


-
木薯(Manihot esculenta)是热带地区主要的粮食作物和经济作物,有着“地下粮仓”、“淀粉之王”的称号[1-2],木薯在我国主要用于各种工业原料的生产[3-5],其市场前景十分巨大。菜豆黄单胞菌木薯萎蔫致病变种(Xanthomonas phaseoli pv. manihotis, Xpm)早已列入我国有害生物名录,它是木薯细菌性枯萎病的病原菌,其能够使木薯叶片褪绿萎蔫甚至全株死亡,可导致木薯减产20%~90%[6-7]。1991年于类囊体腔中首次发现PsbP蛋白,后经过不断研究发现PsbP蛋白广泛存在于藻类、蓝细菌和高等植物中[8-9]。目前,已发现的PsbP蛋白分为3大类,分别是PsbP蛋白、类PsbP蛋白PPL(PsbP-like protein)和PsbP结构域蛋白PPD(PsbP-domain protein)[10-12]。PsbP蛋白是叶绿体光系统Ⅱ(photosystem Ⅱ complex, PSⅡ)的重要组成部分,它在维持钙离子、氯离子浓度方面发挥重要作用;此外,敲除PsbP基因后, PSⅡ的功能将受到一定的影响[13-15]。科学家在拟南芥(Arabidopsis thaliana)中发现,PPL1能参与 PSⅡ复合体的组装,其和植物适合光照强度变化有关[16-17];拟南芥的PPL2缺失突变体中,叶绿体NADH脱氢酶的积累量和活性降低[17]。Roose等[8]于2007年从拟南芥基因组中鉴定出7个PPD蛋白,它们分别是PPD1、PPD2、PPD3、PPD4、PPD5、PPD6、PPD7。已有研究发现,PPD1是PSI复合体的组装因子,在拟南芥中,PPD1缺失突变体能导致PSI复合体无法正常组装,从而影响植物的光合作用[18-19];在拟南芥中,PPD5蛋白和OST1相互作用并被其磷酸化,使细胞中H2O2积累增加并使气孔关闭增强[20]。但目前尚未有过任何关于PPD3蛋白功能的研究报道。
本研究以‘华南8号’木薯为材料,通过qRT-PCR扩增得到MePPD3(Manihot esculenta psbP domain-containing protein 3)基因,并对该基因进行生物信息学分析;同时,对该基因在木薯不同部位和病原菌Xpm CHN11侵染后不同时间的表达模式进行分析;最后利用TRV介导的基因沉默技术沉默MePPD3基因后测定抗病表型,旨在为进一步研究MePPD3基因的分子机理和木薯抗细菌枯萎病的抗病机理提供基础。
-
大肠杆菌DH5α感受态细胞和农杆菌GV3101感受态细胞购自上海唯地生物;菜豆黄单胞菌木薯萎蔫致病变种Xanthomonas phaseoli pv. manihotis CHN11由笔者所在实验室提供。
-
供试的木薯品种为‘华南8号’(‘SC8’),由笔者所在实验室提供。
-
多糖多酚植物总RNA提取试剂盒(RNA prep Pure Plant Kit)和RNA反转录试剂盒(FastKing gDNA Dispelling RT SuperMix)购自天根生化科技有限公司;琼脂糖凝胶回收试剂盒和质粒小提试剂盒购自艾德莱。
-
从phytozome数据库中下载木薯MePPD3基因的CDS序列,使用Primer Primer 5.0分别设计该基因的全长扩增引物、定量分析引物和亚细胞定位引物。本研究所用引物如表1所示。
表 1 本研究所用引物
引物名称 引物序列(5′−3′) 引物用途 EF1α-F TGCCATGTTCCGTGGAAAGATG 内参基因引物 EF1α-R CCCCTAGGTGGAATGTCACAGACAC 内参基因引物 MePPD3-F CATATGATGGCGTCTGTTTCTTTGCTGTC 扩增目的基因 MePPD3-R GAATTCTCATGAAATGAACCTGAAGGATG 扩增目的基因 qpcr-MePPD3-F AGACTGATGAGCTTCGCGTT 定量分析 qpcr-MePPD3-R TGACTTGAACCCGCCGTTAG 定量分析 1300-MePPD3-F TTGATACATATGCCCGTCGACATGGCGTCTGTTTCTTTGC 亚细胞定位 1300-MePPD3-R GCCCTTGCTCACCATGGATCCTGAAATGAACCTGAAGGATG 亚细胞定位 -
取少许木薯叶片于液氮中速冻,用液氮预冷后的研钵进行研磨。使用试剂盒提取木薯RNA并将其反转录为cDNA。以木薯cDNA为PCR模板,以MePPD3-F和MePPD3-R为引物,扩增木薯MePPD3基因序列。待琼脂糖凝胶电泳检测结果无误后,转入植物亚细胞载体pCAMBIA1300-35S-GFP中,挑选阳性单克隆送至楠山生物进行测序。
-
借助SignalP 5.0 Server 对MePPD3蛋白的信号肽进行预测;通过TMHMM Serverv 2.0分析MePPD3蛋白的跨膜结构;利用NetPhos 3.1 Server在线工具(
http://www.cbs.dtu.dk/services/NetPhos/ )预测MePPD3蛋白的磷酸化位点和糖基化位点;分别使用在线网站PSIPRED.(http://bioinf.cs.ucl.ac.uk/psipred/ )和PHYRE2(http://www.sbg.bio.ic.ac.uk/~phyre2/html/page.cgi?id=index )预测MePPD3蛋白的二级结构和三级结构。通过NCBI序列比对,找到MePPD3蛋白在不同植物中的同源蛋白。通过DNAMAN 8.0软件对这9个同源蛋白进行多序列比对;使用MEGA 11.0软件对这些同源蛋白构建NJ进化树;使用MEME在线网站对MePPD3及其同源蛋白进行保守Motif分析。
-
将构建好的植物亚细胞定位载体pCAMBIA1300-35S-GFP-MePPD3与空载pCAMBIA1300-35S-GFP分别转入GV3101菌株。挑取阳性克隆于28 ℃ 200 r·min−1培养过夜,取适量菌液转移至新的LB(含Kan和Rif)液体培养基中扩大培养。调节OD600≈1.0,黑暗静置2.5 h,注射于生长30 d左右的烟草(Nicotiana tabacum)叶片背面,做好标记,培养40 h后于共聚焦显微镜下观察其荧光。
-
使用Primer Primer 5.0设计定量引物qpcr-MePPD3-F/R。分别取长势相同的木薯的顶芽、幼嫩叶片、成熟叶片、叶柄、须根、块根等部位,每个部位至少3个生物学重复,液氮速冻后,使用RNA prep Pure Plant Kit试剂盒提取木薯总RNA,然后再按照FastKing gDNA Dispelling RT SuperMix试剂盒的流程反转录cDNA。
-
用OD600≈0.05的菜豆黄单胞菌木薯萎蔫致病变种Xpm CHN11侵染多株长势相同的木薯,分别于侵染后0 h、6 h、1 d、3 d、5 d随机剪取叶片,液氮速冻后将其研磨提取RNA,并反转录为cDNA。
-
构建沉默载体pCsCMV-MePPD3。将其与正对照pCsCMV-B和负对照pCsCMV-A分别转化入GV3101菌株,在LB液体培养基(含Kan和Rif)中培养阳性克隆至OD600=1.0。然后注射于木薯‘SC8’叶片背面,24 ℃培养30 d。
在LPGA固体培养基(含Chl)上活化Xpm CHN11菌株,活化2次后,用无菌水清洗菌体并稀释至OD600=0.05,用该菌液注射木薯叶片。侵染6 d后,观察叶片表面病斑的扩散情况。
-
以木薯RNA为模板,MePPD3-F/R为上下游引物,通过RT-PCR技术,得到目的基因MePPD3(Manihot esculenta psbP domain-containing protein 3)(图1-a)。基因序列分析发现,MePPD3基因全长807 bp,编码268个氨基酸。ExPasy在线网站分析结果显示,MePPD3相对分子质量为67 522.75,理论等电点为5.11,分子式为C2467H4129N807O1033S184,不稳定系数为49.9,脂肪系数为27.14,亲水系数为0.775。使用NCBI的CDD数据库分析其保守结构域,结果发现,其氨基酸序列第106~266位存在PsbP结构域(图1-b),推测其为PPD3蛋白。

图 1 MePPD3基因全长扩增及蛋白结构分析
-
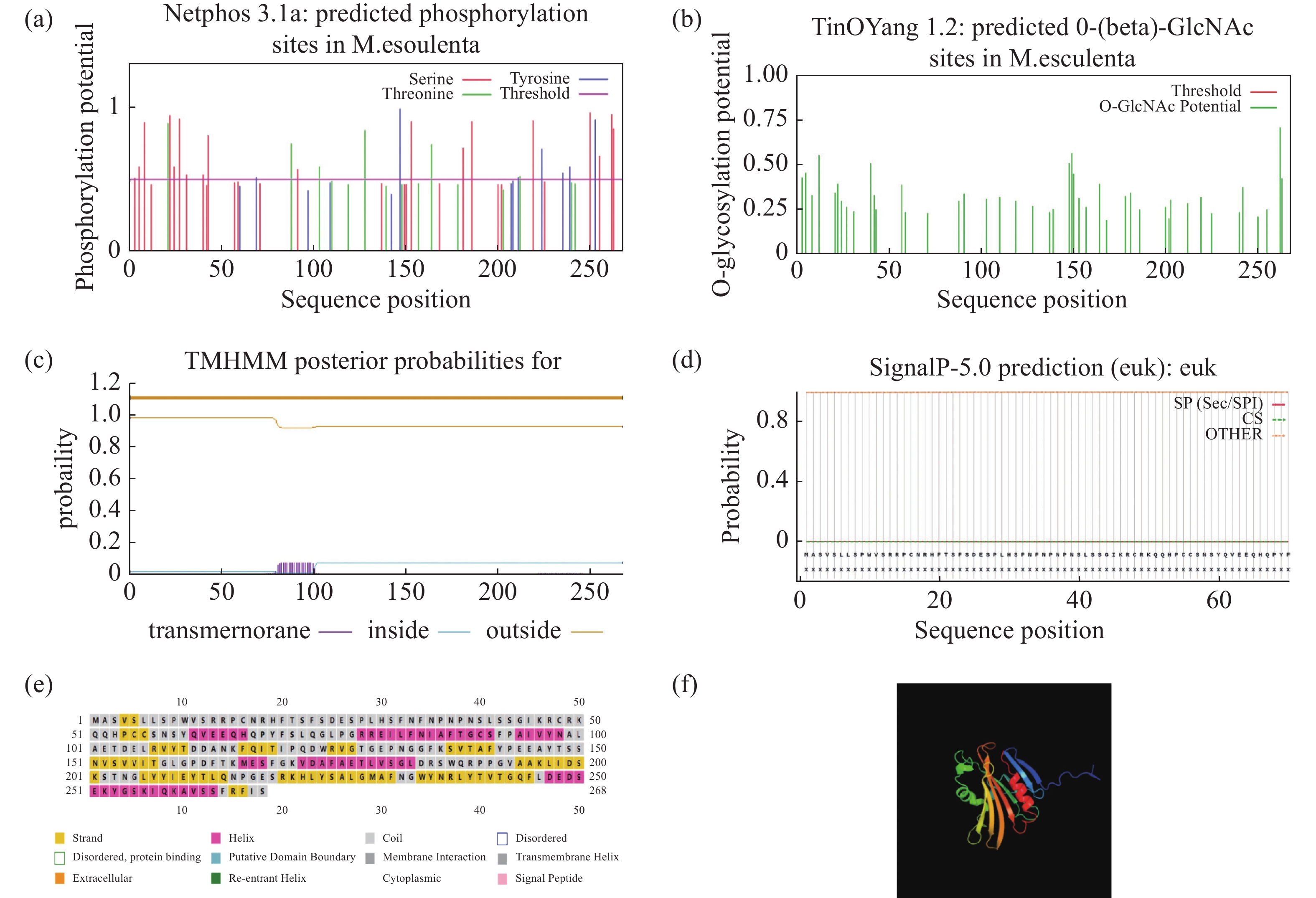
使用NetPhos在线网站,预测MePPD3蛋白有32个磷酸化位点,其中,有19个丝氨酸、6个苏氨酸和7个酪氨酸(图2-a);有5个糖基化位点(图2-b);使用TMHMM在线网站预测发现,其存在跨膜结构域(图2-c);使用SignalIP在线网站发现,其无信号肽(图2-d);使用PSIPRED在线网站预测MePPD3蛋白二级结构,发现其有6处螺旋结构,其中,螺旋占21.3%,折叠占26.5%,无规卷曲占52.2%(图2-e);通过PHYRE2在线网站预测其三级结构,发现其有6个螺旋结构,这符合二级结构预测结果和PsbP蛋白的结构特征(图2-f)。

图 2 MePPD3蛋白的生物信息学分析
-
为研究PPD3蛋白在不同植物中是否具有较高的同源性,首先在NCBI通过BLAST比对,在橡胶(Hevea brasiliensis)、蓖麻(Ricinus communis)、雷公藤(Tripterygium wilfordii)、茶树(Camellia sinensis)、胡杨(Populus euphratica)、芒果(Mangifera indica)、麻风树(Jatropha curcas)、开心果(Pistacia vera)等8种植物中找到了与木薯MePPD3蛋白同源的氨基酸序列(表2)。使用DANMAN 8.0对这些序列进行比对(图3),结果发现,MePPD3蛋白在不同的植物中具有较高的同源性,其中与橡胶中PPD3蛋白的同源性最高。PsbP保守结构域在不同植物中的差别不大(图中红线框),且N端氨基酸序列含有叶绿体/线粒体定位信号(图中蓝线),表明MePPD3蛋白可能定位于叶绿体或线粒体。
表 2 MePPD3在不同植物中的同源蛋白
物种名称 学名 基因号 木薯 Manihot esculenta Manes.05G127800.1 橡胶 Hevea brasiliensis XP_021686524.1 蓖麻 Ricinus communis XP_015573931.1 胡杨 Populus euphratica XP_011041160.1 雷公藤 Tripterygium wilfordii XP_038714823.1 茶树 Camellia sinensis XP_028073751.1 芒果 Mangifera indica XP_044475605.1 麻风树 Jatropha curcas XP_012073570.1 开心果 Pistacia vera XP_031270258.1 
图 3 MePPD3在不同植物中的同源序列比对
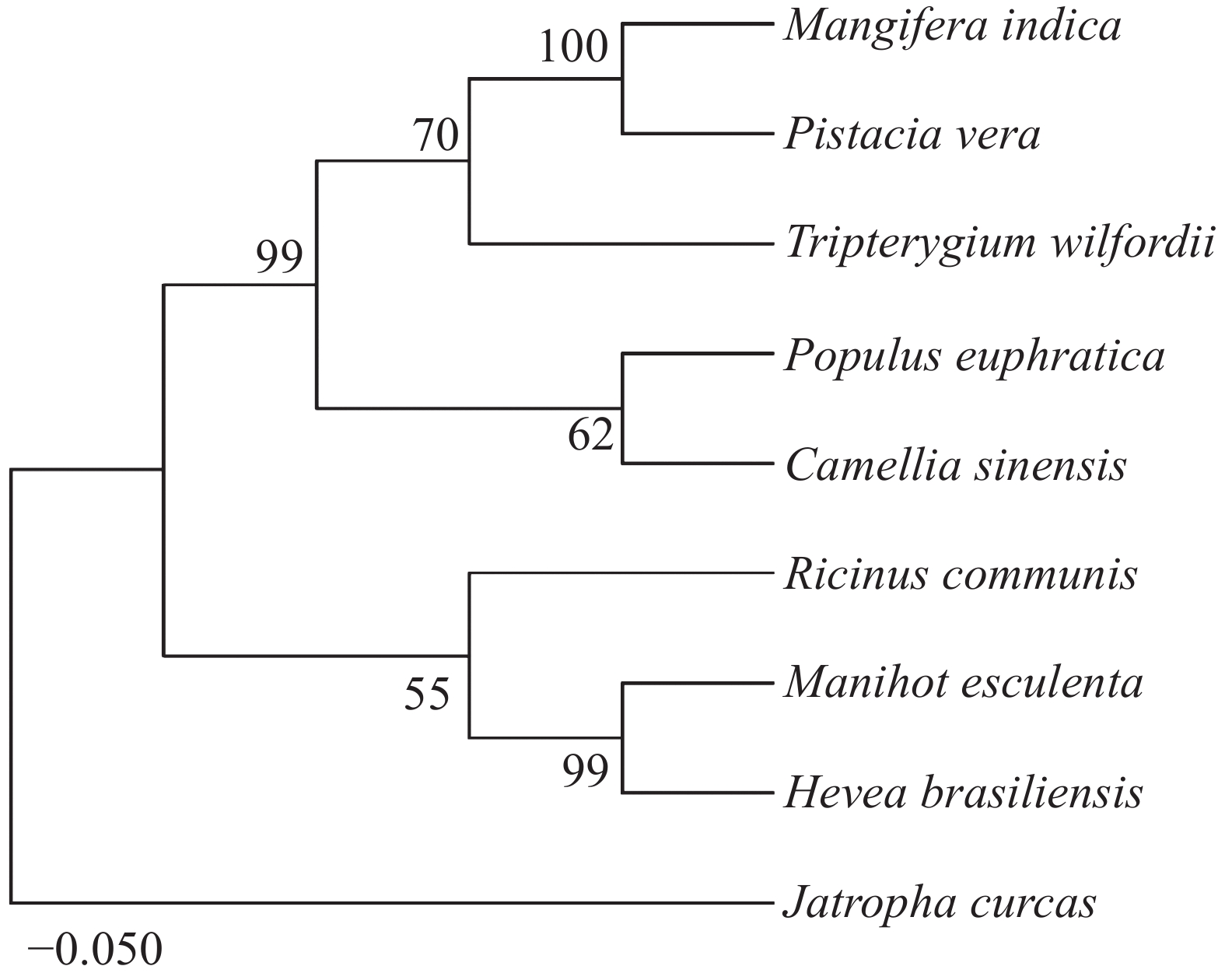
为了进一步验证这些蛋白的亲缘关系,使用MEGA11.0对这9个同源蛋白构建NJ进化树(图4)。结果发现,木薯中的MePPD3蛋白与其他几种植物中的MePPD3蛋白具有较高的亲缘关系,值得一提的是它与橡胶中PPD3蛋白的亲缘关系最接近,可达99%,蓖麻中的PPD3蛋白次之。
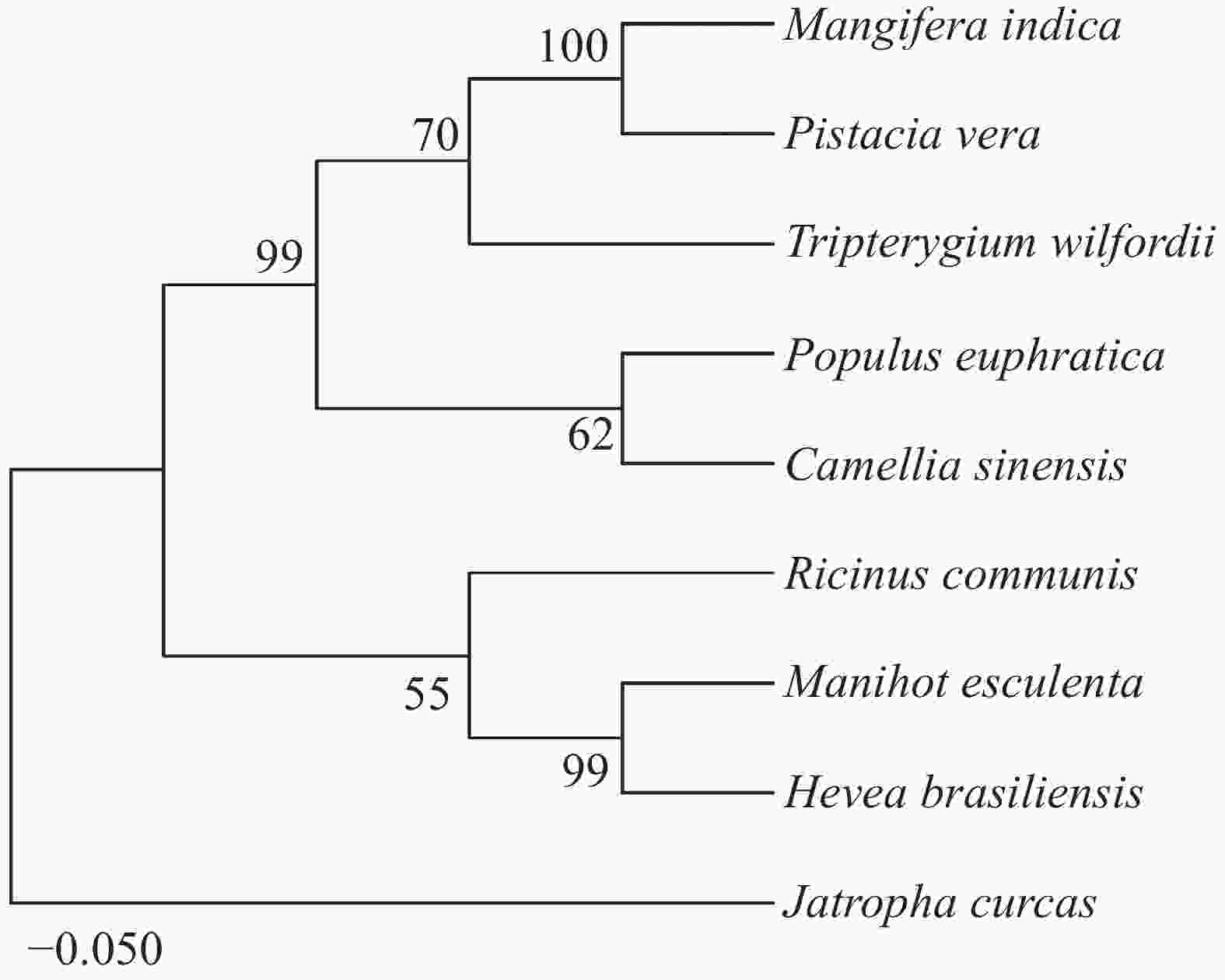
图 4 不同物种间MePPD3同源蛋白的系统进化树分析
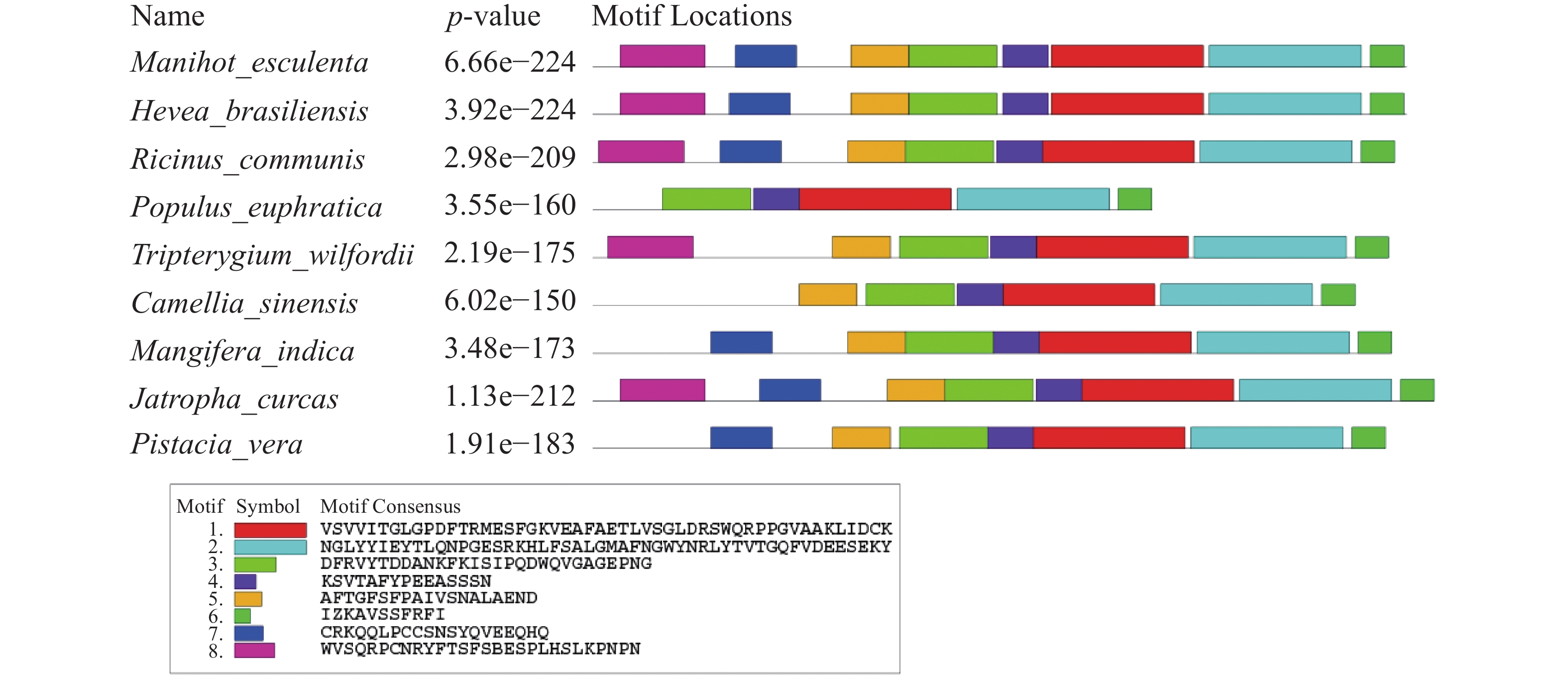
应用MEME在线网站对MePPD3蛋白及其同源蛋白进行保守Motif分析(图5),发现这9个同源蛋白至少含有6个保守Motif,其中,Motif 1、Motif 2、Motif 3、Motif 4和Motif 6具有高度保守性。此外,MePPD3蛋白和橡胶、蓖麻中PPD3蛋白具有Motif的种类和数量一致,均具有这8个Motif。这与同源蛋白多序列比对和进化树分析的结果一致,初步说明,PPD3蛋白在不同植物体内具有较高的保守性,MePPD3蛋白的功能极有可能与其他PPD3蛋白的功能一致。
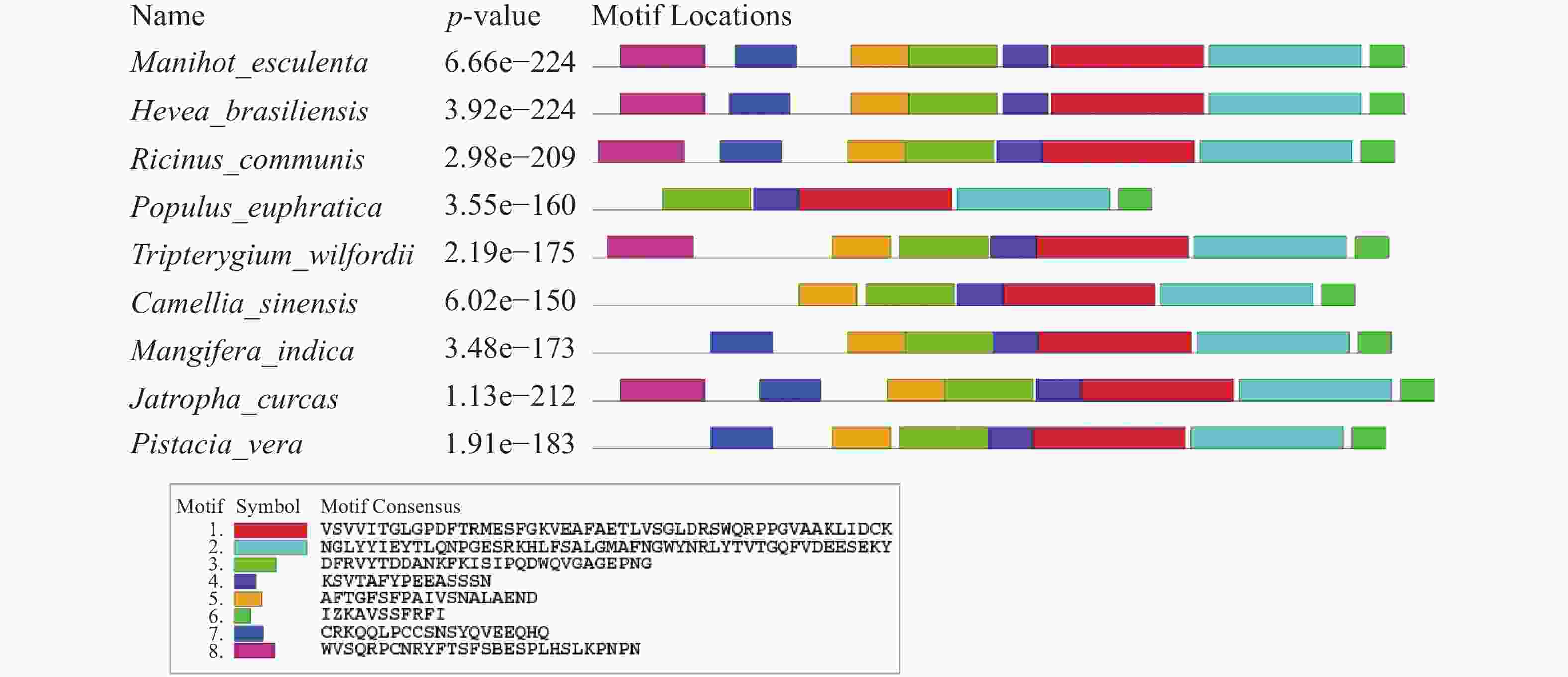
图 5 MePPD3结构域分析
-
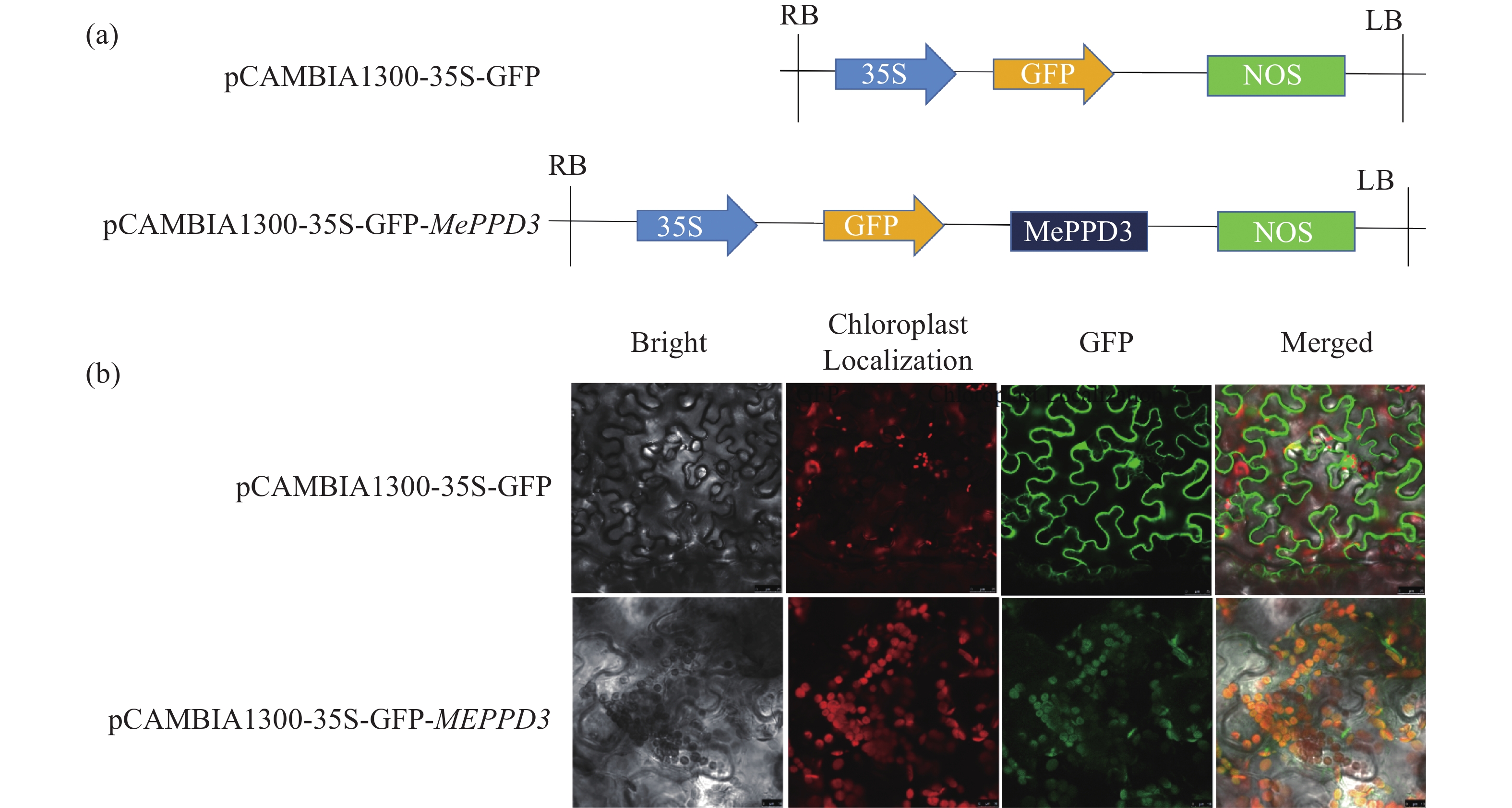
为探究木薯MePPD3蛋白在植物细胞内的亚细胞定位,构建了植物亚细胞定位载体pCAMBIA1300-35S-GFP-MePPD3(图6-a)。使用已提前转进GV3101菌株的亚细胞定位载体pCAMBIA1300-35S-GFP-MePPD3和空载pCAMBIA1300-35S-GFP注射生长约30 d的烟草叶片。40 h后,剪取叶片观察荧光信号(图6-b),结果发现,与对照(GFP空载)相比,试验组在叶绿体中能观测到绿色荧光,且能与叶绿体的红色自发荧光基本重合,表现出黄色的光。说明MePPD3蛋白定位在叶绿体中,符合Psbp蛋白(叶绿体光系统II亚基蛋白)的特点。

图 6 MePPD3蛋白的亚细胞定位
-
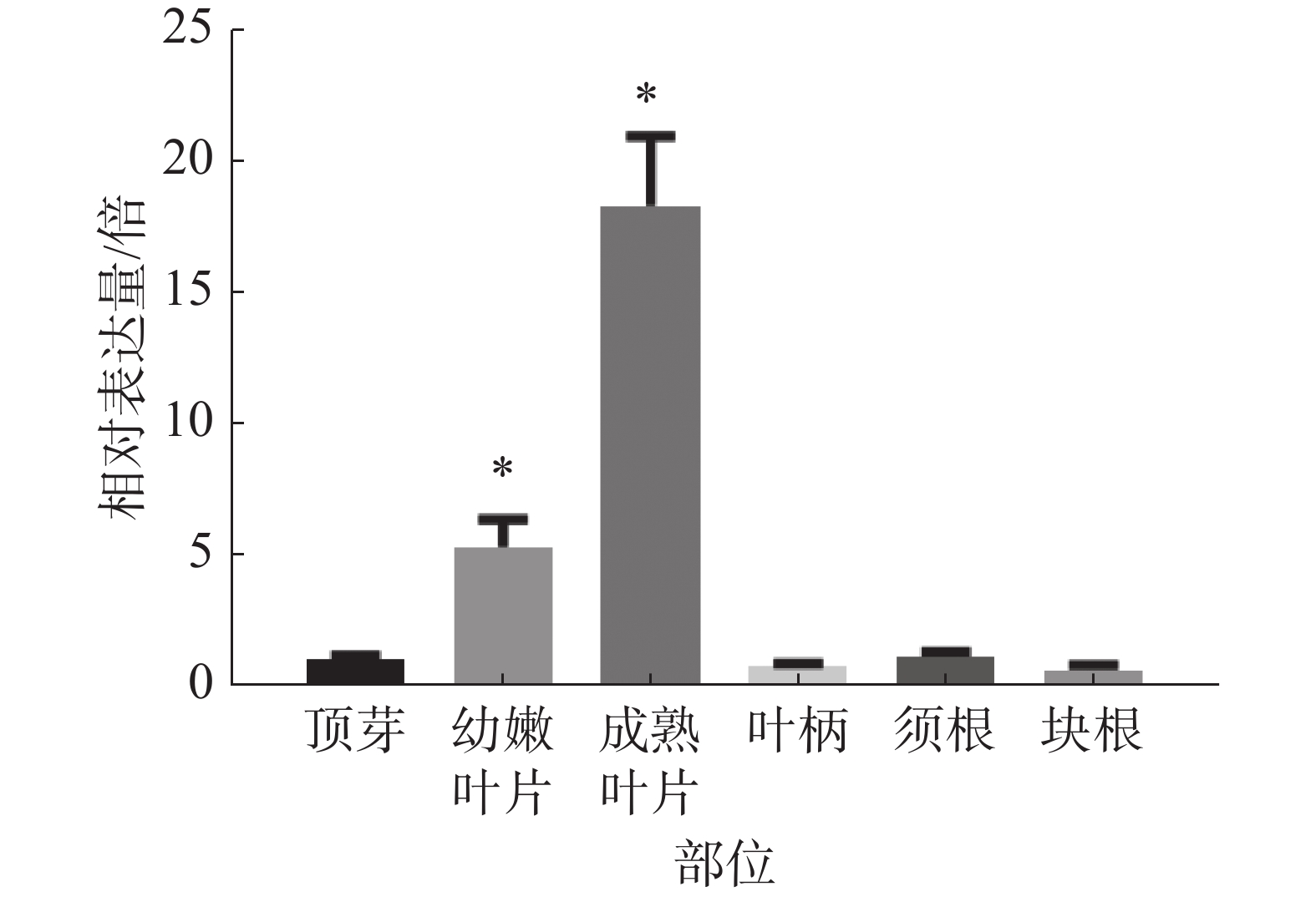
为研究木薯MePPD3基因在木薯不同部位中的表达情况,分别选择同一株‘SC8’木薯的成熟叶片、幼嫩叶片、叶柄、顶芽、块根、须根等部位,对其进行real-time PCR检测MePPD3基因的表达量是否发生变化。观察发现(图7),MePPD3基因在木薯不同组织中均有表达,其在木薯叶片中的表达量最高,在幼嫩叶片中的表达量为顶芽的5倍,在成熟叶片中的表达量为顶芽的18倍;在木薯的叶柄、须根、块根的表达量和顶芽接近。这说明MePPD3基因主要在木薯的叶片中表达,这与其具有的Psbp结构域的功能相符。
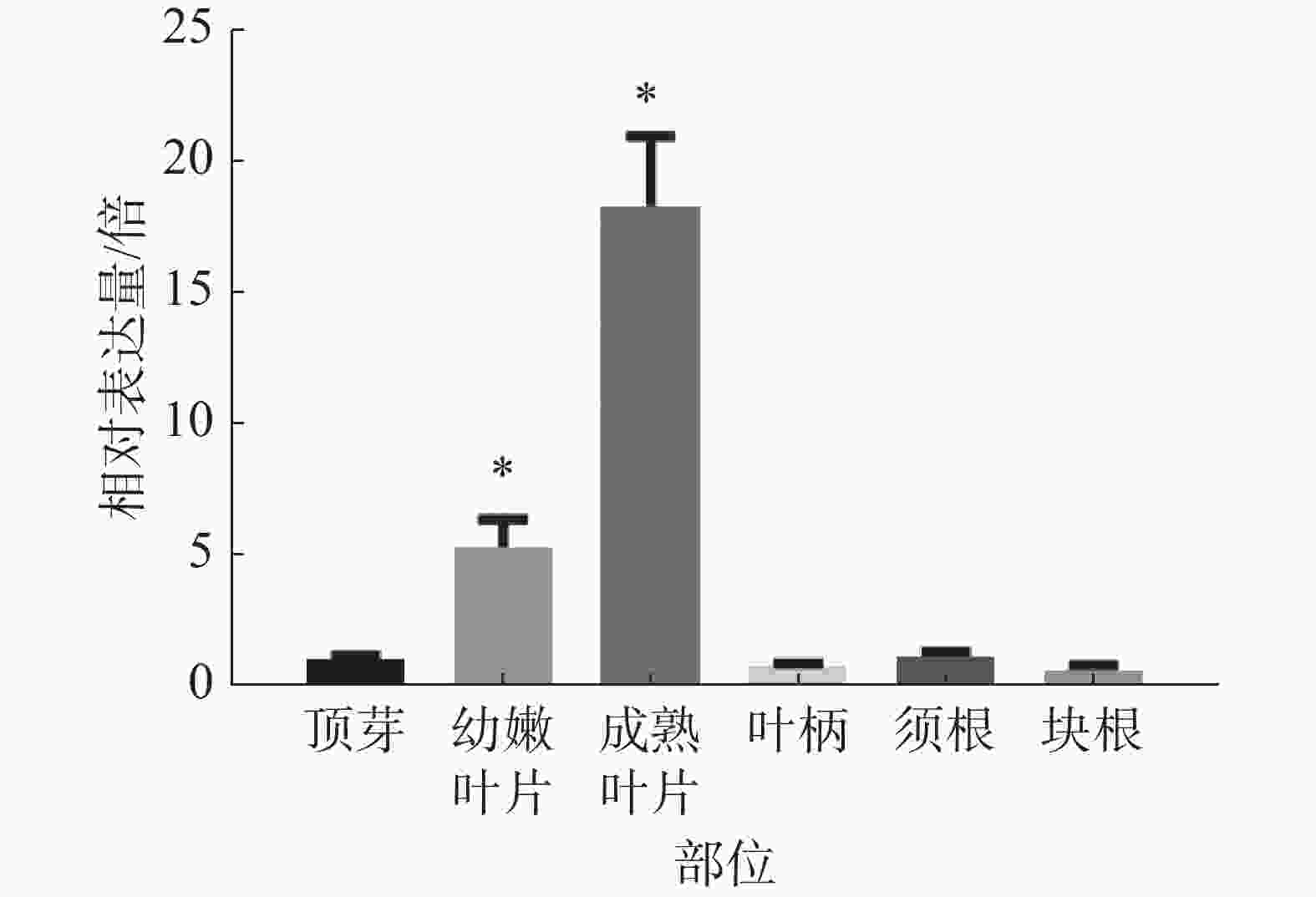
图 7 MePPD3基因在木薯不同部位的表达模式图
-
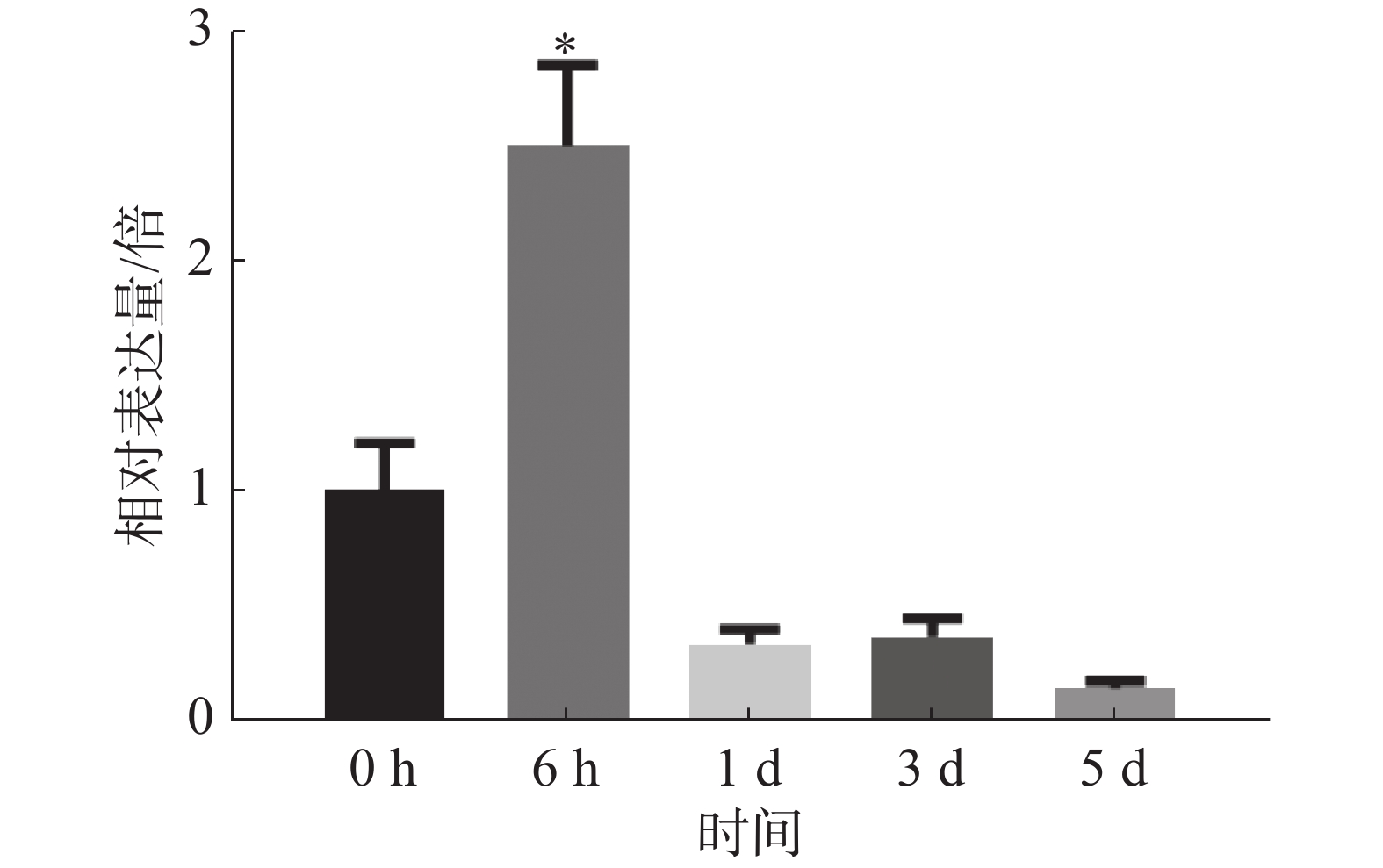
为探究病原菌Xpm CHN11侵染后MePPD3基因在木薯叶片中的表达量是否发生变化,分别取接种0、6 h,1 、3 、5 d的木薯叶片,通过实时荧光定量PCR检测MePPD3基因的表达量变化(图8)。从中可以明显地看出,在Xpm CHN11侵染木薯后的不同时间,MePPD3基因的表达量呈先上升再下降趋势。在6 h时,其表达量上升了2.5倍;在1 、3 和5 d时,其表达量逐渐下降为本底表达量。这说明,MePPD3基因可能在木薯抗病过程中发挥着作用。
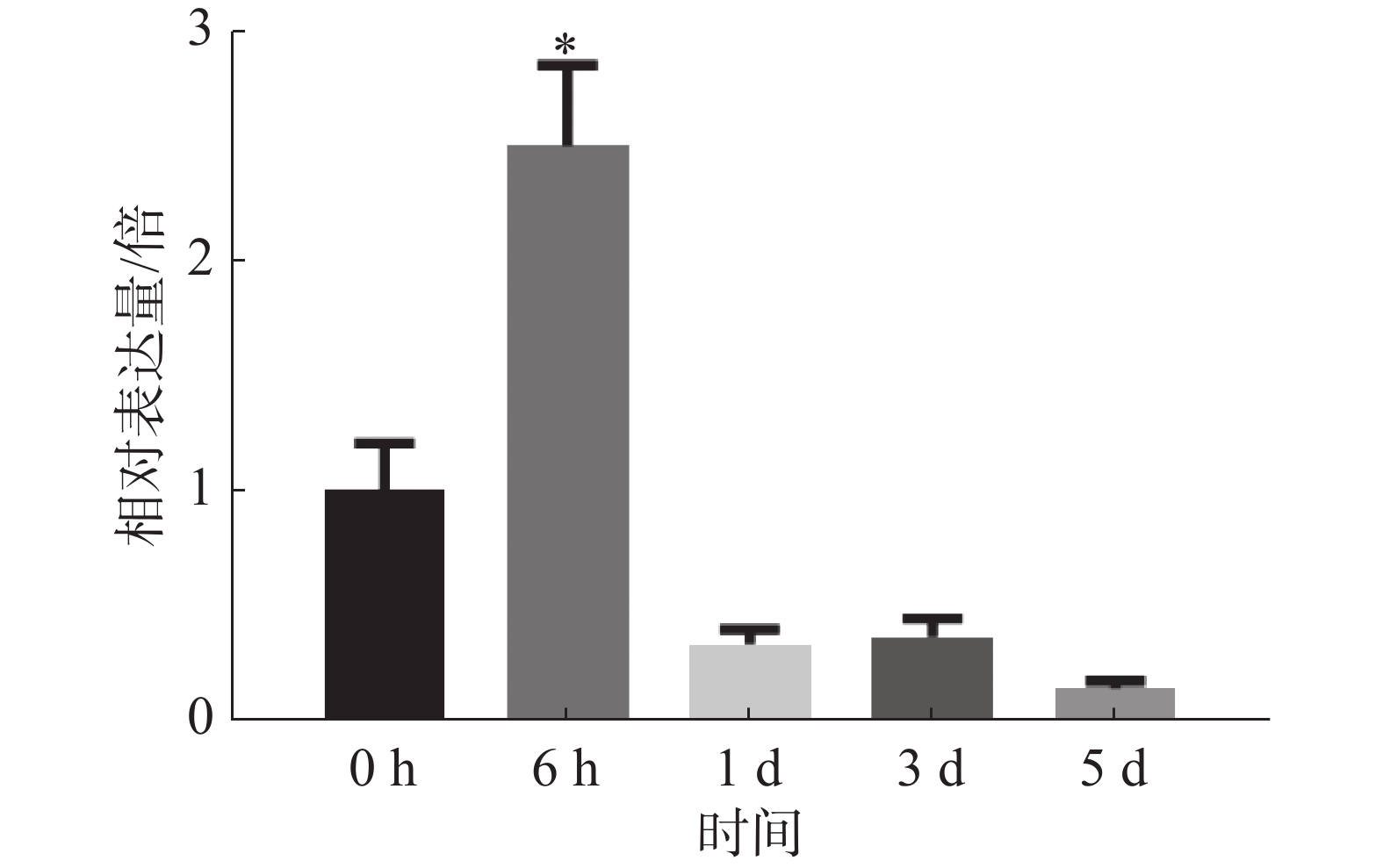
图 8 病原菌侵染后不同时间MePPD3基因的表达量
-
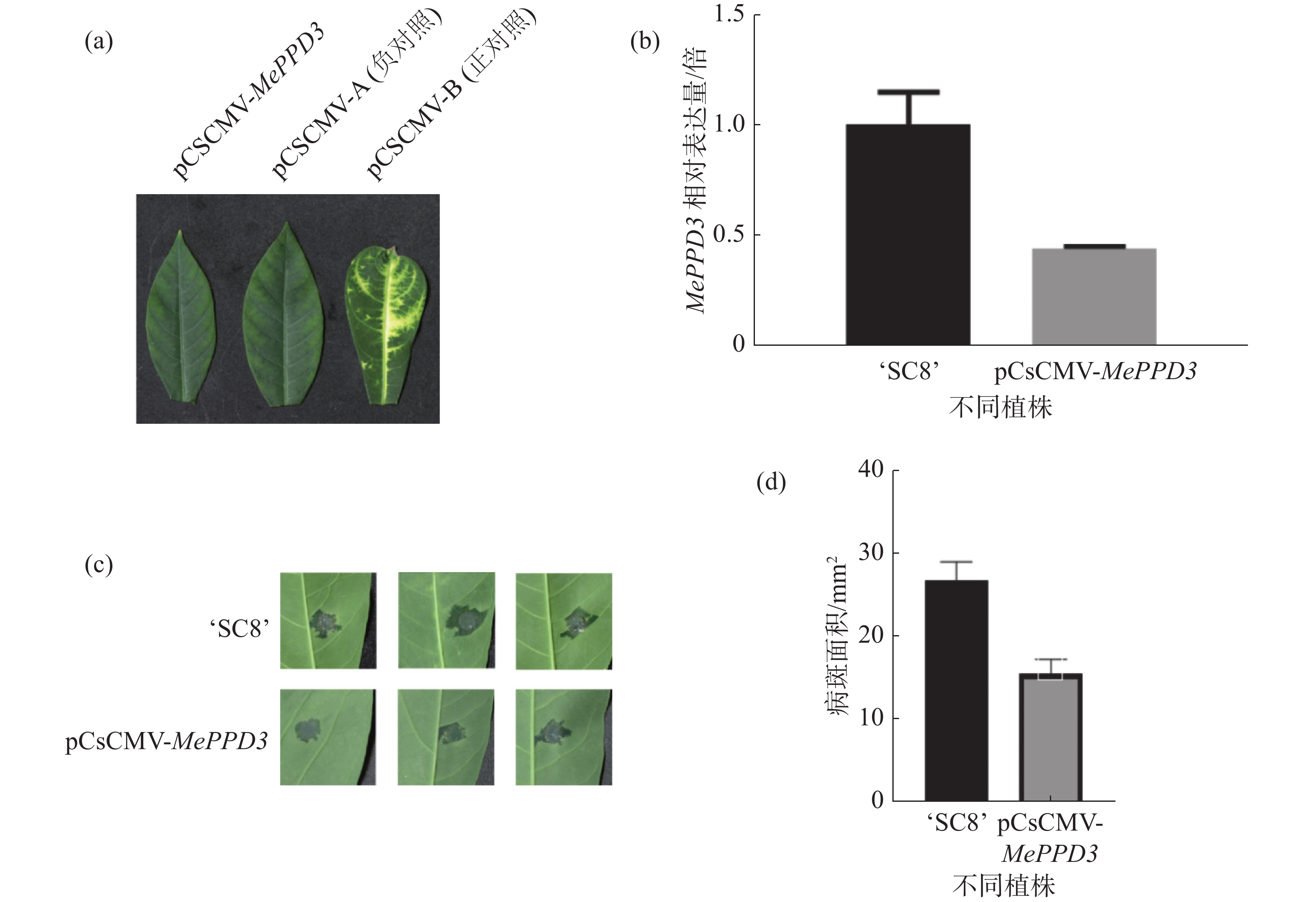
为明确MePPD3基因在木薯抗病过程中的作用,采用VIGS技术沉默MePPD3基因,沉默40 d后,接种正对照植株的叶片出现了白化表型(图9-a)。qRT-PCR结果显示,经过VIGS沉默之后,MePPD3基因的相对表达量显著降低,其表达量下降了2倍(图9-b),说明该基因受到了有效沉默。
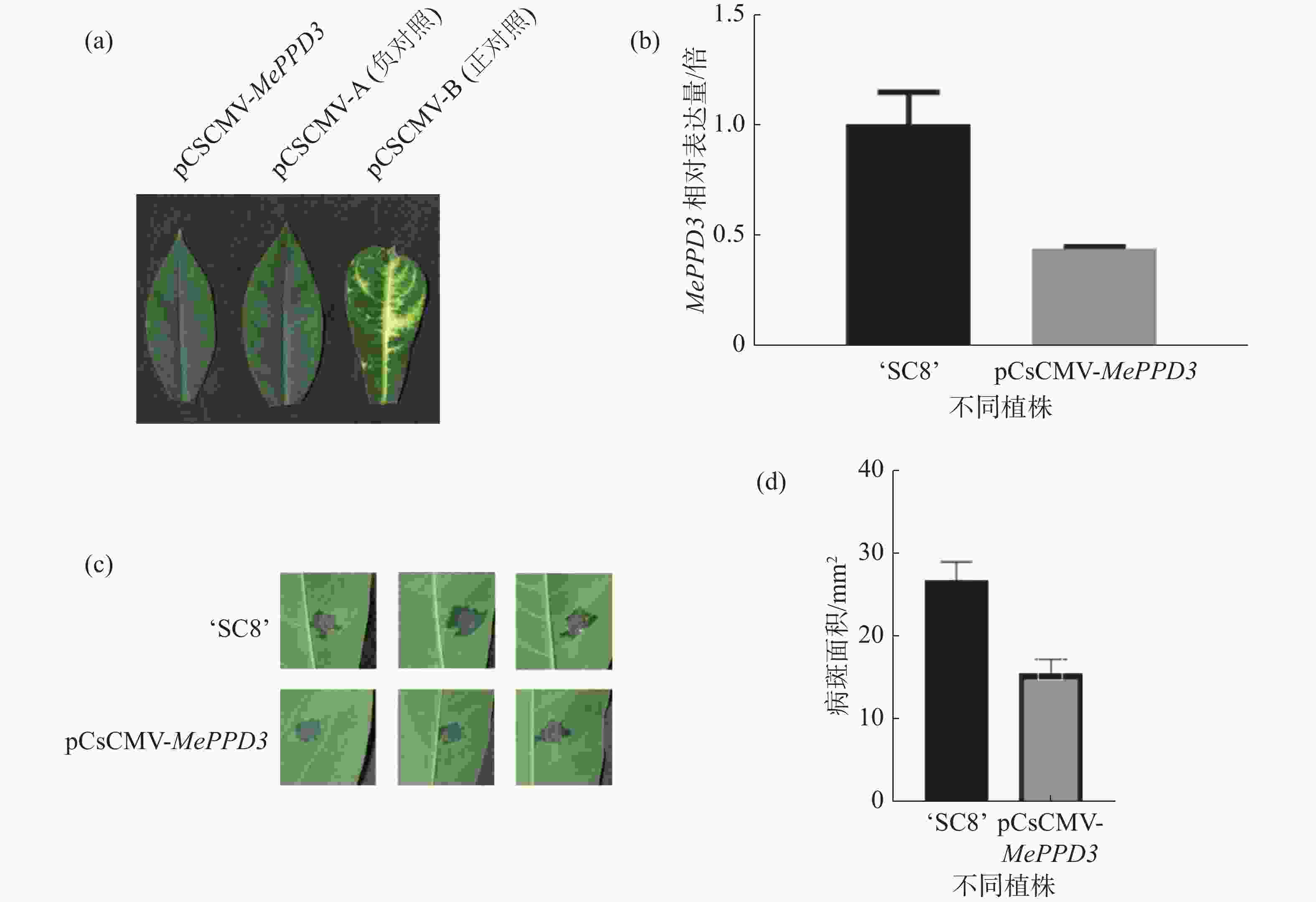
图 9 VIGS沉默植株表型及抗病性检测
分别在沉默植株和野生型植株上接种Xpm CHN11,结果发现,接种后6 d,沉默植株和野生型植株叶片均已产生水渍状病斑,且沉默植株的水渍状病斑明显小于野生型植株(图9-c),统计后发现沉默植株的病斑面积约为对照植株的一半(图9-d),该结果显示MePPD3负调控了木薯的抗病性。
-
叶绿体是植物体所特有的细胞器,它能为植物体的各项生命活动提供能量。PsbP蛋白是叶绿体光系统Ⅱ的重要组成部分,它能参与 PSⅡ的组装[13-15]。此外,葡萄球菌萄球菌RXLR效应因子RXLR31154通过影响Psbp来降低H2O2积累,从而促进葡萄致病[21]。PPD蛋白是PsbP蛋白家族的一员,目前已发现的PPD蛋白有7种。蛋白在PSI组装过程中发挥重要作用[18-19];PPD5蛋白和ROS爆发、气孔关闭有关[19]。以上研究充分说明了Psbp蛋白在植物光合作用和免疫反应中发挥着一定的作用。但目前关于PPD3蛋白的研究却不是很多。
为了增进对PPD3蛋白的了解,本研究首次从木薯数据库中得到MePPD3基因。MePPD3基因CDS序列全长807 bp,编码268个氨基酸序列。对MePPD3基因进行了一系列的生物信息学分析,结果发现:该蛋白第50~100位存在跨膜结构,第106到266位为PsbP保守结构域。其三级结构由6个螺旋组成,符合PsbP蛋白家族的结构特征。选取8种植物中的同源蛋白序列进行多序列比对、进化树和保守结构域分析,结果发现,PPD3蛋白在不同植物中具有较高的保守性,木薯MePPD3蛋白和橡胶中PPD3蛋白具有相同的保守结构域,且两者的相似性可达99%。因此,推测PPD3蛋白在不同植物中起到的作用一致或相近。此外,通过烟草瞬时表达,证实MePPD3定位于叶绿体,这与PPD3蛋白的叶绿体/线粒体定位信号相符,说明MePPD3可能在叶绿体中发挥功能。另外,本研究通过qRT-PCR检测,发现MePPD3基因在木薯成熟叶片中表达量最高;黄单胞菌Xpm CHN11侵染木薯叶片后,MePPD3表达量呈现上调趋势,初步推断,MePPD3可能参与了木薯的抗病途径;VIGS沉默MePPD3基因后接种结果显示,MePPD3负调控木薯的抗病途径。结合前人研究推测,MePPD3可能通过影响气孔开闭和ROS爆发来参与木薯抗病,是否正确仍需要后续研究来证实。
Cloning and physicochemical characterization of MePPD3 gene from cassava (Manihot esculenta)
-
摘要: 为了探究PsbP蛋白是否在木薯(Manihot esculenta)抗病中发挥功能,以木薯‘华南8号’总RNA为模板,通过RT-PCR扩增MePPD3基因(Phytozome数据库编号:Manes.05G127800)。序列分析结果显示,MePPD3基因全长807 bp,编码268个氨基酸,在氨基酸序列第106~266位存在PsbP结构域,推测其为psbP蛋白。蛋白多序列比对、进化树和保守结构域分析表明,木薯MePPD3蛋白与巴西橡胶(Hevea brasiliensis)中PsbP家族蛋白的同源性最高,同源率高达99%。亚细胞定位显示,MePPD3蛋白定位在叶绿体。qRT-PCR结果显示,MePPD3基因在木薯不同组织中的表达量具有较大差异,在叶片特别是成熟叶片中表达量最高;此外,该基因表达量受到菜豆黄单胞菌木薯萎蔫致病变种(Xanthomonas phaseoli pv. manihotis, Xpm)诱导后显著上升。利用VIGS技术沉默MePPD3基因,qRT-PCR结果表明,MePPD3基因沉默成功,其表达量显著下降;Xpm侵染植株后,沉默植株pCsCMV-MePPD3叶片的病斑面积显著小于野生型植株,故推测MePPD3可能负调控木薯对细菌性枯萎病的抗病性。Abstract: The subcellular localization and expression analysis of Psbp protein MePPD3 (Manihot esculenta PsbP domain-containing protein 3, Phytozome database number: Manes.05G127800) were performed to explore whether MePPD3 is involved in disease resistance of cassava. MePPD3 gene was amplified by RT-PCR. Sequence analysis showed that MePPD3 gene was 807bp in length, encoding 268 amino acids, with PsbP domain located at position 106-266 aa. Bioinformatics analysis of MePPD3 protein was conducted by NetPhos 3.1 Server, SignalP 5.0 Server, TMHMM Servervr, PSIPRED and PHYRE2 online, respectively. The results indicated that MePPD3 protein contained 32 phosphorylation sites, 5 glycosylation sites, and 1 transmembrane domain. The secondary structure of the protein was composed of 21.3% Helix (Helix), 26.5% fold (strand) and 52.2% random curl (loop). The multiple sequence alignment, phylogenetic tree analysis and conserved domain analysis indicated that PPD3 protein had a high genetic relationship among different plants. Subcellular localization showed that MePPD3 protein was localized in chloroplast. QRT-PCR results revealed that the expression level of MePPD3 gene in different cassava tissues was significantly different, and was the highest in mature leaves. In addition, the expression of this gene was induced by Xanthomonas phaseoli pv. manihotis (Xpm), indicating MePPD3 is involved in the resistance of cassava to Xpm. After MePPD3 gene was silenced by VIGS technique, the leaf lesion area of silent plants was significantly smaller than that of the control plants, which implies that MePPD3 negatively regulates cassava resistance to bacterial fusarium wilt caused by Xpm.
-
Key words:
- cassava /
- MePPD3 /
- bioinformatics analysis /
- Xpm CHN11
-
图 9 VIGS沉默植株表型及抗病性检测
a:沉默成功表型;b:MePPD3基因沉默效率;c:接菌表型;d:统计病斑面积;‘SC8’为野生型、pCsCMV-MePPD3为沉默植株。
表 1 本研究所用引物
引物名称 引物序列(5′−3′) 引物用途 EF1α-F TGCCATGTTCCGTGGAAAGATG 内参基因引物 EF1α-R CCCCTAGGTGGAATGTCACAGACAC 内参基因引物 MePPD3-F CATATGATGGCGTCTGTTTCTTTGCTGTC 扩增目的基因 MePPD3-R GAATTCTCATGAAATGAACCTGAAGGATG 扩增目的基因 qpcr-MePPD3-F AGACTGATGAGCTTCGCGTT 定量分析 qpcr-MePPD3-R TGACTTGAACCCGCCGTTAG 定量分析 1300-MePPD3-F TTGATACATATGCCCGTCGACATGGCGTCTGTTTCTTTGC 亚细胞定位 1300-MePPD3-R GCCCTTGCTCACCATGGATCCTGAAATGAACCTGAAGGATG 亚细胞定位  下载: 导出CSV
下载: 导出CSV
表 2 MePPD3在不同植物中的同源蛋白
物种名称 学名 基因号 木薯 Manihot esculenta Manes.05G127800.1 橡胶 Hevea brasiliensis XP_021686524.1 蓖麻 Ricinus communis XP_015573931.1 胡杨 Populus euphratica XP_011041160.1 雷公藤 Tripterygium wilfordii XP_038714823.1 茶树 Camellia sinensis XP_028073751.1 芒果 Mangifera indica XP_044475605.1 麻风树 Jatropha curcas XP_012073570.1 开心果 Pistacia vera XP_031270258.1
下载: 导出CSV
-
[1] 黄洁, 李开绵, 叶剑秋, 等. 我国的木薯优势区域概述[J]. 广西农业科学, 2008(1): 104 − 108. [2] 李开绵, 林雄, 黄洁. 国内外木薯科研发展概况[J]. 热带农业科学, 2001(1): 56 − 60. doi: 10.3969/j.issn.1009-2196.2001.01.011 [3] FERRARO V, PICCIRILLO C, TOMLINS K, et al. Cassava (Manihot esculenta Crantz) and yam (Dioscorea spp.) crops and their derived foodstuffs: safety, security and nutritional value[J]. Critical Reviews in Food Science and Nutrition, 2016, 56(16): 2714 − 2727. doi: 10.1080/10408398.2014.922045 [4] PINTO-ZEVALLOS D M, PAREJA M, AMBROGI B G. Current knowledge and future research perspectives on cassava (Manihot esculenta Crantz) chemical defenses: An agroecological view[J]. Phytochemistry, 2016(130): 10 − 21. [5] ZHANG M, XIE L, YIN Z, et al. Biorefinery approach for cassava-based industrial wastes: Current status and opportunities[J]. Bioresource Technology, 2016(215): 50 − 62. [6] LÓPEZ C E, BERNAL A J. Cassava bacterial blight: Using genomics for the elucidation and management of an old problem[J]. Tropical Plant Biology, 2012, 5(1): 117 − 126. [7] ZÁRATE-CHAVES C A, GÓMEZ DE LA CRUZ D, VERDIER V, et al. Cassava diseases caused by Xanthomonas phaseoli pv manihotis and Xanthomonas cassavae[J]. Molecular Plant Pathology, 2021, 22(12): 1520 − 1537. [8] BRICKER T M, ROOSE J L, ZHANG P, et al. The PsbP family of proteins[J]. Photosynthesis Research, 2013, 116(2): 235 − 250. [9] IFUKU K. The PsbP and PsbQ family proteins in the photosynthetic machinery of chloroplasts[J]. Plant Physiology and Biochemistry, 2014, 81: 108 − 114. doi: 10.1016/j.plaphy.2014.01.001 [10] BRICKER T M, FRANKEL L K. Auxiliary functions of the PsbO, PsbP and PsbQ proteins of higher plant Photosystem Ⅱ: A critical analysis[J]. Journal of Photochemistry and Photobiology B-biology, 2011, 104(1/2): 165 − 178. [11] IFUKU K, ISHIHARA S, SHIMAMOTO R, et al. Structure, function, and evolution of the PsbP protein family in higher plants[J]. Photosynthesis Research, 2008, 98(1/2/3): 427 − 437. [12] IFUKU K, NAKATSU T, KATO H, et al. Crystallization and preliminary crystallographic studies on the extrinsic 23 kDa protein in the oxygen-evolving complex of photosystem Ⅱ[J]. Acta Crystallographica Section D, 2003, 59((Pt8)): 1462 − 1463. doi: 10.1107/S0907444903011004 [13] IDO K, IFUKU K, YAMAMOTO Y, et al. Knockdown of the PsbP protein does not prevent assembly of the dimeric PSⅡ core complex but impairs accumulation of photosystem Ⅱ supercomplexes in tobacco[J]. Biochimica et Biophysica Acta (BBA) - Bioenergetics, 2009, 1787(7): 873 − 881. doi: 10.1016/j.bbabio.2009.03.004 [14] IFUKU K, YAMAMOTO Y, ONO T A, et al. PsbP protein, but not PsbQ protein, is essential for the regulation and stabilization of photosystem Ⅱ in higher plants[J]. Plant Physiology, 2005, 139(3): 1175 − 1184. doi: 10.1104/pp.105.068643 [15] YI X, HARGETT S R, LIU H, et al. The PsbP protein is required for photosystem Ⅱ complex assembly/stability and photoautotrophy in Arabidopsis thaliana[J]. Journal of Biological Chemistry, 2007, 282(34): 24833 − 24841. doi: 10.1074/jbc.M705011200 [16] CHE Y, KUSAMA S, MATSUI S, et al. Arabidopsis PsbP-like protein 1 facilitates the assembly of the photosystem Ⅱ super complexes and optimizes plant fitness under fluctuating light[J]. Plant and Cell Physiology, 2020, 61(6): 1168 − 1180. [17] ISHIHARA S, TAKABAYASHI A, IDO K, et al. Distinct functions for the two PsbP-like proteins PPL1 and PPL2 in the chloroplast thylakoid lumen of Arabidopsis[J]. Plant Physiology, 2007, 145(3): 668 − 679. doi: 10.1104/pp.107.105866 [18] LIU J, YANG H, LU Q, et al. PsbP-domain protein1, a nuclear-encoded thylakoid lumenal protein, is essential for photosystem I assembly in Arabidopsis[J]. The Plant Cell, 2012, 24(12): 4992 − 5006. [19] ROOSE J L, FRANKEL L K, BRICKER T M. The PsbP domain protein 1 functions in the assembly of lumenal domains in photosystem[J]. The Journal of Biological Chemistry, 2014, 289(34): 23776 − 23785. doi: 10.1074/jbc.M114.589085 [20] ROOSE J L, FRANKEL L K, BRICKER T M. Developmental defects in mutants of the PsbP domain protein 5 in Arabidopsis thaliana[J]. The Public Library of Science, 2011, 6(12): e28624. [21] LIU R, CHEN T, YIN X, et al. A Plasmopara viticola RXLR effector targets a chloroplast protein PsbP to inhibit ROS production in grapevine[J]. The Plant Journal, 2021, 106(6): 1557 − 1570. doi: 10.1111/tpj.15252 -

 点击查看大图
点击查看大图

图(9) / 表 (2)
计量
- 文章访问数: 1262
- HTML全文浏览量: 791
- PDF下载量: 59
- 被引次数: 0
