


-
木薯(Manihot esculenta)属于大戟科木薯属,是一种多年生淀粉类块根作物[1],具有粗生易长、易栽培和高产等优良特性[2]。是热带及亚热带地区的三大粮食作物之一,同时也是重要的经济及能源作物[3]。
植物内生菌是指定殖于植物组织、器官内部的微生物,寄生过程中宿主植物不表现出明显的症状,并且通过表面彻底消毒后能从植物组织中分离出来,或者可以扩增到微生物核DNA证实其存在的微生物群体[4-5]。植物内生菌在目前已知的高等植物中均有一定分布,具有种类多、数量大等特点。近些年来,通过不断研究,从植物体中分离筛选得到多种内生细菌,并加以鉴定[6-7]。研究者们对植物内生细菌的多样性,种群动态等方面进行了统计研究,并且深入探讨了内生菌与植物体间的相互作用[8-9],同时在研究过程中提出了新的分类单元[10]。内生细菌在各种植物中都是普遍存在的,Lin等[11]利用高通量测序鉴定茶树不同组织微生物群落,证实茶树不同组织内生细菌分布不同。在西藏沙棘(Hippophae tibetama)中,张爱梅等[12]发现5种不同组织内生细菌具有丰富的多样性,但各组织内生细菌群落结构和优势种群有所不同,且不同组织也能够定殖具有多种功能的内生细菌。Liu等[13]对多个柑橘品种进行内生细菌多样性研究,结果表明,不同品种、不同组织等方面存在差异,内生菌群落数量和结构也有很大的差异。王雁南等[14]以葡萄为试验材料对不同部位中的内生细菌进行分离并分析,结果发现,其叶片、果肉等组织中的内生菌数量有一定差别。目前,对木薯内生菌的研究较少,为进一步探究木薯不同组织和品种内生菌种类和多样性差异,采用16S rDNA扩增子高通量测序技术分析了木薯块根、茎和叶等3种组织内生细菌群落分布特征及结构差异,旨在为挖掘有益内生菌资源提供理论支持。
-
供试验材料为木薯‘SC8’和‘SC9’品种的嫩叶、老叶、茎(韧皮部)、茎(木质部)和块根不同组织,分别命名为NL、OL、OS-BZ、OS-WL和RT。‘SC8’为淀粉木薯品种,‘SC9’为可食用木薯品种,目前是生产上的主栽品种。
-
供试木薯栽植于海南省儋州市农业农村部儋州木薯种质资源圃(20°06′N, 110°33′E),属于热带湿润季风气候,年平均太阳总辐射量110~130 kJ·cm−2,年平均降水量1 815 mm,光热资源丰富,年平均气温为23.5 ℃。
-
木薯植株的收集采用不同分布多位点法,取样时将整个植株小心挖出,从每株上分别取嫩叶、老叶、茎(韧皮部)、茎(木质部)和块根不同组织,嫩叶为木薯顶端1~3片叶,老叶为底部叶片,茎进行分段切割后用手术刀片分为韧皮部与木质部,3棵植株组织均匀混合为1个重复,1个处理3个生物学重复。将木薯各组织的样品用无菌水缓流冲洗2 min,然后用75%乙醇浸泡1 min,再用无菌水洗涤3次,擦干后用无菌锡纸包裹,于−80 ℃保存。
-
木薯各样本进行细菌DNA提取参照HiPure Stool DNA提取试剂盒(Magen, Guangzhou, China)指南进行。选择16S rDNA基因V4区设计引物,P1:5′-GTGYCAGCMGCCGCGGTAA-3'、P2:5′-GGACTACNVGGGTWTCTAAT-3'进行PCR扩增。第一轮反应体系:5×Q5@ Reaction Buffer 10.0 μL,5×Q5@ High GC Enhance 10.0 μL,2.5 mmol·L−1 dNTPs 1.5 μL,正向引物1.5 μL,反向引物1.5 μL,Q5@ High-Fidelity DNA Polymer 0.2 μL,Template DNA 50 ng,ddH2O定容至50 μL。PCR扩增条件:95 ℃ 5 min;循环数×(95 ℃ 1 min,60 ℃ 1 min,72 ℃ 1 min);72 ℃ 7 min,30个循环,10 ℃直到反应结束,使用Qubit3.0进行定量操作。第二轮反应体系:5×Q5@ Reaction Buffer 5 μL,2.5 mmol/L dNTPs 1.5 μL,5×Q5@ High GC Enhancer 1.5 μL,Index Primer (10 µmol·L−1) 1 μL,Universal PCR Primer (10 µmol·L−1) 1 μL,Q5@ High-Fidelity DNA Polymerase 1 μL,Template 50 ng,H2O定容至50 μL。第一轮扩增条件循环12次后,使用AMPure XP Beads(Merck KGaA,Germany)纯化后进行DNA浓度测定和电泳检测。将木薯各组织样品送至广州基迪奥生物科技有限公司进行16S rDNA测序。
-
测序结束后,对原始序列进行长度过滤,并筛除嵌合体后获得优化序列,进行聚类后划分新的操作分类单元OTU,将相似性 ≥ 97%的有效序列聚类成为相同的OTU。根据OTU序列与参考数据库(
https://www.ncbi.nlm.nih.gov/ )进行比对,物种注释以及分类学分析。Alpha多样性用来比较单个样品间内部物种存在的多样性,通过Beta多样性对不同样品间物种多样性进行比较。功能途径通过PICRUSt2 (Phylogenetic Investigation of Communities by Reconstruction of Unobserved States)软件进行预测,使用KEGG(Kyoto Encyclopedia of Genes and Genomes)数据库(https://www.kegg.jp/ )与测序得到的16S rDNA基因测序数据进行对比,比较生物代谢通路上的不同功能基因的丰度差异,由此得到木薯所有样品中内生细菌群落功能预测信息。统计检验均以P < 0.05为显著性,采用SPSS 26版本软件进行分析。P < 0.05为差异显著。 -
对木薯不同组织嫩叶、老叶、茎(韧皮部)、茎(木质部)和块根内生细菌产生的原始序列质控后获得V4区有效序列,‘SC8’品种中分别获得28 573、31 299、51 274、46 502和275 590条,‘SC9’品种中分别为28 017、147 902、92 973、50 319和275 229条(表1)。在97%的序列相似性水平上,‘SC8’有效序列产生的OTUs分别是2 894、3 183、3 525、5 559和3 926;‘SC9’分别是2 573、3 658、4 169、5 753和3 995。不同品种各组织内生细菌进行多样性分析结果表明,各组织内生细菌多样性指数存在差异。Chao1指数反映内生细菌群落丰富度,‘SC8’和‘SC9’各样品分别为4381.175~7219.625、3538.192~7544.341,其中,‘SC8’的茎(木质部)的内生细菌群落相较于其他样品表现出显著差异,且丰富度最高;‘SC9’各样品间的内生细菌多样性均表现出显著差异,具体表现为茎(木质部)>茎(韧皮部)>老叶>块根>嫩叶;香农指数反映内生细菌群落丰富度和均匀度,其中,‘SC8’茎(木质部)样品中的内生细菌多样性和其他组织相比明显较高。‘SC9’各样品间的内生细菌丰富度表现为茎(韧皮部)>茎(木质部)>嫩叶>老叶>块根(表1)。可见,木薯植物体样品中均含有丰富的内生细菌种类,且在茎(韧皮部)中的多样性最高,说明木薯内生细菌群落结构与木薯生态位存在着密切关系。
表 1 木薯不同组织内生细菌群落多样性指数分析
品种 样品 有效序
列数OTUs数 香农
指数Chao1
指数覆盖
率/%‘SC8’ NL 28573 2894 7.475 4381.175 95.64 OL 31299 3183 7.851 4562.337 95.76 OS-BZ 51274 3525 8.543 4535.233 97.66 OS-WL 46502 5559 10.273 7219.625 95.98 RT 275590 3926 4.493 4675.875 99.62 ‘SC9’ NL 28017 2573 7.032 3538.192 96.30 OL 147902 3658 6.666 4916.133 99.11 OS-BZ 92973 4169 7.678 5619.683 98.40 OS-WL 50319 5753 9.995 7544.341 95.99 RT 275229 3995 4.362 4818.306 99.60 -
木薯‘SC8’品种不同组织中共检测到48个门,126个纲,281个目,438个科,805个属;‘SC9’品种中存在46个门,130个纲,290个目,390个科,863个属。总体来说,木薯各组织中的内生菌多样性在各分类水平上表现出一定差异,相比于‘SC8’,‘SC9’在各分类水平上丰富度更高。在门水平上,‘SC8’嫩叶、老叶和茎(韧皮部)中的细菌群落丰富度均高于‘SC9’,表现出更高的多样性,而在属水平下,‘SC9’各组织内生细菌群落多样性均高于‘SC8’(表2)。
表 2 木薯不同组织内生细菌群落细菌学分类阶层总数
品种 样品 门 纲 目 科 属 ‘SC8’ NL 44 105 188 274 308 OL 38 99 178 234 328 OS-BZ 41 101 185 247 408 OS-WL 38 100 194 273 455 RT 40 106 200 280 482 ‘SC9’ NL 33 94 172 238 372 OL 37 105 181 254 414 OS-BZ 38 95 184 263 449 OS-WL 40 108 193 290 511 RT 37 98 192 267 488 -
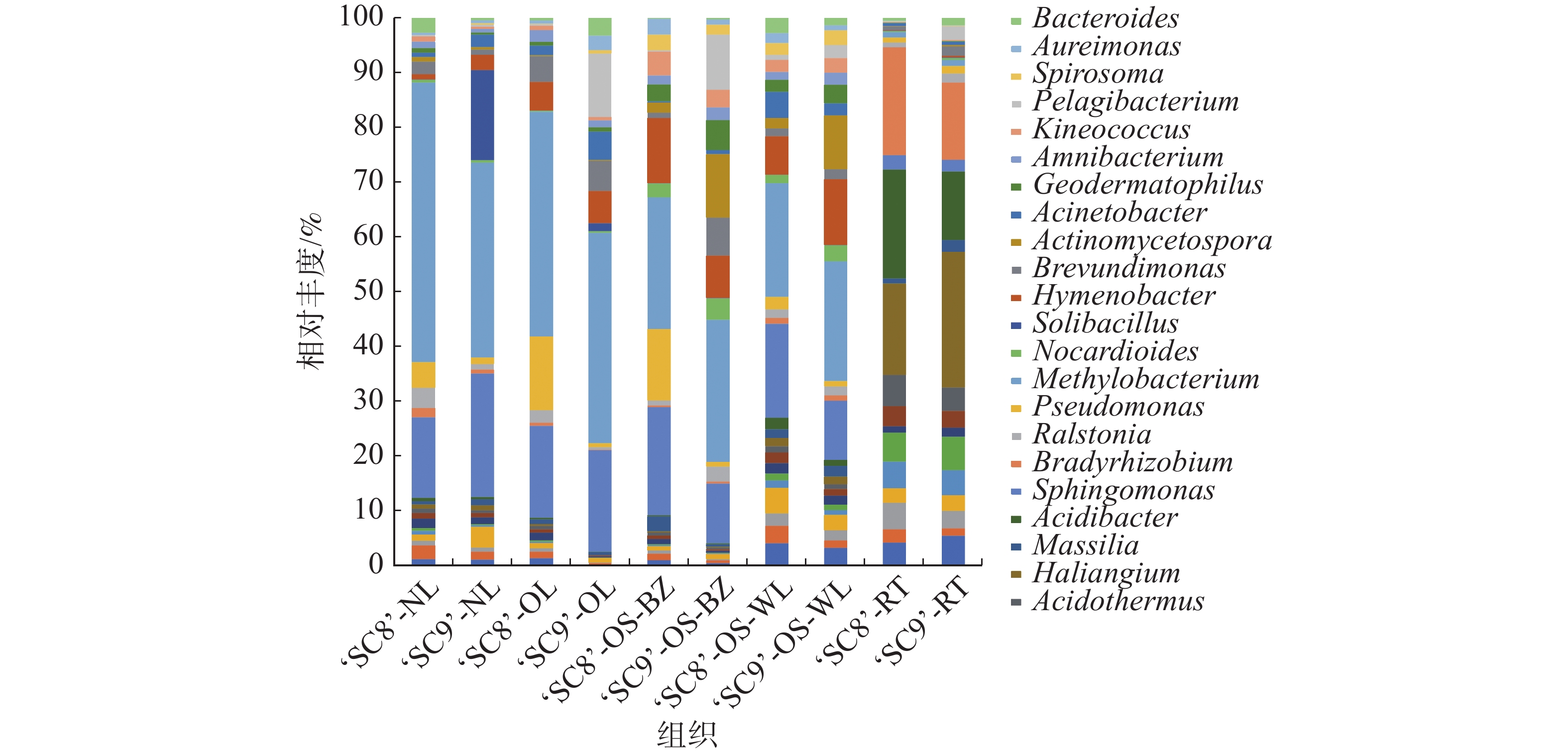
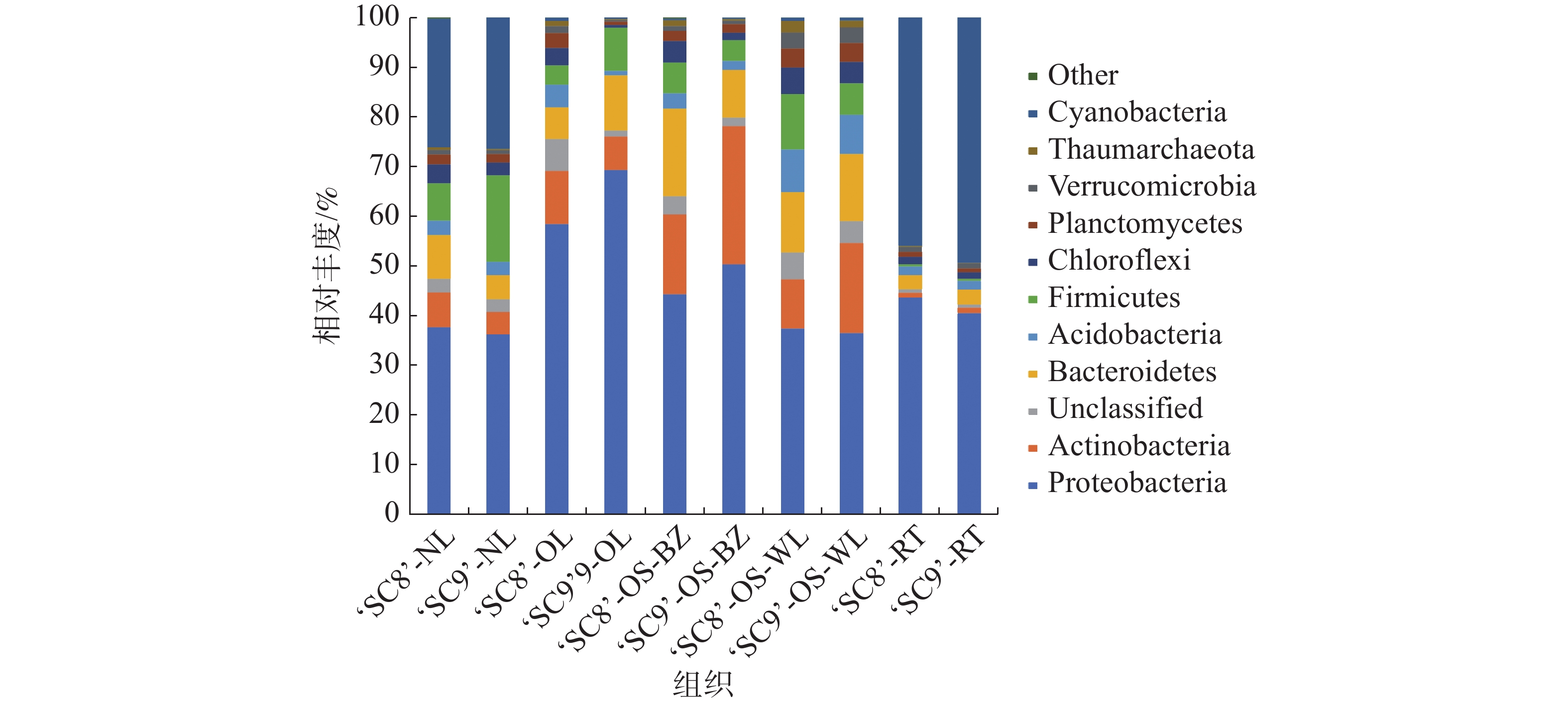
在门水平上,对于丰度高于1%的菌群进行统计,从图1可知,木薯各组织样本内生细菌群落具有相似组成结构,但各菌门所占丰度表现出一定差异。10个组织样品中的优势菌门(可归类,且至少1个组织中的OTU丰度 ≥ 10%)共分布有6个,其平均占比分别为变形菌门41.12%、蓝细菌门12.96%、放线菌门9.33%、拟杆菌门8.06%、厚壁菌门5.95%和酸杆菌门5.36%。‘SC8’嫩叶中的优势菌门为变形菌门和蓝细菌门,分别占比35.64%和24.45%;老叶中的优势菌门包括变形菌门和放线菌门,分别占比56.76%和10.36%;茎(韧皮部)为变形菌门、拟杆菌门和放线菌门,分别占比40.64%、16.16%和14.76%;茎(木质部)为变形菌门、拟杆菌门和厚壁菌门,分别占比35.43%、11.48%和10.53%;块根中为蓝细菌门和变形菌门,分别占比45.29%和43.07%。‘SC9’嫩叶中存在变形菌门、蓝细菌门和厚壁菌门3个优势菌门,分别占比35.68%、25.92%和17.12%;老叶有变形菌门和拟杆菌门2个优势菌门,相对丰度分别为68.41%和10.93%;茎(韧皮部)为变形菌门和放线菌门,相对丰度分别为48.66%和26.75%。茎(木质部)为变形菌门、放线菌门和拟杆菌门,相对丰度分别为34.58%、17.11%和12.80%;块根中的优势菌门为蓝细菌门和变形菌门,分别占比48.68%和40.07%。不同样品中优势细菌门差异很大,其中,变形菌门在所有样品中均为优势菌门,丰度为25.70%~68.41%。
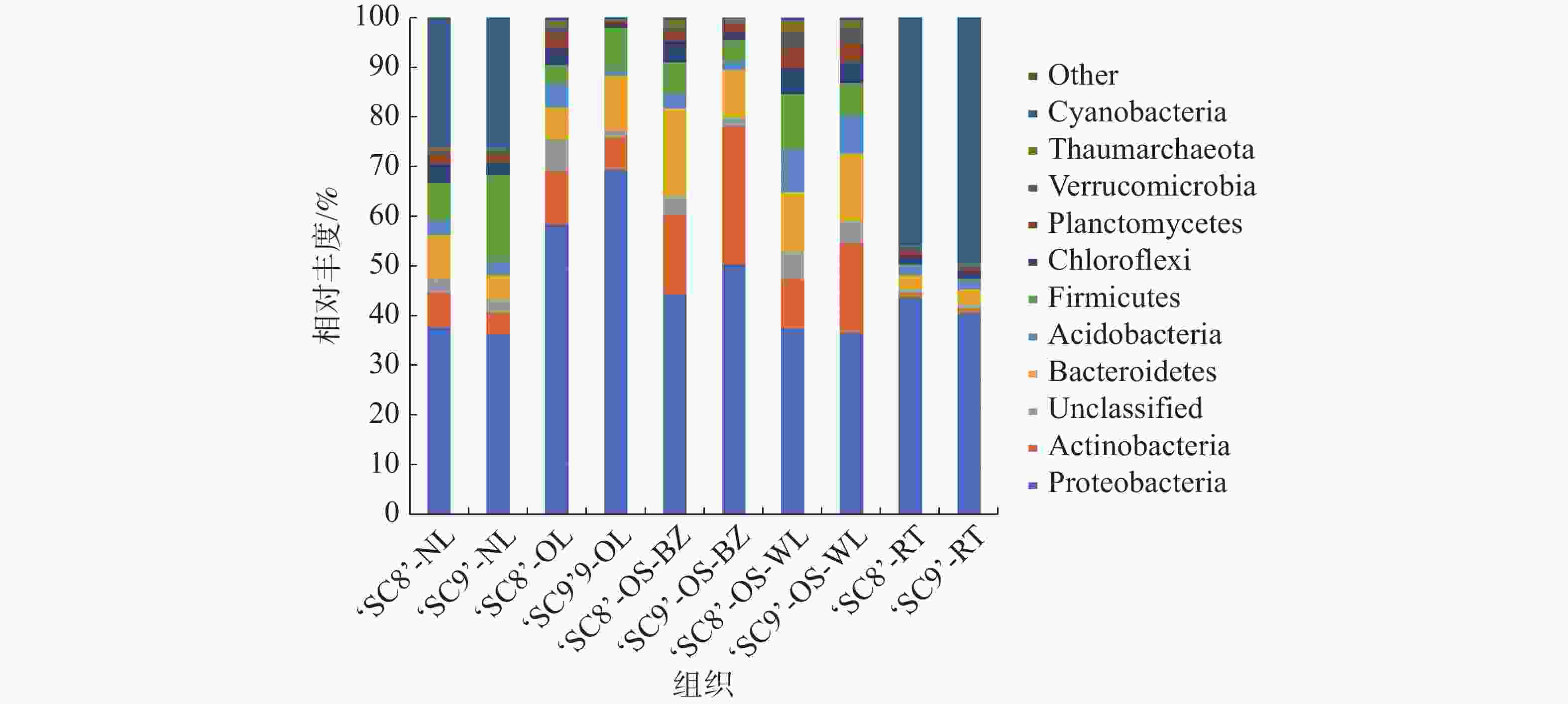
图 1 木薯不同组织内生细菌门水平的相对丰度
-
为了进一步研究木薯内生细菌群落结构组成的影响,分析了不同样品下的各内生细菌在属水平上的相对丰度(图2),以各样本中丰度占比前30名且相对丰度 > 1%的菌属作为优势菌属,‘SC8’嫩叶样本中优势菌属包括甲基杆菌属(Methylobacterium)11.77% 等3个菌属。老叶样本中包括甲基杆菌属22.16%、鞘氨醇单胞菌(Sphingomonas)9.05%、假单胞菌属(Pseudomonas)7.24%等7个菌属。茎(韧皮部)样本中包括甲基杆菌属10.67%、鞘氨醇单胞菌8.76%等10个菌属。茎(木质部)样本中包括甲基杆菌属6.88%、鞘氨醇单胞菌5.68% 等7个菌属。在‘SC9’中,嫩叶样本中包括甲基杆菌属10.26%、鞘氨醇单胞菌6.50%等4个菌属。老叶样本中包括甲基杆菌属25.03%、鞘氨醇单胞菌属12.07%、Pelagibacterium 7.55% 等8个菌属。茎(韧皮部)样本中包括甲基杆菌属15.38%、放线孢菌属(Actinomycetospora)6.85%、鞘氨醇单胞菌属6.48%等12个菌属。茎(木质部)样本中包括甲基杆菌属8.72%等10个菌属。甲基杆菌属为各组织样品中的优势菌属,平均丰度为13.86%。‘SC8’和‘SC9’均为茎(韧皮部)的内生细菌多样性最为丰富,嫩叶中最少。木薯组织间中共有171个共有菌属,占内生细菌所有种类的78.44%,其中,‘SC9’特有菌群118个,‘SC8’为100个(图3)。
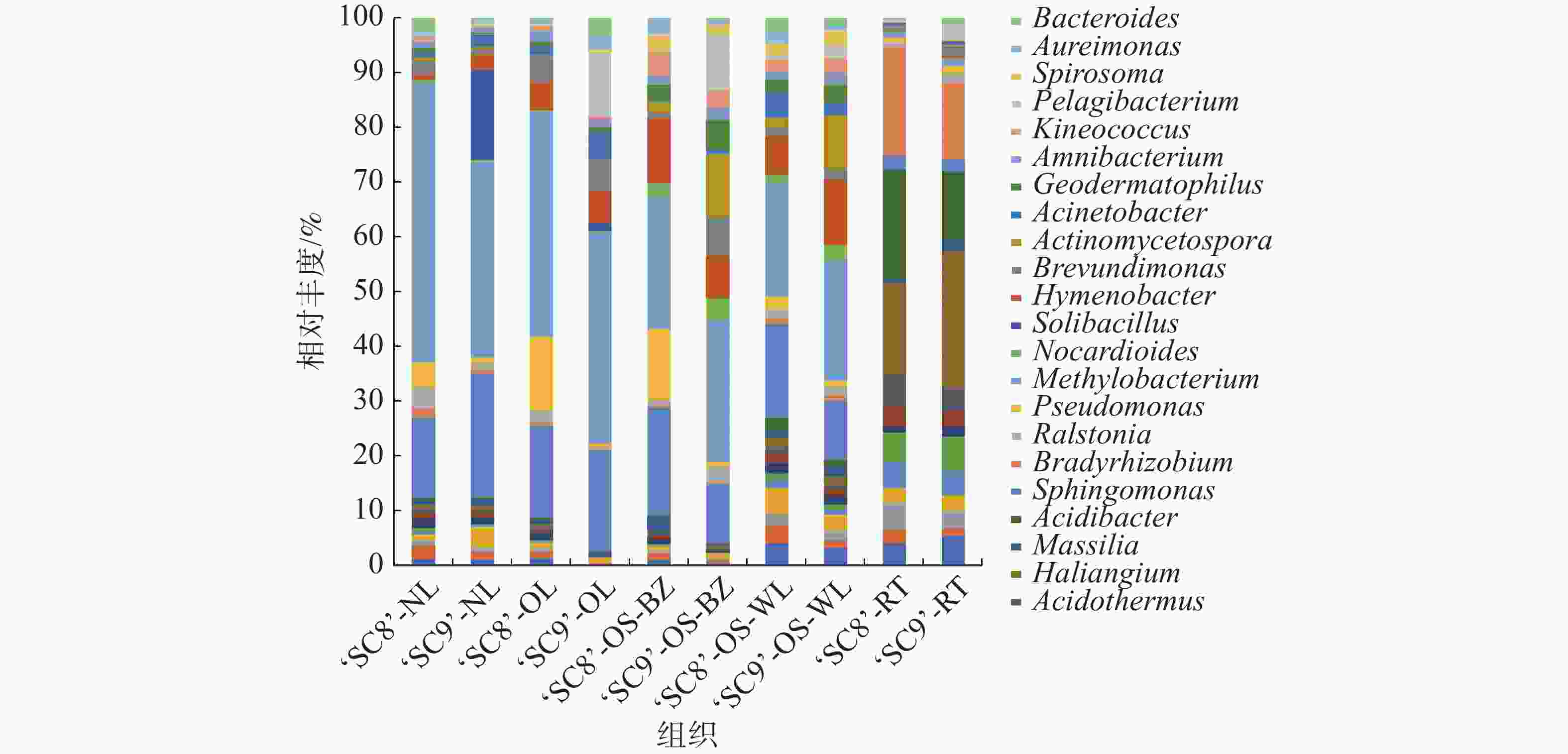
图 2 木薯各组织内生细菌属水平的群落结构组成
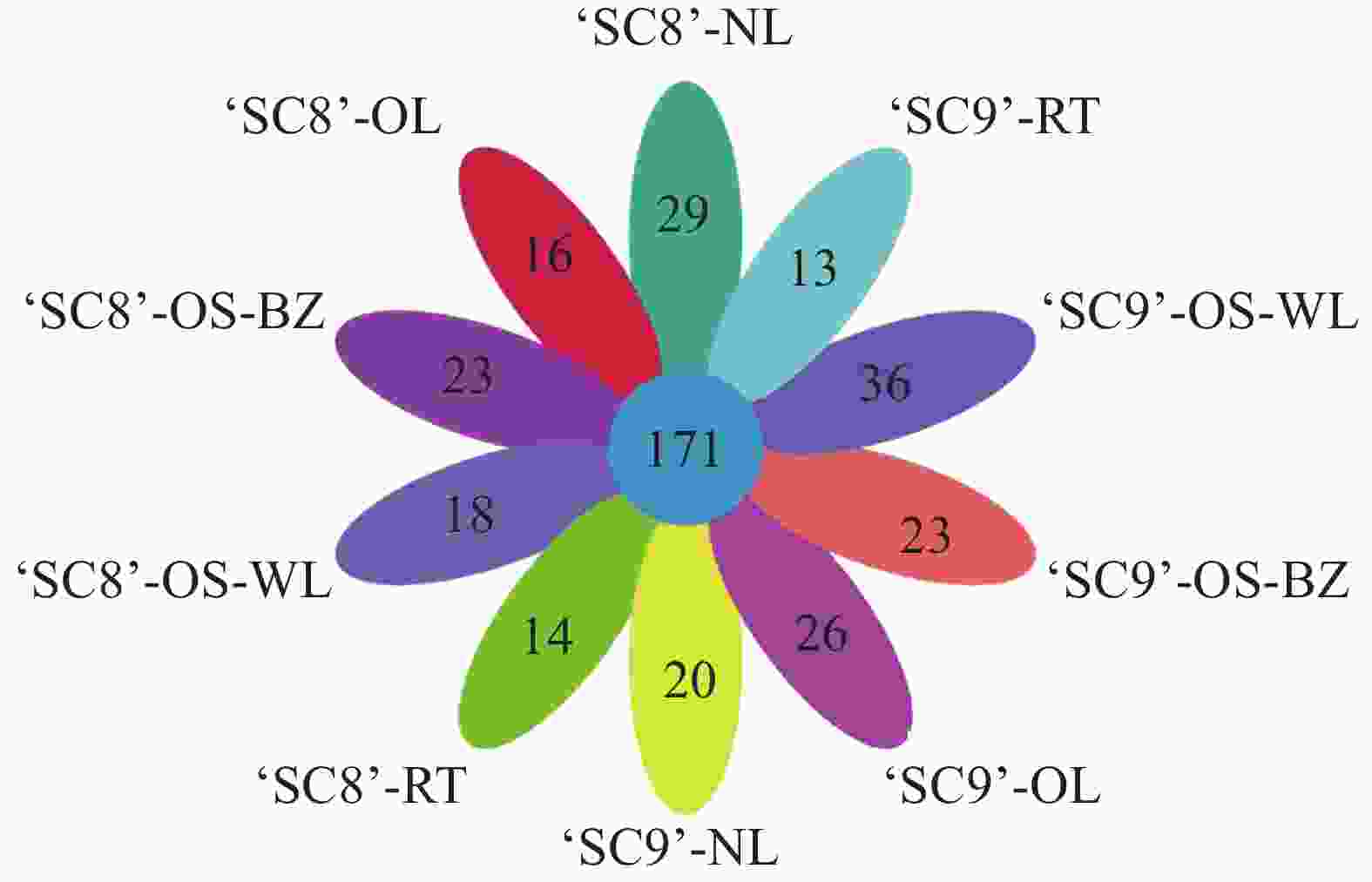
图 3 木薯各组织内生细菌属水平的物种群落结构韦恩图
-
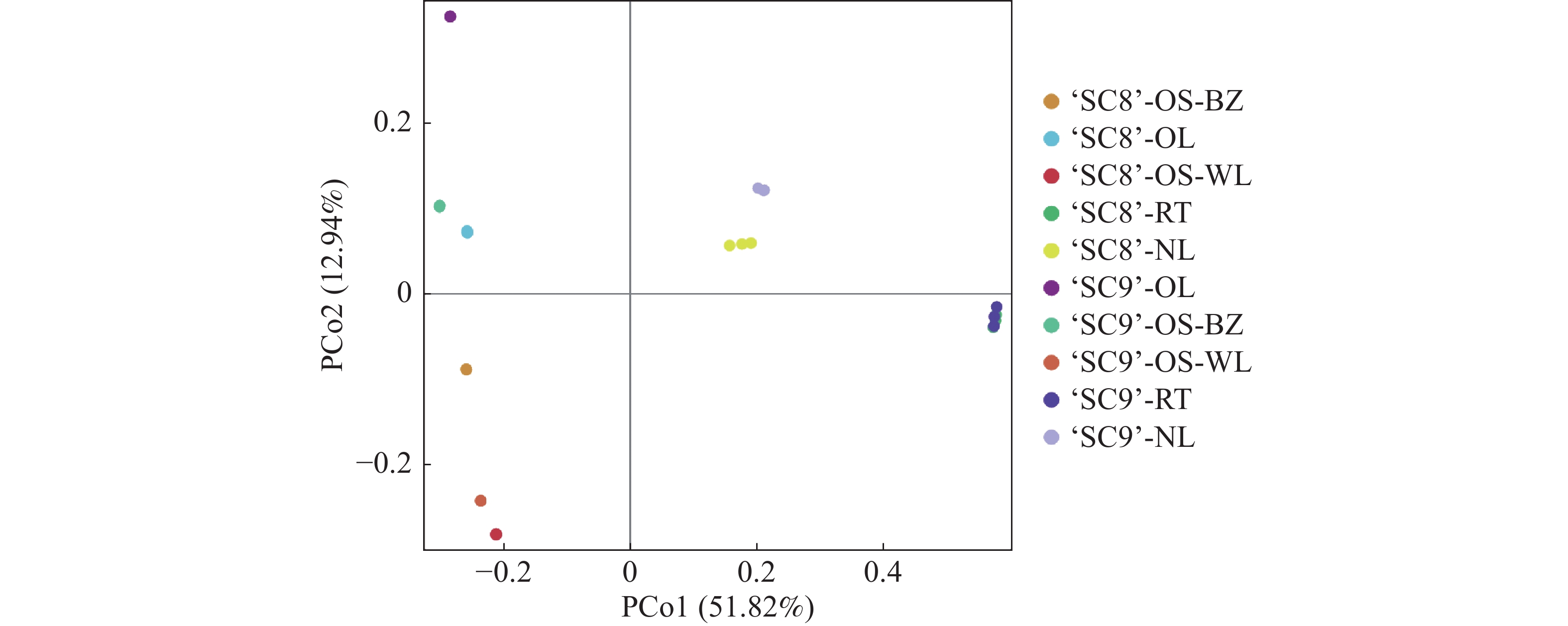
主坐标分析表明(图4),主分析1(PCoA1)和主分析2(PCoA2)的样品差异性贡献率分别达到51.82%和12.94%,合计为64.76%,是差异的主要来源。木薯不同组织内生细菌群落多样性差异较大,但‘SC9’和‘SC8’相同组织差异不明显,其中,嫩叶分布在第一象限,老叶分布在第二象限,茎(木质部)分布在第三象限,块根分布在第四象限。
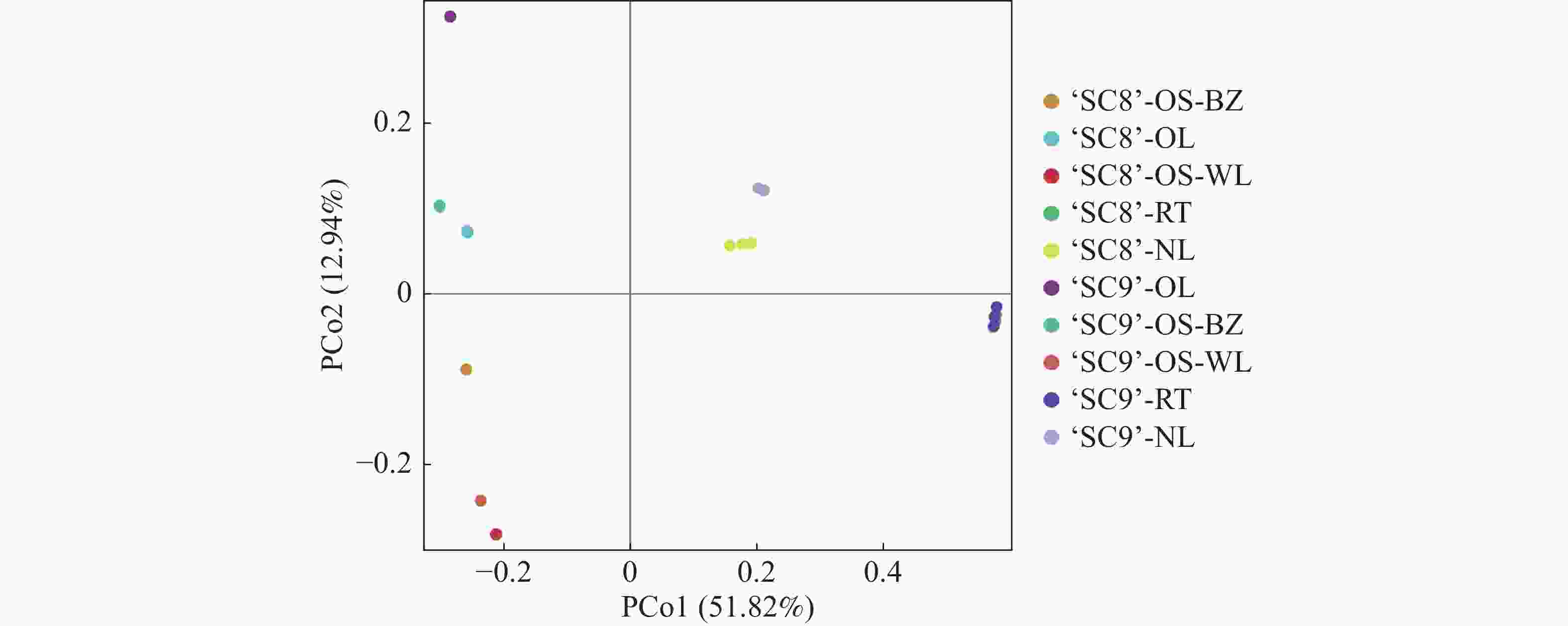
图 4 不同品种木薯内生细菌群落PCoA图
-
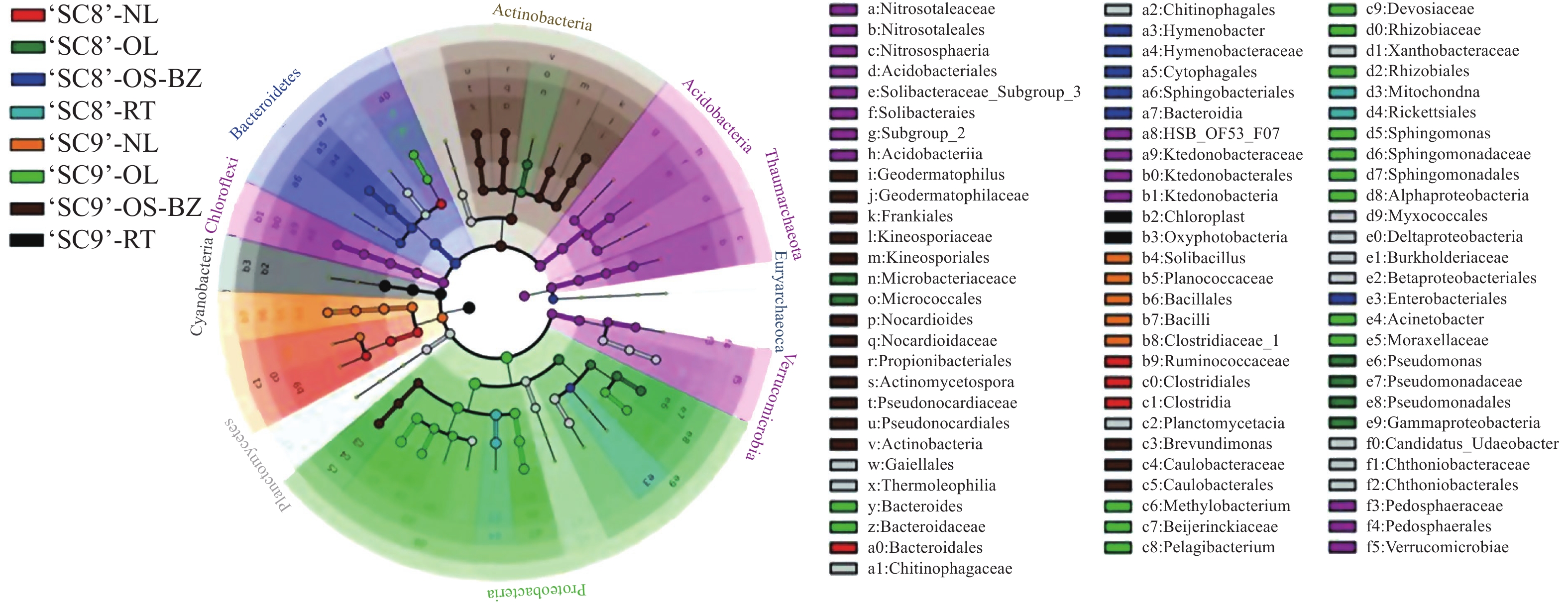
为进一步探究不同品种木薯内生细菌的差异,对供试样品进行了LEfSe(line discriminant analysis effect size)分析,设定LDA(line discriminant analysis) > 4为阈值,在‘SC8’品种中,茎(韧皮部)、老叶、嫩叶、块根得到差异显著单元分别为8、6、4、2个;‘SC9’品种中,茎(韧皮部)、老叶、嫩叶、块根内生细菌分别具有16、15、6、3个差异显著单元,2个品种的块根差异显著单元均表现为最少(图5)。在‘SC8’品种中,茎(韧皮部)内生细菌的8个差异单元,6个分布于拟杆菌门,1个分布于变形菌门,1个分布于广古菌门;老叶内生细菌的6个差异单元,4个分布于变形菌门,2个分布于放线菌门;嫩叶内生细菌的4个差异单元,3个分布于厚壁菌门,1个分布于拟杆菌门;块根内生细菌的2个差异单元均为变形菌门。‘SC9’品种中,茎(韧皮部)内生细菌的16个差异显著单元13个分布于放线菌门,3个分布于变形菌门。老叶内生细菌的差异单元有15个,13个分布于变形菌门,2个分布于拟杆菌门;嫩叶中6个差异单元均属于厚壁菌门;块根中内生细菌的3个差异单元均属于蓝细菌门。由此表明,木薯不同组织部位内生细菌的菌群存在显著差异,以茎(木质部)样品中的内生细菌最为丰富,且‘SC9’比‘SC8’内生细菌丰富。
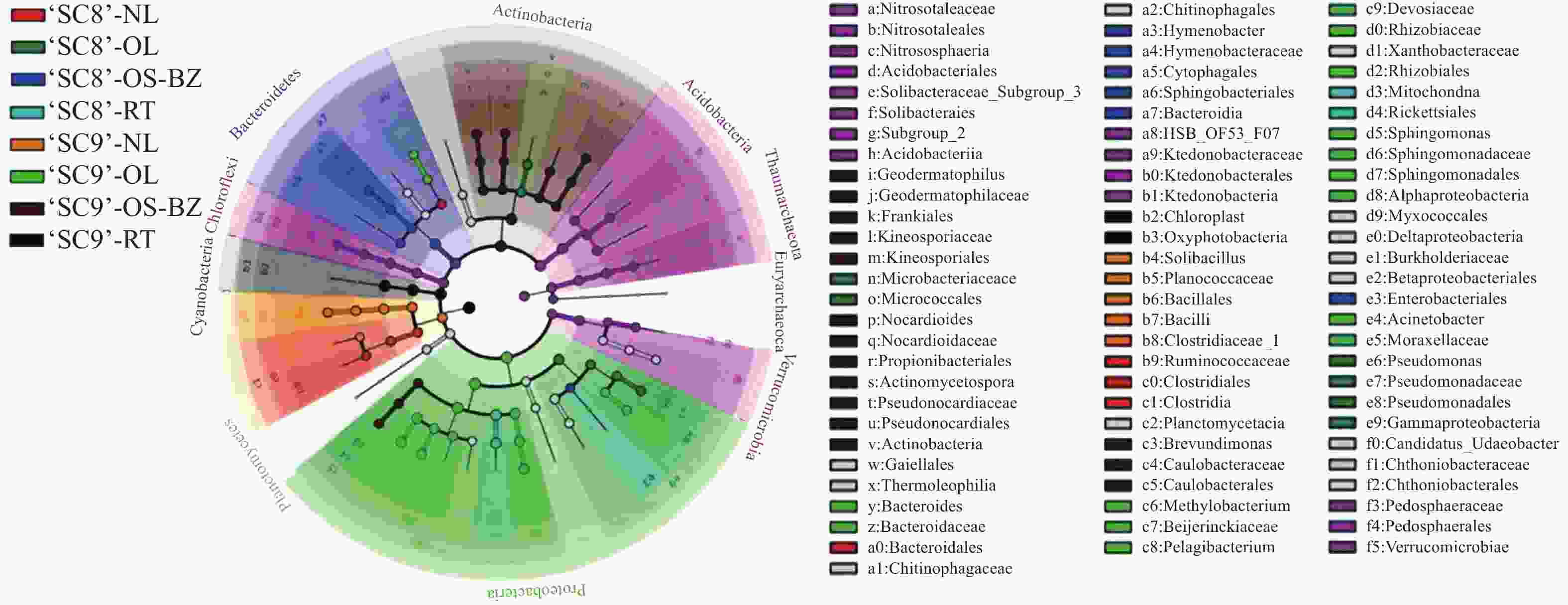
图 5 各样品细菌差异LEfSe分析图
-
通过KEGG数据库系统分析基因组注释和功能信息,发现木薯体内多种相关通路都受到内生细菌的调节,包括各种代谢通路、基因信息过程以及细胞过程等6个相关通路(表3),在‘SC8’中,各个组织在不同的相关通路中都表现出显著差异,块根参与各种功能活动的菌群丰度最高,嫩叶最低;‘SC9’各样品间表现出相同的差异性。其中,代谢相关物种所占比例最高,说明参与植物代谢是内生菌的主要功能;包含参与次级代谢产物(类黄酮、异类黄酮等),各种萜类物质以及各种氨基酸合成通路,辅酶因子、维生素以及氨基酸代谢通路和光合作用相关通路,参与调节植物的生长,间接或直接影响植物抗病能力。
表 3 不同品种木薯内生细菌群落的KEGG代谢通路丰度
品种 类别 嫩叶 老叶 茎(韧皮部) 茎(木质部) 块根 ‘SC8’ 细胞通路 57977 93919 140865 112350 310925 环境信息通路 28094 38493 62180 50147 185638 基因信息通路 156128 179853 313539 275477 1159299 人类疾病通路 2837 7859 12214 8773 27061 代谢通路 1053534 1332532 2260713 1924263 7461881 组织系统通路 4363 4927 15907 7065 42706 未分类 353 384 1034 1190 966 ‘SC9’ 细胞通路 64431 492072 279730 125438 315452 环境信息通路 29597 208048 127368 56210 191782 基因信息通路 158244 861680 596688 306544 1214512 人类疾病通路 5714 44552 29586 11415 25974 代谢通路 1074318 6784692 4650585 2241144 7807924 组织系统通路 4302 23102 14476 7717 45118 未分类 571 3726 1001 790 1035 -
内生菌广泛存在植物各组织和器官中,不仅能够促进植物生长发育[15-17]、降解重金属污染[18]、降解农药残留,还能够防治病虫草害[19]、研制生物有机肥[20]等。已有研究表明,宿主植物不同组织、不同生长阶段的内生菌具有明显差异,与组织本身的状态相关[21-22]。基于微生物16S rDNA基因的高通量测序能快速准确地处理大批数据,与传统的微生物分离、培养和鉴定方法相比,具有明显的优势,被广泛应用于微生物生态学领域[23-26]。本研究利用Illumina MiSeq高通量测序技术对不同品种木薯内生细菌多样性进行分析,结果显示,在木薯组织中,茎的内生细菌多样性大于其他组织,这与张秋玉等[27]对莼菜不同组织内生细菌多样性研究结果相似。本研究选取了木薯‘SC9’和‘SC8’ 2个当前主栽品种,‘SC9’为可食木薯,含有大量胡萝卜素,各类营养元素丰富,口感较好;而‘SC8’则具有较高的淀粉含量,是生产淀粉的主要品种之一,两者均具有较高的经济效益。通过分析得知,‘SC9’的内生菌相较于‘SC8’品种具有较高的多样性,推测菌群多样性的提高,参与改变了木薯内部营养物质的积累,从而改善了其品质,促进了木薯植株营养元素的积累。
本研究发现,木薯内生细菌的优势菌门为变形菌门、放线菌门、拟杆菌门、厚壁菌门、蓝细菌门和酸杆菌门,与其他植物的研究结果相似[27-29]。其中,变形菌门在所有组织中均为优势菌门。变形菌门在植物体内广泛分布,广泛应用于植物病虫害防治、土壤修复、生物固氮等方面[30-32]。放线菌门作为优势菌门之一,可以降解不溶性有机物质,还可以产生大量的、种类繁多的抗生素,是非常重要的生防微生物[33]。进一步统计相对丰度占比大于1%的菌属后,发现甲基杆菌属、鞘氨醇单胞菌和假单胞菌属为木薯各组织样品中的优势菌属。‘SC8’中茎(韧皮部)的内生细菌多样性最为丰富,包含10个菌属,嫩叶中最少,包含3个;‘SC9’中茎(韧皮部)中内生细菌群落多样性最高,包含12个,嫩叶中最少,包括4个。甲基营养菌是一类能够利用包含一个碳原子、或包含多个碳原子化合物作为唯一碳源和能源生长的微生物,具有固氮、溶磷、促进植物生长发育[34]、生物修复[35]等功能。鞘氨醇单胞菌在组织中相对丰度也很高,它能够降解芳香族化合物、合成生物聚合物,在环境生物领域具有广阔前景[36-37]。
植物内生菌是由环境中微生物入侵宿主,经长期协同进化后形成的稳定共生群体,因此特定环境下特定组织中内生细菌分布具有一定的专一性,宿主植物种类、生长发育阶段、区域生态类型等都会影响内生菌的群落结构[38-42]。同一品种不同组织差异较大,不同品种相同组织差异较小。两个木薯品种内生细菌群落功能富集结果显示,代谢通路相关功能基因的丰度最高,意味着内生菌群通过代谢物与寄主之间互惠互利,这与张秋玉等[27]、赵柏霞等[29]研究结果一致。同时由于木薯包含氨基酸类、脂类、多糖、萜类和酮类及其他次生代谢产物等与植物成分生物合成有关的通路,这也说明木薯内生细菌可以利用木薯中代谢产生的各种营养物质[43]。不同组织内优势菌群的差异也会对各组织的功能预测结果造成影响,茎作为营养物质的传递和保存组织,其优势菌门放线菌门丰度与碳水化合物和氨基酸代谢与转运相关基因的含量成正比,其较高丰度使木薯茎对各种营养物质的转运和代谢利用大大提高,从而促进了植物生长[44];叶作为木薯的营养器官,对木薯的生长发育至关重要,其优势菌群丰度较高,其中蓝细菌门作为其优势菌门之一,具有参与调节植物光合作用的功能,同时还可以产生多种生物活性化合物(多糖,萜类化合物,苯丙类化合物等)[45-46];作为叶片优势菌属之一,甲基杆菌的一些品种可以诱导植物叶片和根结节形成,并可通过产生生长素[47]促进植物生长。甲基杆菌大部分是甲基营养体,可以利用甲醇或其他一碳化合物作为能量来源生产蛋白质[48]。这些优势菌群通过分泌生物素或者参与植物生理生化反应,间接或者直接地参与植物生长发育,进一步影响植物的免疫功能,对其抗病性表现出调节能力。同时也发现木薯含有多种活性成分(氨基酸、可溶性糖和总黄酮等),这些活性物质的代谢相关通路同样在功能预测中有所体现。木薯内生细菌与木薯不同组织的有效化学成分合成的基因功能预测信息,可以协调木薯有效成分的合成、运输与分解,为内生细菌资源的挖掘开发和木薯块根品质的提高提供技术支持。因此,解析木薯不同组织中内生细菌与寄主间互惠互利的有效化学成分,阐明其合成通路,将为充分挖掘内生细菌资源,改善木薯抗性和品质,为创制充分利用环境微生物的木薯新种质提供试验依据。
未来将在现有工作的基础上,进一步通过改良培养基成分、培养方法等手段,尽可能多地挖掘出更多的内生细菌菌株;对其进行功能评价,发掘和筛选在促生效应、病虫害生物防治以及环境治理等方面具有较大应用潜力的微生物菌株。
Diversity of entophytic bacteria in different tissues of cassava
-
摘要: 为研究木薯(Manihot esculenta)内生细菌多样性,挖掘木薯内生细菌资源,采用16S rDNA测序技术对‘SC8’和‘SC9’2个木薯品种的块根、茎和叶等不同组织进行了Alpha和Beta多样性分析,分析细菌群落结构组成并对比Kyoto Encyclopedia of Genes and Genomes(KEGG)数据库进行内生细菌菌群基因功能预测。结果显示,从‘SC8’木薯样品中共聚类得到19,087个细菌OTUs(operational taxonomic units)隶属于48个门、126个纲、438个科和805个属;‘SC9’聚类到20,148个细菌OTUs隶属于46个门、130个纲、390个科和863个属。两品种各组织共有内生菌属171个,占属种类的78.44%,‘SC9’特有菌群118个,‘SC8’为100个,茎(韧皮部)内生菌多样性最为丰富。2品种内生细菌的优势菌群有变形菌门、蓝细菌门、放线菌门、拟杆菌门、厚壁菌门和酸杆菌门。木薯内生细菌功能中涉及多种萜类、酮类、氨基酸及维生素等次级代谢产物的生物合成。研究结果表明,木薯植物体样品中均含有丰富的内生细菌种类,且在茎(韧皮部)中的多样性最高;木薯含有多种活性成分,这些活性物质的代谢相关通路同样在功能预测中有所体现。Abstract: In order to study the diversity of endophytic bacteria in cassava (Manihot esculenta) and explore the endophytic bacteria resources, 16S rDNA sequencing technology was used to analyze the Alpha and Beta diversity of tuber, stem and leaf tissues in ‘SC8’ and ‘SC9’ cassava varieties. The structural composition of bacterial community was analyzed and the gene function of endophytic bacteria community was predicted by comparing with Kyoto Encyclopedia of Genes and Genomes (KEGG) database. The results showed that 19,087 bacterial OTUs (operational taxonomic unit) were clustered from ‘SC8’ cassava samples, and belonged to 48 phyla, 126 classes, 438 families and 805 genera, and 20,148 bacterial OTUs belonged to 46 phyla, 130 classes, 390 families and 863 genera in ‘SC9’. There were 171 common bacteria genera in all tissues of cassava, accounting for 78.44% of all endophytic bacteria species. There were 118 unique bacteria in ‘SC9’ and 100 in ‘SC8’, and endophytic bacteria in stem (phloem) had the highest diversity. The dominant bacterial groups of endophytic bacteria from cassava are Proteobacteria, Cyanobacteria, Actinobacteria, Bacteroidetes, Firmicutes and Acidobacteria. Biosynthesis of terpenoids, ketones, amino acids, vitamins and other secondary metabolites is involved in the function of cassava endophytic bacteria. These results showed that there were abundant endophytic bacteria in all cassava plant samples, and the diversity was highest in stem (phloem). Cassava contains a variety of active ingredients, and the metabolism-related pathways of these active ingredients are also reflected in the function prediction.
-
Key words:
- cassava /
- tissues /
- endophytic bacteria /
- diversity
-
表 1 木薯不同组织内生细菌群落多样性指数分析
品种 样品 有效序
列数OTUs数 香农
指数Chao1
指数覆盖
率/%‘SC8’ NL 28573 2894 7.475 4381.175 95.64 OL 31299 3183 7.851 4562.337 95.76 OS-BZ 51274 3525 8.543 4535.233 97.66 OS-WL 46502 5559 10.273 7219.625 95.98 RT 275590 3926 4.493 4675.875 99.62 ‘SC9’ NL 28017 2573 7.032 3538.192 96.30 OL 147902 3658 6.666 4916.133 99.11 OS-BZ 92973 4169 7.678 5619.683 98.40 OS-WL 50319 5753 9.995 7544.341 95.99 RT 275229 3995 4.362 4818.306 99.60  下载: 导出CSV
下载: 导出CSV
表 2 木薯不同组织内生细菌群落细菌学分类阶层总数
品种 样品 门 纲 目 科 属 ‘SC8’ NL 44 105 188 274 308 OL 38 99 178 234 328 OS-BZ 41 101 185 247 408 OS-WL 38 100 194 273 455 RT 40 106 200 280 482 ‘SC9’ NL 33 94 172 238 372 OL 37 105 181 254 414 OS-BZ 38 95 184 263 449 OS-WL 40 108 193 290 511 RT 37 98 192 267 488
下载: 导出CSV
表 3 不同品种木薯内生细菌群落的KEGG代谢通路丰度
品种 类别 嫩叶 老叶 茎(韧皮部) 茎(木质部) 块根 ‘SC8’ 细胞通路 57977 93919 140865 112350 310925 环境信息通路 28094 38493 62180 50147 185638 基因信息通路 156128 179853 313539 275477 1159299 人类疾病通路 2837 7859 12214 8773 27061 代谢通路 1053534 1332532 2260713 1924263 7461881 组织系统通路 4363 4927 15907 7065 42706 未分类 353 384 1034 1190 966 ‘SC9’ 细胞通路 64431 492072 279730 125438 315452 环境信息通路 29597 208048 127368 56210 191782 基因信息通路 158244 861680 596688 306544 1214512 人类疾病通路 5714 44552 29586 11415 25974 代谢通路 1074318 6784692 4650585 2241144 7807924 组织系统通路 4302 23102 14476 7717 45118 未分类 571 3726 1001 790 1035
下载: 导出CSV
-
[1] FALADE K O, AKINGBALA J O. Utilization of cassava for food[J]. Food Reviews International, 2010, 27(1): 51 − 83. doi: 10.1080/87559129.2010.518296 [2] 梁露锋, 玉琼广, 刘洁. 木薯产业发展动态及展望前景[J]. 大众科技, 2011, 13(6): 128 − 130. doi: 10.3969/j.issn.1008-1151.2011.06.057 [3] 孙琪, 陈霞, 贾泽冲, 等. 辐射花粉诱导木薯无融合生殖及鉴定研究[J]. 热带作物学报, 2017, 38(8): 1383 − 1389. doi: 10.3969/j.issn.1000-2561.2017.08.001 [4] 石晶盈, 陈维信, 刘爱媛. 植物内生菌及其防治植物病害的研究进展[J]. 生态学报, 2006, 26(7): 2395 − 2401. doi: 10.3321/j.issn:1000-0933.2006.07.044 [5] 张婉婷, 张灵枝. 茶树内生真菌的分离和鉴定研究进展[J]. 中国茶叶, 2011, 33(6): 7 − 9. doi: 10.3969/j.issn.1000-3150.2011.06.003 [6] 胡萌. 植物内生细菌研究进展[J]. 山东农业大学学报(自然科学版), 2008, 39(1): 148 − 151. [7] 胡泽瑞, 刘媛, 彭长连, 等. 三叶鬼针草内生细菌群体多样性及重金属耐受和吲哚乙酸产生潜力[J]. 微生物学通报, 2019, 46(1): 29 − 41. doi: 10.13344/j.microbiol.china.180314 [8] 曹焜, 王晓楠, 孙宇峰, 等. 植物根部内生细菌多样性及其生防作用研究进展[J]. 农业与技术, 2017, 37(17): 1 − 3, 5. [9] 陈梅春, 朱育菁, 刘波, 等. 基于宏基因组的茉莉花内生细菌多样性分析[J]. 热带亚热带植物学报, 2018, 26(6): 633 − 643. doi: 10.11926/jtsb.3925 [10] 刘元, 文春南, 刘淼, 等. 基于高通量测序技术的怀山药与菜山药内生细菌多样性比较[J]. 浙江农业科学, 2018, 59(10): 1754 − 1759. doi: 10.16178/j.issn.0528-9017.20181008 [11] LIN H Y, LIU C W, PENG Z, et al. Distribution pattern of endophytic bacteria and fungi in tea plants[J]. Frontiers in Microbiology, 2022, 13(9): 872034. [12] 张爱梅, 殷一然, 孔维宝, 等. 西藏沙棘5种不同组织内生细菌多样性[J]. 生物多样性, 2021, 29(9): 1236 − 1244. [13] LIU Q, YUAN M, ZHOU Y, et al. A paralog of the MtN3/saliva family recessively confers race-specific resistance to Xanthomonas oryzae in rice[J]. Plant Cell Environment, 2011, 34(11): 1958 − 1969. doi: 10.1111/j.1365-3040.2011.02391.x [14] 王雁南, 张珣. 北京红地球葡萄内生菌的种群多样性分析[J]. 安徽农业科学, 2016, 44(3): 13 − 15. doi: 10.3969/j.issn.0517-6611.2016.03.005 [15] GUERRERO Z, LÓPEZ L E, RODRÍGUEZ T A, et al. Functional diversity of plant endophytes and their role in assisted phytoremediation [A]//BHARAGAVA R N, SAXENA G. Bioremediation of industrial waste for environmental safety: Volume II: Biological agents and methods for industrial waste management [M]. Singapore: Springer, 2020: 237-255. [16] KOOMNOK C, TEAUMROONG N, RERKASEM B, et al. Diazotroph endophytic bacteria in cultivated and wild rice in Thailand[J]. Science Asia, 2007, 33(4): 429 − 435. doi: 10.2306/scienceasia1513-1874.2007.33.429 [17] THAREK M, DZULAIKHA K, SALWANI S, et al. Ascending endophytic migration of locally isolated diazotroph, Enterobacter sp. strain USML2 in rice[J]. Biotechnology(Faisalabad), 2011, 10(6): 521 − 527. doi: 10.3923/biotech.2011.521.527 [18] KLAYRAUNG S, NIAMSUP P, POONNOY P, et al. Diversity and control of bacterial contamination of plants propagated in temporary immersion bioreactor system[J]. Acta Horticulture, 2017(1115): 439 − 446. [19] 黎起秦, 焦成, 农倩, 等. 广西水稻内生细菌的动态分布及其对水稻纹枯病菌的拮抗作用[J]. 中国生物防治, 2010, 26(3): 312 − 319. [20] DEFEZ R, ANDREOZZI A, BIANCO C. The overproduction of indole-3-acetic acid (IAA) in endophytes upregulates nitrogen fixation in both bacterial cultures and inoculated rice plants[J]. Microbial Ecology, 2017, 74(2): 441 − 452. doi: 10.1007/s00248-017-0948-4 [21] DASTOGEER K M G, LI H, SIVASITHAMPARAM K, et al. Host specificity of endophytic mycobiota of wild Nicotiana plants from arid regions of northern Australia[J]. Microbial Ecology, 2018, 75(1): 74 − 87. doi: 10.1007/s00248-017-1020-0 [22] ZHANG Y, YU X, ZHANG W, et al. Interactions between endophytes and plants: beneficial effect of endophytes to ameliorate biotic and abiotic stresses in plants[J]. Journal of Plant Biology, 2019, 62(1): 1 − 13. doi: 10.1007/s12374-018-0274-5 [23] 郭丽丽, 尹伟伦, 郭大龙, 等. 油用凤丹牡丹不同种植时间根际细菌群落多样性变化[J]. 林业科学, 2017, 53(11): 131 − 144. doi: 10.11707/j.1001-7488.20171115 [24] ABOU-SHANAB R A I, BERKUM P, ANGLE J S, et al. Characterization of Ni-resistant bacteria in the rhizosphere of the hyperaccumulator Alyssum murale by 16S rRNA gene sequence analysis[J]. World Journal of Microbiology & Biotechnology, 2010, 26(1): 101 − 108. [25] WANG F, MEN X, ZHANG G, et al. Assessment of 16S rRNA gene primers for studying bacterial community structure and function of aging flue-cured tobaccos[J]. AMB Express, 2018, 8(1): 182. doi: 10.1186/s13568-018-0713-1 [26] 朱启良, 刘洪凯, 陈旭, 等. 杨树人工林根际古菌群落随细根生长的演变[J]. 应用生态学报, 2019, 30(3): 849 − 856. doi: 10.13287/j.1001-9332.201903.034 [27] 张秋玉, 陈晶, 周雪妹, 等. 莼菜不同组织内生细菌多样性的比较分析[J]. 中南民族大学学报(自然科学版), 2022, 41(3): 285 − 291. [28] 吴燕燕, 徐伟芳, 罗琴, 等. Illumina MiSeq高通量测序分析不同品种桑树内生细菌多样性[J]. 蚕学通讯, 2018, 38(3): 1 − 10. doi: 10.3969/j.issn.1006-0561.2018.03.001 [29] 赵柏霞, 闫建芳. 高通量技术分析‘砂蜜豆’甜樱桃不同组织内生细菌多样性 [J]. 中国农业科技导报: 2023, 25(3): 66−77. [30] BODENHAUSEN N, HORTON M W, BERGELSON J, et al. Bacterial communities associated with the leaves and the roots of Arabidopsis thaliana [J/OL]. PLoS One, 2013, 8(2): e56329. [31] RANGJAROEN C, RERKASEM B, TEAUMROONG N, et al. Comparative study of endophytic and endophytic diazotrophic bacterial communities across rice landraces grown in the highlands of northern Thailand[J]. Archives of Microbiology, 2014, 196(1): 35 − 49. doi: 10.1007/s00203-013-0940-4 [32] SHI Y, YANG H, ZHANG T, et al. Illumina-based analysis of endophytic bacterial diversity and space-time dynamics in sugar beet on the north slope of Tianshan Mountain[J]. Applied Microbiology and Biotechnology, 2014, 98(14): 6375 − 6385. doi: 10.1007/s00253-014-5720-9 [33] MENENDEZ E, CARRO L.Actinobacteria and their role as plant probiotics [M]//GIRI B, PRASAD R, WU Q S, et al. Biofertilizers for Sustainable Agriculture and Environment. Cham: Springer International Publishing, Cham: Springer, 2019: 333−351. [34] 王改萍, 祝长青, 王茹. 一株耐盐甲基杆菌Methylobacterium sp. W-1的分离及促生潜能研究[J]. 微生物学通报, 2021, 48(11): 4134 − 4144. [35] 田丹丹, 宋修鹏, 蒋承健, 等. 1株甲基杆菌Methylobacterium sp. WGM16的鉴定及降解甲醇的最佳培养条件[J]. 微生物学杂志, 2011, 31(1): 28 − 33. doi: 10.3969/j.issn.1005-7021.2011.01.007 [36] 黄海东, 刘云, 刘如林. 合成生物聚合物的重要微生物资源鞘氨醇单胞菌[J]. 微生物学报, 2009, 49(5): 561 − 567. [37] 台喜生, 冯佳丽, 李梅, 等. 鞘氨醇单胞菌在生物降解方面的研究进展[J]. 湖南农业科学, 2011(7): 29 − 33. doi: 10.3969/j.issn.1006-060X.2011.07.007 [38] SHEN S Y, FULTHORPE R. Seasonal variation of bacterial endophytes in urban trees[J]. Frontiers in Microbiology, 2015, 6: 427. [39] DING T, MELCHER U. Influences of plant species season and location on leaf endophytic bacterial communities of non-cultivated plants [J]. PLoS One, 2016, 11(3): e0150895. [40] DE OLIVEIRA COSTA L E, DE QUEIROZ M V, BORGES A C, et al. Isolation and characterization of endophytic bacteria isolated from the leaves of the common bean (Phaseolus vulgaris)[J]. Brazilian Journal of Microbiology, 2012, 43(4): 1562 − 1575. doi: 10.1590/S1517-83822012000400041 [41] SHI Y W, LOU K, LI C, et al. Illumina-based analysis of bacterial diversity related to halophytes Salicornia europaea and Sueada aralocaspica[J]. Journal of Microbiology, 2015, 53(10): 678 − 685. doi: 10.1007/s12275-015-5080-x [42] HAMEED A, YEH M W, HSIEH Y T, et al. Diversity and functional characterization of bacterial endophytes dwelling in various rice (Oryza sativa L.) tissues, and their seed-borne dissemination into rhizosphere under gnotobiotic P-stress[J]. Plant and Soil, 2015, 394(1/2): 177 − 197. [43] 谢红炼, 汪汉成, 蔡刘体, 等. 烟草种子内生细菌群落结构与多样性[J]. 微生物学报, 2020, 60(3): 601 − 616. doi: 10.13343/j.cnki.wsxb.20190275 [44] XU L, NAYLOR D, DONG Z, et al. Drought delays development of the sorghum root microbiome and enriches for monoderm bacteria[J]. Proceedings of the National Academy of sciences of the United States of America, 2018, 115(18): E4284 − E4293. [45] SINGH R K, TIWARI S P, RAI A K, et al. Cyanobacteria: an emerging source for drug discovery[J]. Journal of Antibiotics, 2011, 64(6): 401 − 412. [46] ARMSTRONG L, VAZ M G M V, GENUáRIO D B, et al. Volatile compounds produced by Cyanobacteria isolated from mangrove environment[J]. Current Microbiology, 2019, 76(5): 575 − 582. doi: 10.1007/s00284-019-01658-z [47] KELLY D P, MCDONALD I R, WOOD A P. The family Methylobacteriaceae[J]. Prokaryotes, 2014: 313 − 40. [48] CAO Y R, WANG W, JIN R X, et al. Methylobacterium soli sp. nov. a methanol-utilizing bacterium isolated from the forest soil[J]. Antonie Van Leeuwenhoek, 2011, 99(3): 629 − 634. doi: 10.1007/s10482-010-9535-0 -

 点击查看大图
点击查看大图
图(5) / 表 (3)
计量
- 文章访问数: 1639
- HTML全文浏览量: 983
- PDF下载量: 38
- 被引次数: 0
