


-
香蕉(Musa spp.)是中国重要的热带水果之一,香蕉枯萎病是影响香蕉产业最严重的病害之一。尖孢镰刀菌古巴专化型4号小种(Fusarium oxysporum f. sp. cubense tropical race 4,Foc TR4)是引起该病害的主要病原菌[1],对香蕉产业造成毁灭性危害且难以控制。Foc TR4的分生孢子具有极强的抗逆性,在土壤环境中存活数十年[2],可以通过机械耕作、灌溉、植株迁移等方式迅速传播[3]。Foc TR4侵染香蕉植株的维管束,并渗透到维管束组织内,选择性逃避与外界防控手段的接触,导致传统的防治方法难以奏效[4]。香蕉新型抗病品种的选育被认为是克服香蕉枯萎病的有效方法,但中国并非香蕉原产地,且香蕉主要栽培品种为三倍体,故面临野生抗性资源少且杂交育种困难的问题[5]。目前,市面上尚未有防控香蕉枯萎病的高效农药,而传统农药的大量应用不仅效果甚微,而且造成了土壤环境的污染,加重经济、食品安全和生态环境的担忧[6]。香蕉与甘蔗、马铃薯等作物轮作改变了土壤耕作层的微生物群落结构,对香蕉枯萎病起到了一定的控制作用,但长期轮作势必会造成香蕉减产,制约香蕉产业的发展[7]。生物防治因具有高效、安全、易降解、环境友好等优点,是当前公认的植物病害防治的主要研究方向[8]。而筛选优良的生防菌并研究其抗香蕉枯萎病菌的作用机制,是目前香蕉枯萎病防治研究的热点[9]。近年来报道了从各种生态环境分离筛选出对Foc TR4具有显著抑制活性的生防菌,包括木霉菌[10]、芽孢杆菌[11]、链霉菌[12]、假单胞菌[13]、伯克霍尔德菌[14]等,在实验条件下对香蕉枯萎病的防治都呈现出积极的效果。但有关Foc TR4生防菌产生的活性成分的研究却鲜有报道[15]。本研究以一株具有抗香蕉枯萎病菌活性的海洋链霉菌(编号HNM0301)为研究对象,通过形态特征、16S rRNA基因序列聚类、全基因组序列比较分析鉴定其分类地位,通过基因组挖掘预测其潜在的次级代谢合成基因簇,通过天然产物分离及波谱解析鉴定其活性成分结构,研究结果可为香蕉枯萎病的生物防控提供先导化合物。
-
菌株HNM0301分离自海南东寨港红树林自然保护区红树植物海桑根际土壤,深度5~10 cm,经纬度为110°60'12.73''E 20°0'84.71''N,保存于实验室−80 ℃冰箱;火龙果溃疡病菌(Neoscytaldium dimidiatum)、菠萝黑腐病菌(Ceratocystis paradoxa)、火龙果黑腐病菌(Bipolaris cactivora)、番茄早疫病菌(Alternaria solani)、香蕉枯萎病菌(Fusarium oxysporum f. sp. Cubense tropical race 4)、豇豆枯萎病菌(Fusarium oxysporum Schl. F. sp. Tracheiphlium)、从赤壳菌(Nectria citri)、小麦赤霉病菌(Fusarium graminearum)等植物病原真菌由中国热带农业科学院热带生物研究所的海南省黎药资源天然产物研究与利用重点实验室提供。
-
FEM5培养基:黄豆粉 15.0 g,甘油 20.0 g,碳酸钙 0.4 g,碳酸镁 0.4 g,氯化钠 30.0 g,蒸馏水 1 000 mL,pH7.0;ISP2固体培养基:葡萄糖 4 g,酵母膏 4 g,麦芽浸粉 10.0 g,琼脂 15.0 g,蒸馏水1 000 mL,pH7.0;PDA培养基:土豆 200.0 g,葡萄糖 20.0 g,琼脂 15.0 g,蒸馏水 1 000 mL,pH 7.0。
-
使用平板对峙培养法进行菌株HNM0301的抗病原真菌活性评价。将菌株HNM0301在ISP2固体培养基上活化,28 ℃培养5 d,用打孔器打孔取直径6 mm放线菌菌饼,贴在PDA固体培养基中心的上下左右4个方向各2.5 cm处培养5 d。用打孔器打孔取直径6 mm病原真菌菌饼,贴在平板中心点。只接种病原菌的空白PDA固体培养基作为对照,每个处理重复3次,培养5 d后,测定病原真菌菌落的生长直径。抑制率计算公式:
$$ \begin{split} \text{抑菌率(\%)}=&\text{(对照组菌落直径−处理组菌落直径)/}\\ &\text{对照组菌落直径}\times 100。 \end{split} $$ -
将菌株HNM0301接种在ISP2培养基上,28 ℃培养7 d,用解剖刀切取菌落边缘的菌丝(5 mm
$ \times $ 5 mm),尽量切去琼脂。将切好的菌丝置于2.5%的戊二醛中浸泡4 h固定,再用0.1 M的磷酸缓冲液(PBS)冲洗4次,每次20 min。用乙醇水溶液按照乙醇浓度从低到高逐级脱水,30%、50%脱水2次,70%、90%脱水1次,100%脱水2次,每次10 min。脱水完成后将样品浸泡在叔丁醇中15 min。用二氧化碳临界点干燥仪进行干燥,喷金后,在扫描电子显微镜 (scanning electron microscope,SEM)下观察孢子丝形态。 -
从菌株HNM0301测序获得的全基因中提取全长16S rRNA基因序列,在Ezbiocloud数据库进行序列对比,采用软件MAGA 11.0构建系统发育树。参考在16S rRNA水平上的相似菌株,从NCBI中下载其相似菌株的全基因组序列,通过在线网站Type Strain Genome Server构建全基因组的系统发育树并进行数字 DNA-DNA 杂交(digital DNA-DNA hybridization,dDDH)分析。通过在线网站(
http://www.ezbiocloud.net/sw/oat )比较菌株HNM0301与亲缘关系最近种的全基因组,计算平均核苷酸一致性(average nucleotide identity,ANI)。 -
将菌株HNM0301接种到ISP2液体培养基中,在28 ℃,180 rpm条件下培养4 d,离心弃上清液,获取菌体,用无菌水清洗两次。送至上海美吉生物公司进行DNA提取和全基因组测序工作。使用PacBio SequelⅡ和Ⅰllumina NovaSeq6000测序平台进行全基因组测序,利用PacBio SequelⅡ和Ⅰllumina NovaSeq6000平台生成的数据进行生物信息学分析,所有分析均在上海美吉生物云平台(
http://cloud.majorbio.com )上进行。采用antiSMASH 6.0 (http://antismash.secondarymetabolites.org )在线网站预测菌株HNM0301的次级代谢生物合成基因簇[8]。通过NCBI(https://www.ncbi.nlm.nih.gov )进行staurosporine合成基因簇的比对。 -
菌株HNM0301在FEM5液体培养基中发酵(30 L)结束后,合并发酵液,向发酵液中加入10 L乙酸乙酯,经超声破碎仪进行菌体细胞破碎,萃取3次,合并乙酸乙酯相,减压浓缩后得到乙酸乙酯萃取物(196.0 g)。乙酸乙酯提取物经减压硅胶柱色谱,以石油醚-氯仿和氯仿-甲醇为流动相梯度洗脱,得到13个流份(Fr.1-Fr.13)。Fr.8(20.1 g)经RP-18柱色谱,以甲醇-水梯度洗脱得到21个流份(Fr.8-1-Fr.8-21)。Fr.8-16(552.7 mg)经硅胶柱色谱分离,以石油醚-乙酸乙酯(体积比12∶1-5∶1)进行梯度洗脱分离得12个流份其中Fr.8-16-7(105.7 mg)经Sephadex LH-20凝胶柱色谱,以氯仿-甲醇(V氯仿∶V甲醇=1∶1)、纯甲醇、纯丙酮系统洗脱后,以乙腈-酸水系统(V乙腈∶V酸水=35∶56)为流动相,经高效液相色谱(HPLC-CLS)半制备分离得到化合物1 (tR8.9 min,3.8 mg)。
-
将Foc TR4菌块分别接种于PDA培养基和加有星孢菌素质量浓度为20 μg·mL−1的PDA培养基中央,培养5 d后,观察星孢菌素对香蕉枯萎病的抑制作用,抑制率计算公式:
$$\begin{split} \text{抑菌率(\%)}=&(对照组菌落直径-处理组菌落直径)/\\&对照组菌落直径\times 100。 \end{split} $$ 采用96孔微量滴定法进行星孢菌素抗Foc TR4的MIC值测定[16]。
-
取900 μL Foc TR4孢子悬浮液(106 CFU·mL−1),加入星孢菌素甲醇溶液100 μL,使终浓度为15 μg·mL−1,充分混匀后,取20 μL加到凹玻片上,湿润环境孵育12 h。以添加100 μL甲醇的孢子液为对照,每个处理重复3次。在显微镜下观察孢子萌发情况,当孢子的芽管伸出长度大于孢子的短半径时,即视为萌发。每个处理计数200个孢子,计算星孢菌素对Foc TR4孢子萌发的抑制率。
-
配置PDA固体培养基,趁培养基未完全凝固(45~60 ℃)时,在培养基中加入星孢菌素,使终质量浓度为15 μg·mL−1,Foc TR4接种到培养基中心。PDA固体培养基上正常生长的Foc TR4为对照。生长3 d后,在扫描电镜下观察菌丝形态。
-
菌株HNM0301在平板对峙实验中对香蕉枯萎病菌(Fusarium oxysporum f. sp. Cubense tropical race 4)和豇豆枯萎病菌(Fusarium oxysporum f. sp. tracheiphlium)抑制率达86%,显示出较强抑制作用,对火龙果黑腐病菌(Bipolaris cactivora)、番茄早疫病菌(Alternaria solani)、从赤壳菌(Nectria citri)的抑制率为67% ~71%,对火龙果溃疡病菌(Neoscytaldium dimidiatum)、菠萝黑腐病菌(Ceratocystis paradoxa)和小麦赤霉病菌(Fusarium graminearum)抑制作用不明显(表1,图1)。
表 1 菌株HNM0301抗植物病原菌活性
Table 1. Antifungal activity of strain HNM0301
病原真菌
Pathogenic fungi真菌抑菌率/%
(Antifungal rate/%)从赤壳菌
(Nectria citri)67 豇豆枯萎病菌
(Fusarium oxysporum
f. sp. Tracheiphlium)86 番茄早疫病菌
(Alternaria solani)71 火龙果黑腐病菌
(Bipolaris cactivora)71 火龙果溃疡病菌
(Neoscytaldium dimidiatum)— 菠萝黑腐病菌
(Ceratocystis paradoxa)— 小麦赤霉病菌
(Fusarium graminearum)— 香蕉枯萎病菌
(Fusarium oxysporum
f. sp.cubense tropical race 4)86 注:“—”为没有抑菌效果。
Note: “—” means no antifungal effect.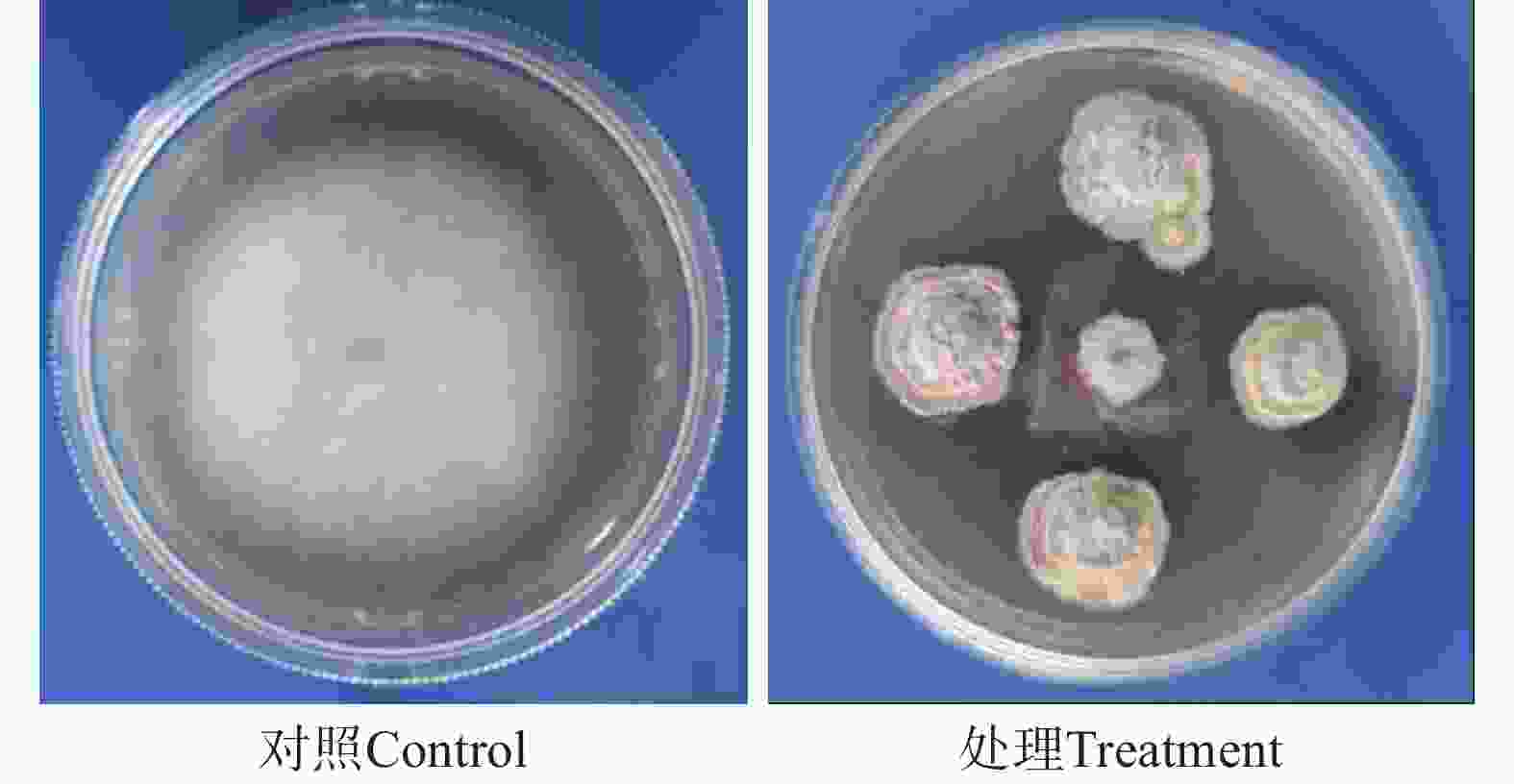
图 1 菌株HNM0301拮抗香蕉枯萎病菌活性
Figure 1. Activity of strain HNM0301 against Foc TR4
-
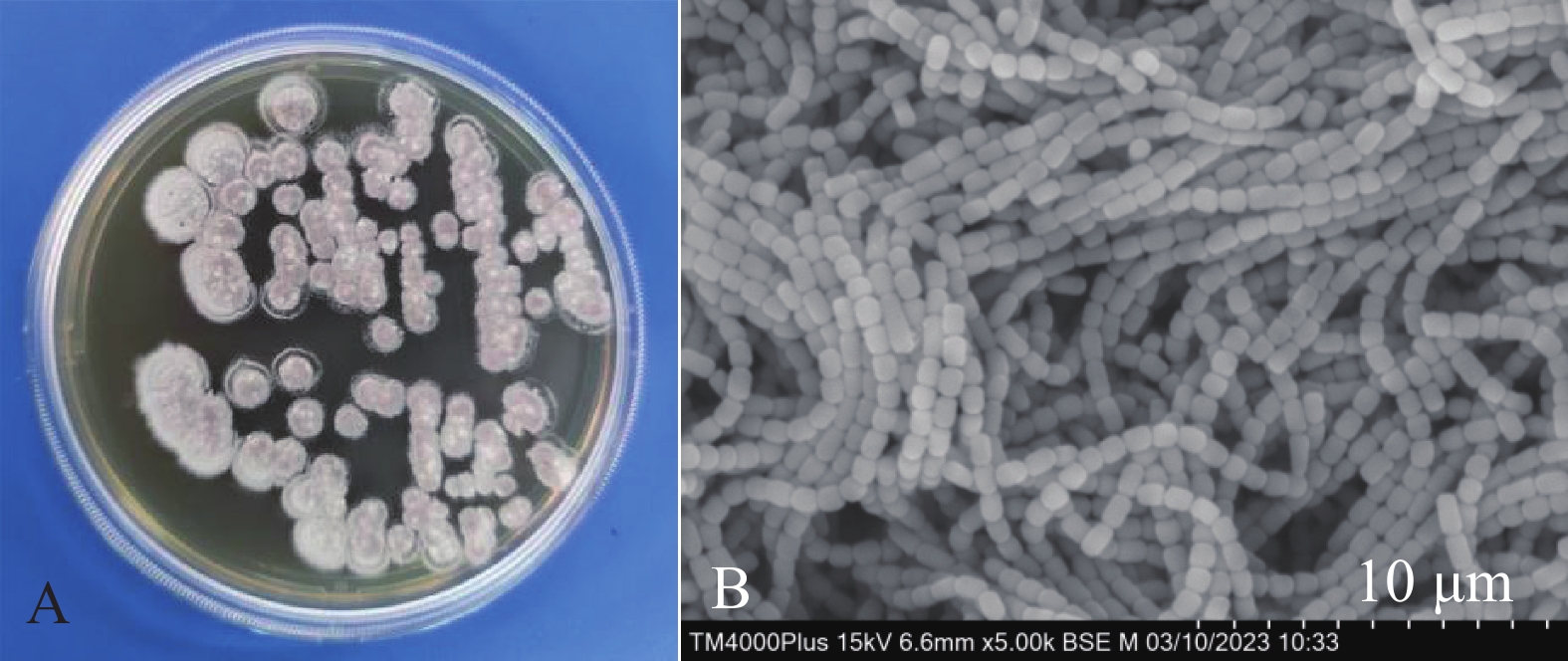
菌株HNM0301在ISP2固体培养基上长势良好,基内菌丝和气生菌丝发达。菌落培养初期呈白色,随时间增长慢慢呈灰白色(图2-A)。孢子丝为多个圆柱状孢子串联成链状结构,孢子成熟后链状菌丝断裂,呈单个圆柱状孢子(图2-B)。
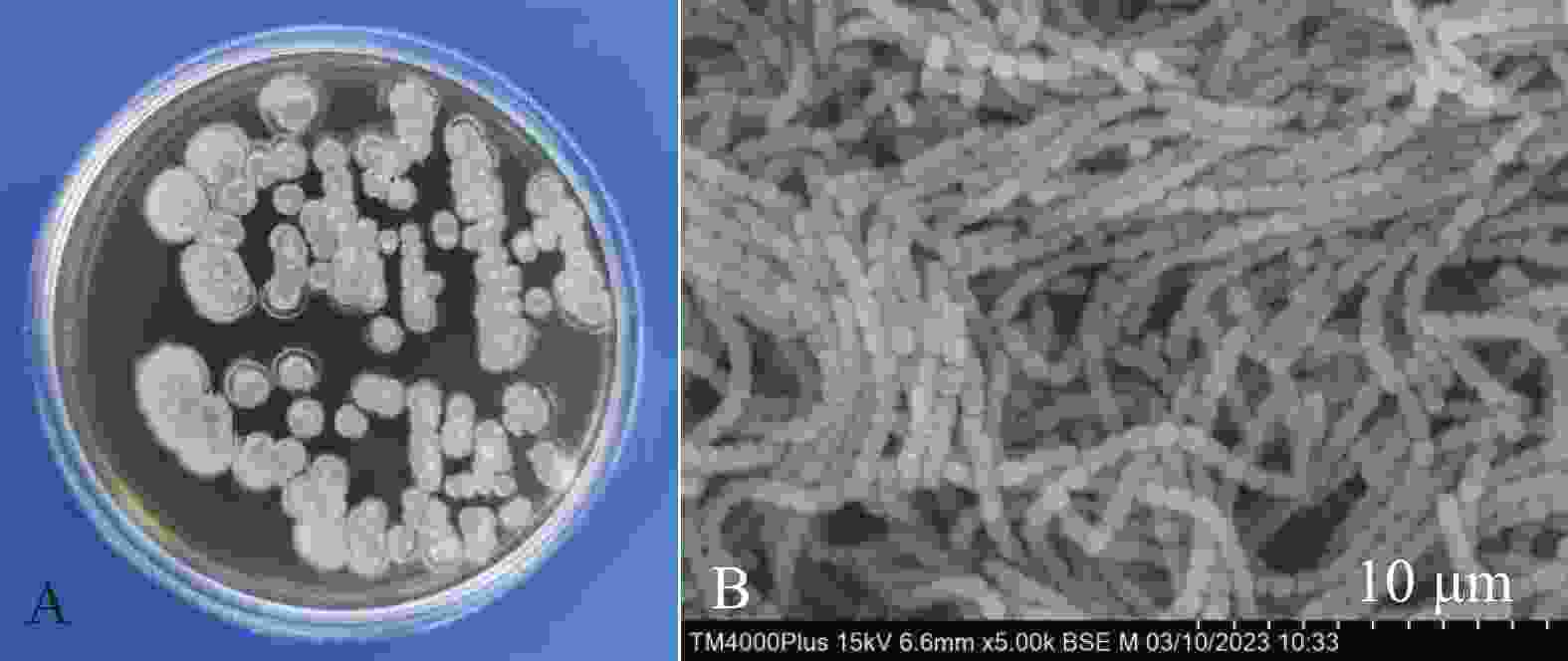
图 2 菌株HNM0301菌落形态(A)和孢子丝形态特征(B)
Figure 2. Colony morphology(A)and spore hypha morphological characteristics(B)of strain HNM0301
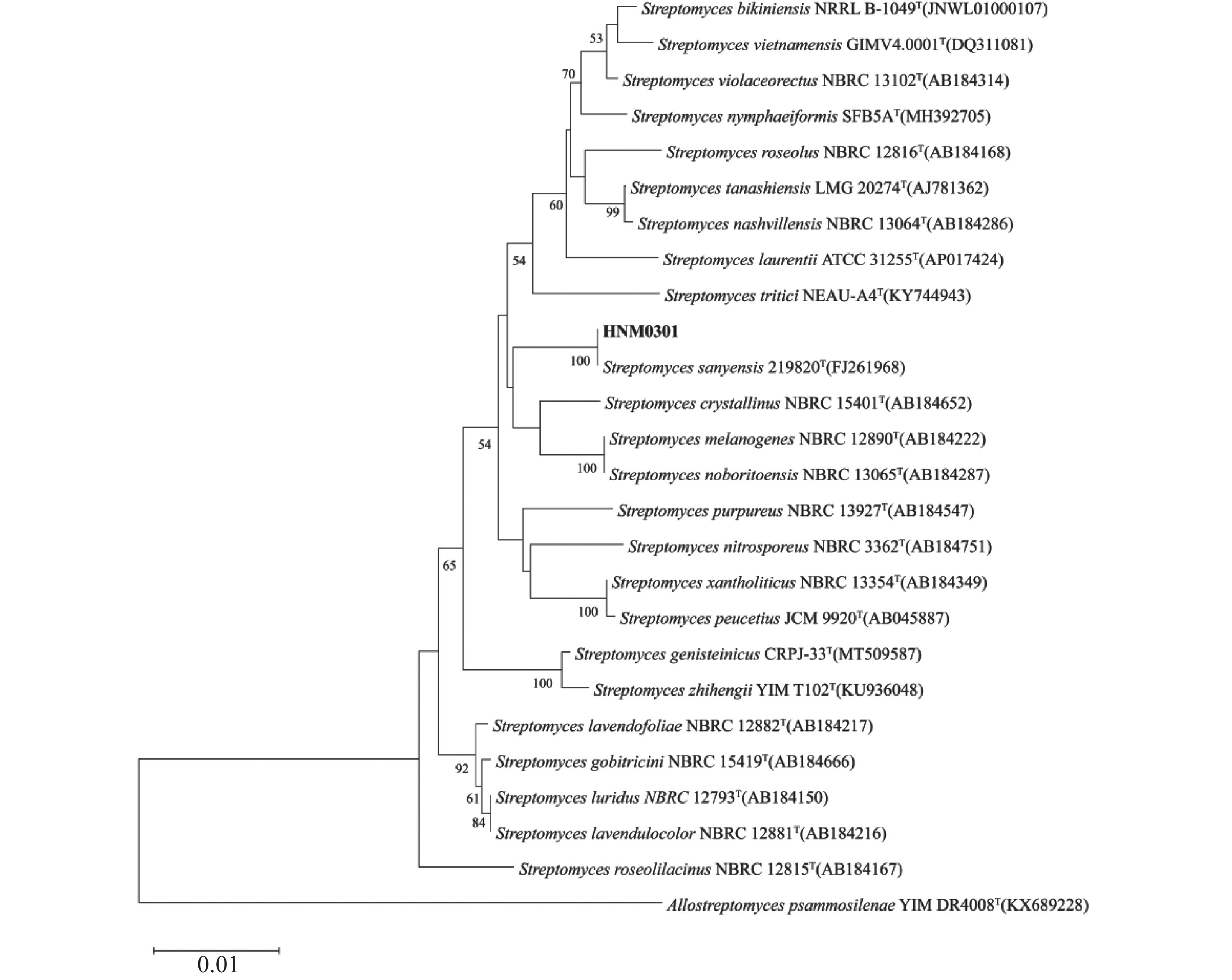
从菌株HNM0301基因组中提取得全长16S rRNA基因序列为1 525 bp,16S rRNA系统发育树结果显示,菌株HNM0301与链霉菌属成员关系紧密,特别是与Streptomyces sanyensis
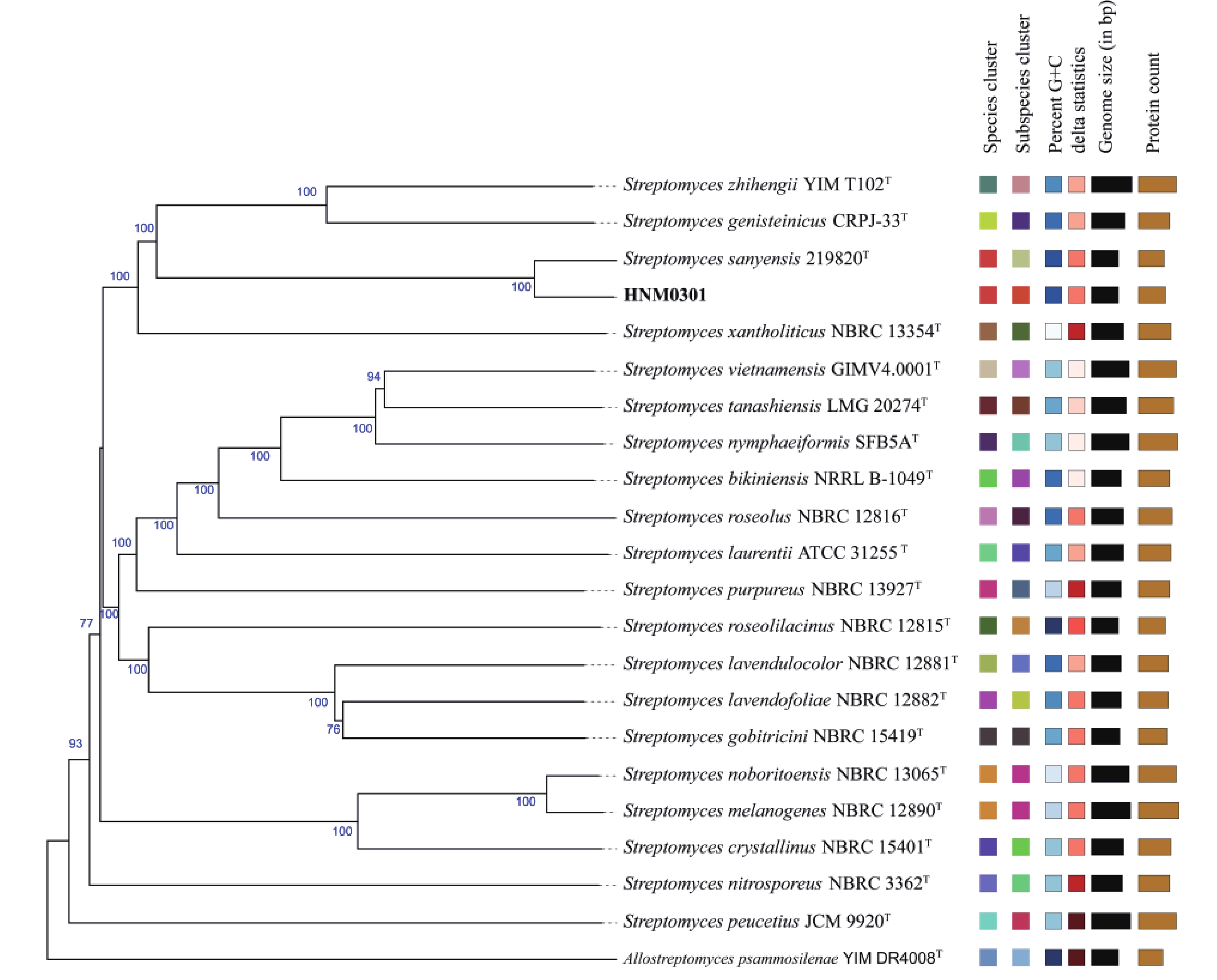
219820 T聚集在同一分支(图3)。基因组系统发育树进一步揭示,菌株HNM0301和Streptomyces sanyensis219820 T同样聚在同一分支,显示出二者存在密切的亲缘联系(图4)。平均核苷酸一致性(ANI)分析得出,菌株HNM0301与Streptomyces sanyensis219820 T的ANI值为96.83,大于95%~96%的种水平阈值;而DNA-DNA杂交(dDDH)测定值为72.4,同样超出了70%的新物种界定标准[17](表2)。因此,基于上述结果,鉴定菌株HNM0301为三亚链霉菌Streptomyces sanyensis。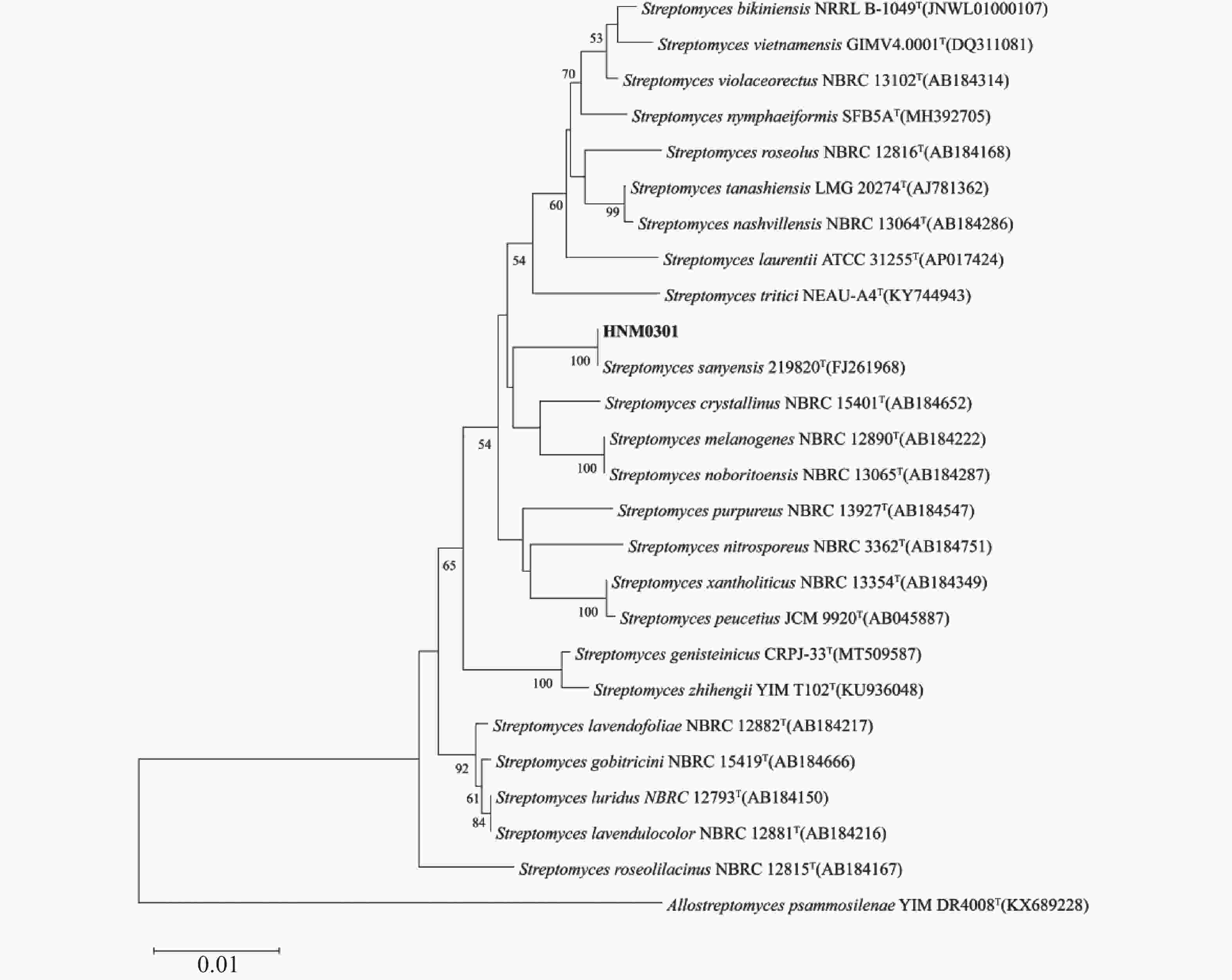
图 3 菌株HNM0301 16S rRNA基因系统发育树
Figure 3. 16S rRNA p\hylogenetic tree of strain HNM0301
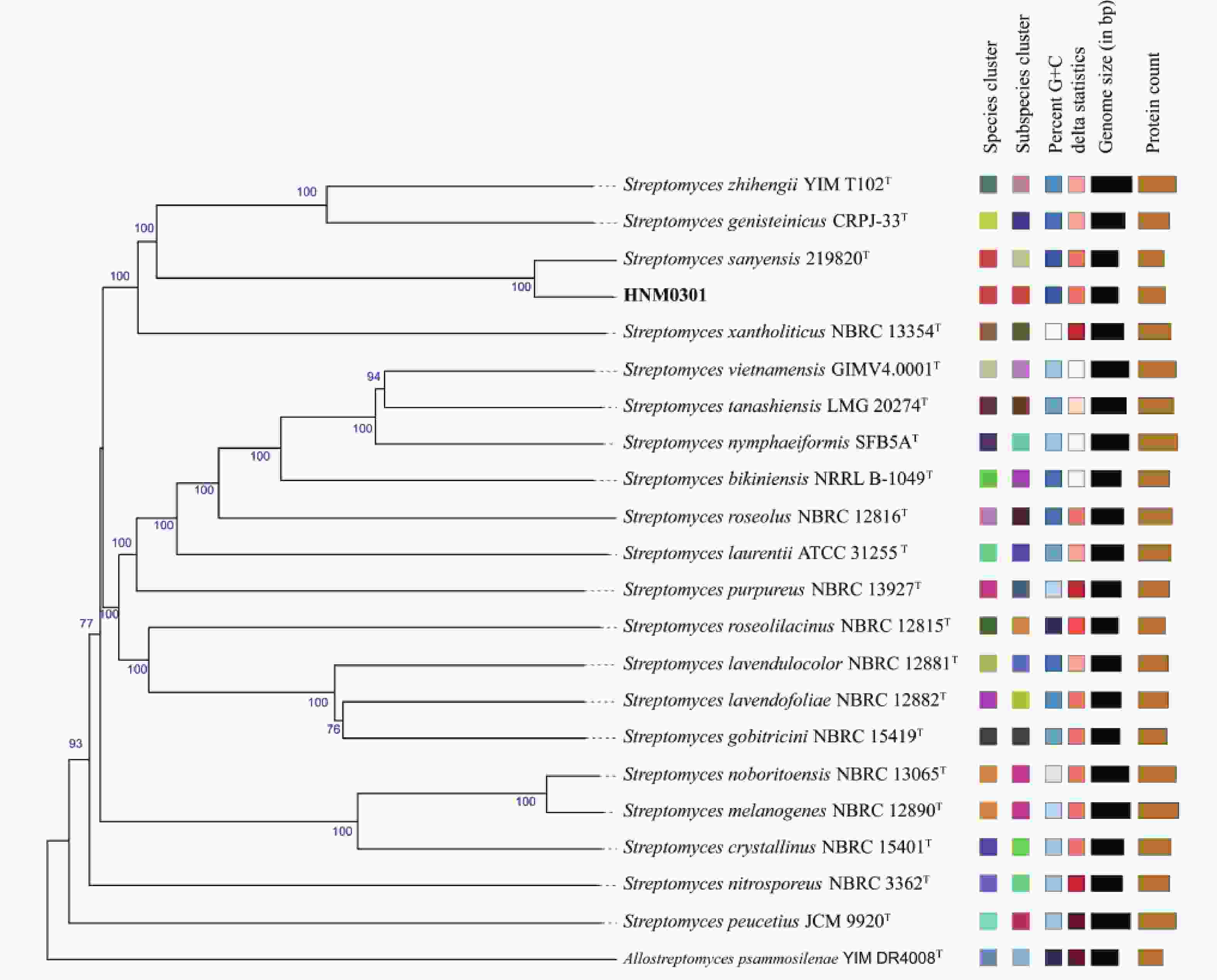
图 4 菌株 HNM0301 的全基因组系统发育树
Figure 4. Whole genome phylogenetic tree of strain HNM0301
表 2 菌株 HNM0301 与最相近种之间的 ANI 值和 dDDH 值
Table 2. Values of ANI and dDDH between HNM0301 and its closest species
模式菌株
(Type strain)HNM0301 ANI dDDH Streptomyces sanyensis JCM 18324 (BAABJV010000001)96.83 72.4 Streptomyces zhihengii YIM T102(JAFEJA010000003) 82.55 26.3 Streptomyces genisteinicus CRPJ-33(CP060825) 82.54 26.3 Streptomyces xantholiticus JCM 4863 (BMWB01000001)80.86 24.7 Streptomyces gobitricini JCM 5062 (BAAASR010000001)80.16 23.9 Streptomyces lavendofoliae JCM 4391 (BMTP01000001)80.08 23.7 Streptomyces purpureus KA281(KB913030) 80.02 23.7 Streptomyces lavendulocolor JCM 4390 (BAAASF010000001)79.99 23.8 Streptomyces bikiniensis NRRL ISP- 5580 (JNXJ01000001)79.92 23.9 Streptomyces roseolilacinus JCM 4335 (BMSV01000001)79.81 23.7 Streptomyces vietnamensis GIM4.0001(CP010407) 79.69 23.7 Streptomyces crystallinus JCM 5067 (BAAACA010000001)79.55 23.3 Streptomyces roseolus JCM 4411 (BMTV01000001)79.44 23.6 Streptomyces tanashiensis Kala(CP084204) 79.42 23.5 Streptomyces noboritoensis JCM 4557 (JBHMQV010000001)79.37 23.4 Streptomyces nymphaeiformis SFB5A(JACHJY010000001) 79.31 23.5 Streptomyces nitrosporeus ATCC 12769 (CP023702)79.21 23 Streptomyces melanogenes JCM 4398 (BMTS01000001)79.16 23.2 Streptomyces laurentii NPDC006966(JBEYHT010000001) 79.15 23.2 Streptomyces peucetius NRRL WC- 3868 (JOCK01000001)77.87 22.6 -
菌株HNM0301的基因组经过测序、组装和注释,得到了全基因组信息(表3)和染色体圈图(图5-A)。菌株HNM0301基因组全长为6 635 340 bp,含有5 859个蛋白质编码基因(CDS)。菌株HNM0301全基因组的GenBank登录号为CP193716,16S rRNA的GenBank登录号为PV933158。
表 3 菌株HNM0301 的基因组特性
Table 3. Genome features of HNM0301
特征 (Feature) 染色体信息 (Chromosome Characteristics) 染色体长度/bp(Chromosome size/bp) 6 635 340 GC含量/%(GC content/%) 73.71 蛋白质编码基因(Protein-coding genes) 5859 基因平均长度/bp(Gene average length/bp) 959.97 具有COG注释的基因(Genes assigned to COG) 4 297 具有KEGG注释的基因(Genes assigned to KEGG) 1 243 具有GO注释的基因(Genes assigned to GO) 2 137 核糖体RNA基因(rRNA genes) 18 转运RNA基因(tRNA genes) 64 微小RNA基因(sRNA genes) 51 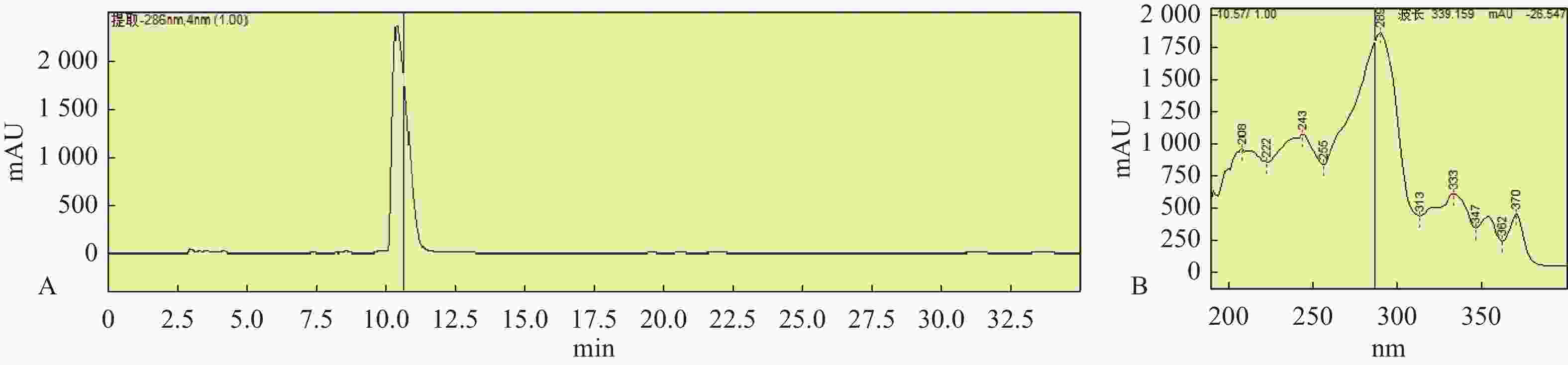
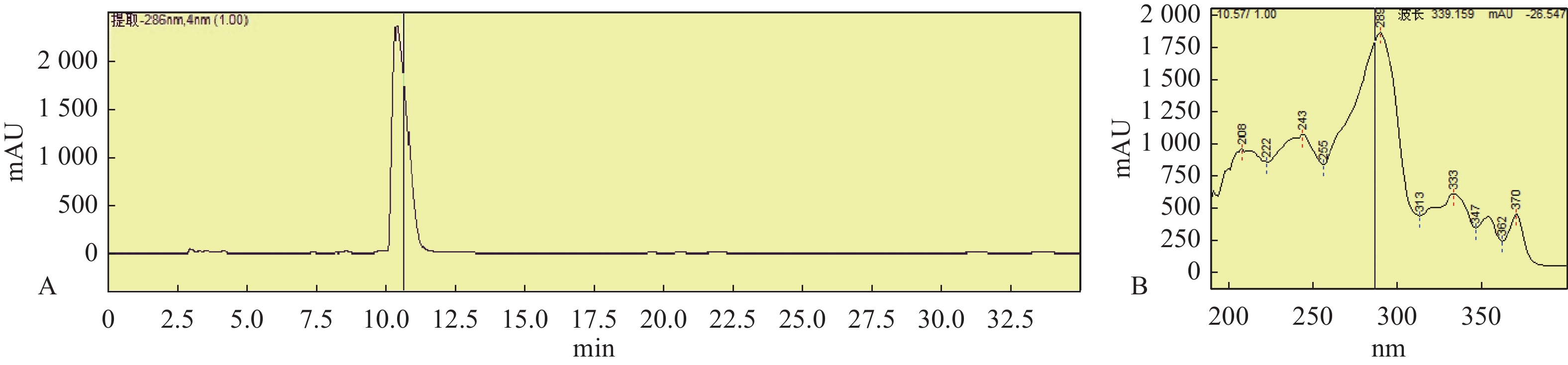
图 5 星孢菌素HPLC图谱(A)和星孢菌素在波长289 nm处吸收峰(B)
Figure 5. Staurosporine HPLC chromatogram(A)and absorption peak of staurosporine at 289 nm (B)
-
通过antiSMASH预测发现菌株HNM0301含有27个次级代谢合成基因簇。其类型包括3个非核糖体肽合成酶(nonribosomal peptide synthetase,NRPS)型、2个聚酮合酶(polyketide synthases,PKSs)型、5个杂合型、5个萜类(terpene)、2个黑色素(melanin)、非NRPS依赖型铁载体(NRPS-independent-siderophore)、Ⅳ类羊毛硫肽(lanthipeptide-class-Ⅳ)、氨基多羧酸(aminopolycarboxylic-acid)、β-内酯(betalactone)、Ⅲ类羊毛硫肽(lanthipeptide-class-Ⅲ)、吲哚(indole)、线性肽类(linaridin)、四氢嘧啶(ectoine)、丁内酯(butyrolactone)各1个,以及其他类型1个(表4)。基因簇的相似度分析显示,5个基因簇与已知基因簇的相似度为100%,分别是Cluster2、12、18、22、26。这说明菌株HNM0301具有产生次级代谢产物委内瑞拉菌素(venezuelin)、黑色素(melanin)、去铁胺B(desferrioxamin B)、四氢嘧啶(ectoine)、土臭素(geosmin)的能力。3个基因簇与已报道过的基因簇相似度大于60%,分别是Cluster3、5、16。这说明,菌株HNM0301具有产生次级代谢产物异海绵烯(isorenieratene)、何帕烯(hopene)、星孢菌素(staurosporine)或者其类似物的潜力。还有15个基因簇与已报道过的基因簇的相似性低于60%,4个基因簇是未知基因簇,这表明菌株HNM0301具有产生一些新的次级代谢产物或分子的潜力。
表 4 菌株HNM0301预测的生物合成基因簇
Table 4. Predicted biosynthetic gene cluster of HNM0301
基因编号
(Cluster)类型
(Type)位置
(Position)已知基因相似度
(Most similar known biosynthetic gene cluster)1 非NRPS依赖型铁载体
(NRPS-independent-siderophore)215 382~227 026 聚克菌素
(Peucechelin)(25%)2 Ⅳ型羊毛硫肽
(lanthipeptide-class-Ⅳ)235 608~258 280 委内瑞拉菌素
(venezuelin)(100%)3 类NRPS-NRPS-萜杂合途径
(NRPS-like,NRPS,terpene)392 445~468 881 异海绵烯
(isorenieratene)(75%)4 非核糖体肽合成酶
(NRPS)497 124~543 572 环吡双环烯/氮杂双环烯B-D
(clipibicyclene/azabicyclene B/
azabicyclene C/azabicyclene D)(22%)5 萜类(terpene) 569 082~594 976 何帕烯(hopene)(69%) 6 Ⅰ型聚酮合酶
(T1PKS)955 184~1 004 505 福克辛A-C
(foxicin A/foxicin B/foxicin C/foxicin)(12%)7 Ⅱ型聚酮合酶(T2PKS) 1 136 46~1 269 167 Sch- 47554 /Sch-47555 )(21%)8 氨基多羧酸
(aminopolycarboxylic-acid)1 330 766~1 346 736 乙二胺羟基乙酸
(EDHA)(33%)9 非NRPS依赖型铁载体
NRPS-independent-siderophore1 432 445~1 445 628 - 10 其他
(other)1 820 060~1 861 973 A- 503083 A/A-503083 B/A-503083 E/A-503083 F
(3%)11 β-内酯类(betalactone) 2 198 156~2 224 871 萘霉素A(naphthomycin A)(6%) 12 黑色素(melanin) 2 293 489~2 300 967 黑色素(melanin)(100%) 13 Ⅲ型羊毛硫肽
(lanthipeptide-class-Ⅲ)2 776 024~2 798 540 - 14 类NRPS-NRPS-类PKS杂合途径
(NRPS-like,NRPS,PKS-like)2 819 955~2 882 442 里莫霉素A-C
(rimomycin A/rimomycin B/rimomycin C)(13%)15 transAT型PKS-NRPS杂合途径
(transAT-PKS,NRPS)3 244 144~3 320 517 拉吉霉素38/39
(largimiycin 38/largimycin 39)(51%)16 吲哚类(indole) 3 947 666~3 971 034 星孢菌素(staurosporine)(93%) 17 萜类(terpene) 4 344 692~4 365 549 波卓霉素A2(bottromycin A2)(24%) 18 非NRPS依赖型铁载体
(NRPS-independent-siderophore)4 404 710~4 416 482 去铁胺B
(desferrioxamin B)(100%)19 萜类(terpene) 4 506 981~4 524 545 - 20 类NRPS-NRPS-萜杂合途径
(NRPS-like,NRPS,betalactone)4 648 053~4 695 032 阿米霉素
(amicetin)(8%)21 线性肽类
(linaridin)4 719 408~4 739 986 链脲佐菌素
(streptozotocin)(15%)22 四氢嘧啶类
(ectoine)5 247 149~5 257 547 四氢嘧啶
(ectoine)(100%)23 羊毛硫肽/噻唑肽/RRE结构域相关肽
(LAP,thiopeptide,RRE-containing)5 803 975~5 838 819 - 24 萜类(terpene) 5 853 508~5 870 658 埃贝内酯(ebelactone)(5%) 25 丁酸内酯类(butyrolactone) 6 133 180~6 144 139 内酯霉素(lactonamycin)(3%) 26 萜类(terpene) 6 284 445~6 306 643 土臭素(geosmin)(100%) 27 黑色素(melanin) 6 391 082~6 401 489 黑色素(melanin)(42%) 比较分析各基因簇预测的次级代谢产物发现,Cluster 16预测的编码产物为星孢菌素(staurosporine),根据文献报道,星孢菌素具有广泛地抗真菌活性[18],因此推测菌株HNM0301的抗香蕉枯萎病活性可能来源于化合物星孢菌素或其类似物的产生。将Cluster 16与来自Streptomyces sp. TP-A0274的staurosporine合成基因簇进行比较,发现两者相似度为93%,且菌株HNM0301含有负责星孢菌素合成的14个基因(图6和表5)。尽管Cluster 16中缺失了调控因子staR基因,但理论上不会影响关键结构的合成。因此,基于以上分析结果,本研究预测菌株HNM0301的抗真菌活性物质可能为星孢菌素类化合物。
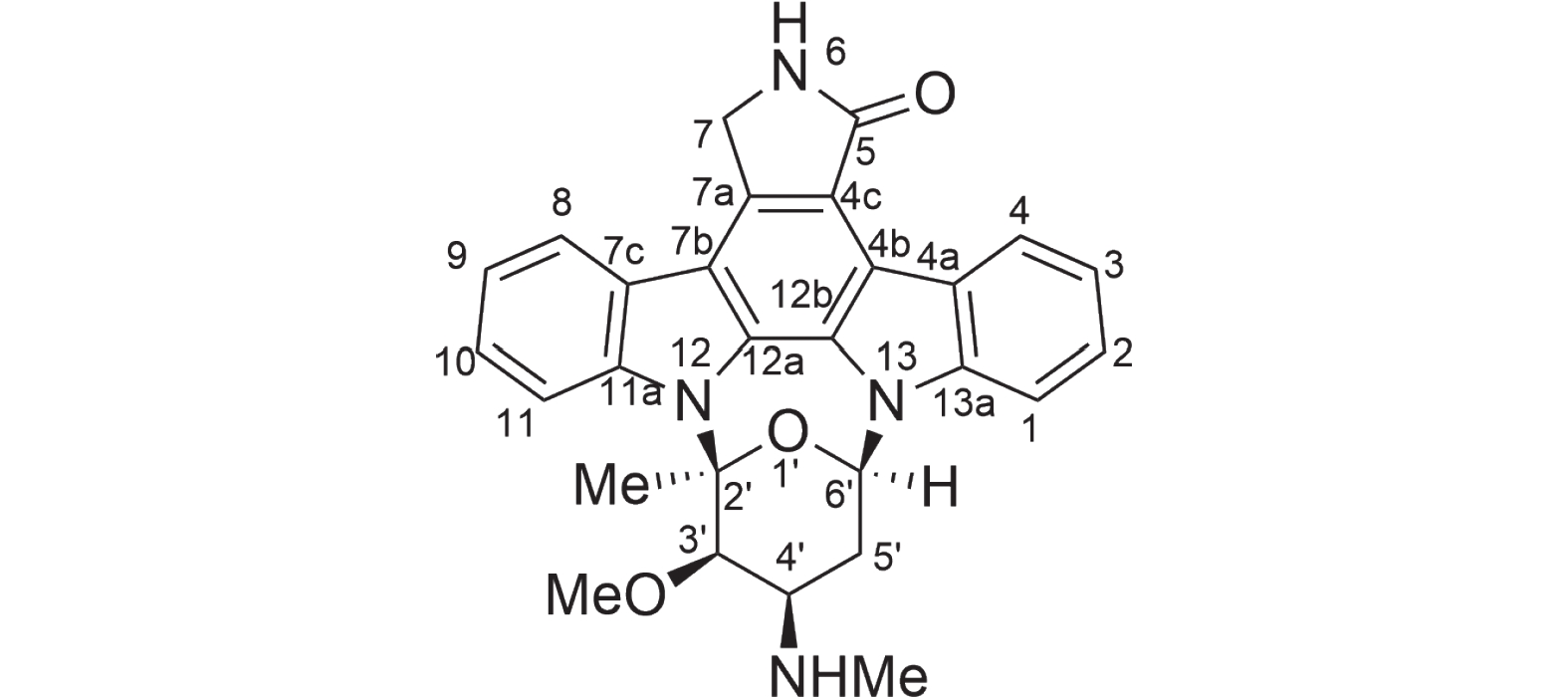
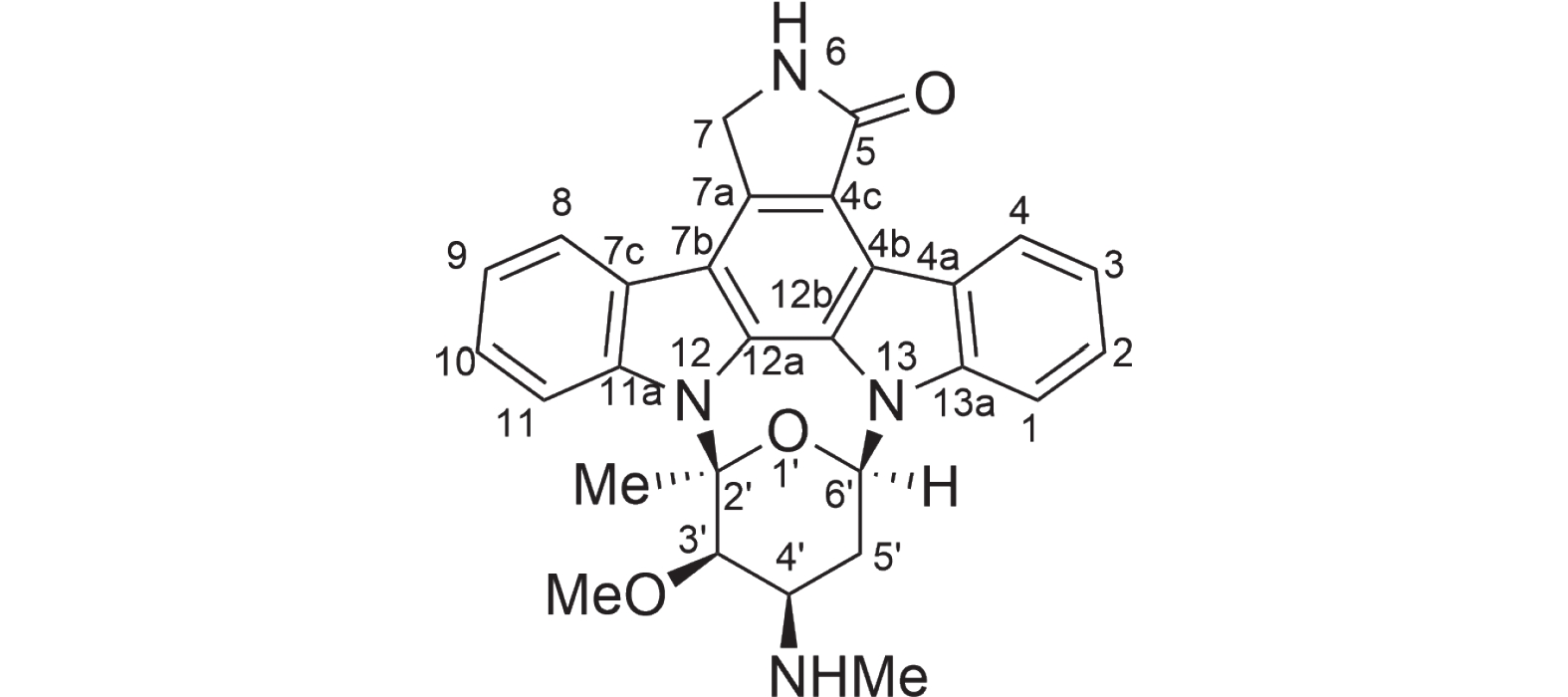
图 6 化合物1星孢菌素结构
Figure 6. Compound 1 structure of staurosporine
表 5 HNM0301的Cluster 16与staurosporine合成基因簇比对
Table 5. Synthetic gene cluster alignment of Cluster 16 and staurosporine
基因ID
(Gene ID)氨基酸数
(Number of amino acids)同源基因ID
(Similar gene ID)氨基酸数
(Number of amino acids)同源基因一致性/%
(Positives/%)ctg1_ 3466 545 staC 535 87 ctg1_ 3467 281 staMB 280 87 ctg1_ 3468 208 staE 206 93 ctg1_ 3469 369 staI 369 91 ctg1_ 3470 331 staK 328 84 ctg1_ 3471 477 staJ 472 85 ctg1_ 3472 277 staMA 276 76 ctg1_ 3473 422 staP 417 77 ctg1_ 3474 1122 staD 1096 76 ctg1_ 3475 504 staO 503 88 ctg1_ 3476 436 staG 446 88 ctg1_ 3477 414 staN 394 87 ctg1_ 3478 350 staA 354 92 ctg1_ 3479 350 staB 334 85 — — staR 943 — 注:“—”为缺失该基因。 Note:“—” means deletion of the gene. -
为了证实基因组分析的结果,对菌株HNM0301进行了发酵培养,提取其发酵粗提物通过萃取、正向硅胶色谱、反向硅胶色谱、半制备分离得到一个单体化合物1,使用HPLC指纹图谱验证其纯度,并检查其吸收峰(图5)。该化合物为墨绿色粉末,通过高分辨电喷雾质谱(high resolution electrospray ionization mass spectroscopy,HRESI-MS)发现该化合物的准分子离子峰[M+H]+ m/z 467.1结合1HNMR和13CNMR谱数据推测其分子式C28H26N4O3。化合物1的核磁数据如下:1H-NMR(500 MHz,CD3OD)δH: 6.97(1H, d, J = 7.8 Hz, H-1), 7.30(1H, t, J = 7.4 Hz, H-2), 7.18(1H, t, J = 7.4 Hz, H-3), 9.16(1H, d, J = 7.8 Hz, H-4), 4.51(1H, d, J = 17.3 Hz, H-7a), 4.09(1H, d, J = 17.3 Hz, H-7b), 7.62(1H, d, J = 7.8 Hz, H-8), 7.32(1H, t, J = 7.4 Hz, H-9), 7.46(1H, t, J =7.8 Hz, H-10), 7.90(1H, d, J = 8.4 Hz, H-11), 4.12(1H, brs, H-3'), 3.86(1H, m, H-4'), 3.12(1H, m, H-5'a), 2.61(1H, m, H-5'b), 6.16(1H, d, J = 9.3 Hz, H-6'), 2.47(3H, s, 2'-CH3), 3.06(3H, m, 3'-OCH3), 2.75(3H, s, 4'-NHCH3); 13C-NMR(125 MHz, CD3OD)δC: 109.3(C-1), 126.2(C-2), 120.5(C-3), 127.1(C-4), 124.3(C-4a), 116.5(C-4b), 119.9(C-4c), 175.0(C-5), 46.6(C-7), 133.5(C-7a), 115.5(C-7b), 125.7(C-7c), 122.6(C-8), 121.8(C-9), 126.4(C-10), 113.3(C-11), 139.2(C-11a), 131.2(C-12a), 127.4(C-12b), 137.4(C-13a), 94.0(C-2'), 81.4(C-3'), 55.8(C-4'), 28.7(C-5'), 81.7(C-6'), 28.6(2'-CH3), 60.3(3'-OCH3), 31.2(4'-NHCH3)。以上数据与文献已报道的化合物星孢菌素的数据一致[19],故化合物1结构被鉴定为吲哚咔唑类化合物星孢菌素(图6)。
-
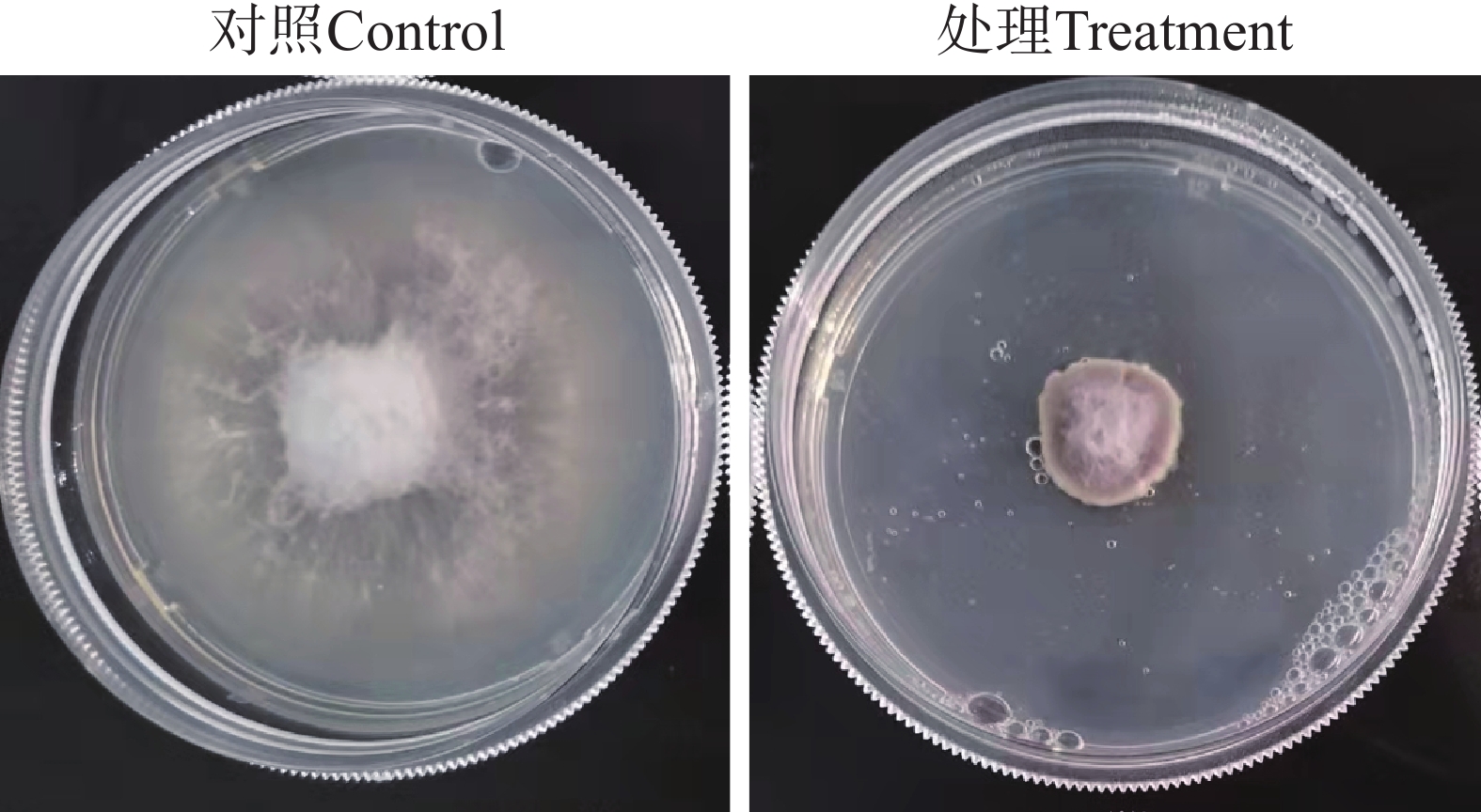
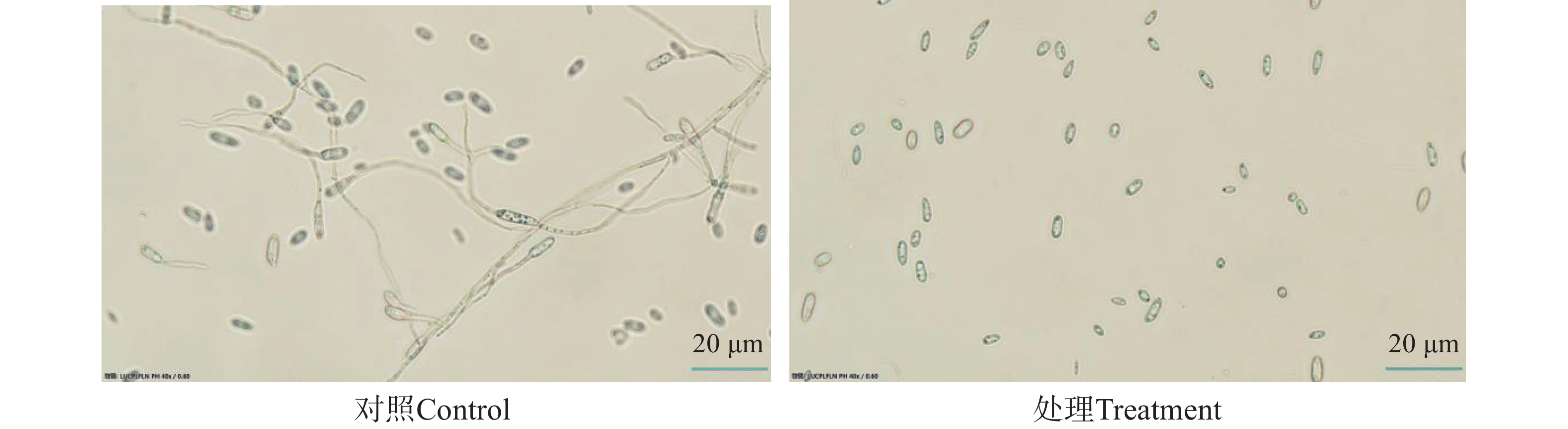
经过检测,星孢菌素在平板上能够显著地抑制Foc TR4菌丝的生长(图7),MIC为1.56 μg·mL−1。扫描电子显微镜观察显示,在对照组中,Foc TR4的菌丝形态表面较光滑,条索清晰,在培养基上总体朝相同方向有规律生长;而在含有星孢菌素的培养基上生长的菌丝则表现出间隔膨胀,弯曲、乱序生长,菌丝衰老干瘪后呈现出表面皱缩破损现象(图8)。此外,星孢菌素质量浓度为15 μg·mL−1时,对Foc TR4孢子的萌发产生较强的抑制作用(图9),抑制率达85.4%。
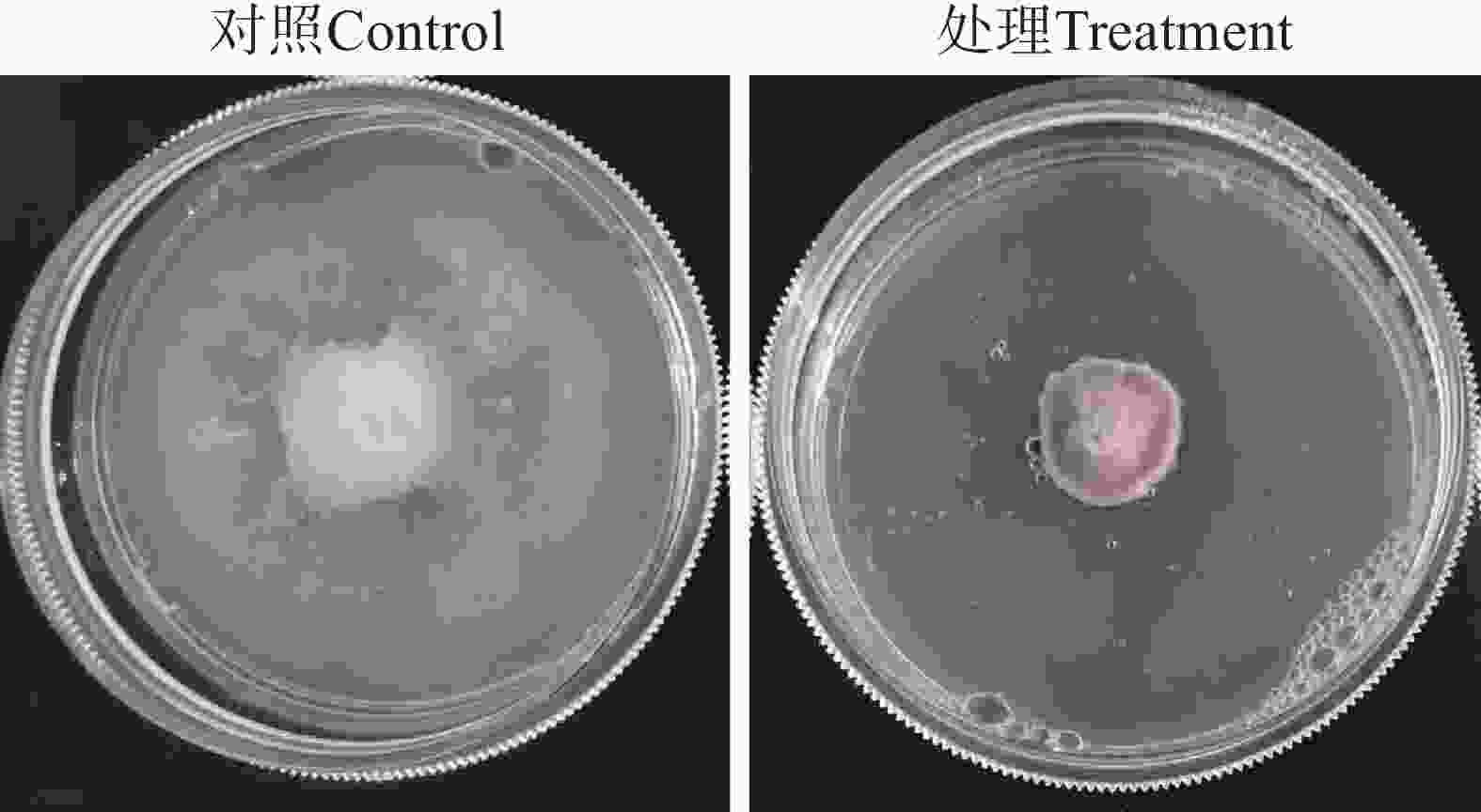
图 7 星孢菌素在平板上对 Foc TR4的抑制效果
Figure 7. Inhibitory effect of staurosporine on Foc TR4

图 8 星孢菌素作用 Foc TR4菌丝的SEM图
Figure 8. SEM image of Foc TR4 hyphae treated with staurosporine

图 9 星孢菌素对Foc TR4孢子的抑制作用
Figure 9. Inhibition of Foc TR4 spores by staurosporine
-
放线菌是自然界中分布最广泛的微生物种类之一,特别是链霉菌属放线菌,可以产生大量的天然活性化合物,是农业和医药抗生素先导化合物筛选的重要来源[20]。红树林作为特殊的海洋生境,具有高盐、高腐殖质、有氧和无氧交替的复杂环境更容易促使微生物产生特殊的代谢途径,产生结构新颖、活性丰富的天然产物[21]。因此,红树林被认为是分离拮抗放线菌和发掘抗菌活性次级代谢产物的理想环境[22 − 23]。本研究从海南东寨港红树林自然保护区红树植物海桑根际土壤分离筛选出了1株对Foc TR4具有良好抑制效果的放线菌菌株HNM0301;通过对其培养特征、16S rRNA基因序列聚类和全基因组序列分析,将菌株HNM0301鉴定为三亚链霉菌Streptomyces sanyensis,该菌不仅对Foc TR4呈现出抗菌活性,还对其他病原真菌具有抑制作用,结果表明,该菌产生的活性代谢物具有广谱抗植物病原真菌活性,具备开发成广谱抗真菌生防剂和农用抗生素的潜力。
三亚链霉菌(Streptomyces sanyensis)最初是洪葵课题组从海南三亚红树林发现的1株具有抗肿瘤活性的海洋放线菌新种[24]。朱伟明课题组从三亚链霉菌FMA 菌株的发酵产物中分离到了具有显著抗肿瘤活性的新颖的吲哚咔唑类化合物streptocarbazoles A和 B[25]。李文利课题组从三亚链霉菌FMA 中从克隆了吲哚咔唑类化合物的生物合成基因簇,并成功在天蓝色链霉菌Streptomyces coelicolor M1152中异源表达获得了包括星孢菌素在内的一列吲哚咔唑类化合物[26]。Cartuche等[27]从西班牙的红树林也分离到三亚链霉菌,并从中分离出星孢菌素,发现该化合物能通过激活线粒体程序性死亡对寄生虫棘阿米巴表现出显著的抗虫活性。
本研究通过基因组测序获得了红树林来源的三亚链霉菌HNM0301的基因组完成图,从中预测到了1个与星孢菌素合成基因簇相似率达93%的基因簇(Cluster 16),并最终通过各种色谱手段分离得到了星孢菌素单体化合物,再一次证实了三亚链霉菌不仅在红树林环境广泛分布,而且是吲哚咔唑类化合物的重要产生菌源。
筛选抗香蕉枯萎病菌活性显著的拮抗菌株是研发香蕉枯萎病生防菌的首要步骤。而探明活性菌株产生的活性化合物是研究生防菌作用机制的重要步骤。Chen等[28]从海洋链霉菌(Streptomyces yongxingensis) 2-11,通过活性追踪分离到了化合物尼菲霉素C,对Foc TR4表现出显著抗性,并通过抑制Foc TR4三羧酸循环关键酶活,造成线粒体的损伤,进而导致香蕉枯萎病菌死亡。随后,Chen等[15]从海洋链霉菌S. solisilvae 2-6中发现了对Foc TR4具有抑制活性的化合物fluvirucin B6,通过抑制Foc TR4细胞毒素的代谢途径发挥抗菌作用。星孢菌素由于其良好的抗肿瘤活性而被视作抗肿瘤先导化合物进行广泛的药理学和机制研究[29]。但近年来其抗菌活性,特别是抗植物病原真菌活性日益受到关注而被广泛地报道,Park等[18]从湖底沉积物中分离的玫瑰孢链霉菌(Streptomyces roseoflavus)中得到星孢菌素,测得星孢菌素对辣椒疫霉菌(Phytophthora capsici)、立枯丝核菌(Rhizoctonia solani)、黄瓜炭疽菌(Colletotrichum orbiculare)、灰霉菌(Botrytis cinerea)黄瓜黑星病菌(Cladosporium cucumerinum)、苦瓜蔓枯病菌(Didymella bryoniae)、番茄枯萎病菌(Fusarium oxysporum f. sp. lycopersici)、番茄疮痂病菌(Xanthomonas campestris)都有较好的抑制效果,表明其在植物真菌病害的防控领域也显示出强大的应用潜力。本研究发现星孢菌素对Foc TR4的菌丝生长和分生孢子萌发都表现出良好的抑制效果,并导致Foc TR4菌丝膨胀、变形弯曲,表明其在香蕉枯萎病的防治上良好的应用前景,但具体抗菌机制还有待进一步研究。
-
本研究将来源于红树林的菌株HNM0301鉴定为Streptomyces sanyensis,参与其抗香蕉枯萎病活性的重要天然产物为星孢菌素,星孢菌素能够抑制Foc TR4子的萌发,并对Foc TR4菌丝有致畸作用,是一株极具开发前景的生防菌株。
Whole-genome sequencing of marine Streptomyces HNM0301 and identification of active compounds against Fusarium oxysporum f. sp. cubense tropical race 4
-
摘要: 为了筛选有效防治香蕉枯萎病菌 (Fusarium oxysporum f. sp. cubense tropical race 4,Foc TR4) 的海洋放线菌及天然产物,本研究通过全基因组序列分析对具有抗Foc TR4活性的海洋放线菌HNM0301进行种水平的分类鉴定,采用antiSMASH 6.0 在线软件对其编码的次级代谢产物合成基因簇进行预测,利用现代天然产物分离鉴定手段对其抗Foc TR4的活性成分进行分离和结构鉴定。结果表明:菌株HNM0301被鉴定为三亚链霉菌(Streptomyces sanyensis),其基因组全长为6 635 340 bp,含有5 859个蛋白质编码基因(CDS),其中2 137、1 243和4 297个CDS分别在GO、KEGG和COG数据库得到功能注释;在基因组中预测到27个次级代谢合成基因簇,其中Cluster 16含有星孢菌素(staurosporine)合成的14个基因;从菌株HNM0301发酵液中分离得到了一个星孢菌素单体化合物,其抗Foc TR4的MIC为1.56 μg·mL−1,对Foc TR4分生孢子萌发的抑制率为85.4%,并导致Foc TR4菌丝膨胀、畸形。菌株HNM0301在抗香蕉枯萎病菌上具有一定的开发应用潜力。Abstract: In order to screen out marine actinomycetes and natural products that can effectively control Fusarium oxysporum f. sp. cubense tropical race 4 (Foc TR4), a marine actinomycete strain HNM0301 isolated from a mangrove and exhibiting anti - Foc TR4 activity was identified taxonomically at a species level through whole - genome sequence analysis. The antiSMASH 6.0 online software was used to predict the biosynthetic gene clusters of secondary metabolites encoded by this strain. Moreover, modern separation and identification techniques for natural products were applied to isolate and elucidate the structures of the active ingredients against Foc TR4. The results demonstrated that the strain HNM0301 was identified as Streptomyces sanyensis. Its genome has a total length of 6,635,340 base pairs (bp) and contains 5,859 protein-coding genes (CDS). Among the protein-coding genes, 2,137, 1,243, and 4,297 CDS were functionally annotated in the Gene Ontology (GO), Kyoto Encyclopedia of Genes and Genomes (KEGG), and Clusters of Orthologous Groups (COG) databases, respectively. A total of 27 biosynthetic gene clusters of secondary metabolites were predicted within the genome, with Cluster 16 containing 14 genes involved in the biosynthesis of staurosporine. A staurosporine monomer compound was successfully isolated from the fermentation broth of the strain HNM0301. The minimum inhibitory concentration (MIC) of this compound against Foc TR4 was determined to be 1.56 μg/mL. This compound exhibited an 85.4% inhibition rate of Foc TR4 conidial germination and also induced swelling and malformation of Foc TR4 mycelia. Evidently, the strain HNM0301 holds certain potentials for development and application in the control of Foc TR4.
-
Key words:
- marine streptomyces /
- genome wide /
- Foc TR4 /
- Staurosporine
-
图 2 菌株HNM0301菌落形态(A)和孢子丝形态特征(B)
Fig. 2 Colony morphology(A)and spore hypha morphological characteristics(B)of strain HNM0301
图 5 星孢菌素HPLC图谱(A)和星孢菌素在波长289 nm处吸收峰(B)
Fig. 5 Staurosporine HPLC chromatogram(A)and absorption peak of staurosporine at 289 nm (B)
图 8 星孢菌素作用 Foc TR4菌丝的SEM图
Fig. 8 SEM image of Foc TR4 hyphae treated with staurosporine
表 1 菌株HNM0301抗植物病原菌活性
Table 1 Antifungal activity of strain HNM0301
病原真菌
Pathogenic fungi真菌抑菌率/%
(Antifungal rate/%)从赤壳菌
(Nectria citri)67 豇豆枯萎病菌
(Fusarium oxysporum
f. sp. Tracheiphlium)86 番茄早疫病菌
(Alternaria solani)71 火龙果黑腐病菌
(Bipolaris cactivora)71 火龙果溃疡病菌
(Neoscytaldium dimidiatum)— 菠萝黑腐病菌
(Ceratocystis paradoxa)— 小麦赤霉病菌
(Fusarium graminearum)— 香蕉枯萎病菌
(Fusarium oxysporum
f. sp.cubense tropical race 4)86 注:“—”为没有抑菌效果。
Note: “—” means no antifungal effect. 下载: 导出CSV
下载: 导出CSV
表 2 菌株 HNM0301 与最相近种之间的 ANI 值和 dDDH 值
Table 2 Values of ANI and dDDH between HNM0301 and its closest species
模式菌株
(Type strain)HNM0301 ANI dDDH Streptomyces sanyensis JCM 18324 (BAABJV010000001)96.83 72.4 Streptomyces zhihengii YIM T102(JAFEJA010000003) 82.55 26.3 Streptomyces genisteinicus CRPJ-33(CP060825) 82.54 26.3 Streptomyces xantholiticus JCM 4863 (BMWB01000001)80.86 24.7 Streptomyces gobitricini JCM 5062 (BAAASR010000001)80.16 23.9 Streptomyces lavendofoliae JCM 4391 (BMTP01000001)80.08 23.7 Streptomyces purpureus KA281(KB913030) 80.02 23.7 Streptomyces lavendulocolor JCM 4390 (BAAASF010000001)79.99 23.8 Streptomyces bikiniensis NRRL ISP- 5580 (JNXJ01000001)79.92 23.9 Streptomyces roseolilacinus JCM 4335 (BMSV01000001)79.81 23.7 Streptomyces vietnamensis GIM4.0001(CP010407) 79.69 23.7 Streptomyces crystallinus JCM 5067 (BAAACA010000001)79.55 23.3 Streptomyces roseolus JCM 4411 (BMTV01000001)79.44 23.6 Streptomyces tanashiensis Kala(CP084204) 79.42 23.5 Streptomyces noboritoensis JCM 4557 (JBHMQV010000001)79.37 23.4 Streptomyces nymphaeiformis SFB5A(JACHJY010000001) 79.31 23.5 Streptomyces nitrosporeus ATCC 12769 (CP023702)79.21 23 Streptomyces melanogenes JCM 4398 (BMTS01000001)79.16 23.2 Streptomyces laurentii NPDC006966(JBEYHT010000001) 79.15 23.2 Streptomyces peucetius NRRL WC- 3868 (JOCK01000001)77.87 22.6
下载: 导出CSV
表 3 菌株HNM0301 的基因组特性
Table 3 Genome features of HNM0301
特征 (Feature) 染色体信息 (Chromosome Characteristics) 染色体长度/bp(Chromosome size/bp) 6 635 340 GC含量/%(GC content/%) 73.71 蛋白质编码基因(Protein-coding genes) 5859 基因平均长度/bp(Gene average length/bp) 959.97 具有COG注释的基因(Genes assigned to COG) 4 297 具有KEGG注释的基因(Genes assigned to KEGG) 1 243 具有GO注释的基因(Genes assigned to GO) 2 137 核糖体RNA基因(rRNA genes) 18 转运RNA基因(tRNA genes) 64 微小RNA基因(sRNA genes) 51
下载: 导出CSV
表 4 菌株HNM0301预测的生物合成基因簇
Table 4 Predicted biosynthetic gene cluster of HNM0301
基因编号
(Cluster)类型
(Type)位置
(Position)已知基因相似度
(Most similar known biosynthetic gene cluster)1 非NRPS依赖型铁载体
(NRPS-independent-siderophore)215 382~227 026 聚克菌素
(Peucechelin)(25%)2 Ⅳ型羊毛硫肽
(lanthipeptide-class-Ⅳ)235 608~258 280 委内瑞拉菌素
(venezuelin)(100%)3 类NRPS-NRPS-萜杂合途径
(NRPS-like,NRPS,terpene)392 445~468 881 异海绵烯
(isorenieratene)(75%)4 非核糖体肽合成酶
(NRPS)497 124~543 572 环吡双环烯/氮杂双环烯B-D
(clipibicyclene/azabicyclene B/
azabicyclene C/azabicyclene D)(22%)5 萜类(terpene) 569 082~594 976 何帕烯(hopene)(69%) 6 Ⅰ型聚酮合酶
(T1PKS)955 184~1 004 505 福克辛A-C
(foxicin A/foxicin B/foxicin C/foxicin)(12%)7 Ⅱ型聚酮合酶(T2PKS) 1 136 46~1 269 167 Sch- 47554 /Sch-47555 )(21%)8 氨基多羧酸
(aminopolycarboxylic-acid)1 330 766~1 346 736 乙二胺羟基乙酸
(EDHA)(33%)9 非NRPS依赖型铁载体
NRPS-independent-siderophore1 432 445~1 445 628 - 10 其他
(other)1 820 060~1 861 973 A- 503083 A/A-503083 B/A-503083 E/A-503083 F
(3%)11 β-内酯类(betalactone) 2 198 156~2 224 871 萘霉素A(naphthomycin A)(6%) 12 黑色素(melanin) 2 293 489~2 300 967 黑色素(melanin)(100%) 13 Ⅲ型羊毛硫肽
(lanthipeptide-class-Ⅲ)2 776 024~2 798 540 - 14 类NRPS-NRPS-类PKS杂合途径
(NRPS-like,NRPS,PKS-like)2 819 955~2 882 442 里莫霉素A-C
(rimomycin A/rimomycin B/rimomycin C)(13%)15 transAT型PKS-NRPS杂合途径
(transAT-PKS,NRPS)3 244 144~3 320 517 拉吉霉素38/39
(largimiycin 38/largimycin 39)(51%)16 吲哚类(indole) 3 947 666~3 971 034 星孢菌素(staurosporine)(93%) 17 萜类(terpene) 4 344 692~4 365 549 波卓霉素A2(bottromycin A2)(24%) 18 非NRPS依赖型铁载体
(NRPS-independent-siderophore)4 404 710~4 416 482 去铁胺B
(desferrioxamin B)(100%)19 萜类(terpene) 4 506 981~4 524 545 - 20 类NRPS-NRPS-萜杂合途径
(NRPS-like,NRPS,betalactone)4 648 053~4 695 032 阿米霉素
(amicetin)(8%)21 线性肽类
(linaridin)4 719 408~4 739 986 链脲佐菌素
(streptozotocin)(15%)22 四氢嘧啶类
(ectoine)5 247 149~5 257 547 四氢嘧啶
(ectoine)(100%)23 羊毛硫肽/噻唑肽/RRE结构域相关肽
(LAP,thiopeptide,RRE-containing)5 803 975~5 838 819 - 24 萜类(terpene) 5 853 508~5 870 658 埃贝内酯(ebelactone)(5%) 25 丁酸内酯类(butyrolactone) 6 133 180~6 144 139 内酯霉素(lactonamycin)(3%) 26 萜类(terpene) 6 284 445~6 306 643 土臭素(geosmin)(100%) 27 黑色素(melanin) 6 391 082~6 401 489 黑色素(melanin)(42%)
下载: 导出CSV
表 5 HNM0301的Cluster 16与staurosporine合成基因簇比对
Table 5 Synthetic gene cluster alignment of Cluster 16 and staurosporine
基因ID
(Gene ID)氨基酸数
(Number of amino acids)同源基因ID
(Similar gene ID)氨基酸数
(Number of amino acids)同源基因一致性/%
(Positives/%)ctg1_ 3466 545 staC 535 87 ctg1_ 3467 281 staMB 280 87 ctg1_ 3468 208 staE 206 93 ctg1_ 3469 369 staI 369 91 ctg1_ 3470 331 staK 328 84 ctg1_ 3471 477 staJ 472 85 ctg1_ 3472 277 staMA 276 76 ctg1_ 3473 422 staP 417 77 ctg1_ 3474 1122 staD 1096 76 ctg1_ 3475 504 staO 503 88 ctg1_ 3476 436 staG 446 88 ctg1_ 3477 414 staN 394 87 ctg1_ 3478 350 staA 354 92 ctg1_ 3479 350 staB 334 85 — — staR 943 — 注:“—”为缺失该基因。 Note:“—” means deletion of the gene.
下载: 导出CSV
-
[1] ZHANG W L, BAI T T, JAMIL A, et al. The interaction between fusarium oxysporum f. sp. cubense tropical race 4 and soil properties in banana plantations in Southwest China[J]. Plant and Soil, 2024, 505(1/2): 779 − 793. doi: 10.1007/s11104-024-06709-4 [2] 黄英兰, 周维, 陈家慧, 等. T系列香蕉抗枯萎病品系在广西的农艺性状及产量初步比较[J]. 中国南方果树, 2024, 53(6): 138 − 142. doi: 10.13938/j.issn.1007-1431.20230543 [3] 辛侃, 赵娜, 邓小垦, 等. 香蕉-水稻轮作联合添加有机物料防控香蕉枯萎病研究[J]. 植物保护, 2014, 40(6): 36 − 41. doi: 10.3969/j.issn.0529-1542.2014.06.007 [4] 吴元立, 杨乔松, 李春雨, 等. 香蕉-尖孢镰刀菌互作机理及抗病育种研究进展[J]. 广东农业科学, 2020, 47(11): 32 − 41. doi: 10.16768/j.issn.1004-874X.2020.11.004 [5] 张妙宜, 周登博, 起登凤, 等. 香蕉枯萎病综合防控研究进展[J]. 中国科学: 生命科学, 2024, 54(10): 1843 − 1852. doi: 10.1360/SSV-2023-0283 [6] 甘林, 代玉立, 刘晓菲, 等. 香蕉枯萎病高效拮抗土著细菌的筛选及其防效[J]. 西北农林科技大学学报(自然科学版), 2024, 52(6): 95 − 105. doi: 10.13207/j.cnki.jnwafu.2024.06.010 [7] 何章飞, 李朝生, 于琴芝, 等. 不同作物与香蕉轮作综合防控枯萎病关键技术[J]. 广西农学报, 2022, 37(6): 21 − 26. doi: 10.3969/j.issn.1003-4374.2022.06.006 [8] RAMAKRISHNAN S, SREENIVAS S S. Biological control of soil-borne fungal and root-knot nematode disease complex in FCV tobacco nursery[J]. Journal of Biological Control, 2015, 29(4): 203 − 206. doi: 10.18641/jbc/29/4/86887 [9] 漆艳香, 谢艺贤, 彭军, 等. 香蕉根际微生态及其与枯萎病防治之间的关系[J]. 生物技术通报, 2024, 40(6): 57 − 67. doi: 10.13560/j.cnki.biotech.bull.1985.2023-1222 [10] 姚遐俊, 谢津, 祁艳华, 等. 一株防治香蕉枯萎病的短密木霉筛选及代谢物木霉素作用评价[J]. 生物工程学报, 2024, 40(1): 211 − 225. doi: 10.13345/j.cjb.230170 [11] 徐杰, 农婉晴, 张耀芳, 等. Bacillus rugosus对香蕉枯萎病的防治及促生效果[J/OL]. 分子植物育种, 2023: 1−19[2025-02-27]. https://link.cnki.net/urlid/46.1068.s.20231025.1308.006. [12] 黄穗萍, 李其利, 韦绍龙, 等. 米修链霉菌TF78对香蕉枯萎病的田间防效及根际土壤微生物的影响[J]. 微生物学通报, 2022, 49(9): 3693 − 3708. doi: 10.13344/j.microbiol.china.220095 [13] 宋秀丽, 赵崇钊, 卓愉林, 等. 绿针假单胞菌桔黄亚种在香蕉枯萎病防治中的应用[J]. 农业资源与环境学报, 2025, 42(1): 206 − 217. doi: 10.13254/j.jare.2023.0339 [14] 朱杰, 郑利乐, 钟鸣鸾, 等. 洋葱伯克霍尔德菌GD1-1的分离鉴定及对香蕉枯萎病的生防潜力[J]. 华南农业大学学报, 2025, 46(1): 72 − 80. doi: 10.7671/j.issn.1001-411X.202404043 [15] CHEN Y F, LI X J, ZHOU D B, et al. Streptomyces-secreted fluvirucin B6 as a potential bio-fungicide for managing banana Fusarium wilt and mycotoxins and modulating the soil microbial community structure[J]. Journal of Agricultural and Food Chemistry, 2024, 72(32): 17890 − 17902. doi: 10.1021/acs.jafc.4c04077 [16] SI SAID Z B O, HADDADI-GUEMGHAR H, BOULEKBACHE-MAKHLOUF L, et al. Essential oils composition, antibacterial and antioxidant activities of hydrodistillated extract of Eucalyptus globulus fruits[J]. Industrial Crops and Products, 2016, 89: 167 − 175. doi: 10.1016/j.indcrop.2016.05.018 [17] THOMPSON C C, CHIMETTO L, EDWARDS R A, et al. Microbial genomic taxonomy[J]. BMC Genomics, 2013, 14: 913. doi: 10.1186/1471-2164-14-913 [18] PARK H J, LEE J Y, HWANG I S, et al. Isolation and antifungal and antioomycete activities of Staurosporine from Streptomyces roseoflavus strain LS-A24[J]. Journal of Agricultural and Food Chemistry, 2006, 54(8): 3041 − 3046. doi: 10.1021/jf0532617 [19] OMURA S, IWAI Y, HIRANO A, et al. A new alkaloid AM-2282 of Streptomyces origin taxonomy, fermentation, isolation and preliminary characterization[J]. The Journal of Antibiotics, 1977, 30(4): 275 − 282. doi: 10.7164/antibiotics.30.275 [20] ZOU N X, ZHOU D B, CHEN Y L, et al. A novel antifungal actinomycete Streptomyces sp. strain H3-2 effectively controls banana Fusarium wilt[J]. Frontiers in Microbiology, 2021, 12: 706647. doi: 10.3389/fmicb.2021.706647 [21] 洪葵. 红树林放线菌及其天然产物研究进展[J]. 微生物学报, 2013, 53(11): 1131 − 1141. [22] 黄媛林, 潘信利, 陆璐, 等. 一株红树林链霉菌所产抑菌活性化合物的分离及其生物合成基因簇的研究[J]. 中国抗生素杂志, 2021, 46(12): 1090 − 1101. doi: 10.3969/j.issn.1001-8689.2021.12.004 [23] 张骏梁, 史蔷, 吴昊, 等. 一株红树林来源放线菌的抗菌活性物质[J]. 深圳大学学报(理工版), 2024, 41(3): 358 − 366. doi: 10.3724/SP.J.1249.2024.03358 [24] SUI J L, XU X X, QU Z, et al. Streptomyces sanyensis sp. nov. , isolated from mangrove sediment[J]. International Journal of Systematic and Evolutionary Microbiology, 2011, 61(7): 1632-1637. doi: 10.1099/ijs.0.023515-0 [25] FU P, YANG C L, WANG Y, et al. Streptocarbazoles A and B, two novel indolocarbazoles from the marine-derived actinomycete strain Streptomyces sp. FMA[J]. Organic Letters, 2012, 14(9): 2422 − 2425. doi: 10.1021/ol3008638 [26] LI T, DU Y Y, CUI Q, et al. Cloning, characterization and heterologous expression of the indolocarbazole biosynthetic gene cluster from marine-derived Streptomyces sanyensis FMA[J]. Marine Drugs, 2013, 11(2): 466 − 488. doi: 10.3390/md11020466 [27] CARTUCHE L, REYES-BATLLE M, SIFAOUI I, et al. Antiamoebic activities of indolocarbazole metabolites isolated from Streptomyces sanyensis cultures[J]. Marine Drugs, 2019, 17(10): 588. doi: 10.3390/md17100588 [28] CHEN Y F, WEI Y Z, CAI B Y, et al. Discovery of Niphimycin C from Streptomyces yongxingensis sp. nov. as a promising agrochemical fungicide for controlling banana Fusarium wilt by destroying the mitochondrial structure and function[J]. Journal of Agricultural and Food Chemistry, 2022, 70(40): 12784 − 12795. doi: 10.1021/acs.jafc.2c02810 [29] REN Z, LI Q, SHEN Y W, et al. Intrinsic relative preference profile of pan-kinase inhibitor drug staurosporine towards the clinically occurring gatekeeper mutations in Protein Tyrosine Kinases[J]. Computational Biology and Chemistry, 2021, 94: 107562. doi: 10.1016/j.compbiolchem.2021.107562 -

 点击查看大图
点击查看大图
计量
- 文章访问数: 568
- HTML全文浏览量: 343
- 被引次数: 0



