


 下载:
下载:








-
木薯(Manihot esculenta Crantz)是大戟科木薯属多年生灌木。作为世界六大粮食作物之一[1],广泛种植于热带地区[2 − 3]。木薯有许多不同于其他粮食作物的优势[4],如无需种子萌发,可通过茎秆扦插进行无性生殖[5],且木薯抗干旱、耐贫瘠、无需精耕细作[6],具有长期发展潜力与研究价值。然而,木薯在生长过程中常受多种病害危害,严重影响其产业的发展,如木薯花叶病毒病(cassava mosaic disease, CMD)。CMD对木薯来说是一种毁灭性病害,感染后会导致田间减产30%~40%,且同一植株会被多种病毒反复感染,对我国乃至全球的木薯生产构成了极大的威胁[7],其中木薯普通花叶病毒(cassava common mosaic virus, CsCMV)是CMD致病病毒之一[8]。该病毒编码5个蛋白,分别为编码RNA依赖性RNA聚合酶(RNA-dependent RNA polymerase, RdRp),3个运动蛋白(triple gene block, TGB)和结构外壳蛋白(coat protein, CP)[9]。而目前,利用生物科学技术分离鉴定抗病基因,解析其抗病机制,创制抗病种质是目前防治CMD最有效的方法[10]。
当植物遭受病害危害或面临复杂环境时,各种胁迫严重影响植物正常生长发育,使细胞代谢异常,导致植物体内产生大量活性氧(reactive oxygen species, ROS)物质,例如超氧阴离子、羟基自由基和过氧化氢等[11],从而破坏生物膜的结构和功能[12 − 13]。为了有效清除细胞内多余的ROS,植物进化出复杂的抗氧化系统,降低和消除氧化胁迫,从而保证植物正常生长发育。植物抗氧化系统大致可分为两类:第一类是抗氧化剂,如抗坏血酸(ascorbic acid, AsA)和谷胱甘肽(glutathione, GSH)等;第二类是抗氧化酶,如超氧化物歧化酶(superoxide dismutase, SOD)等[14]。其中,抗坏血酸过氧化物酶(ascorbate peroxidase, APX)参与抗坏血酸–谷胱甘肽(ascorbate-glutathione, ASC-GSH)循环清除植物体内积累的过氧化氢物质[15]。目前,已有多种植物如盐地碱蓬(Suaeda salsa)[16]、白桦(Betula platyphylla)[17]、黑麦草(Lolium perenne)[18]、番茄(Solanum lycopersicum)等的APX基因被克隆[19],并证实APX在抗氧化胁迫中的重要作用。目前有研究表明木薯遭遇胁迫发生响应时,木薯中APX活性也会发生变化[20]。当ROS的清除速度提升时,木薯对低温的耐受性也会增强[21]。而关于APX在木薯中免疫调控作用仍未见有关报道,木薯如何响应病毒侵染及抵御病毒的免疫系统网络尚未清楚。
本研究克隆MeAPX2并构建其抑制和过表达植株,对其进行木薯花叶病毒侵染,同时对相关基因转录水平及酶活等生理指标进行测定,以期探究MeAPX2在木薯抗普通花叶病毒中的功能,同时解析MeAPX2在木薯抗普通花叶病中的作用机制。其次证实MeAPX2靶向结合CsCMV中的CP,且MeAPX2的酶活可被CP抑制,以期筛选出抗病新品种、确保木薯产业的稳定发展起到一定作用。
-
实验所使用的过表达载体为pEGAD,抑制载体为pTRV1和pTRV2,蛋白表达载体为pET32a,酵母相关载体为pGBKT7、pGADT7。大肠杆菌DH5α感受态(DL1001,上海唯地),农杆菌 GV3101感受态细胞(AC1001,上海唯地),BL21(DE3)感受态细胞(CD601,TRANS)。实验材料所用木薯品种均为‘华南124’,栽培体积比例为v营养土∶v蛭石=2∶1,置于海南三亚崖州独村实验室露天培养。
-
选取15~20 cm,有3~4个芽眼且大小基本一致的木薯茎秆放入花盆中,种植3~4周(平均温度28~32 ℃)。将生长状况一致的木薯苗叶片接种病毒,每个处理设置3个生物学重复,最后将木薯叶片进行取样,用液氮速冻,置于−80 ℃超低温冰箱保存。
-
根据南京建成生物工程研究所抗坏血酸过氧化物酶(APX)测试盒(A123-1-1,南京建成)对蛋白进行酶活性测定。按照说明书配置体系,最后加100 μL待测样品。将上述反应体系混匀后立即在37 ℃条件下水浴2 min,以沸水失活的蛋白为空白对照,最终在290 mm处测定数据,并计算酶活。进行3次生物学重复,并记录相关数值。酶活性定义:每mg重组蛋白每min在每mL体系中催化1 μmol底物为1个活力单位(U)。测定木薯叶片的APX酶活,将处理后的0.1 g叶片置于研钵中,加入缓冲液0.9 mL,冰水浴匀浆,离心后取上清,视作蛋白重复上述操作。
-
根据CTAB粗提取法提取木薯叶片RNA[22],再参照试剂盒(11151ES10,YEASEN)反转录成cDNA。
-
根据 Phytozome数据库中木薯MeAPX2 基因的序列信息,设计全长扩增引物MeAPX2-F(ATGCCGAAGAACTACCCAAAA)和MeAPX2-R(TTACGCCTCAGCAAATCCGA)。以木薯cDNA为模板,扩增MeAPX2的编码区(Coding sequences, CDS),凝胶电泳检测扩增产物。将PCR产物利用胶回收试剂盒回收,连接到所需载体上构建MeAPX2相关载体,进行后续实验。
-
根据序列目的基因设计荧光定量PCR引物(qMeAPXs-F/R),RT-qPCR反应为15 μL,包括:TB Green Premix Ex Taq Ⅱ Fast qPCR(2×)为7.5 μL,正反引物各0.3 μL,cDNA模板2 μL,ddH2O 4.9 μL。扩增程序:95 ℃预变性3 min,95 ℃变性15 s,55 ℃退火30 s,72 ℃延伸20 s,循环48次。以木薯MeEF1a为内参基因,用2−ΔΔCT 法计算MeAPXs基因相对转录水平表达量。
-
过表达和抑制植株 将构建好的pEGAD-MeAPX2和pTRV2-MeAPX2质粒转入根癌农杆菌GV3101菌株中并注射木薯叶片。提取对照组与实验组叶片RNA并反转录成cDNA作为模板,以木薯MeEF1a为内参基因,采用RT-qPCR的方法分析MeAPX2在植株中的相对表达情况[23],确定过表达和抑制植株是否构建成功。
-
将酵母表达质粒pGBKT7-A、pGADT7-B转化到酵母AH109菌株的感受态中,并分别涂在SD/-Leu-Trp(
630417 ,Clontech)和SD/-Leu-Trp-His-Ade(630428 ,Clontech)的平板上,于28 ℃条件下培养。分别挑取SD/-Leu-Trp-His-Ade平板上的酵母单菌落至SD/-Leu-Trp培养液中培养,当菌液OD600达到1时,提取酵母质粒并进行PCR验证。将验证正确的单菌落在超净工作台中梯度稀释并点板至SD/-Leu-Trp和SD/-Leu-Trp-His-Ade板培养基上,28 ℃条件培养4 d后,进行拍照记录[24]。 -
将目的片段通过同源连接到的pET32a载体上并测序验证。验证成功后将质粒转入DE3菌株中,选取阳性克隆,采用LB培养液培育,当OD600=0.6,加入IPTG后继续培养6 h,收集菌体、破碎,离心后上清即所需蛋白[25]。
-
将目的蛋白、A/G磁珠(L00277,GenScript)、抗体混合于离心管中孵育过夜。用PBS缓冲液洗涤混合物,以除去未结合到磁珠上的蛋白。加入上样缓冲液后,将离心管于沸水中煮5 min,最后通过蛋白质印迹检测蛋白质[26]。
-
本研究中所有数据均来自3个生物学重复,每个生物重复的样品均来自多个植株叶片。不同字母代表样本间存在显著性差异,P<0.05。用Graphpad对数据进行处理,用SPSS、Figdraw(
https://www.figdraw.com/static/index.html )进行绘图。 -
为探究木薯中APX酶活在木薯普通花叶病毒侵染过程中的变化,收集并测定病毒侵染后不同时间点(0、3、6、9、12、24 hpi)的木薯叶片的APX酶活,发现APX酶活在3 hpi时有所增加,而随着侵染时间的延长,其活性逐步下降(图1),说明CsCMV的侵染会影响木薯体内APX酶活性。

图 1 CsCMV侵染过程APX的酶活
Figure 1. Enzyme activity of APX in cassava during CsCMV infection
在病毒侵染过程,APX酶活发生改变,推测MeAPXs的转录水平可能也发生改变。经检测木薯体内MeAPXs的表达量发现,随着侵染时间的增加,只有MeAPX2表达量逐渐下降,而MeAPX的家族其他成员(MeAPX1和MeAPX3)的表达量无显著变化(图2)。结果表明,MeAPX2能染够响应CsCMV的侵染,并且可能参与木薯的免疫应答反应。因此选其作为后续研究对象。

图 2 MeAPXs响应CsCMV侵染的相对转录水平
Figure 2. The relative transcript level of MeAPXs in response to CsCMV infection
-
在木薯抗普通花叶病毒中的功能分析 为了探究MeAPX2的生物学功能,首先构建其抑制植株。RT-qPCR结果表明,MeAPX2在MeAPX2-RNAi植株中的转录水平显著低于对照组,抑制率达到40%以上(图3)。而其同家族的MeAPX1和MeAPX3在MeAPX2-RNAi植株中的转录水平相较于对照组无显著变化(图3),说明成功构建MeAPX2-RNAi植株。

图 3 MeAPX2-RNAi植株中MeAPXs的转录水平
Figure 3. Expression level of MeAPXs in the MeAPX2-RNAi plants
为了研究MeAPX2 是否参与调节木薯对CsCMV的免疫反应,在MeAPX2-RNAi和CK叶片中分别注射木薯普通花叶病毒的侵染性克隆,并选取不同侵染时间点(0 、7、14 dpi)样品RT-qPCR分析RdRp和TGB1的相对转录水平。结果发现,MeAPX2-RNAi植株中的RdRp和TGB1 表达量在7 、14 dpi时均显著高于CK,是CK的1.5倍(图4)。同时,MeAPX2-RNAi植株中APX酶活在7、14 dpi时均显著低于CK(图4)。观察表型发现,MeAPX2-RNAi植株整体萎蔫程度比CK组更明显,顶端叶片会更黄且叶片卷曲变形严重(图5)。这些结果表明MeAPX2正调控木薯对CsCMV的免疫反应。

图 4 MeAPX2-RNAi植株中的病毒积累量和APX酶活
Figure 4. The accumulation of virus and APX enzyme activity in the MeAPX2-RNAi plants

图 5 CsCMV侵染MeAPX2-RNAi植株表型
Figure 5. Phenotype of MeAPX2-RNAi plants infected with CsCMV infection
-
与木薯普通花叶病毒编码蛋白互作验证 为了进一步探究MeAPX2在病毒中的靶蛋白,点对点分析其与CsCMV编码的蛋白互作情况。分别构建RdRp-pGBKT7、TGB1-pGBKT7、TGB2-pGBKT7、TGB3-pGBKT7、CP-pGBKT7质粒并与MeAPX2-pGADT7分别进行酵母点对点实验,结果发现,只有共转MeAPX2-pGADT7与CP-pGBKT7组合在SD/-Leu-Trp-His-Ade培养基长出了酵母单菌落,而共转MeAPX2-pGADT7与RdRp-pGBKT7、TGB1-pGBKT7、TGB2-pGBKT7、TGB3-pGBKT7组合则不能在SD/-Leu-Trp-His-Ade培养基生长(图6−A),说明MeAPX2与CP在酵母体内互作。为了进一步验证MeAPX2 与CP之间的相互作用,构建MeAPX2-pET32a、CP-pET32a表达载体,进行Pull-down试验发现,MeAPX2在Input和IP组均可以通过Myc抗体检测出条带,而GFP只能在Input组检测出条带,IP组则无法检测出对应条带(图6−B),结果表明MeAPX2与CP可直接发生互作。

图 6 MeAPX2与CP互作
Figure 6. Interaction of MeAPX2 and CP
-
与木薯普通花叶病毒CP蛋白互作的体外功能分析 为了进一步探究MeAPX2与CP互作的生物学功能,首先分析其对MeAPX2的体外酶活的影响。在50 μL体系中保持MeAPX2总蛋白浓度不变,随着CP蛋白浓度的增加,MeAPX2的酶活逐渐下降,而对照组中GFP蛋白浓度的增加而未改变MeAPX2的酶活(图7)。实验结果表明,CP蛋白会体外抑制MeAPX2的酶活。

图 7 CP体外抑制MeAPX2酶活
Figure 7. CP inhibited the enzyme activity of MeAPX2 in vitro
-
与木薯普通花叶病毒CP蛋白互作的体内功能分析 为了进一步分析CP与MeAPX2互作的体内生物学功能,构建MeAPX2 过表达植株并RT-qPCR分析其表达量,发现MeAPX2在MeAPX2-OE植株中的转录水平显著高于对照组,为对照组的20倍以上(图8)。而MeAPX1和MeAPX3在MeAPX2-OE中的转录水平相较于对照组无显著变化(图8),说明成功构建MeAPX2-OE植株。
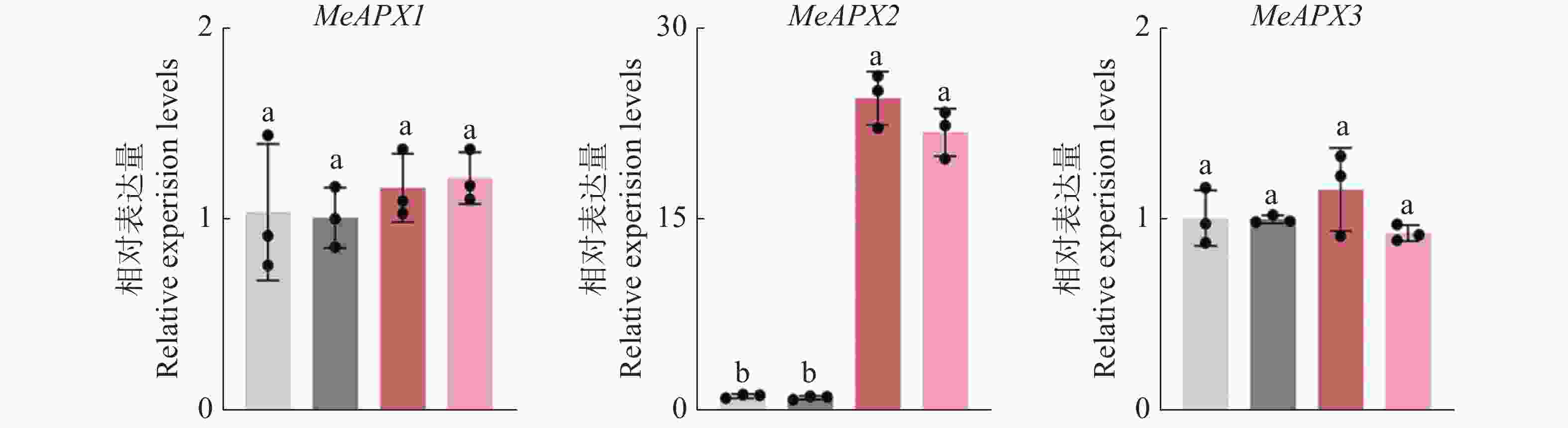
图 8 MeAPX2-OE植株中MeAPXs的表达量
Figure 8. The transcript level in the MeAPXs at MeAPX2-OE plants
在MeAPX2-OE和对照植株中分别喷施等量CP和GFP蛋白后,接种CsCMV并统计病毒积累量发现,外源喷施CP蛋白植株的RdRp和TGB1 转录水平在7 、14 dpi时均显著高于对照组(图9)。同时,外源喷施CP组的APX酶活在7、14 dpi时均显著低于对照组(图9)。观察接种CsCMV 14 dpi的表型发现,喷施CP蛋白的植株顶端叶片卷曲且更黄,而外源施加GFP蛋白组的植株为叶片黄化(图10)。结果表明,CP抑制MeAPX2 介导的免疫反应。

图 9 MeAPX2-OE植株中的病毒积累和APX酶活
Figure 9. The accumulation of virus and APX enzyme activity in the MeAPX2-OE plants

图 10 CP影响MeAPX2-OE植株表型。
Figure 10. The phenotypes of the MeAPX2-OE plants are affected by CPcts
-
研究表明,植物病毒侵染宿主时,会严重影响植物自身代谢和生长发育,可导致活性氧相关的过氧化氢酶和超氧化物歧化酶活性增加,这两种酶会平衡植物组织中活性氧的积累以应对病原体的侵染[27]。APX作为植物体内参与免疫反应重要的过氧化物酶,也可能参与了宿主对病毒的免疫应答。本研究发现,在CsCMV侵染下,APX酶活先上升再下降。在逆境条件下,植物的抗性与其体内APX基因的表达以及酶活性水平具有密切关系[28]。同时有研究表明,APX基因家族成员转录水平的快速应答会维持细胞质中高的APX活性,抵御逆境最初侵染时导致细胞膜质过氧化[29],以保护细胞成分免受ROS诱导的应激氧化损伤[30]。综上所述,APX在植物抵御逆境胁迫时发挥着重要的作用。最初APX活性升高,可维持较高的抗逆性,而后病毒的复制逐渐抑制MeAPX2的表达和APX活性,细胞内动态平衡被破坏,免疫防线逐渐削弱,病毒侵染程度逐渐加重。
为了成功侵染宿主,病毒始终与宿主免疫系统不断并持续地发生“军备竞赛”。在病毒与宿主植物相互作用的过程中,特定病毒蛋白与寄主蛋白直接相互作用发挥着重要作用。而不同病毒的编码蛋白破坏宿主免疫系统的途径和靶蛋白存在差异。在小麦中,小麦黄花叶病毒(Wheat yellow mosaic virus, WYMV)编码的P1蛋白通过干扰钙调蛋白相关的免疫防御,从而促进其在小麦中的侵染[31]。而在本实验中,通过酵母点对点验证和Pull-down实验,证实MeAPX2靶向木薯花叶病毒的编码蛋白CP,且其酶活性被CP抑制。研究结果将为深入解析CsCMV与木薯间的攻防机制提供新线索和木薯抗病分子育种提供候选基因。
随着木薯在世界粮食作物及经济作物的地位逐渐提高,而CMD作为木薯的1种毁灭性病害,严重影响了世界范围内木薯的生产[32 − 33]。根据王国芬等[11]的调查结果,CsCMV的单独侵染的检出率高达78.8%,表明CsCMV引起的病害在我国主要木薯栽培区已经相当普遍,应当引起足够的重视,并开展有关研究,提高木薯产率及稳定木薯品质。前有研究已证明在番茄[34]、拟南芥[35]、苹果[36]、水稻[37]等研究表明APX的可增强植物的抗逆性,但在木薯中抗普通花叶病毒病的功能还未有深入研究,因此探究木薯中MeAPX2抗普通花叶病毒病的生物学功能具有十分重要的意义。同时,研究表明APXs参与多种逆境胁迫,这对后续探究MeAPX2是否参与木薯的其他胁迫反应,以期培育广谱抗性木薯优异种质。
-
本研究首先确定MeAPX2 是木薯抗CsCMV的正向调节因子,并阐明其发挥作用的分子机制。木薯遭受CsCMV侵染时,MeAPX2 的表达被显著抑制,同时APX酶活降低,表明MeAPX2参与木薯免疫系统对抗CsCMV中发挥重要作用。通过酵母双杂交及Pull-down实验证实MeAPX2会靶向结合CsCMV编码蛋白CP,植株体内APX酶活被抑制,破坏木薯免疫系统,成功侵染宿主。
Functional analysis of MeAPX2 gene in cassava resistance to cassava common mosaic disease
-
摘要: 抗坏血酸过氧化物酶(ascorbate peroxidase,APX)在植物抵御逆境胁迫中有显著作用。为了探究APX在木薯抗普通花叶病毒病(cassava common mosaic virus, CsCMV)中的功能,通过分析CsCMV侵染过程中,APX酶活和MeAPXs的表达量发现,APX酶活下降,且MeAPX2的表达量显著下降,因此确定MeAPX2为候选基因进行进一步研究。通过构建MeAPX2抑制植株和指标测定,发现MeAPX2正调控木薯对CsCMV的抗性。进一步蛋白互作分析发现MeAPX2靶向结合CsCMV的外壳蛋白(coat protein,CP),且体内体外实验证实CP可抑制MeAPX2的活性。本研究结果显示,CsCMV侵染木薯过程会抑制APX酶活,MeAPX2通过直接靶向结合CP蛋白正调控木薯对CsCMV的抗性。Abstract: Ascorbate peroxidases (APXs) play a significant role in plant resistance to abiotic stress. An attempt was made to investigate the role of APXs in cassava resistance to cassava common mosaic virus (CsCMV). APX activity and the expression level of MeAPXs during infection of cassava with CsCMV were analyzed. The results showed that APX activity and MeAPX2 transcript level were reduced. Therefore, MeAPX2 was identified as a candidate gene for further analysis. MeAPX2 positively regulated cassava resistance to CsCMV, as determined by the disease index in MeAPX2-silenced plants. Further analysis revealed that MeAPX2 directly interacts with the coat protein (CP) of CsCMV. Moreover, CP was found to inhibit the activity of MeAPX2 both in vivo and in vitro. The results demonstrated that CsCMV suppresses APX activity during infection in cassava. Furthermore, MeAPX2 positively regulates cassava resistance to CsCMV by targeting CP.
-
Key words:
- Manihot esculenta Crantz /
- MeAPX2 /
- cassava common mosaic virus /
- coat protein /
- functional analysis
-
图 2 MeAPXs响应CsCMV侵染的相对转录水平
取CsCMV侵染木薯0、3、6、9、12、24 hpi 的叶片进行MeAPXs转录水平分析。
Fig. 2 The relative transcript level of MeAPXs in response to CsCMV infection
After cassava was infected with CsCMV, the leaves were collected and used for the analysis of the transcript levels of MeAPXs at 0, 3, 6, 9, 12, 24 hpi.
图 3 MeAPX2-RNAi植株中MeAPXs的转录水平
RT-qPCR检测MeAPXs在MeAPX2-RNAi植株中的相对转录水平。MeAPXs在对照组中的转录水平设为1。
Fig. 3 Expression level of MeAPXs in the MeAPX2-RNAi plants
The transcript level of MeAPXs in the MeAPX2-RNAi plants was analysed via RT-qPCR. The relative expression of MeAPXs at the control plant was set as 1.
图 4 MeAPX2-RNAi植株中的病毒积累量和APX酶活
在MeAPX2-RNAi植株中接种CsCMV的侵染性克隆,在0、7、14 dpi时收集叶片并进行生理指标测定。
Fig. 4 The accumulation of virus and APX enzyme activity in the MeAPX2-RNAi plants
The MeAPX2-RNAi plants were infected with the infectious clone of CsCMV, and then the leaves were collected and used to determine the physiological indicators at 0, 7, 14 dpi.
图 5 CsCMV侵染MeAPX2-RNAi植株表型
MeAPX2-RNAi植株在CsCMV侵染14 dpi后生长表型。木薯整株的比例尺为10 cm,木薯叶片的比例尺为1 cm。
Fig. 5 Phenotype of MeAPX2-RNAi plants infected with CsCMV infection
Phenotype of MeAPX2-RNAi plants after infected with CsCMV infection at 14 dpi. The scale bar of the whole cassava plant is 10 cm, and the scale bar of the cassava leaves is 1 cm.
图 6 MeAPX2与CP互作
A:MeAPX2与CP在酵母中互作。共转融合表达质粒的AH109菌株在SD/-Trp/-Leu/-His/-Ade培养基上进行培养筛选。B:Pull-down证实MeAPX2与CP在体外互作。使用Myc抗体检测携带MeAPX2,并使用Flag检测共免疫沉淀的CP-Flag。
Fig. 6 Interaction of MeAPX2 and CP
A:MeAPX2 interacted with CP in yeast. Yeast strain AH109 cotransformed with plasmids expressing the indicated fusion proteins was plated on SD/-Trp/-Leu/-His/-Ade. B: Pull-down assay showed the interaction of MeAPX2 and CP. Myc antibody was used to detect MeAPX2, and Flag antibody was used to detect co-immunoprecipitated CP-Flag.
图 7 CP体外抑制MeAPX2酶活
将定量的MeAPX2蛋白(6 μg)和CP蛋白按不同比例混合并检测MeAPX2的酶活,以GFP蛋白为对照组。
Fig. 7 CP inhibited the enzyme activity of MeAPX2 in vitro
A certain amount(6 μg)of was mixed with different contents of CP protein to test the enzyme activity of MeAPX2 with the GFP protein as the control.
图 8 MeAPX2-OE植株中MeAPXs的表达量
RT-qPCR检测MeAPXs在MeAPX2-OE植株中的相对转录水平。以MeAPXs在对照组中的相对转录水平设为1。
Fig. 8 The transcript level in the MeAPXs at MeAPX2-OE plants
The relative expression levels of MeAPXs in the MeAPX2-OE plants were detected via RT-qPCR. The relative expression of MeAPXs in the control plants was set as 1.
图 9 MeAPX2-OE植株中的病毒积累和APX酶活
在MeAPX2-OE植株中连续喷施CP蛋白7 d后,进行CsCMV处理,并在0、7、14 dpi时取样和生理指标。以等量GFP蛋白为对照。
Fig. 9 The accumulation of virus and APX enzyme activity in the MeAPX2-OE plants
The MeAPX2-OE plants were per-treated with CP protien for 7 d, and then infected with CsCMV. The leaves were collected and used to determine the physiological indicators at 0, 7, 14 dpi. The GFP protein was used as the control.
图 10 CP影响MeAPX2-OE植株表型。
GFP为对照,CP介导MeAPX2过表达植株在接种CsCMV 14 dpi 后生长表型对比图。木薯整株的比例尺为10 cm,木薯叶片的比例尺为1 cm。
Fig. 10 The phenotypes of the MeAPX2-OE plants are affected by CPcts
Pictures of the symptom of the MeAPX2-OE plants mediated by GFP and CP were taken at 14 dpi after CsCMV infection. The scale bars of the whole cassava plant and leaves are 10 cm and 1 cm, respectively.
-
[1] SHAN Z, LUO X, WU M, et al. Genome-wide identification and expression of GRAS gene family members in cassava[J]. BMC Plant Biology, 2020, 20(1): 46. [2] 安飞飞, 罗秀芹, 薛晶晶, 等. 白心与黄心糖木薯代谢物及相关基因表达分析[J]. 江西农业大学学报, 2022, 44(5): 1092 − 1100. [3] 王春华. 木薯在饲料中的应用现状及发展趋势[J]. 中国饲料添加剂, 2018(1): 24 − 25. [4] EL-SHARKAWY M A. Cassava biology and physiology[J]. Plant Molecular Biology, 2004, 56(4): 481 − 501. [5] 蔡赋涵. 木薯渣两相耦合产乙酸工艺研究 [D]. 南宁: 广西大学, 2012. [6] SHAN Z, LUO X, WEI M, et al. Physiological and proteomic analysis on long-term drought resistance of cassava (Manihot esculenta Crantz)[J]. Scientific Reports, 2018, 8(1): 17982. [7] 时涛, 王国芬, 李超萍, 等. 木薯花叶病在中国发生的首次报道[J]. 热带农业科学, 2018, 38(10): 99 − 100. [8] 王国芬, 李超萍, 时涛, 等. 我国木薯花叶病毒病的发生危害及其病原鉴定[J]. 热带作物学报, 2021, 42(6): 1668 − 1677. doi: 10.3969/j.issn.1000-2561.2021.06.023 [9] YU N T, YANG Y, LI J H, et al. Complete genome sequence of a distinct isolate of cassava common mosaic virus (CsCMV) infecting cassava in Hainan, China[J]. European Journal of Plant Pathology, 2020, 158(2): 583 − 587. [10] HOUNGUE J A, ZANDJANAKOU-TACHIN M, NGALLE H B, et al. Evaluation of resistance to cassava mosaic disease in selected African cassava cultivars using combined molecular and greenhouse grafting tools[J]. Physiological and Molecular Plant Pathology, 2019, 105: 47 − 53. [11] WASZCZAK C, CARMODY M, KANGASJäRVI J. Reactive oxygen species in plant signaling[J]. Annual Review of Plant Biology, 2018, 69: 209 − 236. [12] CHEN H, SHI Y, AN L, et al. Overexpression of SlWRKY6 enhances drought tolerance by strengthening antioxidant defense and stomatal closure via ABA signaling in Solanum lycopersicum L[J]. Plant Physiology and Biochemistry, 2024, 213: 108855. [13] LIU X, JI P, YANG H, et al. Priming effect of exogenous ABA on heat stress tolerance in rice seedlings is associated with the upregulation of antioxidative defense capability and heat shock-related genes[J]. Plant Growth Regulation, 2022, 98(1): 23 − 38. [14] ZHANG Z, HOU X, GAO R, et al. CsSHMT3 gene enhances the growth and development in cucumber seedlings under salt stress[J]. Plant Molecular Biology, 2024, 114(3): 52. doi: 10.1007/s11103-024-01451-y [15] 刘备, 宋玉梅, 孙铭, 等. 燕麦劣变种子吸胀过程中线粒体AsA-GSH循环的生理响应[J]. 草地学报, 2021, 29(2): 211 − 219. [16] MA CL, WANG PP, CAO ZY, et al. cDNA cloning and gene expression of APX in Suaeda salsain response to salt stress[J]. Journal of Plant Physiology and Molecular Biology, 2002, 28(4): 261 − 266. [17] WANG C, YANG CP, WANG YC. Cloning and expression analysis of an APX gene from Betula platyphylla[J]. Northeastern Naturalist, 2009, 37(3): 79 − 81. [18] MENG Q, YAN M, ZHANG J, et al. Humic acids enhance salt stress tolerance associated with pyrroline 5-carboxylate synthetase gene expression and hormonal alteration in perennial ryegrass (Lolium perenne L.)[J]. Frontiers in Plant Science, 2023, 14: 1272987. [19] WANG Y, WISNIEWSKI M, MEILAN R, et al. Overexpression of cytosolic ascorbate peroxidase in tomato confers tolerance to chilling and salt stress [J]. Journal of the American Society for Horticultural Science, 130(2): 167−173. [20] SHANG S, TANG Y, DAI J, et al. Genomic analysis of the principal members of antioxidant enzymes in simulated stresses response and postharvest physiological deterioration in cassava[J]. Tropical Plant Biology, 2021, 14(4): 419 − 428. [21] FU H, ZHAO Y, HUANG J, et al. Reduced glutathione and raffinose lengthens postharvest storage of cassava root tubers by improving antioxidant capacity and antibiosis[J]. BMC Plant Biology, 2023, 23(1): 475. [22] 董亚彬, 白玉晶. 木薯过氧化氢酶基因的克隆与原核表达[J]. 分子植物育种, 2024, 22(15): 4912 − 4918. [23] JIANG Z, ZHANG K, LI Z, et al. The Barley stripe mosaic virus γb protein promotes viral cell-to-cell movement by enhancing ATPase-mediated assembly of ribonucleoprotein movement complexes[J]. PLoS Pathogens, 2020, 16(7): e1008709. doi: 10.1371/journal.ppat.1008709 [24] 石波明. 巴西橡胶树橡胶转移酶HRT2互作蛋白的筛选与表达分析 [D]. 海口: 海南大学, 2021. [25] 张也, 韦运谢. 木薯MeLTP基因克隆及原核表达分析[J/OL]. 分子植物育种, 2024, 1-10. [26] 陈耀. 营养应激对TRIB2蛋白稳定性的调控作用及其分子机制研究 [D]. 镇江: 江苏大学, 2020. [27] WU J, KAMANGA B M, ZHANG W, et al. Research progress of aldehyde oxidases in plants[J]. PeerJ, 2022, 10: e13119. doi: 10.7717/peerj.13119 [28] 曾秀存, 孙万仓, 方彦, 等. 白菜型冬油菜抗坏血酸过氧化物酶(APX)基因的克隆、表达及其活性分析[J]. 作物学报, 2013, 39(8): 1400 − 1408. [29] MISHRA P, BHOOMIKA K, DUBEY R S. Differential responses of antioxidative defense system to prolonged salinity stress in salt-tolerant and salt-sensitive Indica rice (Oryza sativa L.) seedlings[J]. Protoplasma, 2013, 250(1): 3 − 19. doi: 10.1007/s00709-011-0365-3 [30] 杜佳庚, 周瑾, 李晓燕, 等. 芹菜APX基因家族鉴定及其表达分析[J]. 干旱地区农业研究, 2023, 41(6): 54 − 63. [31] CHEN D, ZHANG H Y, HU S M, et al. The P1 protein of wheat yellow mosaic virus exerts RNA silencing suppression activity to facilitate virus infection in wheat plants[J]. The Plant Journal, 2023, 116(6): 1717 − 1736. doi: 10.1111/tpj.16461 [32] TUO D, ZHOU P, YAN P, et al. A cassava common mosaic virus vector for virus-induced gene silencing in cassava[J]. Plant Methods, 2021, 17(1): 74. doi: 10.1186/s13007-021-00775-w [33] 吴金山, 王思琦, 黄家权, 等. 5株生防菌定殖能力、促生作用及其对木薯细菌性枯萎病生防潜力的研究[J]. 热带作物学报, 2022, 43(9): 1880 − 1887. [34] CHEN X, HAN H, CONG Y, et al. The protective effect of exogenous ascorbic acid on photosystem inhibition of tomato seedlings induced by salt stress[J]. Plants, 2023, 12(6): 1379. doi: 10.3390/plants12061379 [35] 李泽琴, 李锦涛, 邴杰, 等. 拟南芥APX家族基因在植物生长发育与非生物逆境胁迫响应中的作用分析[J]. 遗传, 2019, 41(6): 534 − 549. [36] LI M, MA F, GUO C, et al. Ascorbic acid formation and profiling of genes expressed in its synthesis and recycling in apple leaves of different ages[J]. Plant Physiology and Biochemistry, 2010, 48(4): 216 − 224. doi: 10.1016/j.plaphy.2010.01.015 [37] 鲁振强. 水稻APX与CAT同工酶的功能特性及其与盐胁迫关系的研究 [D]. 哈尔滨: 东北林业大学, 2005. -

 点击查看大图
点击查看大图
计量
- 文章访问数: 705
- HTML全文浏览量: 360
- 被引次数: 0
