


-
榴莲(Durio zibethinus)为锦葵科热带常绿乔木,原产于马来西亚、文莱、印度尼西亚,多种植于东南亚国家,其中以泰国最多。中国是全球主要的榴莲消费国之一,2022年中国从泰国进口的鲜榴莲数量达78.40万t,随着中国榴莲消费需求不断增加,近年来,中国海南、广东、广西、云南和台湾等省(区)开始尝试榴莲的引种栽培[1]。海南岛北纬18°以南区域是我国唯一规模化种植榴莲的热带地区,受地形与气候双重塑造,形成“三区一带”种植格局。三亚、保亭、乐东等市县是主要种植区,种植面积占全岛78%。截至2024年4月份,海南榴莲种植规模已超过
2666.67 hm2[2]。海南岛北部等高纬度地区,气温较低,易受到寒潮台风等影响,成为限制海南榴莲产业从南向北稳步扩大的主要因素[3]。胚胎发育晚期丰富蛋白(late embryogenesis abundant protein,LEA),最初在棉花(Gossypium hirsutum)中被发现,表明其在种子发育晚期高度表达[4]。有研究发现,干旱、低温、盐胁迫和脱落酸处理均可诱导LEA蛋白表达[5]。目前,LEA基因家族已在多个植物中陆续发现并鉴定,如拟南芥(Arabidopsis thaliana)[6]、水稻(Oryza sativa)[7]、花生(Arachis hypogaea)[8]和茶树(Camellia sinensis)[9]等。基于其序列相似性和保守结构域,LEA蛋白可被划分为8个亚族,分别是:LEA1、LEA2、LEA3、LEA4、LEA5、LEA6、DHN(dehydrin)和SMP(seed maturation protein)[10]。LEA蛋白通常富含亲水性氨基酸(如甘氨酸、丙氨酸、谷氨酸、赖氨酸等),使其在缺水条件下仍能抵抗沉淀或聚集,维持其溶解状态和功能活性;且多数LEA蛋白在常规条件下呈无序构象,其在脱水或胁迫条件下可能形成α-螺旋结构,维持细胞膜结构的稳定,发挥保护功能[11]。由于其上述2种特征,使LEA蛋白在脱水、低温等非生物胁迫条件下,可以稳定细胞膜和蛋白质结构,进而维持细胞功能。本研究对榴莲LEA基因家族成员进行了鉴定和分析,研究成果为探究DzLEAs基因在榴莲抵御低温胁迫中的作用提供了理论支撑。
-
榴莲的基因组序列和注释文件来自figshare在线网站(
https://figshare.com/ articles/dataset/Durian_genome_annotation/25237591 )[12],拟南芥LEAs家族的基因组序列和注释文件来自tair数据库(https://www.arabidopsis.org/ )[6],利用TBtools软件[13](版本:2.326)通过BLAST方法对榴莲LEA基因家族成员进行鉴定。同时,从Pfam数据库(http://pfam.xfam.org/ )中下载了LEA1(PF03760)、LEA2(PF03168)、LEA3(PF03242)、LEA4(PF02987)、LEA5(PF00477)、LEA6(PF10714)、DHN(PF00257)、 和SMP(PF04927)亚家族的隐马尔可夫模型(Hidden Markov Model,HMM)。通过TBtools的simple HMM search程序得到榴莲候选LEAs基因。通过NCBI conserved Domains(http://www. ncbi.nlm.nih.gov/Structure/cdd/wrpsb.cgi )数据库、Pfam数据库(http://pfam-legacy.xfam.org/ ),对BLASTP和HMM两种方法得到的榴莲LEAs候选基因进行结构域的确定。根据LEA家族成员在染色体上的位置,将其依次编号为DzLEA1~DzLEA46用于后续分析。 -
通过TBtools软件Protein Parameter Calc程序[13],分析DzLEAs蛋白的氨基酸长度(Amino acid length)、理论等电点(Theoretical Isoelectric Point)、分子量(Molecular Weight)、不稳定系数(Instability Index)、亲水性平均系数(Grand Average of Hydropathicity,GRAVY)等理化性质,使用WoLF PSORT网站(
https://wolfpsort.hgc.jp/ )预测DzLEAs蛋白的亚细胞定位[14]。 -
通过MEGA12软件中最大似然法(Maximum Likelihood, ML)构建系统进化树,Bootstrap重复值设置为1 000,使用iTOL(
https://itol.embl.de/ )在线工具对生成的系统进化树进行美化[15]。 -
在NCBI保守结构域数据库(
https://www.ncbi.nlm.nih.gov/Structure/bwrpsb/bwrpsb.cgi )分析榴莲LEA基因家族的保守结构域;使用MEME套件(https://meme-suite.org/meme/ )分析榴莲LEA基因家族的保守基序,搜索motif数量设置为20个;根据榴莲的基因组注释,分析DzLEAs的基因结构。最后,通过TBtools的Gene Structure view(Advanced)程序对DzLEAs的进化树、保守基序、保守结构域和基因结构进行可视化[13]。 -
通过TBtools软件获取DzLEAs基因起始密码子上游2 000 bp的序列,在PlantCARE网站(
https://bioinformatics.psb.ugent.be/webtools/plantcare/html/ )对获取的序列进行分析,最后通过TBtools软件对分析结果进行可视化[13]。 -
基于榴莲和拟南芥的基因组文件和注释文件[12],通过TBtools软件[13]获取其染色体长度、基因密度和位置信息,通过MCScanX对榴莲与拟南芥LEA基因家族进行物种内和物种间共线性分析,采用默认参数;最后通过Advanced Circos程序对共线性分析结果进行可视化。
-
在eggNOG-mapper网站(
http://eggnog-mapper.embl.de/ )获取榴莲的GO和KEGG注释文件[16],之后通过TBtools软件中GO Enrichment和KEGG Enrichment Analysis程序中对DzLEAs基因进行富集分析,最后通过Enrichment Bar Plot程序对分析结果进行可视化[13]。 -
以‘猫山王’和‘金枕’2个榴莲品种为研究对象,榴莲苗高为1 m,苗龄为12个月,其中,处理组置于恒温培养箱中进行培养,光照设置为12 h光照和12 h黑暗循环,温度设置为10℃;对照组置于室内空调屋进行培养,空气相对湿度约为45%,温度设置为26℃。取26℃常温、10℃低温处理1 d、10℃低温处理5 d叶片样品进行转录组数据分析,每个处理重复3次。
分别取‘猫山王’和‘金枕’在26℃常温、10℃处理1 d、10℃处理3 d、10℃处理5 d、10℃处理7 d的叶片样品进行qRT-PCR实验,生物学重复3次取样,混样分3次技术重复进行后续qRT-PCR实验。采用十六烷基三甲基溴化铵法提取叶片样品RNA,通过超微紫外分光光度计(ND-100C)测定提取的RNA浓度。根据所测样品RNA浓度统一其含量,使用含gDNA清除剂的SPARKscript Ⅱ RT Plus Kit试剂盒(SparkJade)合成第1链互补DNA(cDNA)。qRT-PCR的引物(表1)通过NCBI中的Primer-BLAST(
https://www.ncbi.nlm.nih.gov/tools/primer-blast/ )工具设计[17]。通过Applied Biosystems 7500实时定量PCR系统(Life Technologies,USA)进行qRT-PCR,反应体系包括10 μL 2×SYBR qPCR Mix、1 μL cDNA、0.4 μL Forward Primer、0.4 μL Reverse Primer、0.4 μL ROX Reference Dye Ⅱ、7.8 μL RNase Free H2O,总体积为20 μL。采用相对定量法(ΔΔCt法)计算样品的相对表达量。表 1 实时荧光定量PCR引物及其碱基序列
Table 1. Sequences of primers for quantitative real-time PCR
基因名称
Gene基因编号
Sequence ID正向引物序列(5′−3′)
Forward primer sequence(5′−3′)反向引物序列(5′−3′)
Reverse primer sequence(5′−3′)DzLEA5 evm.model.GRLG01.2002 ACCCCATGAAGGTGAGACGA TGGGAAGCGCTCTCCTTTTC DzLEA18 evm.model.GRLG16.937 GCCAAGGACCGAGGGTTG TCCTTGCGCTCCTCCTCT DzLEA21 evm.model.GRLG17.729 GGAGCAGCAAGGAACGCT CTCGGCCACGTCGATCTC DzLEA26 evm.model.GRLG18.174 GGATATGCTGCGGGATCACA GTCCGGAACCCATGAAACCT DzLEA39 evm.model.GRLG24.892 CGGCAACAAACTCTGCGG CTCGGCCACGTCGATCTC DzLEA45 evm.model.GRLG27.783 GTGGCGCCTAAGGATGCA TGCATGGCAGCAGATTCCA Dz04.19 evm.model.GRLG04.19 CGACGAGCTCAAGGAGGC ATGGCACGAGCAGCGAAT Dz27.910 evm.model.GRLG27.910 GCAAGTAGCGGCTGTCCA GCACTTCAACGGTCCCGA Dz25.635 evm.model.GRLG25.635 TGCGGCTGCCATTGATGA GCACTGTCTGCTGCTCAA Dz20.232 evm.model.GRLG20.232 AGCATCAAGACACGCAAGGA ATGTCGTTTGGCCGCCAT Dz4GAPDH09.1422 evm.model.GRLG09.1422 TGGGCTGAGACAGGAGCT CCACAACAAACATGGGGGC -
基于BLAST和HMM两种方法,从榴莲基因组中共鉴定出46个LEA基因家族成员,根据其在染色体上的位置分别命名为DzLEA1~DzLEA46(图1)。
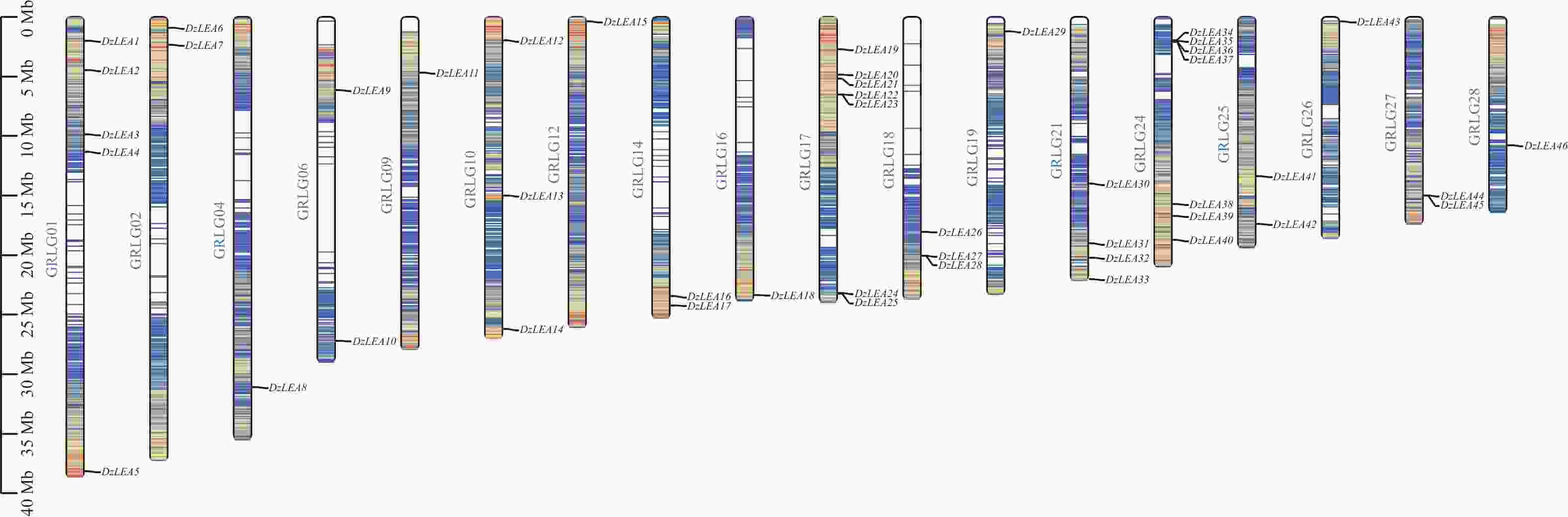
图 1 DzLEAs基因的染色体定位
Figure 1. Chromosomal localization of the DzLEAs gene
由表2可知,46个榴莲LEA基因家族成员编码的蛋白氨基酸序列长度在65~467个之间,蛋白质的理论分子质量范围在6.965 8~52.080 58 kDa之间。理论等电点在4.33~9.79之间,其中,27个(59%)DzLEA蛋白的等电点小于7,为酸性蛋白;19个(41%)DzLEA蛋白的等电点大于7,为碱性蛋白。46个DzLEA蛋白中,稳定蛋白和不稳定蛋白均占50%(23个)。脂肪族系数在16.67~107.81之间,且大多数DzLEA蛋白为亲水性蛋白(44个),仅DzLEA20和DzLEA30为疏水性蛋白。
表 2 榴莲DzLEA基因家族特征分析
Table 2. Features of DzLEA gene family
基因名称
Gene基因编号
Sequence ID基因
家族
Gene
family氨基酸长度
Amino acid length分子质量
Molecular weight理论等电点
Theoretical pI不稳定系数
Instability Index脂肪族系数
Aliphatic Index亲水性
平均系数
Grand
average of hydropathicity亚细胞定位
Subcellular localizationDzLEA1 evm.model.
GRLG01.168LEA1 148 15 343.01 8.07 32.05 52.97 −0.8 细胞核 DzLEA2 evm.model.
GRLG01.459LEA4 424 47 446.41 5.44 37.00 46.98 −1.29 细胞核 DzLEA3 evm.model.
GRLG01.897LEA5 134 15 000.93 9.79 44.68 70.6 −0.657 细胞核 DzLEA4 evm.model.
GRLG01.981LEA2 314 34 879.84 4.82 23.95 97.74 −0.329 细胞质 DzLEA5 evm.model.
GRLG01.2002DHN 217 24 476.28 5.21 56.28 50.28 −1.469 细胞核 DzLEA6 evm.model.
GRLG02.113LEA6 119 13 380.79 5.67 39.91 63.19 −0.873 细胞核 DzLEA7 evm.model.
GRLG02.284DHN 129 14 547.17 5.92 38.65 48.45 −1.479 细胞质 DzLEA8 evm.model.
GRLG04.1406LEA4 68 7 164.88 6.17 49.42 21.91 −1.128 细胞核 DzLEA9 evm.model.
GRLG06.455LEA3 92 10 608.17 7.97 39.78 75.33 −0.738 线粒体 DzLEA10 evm.model.
GRLG06.838LEA4 89 9 788.14 9.44 36.21 67.08 −0.84 线粒体 DzLEA11 evm.model.
GRLG09.393LEA6 467 52 080.58 8.43 54.41 57.24 −0.995 细胞核 DzLEA12 evm.model.
GRLG10.285LEA4 219 24 550.1 5.21 47.73 57.12 −1.093 线粒体 DzLEA13 evm.model.
GRLG10.1061LEA5 154 16 970.59 5.73 40.03 49.48 −1.089 细胞核 DzLEA14 evm.model.
GRLG10.1609SMP 255 26 106.87 4.76 44.96 72.24 −0.37 细胞核 DzLEA15 evm.model.
GRLG12.47LEA5 148 16 148.01 9.76 53.95 47.5 −1.164 细胞核 DzLEA16 evm.model.
GRLG14.853DHN 148 16 966.69 9.22 60.67 38.92 −1.568 细胞核 DzLEA17 evm.model.
GRLG14.963DHN 175 19 794.94 6.23 33.38 42.46 −1.727 细胞核 DzLEA18 evm.model.
GRLG16.937DHN 225 25 603.54 5.39 64.15 52 −1.455 细胞核 DzLEA19 evm.model.
GRLG17.386LEA2 151 16 479.04 5.13 22.74 107.81 −0.004 细胞质 DzLEA20 evm.model.
GRLG17.677LEA2 213 23 484.56 9.63 40.25 86.67 0.306 质膜 DzLEA21 evm.model.
GRLG17.729LEA3 98 10 476.93 9.39 32.05 75.71 −0.218 叶绿体 DzLEA22 evm.model.
GRLG17.907SMP 199 20 782.97 4.33 29.71 83.52 −0.239 细胞质 DzLEA23 evm.model.
GRLG17.908SMP 209 21 511.99 5.58 19.87 73.44 −0.346 叶绿体 DzLEA24 evm.model.
GRLG17.1800LEA4 68 7 255.97 6.55 44.54 27.65 −1.144 细胞核 DzLEA25 evm.model.
GRLG17.1806LEA4 66 7 012.69 8.06 45.65 26.82 −1.162 细胞核 DzLEA26 evm.model.
GRLG18.174LEA3 98 10 468.01 9.51 27.63 79.59 −0.136 叶绿体 DzLEA27 evm.model.
GRLG18.273SMP 218 22 421.95 4.53 27.47 77.25 −0.141 细胞质 DzLEA28 evm.model.
GRLG18.274SMP 235 24 596.15 5.26 28.91 69.79 −0.565 线粒体 DzLEA29 evm.model.
GRLG19.74LEA5 188 21 184.08 9.32 56.22 74.2 −0.728 细胞核 DzLEA30 evm.model.
GRLG21.580SMP 138 14 227.09 4.85 37.66 104.06 0.151 细胞质 DzLEA31 evm.model.
GRLG21.828LEA4 89 9 524.33 6.13 51.25 36.4 −1.126 细胞核 DzLEA32 evm.model.
GRLG21.971LEA3 104 11 155.67 7.92 34.59 73.27 −0.207 叶绿体 DzLEA33 evm.model.
GRLG21.1142LEA4 160 17 803.70 9.23 34.74 44.06 −1.263 细胞核 DzLEA34 evm.model.
GRLG24.39LEA4 68 7 230.00 5.31 54.78 17.65 −1.131 细胞核 DzLEA35 evm.model.
GRLG24.42LEA4 66 7 046.73 7.86 42.34 16.67 −1.312 线粒体 DzLEA36 evm.model.
GRLG24.44LEA4 66 7 028.69 7.86 41.05 22.58 −1.283 线粒体 DzLEA37 evm.model.
GRLG24.45LEA4 66 7 046.73 7.86 42.34 16.67 −1.312 线粒体 DzLEA38 evm.model.
GRLG24.756SMP 349 36 686.51 4.5 27.85 72.06 −0.367 细胞质 DzLEA39 evm.model.
GRLG24.892LEA3 98 10 348.88 9.51 27.74 76.63 −0.129 叶绿体 DzLEA40 evm.model.
GRLG24.1149LEA2 206 22 466.73 4.92 45.57 96.12 −0.233 细胞质 DzLEA41 evm.model.
GRLG25.862LEA3 86 9 907.57 9.69 38.16 73.72 −0.614 叶绿体 DzLEA42 evm.model.
GRLG25.1244LEA4 108 11 383.16 5.04 46.33 30 −1.132 细胞核 DzLEA43 evm.model.
GRLG26.6LEA4 310 33 834.60 5.67 36.56 52.77 −0.992 细胞外 DzLEA44 evm.model.
GRLG27.782SMP 177 18 410.53 6.75 37.62 78.53 −0.323 细胞核 DzLEA45 evm.model.
GRLG27.783SMP 65 6 965.80 4.35 59.62 57.23 −0.497 线粒体 DzLEA46 evm.model.
GRLG28.782LEA4 111 11 764.55 5.65 40.11 31.89 −1.196 细胞核 DzLEA蛋白的亚细胞定位预测表明:LEA1亚家族的1个成员(DzLEA1)、LEA5亚家族的3个成员(DzLEA13、DzLEA15、DzLEA29)和LEA6亚家族的2个成员(DzLEA6、DzLEA11)均位于细胞核内;LEA2亚家族的1个成员(DzLEA20)位于质膜内,其余3个成员(DzLEA4、DzLEA19、DzLEA40)位于细胞质内;LEA3亚家族的1个成员(DzLEA9)位于线粒体内,其余5个成员(DzLEA21、DzLEA26、DzLEA32、DzLEA39、DzLEA41)位于叶绿体内;LEA4亚家族的1个成员(DzLEA43)位于细胞外,5个成员(DzLEA10、DzLEA12、DzLEA35、DzLEA36、DzLEA37)位于线粒体内,其余均位于细胞核内;DHN亚家族的1个成员位于细胞质内,其余DHN成员位于细胞核内;SMP亚家族的成员在细胞核、细胞质、线粒体和叶绿体中均有存在。
-
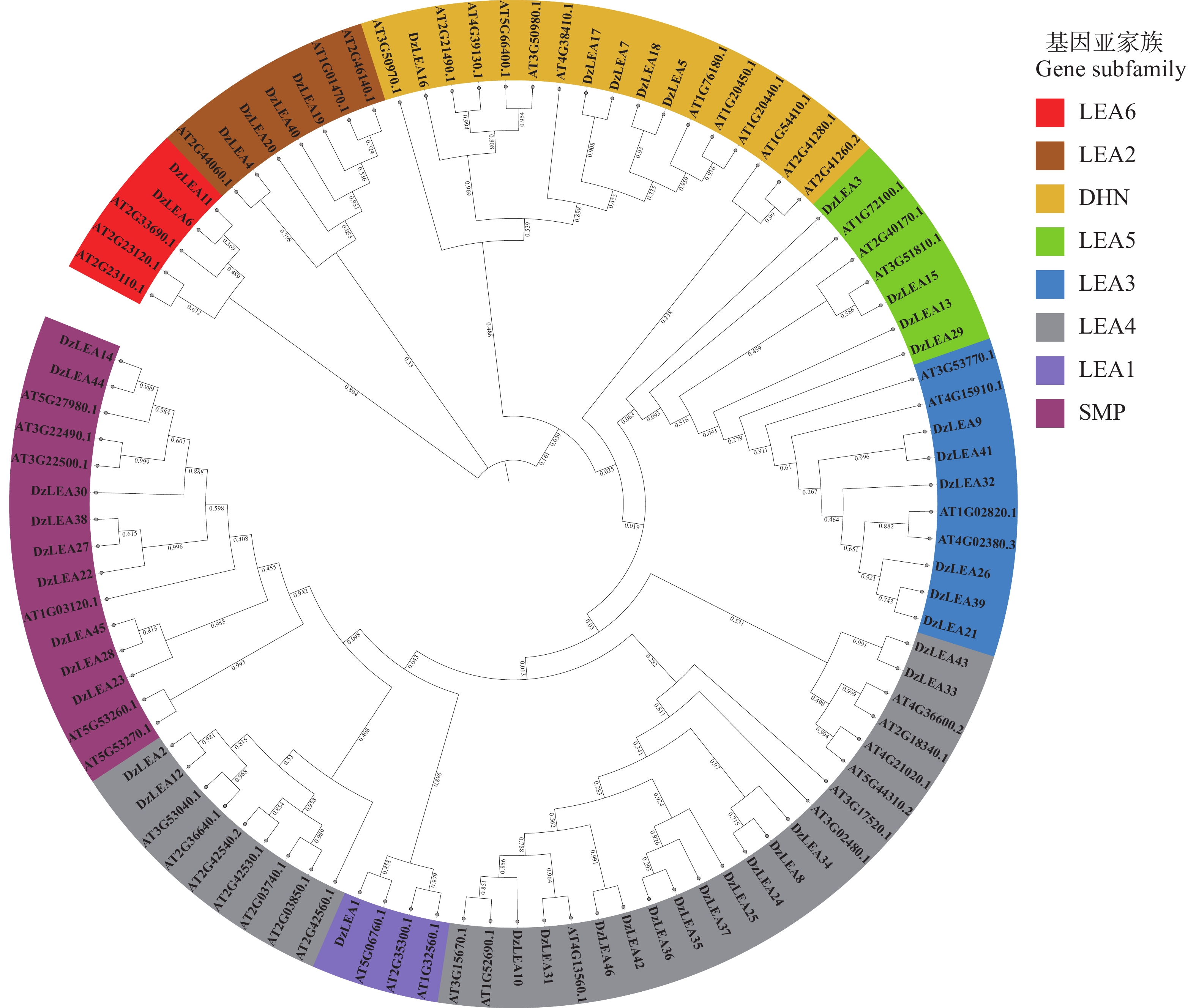
将榴莲与拟南芥LEA家族成员共同构建系统进化树(图2),参考拟南芥中LEA蛋白亚家族分类结果[6],发现DzLEAs共分为8个亚家族:LEA1、LEA2、LEA3、LEA4、LEA5、LEA6、DHN和SMP。其中,LEA4亚家族成员最多为15个,LEA2、LEA3、LEA5、LEA6、DHN和SMP亚家族成员分别有4、6、4、2、5和9个,LEA1亚家族成员最少为1个。榴莲与拟南芥的蛋白分类几乎一致,说明榴莲LEA基因在长期进化的过程中,这些蛋白质没有遗失。
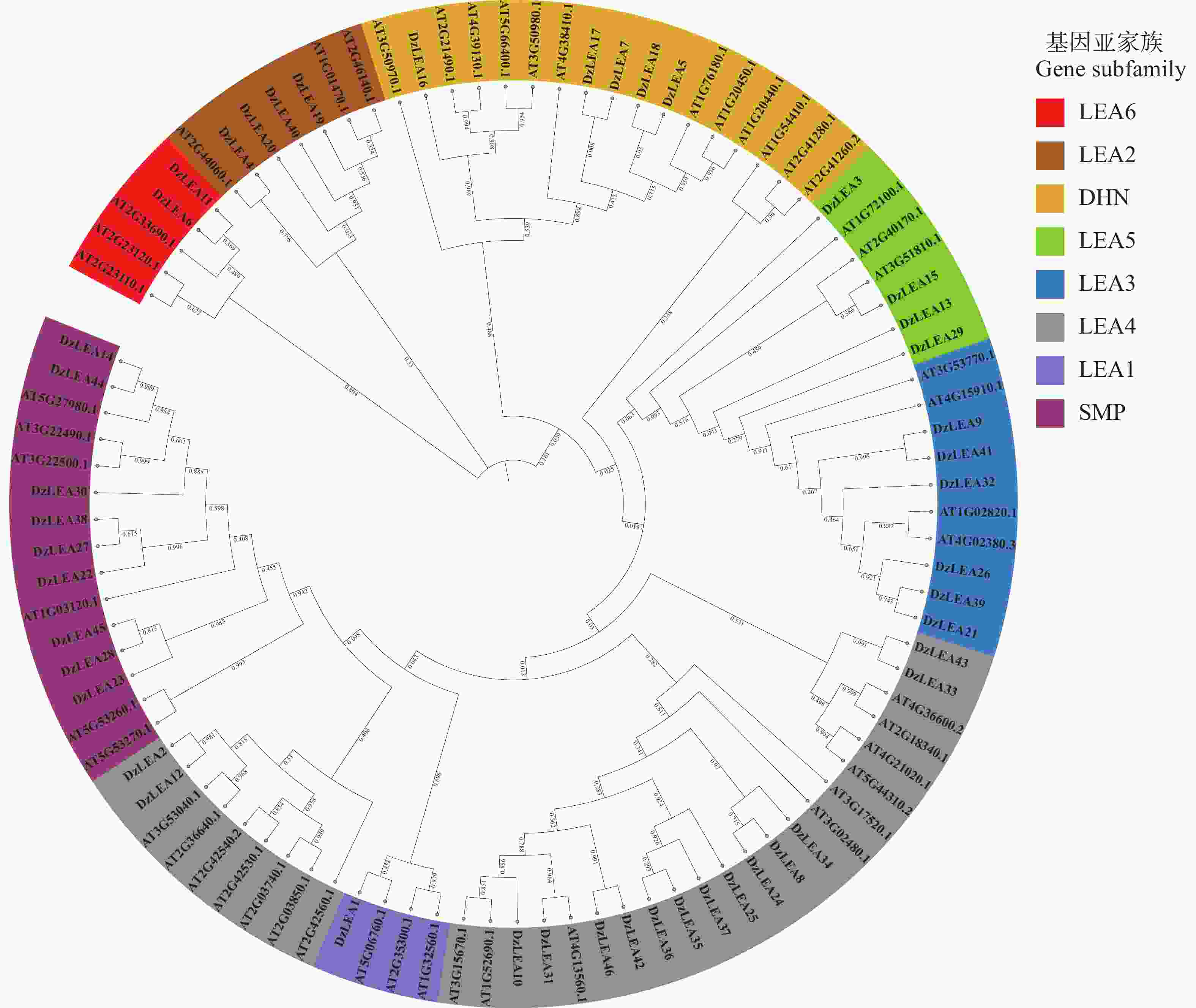
图 2 榴莲和拟南芥LEA蛋白的系统发育树和亚科分类
Figure 2. Phylogenetic tree and subfamily classification of LEA proteins in durian and Arabidopsis
-
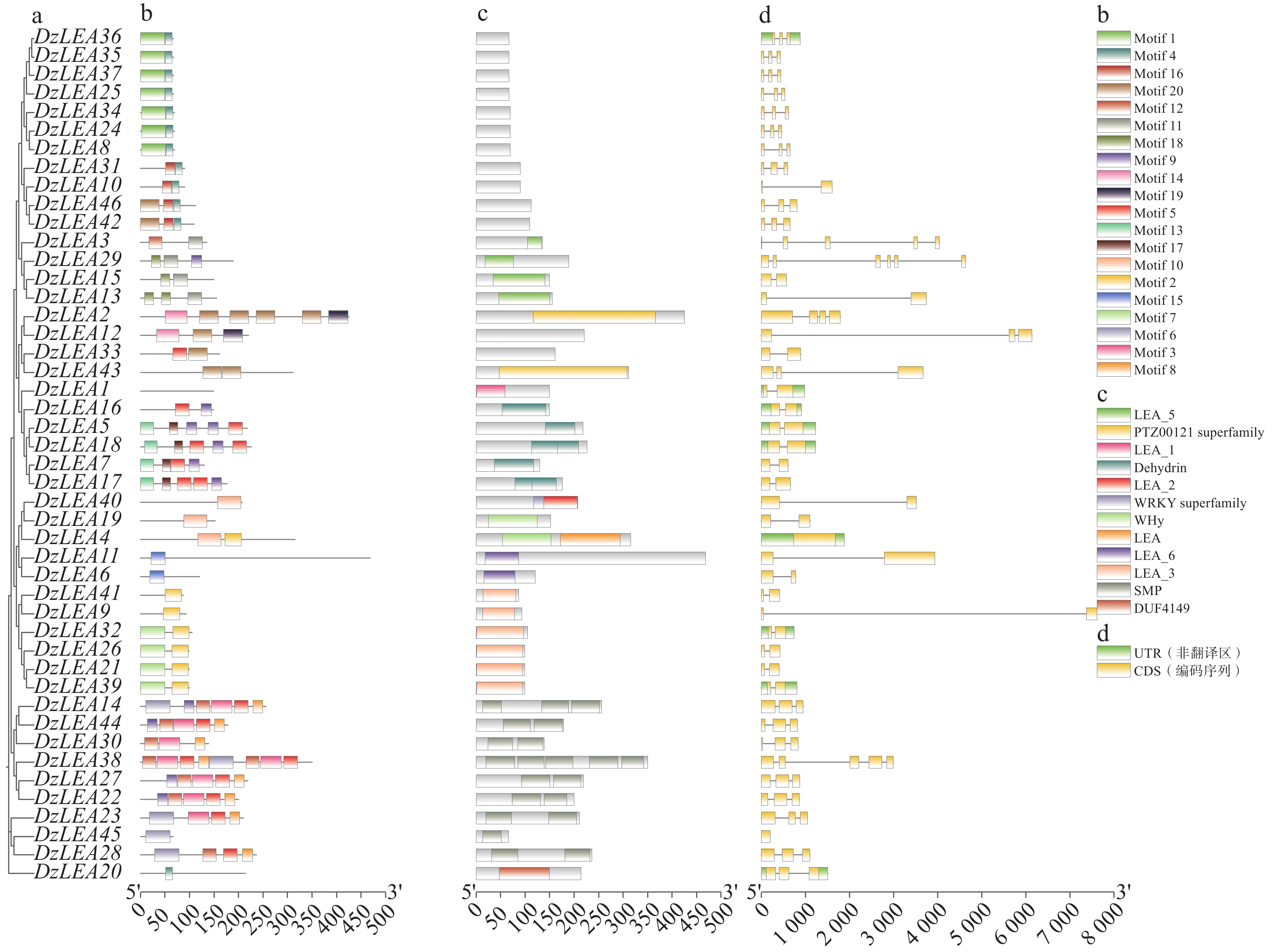
本研究对46个DzLEAs基因的保守基序进行分析,共鉴定出20个motif(motif1~motif20)(图3-b)。结果表明,LEA1亚家族成员不包含任何motif;LEA2亚家族成员除DzLEA20外,其余成员均包含motif10;LEA3亚家族成员主要包含motif2和motif7;LEA4亚家族中,除了4位成员(DzLEA2、DzLEA12、DzLEA33、DzLEA43),其余成员均包含motif4,且绝大多数成员包含motif1,少部分成员包含motif20;LEA5亚家族成员均包含motif11,且除了DzLEA3外,其余成员均包含motif18;LEA6亚家族成员均包含motif15;DHN亚家族成员主要包含motif5、motif9、motif13和motif17;SMP亚家族成员主要包含motif3、motif5、motif6、motif8、motif9和motif12。DzLEAs基因的保守结构域分析显示(图3-c),不同亚家族成员均含有其特定保守结构域。本研究发现,不同亚家族成员的motif类型和分布存在较大差异,而同一亚家族成员之间motif结构较为类似,表明相同亚家族成员之间功能具有相似性。
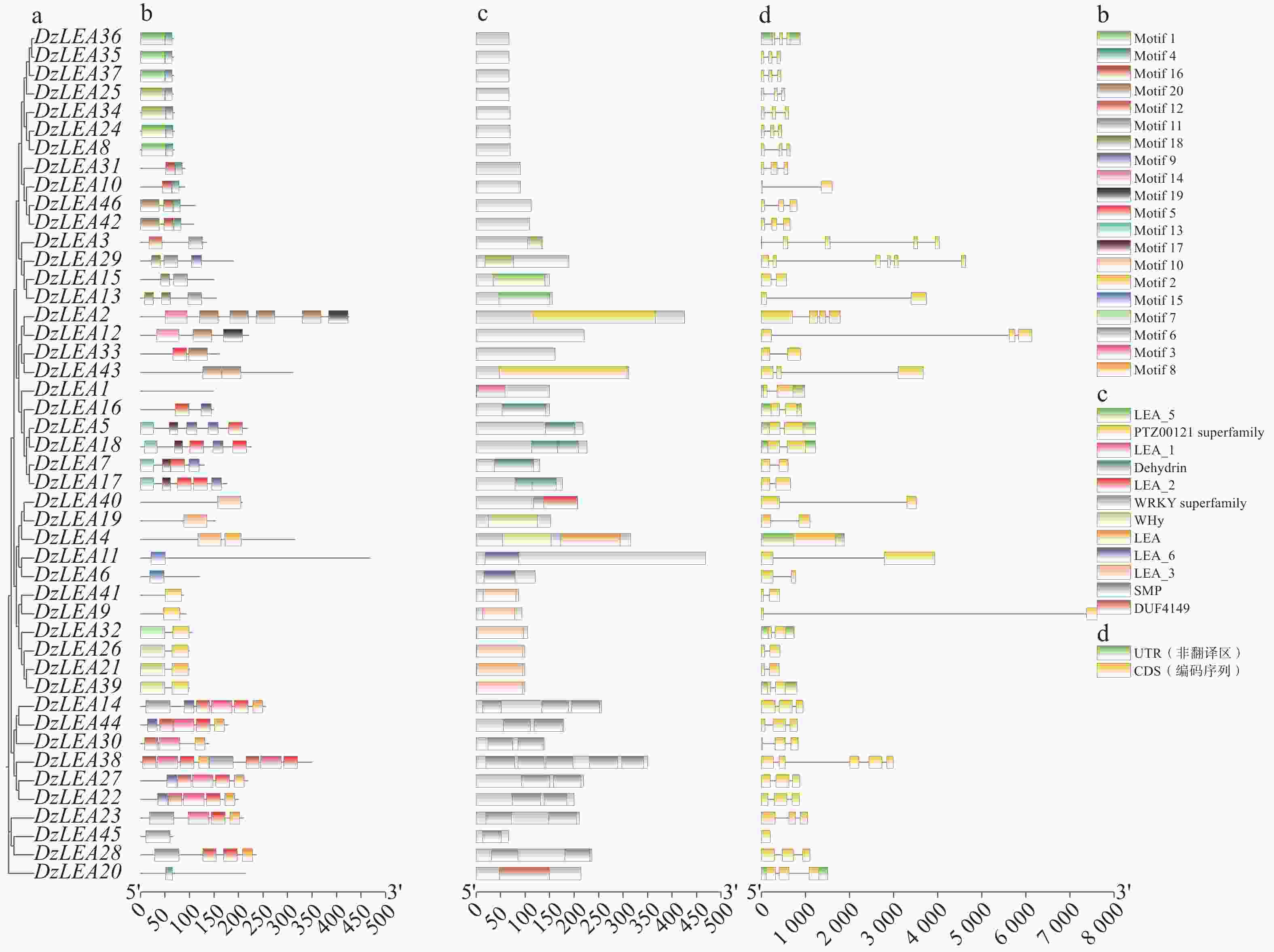
图 3 DzLEAs基因的系统进化树(a)、保守基序(b)、蛋白保守结构域(c)和基因结构(d)
Figure 3. The phylogenetic tree (a), conserved motifs (b), protein domains (c), and gene structures (d)of DzLEA genes.
榴莲LEA基因家族成员基因结构分析显示(图3-d),LEA1亚家族成员包含1个内含子;LEA2亚家族成员含有0~2个内含子,其中DzLEA4基因不含有内含子,DzLEA19和DzLEA40均含有1个内含子,DzLEA20含有2个内含子;LEA3亚家族成员均含有1个内含子;LEA4亚家族成员除3个成员外(DzLEA2、DzLEA10、DzLEA33)均含有2个内含子;LEA5亚家族成员中,DzLEA29含有5个内含子,DzLEA3含有4个内含子,其余2个成员(DzLEA13、DzLEA15)含有1个内含子;DHN亚家族成员均含有1个内含子;SMP亚家族成员含有0~4个内含子,其中DzLEA45不含有内含子,DzLEA38含有4个内含子,其余成员均含有2个内含子。在同一亚家族成员中,具有相似的基因结构,并且进化关系密切的成员之间表现出更为相似的外显子-内含子结构、外显子数量和基因长度。
-
对46个榴莲LEA基因进行了顺式作用元件分析(图4),结果表明,发现了5个与光响应相关的元件,分别为3-AF1 binding site(5个成员)、ACE(9个成员)、G-box(37个成员)、GT1-motif(28个成员)和Sp1(8个成员);3个与赤霉素响应相关的元件,分别为GARE-motif(10个成员)、P-box(13个成员)和TATC-box(7个成员);茉莉酸甲酯响应元件CGTCA-motif(36个成员)和TGACG-motif(37个成员);生长素响应元件AuxRR-core(4个成员)和TGA-element(11个成员);脱落酸响应元件ABRE(38个成员);水杨酸响应元件TCA-element(25个成员);与低温响应的元件LTR(16个成员);参与防御和应激反应的元件TC-rich repeats(15个成员);参与干旱响应的元件MBS(22个成员)。除此之外,还发现了与厌氧诱导相关的元件(ARE)、与分生组织表达相关的元件(CAT-box)、MYB结合位点(MRE、CCAAT-box)、昼夜节律控制相关的元件(circadian)、胚乳表达相关的元件(GCN4_motif)、玉米醇溶蛋白代谢调控相关的元件(O2-site)、种子特异性调控元件(RY-element)和伤口响应元件(WUN-motif)。这些结果表明,DzLEAs基因可能参与多种植物生长发育过程,并且在逆境胁迫中发挥重要作用。

图 4 DzLEAs基因启动子上的顺式调控元件分布情况
Figure 4. Distribution of cis-regulatory elements in the promoters of DzLEA genes.
-
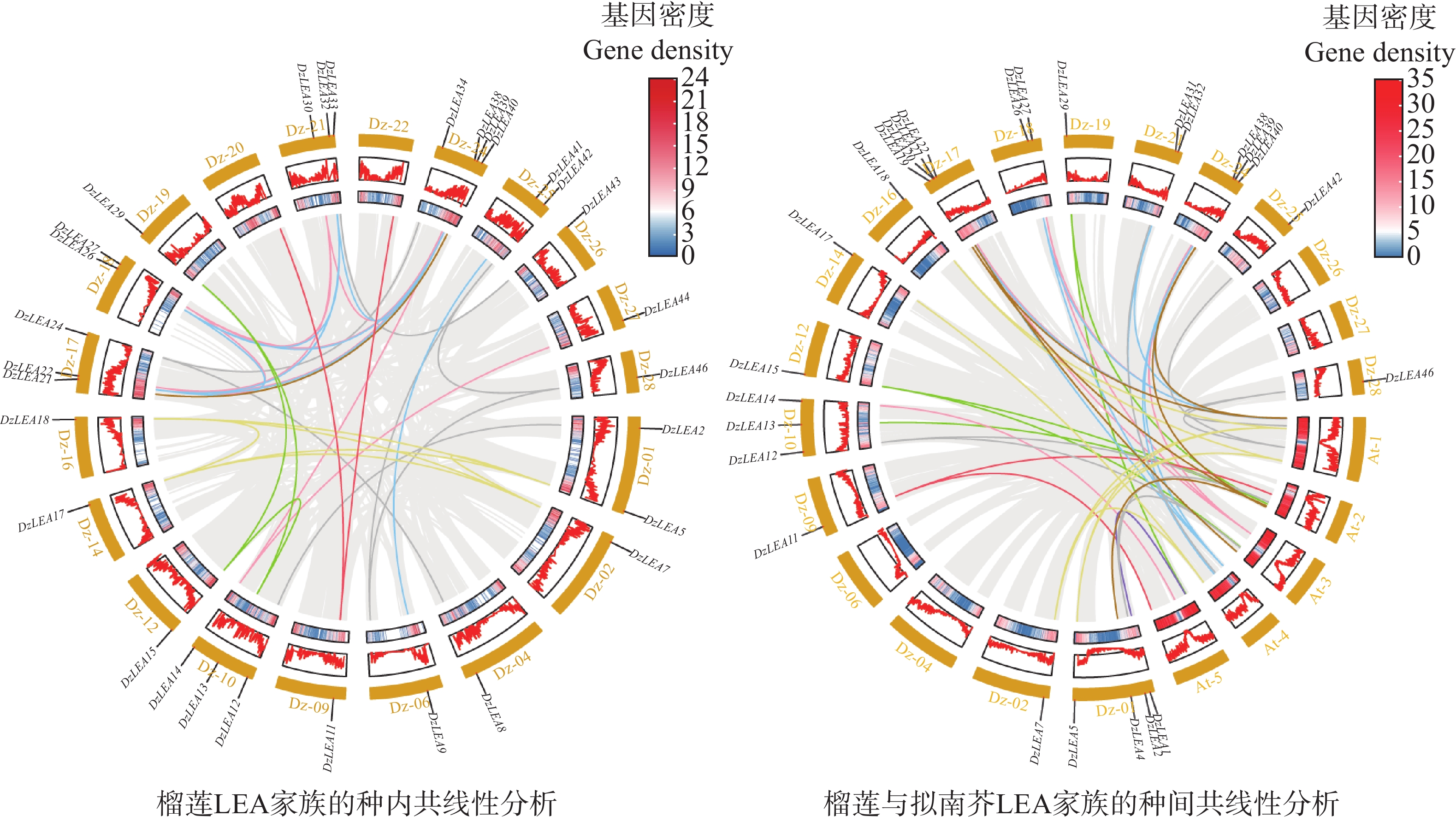
对DzLEA家族成员在染色体上的位置进行分析(图1),46个DzLEA家族成员分布在18条染色体上,且呈现不均匀分布。其中,17号和24号染色体均含有7个DzLEAs基因;1号和21号染色体上分别含有5个和4个DzLEAs基因;10号和18号染色体上均含有3个DzLEAs基因;2号、6号、14号、25号和27号染色体均含有2个DzLEAs基因;4号、9号、12号、16号、19号、26号和28号染色体各含有1个DzLEAs基因。并且,通过染色体定位图显示,DzLEA22和DzLEA23、DzLEA27和DzLEA28、DzLEA36和DzLEA37、DzLEA44和DzLEA45,这四组基因距离相近,推测以上成员存在遗传连锁和多种复制事件的可能性。
同时对DzLEAs基因进行了物种内共线性分析(图5),共发现28组共线性基因对,结果显示榴莲LEA基因家族至少发生了19次片段复制事件,但并未发现串联复制事件,这导致榴莲LEA家族数量上的扩增。同时,发现部分DzLEAs基因与多个基因存在共线性关系,如DzLEA5和DzLEA7、DzLEA17、DzLEA18;DzLEA21和DzLEA26、DzLEA32、DzLEA39;DzLEA22和DzLEA27、 DzLEA30、DzLEA38,这些基因分别聚类于DHN、LEA3和SMP亚家族,说明其在功能与结构上可能具有相似性。
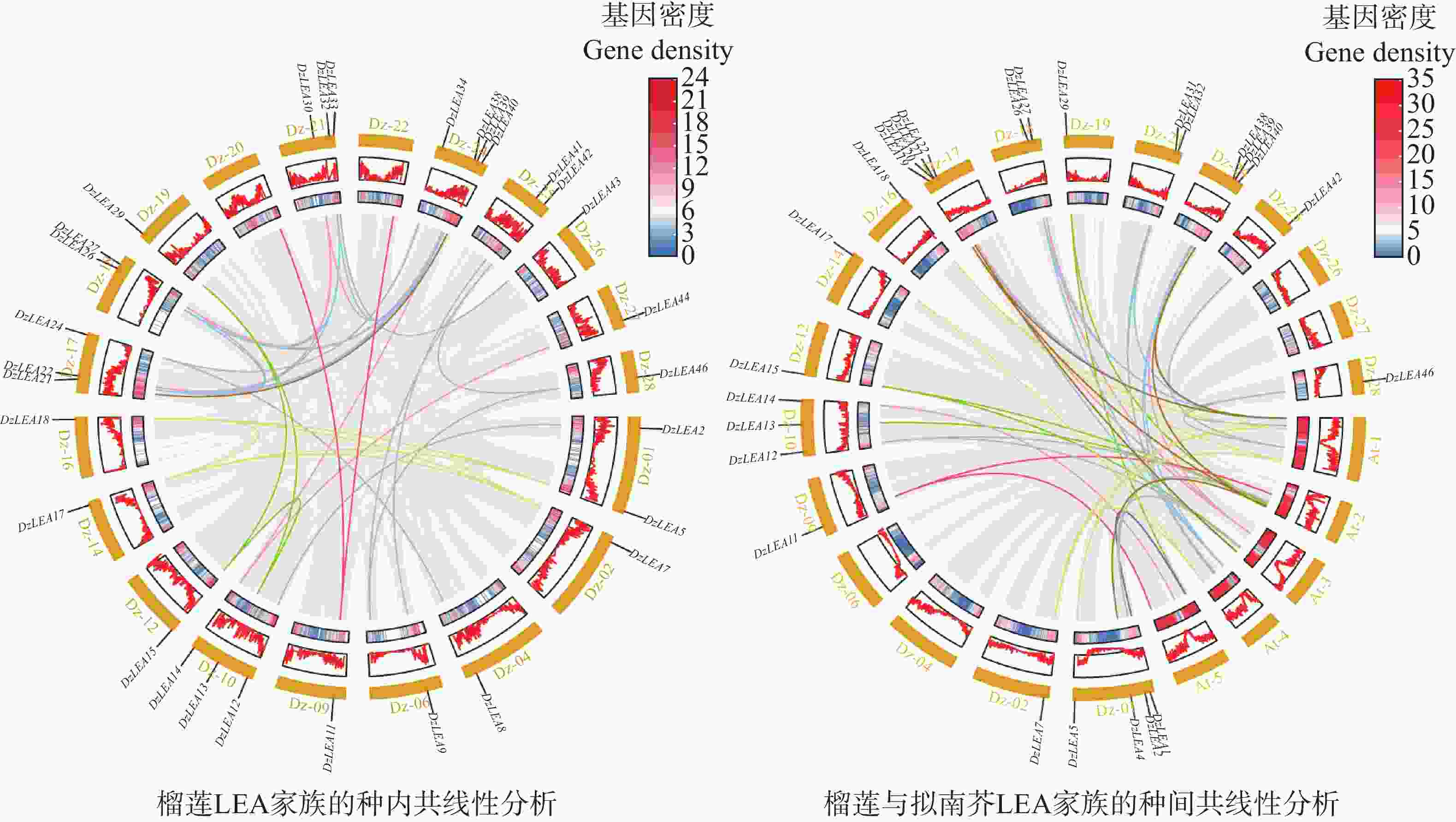
图 5 榴莲LEA基因家族的种内(左)与种间(右)共线性分析
Figure 5. Intra-species (left)and inter-species (right)synteny analysis of the LEA gene family in durian.
另外,对榴莲和拟南芥LEA基因家族进行了物种间的共线性分析(图5),共鉴定出46对同源基因,具体表现为:LEA1亚家族中,DzLEA1与AT5G06760.1;LEA2亚家族中,DzLEA4与AT2G44060.1,DzLEA19与AT1G01470.1、AT2G46140.1,DzLEA20与AT3G62580.1,DzLEA40与AT1G01470.1、AT2G46140.1;LEA3亚家族中,DzLEA21与AT4G02380.3、AT1G02820.1,DzLEA26与AT1G02820.1、AT4G02380.3,DzLEA32与AT1G02820.1、AT4G15910.1,DzLEA39与AT1G02820.1、AT4G15910.1、AT4G02380.3;LEA4亚家族中,DzLEA2与AT2G36640.1、AT3G53040.1,DzLEA12与AT2G36640.1、AT3G53040.1,DzLEA31与AT4G13560.2,DzLEA42与AT1G52680.1,DzLEA46与AT1G15415.1、AT1G52680.1;LEA5亚家族中,DzLEA13与AT2G40170.1,DzLEA15与AT2G40170.1、AT3G51810.1,DzLEA29与AT2G40170.1、AT5G05560.2;LEA6亚家族中,DzLEA11与AT2G23110.1、AT5G67350.1;DHN亚家族中,DzLEA5与AT1G76180.1、AT1G20440.1,DzLEA7与AT1G76180.1、AT1G20440.1、AT4G38410.1,DzLEA17与AT4G38410.1,DzLEA18与AT1G76180.1、AT1G20440.1;SMP亚家族中,DzLEA14与AT5G27980.1,DzLEA22与AT1G03120.1、AT3G22490.1,DzLEA27与AT1G03120.1、AT3G22490.1,DzLEA38与AT1G03120.1、AT3G22490.1。这表明这些基因可能具有某些特定功能和进化的保守性。
-
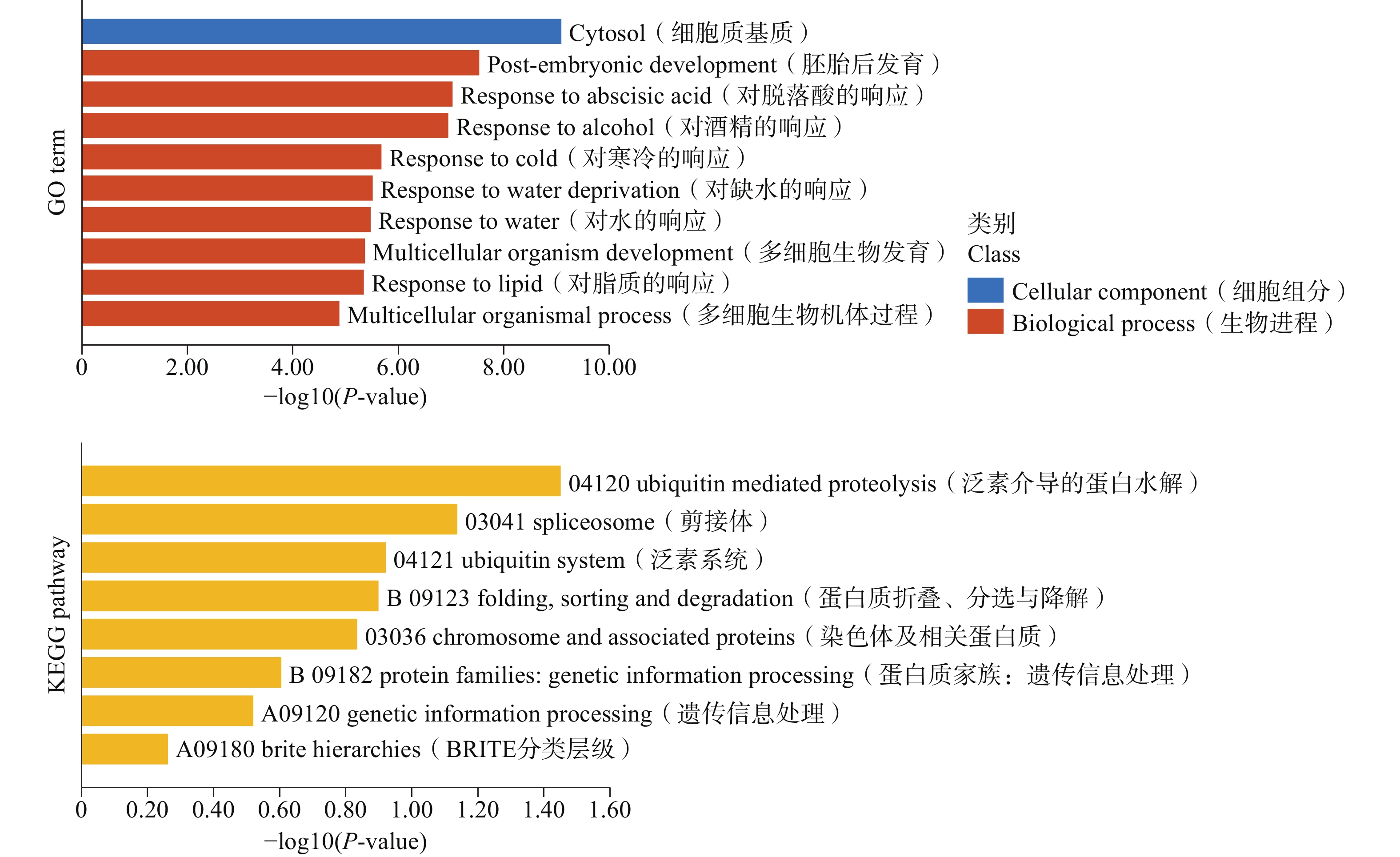
为了进一步研究LEA基因的具体功能,对榴莲DzLEAs基因进行基于超几何分布的GO和KEGG分析(图6)。GO富集分析结果显示,在生物进程类别中,6个LEA基因家族成员均在“对寒冷的反应”以及“对缺水的反应”显著富集,分别是DzLEA5、 DzLEA16、 DzLEA18、 DzLEA21、 DzLEA26、 DzLEA39。
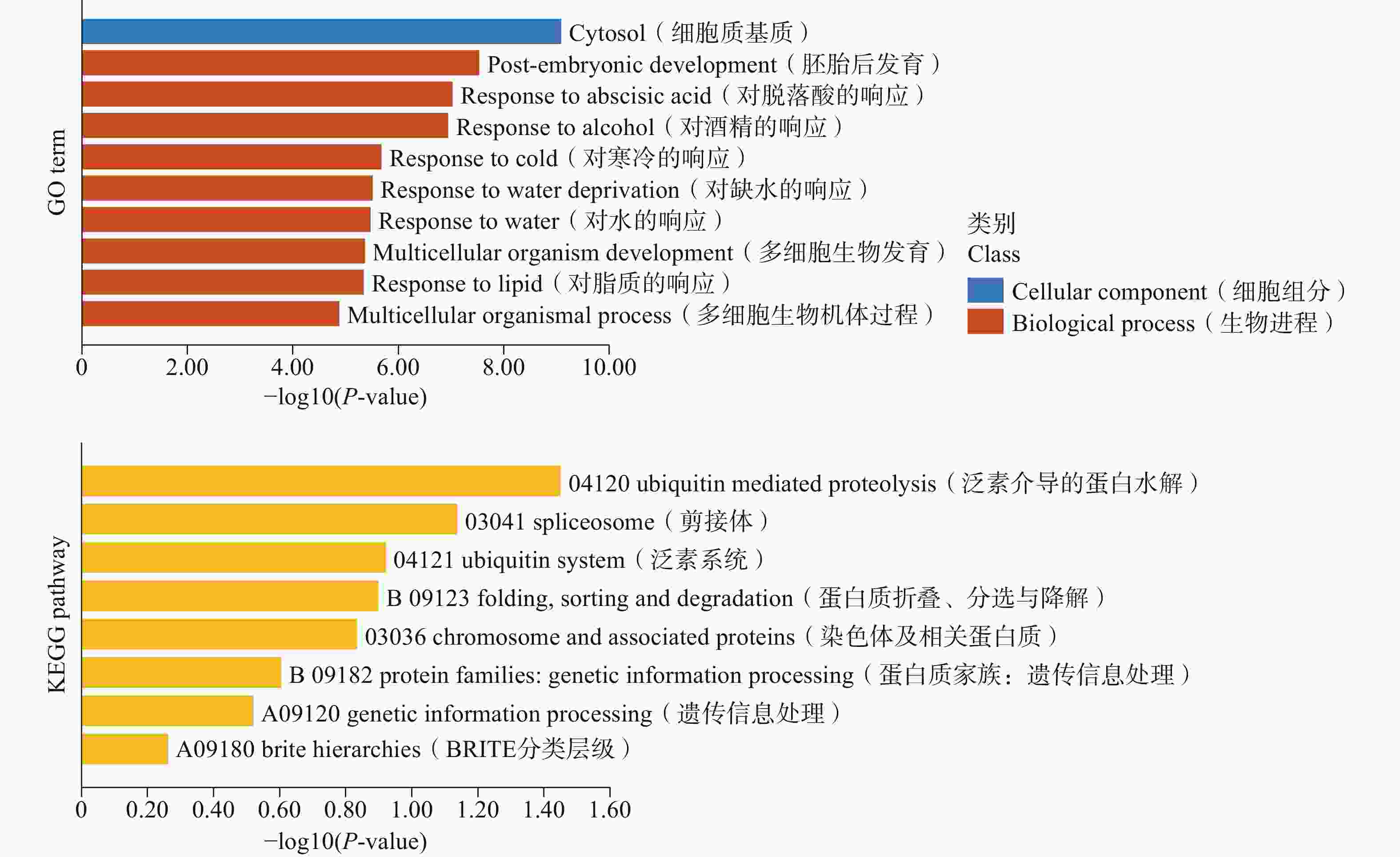
图 6 榴莲LEA基因家族的GO分析(上侧)和KEGG(下侧)分析条状图
Figure 6. Bar graphs of GO (top)and KEGG (bottom)enrichment analysis for the durian LEA gene family.
在KEGG富集分析中,LEA基因家族成员主要富集在泛素介导的蛋白水解,剪接体、泛素系统,蛋白质的折叠、分选与降解,染色体及相关蛋白质功能通路上。
-
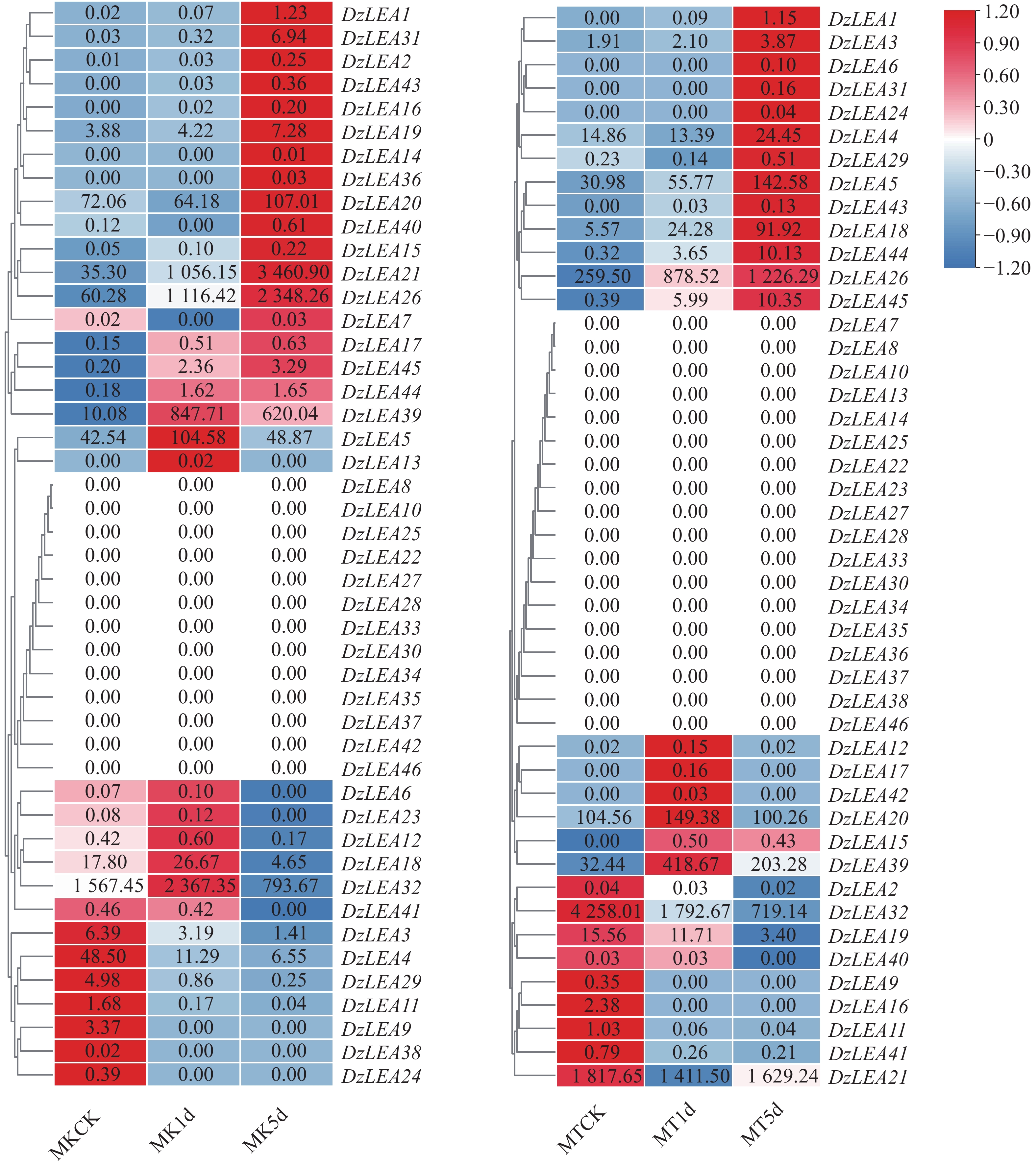
转录组学分析表明(图7),部分DzLEAs基因对低温胁迫具有较高敏感性,具体表现为,与常温下榴莲对照相比10℃低温胁迫1d条件下,猫山王榴莲品种有7个基因显著上调表达,3个基因显著下调表达,这其中两个基因(DzLEA21和DzLEA26)随着低温胁迫处理时间的增长,呈现持续显著上调表达,另有两个基因(DzLEA11和DzLEA29)随着低温胁迫处理时间的增长,呈现持续显著下调表达;与常温下榴莲对照相比10℃低温胁迫1d条件下,金枕榴莲品种有5个基因显著上调表达,3个基因显著下调表达,这其中3个基因(DzLEA18、DzLEA26和DzLEA44)随着低温胁迫处理时间的增长,呈现持续显著上调表达,另有2个基因(DzLEA11和DzLEA32)随着低温胁迫处理时间的增长,呈现持续显著下调表达。
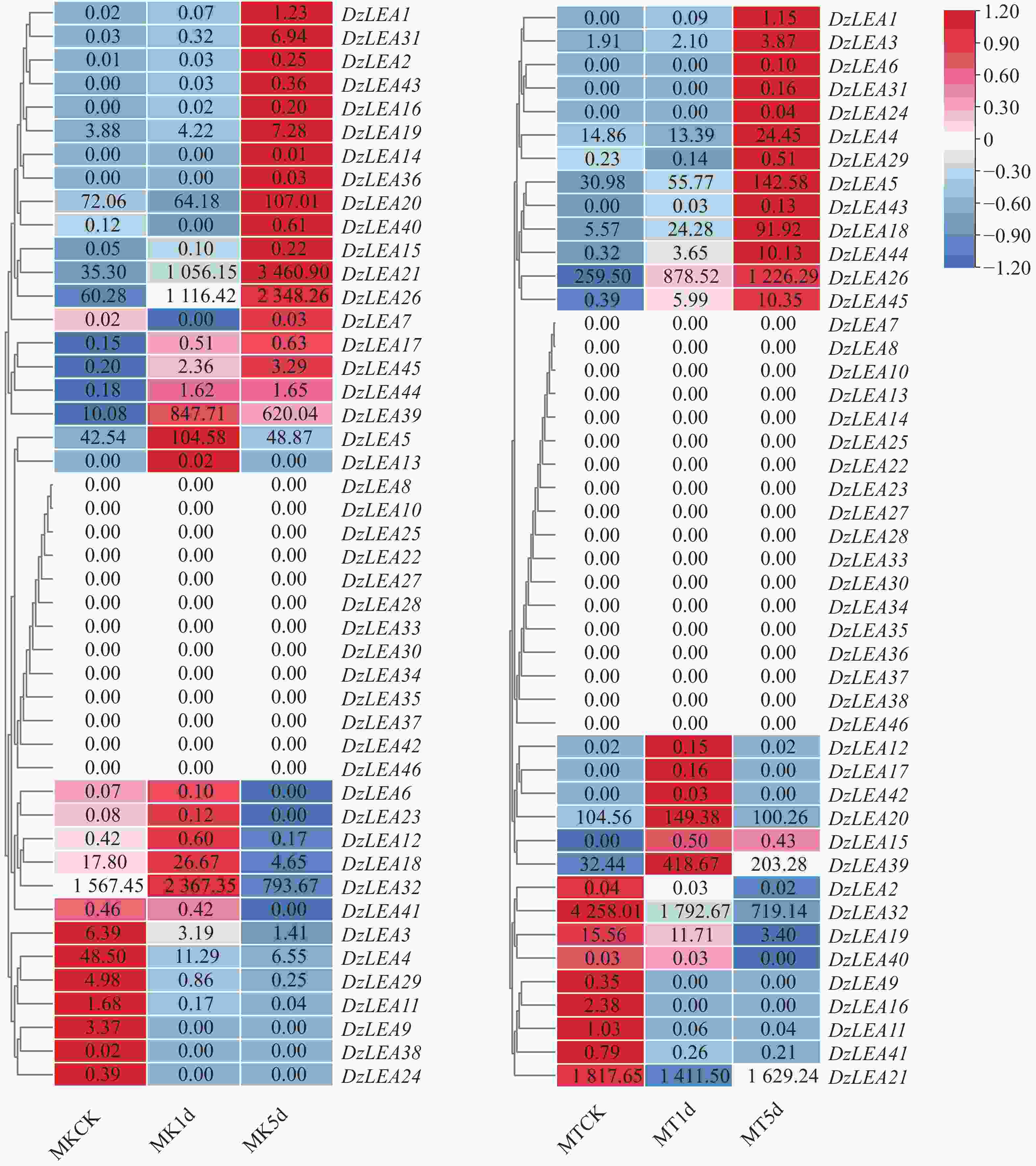
图 7 猫山王(左)与金枕(右)榴莲低温胁迫下LEA基因家族成员转录组表达情况
Figure 7. Transcriptomic expression profiles of LEA gene family members in durian cultivars Musang King (left)and Golden Pillow (right)under cold stress.
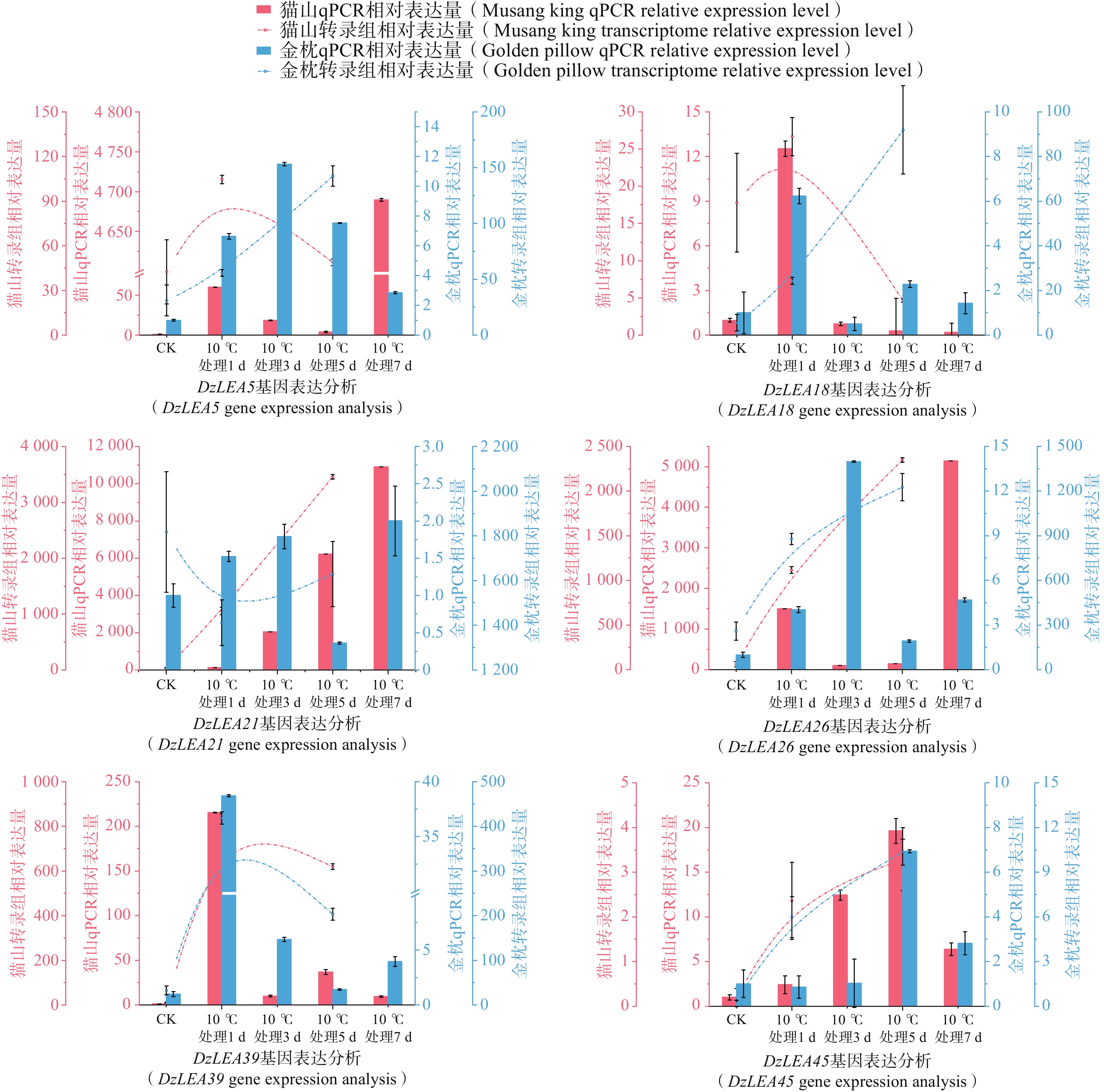
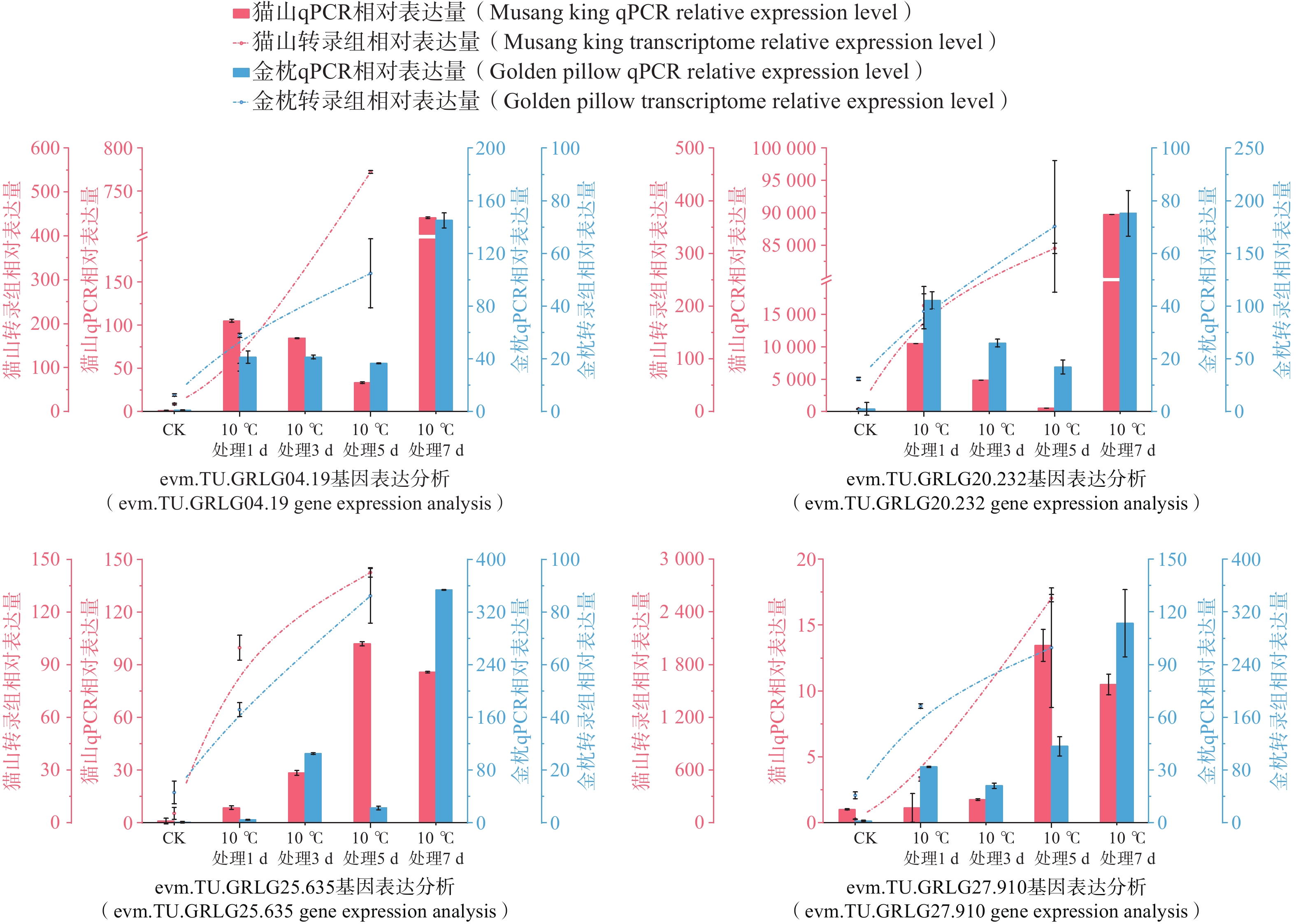
为了进一步分析DzLEAs基因在低温胁迫中的作用,基于低温处理下的榴莲叶片转录组数据,挑选出6个低温处理(10℃)下表达量升高的DzLEAs基因(图8)和4个非DzLEAs基因(图9),通过qRT-PCR测定榴莲叶片在四个低温处理时期下的表达量变化。结果显示,DzLEAs基因均在10℃低温处理下呈现不同程度上调表达,其中不同品种的DzLEA18和DzLEA39均在10℃低温处理1d表达量最高,而后表达量逐渐降低;猫山王品种DzLEA5表达量随低温处理时间增加而逐步上升,并在10℃低温处理7d表达量最高,金枕品种DzLEA5表达量呈现先升高而后降低趋势,在10℃低温处理3 d表达量最高;不同品种的DzLEA21表达量均随低温处理时间增加而逐步上升,且在10℃低温处理7 d表达量最高;猫山王品种DzLEA26表达量随低温处理时间增加而逐步上升,并在10℃低温处理7 d表达量最高,金枕品种DzLEA5表达量呈现先升高而后降低趋势,在10℃低温处理3 d表达量最高;不同品种的DzLEA45表达量均呈现先升高而后降低趋势,且在10℃低温处理5 d表达量最高。非DzLEAs基因在10℃低温处理下表达模式有所不同,evm.model.GRLG04.19和evm.model.GRLG20.232表达模式类似,在不同品种中均呈现先升高后降低而后再升高的趋势,且均在10℃低温处理7 d表达量最高;evm.model.GRLG25.635和evm.model.GRLG27.910表达模式类似,猫山王品种中呈现先升高而后降低的趋势,在10℃低温处理5 d表达量最高,金枕品种中呈现逐步升高的趋势,在10℃低温处理7 d表达量最高。综上,这些基因的表达模式在不同品种之间相近,且与转录组表达结果基本一致,呈现较为明显的上调趋势,表明这些基因可能在榴莲响应低温胁迫过程中发挥了关键作用。
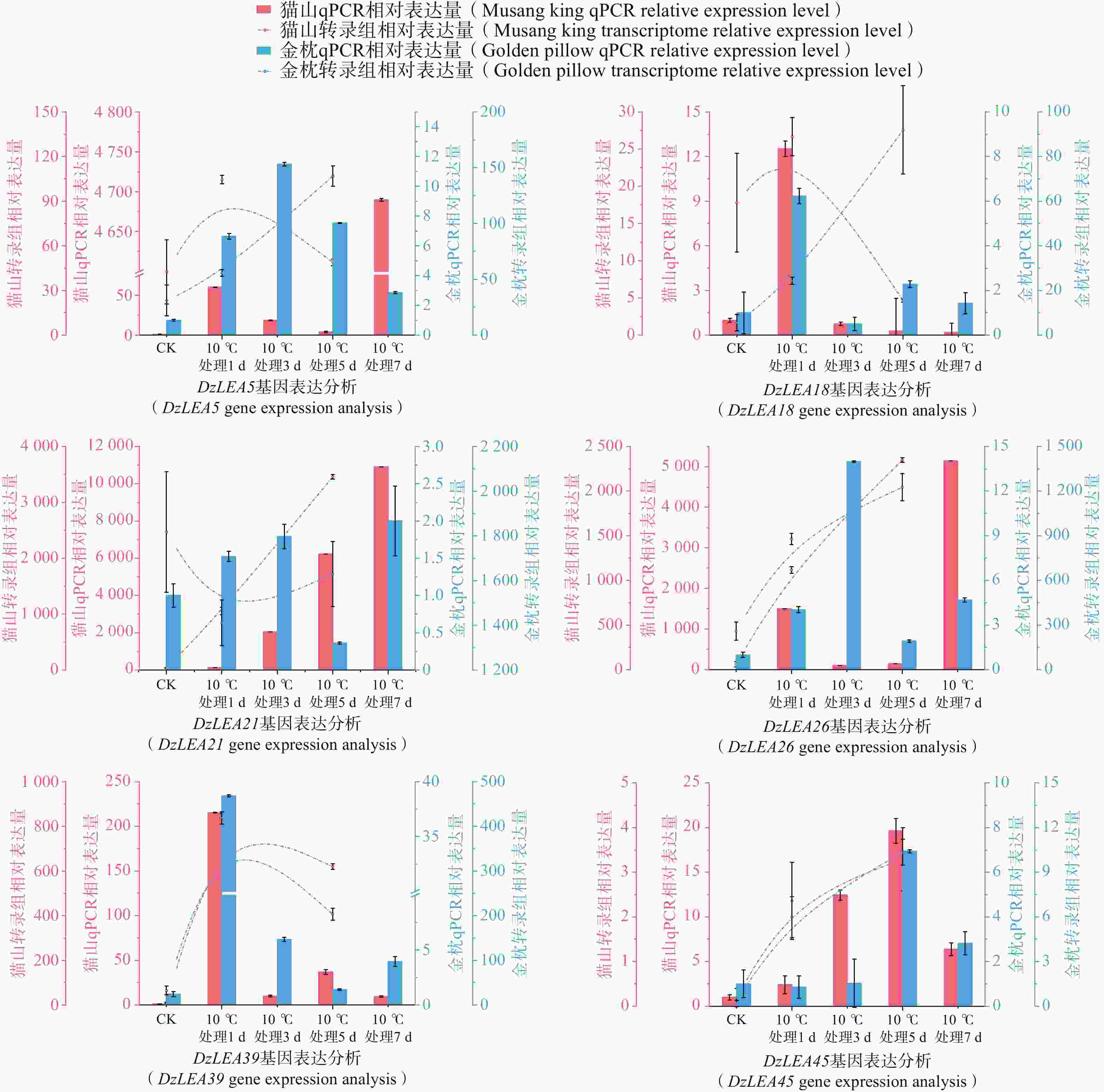
图 8 2个榴莲品种低温胁迫下DzLEAs基因相对表达量
Figure 8. Relative expression levels of DzLEA genes in the two durian cultivars under cold stress.
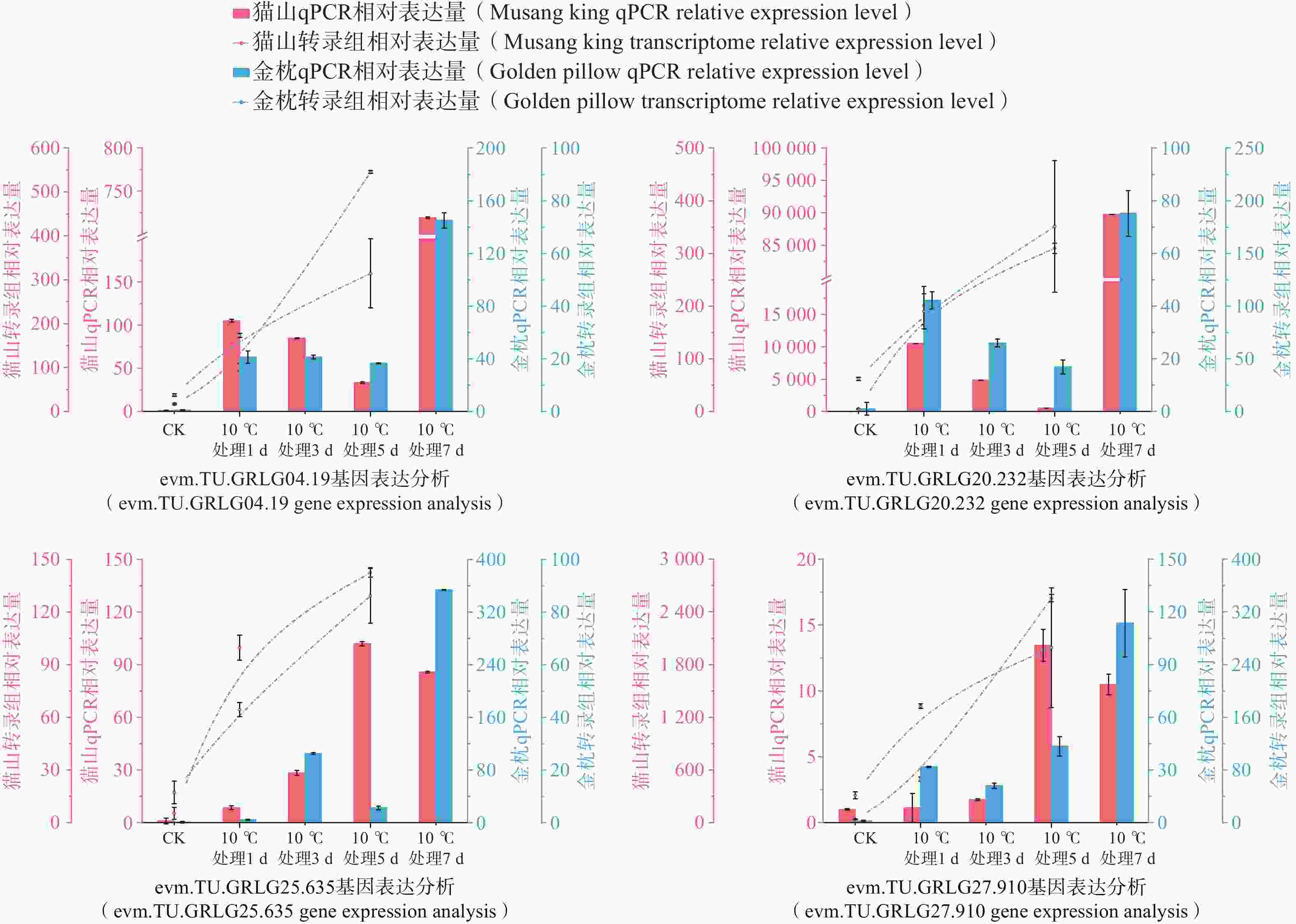
图 9 2个榴莲品种低温胁迫下非DzLEAs基因相对表达量
Figure 9. Relative expression levels of non-DzLEA genes in the two durian cultivars under cold stress.
-
本研究对榴莲LEA基因家族进行了鉴定和生物信息学分析,共鉴定了46个DzLEAs基因,分为8个亚族,分布在18条染色体上。榴莲LEA基因家族成员数量相较于茶树(48个)[9]、甜橙(72个)[18]和拟南芥(51个)[6]经历了一定程度的收缩,相较于木薯(26个)[19]、香蕉(23个)[20]和石斛(17个)[21]经历了一定程度的扩张。通过对其进行理化性质分析,结果显示,大多数DzLEA蛋白为亲水性蛋白(44个),这与前人的研究一致,高度亲水性有利植物在失水胁迫下高效捕获水分,从而保护细胞膜和蛋白质[22]。且有研究表明,大多数LEA蛋白被预测为固有无序蛋白,且在干燥过程中形成两亲性α螺旋,这些特性能够使LEA蛋白在冷冻胁迫下阻止保护酶聚集和失活,并且提高膜脂的稳定性[23]。对榴莲LEA基因家族的结构分析显示,2个成员缺乏内含子、20个成员只有1个内含子、20个成员有两个内含子、4个成员有3个或3个以上内含子,较少的内含子结构缩短了从转录到翻译的防御胁迫时间,这有利于植物对环境胁迫引起的刺激迅速作出反应[24]。
在研究过程中发现,榴莲在10℃低温胁迫下,短时间(1d)就会发生大面积正常颜色叶片脱落情况。我们推测这与其长期进化过程中的生存策略有关,其生存策略是“快速生长、高效繁殖”,低温环境下,榴莲会迅速响应并引起脱落酸和乙烯水平增高,丢弃耗能器官(叶片),将资源集中于维持主干生命,等待环境回暖。榴莲叶片具有厚角质层、厚蜡质层和高糖代谢特征,且油脂成分主要为结构性脂质,不饱和脂肪酸比例较低,相变温度较高。其结构和成分特征与抵御热带地区强光、暴雨、高温和病虫害相适应,但表现出极强的冷敏感性,其厚角质层和蜡质层使其在面对低温时不易回温,易发生光合系统受损和膜脂过氧化。
对榴莲LEA基因家族进行生物信息学分析过程中,发现4个基因(DzLEA5、DzLEA21、DzLEA26和DzLEA39)启动子上均存在与低温响应相关的元件LTR,且在GO分析中,在生物进程类别中,该4个基因均与“对寒冷的反应”以及“对缺水的反应”有显著关联。将该四个基因与转录组中低温胁迫下显著上调表达的两个基因(DzLEA18和DzLEA45)共同进行表达模式分析。发现这6个基因均在低温胁迫条件下呈现不同程度的上调表达,表明其可作为后续研究榴莲低温胁迫响应的分子机制及榴莲抗寒品种选育的候选基因。针对这些基因,后续可通过在模式植物(拟南芥)异源过表达,结合表型和生理指标数据进行进一步验证;并通过多品种的重测序,发掘其单核苷酸多态性(SNP)或插入/缺失(InDel)变异,开发与低温响应性状关联的分子标记。这些标记可用于榴莲种质资源的抗寒性鉴定,加速抗寒品种的筛选与选育进程。
Identification of the durian LEA gene family and expression analysis under low temperature stress
-
摘要: 为了分析榴莲(Durio zibethinus)LEA基因家族的特征和潜在功能,探究DzLEAs基因在榴莲抵御低温胁迫中的作用,通过生物信息学方法对榴莲LEA基因家族成员进行全基因组鉴定,并对其理化性质、系统进化关系、染色体定位、亚细胞定位、共线性、启动子顺式作用元件、GO和KEGG富集,以及在低温胁迫处理后的表达情况进行分析。结果表明,从榴莲基因组中共鉴定到46个基因家族成员,分为LEA1、LEA2、LEA3、LEA4、LEA5、LEA6、DHN和SMP8个亚族,分布于18条染色体上。家族成员的编码蛋白含有65~467个氨基酸;等电点介于4.33~9.79之间;大多数LEA蛋白为小分子亲水性蛋白,在细胞核、细胞质、线粒体、叶绿体、质膜和细胞外均有分布。基因结构分析表明,91%DzLEAs基因具有较少内含子(<3)。在16个DzLEAs基因的启动子区存在与低温胁迫相关的元件(LTR)。榴莲种内以及榴莲与拟南芥种间共线性分析分别发现28和46对同源基因。GO富集分析表明,有6个LEA基因家族成员均在“对寒冷的反应”和“对缺水的反应”显著富集。qRT-PCR验证结果显示,6个DzLEAs基因和4个非DzLEAs基因表达趋势与转录组结果基本一致,均响应低温胁迫呈现不同程度上调表达。研究结果为进一步探讨榴莲DzLEAs基因响应低温胁迫的分子机制提供参考。
-
关键词:
- 榴莲 /
- 胚胎发育晚期丰富蛋白基因家族 /
- 生物信息学 /
- 低温胁迫
Abstract: To elucidate the characteristics, potential functions, and role of the late embryogenesis abundant (LEA) gene family in durian (Durio zibethinus) (DzLEAs) in response to low-temperature stress, a genome-wide identification of the LEA gene family in durian was conducted using bioinformatics approaches. Comprehensive analyses were performed, including assessments of physicochemical properties, phylogenetic relationships, chromosomal localization, subcellular localization, synteny, promoter cis-acting elements, Gene Ontology (GO) and Kyoto Encyclopedia of Genes and Genomes (KEGG) enrichment, as well as expression profiles under cold stress. A total of 46 DzLEAs members were identified and classified into eight subgroups: LEA1, LEA2, LEA3, LEA4, LEA5, LEA6, DHN, and SMP. These genes were distributed across 18 chromosomes. The encoded proteins ranged from 65 to 467 amino acids in length, with isoelectric points (pI) varying between 4.33 and 9.79. Most DzLEA proteins were predicted to be small hydrophilic molecules localized to multiple cellular compartments, including the nucleus, cytoplasm, mitochondria, chloroplast, plasma membrane, and extracellular space. Gene structure analysis revealed that 91% of DzLEAs possessed a relatively low number of introns (<3). Promoter analysis identified low-temperature-responsive cis-acting elements (LTRs) in 16 DzLEAs genes. Synteny analysis detected 28 and 46 pairs of homologous genes within the durian genome and between durian and Arabidopsis thaliana, respectively. GO enrichment analysis indicated that six DzLEAs were significantly enriched in the biological processes, "response to cold" and "response to water stress." Quantitative real-time PCR (qRT-PCR) validation showed that the expression patterns of six selected DzLEAs and four non-DzLEA genes were largely consistent with the transcriptomic data, with all genes being up-regulated to varying degrees under cold stress. These findings provide a valuable foundation for further exploration of the molecular mechanisms underlying the response of DzLEAs genes to low-temperature stress in durian.-
Key words:
- durian /
- late embryogenesis abundant protein /
- bioinformatics /
- low-temperature stress
-
图 2 榴莲和拟南芥LEA蛋白的系统发育树和亚科分类
Fig. 2 Phylogenetic tree and subfamily classification of LEA proteins in durian and Arabidopsis
图 3 DzLEAs基因的系统进化树(a)、保守基序(b)、蛋白保守结构域(c)和基因结构(d)
Fig. 3 The phylogenetic tree (a), conserved motifs (b), protein domains (c), and gene structures (d)of DzLEA genes.
图 4 DzLEAs基因启动子上的顺式调控元件分布情况
Fig. 4 Distribution of cis-regulatory elements in the promoters of DzLEA genes.
图 5 榴莲LEA基因家族的种内(左)与种间(右)共线性分析
Fig. 5 Intra-species (left)and inter-species (right)synteny analysis of the LEA gene family in durian.
图 6 榴莲LEA基因家族的GO分析(上侧)和KEGG(下侧)分析条状图
Fig. 6 Bar graphs of GO (top)and KEGG (bottom)enrichment analysis for the durian LEA gene family.
图 7 猫山王(左)与金枕(右)榴莲低温胁迫下LEA基因家族成员转录组表达情况
Fig. 7 Transcriptomic expression profiles of LEA gene family members in durian cultivars Musang King (left)and Golden Pillow (right)under cold stress.
图 8 2个榴莲品种低温胁迫下DzLEAs基因相对表达量
Fig. 8 Relative expression levels of DzLEA genes in the two durian cultivars under cold stress.
图 9 2个榴莲品种低温胁迫下非DzLEAs基因相对表达量
Fig. 9 Relative expression levels of non-DzLEA genes in the two durian cultivars under cold stress.
表 1 实时荧光定量PCR引物及其碱基序列
Table 1 Sequences of primers for quantitative real-time PCR
基因名称
Gene基因编号
Sequence ID正向引物序列(5′−3′)
Forward primer sequence(5′−3′)反向引物序列(5′−3′)
Reverse primer sequence(5′−3′)DzLEA5 evm.model.GRLG01.2002 ACCCCATGAAGGTGAGACGA TGGGAAGCGCTCTCCTTTTC DzLEA18 evm.model.GRLG16.937 GCCAAGGACCGAGGGTTG TCCTTGCGCTCCTCCTCT DzLEA21 evm.model.GRLG17.729 GGAGCAGCAAGGAACGCT CTCGGCCACGTCGATCTC DzLEA26 evm.model.GRLG18.174 GGATATGCTGCGGGATCACA GTCCGGAACCCATGAAACCT DzLEA39 evm.model.GRLG24.892 CGGCAACAAACTCTGCGG CTCGGCCACGTCGATCTC DzLEA45 evm.model.GRLG27.783 GTGGCGCCTAAGGATGCA TGCATGGCAGCAGATTCCA Dz04.19 evm.model.GRLG04.19 CGACGAGCTCAAGGAGGC ATGGCACGAGCAGCGAAT Dz27.910 evm.model.GRLG27.910 GCAAGTAGCGGCTGTCCA GCACTTCAACGGTCCCGA Dz25.635 evm.model.GRLG25.635 TGCGGCTGCCATTGATGA GCACTGTCTGCTGCTCAA Dz20.232 evm.model.GRLG20.232 AGCATCAAGACACGCAAGGA ATGTCGTTTGGCCGCCAT Dz4GAPDH09.1422 evm.model.GRLG09.1422 TGGGCTGAGACAGGAGCT CCACAACAAACATGGGGGC  下载: 导出CSV
下载: 导出CSV
表 2 榴莲DzLEA基因家族特征分析
Table 2 Features of DzLEA gene family
基因名称
Gene基因编号
Sequence ID基因
家族
Gene
family氨基酸长度
Amino acid length分子质量
Molecular weight理论等电点
Theoretical pI不稳定系数
Instability Index脂肪族系数
Aliphatic Index亲水性
平均系数
Grand
average of hydropathicity亚细胞定位
Subcellular localizationDzLEA1 evm.model.
GRLG01.168LEA1 148 15 343.01 8.07 32.05 52.97 −0.8 细胞核 DzLEA2 evm.model.
GRLG01.459LEA4 424 47 446.41 5.44 37.00 46.98 −1.29 细胞核 DzLEA3 evm.model.
GRLG01.897LEA5 134 15 000.93 9.79 44.68 70.6 −0.657 细胞核 DzLEA4 evm.model.
GRLG01.981LEA2 314 34 879.84 4.82 23.95 97.74 −0.329 细胞质 DzLEA5 evm.model.
GRLG01.2002DHN 217 24 476.28 5.21 56.28 50.28 −1.469 细胞核 DzLEA6 evm.model.
GRLG02.113LEA6 119 13 380.79 5.67 39.91 63.19 −0.873 细胞核 DzLEA7 evm.model.
GRLG02.284DHN 129 14 547.17 5.92 38.65 48.45 −1.479 细胞质 DzLEA8 evm.model.
GRLG04.1406LEA4 68 7 164.88 6.17 49.42 21.91 −1.128 细胞核 DzLEA9 evm.model.
GRLG06.455LEA3 92 10 608.17 7.97 39.78 75.33 −0.738 线粒体 DzLEA10 evm.model.
GRLG06.838LEA4 89 9 788.14 9.44 36.21 67.08 −0.84 线粒体 DzLEA11 evm.model.
GRLG09.393LEA6 467 52 080.58 8.43 54.41 57.24 −0.995 细胞核 DzLEA12 evm.model.
GRLG10.285LEA4 219 24 550.1 5.21 47.73 57.12 −1.093 线粒体 DzLEA13 evm.model.
GRLG10.1061LEA5 154 16 970.59 5.73 40.03 49.48 −1.089 细胞核 DzLEA14 evm.model.
GRLG10.1609SMP 255 26 106.87 4.76 44.96 72.24 −0.37 细胞核 DzLEA15 evm.model.
GRLG12.47LEA5 148 16 148.01 9.76 53.95 47.5 −1.164 细胞核 DzLEA16 evm.model.
GRLG14.853DHN 148 16 966.69 9.22 60.67 38.92 −1.568 细胞核 DzLEA17 evm.model.
GRLG14.963DHN 175 19 794.94 6.23 33.38 42.46 −1.727 细胞核 DzLEA18 evm.model.
GRLG16.937DHN 225 25 603.54 5.39 64.15 52 −1.455 细胞核 DzLEA19 evm.model.
GRLG17.386LEA2 151 16 479.04 5.13 22.74 107.81 −0.004 细胞质 DzLEA20 evm.model.
GRLG17.677LEA2 213 23 484.56 9.63 40.25 86.67 0.306 质膜 DzLEA21 evm.model.
GRLG17.729LEA3 98 10 476.93 9.39 32.05 75.71 −0.218 叶绿体 DzLEA22 evm.model.
GRLG17.907SMP 199 20 782.97 4.33 29.71 83.52 −0.239 细胞质 DzLEA23 evm.model.
GRLG17.908SMP 209 21 511.99 5.58 19.87 73.44 −0.346 叶绿体 DzLEA24 evm.model.
GRLG17.1800LEA4 68 7 255.97 6.55 44.54 27.65 −1.144 细胞核 DzLEA25 evm.model.
GRLG17.1806LEA4 66 7 012.69 8.06 45.65 26.82 −1.162 细胞核 DzLEA26 evm.model.
GRLG18.174LEA3 98 10 468.01 9.51 27.63 79.59 −0.136 叶绿体 DzLEA27 evm.model.
GRLG18.273SMP 218 22 421.95 4.53 27.47 77.25 −0.141 细胞质 DzLEA28 evm.model.
GRLG18.274SMP 235 24 596.15 5.26 28.91 69.79 −0.565 线粒体 DzLEA29 evm.model.
GRLG19.74LEA5 188 21 184.08 9.32 56.22 74.2 −0.728 细胞核 DzLEA30 evm.model.
GRLG21.580SMP 138 14 227.09 4.85 37.66 104.06 0.151 细胞质 DzLEA31 evm.model.
GRLG21.828LEA4 89 9 524.33 6.13 51.25 36.4 −1.126 细胞核 DzLEA32 evm.model.
GRLG21.971LEA3 104 11 155.67 7.92 34.59 73.27 −0.207 叶绿体 DzLEA33 evm.model.
GRLG21.1142LEA4 160 17 803.70 9.23 34.74 44.06 −1.263 细胞核 DzLEA34 evm.model.
GRLG24.39LEA4 68 7 230.00 5.31 54.78 17.65 −1.131 细胞核 DzLEA35 evm.model.
GRLG24.42LEA4 66 7 046.73 7.86 42.34 16.67 −1.312 线粒体 DzLEA36 evm.model.
GRLG24.44LEA4 66 7 028.69 7.86 41.05 22.58 −1.283 线粒体 DzLEA37 evm.model.
GRLG24.45LEA4 66 7 046.73 7.86 42.34 16.67 −1.312 线粒体 DzLEA38 evm.model.
GRLG24.756SMP 349 36 686.51 4.5 27.85 72.06 −0.367 细胞质 DzLEA39 evm.model.
GRLG24.892LEA3 98 10 348.88 9.51 27.74 76.63 −0.129 叶绿体 DzLEA40 evm.model.
GRLG24.1149LEA2 206 22 466.73 4.92 45.57 96.12 −0.233 细胞质 DzLEA41 evm.model.
GRLG25.862LEA3 86 9 907.57 9.69 38.16 73.72 −0.614 叶绿体 DzLEA42 evm.model.
GRLG25.1244LEA4 108 11 383.16 5.04 46.33 30 −1.132 细胞核 DzLEA43 evm.model.
GRLG26.6LEA4 310 33 834.60 5.67 36.56 52.77 −0.992 细胞外 DzLEA44 evm.model.
GRLG27.782SMP 177 18 410.53 6.75 37.62 78.53 −0.323 细胞核 DzLEA45 evm.model.
GRLG27.783SMP 65 6 965.80 4.35 59.62 57.23 −0.497 线粒体 DzLEA46 evm.model.
GRLG28.782LEA4 111 11 764.55 5.65 40.11 31.89 −1.196 细胞核
下载: 导出CSV
-
[1] 辛曙丽, 刘永华, 陈庆河. 中国榴梿产业发展现状·研究进展及展望[J]. 安徽农业科学, 2025, 53(24): 21−26. [2] 谢圣华, 李向宏, 张晶, 等. 海南省榴莲病虫害调查初报[J]. 中国果树, 2025(1): 118−126. https://doi.org/10.16626/j.cnki.issn1000-8047.2025.01.018 doi: 10.16626/j.cnki.issn1000-8047.2025.01.018 [3] 何红照, 周兆禧, 李新国, 等. 低温胁迫下不同品种榴莲幼苗叶片解剖结构及光合特性分析[J/OL]. 中国南方果树, 1-156-01-06]. https://doi.org/10.13938/j.issn.1007-1431.20240623. (in Chinese 6-01-06]. https://doi.org/10.13938/j.issn.1007-1431.20240623. [4] Galau G A, Dure III L. Developmental biochemistry of cottonseed embryogenesis and germination: changing messenger ribonucleic acid populations as shown by reciprocal heterologous complementary deoxyribonucleic acid-messenger ribonucleic acid hybridization [J]. Biochemistry, 1981, 20(14): 4169−4178. https://doi.org/10.1021/bi00517a034 doi: 10.1021/bi00517a034 [5] Zou Z, Li M Y, Jia R Z, et al. Genes encoding light-harvesting chlorophyll a/b-binding proteins in papaya (Carica papaya L. ) and insight into lineage-specific evolution in brassicaceae [J]. Gene, 2020, 748: 144685. https://doi.org/10.1016/j.gene.2020.144685 doi: 10.1016/j.gene.2020.144685 [6] Hundertmark M, Hincha D K. Lea (Late Embryogenesis Abundant) proteins and their encoding genes in Arabidopsis thaliana [J]. BMC Genomics, 2008, 9: 118. https://doi.org/10.1186/1471-2164-9-118 doi: 10.1186/1471-2164-9-118 [7] Wang X S, Zhu H B, Jin G L, et al. Genome-scale identification and analysis of LEA genes in rice (Oryza sativa L. ) [J]. Plant Science, 2007, 172(2): 414−420. https://doi.org/10.1016/j.plantsci.2006.10.004 doi: 10.1016/j.plantsci.2006.10.004 [8] Su L, Zhao C Z, Bi Y P, et al. Isolation and expression analysis of lea genes in peanut (Arachis hypogaea L. ) [J]. Journal of Biosciences, 2011, 36(2): 223−228. https://doi.org/10.1007/s12038-011-9058-5 doi: 10.1007/s12038-011-9058-5 [9] Jin X F, Cao D, Wang Z J, et al. Genome-wide identification and expression analyses of the lea protein gene family in tea plant reveal their involvement in seed development and abiotic stress responses [J]. Scientific Reports, 2019, 9(1): 14123. https://doi.org/10.1038/s41598-019-50645-8 doi: 10.1038/s41598-019-50645-8 [10] Artur M A S, Zhao T, Ligterink W, et al. Dissecting the genomic diversification of late embryogenesis abundant (LEA) protein gene families in plants [J]. Genome Biology and Evolution, 2019, 11(2): 459−471. https://doi.org/10.1093/gbe/evy248 doi: 10.1093/gbe/evy248 [11] Garay-Arroyo A, Colmenero-Flores J M, Garciarrubio A, et al. Highly hydrophilic proteins in prokaryotes and eukaryotes are common during conditions of water deficit [J]. Journal of Biological Chemistry, 2000, 275(8): 5668−5674. https://doi.org/10.1074/jbc.275.8.5668 doi: 10.1074/jbc.275.8.5668 [12] Li W W, Chen X, Yu J N, et al. Upgraded durian genome reveals the role of chromosome reshuffling during ancestral karyotype evolution, lignin biosynthesis regulation, and stress tolerance [J]. Science China Life Sciences, 2024, 67(6): 1266−1279. https://doi.org/10.1007/s11427-024-2580-3 doi: 10.1007/s11427-024-2580-3 [13] Chen C J, Chen H, Zhang Y, et al. TBtools: an integrative toolkit developed for interactive analyses of big biological data [J]. Molecular Plant, 2020, 13(8): 1194−1202. https://doi.org/10.1016/j.molp.2020.06.009 doi: 10.1016/j.molp.2020.06.009 [14] Horton P, Park K J, Obayashi T, et al. WoLF PSORT: protein localization predictor [J]. Nucleic Acids Research, 2007, 35(suppl_2): W585−W587. https://doi.org/10.1093/nar/gkm259 doi: 10.1093/nar/gkm259 [15] Letunic I, Bork P. Interactive tree of life (iTOL)v6: recent updates to the phylogenetic tree display and annotation tool [J]. Nucleic Acids Research, 2024, 52(W1): W78−W82. https://doi.org/10.1093/nar/gkae268 doi: 10.1093/nar/gkae268 [16] Cantalapiedra C P, Hernández-Plaza A, Letunic I, et al. EggNOG-mapper v2: functional annotation, orthology assignments, and domain prediction at the metagenomic scale [J]. Molecular Biology and Evolution, 2021, 38(12): 5825−5829. https://doi.org/10.1093/molbev/msab293 doi: 10.1093/molbev/msab293 [17] Ye J, Coulouris G, Zaretskaya I, et al. Primer-BLAST: a tool to design target-specific primers for polymerase chain reaction [J]. BMC Bioinformatics, 2012, 13: 134. https://doi.org/10.1186/1471-2105-13-134 doi: 10.1186/1471-2105-13-134 [18] Pedrosa A M, De Paula Santos Martins C, Gonçalves L P, et al. Late Embryogenesis Abundant (LEA) constitutes a large and diverse family of proteins involved in development and abiotic stress responses in sweet orange (Citrus sinensis L. Osb. ) [J]. PLoS One, 2015, 10(12): e0145785. https://doi.org/10.1371/journal.pone.0145785 doi: 10.1371/journal.pone.0145785 [19] Wu C L, Hu W, Yan Y, et al. The late embryogenesis abundant protein family in cassava (Manihot esculenta crantz): genome-wide characterization and expression during abiotic stress [J]. Molecules, 2018, 23(5): 1196. https://doi.org/10.3390/molecules23051196 doi: 10.3390/molecules23051196 [20] 赵东方, 苗红霞, 刘菊华, 等. 香蕉胚胎发育晚期丰富蛋白(MaLEA)家族生物信息学分析[J]. 分子植物育种, 2020, 18(12): 3893−3901. https://doi.org/10.13271/j.mpb.018.003893 doi: 10.13271/j.mpb.018.003893 [21] Ling H, Zeng X, Guo S X. Functional insights into the late embryogenesis abundant (LEA) protein family from Dendrobium officinale (orchidaceae) using an Escherichia coli system [J]. Scientific Reports, 2016, 6: 39693. https://doi.org/10.1038/srep39693 doi: 10.1038/srep39693 [22] Candat A, Paszkiewicz G, Neveu M, et al. The ubiquitous distribution of late embryogenesis abundant proteins across cell compartments in Arabidopsis offers tailored protection against abiotic stress [J]. The Plant Cell, 2014, 26(7): 3148−3166. https://doi.org/10.1105/tpc.114.127316 doi: 10.1105/tpc.114.127316 [23] Hincha D K, Thalhammer A. LEA proteins: IDPs with versatile functions in cellular dehydration tolerance [J]. Biochemical Society Transactions, 2012, 40(5): 1000−1003. https://doi.org/10.1042/BST20120109 doi: 10.1042/BST20120109 [24] Defoort J, Van De Peer Y, Carretero-Paulet L. The evolution of gene duplicates in angiosperms and the impact of protein-protein interactions and the mechanism of duplication [J]. Genome Biology and Evolution, 2019, 11(8): 2292−2305. https://doi.org/10.1093/gbe/evz156 doi: 10.1093/gbe/evz156 -

 点击查看大图
点击查看大图
计量
- 文章访问数: 630
- HTML全文浏览量: 266
- 被引次数: 0
