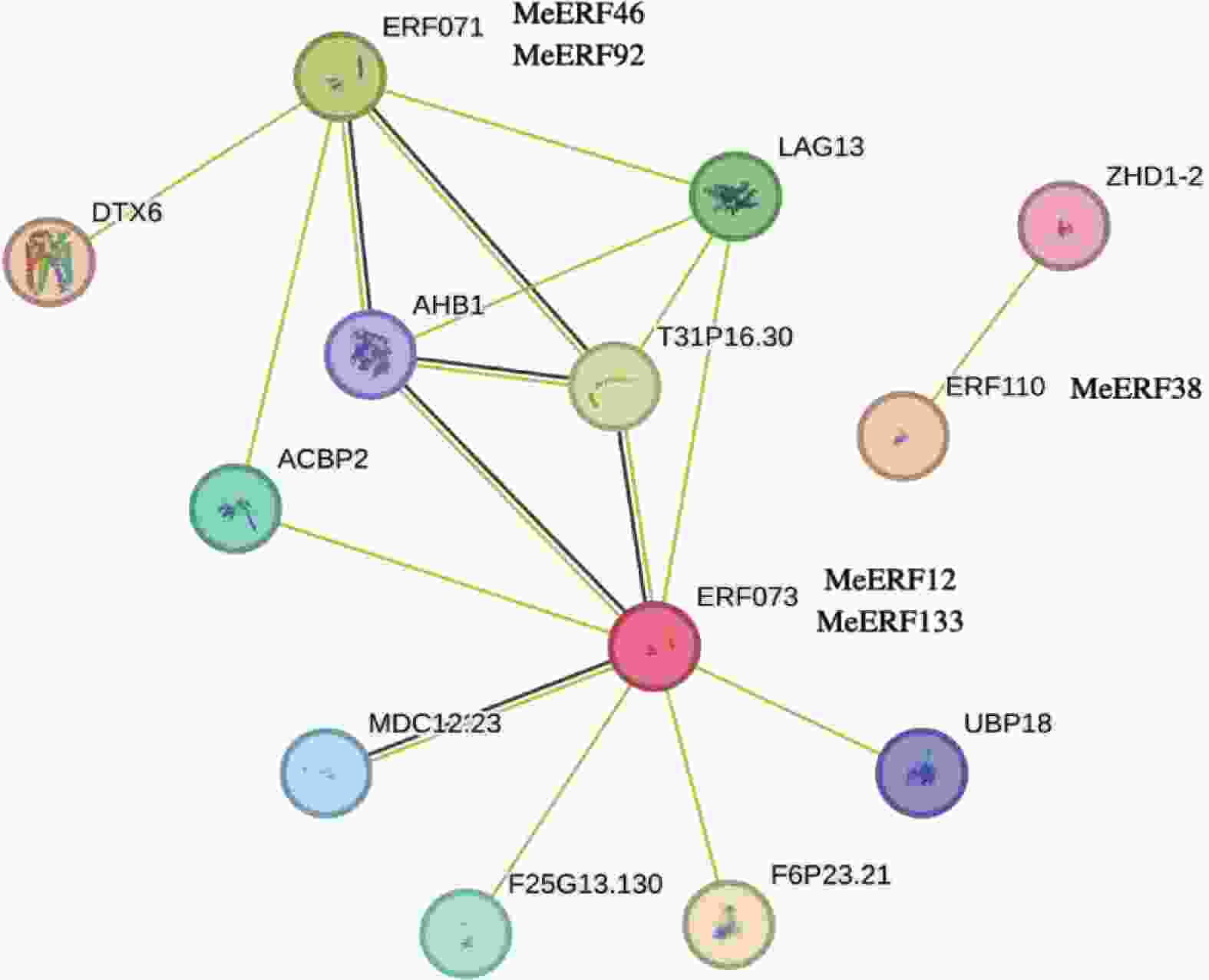
图 6 木薯ERF基因家族Ⅶ亚族的互作蛋白网络图
Figure 6. Protein-protein interaction of cassava ERF Ⅶ subfamily
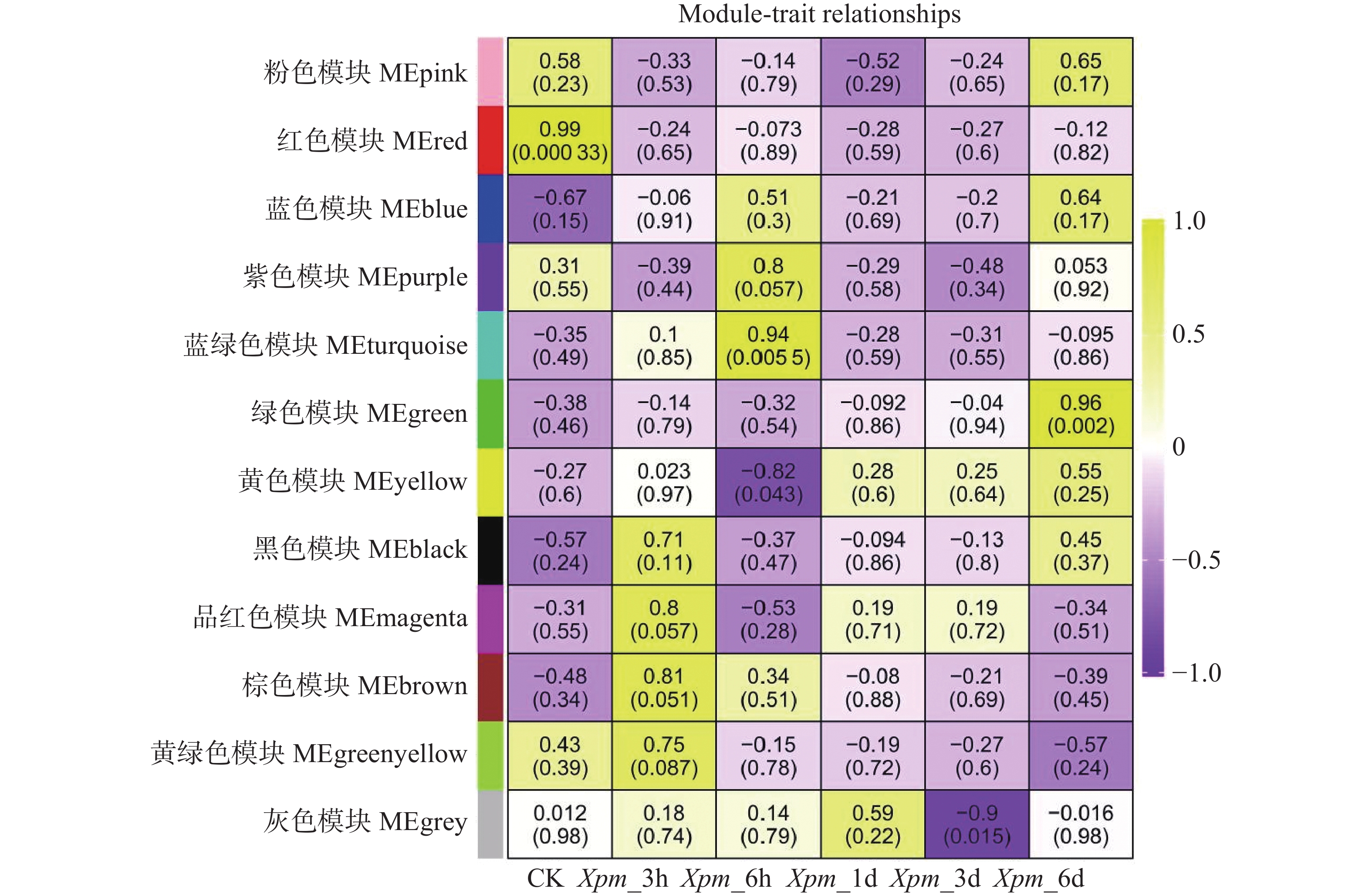
利用PlantTFDB网站对木薯ERF基因家族Ⅶ亚族基因进行靶基因预测,仅检索到MeERF133和MeERF92的所有靶基因信息。预测获得的所有靶基因分布在1号到18号染色体上,其中MeERF133的靶基因有1 902个,MeERF92的靶基因有4 147个,其中979个为共有靶基因。利用WGCNA包构建共表达网络,并根据推荐阈值将其分为12个模块(图7),其中MEturquoise组和MEgreen组的基因分别在Xpm_6h和Xpm_6d处理组中具有很高的正相关性。该结果表明这2组中的基因可能在不同时间点协同表达,以响应Xpm侵染。MEyellow和MEgrey组的基因分别在Xpm_6h和Xpm_3d处理组中具有很高的负相关性,表明这两组中的基因可能在不同时间点响应Xpm的侵染。
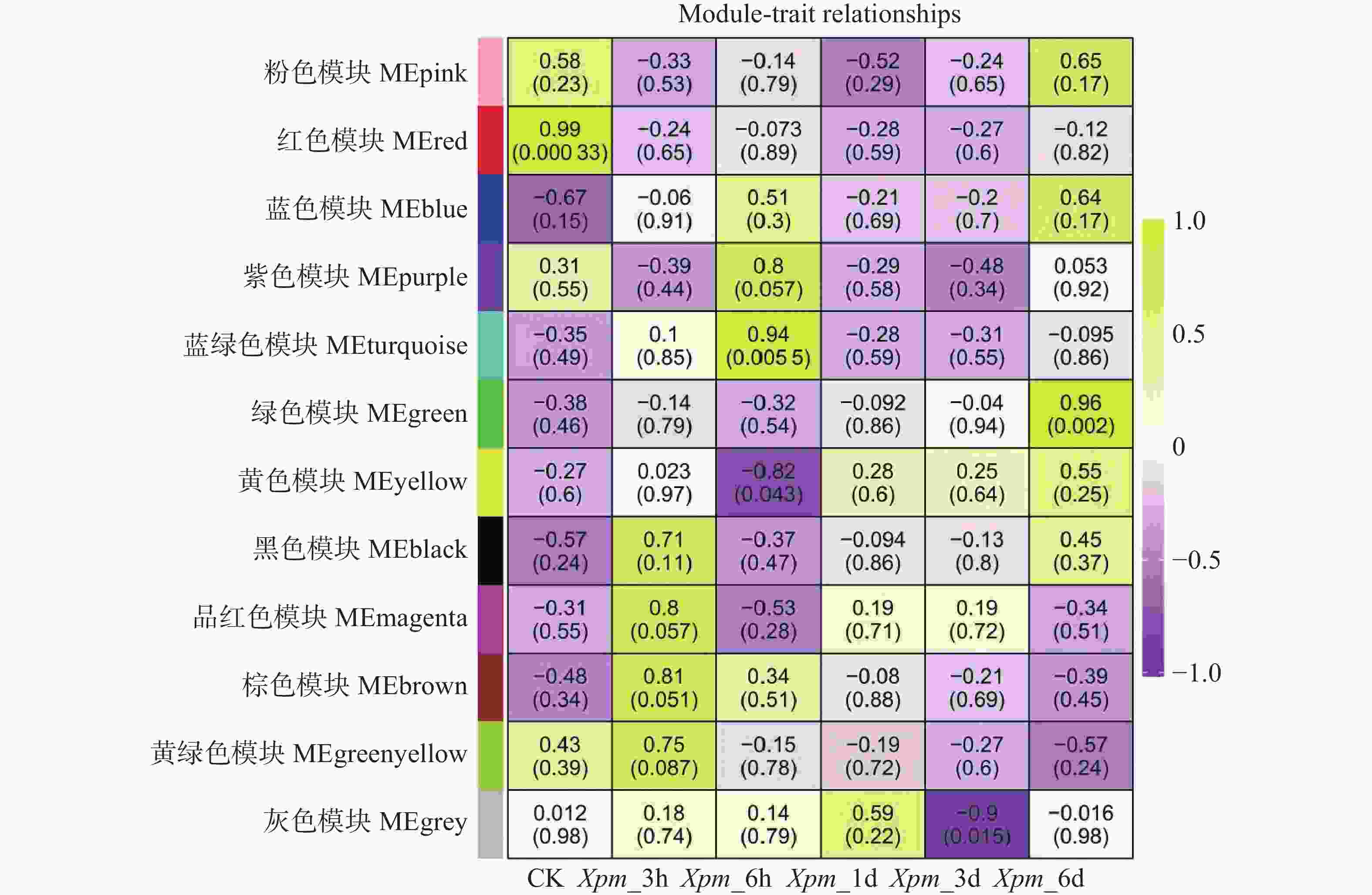
图 7 木薯ERF基因家族Ⅶ亚族基因共表达网络分析
Figure 7. Co-expression network of Ⅶ subfamily genes in cassava ERF gene family
-
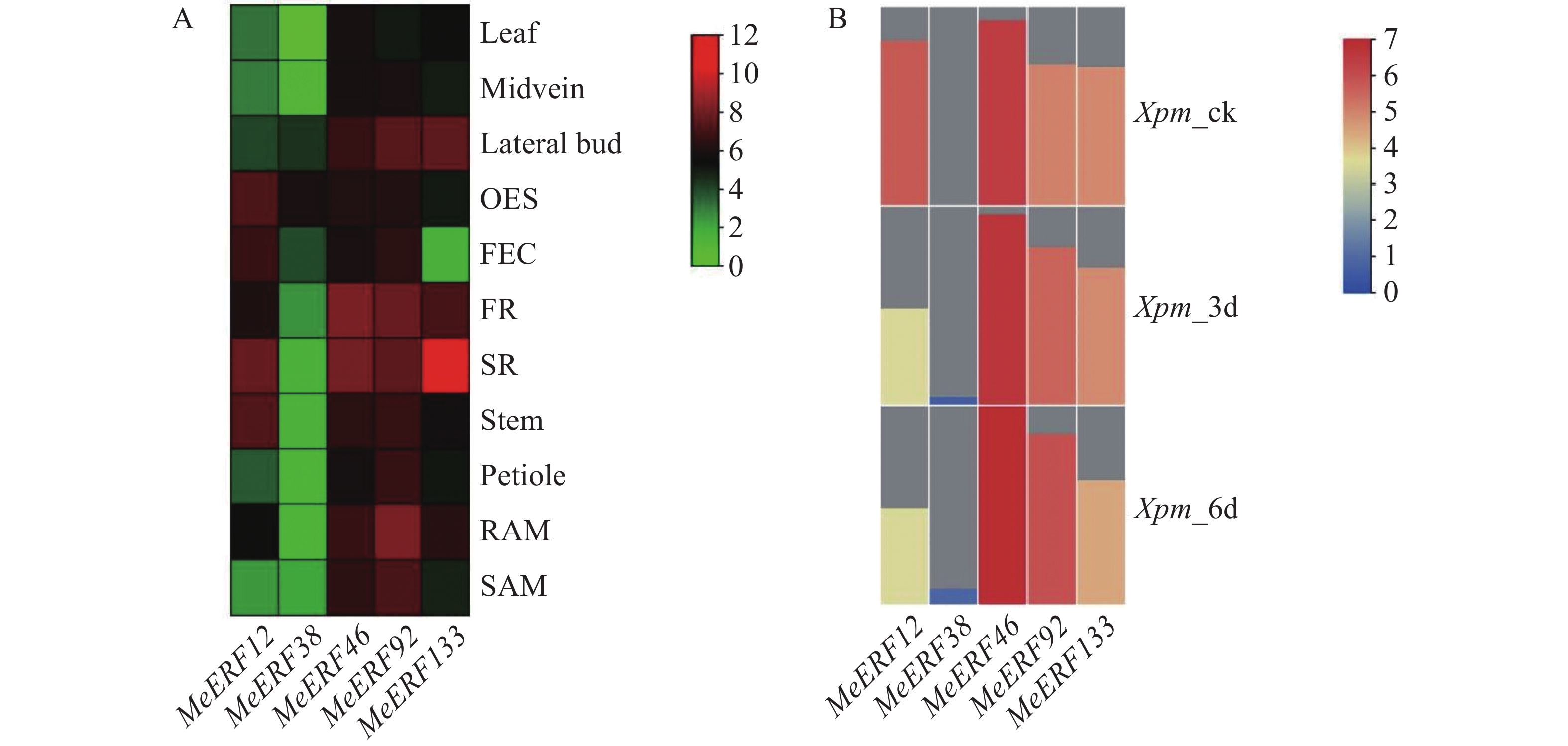
为了分析MeERFs在木薯不同组织的表达情况,利用下载于GEO数据库的转录组数据绘制表达热图。表达谱显示,所有基因至少在一种组织中表达,其中MeERF46和MeERF92近乎在所有组织中均有较高表达量,MeERF133在木薯块根中的转录水平呈现高于其余基因在任意组织中的情况(图8-A)。利用本实验室前期获得的木薯受到Xpm侵染后的转录组数据构建基因表达热图,以探究Ⅶ亚族MeERFs基因在抗病中的作用。表达谱显示,除MeERF38基因外,其余基因均有明显表达,MeERF12、MeERF92和MeERF133在病原菌侵染后表达量升高,其中MeERF92表达量变化最显著(图8-B)。MeERF46在病原菌侵染前后的表达量均高于其余基因,而MeERF38的表达量则相反。在5个基因中,MeERF46在不同组织中的表达量较稳定,且相较于亚族内其他成员在叶片中的表达量较高,同时其表达量受Xpm的诱导升高约1倍。基于表达模式分析结果推测,MeERF46可能与木薯正常生长发育及病原菌防御均相关,MeERF38可能积极响应Xpm侵染,MeERF133则主要与块根中的生理功能及响应Xpm的侵染相关。
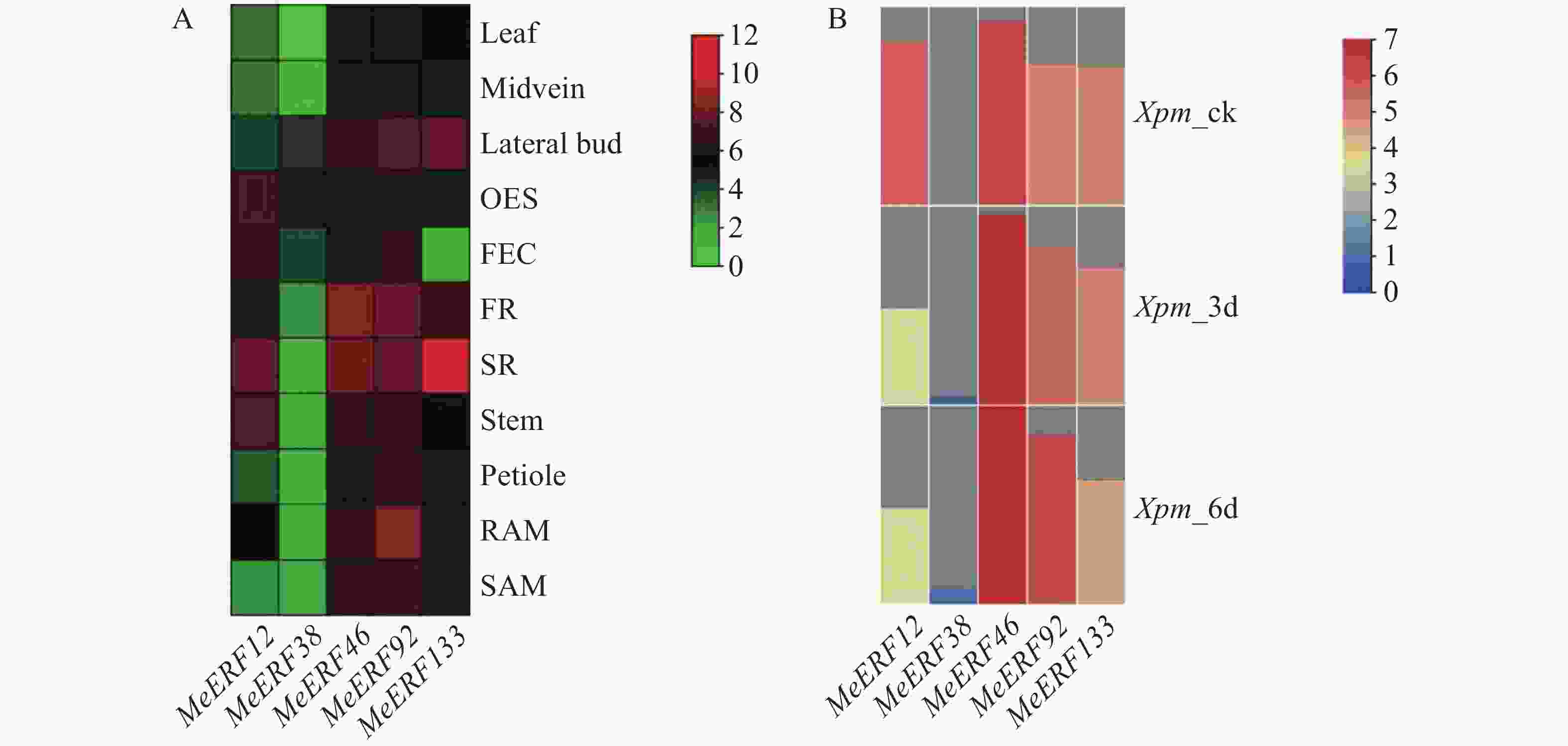
图 8 木薯ERF基因家族Ⅶ亚族基因在木薯不同组织中和受Xpm侵染后的表达模式图
Figure 8. Expression pattern of Ⅶ subfamily genes of cassava ERF gene family in different tissues of cassava and after Xpm infection
-
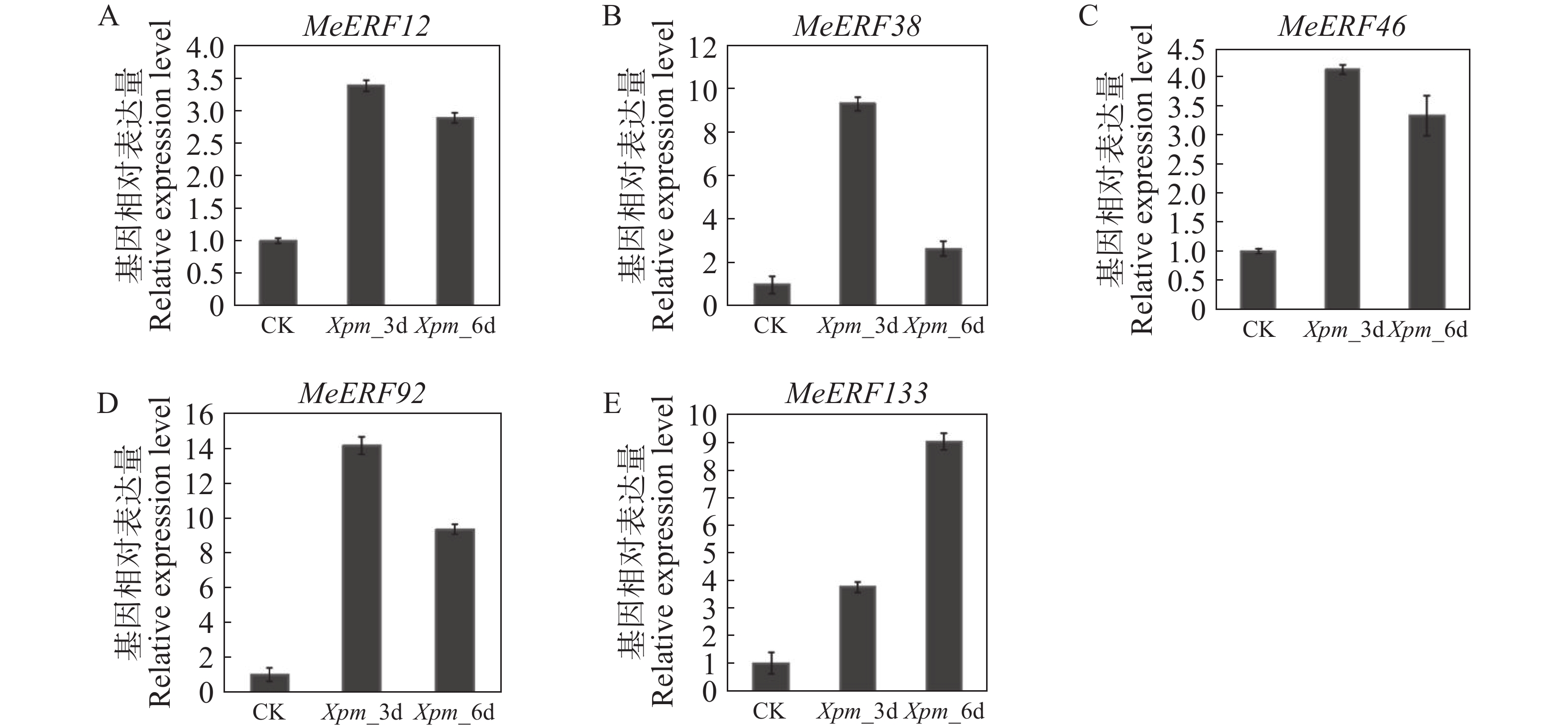
为了进一步验证木薯ERF基因家族Ⅶ亚族成员在木薯遭受病原菌侵染时的转录组水平变化,对木薯ERF基因家族Ⅶ亚族进行定量分析。除了MeERF133,木薯ERF基因家族Ⅶ亚族所有成员基因的表达量均在Xpm侵染3 d后显著上调,随后在Xpm侵染6 d后下调(图9)。在Xpm侵染第3天时,MeERF92的表达量变化最显著,其次是MeERF38。然而,MeERF38的表达量在Xpm侵染第6天后迅速下降。MeERF133的表达量呈现逐渐升高的趋势,并且在Xpm侵染第6天达到顶峰,推测其表达量有可能继续上升。
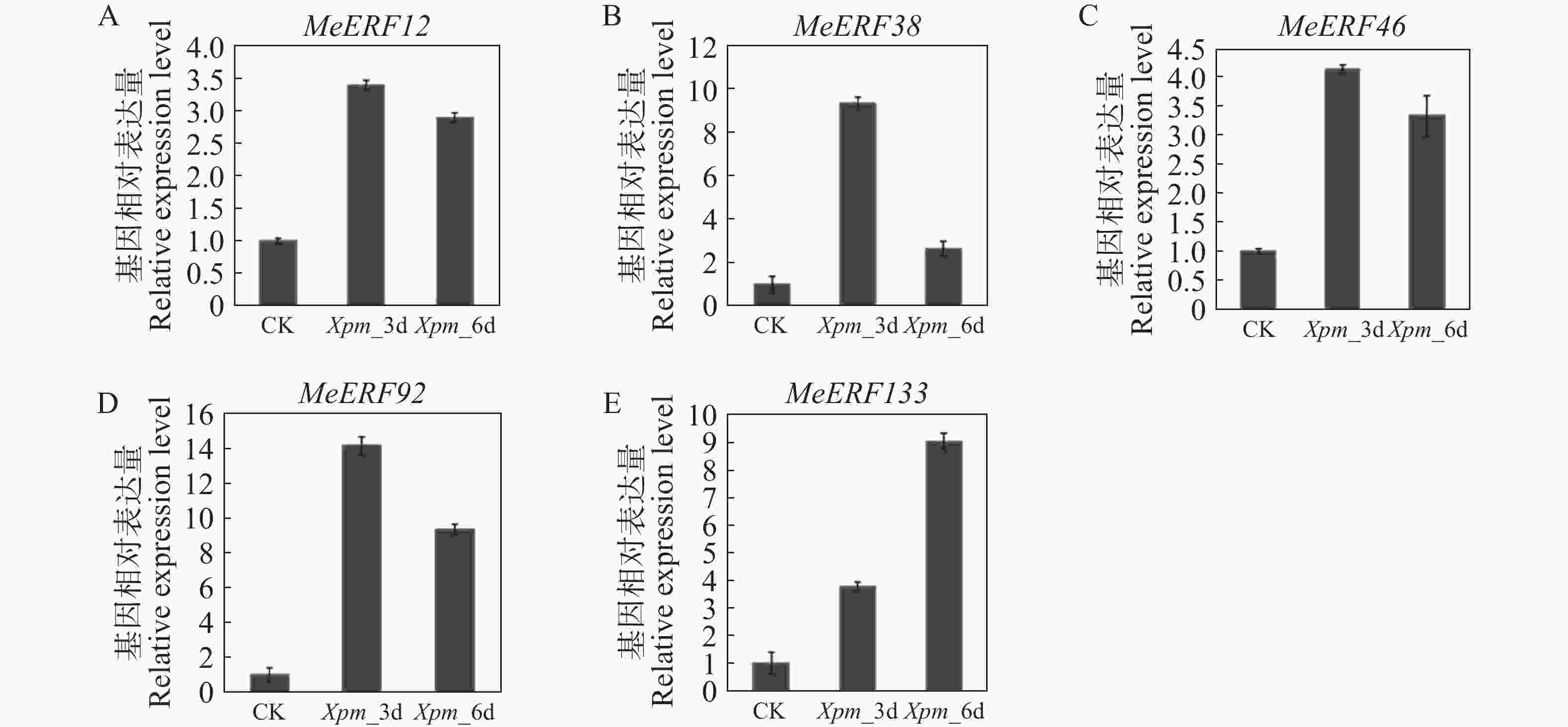
图 9 木薯ERF基因家族Ⅶ亚族基因在Xpm处理下的表达模式
Figure 9. Expression patterns of Ⅶ subfamily genes of cassava ERF gene family after Xpm infection
-
在自然界中,植物进化出一系列应对生物和非生物胁迫的功能,其中转录因子是植物抗逆的关键[20]。转录因子可以特异结合多种基因启动子区的顺式元件,从而调控植物的生长发育和逆境应答[21]。属于AP2/ERF超家族的ERF转录因子在植物生长发育和多种胁迫应答中发挥重要作用[22]。在不同植物中,ERF不仅参与许多信号传导过程,还参与响应病原菌侵染[23]。已有研究表明,过表达OsERFBP1水稻植株不仅可以提升对水稻黄单胞菌水稻致病变种(Xanthomonas oryzae pv. Oryzae,Xoo)的抗性,还可以提升水稻抗旱耐涝的能力[24];玉米ZmERF105可以积极响应玉米大斑病菌(Exserohilum turcicum)的侵染[25]。拟南芥AT4g13040和拟南芥AtERF71分别正向调控其对丁香假单胞菌和禾谷镰刀菌的抗性,其中AtERF71属于第Ⅶ亚族[26-27]。月季RcERF099正向调控其对葡萄孢菌的抗性[28]。TFIIE和ASHR1分别是RNA聚合酶Ⅰ/Ⅱ的转录因子(transcription factor IE/IIE)和组蛋白赖氨酸甲基化转移酶(histone-lysine N-methyltransferase)。鉴定于木薯基因组v6.1中的MeERF10对MeTFIIE和MeASHR1均有正调控作用;MeERF137对MeTFIE呈负调控作用,但对MeASHR1呈正调控作用;MeERF58仅对MeTFIIE有正调控作用;而MeERF35仅负调控MeASHR1,推测这4种木薯ERFs参与抗木薯细菌性枯萎病的应答[29]。
本研究在木薯基因组v8.1中挖掘了161个木薯ERF基因家族成员,并分析其在抗病中的潜在功能。161个MeERFs被分为13个亚族,其中第Ⅶ亚族的作用与抗病、盐胁迫和耐寒相关[30]。木薯ERF基因家族Ⅶ亚族成员均包含motif2、motif6、motif8和motif10,以及包含了AP2结构域的motif1,说明该亚族蛋白高度保守且具有相似生物学功能。在植物不同发育阶段,不同激素能诱导植物自身产生应对生物及非生物胁迫的信号[31],其中茉莉酸甲酯(methyl jasmonate,MeJA)可以增强植物的抗病能力[32-33]。木薯ERF基因家族Ⅶ亚族成员的启动子区域含有参与植物的生长发育、光响应、激素响应和环境胁迫响应的顺式作用元件。除MeERF12外,所有MeERFs启动子区均含有赤霉素响应元件,推测Ⅶ亚族大部分成员均能响应水淹胁迫,该结果与研究团队鉴定出的深水水稻的ERF基因家族SK1(SNORKEL1)和SK2(SNORKEL2)基因所编码的ERF转录因子作用一致[34]。ERF能够通过水杨酸激素的信号传导响应生物胁迫[35],推测2个MeERFs(MeERF38和MeERF133)可能通过水杨酸激素的信号传导途径以响应病原菌的侵染。MeJA正调控柑橘对轮斑病菌的抗病性,且外源MeJA能够进一步增强柑橘的抗病性[36]。在激素响应元件中,MeJA响应元件数量最多(54%),推测该亚族成员可能主要通过MeJA途径正调控木薯抗病性。
互作蛋白和靶基因的预测以及共表达网络分析有助于解析木薯ERF的抗病通路。AHB1蛋白具有氧有高亲和性,且在响应生物胁迫过程中发挥作用[37]。ACBP2蛋白属于酰基辅酶A结合蛋白,是一类脂类载体蛋白,在植物调节生长发育和响应非生物及生物胁迫过程中发挥重要作用[38-39]。属于同源异形盒蛋白家族的ZHD1-2,也称ZF-HD蛋白,该类蛋白在植物生长发育和响应多种生物和非生物胁迫过程中发挥作用[40]。依据蛋白互作的结果推测,4个MeERF(MeERF46、MeERF92、MeERF12和MeERF133)的互作蛋白可能是AHB1和ACBP2,MeERF38的互作蛋白则可能是ZHD1-2。Zhou等[41]通过实验验证表明,属于同源异形盒蛋白家族的ACBP2蛋白与拟南芥ERF-Ⅶ有直接相互作用,其互作能增强拟南芥介导缺氧信号的传导从而响应水淹胁迫。根据共表达分析结果,MeERF46属于MEturquoise模块,是MeERFs与Xpm正响应相关的基因,且在受到Xpm侵染6 h时相关性最高。以上结果表明,木薯ERF基因家族Ⅶ亚族基因可以直接或间接地调控下游基因,参与木薯响应Xpm侵染。
木薯ERF基因家族Ⅶ亚族成员在不同组织中表达差异明显,2个基因在Xpm病原菌侵染时表达量上调。不同组织的表达谱显示,MeERF46基因在叶片中的表达量最高,MeERF133和MeERF12分别在块根和茎中的表达量最高,其分别是其余基因在叶片中表达量的60倍,在块根中表达量的660倍和在茎中表达量的160倍。MeERF46、MeERF92和MeERF133基因的表达量显著受到Xpm的诱导,其转录水平除MeERF92上升2倍外,MeERF46和MeERF133分别上升和下降约1倍。在5个木薯ERF基因家族Ⅶ亚族成员中,MeERF133基因在块根中的表达量较高,且受到Xpm的侵染后,其表达量在第6天提高9倍,推测该基因主要在木薯块根中发挥抗病功能。有趣的是,5个木薯ERFs的qPCR结果与转录组的表达谱不完全一致,推测不同品种中的该亚族基因对病原菌的响应程度稍有区别。然而,MeERF133基因的表达可能在不同品种中均能受到Xpm的显著诱导。
-
本研究在木薯基因组v8.1中挖掘获得了161个木薯ERF基因家族成员,并将其分为13个亚族。本研究采用生物信息学分析方法对Ⅶ亚族成员进行了保守结构域、基因结构及保守基序分析、启动子元件分析、互作蛋白和靶基因预测,以及表达模式分析,以推测该亚族成员可能在抗Xpm侵染中的作用。研究发现,该亚族成员在结构上高度保守,且含有多种参与植物生长发育、光响应、激素响应和环境胁迫响应的顺式作用元件;AHB1、ACBP2和ZHD1-2可能是该亚族成员的互作蛋白;MeERF46在Xpm侵染后6 h正响应的相关性最高。该亚族成员在木薯不同组织中的表达丰度和受到Xpm侵染后表达量存在差异,MeERF46、MeERF133和MeERF12分别在叶片、块根、茎组织中高度表达,MeERF92表达量显著受到Xpm的诱导。实时荧光定量的结果进一步证明,MeERF92和MeERF133与木薯抗病性正相关,MeERF92的表达量在第3天上升了6倍,而MeERF133的表达量在第6天上升了9倍。本研究表明,该亚族成员在木薯的疾病防御机制中具有潜在作用,且MeERF133、MeERF92和MeERF46与木薯抵抗Xpm的侵染具有十分重要的意义,该结果可为进一步研究木薯的抗病机制和抗病育种提供理论依据。
木薯ERF基因家族Ⅶ亚族的鉴定与表达分析
DOI: 10.15886/j.cnki.rdswxb.20250035
 CSTR: 32425.14.j.cnki.rdswxb.20250035
CSTR: 32425.14.j.cnki.rdswxb.20250035
Bioinformatics analysis and functional study of cassava ERF gene family Ⅶ subfamily
-
摘要: 为了探究木薯(Manihot esculenta)ERF基因家族Ⅶ亚族在木薯应对生物胁迫中的潜在功能,本研究对该家族进行了全基因组扫描,在木薯基因组中鉴定了161个ERF基因家族成员,将其分为13个亚族。对Ⅶ亚族成员进行保守结构域、启动子顺式作用元件、互作蛋白、靶基因及表达模式分析。结果表明,该亚族成员均含有一个保守的AP2结构域,其启动子区域包含13种与植物生长发育、环境胁迫响应相关的顺式作用元件。表达模式分析发现,MeERF46、MeERF133、MeERF92基因均积极响应Xpm侵染,其中MeERF92表达量变化最显著;WGCNA和蛋白互作网络分析显示MeERF92可能参与了氧化应激过程,而MeERF133广泛参与了糖酵解过程。以上结果为深入研究ERF在木薯应答生物胁迫中的功能和作用机制提供了候选基因。Abstract: In order to investigate the potential functions of the cassava (Manihot esculenta) Ⅶ subfamily of ERF gene family in response to biotic stress, we identified 161 ERF genes in the cassava genome, which were divided into 13 subfamilies. Analyses were performed on the conserved domains, cis-acting elements in promoter regions, interacting proteins, target genes, and expression patterns of the Ⅶ subfamily members. The results showed that all members of this subfamily contained a conserved AP2 domain, and their promoter regions included 13 types of cis-acting elements related to plant growth and development, and environmental stress responses. Expression pattern analysis revealed that the MeERF46, MeERF133, and MeERF92 genes responded actively to Xpm infection among them, the change of MeERF92 expression was the most significant. WGCNA and protein-protein interaction network analysis indicated that MeERF92 might be involved in the process of oxidative stress, while MeERF133 was widely involved indisease resistance related processes. These findings provide candidate genes for further research into the functions and mechanisms of ERF in cassava's response to biotic stress.
-
Key words:
- Cassava /
- ERF gene family /
- bioinformatics analysis /
- Expressionanalysis
-
图 2 木薯ERF基因家族Ⅶ亚族基因结构
Fig. 2 Gene structure of Ⅶ subfamily genes in cassava ERF gene family
图 3 木薯ERF基因家族Ⅶ亚族保守基序分析
Fig. 3 Conserved motif analysis of Ⅶ subfamily of cassava ERF gene family
图 5 木薯ERF基因家族Ⅶ亚族启动子顺式作用元件分析
Fig. 5 Analysis of cis acting elements of the Ⅶ subfamily promoter of cassava ERF gene family
图 6 木薯ERF基因家族Ⅶ亚族的互作蛋白网络图
Fig. 6 Protein-protein interaction of cassava ERF Ⅶ subfamily
图 7 木薯ERF基因家族Ⅶ亚族基因共表达网络分析
Fig. 7 Co-expression network of Ⅶ subfamily genes in cassava ERF gene family
图 8 木薯ERF基因家族Ⅶ亚族基因在木薯不同组织中和受Xpm侵染后的表达模式图
Fig. 8 Expression pattern of Ⅶ subfamily genes of cassava ERF gene family in different tissues of cassava and after Xpm infection
图 9 木薯ERF基因家族Ⅶ亚族基因在Xpm处理下的表达模式
Fig. 9 Expression patterns of Ⅶ subfamily genes of cassava ERF gene family after Xpm infection
表 1 木薯ERF转录因子家族信息
Table 1 Information on cassava ERF transcription factor family
重命名
Rename基因编号
Gen ID蛋白长度/aa
Protein length相对分子质量
Mw等电点
Theoretical pIMeERF1 Manes.01G000062.1 338 35665.73 5.18 MeERF2 Manes.01G012200.6 308 35086.46 5.21 MeERF3 Manes.01G022500.1 220 24351.21 9.49 MeERF4 Manes.01G046200.1 337 37280.05 4.93 MeERF5 Manes.01G067400.1 243 26965.04 5.82 MeERF6 Manes.01G067500.1 233 26178.39 5.27 MeERF7 Manes.01G085100.1 260 28968.13 7.67 MeERF8 Manes.01G085200.1 315 35275.61 6.56 MeERF9 Manes.01G085400.1 289 30917.88 6.36 MeERF10 Manes.01G085800.1 318 36809.66 9.41 MeERF11 Manes.01G101000.1 402 45128.87 4.59 MeERF12 Manes.01G222600.1 309 34577.41 6.06 MeERF13 Manes.01G245200.1 187 20860.34 5.54 MeERF14 Manes.01G249600.1 310 35421.18 4.97 MeERF15 Manes.01G262100.2 283 31426.76 9.07 MeERF16 Manes.01G271300.1 251 27577.35 5.45 MeERF17 Manes.02G010700.1 308 34347.42 8.80 MeERF18 Manes.02G013200.1 335 37300.15 5.26 MeERF19 Manes.02G154400.1 355 39070.65 6.21 MeERF20 Manes.02G161700.1 232 24812.79 6.11 MeERF21 Manes.02G182000.1 458 48601.51 6.39 MeERF22 Manes.02G208000.2 176 19697.98 5.43 MeERF23 Manes.02G208500.1 174 19131.18 5.74 MeERF24 Manes.03G011000.1 201 22830.86 8.69 MeERF25 Manes.03G015400.1 189 21225.84 6.97 MeERF26 Manes.03G025200.1 279 30468.31 5.30 MeERF27 Manes.03G044100.1 206 23073.70 4.94 MeERF28 Manes.03G056100.1 209 23281.35 4.81 MeERF29 Manes.03G058500.1 269 29083.08 4.84 MeERF30 Manes.03G090396.1 374 43206.81 9.13 MeERF31 Manes.03G098000.1 288 32246.12 8.14 MeERF32 Manes.03G106300.1 228 24636.33 9.47 MeERF33 Manes.03G147500.1 332 37682.17 7.25 MeERF34 Manes.03G165100.1 219 25037.03 5.61 MeERF35 Manes.03G165200.1 150 17202.76 6.12 MeERF36 Manes.03G165300.1 140 15483.99 6.31 MeERF37 Manes.03G192400.1 331 37005.58 5.26 MeERF38 Manes.03G199300.1 259 28942.74 6.54 MeERF39 Manes.04G027000.8 504 55441.53 7.13 MeERF40 Manes.04G094600.1 265 29036.53 4.79 MeERF41 Manes.04G100700.1 330 37395.26 8.61 MeERF42 Manes.04G105000.1 179 20194.42 7.13 MeERF43 Manes.04G114000.1 178 20154.76 8.94 MeERF44 Manes.04G131900.1 231 25233.03 4.75 MeERF45 Manes.04G144500.1 573 63762.94 6.18 MeERF46 Manes.04G150100.4 381 42281.90 4.75 MeERF47 Manes.05G040000.1 302 33130.72 8.92 MeERF48 Manes.05G052000.1 244 27100.90 5.57 MeERF49 Manes.05G054200.1 222 24764.44 4.80 MeERF50 Manes.05G054300.1 222 24887.60 4.97 MeERF51 Manes.05G054400.1 230 25481.27 5.35 MeERF52 Manes.05G106400.1 368 41238.68 4.77 MeERF53 Manes.05G120878.1 319 36175.14 4.59 MeERF54 Manes.05G153700.1 233 25772.96 5.32 MeERF55 Manes.05G175800.1 152 17157.18 9.12 MeERF56 Manes.05G184000.1 661 72872.84 6.60 MeERF57 Manes.06G032500.1 353 40399.73 6.33 MeERF58 Manes.06G058200.1 211 23455.48 6.97 MeERF59 Manes.06G068900.1 350 38842.80 4.82 MeERF60 Manes.06G071700.1 369 41012.18 9.07 MeERF61 Manes.06G127000.1 247 27672.40 9.24 MeERF62 Manes.06G131100.1 248 27664.05 7.07 MeERF63 Manes.06G131200.1 285 31753.29 9.21 MeERF64 Manes.06G131300.1 212 23612.22 5.67 MeERF65 Manes.06G143102.1 231 25542.74 5.49 MeERF66 Manes.06G143200.1 230 24770.37 5.39 MeERF67 Manes.06G156400.1 372 41907.57 4.70 MeERF68 Manes.07G114500.1 236 25810.98 9.26 MeERF69 Manes.07G131900.1 344 37015.33 6.38 MeERF70 Manes.07G135300.2 388 42958.18 4.94 MeERF71 Manes.08G021600.1 199 21599.19 5.42 MeERF72 Manes.08G022000.1 202 22777.86 6.66 MeERF73 Manes.08G048500.1 294 33611.76 6.02 MeERF74 Manes.08G096900.1 421 47903.26 6.08 MeERF75 Manes.08G099800.1 274 29891.50 4.68 MeERF76 Manes.08G101300.1 250 28280.07 5.81 MeERF77 Manes.08G137700.1 222 25471.20 4.91 MeERF78 Manes.08G145400.1 277 31562.01 5.52 MeERF79 Manes.09G020700.1 427 47250.60 8.53 MeERF80 Manes.09G058852.1 204 21585.89 5.64 MeERF81 Manes.09G080100.1 458 51437.98 6.18 MeERF82 Manes.09G146100.1 200 21356.54 5.12 MeERF83 Manes.09G149000.1 222 25191.10 5.15 MeERF84 Manes.09G173200.1 232 25325.83 4.51 MeERF85 Manes.09G188100.1 231 26127.79 6.30 MeERF86 Manes.10G010300.1 332 35806.27 6.20 MeERF87 Manes.10G031600.1 240 26187.29 8.84 MeERF88 Manes.10G041100.3 478 53902.31 7.10 MeERF89 Manes.10G056300.1 263 29211.72 8.22 MeERF90 Manes.10G063200.1 422 47166.90 5.12 MeERF91 Manes.10G119500.4 513 57285.11 5.95 MeERF92 Manes.11G013700.7 385 42590.45 4.86 MeERF93 Manes.11G020300.1 560 62316.43 6.53 MeERF94 Manes.11G035600.1 236 25593.38 5.11 MeERF95 Manes.11G055000.1 196 21564.58 9.62 MeERF96 Manes.11G064100.1 178 20254.65 6.31 MeERF97 Manes.11G064200.1 178 20096.35 6.17 MeERF98 Manes.11G074500.1 278 30326.10 4.78 MeERF99 Manes.11G076500.1 280 31229.21 6.25 MeERF100 Manes.11G158533.1 217 23760.89 8.46 MeERF101 Manes.11G158600.1 217 23760.89 8.46 MeERF102 Manes.12G002400.1 706 77507.62 6.04 MeERF86 Manes.10G010300.1 332 35806.27 6.2 MeERF87 Manes.10G031600.1 240 26187.29 8.84 MeERF88 Manes.10G041100.3 478 53902.31 7.10 MeERF89 Manes.10G056300.1 263 29211.72 8.22 MeERF90 Manes.10G063200.1 422 47166.90 5.12 MeERF91 Manes.10G119500.4 513 57285.11 5.95 MeERF92 Manes.11G013700.7 385 42590.45 4.86 MeERF93 Manes.11G020300.1 560 62316.43 6.53 MeERF94 Manes.11G035600.1 236 25593.38 5.11 MeERF95 Manes.11G055000.1 196 21564.58 9.62 MeERF96 Manes.11G064100.1 178 20254.65 6.31 MeERF97 Manes.11G064200.1 178 20096.35 6.17 MeERF98 Manes.11G074500.1 278 30326.10 4.78 MeERF99 Manes.11G076500.1 280 31229.21 6.25 MeERF100 Manes.11G158533.1 217 23760.89 8.46 MeERF101 Manes.11G158600.1 217 23760.89 8.46 MeERF102 Manes.12G002400.1 706 77507.62 6.04 MeERF103 Manes.12G022600.3 414 45296.72 6.21 MeERF104 Manes.12G047500.5 232 26223.60 9.94 MeERF105 Manes.12G087500.1 471 52295.81 6.02 MeERF106 Manes.12G106400.1 510 56821.22 6.96 MeERF107 Manes.12G117800.1 677 75540.73 6.42 MeERF108 Manes.12G125500.1 496 54660.26 5.81 MeERF109 Manes.12G136800.1 361 40158.47 6.08 MeERF110 Manes.12G146000.1 349 38119.39 5.89 MeERF111 Manes.13G001800.1 701 76935.96 6.21 MeERF112 Manes.13G023700.1 403 43709.99 6.43 MeERF113 Manes.13G033000.1 117 13330.77 5.39 MeERF114 Manes.13G033100.1 146 16311.89 8.96 MeERF115 Manes.13G049600.3 235 26376.60 9.73 MeERF116 Manes.13G081100.1 316 34021.08 8.95 MeERF117 Manes.13G093300.1 358 39861.37 5.90 MeERF118 Manes.13G100600.1 503 55506.27 5.86 MeERF119 Manes.13G108800.1 632 70853.51 6.27 MeERF120 Manes.13G120400.1 191 21567.66 8.70 MeERF121 Manes.13G120700.1 484 53738.25 8.68 MeERF122 Manes.13G143500.1 215 23535.44 9.30 MeERF123 Manes.13G148300.1 474 52704.53 6.28 MeERF124 Manes.14G000900.2 544 59966.77 6.12 MeERF125 Manes.14G019032.1 345 38561.03 5.07 MeERF126 Manes.14G029500.2 230 24322.77 4.78 MeERF127 Manes.14G029600.1 219 24140.19 5.11 MeERF128 Manes.14G038900.1 236 26378.58 6.00 MeERF129 Manes.14G044700.1 243 26918.67 9.80 MeERF130 Manes.14G098000.1 366 40348.38 9.24 MeERF131 Manes.14G101900.1 358 40001.20 5.08 MeERF132 Manes.14G154900.1 562 61094.18 6.95 MeERF133 Manes.15G009900.1 269 29932.67 6.03 MeERF134 Manes.15G014600.1 315 34961.60 5.28 MeERF135 Manes.15G039500.1 132 14731.29 6.37 MeERF136 Manes.15G039600.1 162 18420.07 7.97 MeERF137 Manes.15G039700.1 210 23781.70 5.77 MeERF138 Manes.15G066800.1 236 25840.91 8.91 MeERF139 Manes.15G066900.1 233 25825.94 9.42 MeERF140 Manes.15G092200.1 142 15906.60 6.31 MeERF141 Manes.15G102800.1 371 42235.59 8.78 MeERF142 Manes.15G108400.1 294 32691.73 8.79 MeERF143 Manes.16G030400.1 162 17410.50 8.07 MeERF144 Manes.16G030600.1 252 27628.79 9.24 MeERF145 Manes.16G075800.1 264 28999.09 5.13 MeERF146 Manes.16G090900.1 159 17690.76 9.20 MeERF147 Manes.16G121500.1 192 21619.25 6.59 MeERF148 Manes.16G126900.1 203 22932.96 8.99 MeERF149 Manes.17G048200.1 161 17432.51 9.62 MeERF150 Manes.17G048400.1 248 27075.57 9.47 MeERF151 Manes.17G051600.1 186 20613.92 9.29 MeERF152 Manes.17G051700.1 280 31615.20 6.34 MeERF153 Manes.18G040400.1 152 17222.06 8.55 MeERF154 Manes.18G050000.1 683 75301.40 6.23 MeERF155 Manes.18G075600.1 222 23871.83 5.45 MeERF156 Manes.18G093800.1 410 43467.85 5.91 MeERF157 Manes.18G097200.1 150 17004.09 6.73 MeERF158 Manes.18G118800.1 192 21220.90 4.85 MeERF159 Manes.18G118900.3 193 21114.49 5.41 MeERF160 Manes.18G119500.1 178 19721.91 7.01 MeERF161 Manes.18G125700.1 267 30424.46 6.27  下载: 导出CSV
下载: 导出CSV
表 2 木薯ERF基因家族Ⅶ亚族保守基序
Table 2 Conserved motifs of cassava ERF Ⅶ subfamily genes
保守基序 Motif 基序氨基酸序列 Sequence Motif1 IYRGIRQRPWGKWAAEIRDPRKGVRVWLGTFNTAEEAARAYDEEARRIRG Motif2 MCGGAIISDFIPVTRARR Motif3 QLGGSSQDGGEPVDLWSFDDLPNHVGGQR Motif4 LMDSFPANGDGANKSIPPCDNVPMFFNSDQGSNSFECSDFGWGEQASKTP Motif5 NQMNFQMPYLEGSWE Motif6 LDTYNKFLGLDYLBNABPDYFSTQGSL Motif7 FQEFKDESDVDEEDDVLFDVKPFAFSATASPPPRNRSPSRGSTAVKSVEF Motif8 KKAKVNFPDE Motif9 EIDESLFMEDANPKKKIKADSENAVPIEENNGKSPSEELLAF Motif10 RLTADFLAPPLKKPIGVGY
下载: 导出CSV
-
[1] Díaz T P A, Herrera C M, Ochoa C J C, et al. The overexpression of RXam1, a cassava gene coding for an RLK confers disease resistance to Xanthomonas axonopodis pv. manihotis [J]. Planta, 2018, 247(4): 1031−1042. https://doi.org/10.1007/s00425-018-2863-4 doi: 10.1007/s00425-018-2863-4 [2] López C E, Bernal A J. Cassava bacterial blight: using genomics for the elucidation and management of an old problem [J]. Tropical Plant Biology, 2012, 5(1): 117−126. https://doi.org/10.1007/s12042-011-9092-3 doi: 10.1007/s12042-011-9092-3 [3] Brown R L, Kazan K, Mcgrath K C, et al. A role for the GCC-box in jasmonate-mediated activation of the PDF1.2 gene of Arabidopsis [J]. Plant Physiology, 2003, 132(2): 1020−1032. https://doi.org/10.1104/pp.102.017814 doi: 10.1104/pp.102.017814 [4] Fujimoto S Y, Ohta M, Usui A, et al. Arabidopsis ethylene-responsive element binding factors act as transcriptional activators or repressors of GCC box-mediated gene expression [J]. The Plant Cell, 2000, 12(3): 393−404. [5] 葛宝宇, 林轶, 侯和胜. ERF类转录因子的结构与功能[J]. 安徽农学通报, 2007, 13(20): 32−35. https://doi.org/10.3969/j.issn.1007-7731.2007.20.013 doi: 10.3969/j.issn.1007-7731.2007.20.013 [6] Feng K, Hou X L, Xing G M, et al. Advances in AP2/ERF super-family transcription factors in plant [J]. Critical Reviews in Biotechnology, 2020, 40(6): 750−776. https://doi.org/10.1080/07388551.2020.1768509 doi: 10.1080/07388551.2020.1768509 [7] Rashid M, He G, Yang G, et al. AP2/ERF transcription factor in rice: genome-wide canvas and syntenic relationships between monocots and eudicots [J]. Evolutionary Bioinformatics Online, 2012, 8: 321−355. [8] 黄奕孜, 钱旺, 邱姗, 等. 光皮桦AP2/ERF基因家族鉴定与表达分析[J]. 浙江农林大学学报, 2022, 39(6): 1183−1193. https://doi.org/10.11833/j.issn.2095-0756.20220331 doi: 10.11833/j.issn.2095-0756.20220331 [9] 洪雨慧. 木薯ERF转录因子家族全基因组分析[D]. 海口: 海南大学, 2018. [10] Sakuma Y, Liu Q, Dubouzet J G, et al. DNA-binding specificity of the ERF/AP2 domain of Arabidopsis DREBs, transcription factors involved in dehydration- and cold-inducible gene expression [J]. Biochemical and Biophysical Research Communications, 2002, 290(3): 998−1009. https://doi.org/10.1006/bbrc.2001.6299 doi: 10.1006/bbrc.2001.6299 [11] Zhu Z, Shi J, Xu W, et al. Three ERF transcription factors from Chinese wild grapevine Vitis pseudoreticulata participate in different biotic and abiotic stress-responsive pathways [J]. Journal of Plant Physiology, 2013, 170(10): 923−933. https://doi.org/10.1016/j.jplph.2013.01.017 doi: 10.1016/j.jplph.2013.01.017 [12] 吴红漫, 林世扬, 颜彦, 等. 木薯ERF转录因子基因MeERF5克隆及表达分析[J]. 南方农业学报, 2022, 53(9): 2510−2517. https://doi.org/10.3969/j.issn.2095-1191.2022.09.013 doi: 10.3969/j.issn.2095-1191.2022.09.013 [13] 张兴龙, 张亚文, 王晓彤, 等. 基因编辑MeERF127提高木薯抗旱和耐盐性[J]. 热带作物学报, 2024, 45(9): 1780−1790. https://doi.org/10.3969/j.issn.1000-2561.2024.09.002 doi: 10.3969/j.issn.1000-2561.2024.09.002 [14] 廖明馨, 李可, 李梦桃, 等. 过表达MeERF72对C3木薯淀粉合成的影响[J]. 分子植物育种, 2024, 22(16): 5335−5341. [15] 罗数, Maliwan Naconsie, 张鹏. 木薯转录因子MeERF109促进淀粉积累并负调控低温耐受[C]// 2019年全国热带作物学术年会论文集. 西安: 中国热带作物学会, 西北农林科技大学, 2019. https://doi.org/10.26914/c.cnkihy.2019.042836 [16] Ma G, Zhu B, Zhang Y, et al. CPK1-mediated ERF72 protein phosphorylation confers improved disease resistance to cassava bacterial blight [J]. Plant Biotechnology Journal, 2023, 21(11): 2166−2168. https://doi.org/10.1111/pbi.14151 doi: 10.1111/pbi.14151 [17] Finn R D, CoggillP, Eberhardt R Y, et al. The pfam protein families database: towards a more sustainable future [J]. Nucleic Acids Research, 2016, 44(D1): D279−D285. https://doi.org/10.1093/nar/gkv1344 doi: 10.1093/nar/gkv1344 [18] Zafar M M, Rehman A, Razzaq A, et al. Genome-wide characterization and expression analysis of Erf gene family in cotton [J]. BMC Plant Biology, 2022, 22(1): 134. https://doi.org/10.1186/s12870-022-03521-z doi: 10.1186/s12870-022-03521-z [19] Lai Z, Wang J, Fu Y, et al. Revealing the role of CCoAOMT1: fine-tuning bHLH transcription factors for optimal anther development [J]. Science China Life Sciences, 2024, 67(3): 565−578. https://doi.org/10.1007/s11427-023-2461-0 doi: 10.1007/s11427-023-2461-0 [20] Hernandez-garcia C M, Finer J J. Identification and validation of promoters and Cis-acting regulatory elements [J]. Plant Science, 2014, 217: 109−119. [21] Shoji T, Yuan L. ERF gene clusters: working together to regulate metabolism [J]. Trends in Plant Science, 2021, 26(1): 23−32. https://doi.org/10.1016/j.tplants.2020.07.015 doi: 10.1016/j.tplants.2020.07.015 [22] Mitsuda N, Ohme-takagi M. Functional analysis of transcription factors in Arabidopsis [J]. Plant & Cell Physiology, 2009, 50(7): 1232−1248. [23] Jofuku K D, Den boer B G, Van montagu M, et al. Control of Arabidopsis flower and seed development by the homeotic gene APETALA [J]. The Plant Cell, 1994, 6(9): 1211−1225. [24] Jisha V, Dampanaboina L, Vadassery J, et al. Overexpression of an AP2/ERF type transcription factor OsEREBP1 confers biotic and abiotic stress tolerance in rice [J]. PLoS One, 2015, 10(6): e0127831. https://doi.org/10.1371/journal.pone.0127831 doi: 10.1371/journal.pone.0127831 [25] Zang Z, Lv Y, Liu S, et al. A novel ERF transcription factor, ZmERF105, positively regulates maize resistance to Exserohilum turcicum [J]. Frontiers in Plant Science, 2020, 11: 850. https://doi.org/10.3389/fpls.2020.00850 doi: 10.3389/fpls.2020.00850 [26] Giri M K, Swain S, Gautam J K, et al. The Arabidopsis thaliana At4g13040 gene, a unique member of the AP2/EREBP family, is a positive regulator for salicylic acid accumulation and basal defense against bacterial pathogens [J]. Journal of Plant Physiology, 2014, 171(10): 860−867. https://doi.org/10.1016/j.jplph.2013.12.015 doi: 10.1016/j.jplph.2013.12.015 [27] Yelli F, Kato T, Nishiuchi T. The possible roles of AtERF71 in the defense response against the Fusarium graminearum [J]. Plant Biotechnology, 2018, 35(3): 187−192. https://doi.org/10.5511/plantbiotechnology.18.0501b doi: 10.5511/plantbiotechnology.18.0501b [28] Li D, Liu X, Shu L, et al. Global analysis of the AP2/ERF gene family in rose (Rosa chinensis) genome unveils the role of RcERF099 in Botrytis resistance [J]. BMC Plant Biology, 2020, 20(1): 533. https://doi.org/10.1186/s12870-020-02740-6 doi: 10.1186/s12870-020-02740-6 [29] Hong Y, Xiao Y, Song N, et al. Identification and characterization of MeERF genes and their targets in pathogen response by cassava (Manihot esculenta) [J]. The Crop Journal, 2021, 9(5): 1145−1153. https://doi.org/10.1016/j.cj.2020.10.017 doi: 10.1016/j.cj.2020.10.017 [30] Nakano T, Suzuki K, Fujimura T, et al. Genome-wide analysis of the ERF gene family in Arabidopsis and rice [J]. Plant Physiology, 2006, 140(2): 411−432. https://doi.org/10.1104/pp.105.073783 doi: 10.1104/pp.105.073783 [31] 蒋科技, 皮妍, 侯嵘, 等. 植物内源茉莉酸类物质的生物合成途径及其生物学意义[J]. 植物学报, 2010, 45(2): 137−148. https://doi.org/10.3969/j.issn.1674-3466.2010.02.001 doi: 10.3969/j.issn.1674-3466.2010.02.001 [32] 杨婧,苏顺雨,赵添琦, 等. 外源茉莉酸对脂氧合酶基因LOX3敲除的粳稻防御响应的影响[J]. 南方农业学报, 2024, 55(2): 397−410. https://doi.org/10.3969/j.issn.2095-1191.2024.02.010 doi: 10.3969/j.issn.2095-1191.2024.02.010 [33] Yang J, Ma Y, Zeng T, et al. Molecular and metabolic insights into the mechanism of exogenous methyl jasmonate in enhancing the postharvest resistance of kiwifruit to Botrytis cinerea [J]. Postharvest Biology and Technology, 2025, 219: 113226. https://doi.org/10.1016/j.postharvbio.2024.113226 doi: 10.1016/j.postharvbio.2024.113226 [34] Hattori Y, Nagai K, Furukawa S, et al. The ethylene response factors SNORKEL1 and SNORKEL2 allow rice to adapt to deep water [J]. Nature, 2009, 460(7258): 1026−1030. https://doi.org/10.1038/nature08258 doi: 10.1038/nature08258 [35] 唐征, 陈思雀, 徐谦, 等. AP2/ERF在青花菜苗期响应黑腐病的功能研究[J]. 园艺学报, 2024: 2523−2539. [36] 刘凤娇. CsLOB1与CsERF027互作并介导CsRAP2.3-CsERF1级联转录调控柑橘对轮斑病菌的抗性[D]. 西南大学, 2024. [37] 尹虹. AtNUDX6/7基因缺失型拟南芥基因芯片表达谱的生物信息学分析[J]. 锦州医科大学学报, 2021, 42(5): 30−35. [38] Hurlock A K, Roston R L, Wang K, et al. Lipid trafficking in plant cells [J]. Traffic, 2014, 15(9): 915−932. https://doi.org/10.1111/tra.12187 doi: 10.1111/tra.12187 [39] 王健强, 韩林林, 李佳, 等. 苹果轮纹病抗性候选基因的表达分析[J]. 分子植物育种, 2020, 18(8): 2484−2489. [40] 龙姣卉. 宁夏小麦近缘属物种的收集鉴定及黑麦ZF-HD基因家族分析[D]. 银川: 宁夏大学, 2023. [41] Zhou Y, Tan W J, Xie L J, et al. Polyunsaturated linolenoyl-CoA modulates ERF-Ⅶ-mediated hypoxia signaling in Arabidopsis [J]. Journal of Integrative Plant Biology, 2020, 62(3): 330−348. -

 点击查看大图
点击查看大图
计量
- 文章访问数: 1398
- HTML全文浏览量: 575
- PDF下载量: 94
- 被引次数: 0
