

 下载:
下载:


-
火龙果( Hylocereus undatus )原产中美洲热带地区,是仙人掌科(Cactaceae)量天尺属(Hylocereus)或蛇鞭柱属(Selenicereus)植物,喜光喜热。火龙果在我国的种植面积已达到6.7万公顷,在热区经济中已占有重要地位[1]。火龙果是长日照作物,在我国因冬季光照时长不足,不能开花,为此在生产上可采用夜晚补光的方式诱导火龙果开花结果。bHLH转录因子蛋白,包含1个碱性DNA结合区和1个螺旋-环-螺旋(HLH)区,能通过与顺式启动子元件相结合来调节基因表达,参与植物开花、种子发芽、矿质营养与非生物胁迫、应激反应、光信号、激素信号、光和植物激素之间的相互作用等多种生理过程[2]。
bHLH基因家族参与调控植物的开花调控,在拟南芥、番茄等植物中已有广泛的研究。在成花起始阶段,拟南芥的FBH蛋白(bHLH类型转录因子),可与光周期信号途径的关键基因CO (CONSTANS)启动子的E-box顺式元件结合,激活光周期开花基因的共转录[3]。在花发育阶段,与CIB1同源的bHLH蛋白,可以作为激活子直接促进成花素基因FT的转录[4]。bHLH家族的光敏色素互作因子PIFs可以与光信号直接作用,PIF3在光感受器信号网络中直接与光敏色素反应,PIF4在高温下直接激活成花素基因FT[5]。bHLH48和bHLH60与PIF7联合,可以作为赤霉素途径中的正调控因子,正调控GA介导的开花[6]。bHLH38/100/101会干扰CO的转录,调节FT的表达,进而影响拟南芥的开花[7]。bHLH基因家族成员SlbHLH22的过表达,会促进番茄早开花[8]。在茉莉酸途径中,JA激活的bHLH转录因子MYC2, MYC3和MYC4冗余调控拟南芥的开花[9]。拟南芥的bHLH家族基因BEE1具有诱导开花起始的功能,BES1-BEE1-FT可以调节光周期并与FT的激活因子CO互作来影响植物开花[10]。bHLH基因家族CRY2、CIB1可以响应蓝光,能将生物钟信号转化为开花信号,促进CO基因形成蛋白质复合物,诱导花的形成[11]。除了调节开花时间外,bHLH基因家族成员的SPT基因可以在花器官形成中起作用[12]。
目前,bHLH基因家族在拟南芥(Arabidopsis thaliana)[13]、小麦(Triticum aestivum L.)[14]、谷子(Setaria italica L.)[15]、烟草(Nicotiana tabacum L.)[16]、柚子(Citrus maxima)[17]等植物中进行了全基因组鉴定。火龙果作为一种重要的热带水果,有独特的开花习性,bHLH家族基因可能发挥重要作用,但目前还未见火龙果HubHLH基因家族相关系统分析的研究报道。火龙果(Hylocereus undatus)全基因组序列的发表,为鉴定火龙果HubHLH基因家族提供了条件[18]。本研究拟利用火龙果(Hylocereus undatus)基因组数据,对火龙果HubHLH转录因子家族进行全基因组筛选,并进行理化性质、系统进化、基因表达等方面的比较分析;利用火龙果补光诱导开花的转录组数据,研究HubHLH基因家族在火龙果冬季补光诱导开花过程的表达响应,旨在解析火龙果冬季补光诱导开花的分子机制。
-
分别在
http://www.pitayagenomic.com/index.php 网址和Ensembal plants核酸数据库(https://plants.ensembl.org/index.html )中下载火龙果和拟南芥的基因组及注释文件。从已报道的文章中查找拟南芥bHLH 蛋白质序列作为种子序列,blast后去掉重复的火龙果基因号,提取蛋白质序列,通过注释信息,筛选得到火龙果候选bHLH基因。从 PFAM 数据库(http://pfam.xfam.org/ )中下载bHLH (PF00010)基因家族的隐马尔科夫模型,在HMMER 3.0软件中比对,得到火龙果候选bHLH基因。取BLASTP和HMM两种方式获得的bHLH基因交集,并通过NCBI conserved Domains(http://www.ncbi.nlm.nih.gov/Structure/cdd/wrpsb.cgi )数据库、SMART (http://smart.ebl-heidelberg.de/ )数据库,进一步确定所有候选bHLH基因的结构域。根据bHLH家族成员在染色体的位置,将其依次编号为HubHLH1~HubHLH153用于后续分析。 -
使用 ExPASy蛋白质组服务器上的ProtParam tool(
https://web.expasy.org/protparam/ )工具[19],预测火龙果bHLH基因家族蛋白的基本理化性质,用 Plant-mPLoc(http://www.csbio.sjtu.edu.cn/bioinf/plant-multi/ )在线网站,进行亚细胞定位预测[20]。 -
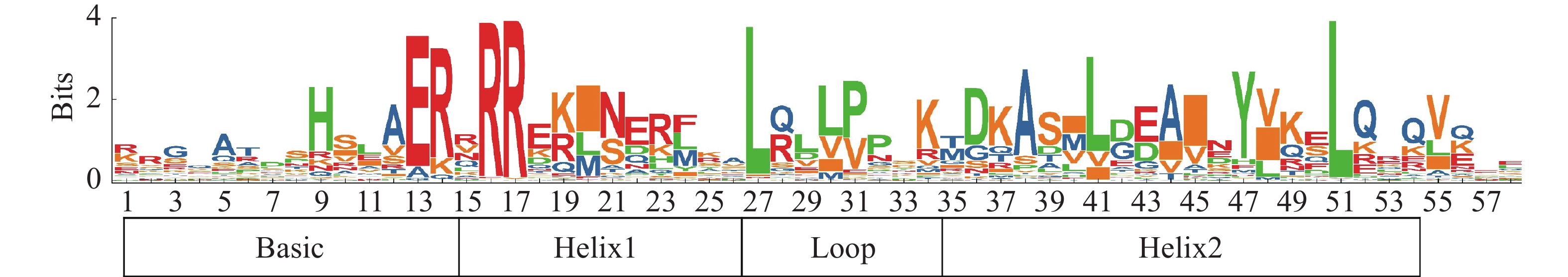
用MEGA 7.0自带的Clustal W工具进行153个HubHLHs的多序列比对,将比对结果上传到WEBLOGO在线程序(
http://weblogo.berkeley.edu )分析其序列(图1),展示出共同的序列标识[21]。同时,用TBTOOLS软件的Trimmer程序,将获得的比对结果修剪,将火龙果和拟南芥的bHLH蛋白序列共同构建进化树,选择最大似然法,校验参数Bootstrap重复1000次,其他使用默认参数。进化树美化用ITOL(http://itol.embl.de/ )网址绘制[22]。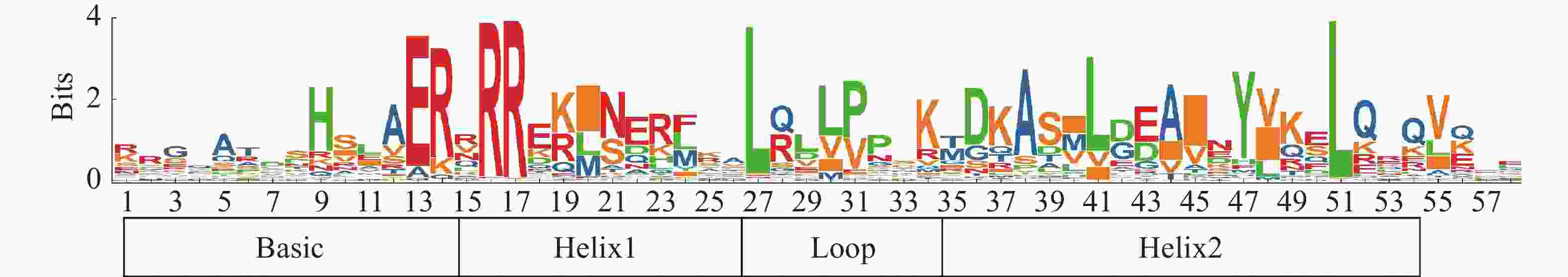
图 1 火龙果bHLH保守结构域序列LOGO
-
根据火龙果的基因组注释,提取HubHLH的基因结构,采用MEME(
http://meme-suite.org/tools/meme )程序,设置默认参数,预测HubHLH蛋白的保守motif。使用TBTOOLS的 Gene Structure view(Advanced)程序展示153个HubHLHs基因的进化树、保守motif以及基因结构的组合图[23]。 -
基于火龙果基因组,使用TBTOOLs 提取153个HubHLHs基因在染色体的物理位置,使用Gene Density Profile程序提取染色体的基因密度信息,在TBTOOLs 的Gene Location Visualize from GFF程序中一起作图展示。
-
采用多重共线性扫描工具包(MCScanX)分析基因重复事件,使用默认参数,利用TBTOOLs 的Fasta Stats功能得到火龙果染色体长度信息,利用 TBtools的Advanced Circos程序进行HubHLH基因家族共线性的可视化。
-
利用笔者所在的课题组前期研究的高质量火龙果转录组[24]进行分析,筛选出全部注释到 Nr、 Swissprot、KOG(evalue<0.00001)、KEGG蛋白质数据库的 bHLH 基因,并提取蛋白质序列,在SMART数据库中比对,进一步确定候选基因。将候选基因的蛋白质序列在基因组数据库进行本地BLAST比对,实现转录组和基因组中的ID转换。RNA-Seq 数据中使用 RPKM 法计算基因表达量,可直接用于比较不同样品间的基因表达差异。使用 TBTOOLs的 Heatmap 工具,展示bHLH基因在NL、L0、L1、L2(NL:火龙果未补光未开花,L0:补光了未开花,L1:补光后刚现蕾,L2:补光后现蕾一周)4个时期的表达变化,并设置筛选条件为 FDR<0.05 且 |log2FC|>1,展示差异基因的表达模式。利用eggNOG-mapper平台,得到火龙果基因功能注释信息,利用TBTOOLs整理结果,进行GO和KEGG富集分析,并以条状图进行可视化。
-
火龙果全基因组筛选得到153个HubHLH基因家族成员,占总基因的比率约为0.55%。对其蛋白质序列在ProtParam tool上分析,得到蛋白序列长度最小的HubHLH143有176个氨基酸,长度最大的HubHLH48有687个氨基酸,蛋白质的分子量范围为19.28~74.44 kDa,理论等电点范围为4.81(HubHLH68)至9.88(HubHLH143),平均值为6.86。所有的HubHLHs全部为亲水蛋白,总平均亲水性全部为负值。除了HubHLH143外,其他基因的不稳定系数均大于40,为不稳定蛋白。亚细胞定位预测显示,有140个基因定位到细胞核,7个位于细胞质中,6个位于叶绿体中,符合转录因子主要在细胞核中发挥作用的特点。
-
将获得的153个火龙果HubHLH家族成员进行同源序列比对后,在WEBLOGO网址以图形的形式展示bHLH保守结构域氨基酸的序列,可以得到所鉴定的HubHLH家族成员,具有明显保守的火龙果HubHLH结构域,且HubHLH的氨基酸数量约为60。
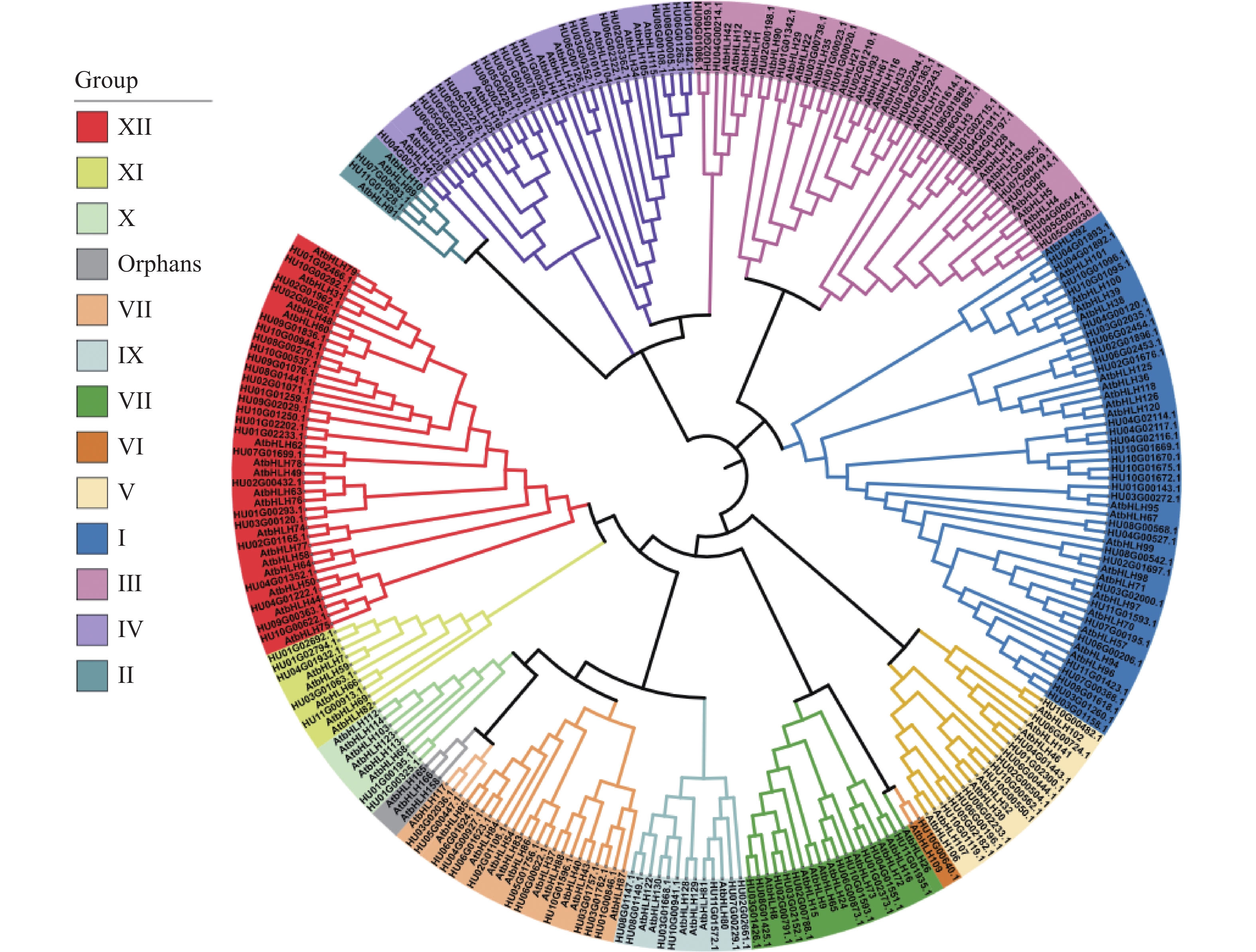
基于153个火龙果bHLH和120个拟南芥bHLH的蛋白序列共同构建有根系统发育树(图2),参考拟南芥中的bHLH蛋白亚家族分类结果,发现273个bHLH蛋白被分为25个亚家族,除了拟南芥中一个单独的组,其他拟南芥和火龙果中的bHLH共同聚集到12个组别(Ⅰ-Ⅻ),火龙果与拟南芥的蛋白分类几乎一致,说明火龙果HubHLH基因在长期进化的过程中,这些蛋白质没有遗失。
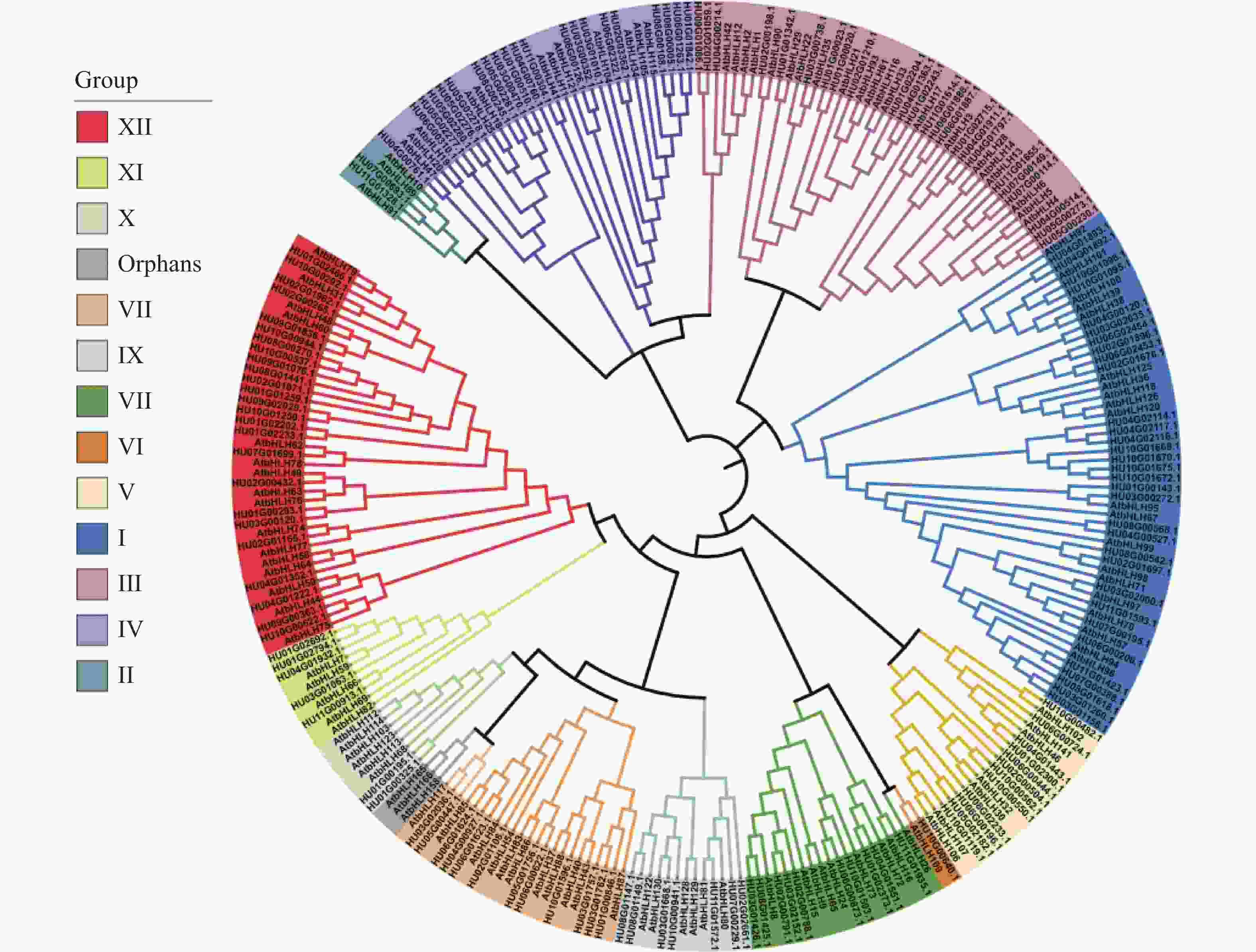
图 2 火龙果和拟南芥bHLH蛋白的系统发育树和亚科分类
-
基于153个HubHLH蛋白序列构建的矩形系统发育树(图3-A),系统发育树在外结点的自展值几乎都大于70,所以分支比较可靠,HubHLH基因被分成12组(Ⅰ-Ⅻ),包含25个亚家族。其中,GroupI中包含的火龙果bHLH基因家族成员数最多,有32个,占基因家族总数的20.92%,其次为Group Ⅻ有25个bHLH (16.34%),第三为Group Ⅲ,含有24个bHLH (15.69%),Group Ⅵ含有的bHLH成员最少,仅有1个(0.66%)。其他组别Group Ⅱ和Group Ⅹ均只含有2个,Group Ⅳ有21个,Group V含有12个,Group Ⅶ含有10个,Group Ⅷ有12个,Group Ⅸ有7个,Group Ⅺ有5个。这与拟南芥中Group Ⅵ、Group Ⅱ含有bHLH基因较少,Group Ⅰ、Group Ⅲ、Group Ⅻ含有bHLH较多的结果一致[13]。
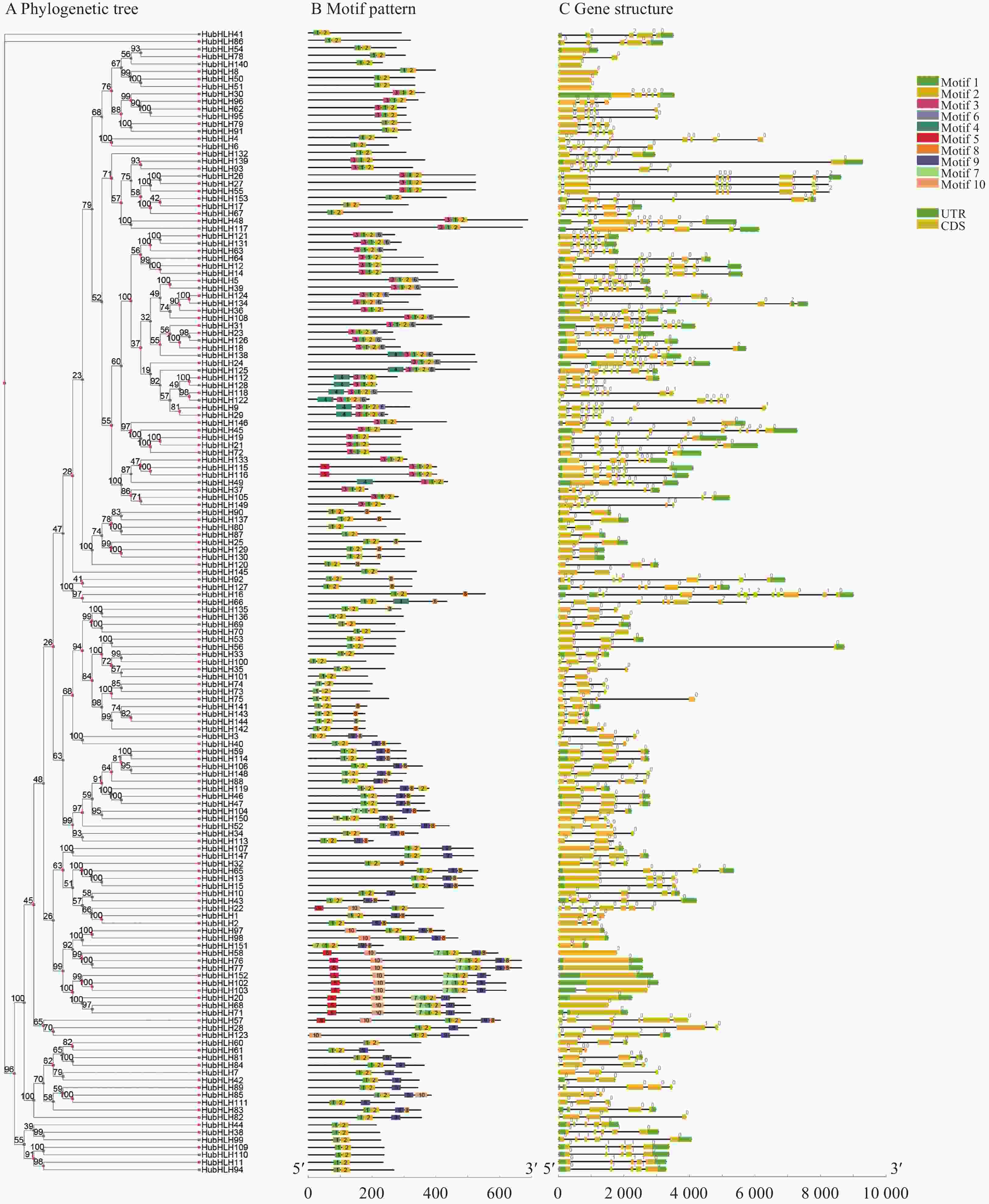
图 3 HubHLHs的motif分布和基因结构图
使用TBtools展示MEME在线软件预测的153个HubHLH蛋白的保守motif(图3-B),结果表明,所有的HubHLH家族成员都包含保守Motif 1:AERRRREKJNERFKALRSLVP和Motif 2:NVKKTDKASILDEAINYVKELQRQVQELS;但不同HubHLH成员所包含的保守motif数目及种类是存在差异的,含有motif3~motif10的基因数量依次为48、10、15、18、10、45、49、15。本研究发现,同一组中的HubHLH成员具有相似的motif组成,有些motif仅存在于特定的亚家族中,例如,motif6只存在Ⅻ成员中,motif5除了HubHLH115和HubHLH116外,剩余全部在第Ⅲ组。通过比较基因组DNA序列,并根据系统发育分类的顺序,显示所有HubHLH的内含子和外显子结构,通过比较其数量和位置,发现153个HubHLH 基因具有不同数量的外显子,从1到11不等(图3-C)。同一亚家族的基因外显子数量及位置相近,同一分类的基因结构也极为相似,例如VIIIb成员都只包含1个外显子,IVa含有3或4个外显子,IVc中都有5个外显子和4个内含子,这与某些亚家族具有特定的功能有关。
-
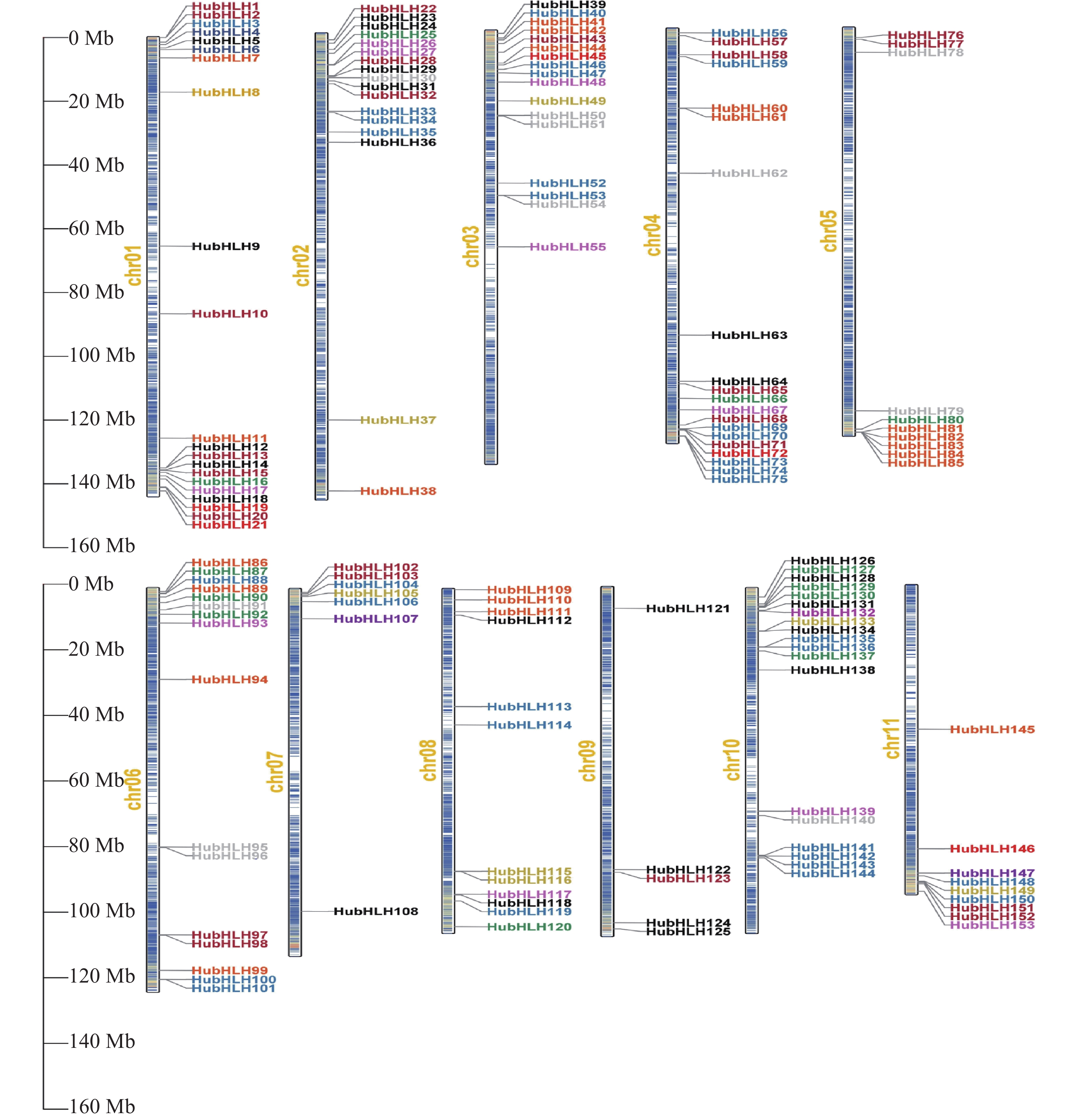
基于火龙果基因组数据库,创建HubHLH基因在染色体的分布示意图(图4),分布映射结果表明,大多数HubHLH基因分布在染色体的末端,染色体两端的基因密度较大。此外,153个HubHLH基因在染色体1至11上的分布不均匀,Chr1含有最多的HubHLH基因(21个,约13.73%),其次是Chr4(20个,约13.07%),然后是Chr10(19个,12.42%),而Chr9含有最少的基因(5个,约3.27%),Chr2和Chr3都含有17个(约11.11%),Chr6、Chr8、Chr5、Chr11、Chr7从多到少依次包含16个(10.46%)、12个(7.84%)、10个(6.54%)、9个(5.89%)、7个(4.58%)HubHLH基因。以不同的颜色展示HubHLH所属的12个组别,显然,除了少数的基因簇,每个组别在染色体的位置没有特殊的指向,且不同HubHLH亚家族的一些成员位于同一小染色体区域内,而同一亚家族的某些成员分布在不同的染色体区域,表明HubHLH基因广泛分布于火龙果基因组中。
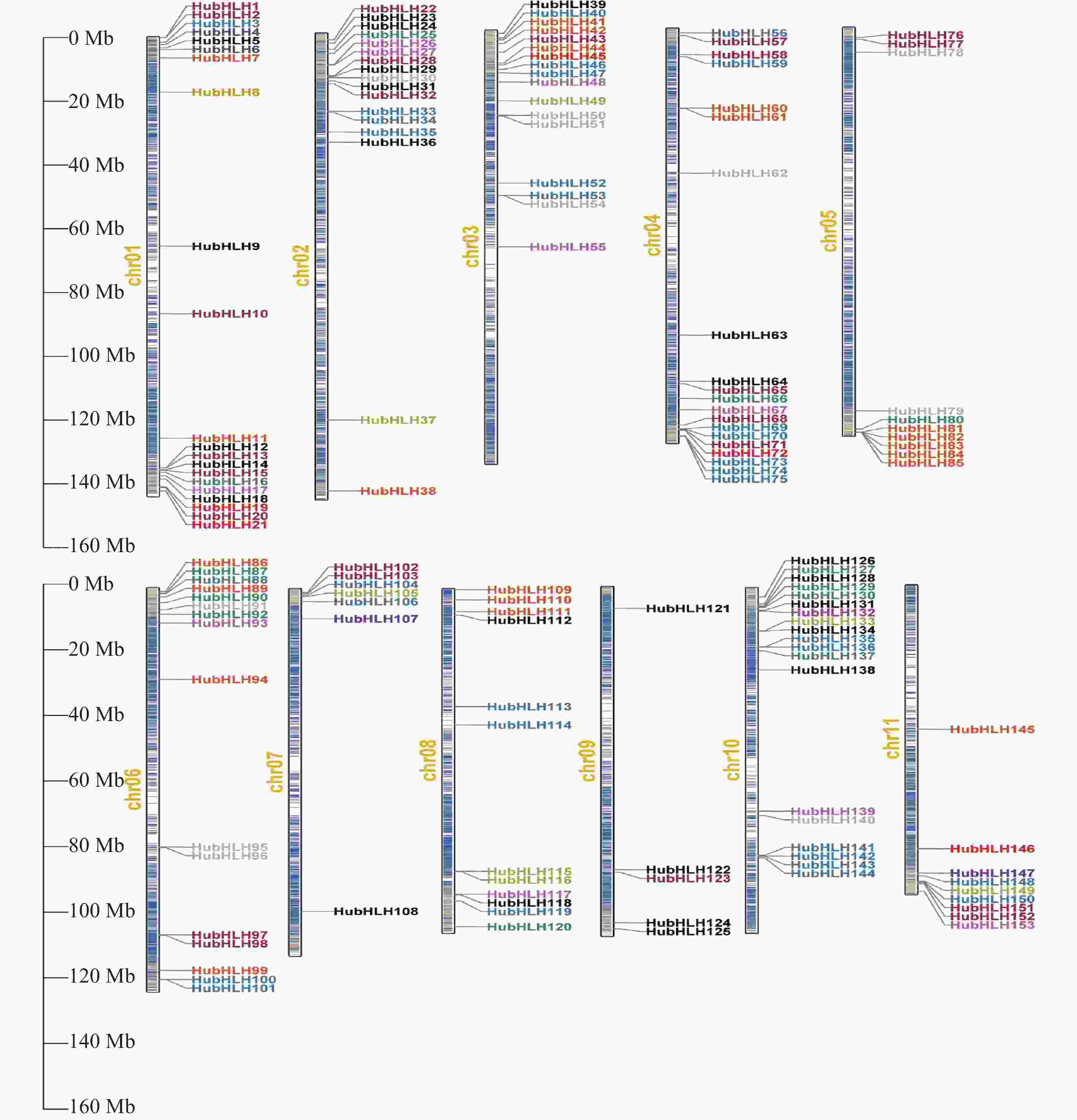
图 4 HubHLHs基因的染色体定位
-
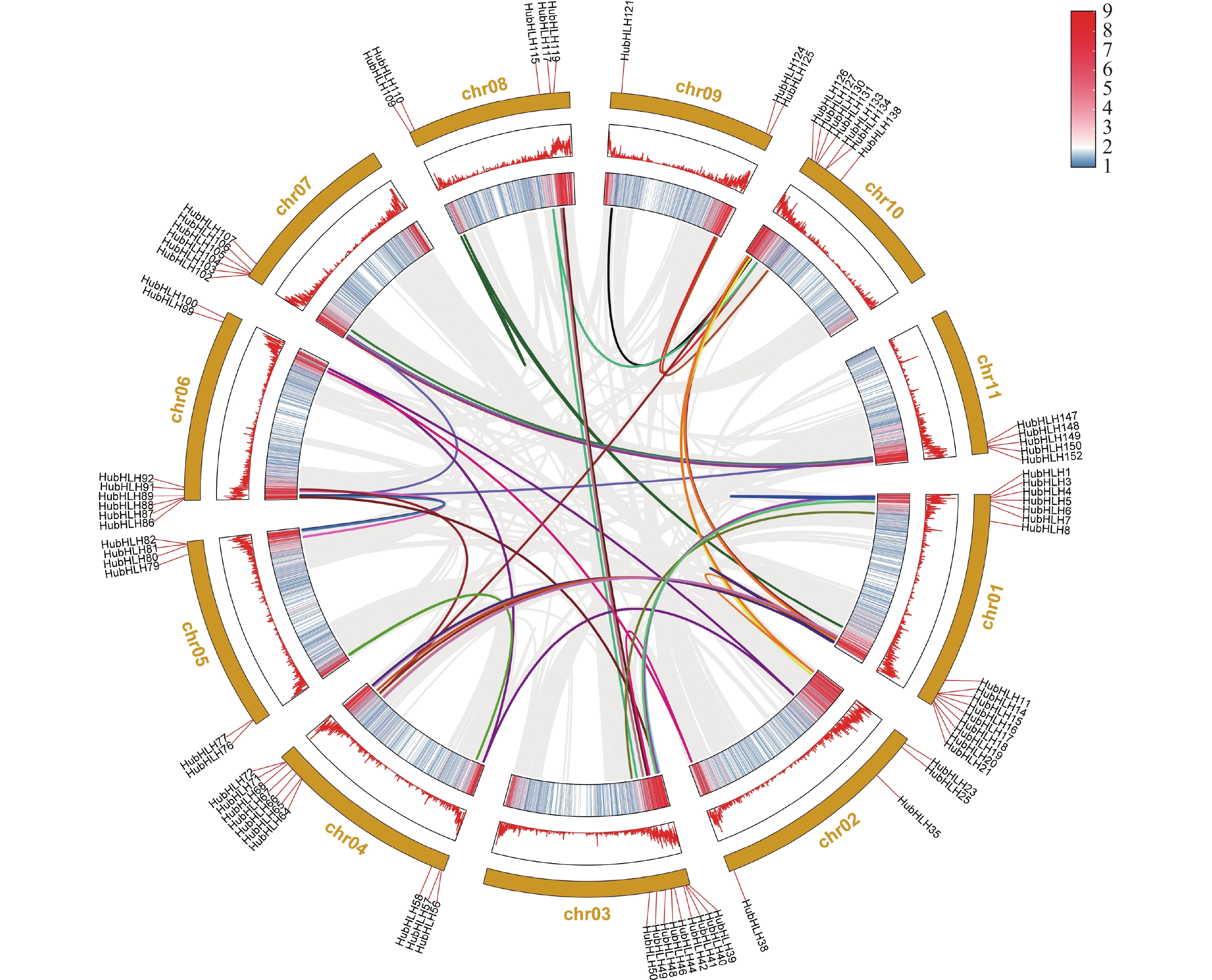
通过MCScanX分析火龙果的基因组,共线性结果(图5)显示,在11条染色体上,火龙果bHLH基因家族,在全基因组水平存在大量的片段重复(segmental duplication, SD)基因对,这将导致火龙果bHLH基因家族数量上的扩增,图5中共有78个HubHLH基因聚集在49个片段复制事件。由于基因家族和基因组的扩展是植物进化过程的一部分,说明片段重复事件是驱动HubHLH基因家族进化的主要力量,这也同样促进了火龙果表型的多样化和新功能的产生。本研究还发现了7对串联重复基因对(HubHLH69/HubHLH70,HubHLH74/75,HubHLH84/85,HubHLH95/96,HubHLH97/98,HubHLH135/HubHLH136,HubHLH141/HubHLH142),可能通过串联复制(tandem duplication, TD)产生,每对都聚集在同一个亚家族中,产生这种现象的原因可能有基因加倍、染色体片段的交换等。
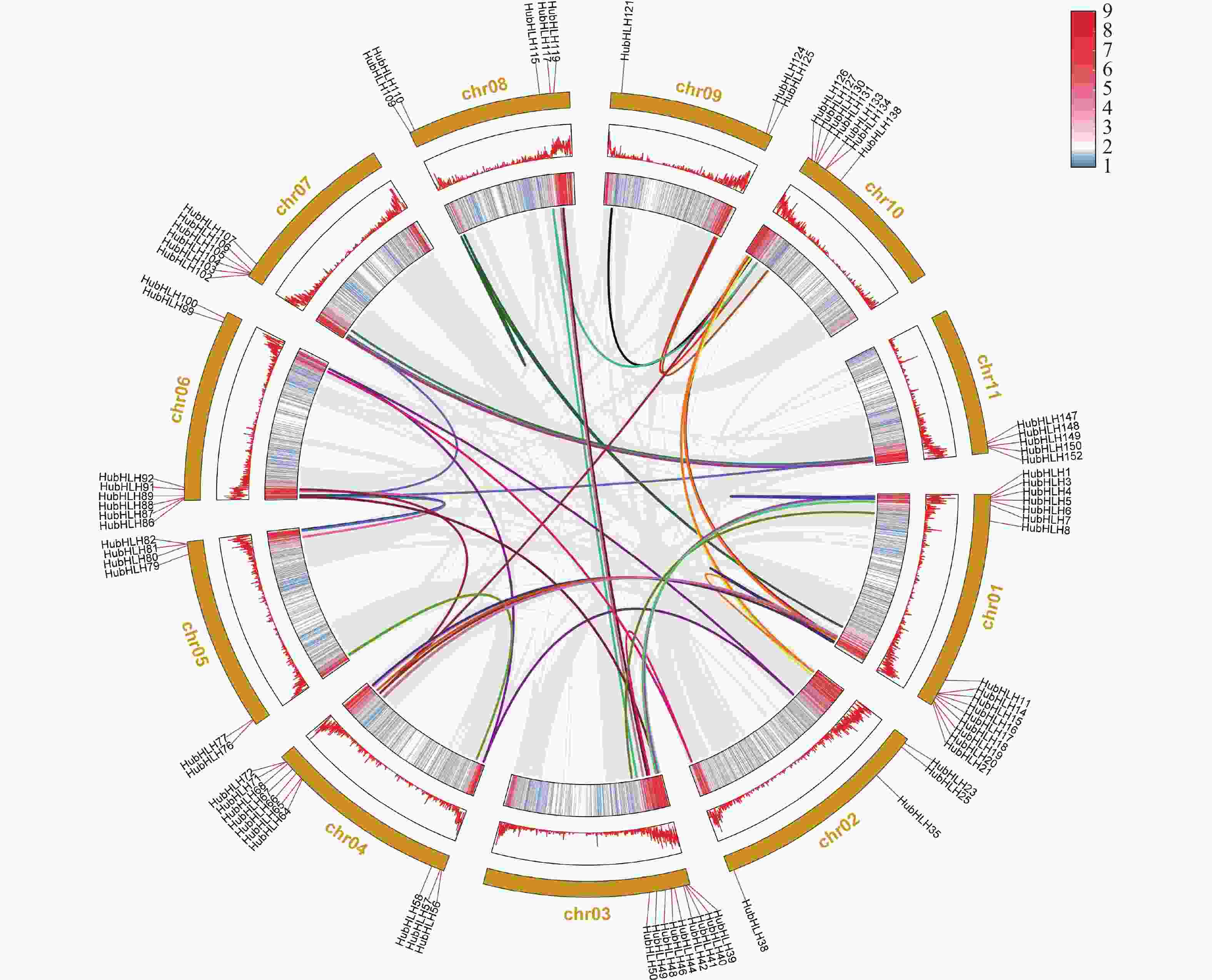
图 5 火龙果HubHLHs基因家族的基因复制和同步分析
-
对火龙果补光诱导开花的转录组数据进行分析,共注释到142个bHLH基因,与基因组数据库比对得到87个HubHLH基因,依据FDR<0.05 且 |log2FC|>1,挑选差异表达基因,筛选到59个差异表达基因,以热图展示差异表达情况(图6)。重点关注补光诱导前后bHLH基因的变化,在L0至L1时期,差异表达HubHLH基因有6个显著下调,34个显著上调;在NL和L1时期,有8个差异表达HubHLH基因下调,33个上调;在L1至L2花发育过程,HubHLH差异基因有15个下调,7个上调。总之,HubHLH基因在补光前后呈现较为明显的上下调趋势,这些基因在冬季补光诱导开花中可能发挥了作用。

图 6 转录组中HubHLH基因的表达情况
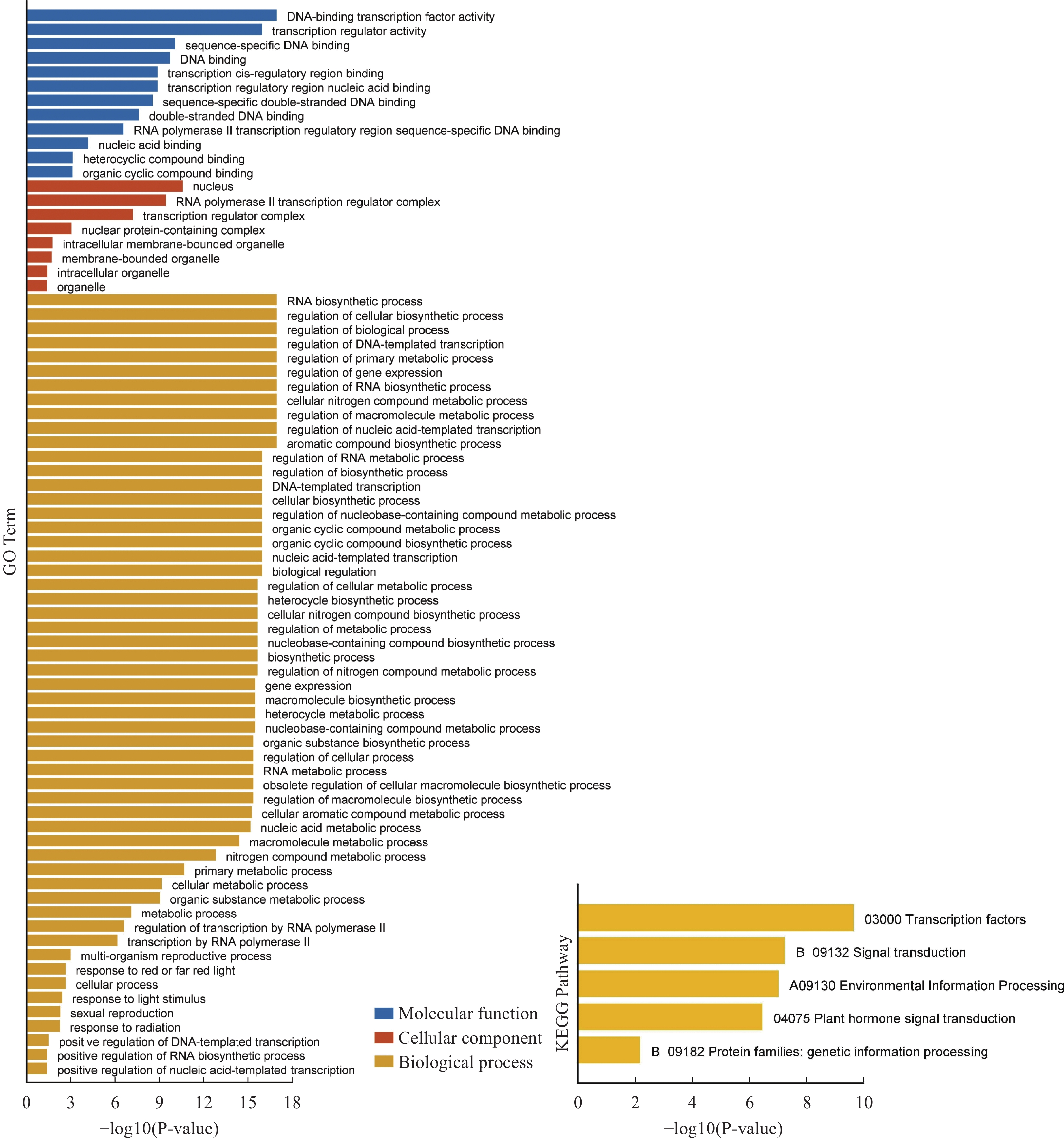
为了解HubHLH差异表达基因的偏好功能,对它们进行基于超几何分布的GO和KEGG分析(图7)。重点关注生物进程,发现HubHLH差异基因,在红光或远红光的反应、对光刺激的反应、有性生殖功能、对辐射的反应等功能上均有富集。在KEGG分析中,主要富集在转录因子,信号转导,环境信息处理,植物激素信号转导功能通路上。
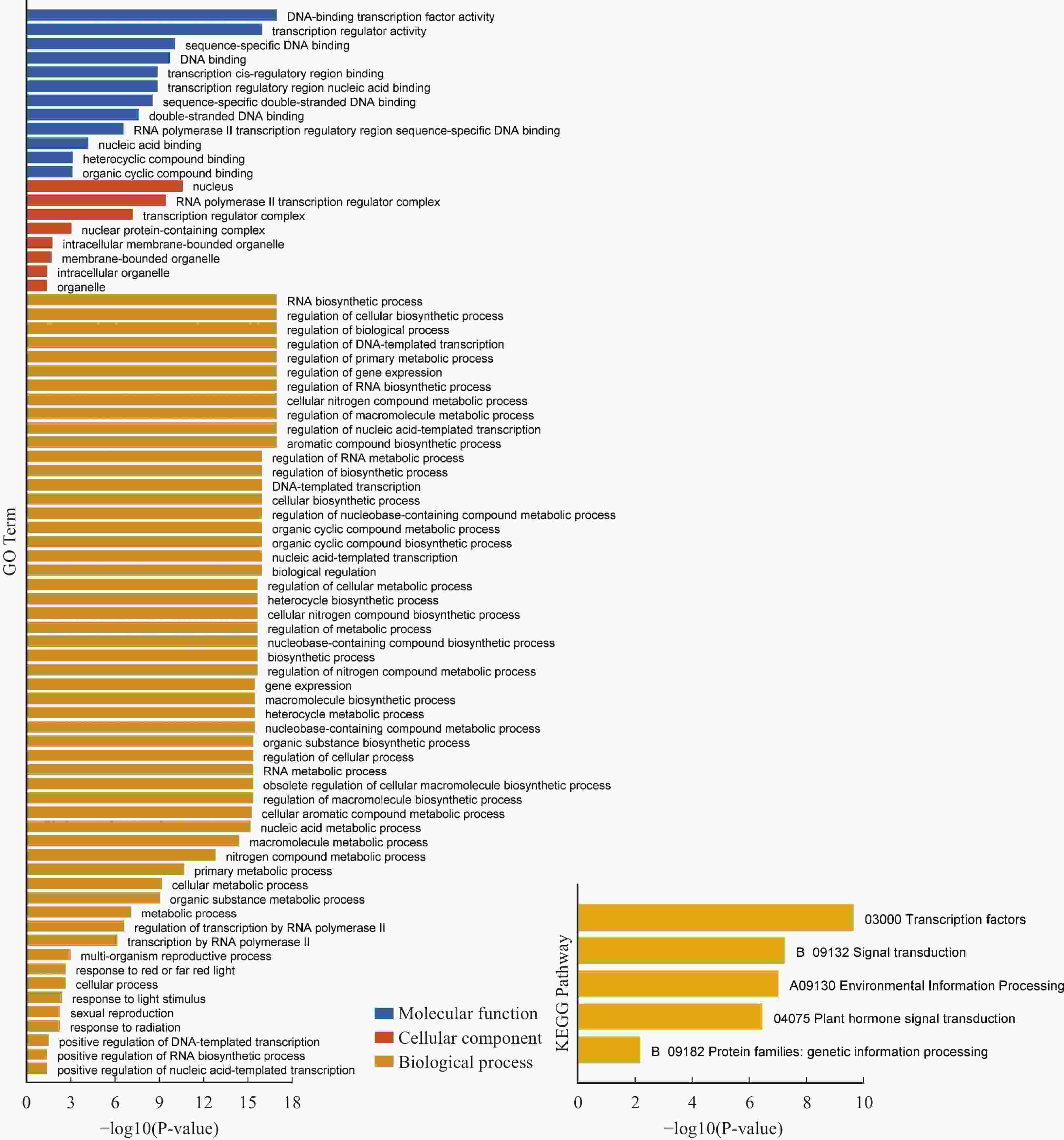
图 7 差异HubHLH基因的GO和KEGG分析条状图
-
bHLH作为植物中第二大类转录因子,近几年在菠萝(Ananas comosus L.)[25]、香蕉(Musa acuminata L.)[26]、木薯(Manihot esculenta crantz)[27]等热带植物中实现了全基因组鉴定,分别鉴定出121、259、148个bHLH基因。火龙果全基因组序列的发表,为进一步检索、克隆和验证火龙果的bHLH基因提供了条件。同时,冬季补光诱导开花过程是严密而复杂的过程,bHLH基因被证实参与了植物生长发育的很多过程。因此对火龙果进行bHLH基因家族的全基因组筛选和分析,为后续它们对冬季补光诱导开花的响应过程中的功能研究很有必要。 本研究首次对火龙果bHLH基因家族进行了全基因组鉴定,得到153个火龙果HubHLH基因,对它们的理化性质全面预测,得到火龙果HubHLH的分子量、氨基酸长度、等电点等理化性质,与其他植物中的bHLH差异不大,亚细胞定位预测主要分布在细胞核。
目前,植物中bHLH基因还未有特别明确的分类标准,通常选择与模式植物或亲缘关系相近的植物,共同构建有根系统发育树进行分类,本研究选择长日照植物的拟南芥,将它的120个bHLH蛋白序列和鉴定到的153个火龙果HubHLH蛋白序列共同构建有根系统发育树,通过与拟南芥的系统发育关系比较,火龙果bHLH分类结果和拟南芥几乎一致,都得到了12个主要的组别(Ⅰ-Ⅻ)[28]。进行火龙果HubHLH基因家族的保守motif和基因结构分析,153个HubHLH基因都含有motif 1和motif 2,同一分组的基因结构相似,证明了系统发育分析结果的可靠性。motif位点的识别有助于鉴定其家族同源性和预测蛋白质的功能,而系统发育树显示,一些火龙果中的HubHLH与拟南芥的bHLH基因可能同源,并具有相似功能,因此可以通过发现聚集到同一亚家族的拟南芥中功能较为明确的bHLH蛋白,发现火龙果中bHLH的可能功能。例如在亚家族IIIe中的成员与茉莉酸介导的开花相关,亚家族Ⅶ的成员多与光信号有关[29],成员较多的Ⅻ亚家族中有些调节光周期开花[30],有些与光和植物激素之间的相互作用有关[31]。
-
基于转录组数据展示HubHLH基因家族成员在火龙果冬季夜晚补光诱导成花过程中NL、L0、L1、L2四个时期的差异表达情况,其中有注释到JA途径中调节开花时间的MYC2基因,在补光后表达含量急速增加;还有响应光和温度,在花形态发生过程中起作用的光稳定SPATULA基因,也在L0-L1阶段表达出上调趋势,其他HubHLH基因对补光诱导也有不同程度的响应。另外,根据HubHLH差异基因的GO富集结果,发现在有性生殖,即火龙果成花过程,对光反应和光刺激,即补光过程,都得到了功能富集。HubHLH基因家族可能在冬季补光诱导火龙果成花过程起到了作用。总之,开花时间的调控是严格而复杂的,在冬季通过补光诱导火龙果开花具有重要的生产实践价值,本研究利用火龙果基因组信息和转录组信息,对火龙果HubHLH基因家族进行了全面分析研究,为火龙果冬季补光诱导开花,提供了许多潜在的候选基因。
火龙果HubHLH基因家族的全基因组分析及其对冬季补光诱导开花的表达响应
DOI: 10.15886/j.cnki.rdswxb.20220108
 CSTR: 32425.14.j.cnki.rdswxb.20220108
CSTR: 32425.14.j.cnki.rdswxb.20220108
Whole-genome analysis of the HubHLH gene family in Pitaya and its differential expression in response to supplementary light-induced flowering in winter
-
摘要: 为了获得较完整的候选基因,探讨HubHLH基因在火龙果( Hylocereus undatus )冬季补光诱导开花过程的表达响应,对火龙果HubHLH基因家族进行全基因组分析。鉴定出153个HubHLH基因;这些基因的编码蛋白含有176~687个氨基酸,分子量大小为19.28~74.44 kDa ,等电点(pI)为4.81~9.88,均为亲水蛋白,亚细胞定位预测大多定位到细胞核。为鉴定HubHLH基因家族的同源性,本研究将153个火龙果HubHLH和120个拟南芥AtbHLH蛋白进行系统发育比较分析。系统发育比较分析结果:火龙果HubHLH基因家族成员被分为12个组,25个亚族;对HubHLH基因家族的保守motif、基因结构及在染色体的位置分布的分析结果表明,同一亚族的基因具有相似的基序组成和基因结构。对HubHLH基因家族的内部复制事件的分析结果表明,有78条片段复制被鉴定为片段重复基因,说明片段复制是HubHLH基因家族的主要扩张力量。此外,基于已测定的关于火龙果冬季补光诱导开花的4个时期转录组数据,筛选到59个HubHLH基因在冬季补光诱导成花过程中有差异表达,随后对这59个HubHLH基因进行GO功能富集,发现它们在红光或远红光的反应、对光刺激的反应、有性生殖功能、对辐射的反应等功能上均有富集,说明HubHLH基因家族可能在冬季补光诱导火龙果成花过程中起到了调控作用。Abstract: In order to obtain relative complete candidate genes, the HubHLH gene expression response in Pitaya during the flowering process induced by supplementary light in winter was investigated, and the whole genome of HubHLH gene family in Pitaya was analyzed. There were 153 pitaya HubHLH genes identified, and their encoding proteins contained 176-687 amino acids with their molecular weight being 19.28 – 74.44 kDa, pI 4.81-9.88. These proteins were all hydrophilic. The prediction of subcellular localization showed that most of the proteins were localized in the nucleus. The pitaya HubHLH proteins were compared with 120 Arabidopsis AtbHLH proteins for phylogenetic analysis. Phylogenetic analysis showed that the pitaya HubHLH proteins were divided into 12 groups and 25 subfamilies. Analysis of conserved motif, gene structure and location distribution in chromosomes showed that genes of the same subfamily had similar motif composition and gene structure. Analysis of the internal replication events of the pitaya HubHLH gene family found that 78 genes were identified as fragment repeating genes, indicating that fragment replication was the main expansion force of the pitaya HubHLH gene family. In addition, based on the transcriptomic data of the four phases of supplementary light-induced flowering of Pitaya in winter, 59 HubHLH genes were differentially expressed during the process of supplementary light induced flowering in winter. GO function enrichment showed that they were enriched in response to red or far-red light, light stimulation, sexual reproduction function, and radiation. The HubHLH gene family may play a regulatory role in the flowering process of Pitaya induced by supplementary light in winter. In this study, we conducted a comprehensive genome analysis of the HubHLH gene family in Pitaya, obtained relatively complete candidate genes, and preliminarily investigated the expression response of HubHLH genes in the winter flowering process induced by supplementary light in Pitaya.
-
图 3 HubHLHs的motif分布和基因结构图
图3-A是HubHLH蛋白序列构建的矩形系统发育树,每个节点上的数值代表1000个重复的自展值;图3-B的彩色矩形框代表HubHLH蛋白的保守motif;图3-C中黄色矩形和黑色线条分别表示外显子(CDS,coding sequence)和内含子(introns),绿色的矩形展示了非翻译区域(UTR,untranslated region),横坐标的黑线表示相对蛋白质长度。
图 4 HubHLHs基因的染色体定位
根据其在火龙果Chr1至Chr11上,从上到下的物理位置,依次命名为HubHLH1~HubHLH153,图4中左边的刻度尺表示染色体长度,单位为Mb。右侧细的垂直条代表火龙果的染色体,染色体编号显示在每条染色体的左侧,内部颜色代表染色体上的基因密度,颜色越白,表示该位置的基因密度越小,颜色越红,表示该位置的基因密度越大。
图 5 火龙果HubHLHs基因家族的基因复制和同步分析
HubHLH基因的染色体位置由圆圈上的短红线连接,背景的灰色线表示火龙果基因组中的所有共线性区块,彩色线表示片段重复的HubHLH基因对,相同片段重复基因对的两个基因以相同的颜色标记。并将染色体的基因密度分别以热图和线图的形式在外圈展示。
-
[1] 刘成立, 王猛, 郭攀阳, 等. 火龙果花和果实的动态发育规律研究[J]. 海南大学学报(自然科学版), 2020, 38(2): 147 − 152. doi: 10.15886/j.cnki.hdxbzkb.2020.0021 [2] HAO Y, ZONG X, REN P, et al. Basic helix-loop-helix (bHLH) transcription factors regulate a wide range of functions in Arabidopsis[J]. Int J Mol Sci, 2021, 22(13): 7152. doi: 10.3390/ijms22137152 [3] ITO S, SONG Y H, JOSEPHSON-DAY A R, et al. FLOWERING BHLH transcriptional activators control expression of the photoperiodic flowering regulator CONSTANS in Arabidopsis[J]. Proc Natl Acad Sci U S A, 2012, 109(9): 3582 − 3587. doi: 10.1073/pnas.1118876109 [4] LIU H, YU X, LI K, et al. Photoexcited CRY2 interacts with CIB1 to regulate transcription and floral initiation in Arabidopsis[J]. Science, 2008, 322(5907): 1535 − 1539. doi: 10.1126/science.1163927 [5] KUMAR S V, LUCYSHYN D, JAEGER K E, et al. Transcription factor PIF4 controls the thermosensory activation of flowering[J]. Nature, 2012, 484(7393): 242 − 245. doi: 10.1038/nature10928 [6] LI Y, WANG H, LI X, et al. Two DELLA-interacting proteins bHLH48 and bHLH60 regulate flowering under long-day conditions in Arabidopsis thaliana[J]. J Exp Bot, 2017, 68(11): 2757 − 2767. doi: 10.1093/jxb/erx143 [7] CHEN W, ZHAO L, LIU L, et al. Iron deficiency-induced transcription factors bHLH38/100/101 negatively modulate flowering time in Arabidopsis thaliana[J]. Plant Sci, 2021, 308: 110929. doi: 10.1016/j.plantsci.2021.110929 [8] WASEEM M, LI N, SU D, et al. Overexpression of a basic helix-loop-helix transcription factor gene, SlbHLH22, promotes early flowering and accelerates fruit ripening in tomato (Solanum lycopersicum L.)[J]. Planta, 2019, 250(1): 173 − 185. doi: 10.1007/s00425-019-03157-8 [9] WANG H, LI Y, PAN J, et al. The bHLH Transcription Factors MYC2, MYC3, and MYC4 are required for jasmonate-mediated inhibition of flowering in Arabidopsis[J]. Mol Plant, 2017, 10(11): 1461 − 1464. doi: 10.1016/j.molp.2017.08.007 [10] WANG F, GAO Y, LIU Y, et al. BES1-regulated BEE1 controls photoperiodic flowering downstream of blue light signaling pathway in Arabidopsis[J]. New Phytol, 2019, 223(3): 1407 − 1419. doi: 10.1111/nph.15866 [11] LIU Y, LI X, MA D, et al. CIB1 and CO interact to mediate CRY2-dependent regulation of flowering[J]. EMBO Rep, 2018, 19(10): e45762. doi: 10.15252/embr.201845762 [12] REYMOND M C, BRUNOUD G, CHAUVET A, et al. A light-regulated genetic module was recruited to carpel development in Arabidopsis following a structural change to SPATULA[J]. Plant Cell, 2012, 24(7): 2812 − 2825. doi: 10.1105/tpc.112.097915 [13] HEIM M A, JAKOBY M, WERBER M, et al. The basic helix-loop-helix transcription factor family in plants: a genome-wide study of protein structure and functional diversity[J]. Mol Biol Evol, 2003, 20(5): 735 − 747. doi: 10.1093/molbev/msg088 [14] GUO X J, WANG J R. Global identification, structural analysis and expression characterization of bHLH transcription factors in wheat[J]. BMC Plant Biol, 2017, 17(1): 90. doi: 10.1186/s12870-017-1038-y [15] FAN Y, LAI D, YANG H, et al. Genome-wide identification and expression analysis of the bHLH transcription factor family and its response to abiotic stress in foxtail millet (Setaria italica L.)[J]. BMC Genomics, 2021, 22(1): 778. doi: 10.1186/s12864-021-08095-y [16] BANO N, PATEL P, CHAKRABARTY D, et al. Genome-wide identification, phylogeny, and expression analysis of the bHLH gene family in tobacco (Nicotiana tabacum)[J]. Physiol Mol Biol Plants, 2021, 27(8): 1747 − 1764. doi: 10.1007/s12298-021-01042-x [17] ZHANG X Y, QIU J Y, HUI Q L, et al. Systematic analysis of the basic/helix-loop-helix (bHLH) transcription factor family in pummelo (Citrus grandis) and identification of the key members involved in the response to iron deficiency[J]. BMC Genomics, 2020, 21(1): 233. doi: 10.1186/s12864-020-6644-7 [18] CHEN J Y, XIE F F, CUI Y Z, et al. A chromosome-scale genome sequence of pitaya (Hylocereus undatus) provides novel insights into the genome evolution and regulation of betalain biosynthesis[J]. Horticulture Research, 2021, 8: 164. doi: 10.1038/s41438-021-00612-0 [19] DUVAUD S, GABELLA C, LISACEK F, et al. Expasy, the Swiss Bioinformatics Resource Portal, as designed by its users[J]. Nucleic Acids Research, 2021, 49(W1): W216 − W227. doi: 10.1093/nar/gkab225 [20] CHOU K C, SHEN H B. Plant-mPLoc: a top-down strategy to augment the power for predicting plant protein subcellular localization[J]. PLoS One, 2010, 5(6): e11335. doi: 10.1371/journal.pone.0011335 [21] CROOKS G E, HON G, CHANDONIA J M, et al. Weblogo: a sequence logo generator[J]. Genome Research, 2004, 14(6): 1188 − 1190. doi: 10.1101/gr.849004 [22] LETUNIC I, BORK P. Interactive Tree Of Life (iTOL): an online tool for phylogenetic tree display and annotation[J]. Bioinformatics, 2007, 23(1): 127 − 128. doi: 10.1093/nar/gkab301 [23] CHEN C, CHEN H, ZHANG Y, et al. TBtools: An integrative toolkit developed for interactive analyses of big biological data[J]. Molecular Plant, 2020, 13(8): 1194 − 1202. doi: 10.1016/j.molp.2020.06.009 [24] XIONG R, LIU C, XU M, et al. Transcriptomic analysis of flower induction for long-day pitaya by supplementary lighting in short-day winter season[J]. BMC Genomics, 2020, 21(1): 329. doi: 10.1186/s12864-020-6726-6 [25] MOHAMMAD A, HASSAN J B, BEENISH F, et al. Genome-wide study of pineapple (Ananas comosus L.) bHLH transcription factors indicates that cryptochrome-interacting bHLH2 (AcCIB2) participates in flowering time regulation and abiotic stress response[J]. BMC Genomics, 2020, 21(1): 735. doi: 10.1186/s12864-020-07152-2 [26] WANG Z, JIA C, WANG J, et al. Genome-wide analysis of basic Helix-loop-Helix transcription factors to elucidate candidate genes related to fruit ripening and stress in banana (Musa acuminata L. AAA group, cv. Cavendish)[J]. Front Plant Sci, 2020, 11: 650. doi: 10.3389/fpls.2020.00650 [27] AN F, XIAO X, CHEN T, et al. Systematic analysis of bHLH transcription factors in cassava uncovers their roles in postharvest physiological deterioration and cyanogenic glycosides biosynthesis[J]. Frontiers in Plant Science, 2022, 13: 901128. doi: 10.3389/fpls.2022.901128 [28] TOLEDO-ORTIZ G, HUQ E, QUAIL P H. The Arabidopsis basic/Helix-loop-Helix transcription factor family[J]. The Plant Cell, 2003, 15(8): 1749 − 1770. doi: 10.1105/tpc.013839 [29] HAO Y, OH E, CHOI G, et al. Interactions between HLH and bHLH factors modulate light-regulated plant development[J]. Molecular Plant, 2012, 5(3): 688 − 697. doi: 10.1093/mp/sss011 [30] SHARMA N, XIN R, KIM D H, et al. NO FLOWERING IN SHORT DAY (NFL) is a bHLH transcription factor that promotes flowering specifically under short-day in Arabidopsis[J]. Development, 2016, 143(4): 682 − 690. [31] WU M, UPRETI S, YAN A, et al. SPATULA regulates floral transition and photomorphogenesis in a PHYTOCHROME B-dependent manner in Arabidopsis[J]. Biochemical and Biophysical Research Communications, 2018, 503(4): 2380 − 2385. doi: 10.1016/j.bbrc.2018.06.165 -

 点击查看大图
点击查看大图
计量
- 文章访问数: 2540
- HTML全文浏览量: 747
- PDF下载量: 55
- 被引次数: 0
