


-
内源性醛分子是一些生物合成与代谢途径的中间产物,这些途径包括碳水化合物、维生素、类固醇、氨基酸和脂类的代谢过程[1]。植物在逆境胁迫下,会产生醛类物质,将对植物产生毒害作用。醛脱氢酶(ALDHs)是NAD(P)+依赖性酶,其可以将有毒醛类物质氧化成羧酸,并降低脂质过氧化,提高植物对逆境的耐受性,这是植物体内重要的解毒机制[2]。据报道,在拟南芥中发现14个ALDH基因拷贝[3],拟南芥的ALDH3I1和ALDH7B4在提高植物耐受生物和非生物胁迫方面发挥着重要作用[4-5];拟南芥中ALDH3F1编码产生乙酸盐的醛氢酶,通过调节FLC位点上的H3K9Ac来控制开花时间,ALDH3F1突变导致开花时间早,ALDH3F1过表达导致开花晚[6];拟南芥中ALDH3家族的其他2个基因ALDH3I1与ALDH3H1在外源ABA、高盐、脱水和重金属等环境下诱导表达,证明其在氧化应激反应中发挥作用。ALDH3I1在叶片中特异表达,而在渗透压与外源ABA处理下,ALDH3H1在根中被诱导表达[3]。在水稻中共有19个ALDH基因拷贝[7],对水稻幼苗进行干旱、高盐及脱落酸处理,发现OsALDH2-2、OsALDH3-1和OsALDH3-2在干旱和高盐胁迫下,下调表达,而OsALDH2-4、OsALDH3-4、OsALDH7、OsALDH10-1和OsALDH18-2在干旱和高盐胁迫下诱导表达,OsALDH5和OsALDH12仅受干旱胁迫诱导表达,OsALDH18-1仅在高盐胁迫下上调表达;水稻胚乳中ALDH7功能的丧失将导致种子干燥和贮藏过程中的褐变,表明OsALDH7对种子采后保持活力至关重要。AKAMURA[8]发现,BADH基因编码的甜菜碱脱氢酶能催化甘氨酸合成甜菜碱(GB),甜菜碱作为一种无毒或保护性的细胞质渗透压调节剂,在叶片中积累,可以抵御非生物胁迫,从而增强植物的抗逆性。BRADBURY [9]研究结果显示,水稻8 号染色体上编码BADH基因与香味相关,水稻中常见的BADH突变类型包括badh2-E7[10]、badh2-E2[11]、bahd2-E4-5[12]等,该基因的突变导致催化GB合成的酶活性丧失,从而使得上游底物γ-氨基丁醛(GABald)自发环化形成 Δ1-吡咯啉,这是香味物质2-AP的关键前体[10]。此外,ALDH基因家族成员在果树中也被陆续鉴定,如在葡萄基因组中,鉴定出23个ALDH家族成员[13],在苹果中共鉴定39个ALDH基因拷贝[14],它们在生物和非生物胁迫下发挥重要作用。
椰子(Cocos nucifera L., 2n=32)属于棕榈科椰子属,单子叶多年生常绿乔木,是一种重要的热带水果和油料作物,具有极高的综合利用经济价值,被称为“宝树”[15]。椰子作为一种滨海经济作物,具有很好的逆境耐受性,我国培育的“文椰4号”椰子新品种,有一种特殊的芋头香味,深受消费者喜爱。ALDH基因家族在逆境调控以及香味调控中发挥重要作用,但在椰子中鲜有研究报道。本研究旨在鉴定椰子的ALDH基因家族,分析其基因结构、保守结构域、系统发育树及其表达模式,为椰子ALDH家族的基因功能阐释奠定基础。
-
在NCBI (National Center for Biotechnology Information) 网站上下载水稻ALDH基因家族的蛋白质序列,利用BLAST软件将水稻OsALDH基因家族蛋白序列与椰子CDS序列数据库进行比对,阈值E-value设置为1e−10,筛选椰子ALDH基因家族的CDS序列,共鉴定13个椰子CnALDH基因序列。应用TB tools软件将CDS序列翻译成蛋白质,在Pfma(
http://pfam.xfam.org/ search )数据库中进行结构域的预测,去除不含有ALDEH结构域的序列,最终获得椰子ALDH基因家族成员。 -
在ExPASy数据库(
https://web.expasy.org/protparam/ )中找到“ProtParam”选项,对椰子ALDH蛋白家族的氨基酸大小、分子质量、等电点基本信息进行分析,并通过WOLFPSORT网站对ALDH基因家族成员进行亚细胞定位信息的预测。 -
利用TB tools绘制椰子ALDH基因拷贝的结构图。使用MEGA X软件将椰子ALDH基因家族的蛋白序列与水稻OsALDH基因家族序列进行多序列比对,并用Neighbor-Joining方法构建进化树,Bootstrap设置为 1 000 次。
-
通过GeneDoc软件对椰子ALDH基因家族的蛋白质序列进行多序列分析,再利用 MEME(
http://meme-suite.org/tools/meme )在线工具对比得到的椰子蛋白序列与水稻OsALDH基因家族序列进行 motif 搜索,将预测的motif数值设定为10。在Weblogo3(http://weblogo.threeplusone.com/ )对所得的多序列分析结果绘制相应保守域结构的logo。在SWISS-MODEL上对CnAMADH2基因进行蛋白质三级结构同源建模。 -
在NCBI (National Center for Biotechnology Information) 网站上下载水稻和拟南芥BADH基因的蛋白质序列,利用BLAST软件将2个物种的BADH基因的蛋白序列与椰子CDS序列数据库进行比对,比对得出的CnAMADH2基因全长为16 359 bp,有14个外显子。针对该基因的第14个外显子设计引物,在香水椰子、高种椰子中扩增该基因的第14外显子并测序,在群体中验证是否存在碱基突变。
-
在 NCBI (National Center for Biotechnology Information)数据库的SRA (Sequence Read Archive)数据库下载不同品种椰子组织(SRR1063404、SRR1063407、SRR1125016、SRR-1173229、SRR1273180)与椰子胚(SRR1273070)、胚乳(SRR1265939)及胚愈伤组织(SRR1137438)的转录组原始数据,所得结果用TB Tools进行聚类并绘制成热图。
-
通过椰子ALDH基因家族鉴定,获得12个ALDH基因家族成员。根据AGNC注释标准,氨基酸序列与先前鉴定的ALDH序列相同超过40%的ALDH组成1个家族,而序列相同超过60%的ALDH组成1个亚家族,将12个ALDH基因家族成员分为9个家族。其蛋白质理化性质及亚细胞定位如下(表1)。
表 1 CnALDH基因家族理化性质分析
基因名称 氨基酸数量/个 分子量/Du 等电点 亚细胞定位 nucl mito Chol mito cysk chlo plas golg cyto CnALDH2_B4 535 58 046.51 6.1 11.5 7.5 2.5 CnALDH2_B7 543 58 891.36 6.03 11 3 CnALDH2_C4 500 54 724.76 5.95 14 1 1 CnALDH3_F1 814 91 359.59 8.68 2 6 CnALDH3_H1 495 54 206.91 8.59 1 4 1 1 7 CnALDH5_F1 499 53 897.15 8.30 4 10 CnALDH6_B2 537 57 488.84 7.13 11 3 CnALDH7_B4 509 54 587.40 6.57 2 1 2 2 2 CnALDH10_A8 503 54 919.05 5.35 1.5 2 1.5 CnALDH11_A3 496 53 273.64 6.76 6 6 2 CnALDH12_A1 555 61 854.02 6.07 1 7.5 5 4.5 CnALDH22_A1 590 65 312.80 7.87 2 2 结果表明,CnALDH基因家族氨基酸的分子量在53 273.64 ~ 91 359.59 Du之间。等电点在5.35~8.68之间,其中CnALDH2_B4、CnALDH2_B7、CnALDH2_C4、CnALDH7_B4、CnALDH10_A8、CnALDH11_A3、CnALDH12_A1为酸性蛋白质,其他CnALDH为碱性蛋白质。CnALDH基因家族编码氨基酸的数量在495~814之间,大多数基因家族成员编码的氨基酸数量差别不大,其中CnALDH3_F1编码的氨基酸数量最长,为814个氨基酸。CnALDH3_F1与拟南芥ALDH3_F1同源性较高,而拟南芥ALDH3_F1调节植物开花时间,并不被外界环境胁迫所诱导,CnALDH基因家族成员大多数分布在叶绿体中。
-
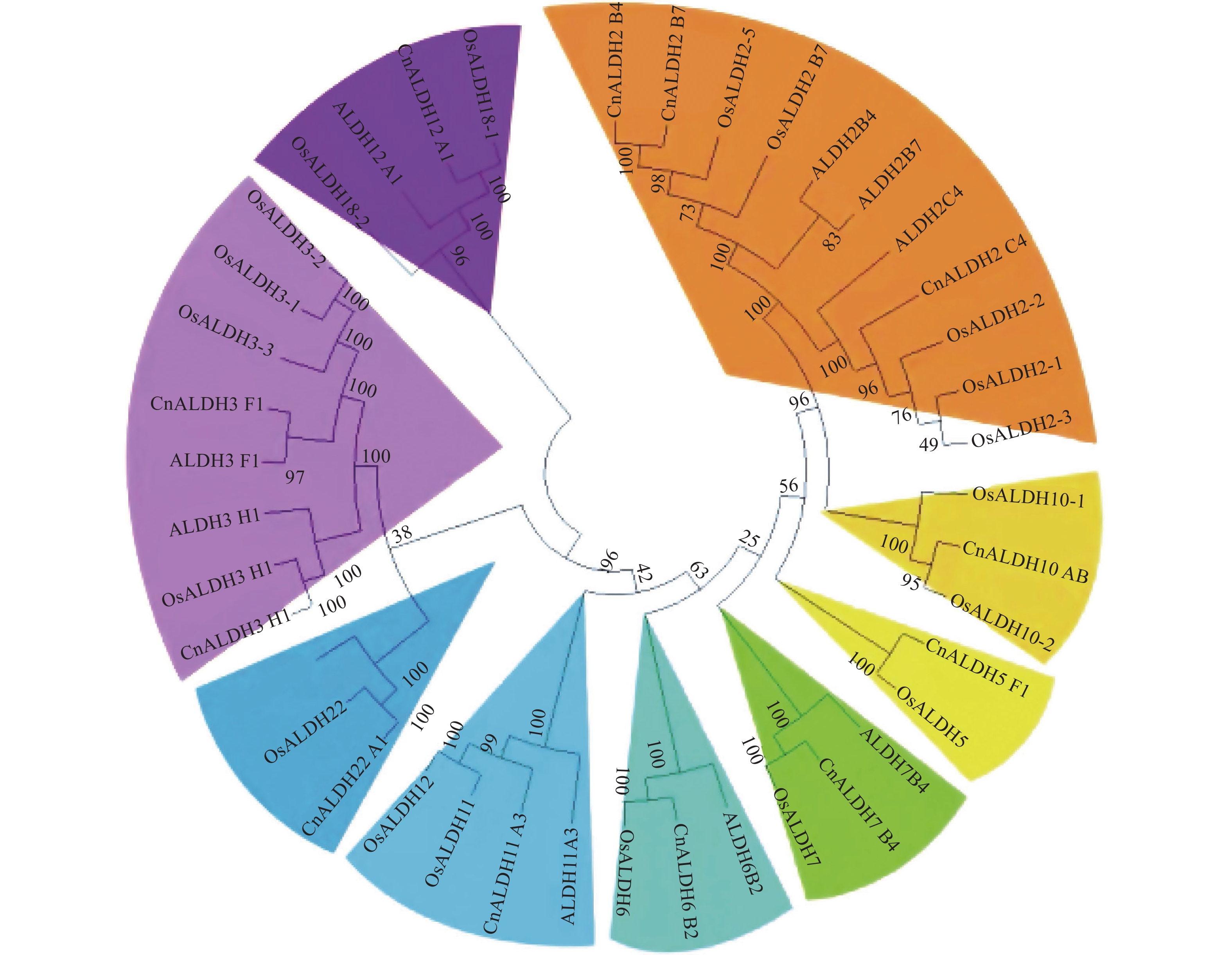
将椰子CnALDH基因家族的外显子和内含子相关数据导入TB Tools。随后使用MEGA X软件将椰子、水稻的ALDH基因进行多序列比对,使用邻接法构建系统进化树(图1),椰子ALDH基因家族结构域水稻和拟南芥的结构相似。此外还对椰子ALDH基因家族的外显子-内含子结构进行了研究。
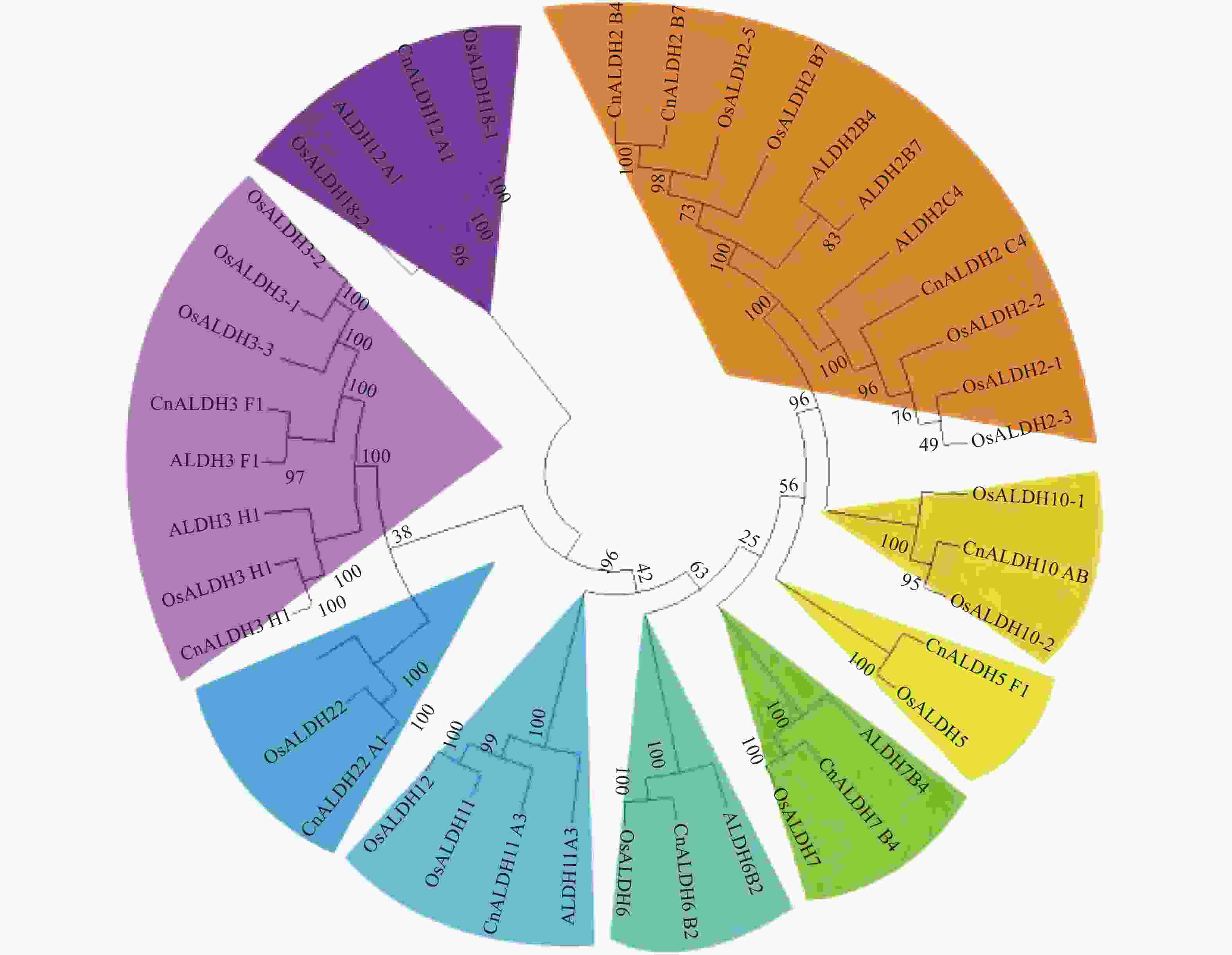
图 1 椰子、拟南芥和水稻ALDH基因家族的系统发育树
基因结构分析显示,椰子ALDH家族外显子个数变异为9~17。家族内外显子-内含子结构差异在多基因家族的进化中起关键作用。结合系统进化树结果分析发现,亲缘关系相近的基因具有相似的基因结构,如CnALDH2-B4与CnALDH2_B7均含有11个外显子;不同基因家族成员基因结构则相差较大(图2)。椰子基因组中共鉴定出12个基因家族成员,分属于9个不同的亚族。
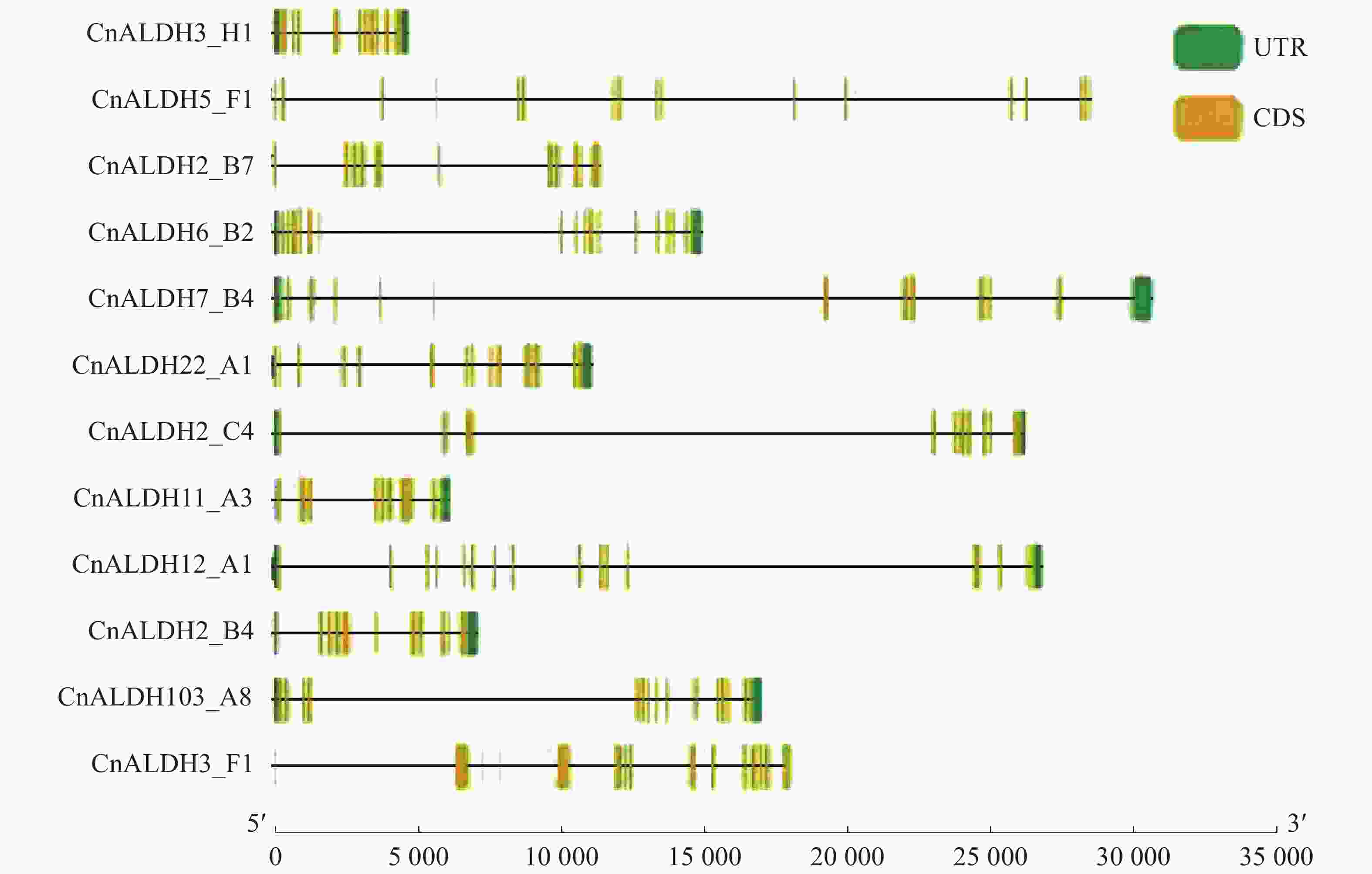
图 2 椰子CnALDH基因家族结构
-
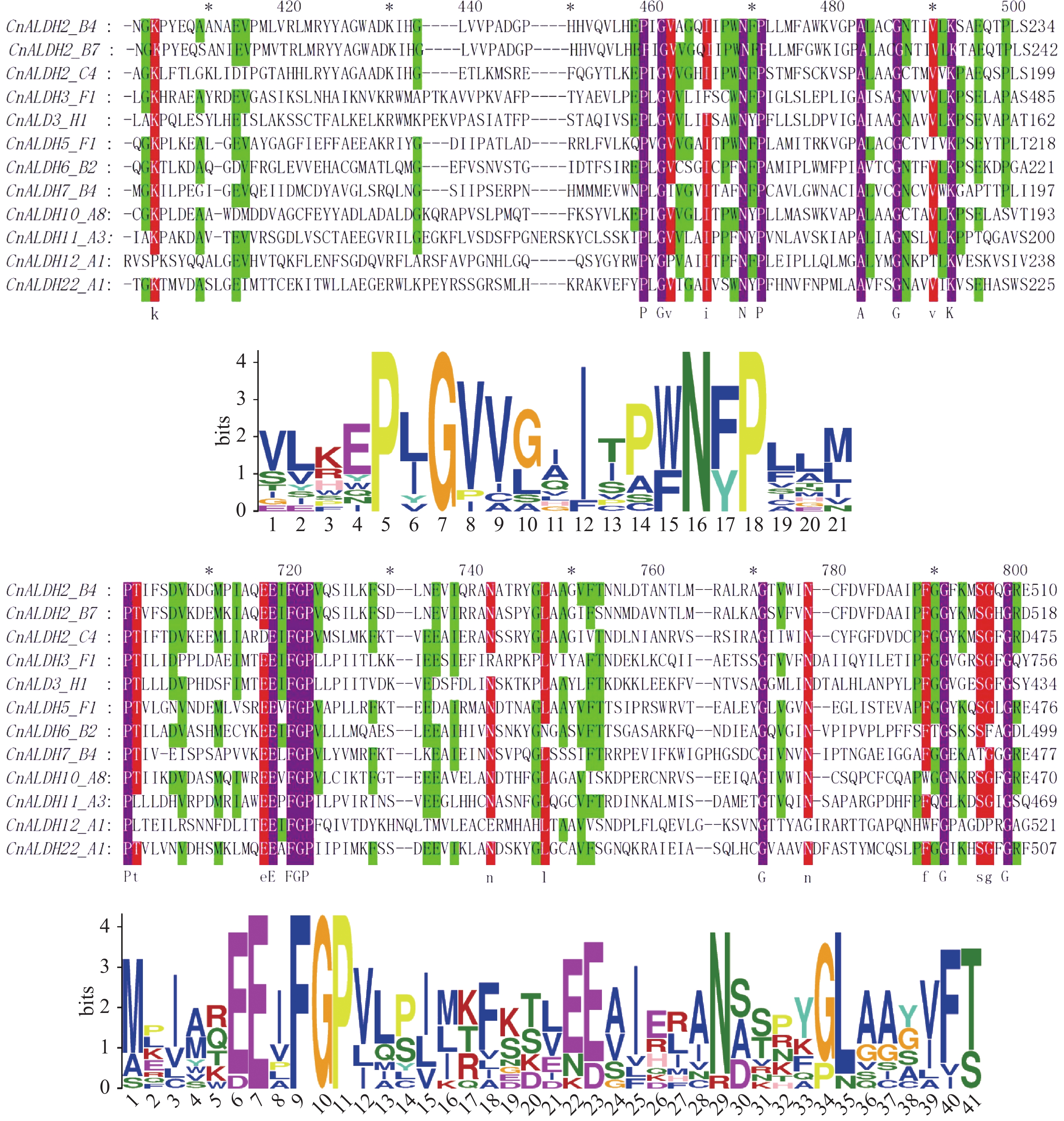
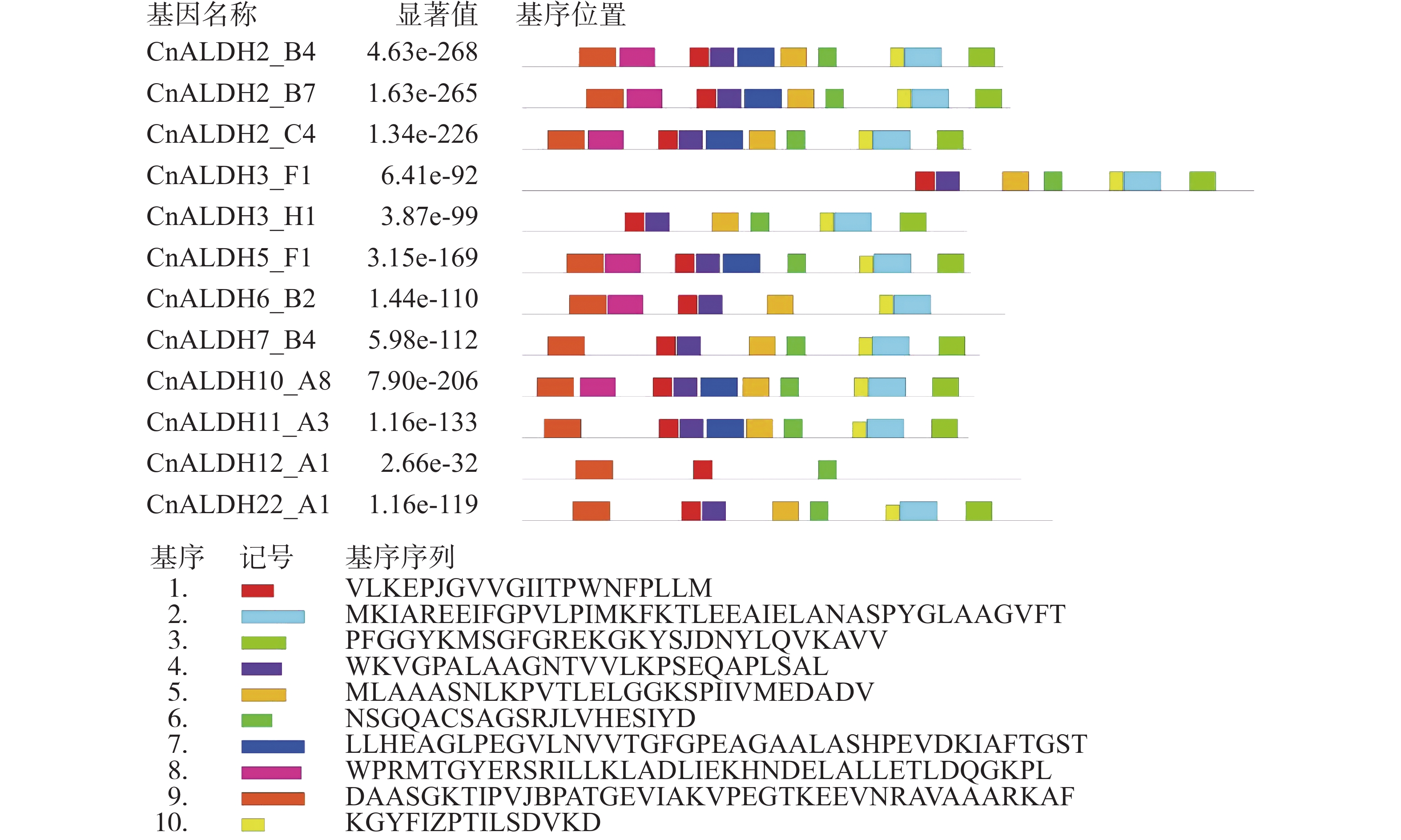
采用MEME软件,分析了ALDH蛋白质的保守结构域(图3)。共搜索到10个motif(图4),其中最保守的结构域为motif 1。CnALDH2_B4、CnALDH2_B7、CnALDH2_C4均属于同一个亚族,他们的motif构成一致。CnALDH3_F1与CnALDH3_H1均不含有motif 8和motif 9。与其他ALDH基因家族motif构成相比,CnALDH12_A1 motif构成差别较大,根据水稻中对该亚家族的研究结果,推测椰子中这两个基因家族成员可能行使不同的功能。

图 3 椰子CnALDH基因家族保守域多序列比对

图 4 椰子CnALDH基因家族 motif 分析
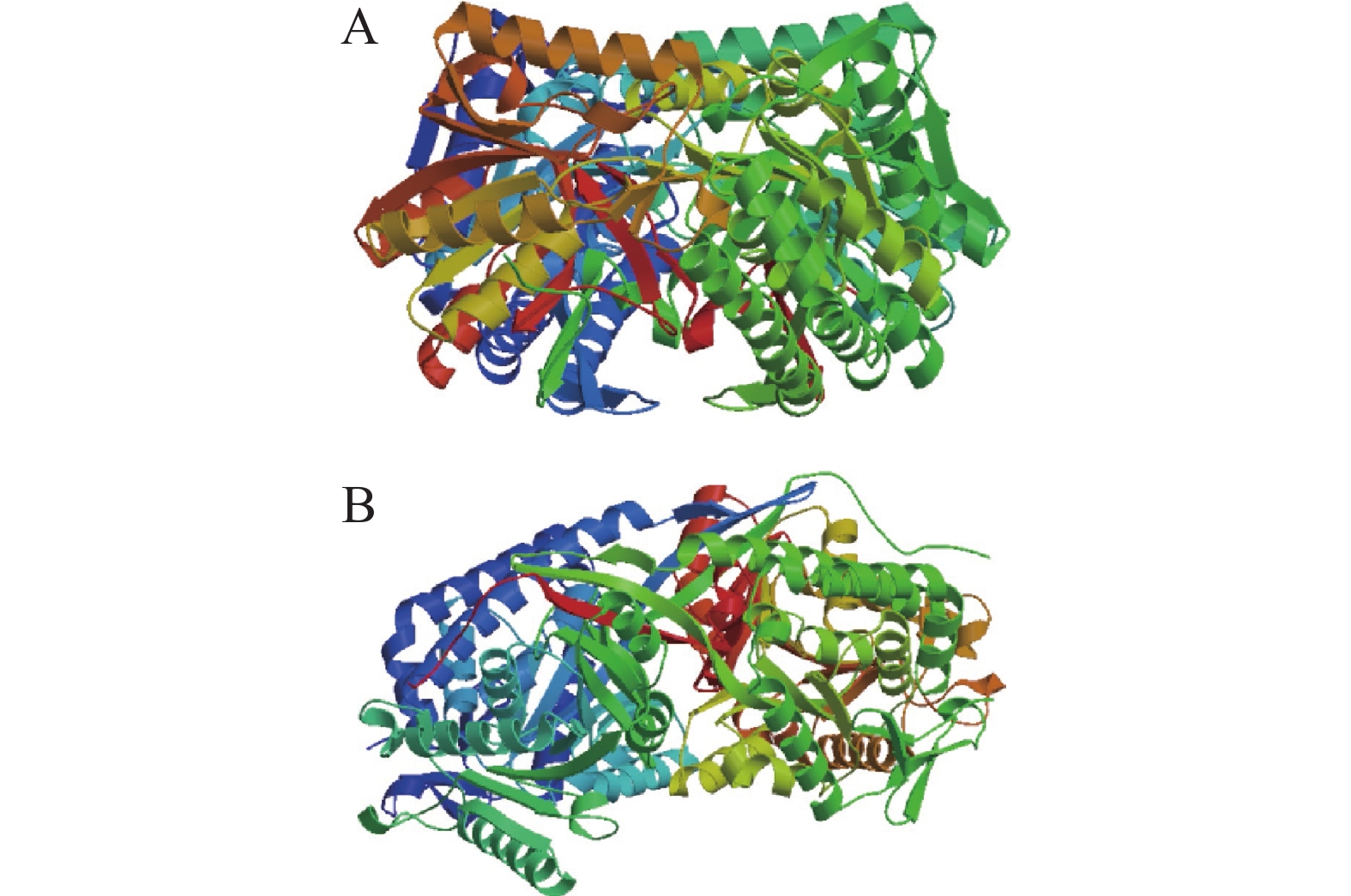
本研究利用 SWISS-MODEL 在线对椰子CnAMADH2基因以及该基因的变异序列进行三维结构同源建模。图5-A为非香型椰子CnAMADH2基因的蛋白质结构,图5-B为香水椰子中CnAMADH2基因的蛋白质结构。该基因第14外显子上的G/C突变导致蛋白质结构发生变化,引起CnAMADH2基因的功能丧失。

图 5 椰子 CnAMADH2基因三维结构模型
-
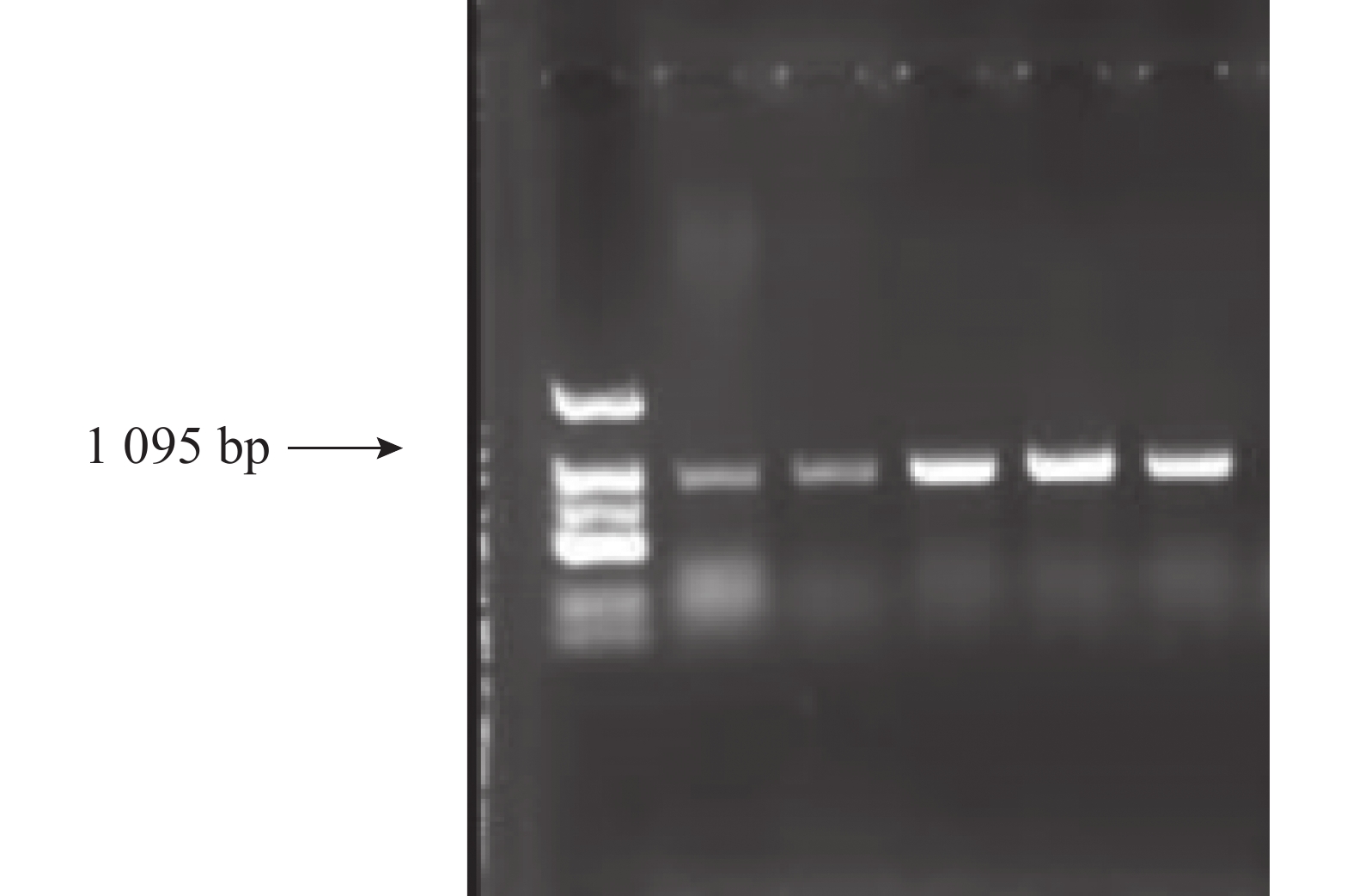
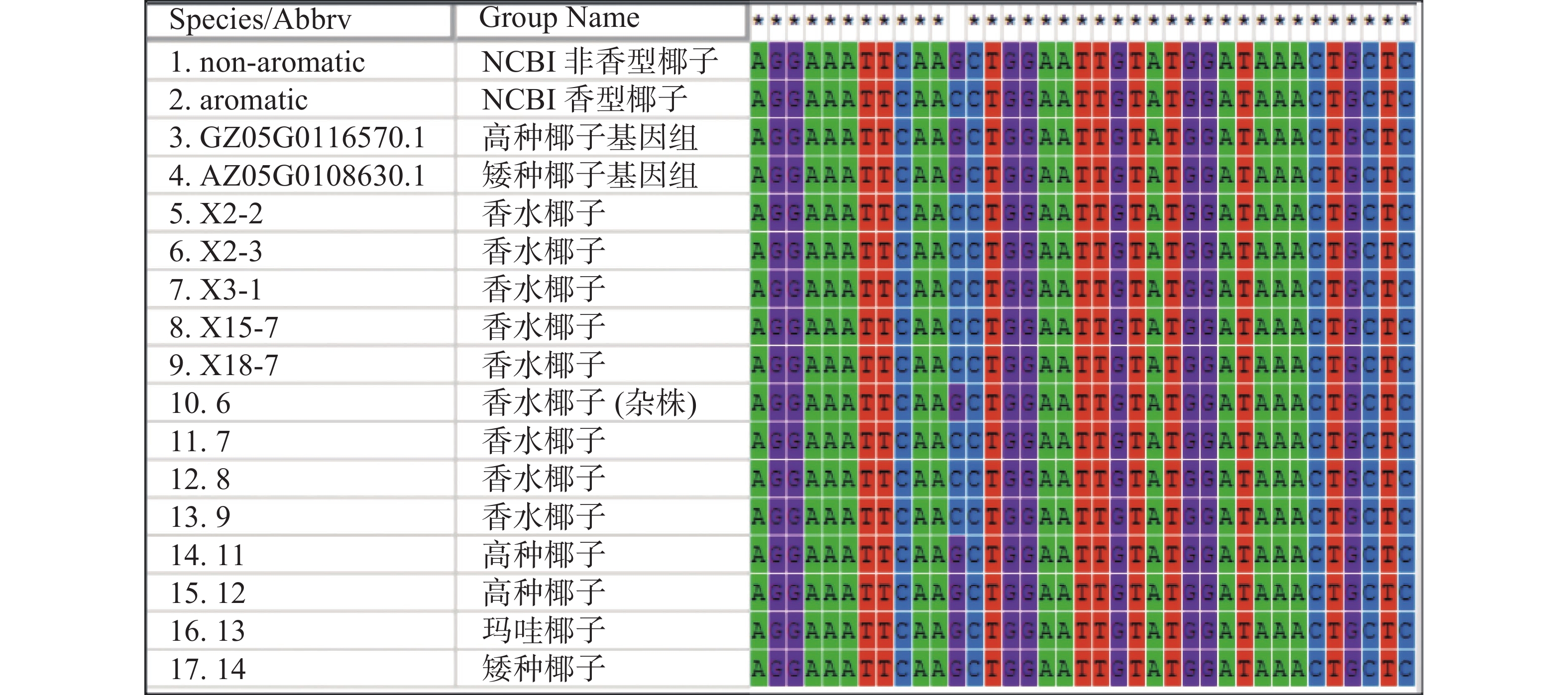
根据目的基因序列设计引物,左引物(CnA-exon11-F:5′AGTATGAGAAGATCAAAAAGTTCA3′)设计在第11外显子上,右引物(CnA-exon14-R:5′CCATTCCCCAAGTTCACG3′)设计在第14外显子,全长为1 095 bp(图6)。取13个样品的新鲜嫩叶提取基因组DNA,扩增后得到的测序结果如图7所示,香型椰子基因型为CC纯合,非香型椰子基因型为GG纯合,而样品6号为GC杂合香水椰,该测序结果与NCBI测定结果一致。
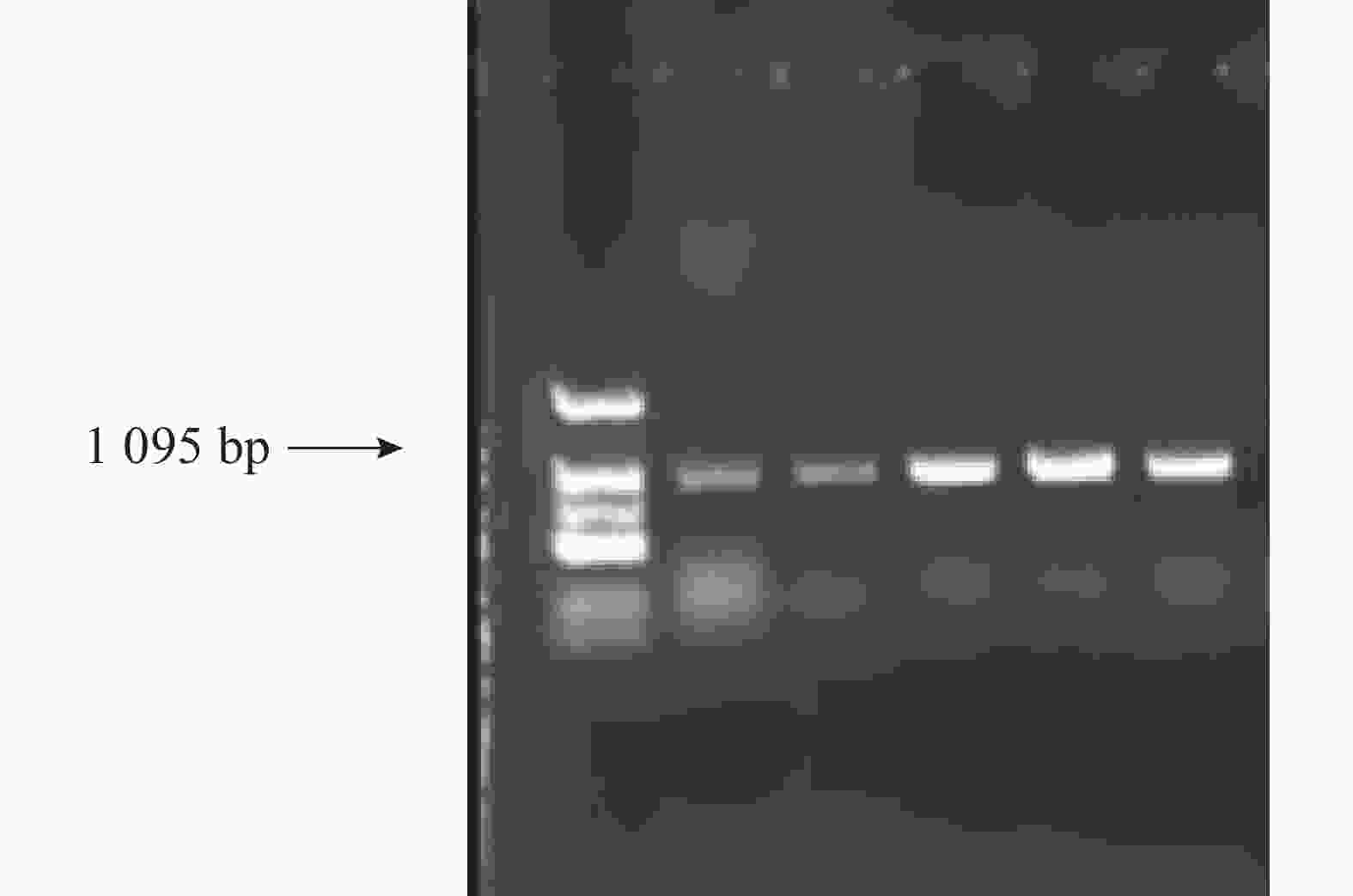
图 6 椰子 CnAMADH2第14外显子扩增结果
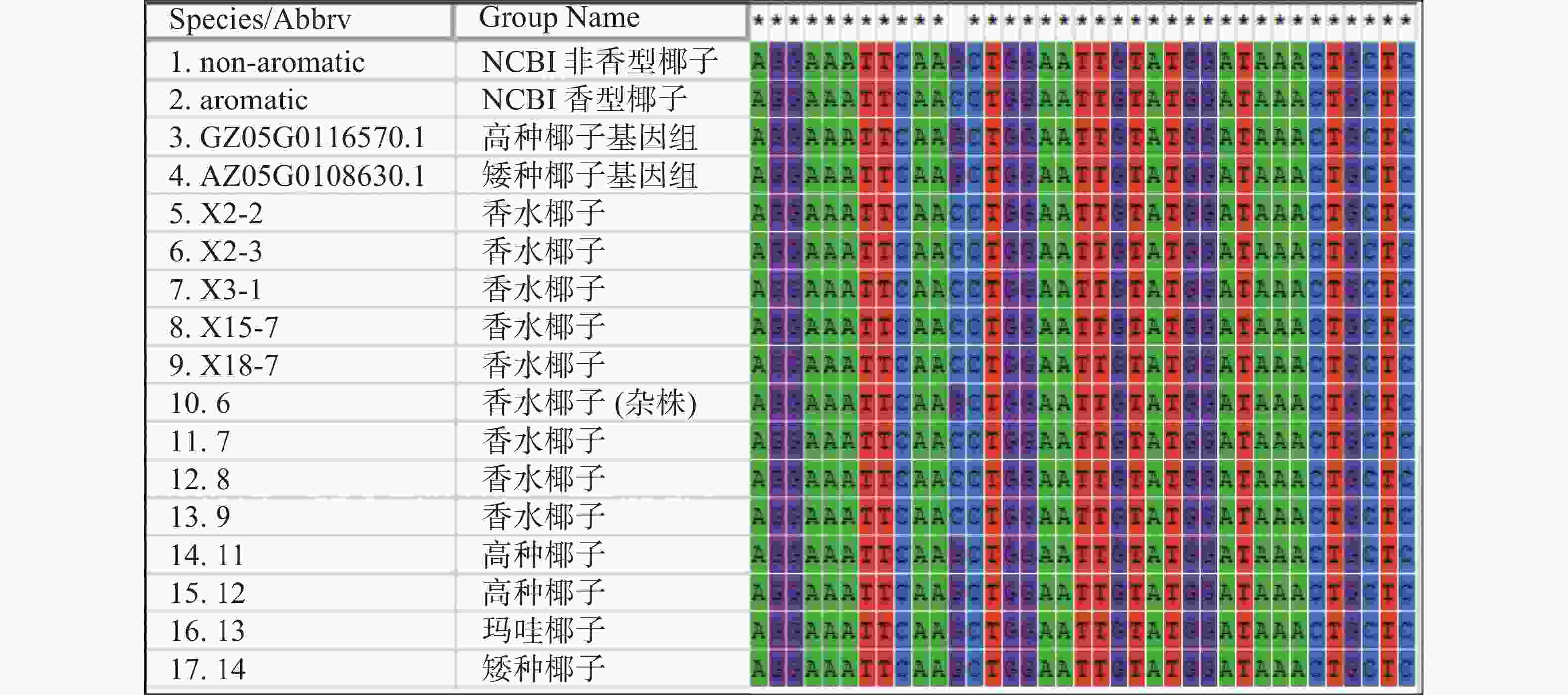
图 7 椰子 CnAMADH2第14外显子测序结果
-
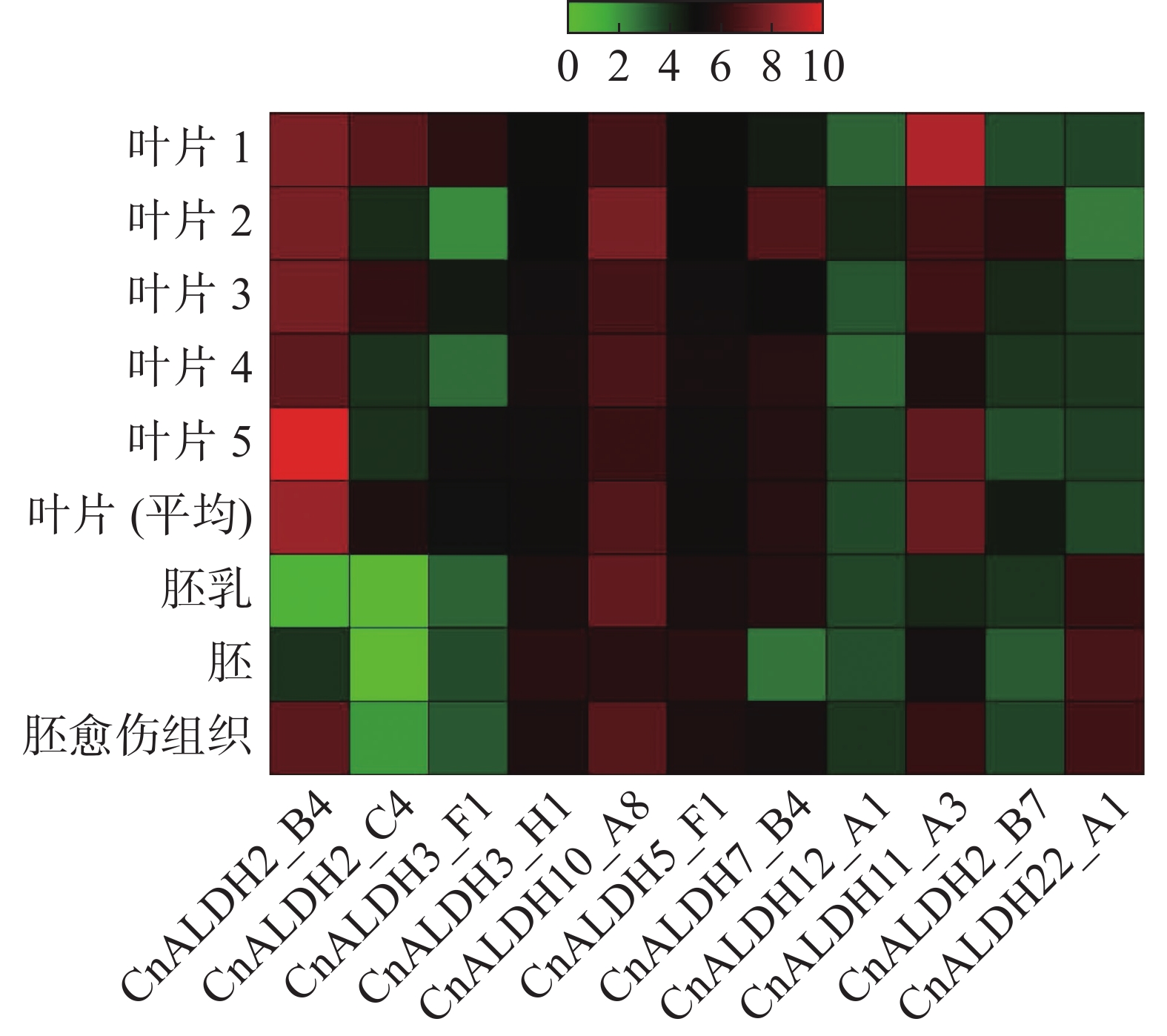
在 NCBI 数据库中下载不同品种椰子叶片、椰子胚、胚乳及胚愈伤组织的转录组数据。通过计算RPKM值,得到该家族成员在不同组织中的表达量,采用TBtools绘制热图(图8)。除CnALDH12_A1和CnALDH22_A1外,其他的ALDH基因家族成员均在叶片中有高水平表达。CnALDH22_A1在胚、胚乳与胚愈伤组织中表达。根据椰子与水稻进化树分析得出CnALDH12_A1与水稻OSALDH18家族的同源性较高,而水稻OsALDH18编码P5CS,该物质可以合成脯氨酸抵抗渗透胁迫,但在椰子中几乎不表达。CnALDH10在各个时期均有高水平的表达,说明其在椰子整个生长周期应对环境胁迫,这可能与椰子本身较强的抗逆性有关。CnALDH2_B4、CnALDH2_B7、CnALDH2_C4均属于椰子CnALDH2家族,但在椰子不同生长时期发挥作用,CnALDH2_C4仅在叶片中表达。CnALDH7_B4在愈伤组织、胚乳及叶片中表达量较高。
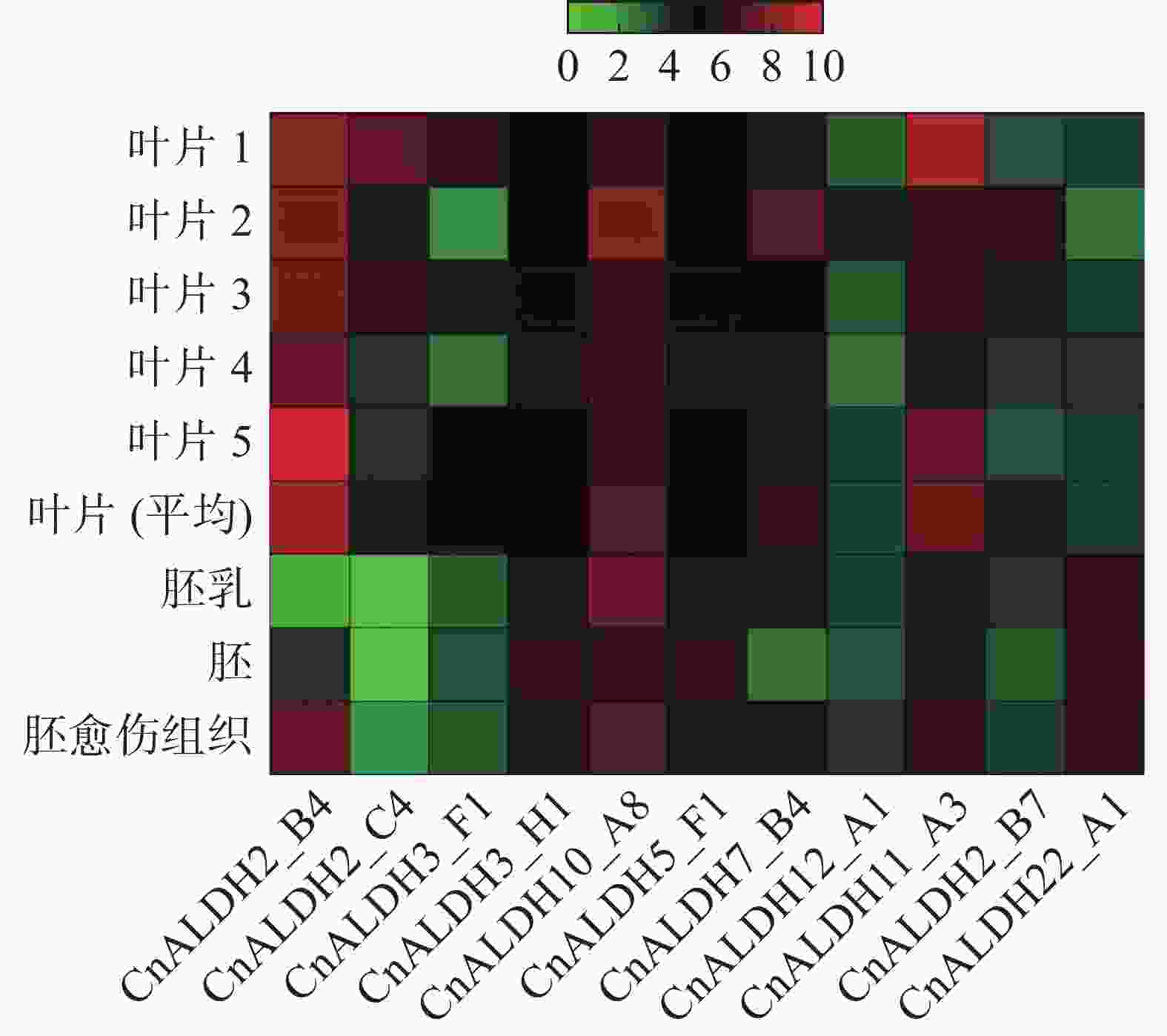
图 8 椰子CnALDH基因家族在不同组织器官中的表达谱分析
-
本研究通过生物信息学方法,鉴定椰子ALDH基因家族成员。研究结果表明,水稻基因组包括19个ALDH基因家族成员,而椰子中仅鉴定出12个。分析椰子ALDH基因家族的蛋白理化性质发现。除CnALDH3_F1,其他基因编码的氨基酸数量相差不大。对椰子motif分析发现,所有椰子ALDH基因家族成员均含有motif1。
研究椰子ALDH基因家族在叶片、胚、胚乳、胚愈伤组织表达模式发现,CnALDH10_A8在植物各个时期均有表达。ALDH10家族的作用是催化甜菜醛脱氢为甘氨酸甜菜碱,甘氨酸甜菜碱作为一种渗透调节物质,可以帮助植物抵抗缺水、脱水的不良环境[8]。在大豆中,非生物胁迫使大豆中二胺氧化酶活性增加,迫使根系合成大量
$ \gamma $ -氨基丁酸[7]。对椰子抗逆性研究发现,高种椰子比香水椰子抵抗逆境的能力强。BRADBURY[10]发现香水椰子中编码BADH基因14外显子上一个碱基的缺失阻断了GB的合成,由此可以合理推测该基因的突变是导致香水椰子不耐逆境的其中一个原因。ALDH12和ALDH18家族均参与脯氨酸的代谢,该基因家族受干旱诱迫诱导,介导植物的应激反应和ROS的积累[16-17]。拟南芥中,外源脯氨酸和高盐诱导ALDH12基因的表达。CnALDH12_A1在椰子转录组样本中表达量较少,其表达量是否与脯氨酸代谢有关需要进一步的实验证明。ABA能诱导脱水耐性的获得[18]。KIRCH等[19]报道了拟南芥中ALDH7_B4基因的表达是由脱落酸激活的,它调节脱水应激反应和种子脱水通路。研究证明,OsALDH7是种子成熟的关键基因,胚乳中该基因的缺失导致种子在干燥和贮藏中发生褐变[20]。根据CnALDH7_B4的表达谱推测该基因在胚乳中的表达也与种子成熟有关。植物中与渗透胁迫诱导有关的ALDH基因家族为ALDH3、ALDH7、ALDH10、ALDH11和ALDH12中的大部分成员,ALDH3_H1在拟南芥的叶片中低水平表达[1],而ALDH3_F1的表达则与开花有关[6],ALDH11_A3催化植物特有的一种经典的糖酵解“旁路”反应[1]。此外,在果树中也陆续发现ALDH基因家族的存在。对葡萄ALDH基因家族的研究发现,该基因家族除了在干旱等非生物胁迫发生作用,霜霉病侵染时,部分抗病品种的ALDH基因家族成员表达量发生显著变化[13]。椰子中ALDH基因的表达是否也受病害的诱导还未知。
椰子基因组的完整序列极大地帮助了研究人员将椰子作为一种经济作物进行改良,为提高椰子的抗逆性以及筛选优良树种奠定了分子基础。椰子基因组包含12个ALDH超家族基因,编码了9个不同的蛋白质家族,其中大多数家族都与应激条件有关。目前,ALDH基因家族在拟南芥和水稻模式植物中研究较为透彻,在葡萄、苹果、香蕉等[14-15]果树的ALDH基因家族也被陆续研究,而在椰子中对ALDH基因家族的研究鲜见报道。分析该基因家族的序列信息、基因结构,与水稻[5]等作物系统发育关系及在椰子不同组织的表达模式,可预测或识别椰子中各个ALDH蛋白或蛋白质家族发挥作用的途径,并为进一步研究ALDH基因家族的生物学功能奠定基础。
椰子ALDH基因家族的鉴定及生物信息学分析
DOI: 10.15886/j.cnki.rdswxb.2022.06.001
 CSTR: 32425.14.j.cnki.rdswxb.2022.06.001
CSTR: 32425.14.j.cnki.rdswxb.2022.06.001
Identification and bioinformatics analysis of ALDH gene family in coconut
-
摘要: 为了鉴定椰子的ALDH基因家族,分析其基因结构、保守结构域、系统发育树以及其表达模式,通过下载OsALDH基因家族的蛋白质序列,并与椰子的蛋白质的数据库进行比对,鉴定出12个CnALDH基因,分属于9个亚族。结果表明,ALDH编码的氨基酸等电点为5.35~8.68,多数分布在叶绿体中。表达谱分析结果显示,除CnALDH12_A1和CnALDH22_A1外,其他的CnALDH基因家族成员均在叶片中有高水平表达。而CnALDH10_A8在不同组织以及不同时期展现组成性表达,推测该基因参与椰子的一些基础性功能,是不可或缺的基因。Abstract: Aldehyde dehydrogenase (ALDH), which dehydrogenates aldehydes into carboxylic acids, plays an important role in plant stress, development and fragrance synthesis. The protein sequences of OsALDH gene were downloaded and compared with the protein database of coconut, and 12 CnALDH genes were identified, which belonged to 9 subfamilies. The results showed that the isoelectric points of amino acids encoded by ALDH ranged from 5.35 to 8.68, and most of them were distributed in chloroplasts. Expression profile analysis showed that all ALDH gene family members were highly expressed in coconut young leaves except CnALDH12_A1 and CnALDH22_A1. CnALDH10_A8 showed constitutive expression in different tissues and at different periods, suggesting that CnALDH10_A8 is an indispensable gene involved in some basic functions in coconut.
-
Key words:
- coconut /
- ALDH gene family /
- bioinformatics analysis
-
表 1 CnALDH基因家族理化性质分析
基因名称 氨基酸数量/个 分子量/Du 等电点 亚细胞定位 nucl mito Chol mito cysk chlo plas golg cyto CnALDH2_B4 535 58 046.51 6.1 11.5 7.5 2.5 CnALDH2_B7 543 58 891.36 6.03 11 3 CnALDH2_C4 500 54 724.76 5.95 14 1 1 CnALDH3_F1 814 91 359.59 8.68 2 6 CnALDH3_H1 495 54 206.91 8.59 1 4 1 1 7 CnALDH5_F1 499 53 897.15 8.30 4 10 CnALDH6_B2 537 57 488.84 7.13 11 3 CnALDH7_B4 509 54 587.40 6.57 2 1 2 2 2 CnALDH10_A8 503 54 919.05 5.35 1.5 2 1.5 CnALDH11_A3 496 53 273.64 6.76 6 6 2 CnALDH12_A1 555 61 854.02 6.07 1 7.5 5 4.5 CnALDH22_A1 590 65 312.80 7.87 2 2  下载: 导出CSV
下载: 导出CSV
-
[1] HANS-HUBERT K, DOROTHEA B. The ALDH gene superfamily of Arabidopsis [J]. Trends in Plant Science, 2004, 6(9): 371 − 377. [2] KOTCHONI S O, KUHNS C, KIRCH H H, et al. Overexpression of different aldehyde dehydrogenase genes in Arabidopsis thaliana confers tolerance to abiotic stress and protects plants against lipid peroxidation and oxidative stress [J]. Plant Cell Environ, 2006, 29: 1033 − 1048. doi: 10.1111/j.1365-3040.2005.01458.x [3] SUNKAR R, BARTELS D, KIRCH H H. Overexpression of a stress‐inducible aldehyde dehydrogenase gene from Arabidopsis thaliana in transgenic plants improves stress tolerance [J]. Plant, 2003, 35: 452 − 464. [4] KIM JM, TO TK, MATSUI A, et al. Acetate‐mediated novel survival strategy against drought in plants [J]. Nature Plants, 2017, 3: 17097. doi: 10.1038/nplants.2017.97 [5] GAO C X, HAN B. Evolutionary and expression study of the aldehyde dehydrogenase (ALDH) gene superfamily in rice (Oryza sativa) [J]. Gene, 2009, 431: 86 − 94. doi: 10.1016/j.gene.2008.11.010 [6] XU D Y, LIU Q, CHEN G, et al. Aldehyde dehydrogenase ALDH3F1 involvement in flowering time regulation through histone acetylation modulation on flowering locus [J]. Journal of Integrative Plant Biology, 2019, 62: 1080 − 1092. [7] KISHITANI S, TAKANAMI T, SUZUKI. M, et al. Compatibility of glycinebetaine in rice plants: evaluation using transgenic rice plants with a gene for peroxisomalbetaine aldehyde dehydrogenase from barley [J]. Plant, Cell and Environment, 2000, 23: 107 − 114. doi: 10.1046/j.1365-3040.2000.00527.x [8] NAKAMURA. , YOKOTA S, MURAMOTO Y, et al. Expression of a betaine aldehyde dehydrogenase gene in rice, a glycinebetaine nonaccumulator, and possible localization of its protein in peroxisomes [J]. Plant Journal, 1997, 11: 1115 − 1120. doi: 10.1046/j.1365-313X.1997.11051115.x [9] BRADBURY L M T, FITZGERALD T. L, HENRY R J, et al. The gene for fragrance in rice [J]. Plant Biotechnol, 2005, 3: 363 − 370. doi: 10.1111/j.1467-7652.2005.00131.x [10] BRADBURY L M T, GILLIES S A, BRUSHETT D J, et al. Inactivation of an aminoaldehyde dehydrogenase is responsible for fragrance in rice [J]. Plant Mol Biol, 2008, 68: 439 − 449. doi: 10.1007/s11103-008-9381-x [11] SHI W W YANG Y, CHEN S H, et al. Discovery of a new fragrance allele and the development of functional markers for the breeding of fragrant rice varieties [J]. Molecular Breeding, 2008, 22: 185 − 192. doi: 10.1007/s11032-008-9165-7 [12] SHAO G N, TANG S Q, CHEN M L, et al. Haplotype variation at Badh2, the gee determining fragrance in rice [J]. Genomics, 2003, 101: 157 − 162. [13] ZHANG Y, MAO L, WANG H, et al. Genome-wide identification and analysis of grape aldehyde dehydrogenase (ALDH) gene superfamily [J]. PLoS One, 2012, 7: e32153. doi: 10.1371/journal.pone.0032153 [14] 张乐, 陈宣钦, 李昆志. 植物醛脱氢酶基因家族[J]. 生命的化学, 2013, 33(3): 299 − 306. [15] 刘莹. 椰子全身都是宝[J]. 解放军健康, 2002(4): 31 − 31. [16] DEUSCHLE K, FUNCK D, HELLMANN H, et al. A nuclear gene encoding mitochondrial delta-pyrroline-5-carboxylate dehydrogenase and its potential role in protection from proline toxicity [J]. The Plant Journal, 2001, 27: 345 − 356. [17] YOSHIBA Y, KIYOSUE T, NAKASHIMA K, et al. Regulation of levels of proline as an osmolyte in plants under water stress [J]. Plant and Cell Physiology, 1997, 38: 1095 − 1102. doi: 10.1093/oxfordjournals.pcp.a029093 [18] 宋松泉, 傅家瑞. 成熟脱水对种子发育和萌发的作用[J]. 植物学通报, 1998, 15(2): 23 − 32. [19] KIRCH H H, SCHLINGENSIEPEN S, KOTCHONI S, et al. Detailed expression analysis of selected genes of the aldehyde dehydrogenase (ALDH) gene superfamily in Arabidopsis thaliana [J]. Plant Mol Biol, 2005, 57: 315 − 332. doi: 10.1007/s11103-004-7796-6 [20] JUN-HYE SHIN, SUNG-RYUL KIM, GYNHEUNG AN. Rice ALDH7 Is needed for seed maturation and viability [J]. Plant Physiol, 2009, 149(2): 905 − 915. doi: 10.1104/pp.108.130716 -

 点击查看大图
点击查看大图
图(8) / 表 (1)
计量
- 文章访问数: 2337
- HTML全文浏览量: 624
- PDF下载量: 61
- 被引次数: 0

