




-
植物在长期进化过程中形成了复杂的信号传导网络,以应对多变的环境和生理需求。其中,茉莉酸(Jasmonate,JA)作为一种重要的植物激素,在调节植物的生长发育、逆境响应和次生代谢中发挥着关键作用[1]。随着对JA信号通路的深入研究,MYC2基因作为该通路中的核心转录因子,受到了广泛关注。
MYC2属于bHLH(碱性螺旋−环−螺旋)转录因子家族成员,是JA信号传导的关键调控因子[2]。研究表明,MYC2通过识别并结合下游基因启动子中的G-box元件,调控多种与JA响应相关的基因表达,从而影响植物的防御反应、生长发育和次生代谢产物的合成[3]。在植物次生代谢方面,MYC2能够直接激活拟南芥中倍半萜烯合酶基因的转录,从而提高倍半萜烯的合成[4]。在蒲公英中,TaMYC2通过与角鲨烯合酶基因启动子区域的E-box元件结合来激活该基因的表达,从而促进三萜类化合物蒲公英甾醇和蒲公英萜醇的积累[5]。
油茶是山茶科(Theaceae)山茶属(Camellia L.)一类木本油料植物的总称,包括普通油茶(C. oleifera Abel.)、越南油茶(C. drupifera Lour.)等[6-7]。越南油茶是海南油茶的主要栽培种,茶油富含不饱和脂肪酸及丰富的活性成分,具有较高的营养价值[8]。三萜皂苷是油茶的主要活性成分之一, 具有降血糖和降血脂、抗炎、抗病毒、抗肿瘤、保肝和增强免疫等药理活性,广泛的药理作用使其成为了近几年的研究热点[9]。三萜皂苷在植物体内的积累易受环境因素如光照、水分、干旱和外源激素的影响[10]。这些因素通过调节关键酶基因(HMGR、SS等)的表达和代谢中间体的浓度来控制三萜皂苷的合成,当植物遭受外界刺激时,会激活这些酶基因的表达,从而增强次生代谢物的产生以抵御胁迫。
目前,植物次生代谢合成机制的研究已成为提高目标物质产量的主要手段,特别是通过调控转录因子来影响相关基因的表达,进而控制植物次生代谢的积累。然而,关于CdMYC2转录因子在越南油茶中的研究未见报道。本研究基于越南油茶转录组数据,运用基因克隆技术,从越南油茶中分离并克隆CdMYC2基因,并利用生物信息学方法对其理化性质、进化关系和蛋白质结构进行分析。通过RT-qPCR技术,研究CdMYC2基因在越南油茶不同组织和果实不同成熟发育期中的表达模式,为深入解析CdMYC2转录因子对越南油茶三萜皂苷生物合成积累的调控作用奠定了基础。
-
越南油茶样本采集自海南大学儋州校区的油茶育苗圃(19°30'28" N, 109°29'45" E),不同成熟期果实采摘时间分别为8、9和10月。挑选生长状况优良的油茶植株,取其根、茎、叶和花部分作为提取RNA的样本。采用天根多糖多酚RNA提取试剂盒(RT411)对相应样品进行RNA的提取,而剩余的材料在液氮中冷冻,并在−80 °C下储存。提取的样品RNA经Nanodrop Lite核酸测定仪(thermo scientific, USA)检测浓度和质量分数1%的琼脂糖凝胶电泳检测质量,合格后按照MonScript™RTIII All in One Mix with dsDNase试剂盒(MR05101)进行反转录获得第一链cDNA,然后-20℃保存。后续用于CdMYC2基因的扩增和实时荧光定量PCR。
-
根据课题组已有的转录组数据,找到CdMYC2基因相关的序列,并使用primer premier 5软件设计特异性引物(表1)。按照 PrimeSTAR®系列高保真 PCR 酶试剂(R045A)说明书设置反应体系进行扩增目的片段。用1%质量分数琼脂糖凝胶电泳检测扩增产物,并回收符合大小的目的条带,回收产物的连接和转化依照pBM16A Toposmart克隆试剂盒(CL071-01)和大肠杆菌DH5α感受态细胞(DF1001)说明书进行。
表 1 用于基因克隆和RT-qPCR 反应的引物序列
Table 1. Primer sequences for gene cloning and RT-qPCR reaction
引物 Primer 序列(5′—3′)Sequence (5′—3′) CdMYC2-F ATGGATGAATTAATCATATCGTCAT CdMYC2-R CTAACAAGTCTGTCCTACCCC CdMYC2-QF CATCCAACAAAAGATAGGTG CdMYC2-QR CTTCTCTGTGTGTAAAGCCA Actin-F TTGGCATCGTTGAGGGTCT Actin-R CAGTGGGAACACGGAAAGC -
采用open reading frame finder对CdMYC2的全长序列进行开放阅读框分析;通过NCBI-Conserved Domain 预测CdMYC2的保守结构域;采用NCBI(
http://blast.ncbi.nlm.nih.gov )数据库下载不同植物的MYC2氨基酸序列;使用DNAMAN v8.0 软件对CdMYC2和其他物种同源蛋白序列进行比对;采用MEGA11邻接法构建系统进化树,重复次数为1 000;利用ExPASy - ProtParam tool在线分析CdMYC2编码的蛋白质的理化性质;利用SignalP v3.0在线软件预测CdMYC2蛋白信号肽;通过TMHMM v2.0分析CdMYC2的跨膜结构;NetPhos v3.1在线软件预测CdMYC2的磷酸化位点;运用Plant - mPLoc在线网站预测亚细胞定位;SOPMA在线工具预测CdMYC2的蛋白质二级结构;运用SWISS-MODEL在线工具预测CdMYC2的蛋白质三级结构。 -
以越南油茶的不同组织部位和果实不同成熟期的cDNA为模板,扩增引物通过 Primer v5.0 软件设计(表1)。使用PerfectStart® Green qPCR SuperMix试剂盒(AQ601)的两步法进行荧光定量 PCR 反应,按照说明书中的反应体系和程序进行,相对表达量计算采用2−ΔΔCt 法,利用Graphpad Prism v10.1.2绘图。
-
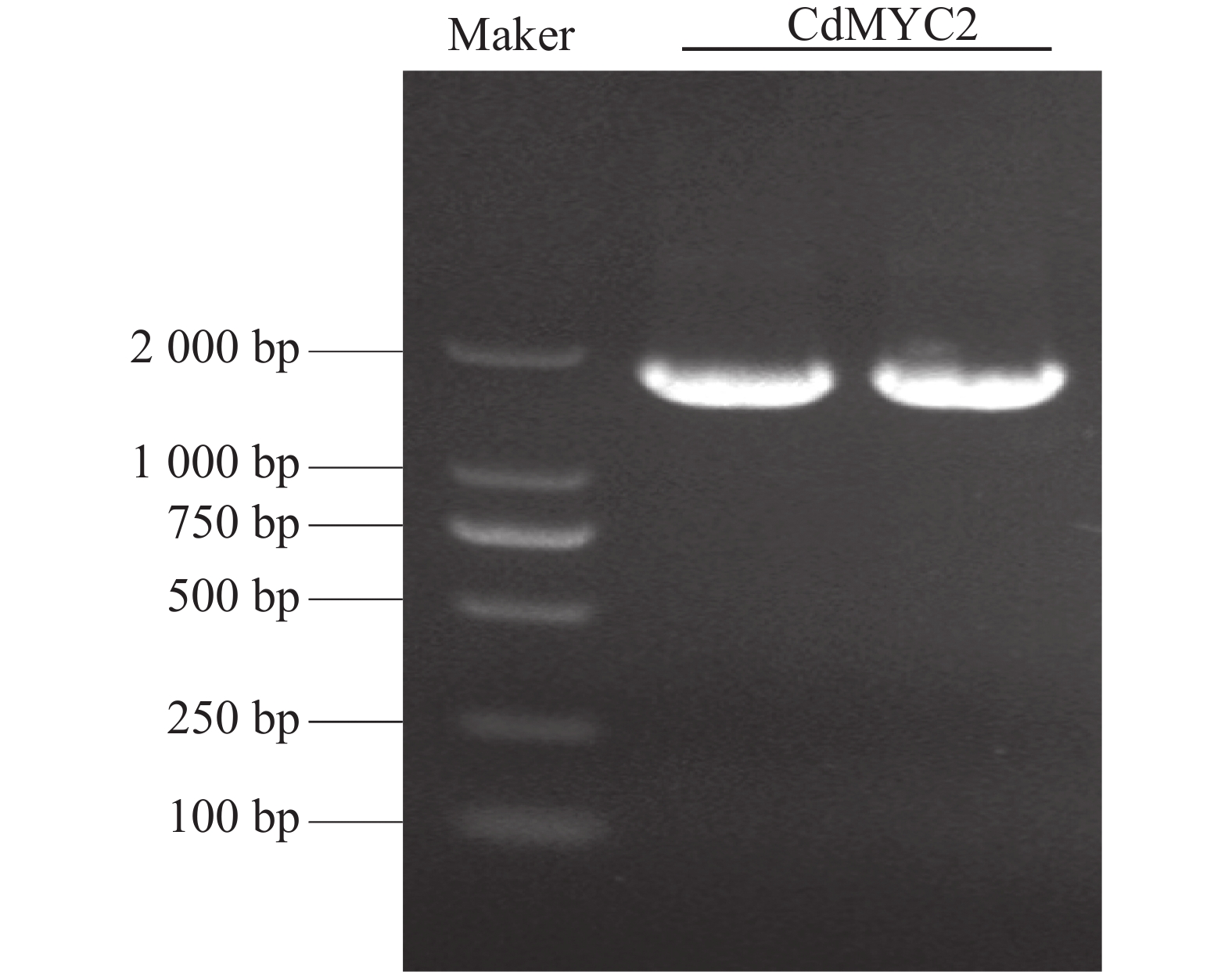
根据课题组的越南油茶转录组数据,设计特异性引物,并以越南油茶叶片cDNA为模板,使用PrimeSTAR®系列高保真PCR 酶扩增CdMYC2基因,获得全长序列长度约为1 850 bp(图1)。测序结果与转录本序列一致。通过ORF finder在线工具对克隆出的CdMYC2的全长序列进行开放阅读框分析,编码序列(CDS)长度为
1 515 bp,编码504个氨基酸(图2)。
图 1 越南油茶CdMYC2基因PCR凝胶电泳图
Figure 1. PCR gel electrophoresis of CdMYC2 gene in C. drupifera

图 2 CdMYC2基因CDS核酸及氨基酸序列
Figure 2. CDS nucleic acid and amino acid sequence of CdMYC2 gene
-
通过NCBI-Conserved Domain 对CdMYC2的氨基酸序列进行保守结构域分析。结果发现,CdMYC2基因具有两个保守结构域家族即bHLH-MYC_N super family (25~208个aa)和bHLH_SF super family (298~371个aa)。bHLH_SF super family是一种碱性螺旋–环–螺旋(bHLH)结构域超家族,包括DNA 结合位点和多肽结合位点(图3)。

图 3 CdMYC2保守结构域分析
Figure 3. Analysis of conserved domains in CdMYC2
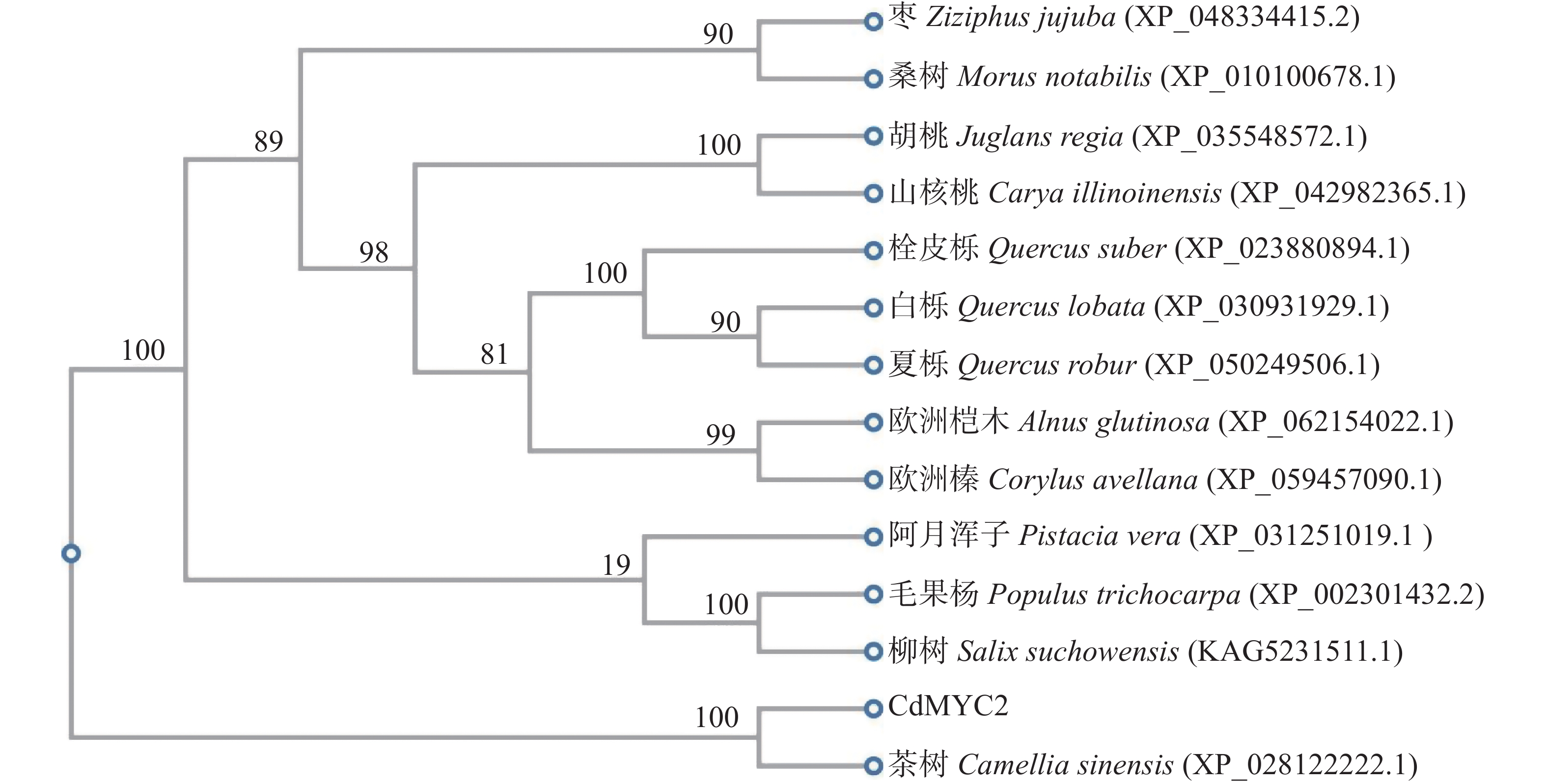
多序列比对和系统发育分析是多物种同源物功能研究的重要方法。将CdMYC2的蛋白序列与从NCBI下载的不同木本植物中的同源蛋白序列进行多序列比对。结果显示,CdMYC2在大多数MYC2中具有高度保守的bHLH结构域(图4)。因此,表明扩增的CdMYC2属于bHLH家族中的MYC2转录因子。系统发育树结果显示,CdMYC2与同科同属植物茶树(C. sinensis)的同源蛋白距离最近;与壳斗科栎属植物白栎(Quercus lobata)、夏栎(Q. robur)的距离最远(图5)。

图 4 CdMYC2与不同物种同源蛋白序列比对
Figure 4. Sequence alignment of CdMYC2 with homologous proteins of different species
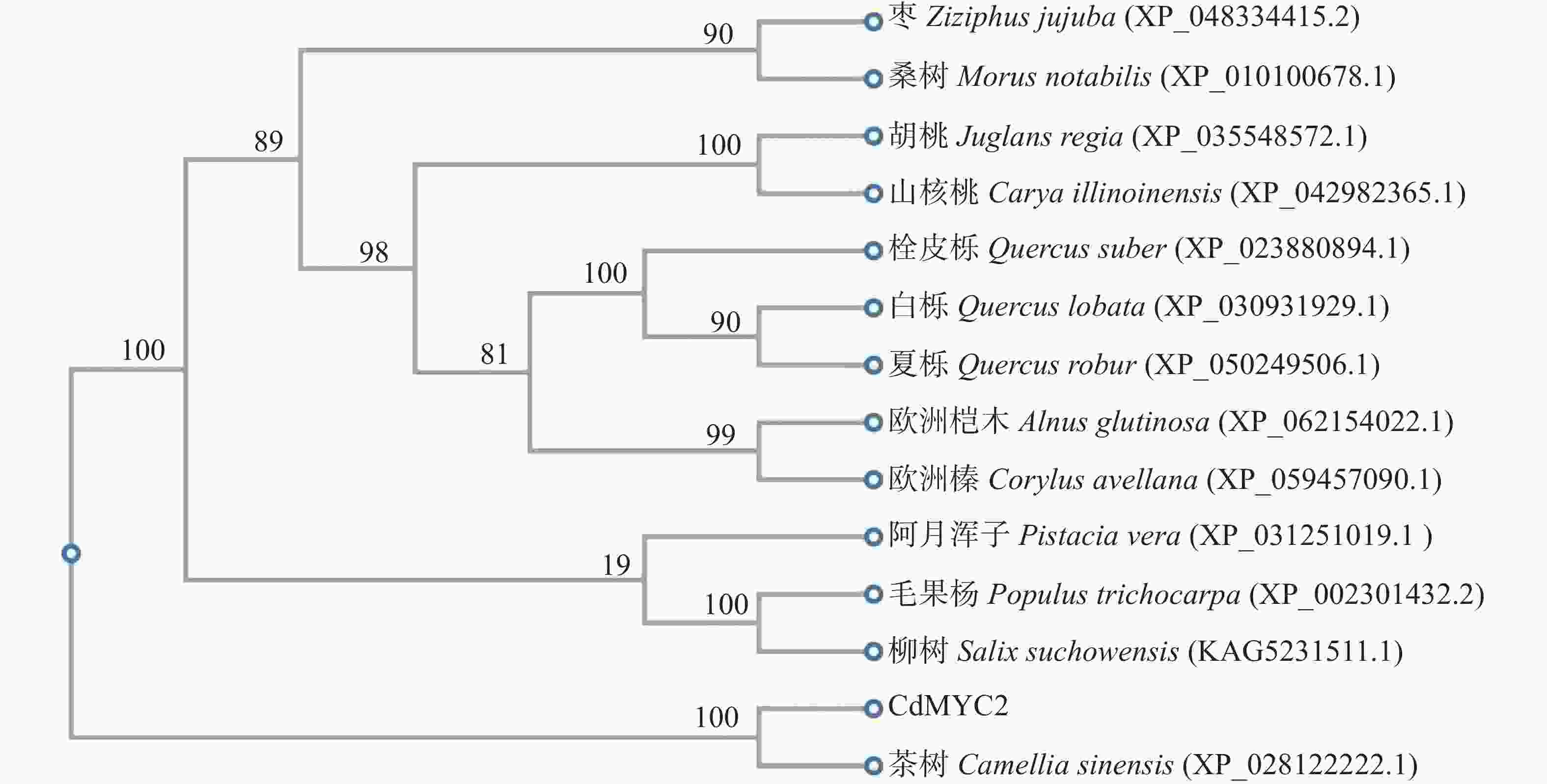
图 5 CdMYC2蛋白的系统发育树分析
Figure 5. Phylogenetic tree analysis of CdMYC2 protein
-
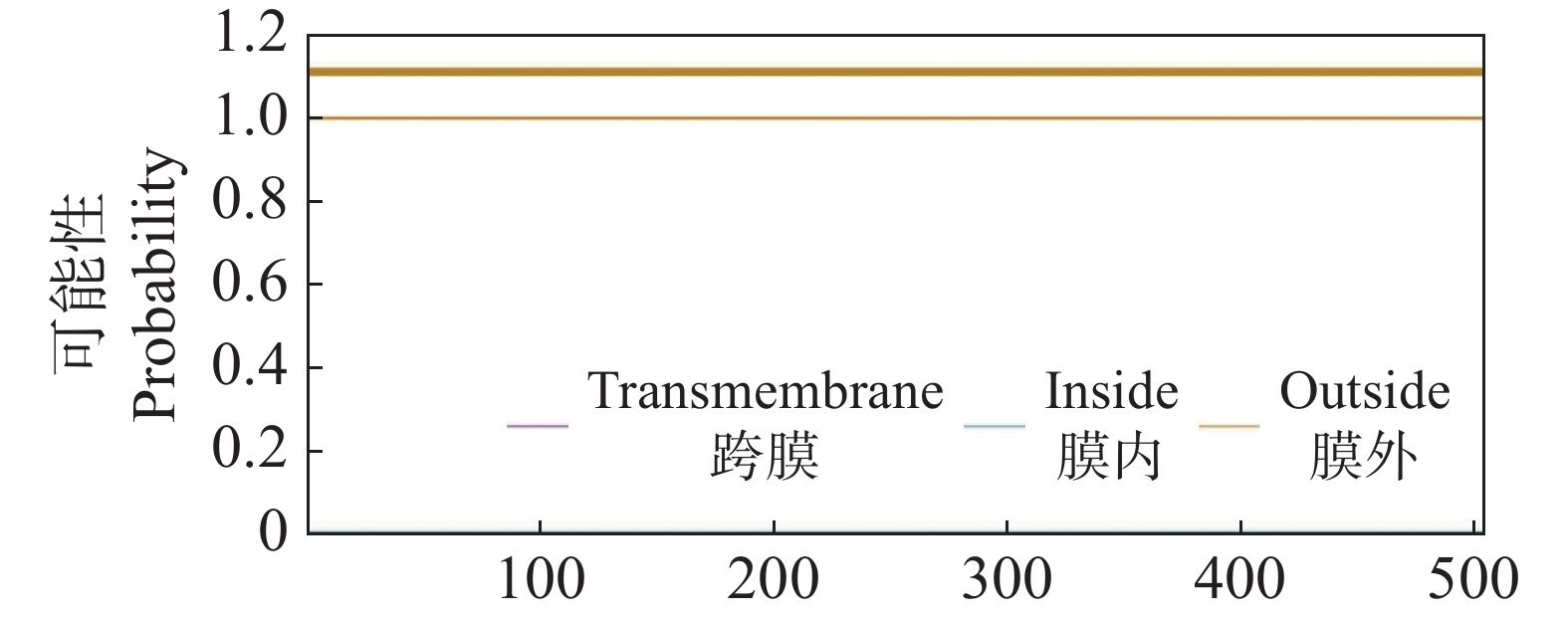
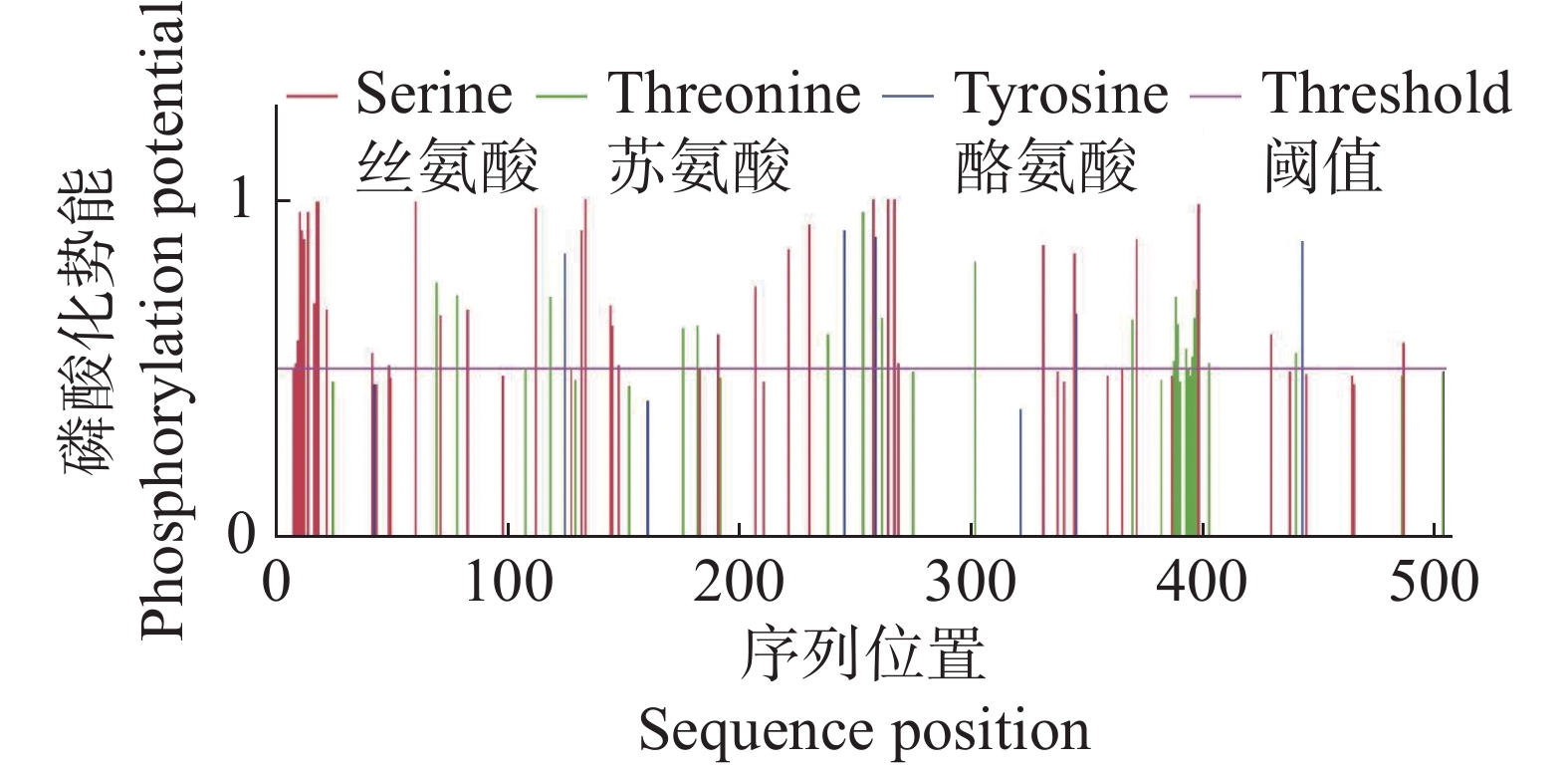
利用SignalP v3.0在线软件预测CdMYC2蛋白信号肽,结果显示CdMYC2蛋白为非分泌蛋白,表明CdMYC2不会通过细胞的分泌途径被释放到细胞外,而是留在细胞内执行其功能。经过TMHMM v2.0的分析,发现CdMYC2不具有跨膜区域(图6)。通过亚细胞定位网站在线预测,结果显示CdMYC2定位在细胞核,与作为转录因子发挥作用位置一致。NetPhos v3.1预测CdMYC2的磷酸化位点,发现总共有60个磷酸化位点,其中丝氨酸磷酸化位点有35个,苏氨酸磷酸化位点有20个,酪氨酸磷酸化位点有5个(图7)。
图 6 CdMYC2蛋白跨膜结构预测
Figure 6. Prediction of transmembrane structure of CdMYC2 protein
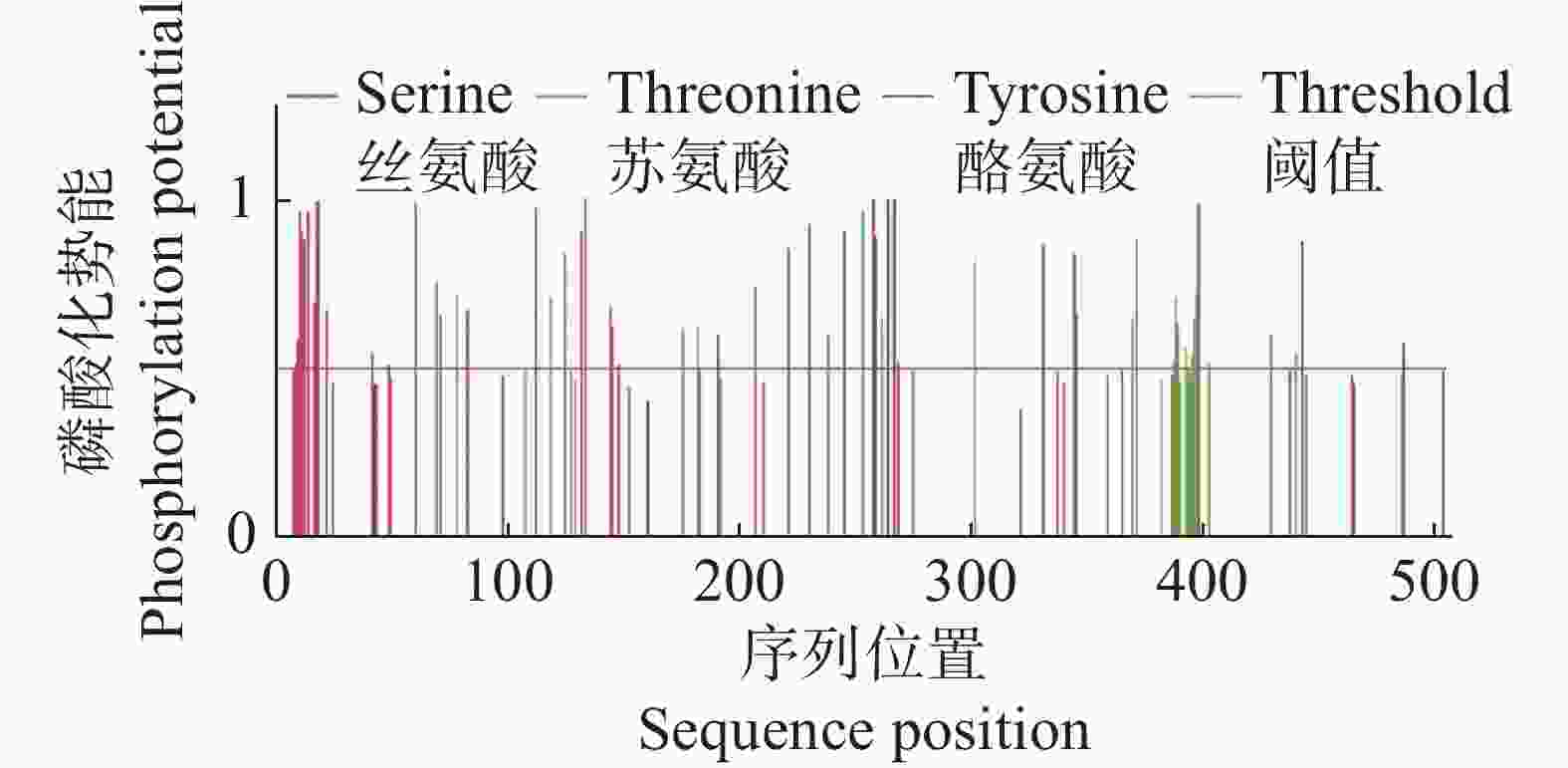
图 7 CdMYC2蛋白磷酸化位点预测结果
Figure 7. Prediction results of CdMYC2 protein phosphorylation sites
-
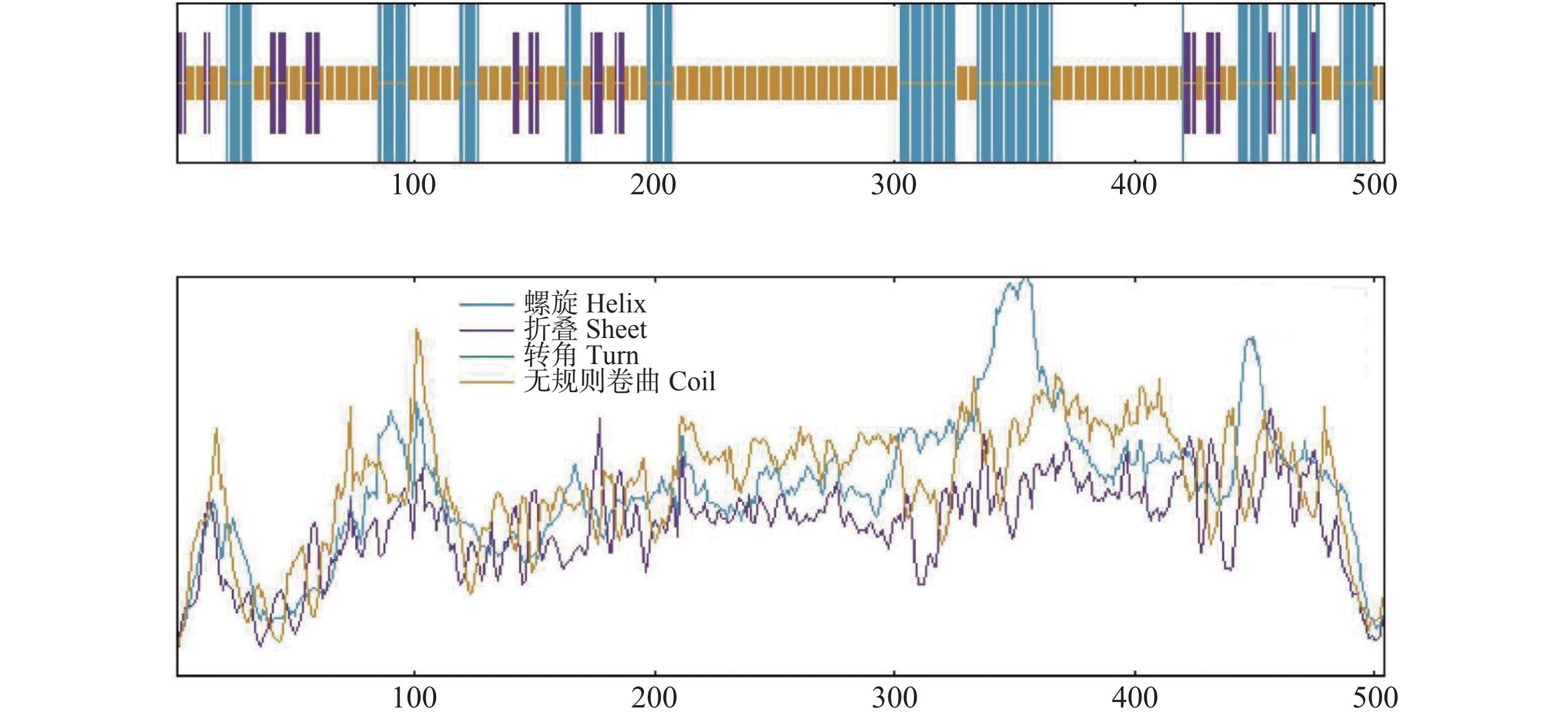
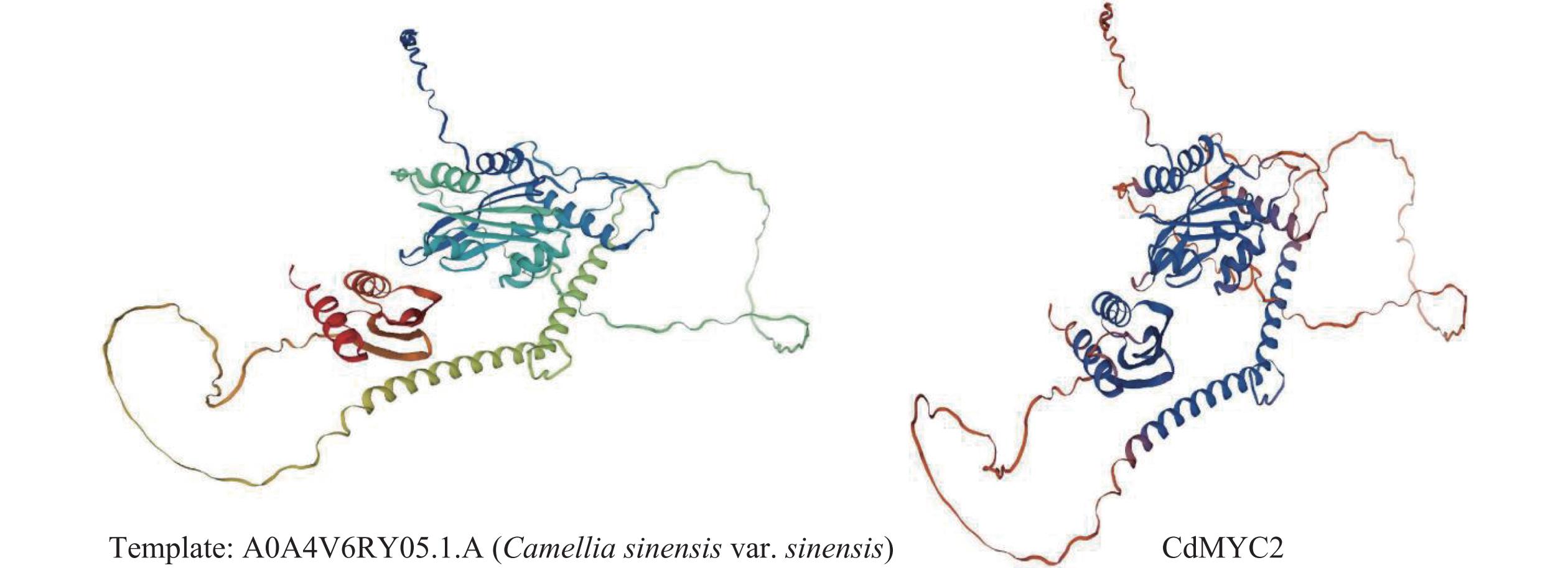
利用SOPMA 在线工具对越南油茶CdMYC2蛋白质的二级结构进行分析,结果其主要由无规则卷曲(random coil)、α−螺旋(alpha helix)与延伸链(extended strand)构成,其中无规则卷曲结构由308个氨基酸残基组成,占比61.11%;α−螺旋结构由146个氨基酸残基组成,占比28.97%;延伸链结构由50个氨基酸残基组成,占比9.92%(图8)。通过SWISS-MODEL在线软件预测CdMYC2蛋白质的三级结构,得到与茶树(C. sinensis)中的bHLH转录因子 (A0A4V6RY05.1.A)结构相似度(98.41%)较高的蛋白质三级结构模型,该结果与进化树分析结果一致,这一高相似度表明两者在结构层面上可能存在紧密的功能关联(图9)。
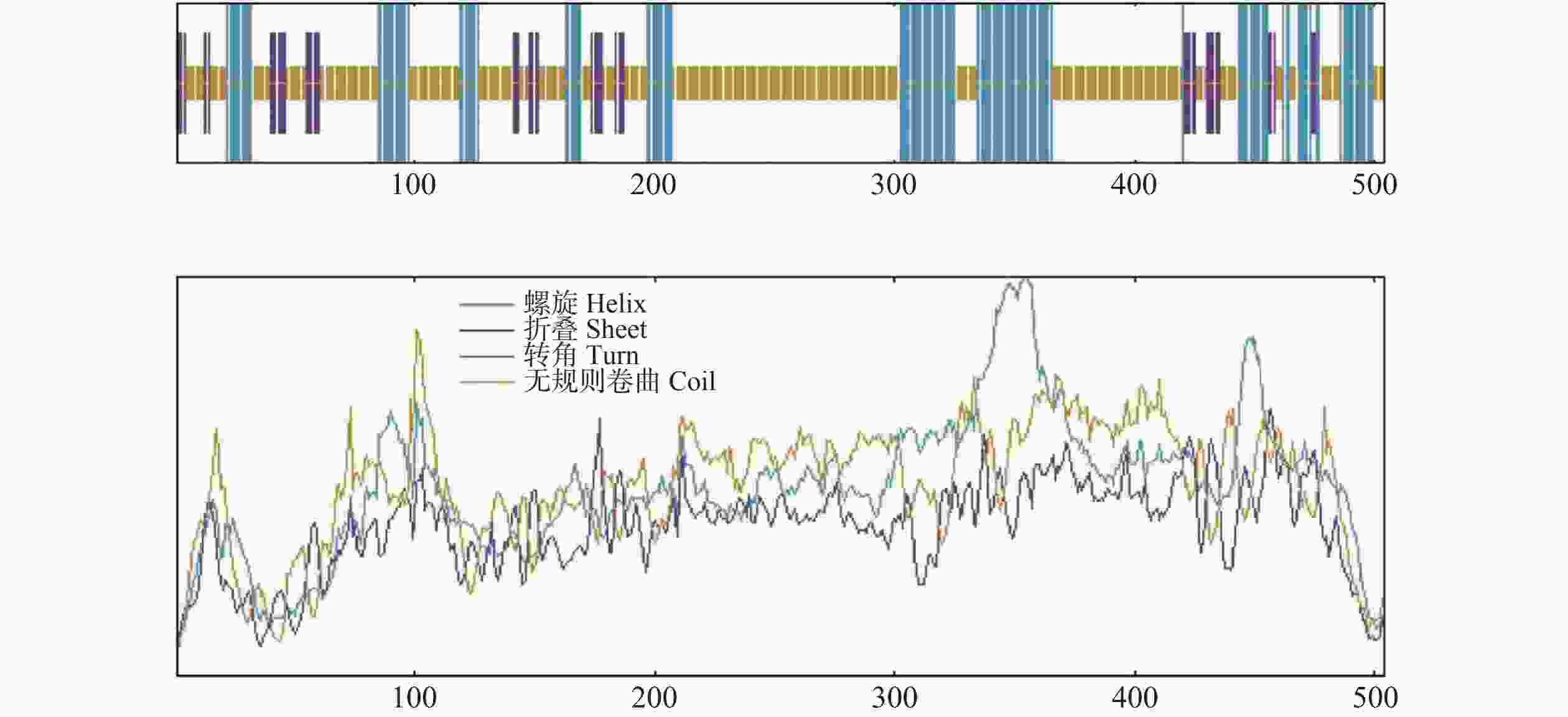
图 8 CdMYC2蛋白二级结构预测图
Figure 8. Secondary structure prediction of CdMYC2 protein
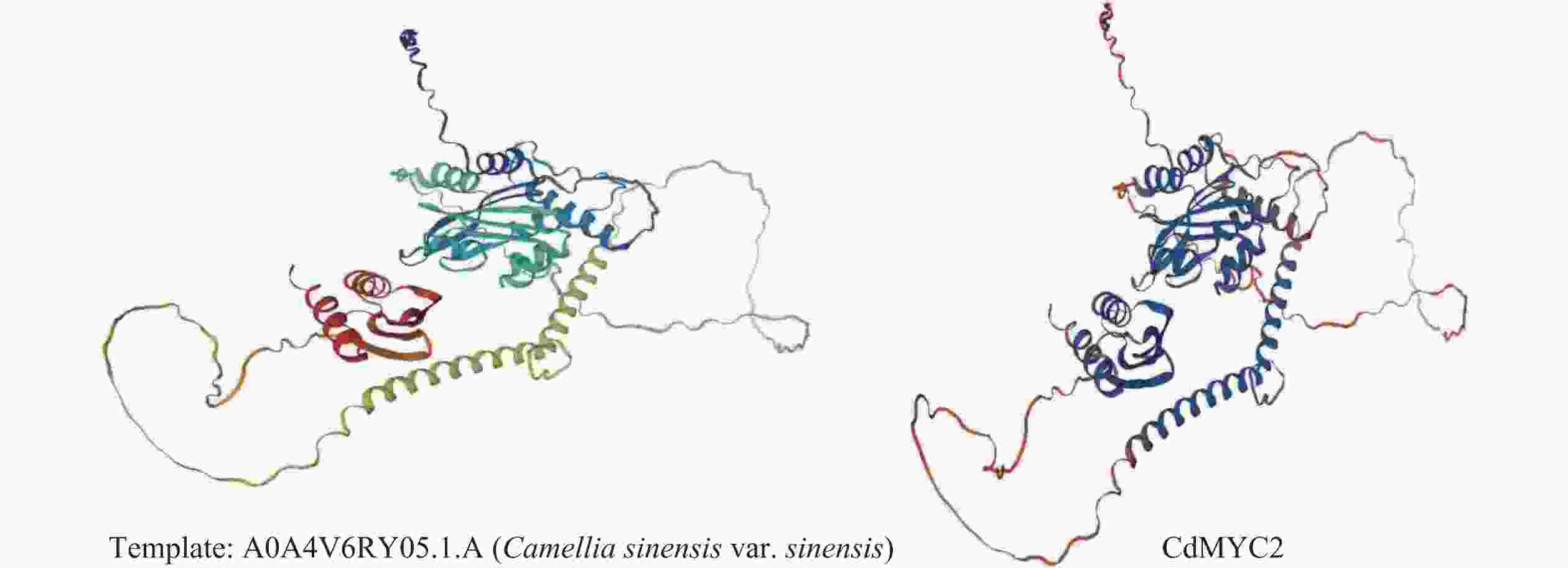
图 9 CdMYC2蛋白三级结构预测模型
Figure 9. The tertiary structure prediction model of CdMYC2 protein
-
利用ExPASy-ProtParam tool在线分析CdMYC2编码的蛋白质的理化性质。结果显示,CdMYC2分子式(formula)为C2424H3851N713O789S21;分子质量(molecular weight)为
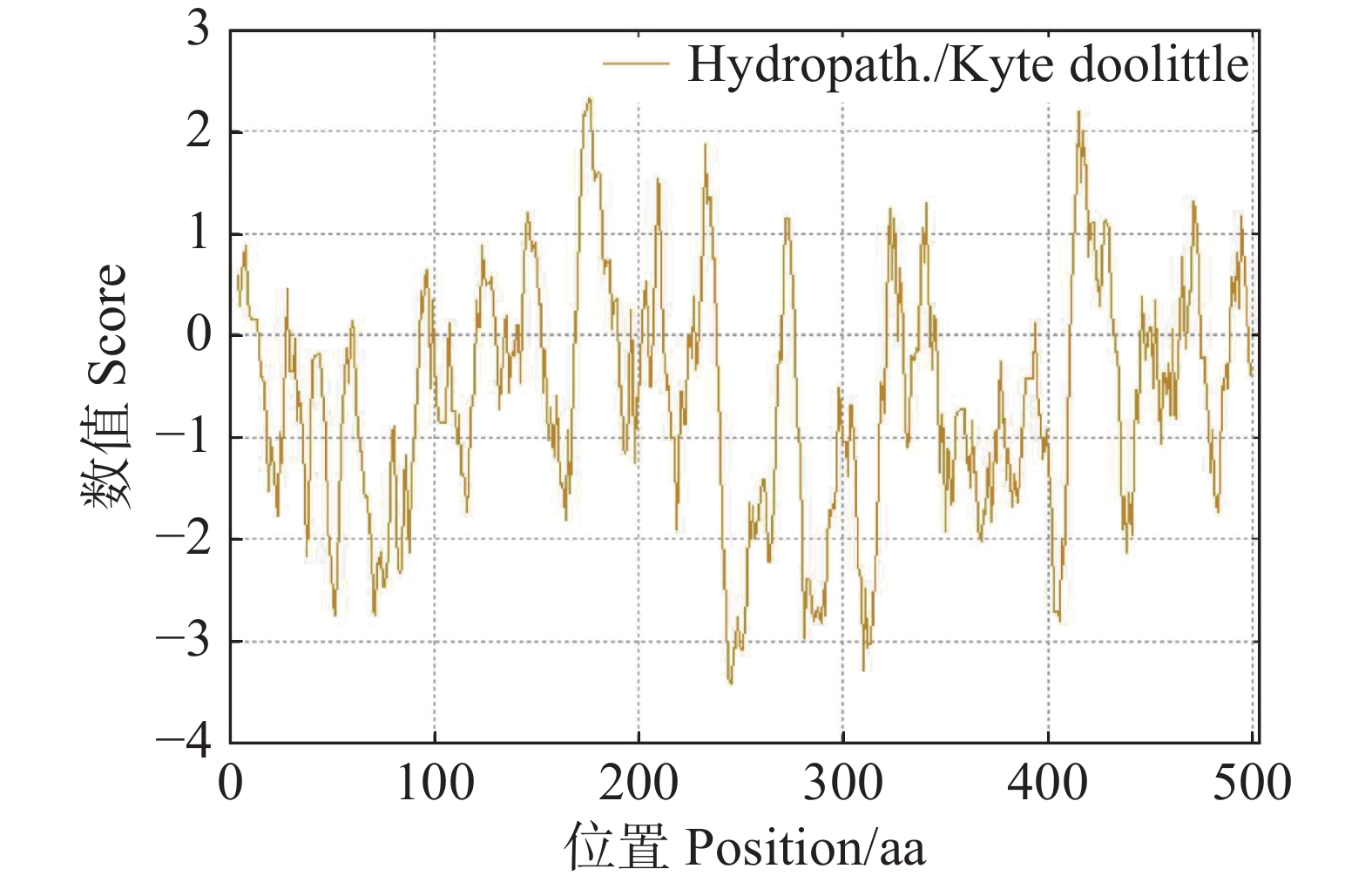
56279.81 ;等电点(theoretical pI)为5.72;在构成该蛋白质的氨基酸中,丝氨酸(Ser)的占比最高,达到了10.1%,不稳定系数(instability index)为46.62,属于不稳定蛋白质,脂溶指数(aliphatic index)为73.10,亲水性系数(GRAVY)为−0.608,属于亲水性蛋白。使用 ExPASy ProtScale 在线软件对CdMYC2蛋白亲疏水性进行评估,结果显示CdMYC2的亲疏水性值在−3.411~2.333,分布总体上呈现出小于0的趋势,这表明CdMYC2更倾向于亲水性(图10)。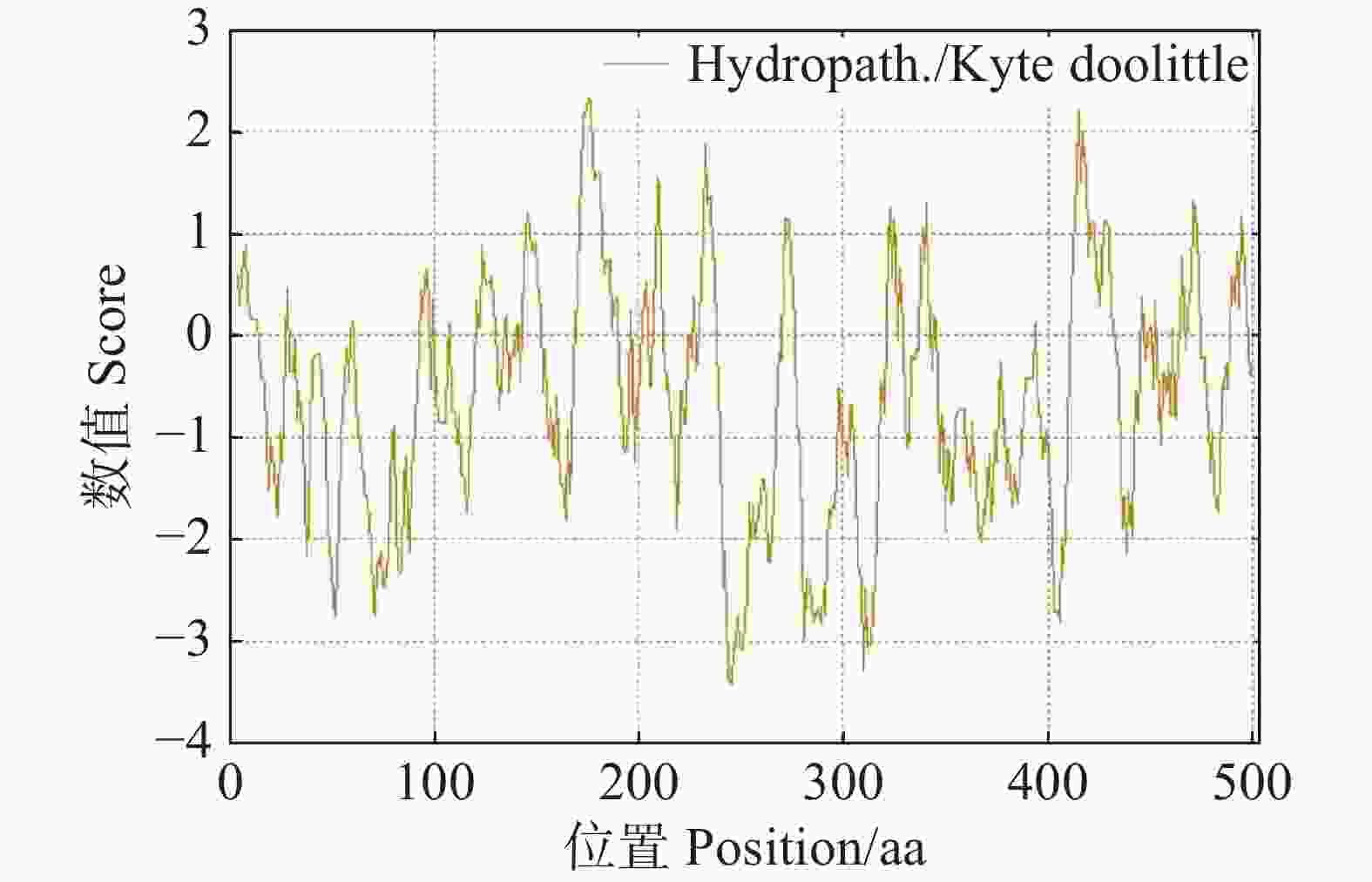
图 10 CdMYC2蛋白的亲疏水性预测图
Figure 10. Prediction of hydrophilicity and hydrophobicity of CdMYC2 protein
-
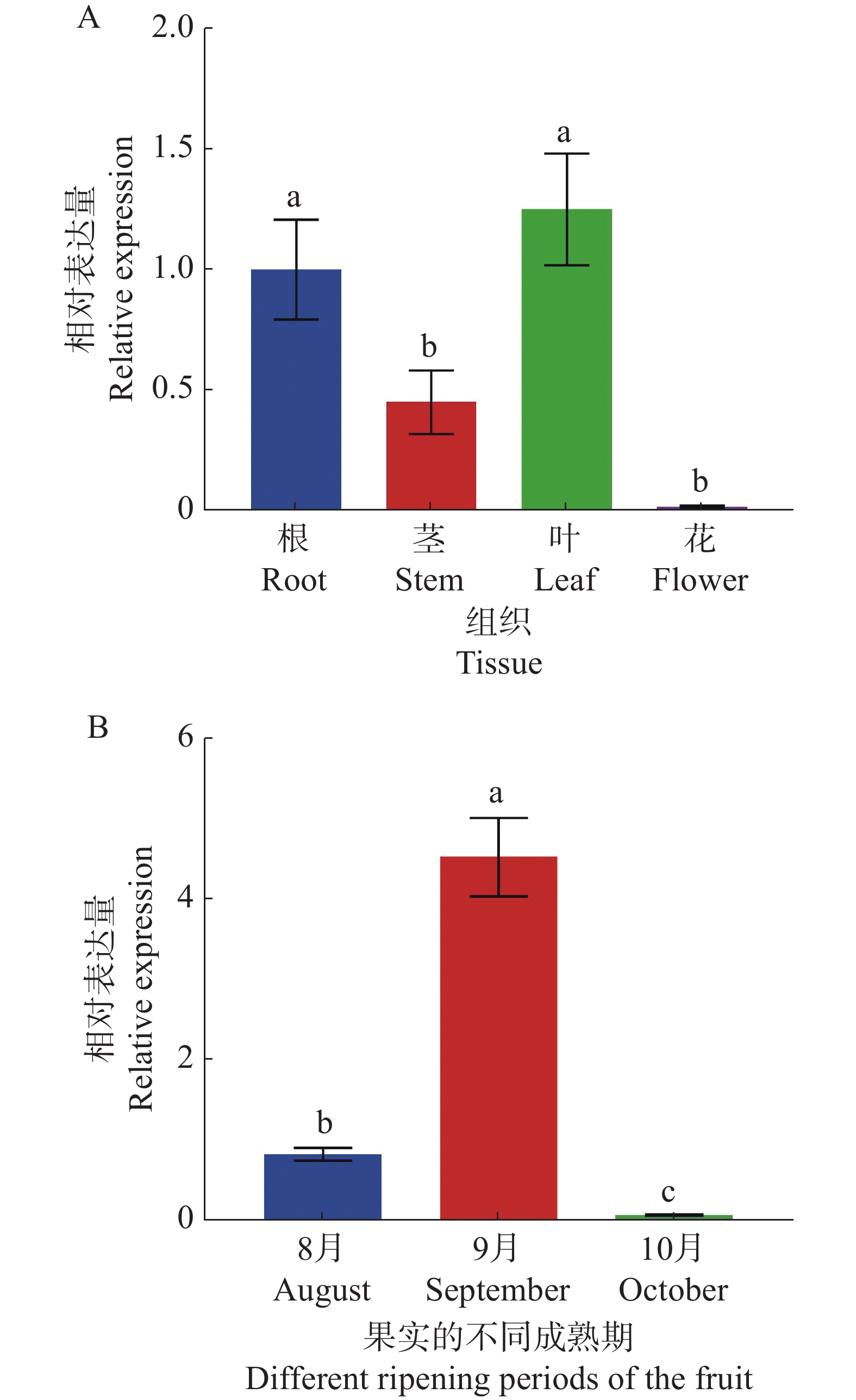
为了深入了解CdMYC2的表达模式,使用实时荧光定量PCR(RT-qPCR)技术分析其在越南油茶不同组织部位以及果实不同成熟阶段的表达模式。结果表明,在植物的不同组织中,CdMYC2基因在叶片中的表达最为活跃,其次是根和茎中的表达,但在花中几乎没有表达(图11-A)。而在果实的不同成熟期中,CdMYC2基因在S2时期表达量最高,其次是S1时期,在S3时期的表达量过低(图11-B)。因此,结果表明CdMYC2基因的表达模式具有组织特异性和发育阶段特异性。
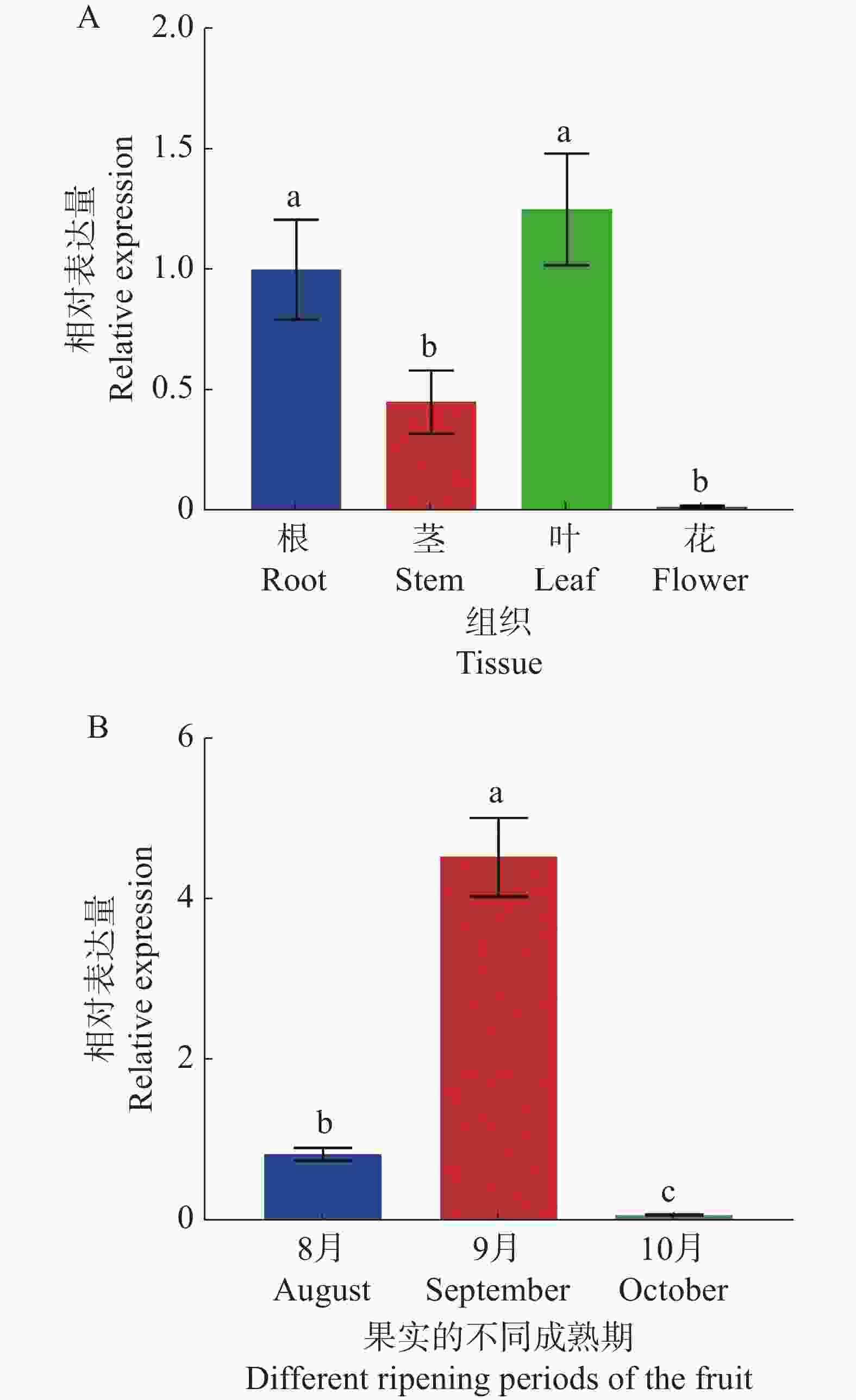
图 11 越南油茶CdMYC2基因的相对表达水平
Figure 11. Relative expression level of CdMYC2 gene in C. drupifera
-
越南油茶作为山茶科山茶属植物,其体内含有多种活性成分,其中三萜皂苷是主要活性成分之一。研究表明MYC2能够调控植物中三萜皂苷的合成。然而,越南油茶CdMYC2是否调控三萜皂苷的研究尚未报道。
MYC2属于碱性螺旋–环–螺旋 (bHLH)家族成员,其C末端含有高度保守的bHLH结构域,该结构域是bHLH家族蛋白的一个标志性特征[2]。在本研究中,对获得的越南油茶CdMYC2进行保守结构域预测,结果显示其含有高度保守的bHLH结构域。进一步通过与其他物种同源蛋白序列的比对分析,笔者确认了CdMYC2与多数木本植物中的MYC2一样,均具备这一高度保守的bHLH结构域。该结构域同样存在于银杏[11]、北柴胡[12]、蒲公英等药用植物MYC2中。这一结果不仅证实了CdMYC2属于bHLH家族的转录因子,还表明其bHLH结构域在进化过程中具有保守性。然而,多序列比对结果也揭示了越南油茶CdMYC2保守结构域中氨基酸的数目和种类仍与其他物种存在差异,其中,与同科同属茶树的同源蛋白在碱基序列上差异小,相似度占比97.24%,而与其余物种的同源蛋白在碱基序列上相似度均在70%以下,这说明在不同物种的进化中MYC2基因并非绝对静态,而是在不同物种间随着亲缘关系的疏远而展现出不同程度的碱基缺失与突变,进化树分析结果进一步印证了这一点,揭示了不同物种MYC2基因间的亲缘关系远近与它们的保守性程度密切相关[13]。
基因在不同组织或发育阶段的表达水平,往往与其在生物体内的角色密切相关。通过RT-qPCR检测发现,CdMYC2基因表达具有显著的组织特异性和发育阶段特异性。在组织特异性方面,CdMYC2基因在叶片中的表达最为活跃,其次是根部和茎部,而在花中几乎不表达。由此,推测CdMYC2基因在叶片中具有较高的表达量可能是因为需要合成更多的三萜皂苷来抵御环境压力或参与光合作用等生理活动;相对而言,花中对这种化合物的需求较低,因此在花中的表达量最低。对此,已有研究表明,药用植物中三萜皂苷合成与受到环境因子影响有关,同时,bHLH转录因子在植物合成次生代谢物(包括黄酮类、萜类、生物碱类等的生物合成)以应对环境压力过程中发挥了重要作用[2,14]。此外,有研究还指出光反应产物是初生代谢与次生代谢过程中的重要物质,在三萜皂苷的生物合成途径中,多个关键步骤(如MVA的形成、MVA磷酸化生成MVAP、SS酶催化FPP缩合)均依赖于ATP和NADP(H)的参与[15]。由于植物叶片是光合作用的主要场所,合理推测CdMYC2基因在越南油茶中表现出的组织特异性表达模式可能与其不同组织对三萜皂苷的具体需求和功能紧密相关。而这一推测与代佳妮[7]、郑蔚等[16]的研究发现越南油茶三萜皂苷生物合成途径中的关键酶基因(CdHMGCR、CdSQS、CdSQE)在叶片中的表达量最高,而在花中的表达量则最低的成果相吻合。在油茶果实的发育过程中,CdMYC2基因在9月份表达量最高,其次是8月份,在10月份的表达量过低。据有关报道,在油茶果成熟过程中籽仁三萜皂苷含量呈现出先增加、后下降的趋势,9月的三萜皂苷含量达最高峰,随后在10月份下降[17]。由此推测CdMYC2基因可能在调控三萜皂苷的生物合成中扮演着重要角色。
综上所述,本研究首次从越南油茶叶片中获得CdMYC2基因序列,为后续深入解析MYC2转录因子对越南油茶三萜皂苷生物合成积累的调控作用奠定了基础。
越南油茶CdMYC2基因克隆、生物信息学分析与表达模式研究
DOI: 10.15886/j.cnki.rdswxb.20250031
 CSTR: 32425.14.j.cnki.rdswxb.20250031
CSTR: 32425.14.j.cnki.rdswxb.20250031
Cloning, bioinformatics analysis and expression pattern of CdMYC2 gene in Camellia drupifera
-
摘要: MYC2是一种重要的转录因子,可以调控植物萜类次生代谢产物的合成。然而,关于越南油茶(Camellia drupifera Lour.)中的CdMYC2转录因子知之甚少。本研究基于越南油茶的转录组注释数据获取了CdMYC2基因的相关序列信息。首次从越南油茶叶片中成功分离并克隆了一个CdMYC2基因,其CDS序列长度为1 515 bp,编码504个氨基酸,具有典型的bHLH-MYC_N super family和bHLH_SF super family保守结构域,通过预测该基因位于细胞核中。多重序列比对结果进一步证实了扩增的CdMYC2属于MYC2家族转录因子。系统进化树显示,CdMYC2与同科同属植物茶树(Camellia sinensis)聚为一支,而与壳斗科栎属植物白栎(Quercus lobata)、夏栎(Q. robur)的分支距离远。RT-qPCR结果表明,CdMYC2基因在叶片中的表达水平高于其他组织部位;而在果实的不同发育阶段中,9月份果实的CdMYC2表达量达到最高。本研究为后续探究MYC2转录因子在调控越南油茶三萜皂苷生物合成途径中的功能提供了重要的参考依据。Abstract: MYC2 is an important transcription factor that regulates the synthesis of terpenoid secondary metabolites in plants. However, little is known about the CdMYC2 transcription factor in C. drupifera. In this study, the related sequence information of CdMYC2 gene was obtained based on the transcriptome annotation data of C. drupifera. A CdMYC2 gene was successfully isolated and cloned from the leaves of C. drupifera for the first time. Its CDS sequence length was
1515 bp, encoding 504 amino acids, with typical bHLH-MYC_N super family and bHLH_SF super family conserved domains. This gene was predicted to be located in the nucleus. Multiple sequence alignment further confirmed that the amplified CdMYC2 belongs to the MYC2 family transcription factor. The phylogenetic tree analysis showed that CdMYC2 was clustered into a clade with Camellia sinensis, a plant belonging to the same family and genus, while it was far away from the clades of Quercus lobata and Q. robur. RT-qPCR results showed that the expression level of CdMYC2 gene was higher in the leaves than in other tissues. Among the different developmental stages of the fruit, the expression of CdMYC2 in the fruit reached the highest in September. This study provides an important reference for the subsequent exploration of the function of MYC2 transcription factor in the regulation of triterpenoid saponin biosynthesis pathway in C. drupifera.-
Key words:
- Camellia drupifera /
- CdMYC2 /
- gene cloning /
- bioinformatics /
- expression analysis
-
图 2 CdMYC2基因CDS核酸及氨基酸序列
Fig. 2 CDS nucleic acid and amino acid sequence of CdMYC2 gene
图 4 CdMYC2与不同物种同源蛋白序列比对
注:*代表DNA结合位点。
Fig. 4 Sequence alignment of CdMYC2 with homologous proteins of different species
Note: * represents a DNA binding site.
图 7 CdMYC2蛋白磷酸化位点预测结果
Fig. 7 Prediction results of CdMYC2 protein phosphorylation sites
图 10 CdMYC2蛋白的亲疏水性预测图
Fig. 10 Prediction of hydrophilicity and hydrophobicity of CdMYC2 protein
图 11 越南油茶CdMYC2基因的相对表达水平
注:不同小写字母表示具有显著性差异(P<0.05)。
Fig. 11 Relative expression level of CdMYC2 gene in C. drupifera
Note: Different lowercase letters indicate significant differences (P < 0.05).
表 1 用于基因克隆和RT-qPCR 反应的引物序列
Table 1 Primer sequences for gene cloning and RT-qPCR reaction
引物 Primer 序列(5′—3′)Sequence (5′—3′) CdMYC2-F ATGGATGAATTAATCATATCGTCAT CdMYC2-R CTAACAAGTCTGTCCTACCCC CdMYC2-QF CATCCAACAAAAGATAGGTG CdMYC2-QR CTTCTCTGTGTGTAAAGCCA Actin-F TTGGCATCGTTGAGGGTCT Actin-R CAGTGGGAACACGGAAAGC  下载: 导出CSV
下载: 导出CSV
-
[1] Wasternack C, Hause B. Jasmonates: biosynthesis, perception, signal transduction and action in plant stress response, growth and development. An update to the 2007 review in Annals of Botany[J]. Annals of Botany, 2013, 111(6): 1021−1058. https://doi.org/10.1093/aob/mct067 doi: 10.1093/aob/mct067 [2] 郑嘉瑞, 杨晓燕, 叶家保, 等. MYC2转录因子在植物中的功能研究进展[J]. 园艺学报, 2023, 50(4): 896−908. https://doi.org/10.16420/j.issn.0513-353x.2022-0102 doi: 10.16420/j.issn.0513-353x.2022-0102 [3] Kazan K, Manners J M. MYC2: the master in action[J]. Molecular Plant, 2013, 6(3): 686−703. https://doi.org/10.1093/mp/sss128 doi: 10.1093/mp/sss128 [4] Hong G J, Xue X Y, Mao Y B, et al. Arabidopsis MYC2 interacts with DELLA proteins in regulating sesquiterpene synthase gene expression[J]. The Plant Cell, 2012, 24(6): 2635−2648. https://doi.org/10.1105/tpc.112.098749 doi: 10.1105/tpc.112.098749 [5] Liu T, Liao J, Shi M, et al. A jasmonate-responsive bHLH transcription factor TaMYC2 positively regulates triterpenes biosynthesis in Taraxacum antungense Kitag[J]. Plant Science, 2023, 326: 111506. https://doi.org/10.1016/j.plantsci.2022.111506 doi: 10.1016/j.plantsci.2022.111506 [6] 刘进平, 周扬, 胡海燕, 等. 热带油茶种质资源综述[J]. 热带林业, 2022, 50(4): 70−76. https://doi.org/10.3969/j.issn.1672-0938.2022.04.015 doi: 10.3969/j.issn.1672-0938.2022.04.015 [7] 代佳妮. 越南油茶茶皂素合成关键酶基因CvSQS、CvSQE的克隆与功能分析 [D]. 海口: 海南大学, 2021. https://doi.org/10.27073/d.cnki.ghadu.2021.000501 [8] 刘运锋, 邱兆志, 申新刚, 等. 茶油的化学成分和药理作用研究进展[J]. 南方农业, 2020, 14(9): 132−134. https://doi.org/10.19415/j.cnki.1673-890x.2020.09.067 doi: 10.19415/j.cnki.1673-890x.2020.09.067 [9] 黄运安. 油茶中三萜皂苷及其生物活性研究进展[J]. 食品安全质量检测学报, 2024, 15(13): 59−67. https://doi.org/10.19812/j.cnki.jfsq11-5956/ts.20240312008 doi: 10.19812/j.cnki.jfsq11-5956/ts.20240312008 [10] Li Y, Wang J, Li L, et al. Natural products of pentacyclic triterpenoids: from discovery to heterologous biosynthesis[J]. Natural Product Reports, 2023, 40(8): 1303−1353. https://doi.org/10.1039/D2NP00063F doi: 10.1039/D2NP00063F [11] Zheng J, Liao Y, Ye J, et al. The transcription factor MYC2 positively regulates terpene trilactone biosynthesis through activating GbGGPPS expression in Ginkgo biloba[J]. Horticulture Research, 2024, 11(10): uhae228. https://doi.org/10.1093/hr/uhae228 doi: 10.1093/hr/uhae228 [12] 王晨, 安立成, 李剑超, 等. 北柴胡MYC2转录因子的克隆及茉莉酸诱导的调控分析[J]. 植物生理学报, 2021, 57(2): 439−450. https://doi.org/10.13592/j.cnki.ppj.2020.0381 doi: 10.13592/j.cnki.ppj.2020.0381 [13] Alix K, Gérard P R, Schwarzacher T, et al. Polyploidy and interspecific hybridization: partners for adaptation, speciation and evolution in plants[J]. Annals of Botany, 2017, 120(2): 183−194. https://doi.org/10.1093/aob/mcx079 doi: 10.1093/aob/mcx079 [14] 焦晓林, 高微微. 环境因子对药用植物三萜皂苷合成影响的研究进展[J]. 中草药, 2011, 42(2): 398−402. [15] Dellas N, Thomas S T, Manning G, et al. Discovery of a metabolic alternative to the classical mevalonate pathway[J]. eLife, 2013, 2: e00672. https://doi.org/10.7554/eLife.00672 doi: 10.7554/eLife.00672 [16] 郑蔚, 代佳妮, 严和琴, 等. 越南油茶HMGCR基因的克隆及表达分析[J]. 分子植物育种, 2022, 20(13): 4296−4304. https://doi.org/10.13271/j.mpb.020.004296 doi: 10.13271/j.mpb.020.004296 [17] 李好, 方学智, 钟海雁, 等. 油茶果成熟过程茶皂素含量分布状况[J]. 粮食与油脂, 2013, 26(9): 24−26. https://doi.org/10.3969/j.issn.1008-9578.2013.09.007 doi: 10.3969/j.issn.1008-9578.2013.09.007 -

 点击查看大图
点击查看大图
计量
- 文章访问数: 698
- HTML全文浏览量: 512
- PDF下载量: 11
- 被引次数: 0
