

-
中国是全球最大的天然橡胶消费国和进口国。橡胶树(Hevea brasiliensis)几乎是商业天然橡胶的唯一来源,也是中国热带农业经济的支柱产业,但天然橡胶供应安全长期面临严峻挑战[1-3]。由胶孢炭疽菌(Colletotrichum gloeosporioides)引起的橡胶树炭疽病是导致橡胶产量与品质下降的主要病害之一[4-5]。目前,对橡胶炭疽病的防控仍依赖化学杀菌剂,如多菌灵、代森锰锌等[4-6]。此外,长期单一用药易使部分地区病原菌的抗药性上升[7],且过度使用化学农药还可能造成环境污染危及生态安全。因此,针对胶孢炭疽菌致病机制开发绿色、靶向性防控新策略,已成为发展橡胶产业的迫切需求。超长链脂肪酸(Very Long-Chain Fatty Acids, VLCFAs),一般指含有16个及以上碳原子数的脂肪酸[8-9]。作为细胞膜鞘脂和磷脂的重要组成部分,VLCFAs不仅是细胞生存的必需成分,还在维持膜动态平衡、诱导因VLCFAs积累而触发的细胞自噬,及提高生物对应激源的敏感性等方面发挥重要作用[10-12]。在植物病原菌的研究中发现,VLCFAs具有维持细胞膜稳定性的作用[13],并参与附着胞膨压形成及穿透能力的调控[14],因此VLCFAs被认为是调控植物病原真菌致病力的重要组分。超长链脂肪酸延伸酶(Very Long-Chain Fatty Acids Elongase, Elo)是催化VLCFAs合成的关键限速酶,在动、植物及微生物等生命体中均有表达。哺乳动物体内存在7种超长链脂肪酸延伸酶(常用ELOVL1-7表示)[15-17],并且各ELOVL的功能不完全相同。ELOVL1在免疫系统相关途径中发挥重要作用[18];ELOVL2与男性睾丸中超长链多不饱和脂肪酸含量相关,缺失ELOVL2可能导致不育[19];ELOVL4与视网膜功能关系密切[20-21],且缺失ELOVL4会干扰神经酰胺合成,使小鼠皮肤通透性出现异常,并在新生期死亡[22];ELOVL5参与调控肝脏中18个以上碳原子脂肪酸的延伸[23]。另有研究发现Elo与酵母维持细胞膜上脂肪酸的平衡有关,且Elo能够影响酵母中VLCFAs的积累、油酸诱导的脂毒性细胞死亡等[11]。在裂殖酵母(Schizosaccharomyces pombe)中Elo2定位于核膜上催化合成神经酰胺,能够防止因核膜破裂导致的致死现象,对维持细胞活性至关重要[24]。同时,在植物病原菌中Elo也发挥着重要作用,Elo2参与调控禾谷镰刀菌(Fusarium graminearum)的发育、对胁迫的响应和毒力发生[25];在稻瘟菌(Magnaporthe oryzae)中缺失Elo1能够阻断附着胞内Septin环组装和干扰附着胞发育进而影响对水稻的入侵[14]。这些报道为深入探究Elo在调控胶孢炭疽菌的VLCFAs合成及入侵等环节的分子机制奠定了理论基础。鉴于橡胶树表皮具有较厚的蜡质,胶孢炭疽菌的成功侵染与其附着胞的功能密切相关,而这一过程很有可能依赖于VLCFAs对细胞膜完整性与稳定性的维持作用[10]。因此,本研究聚焦于胶孢炭疽菌的CgElo3,通过构建基因敲除突变体,分析其对胶孢炭疽菌生理表型、致病力及超长链脂肪酸积累等的影响。本研究旨在填补胶孢炭疽菌中Elo调控VLCFAs合成研究的空缺,既为阐明橡胶树炭疽病的致病分子机制提供新视角,同时也为实现橡胶树炭疽病从化学防控向绿色精准防控的转型奠定坚实基础。
-
胶孢炭疽菌(C. gloeosporioides Wild Type,WT)由本实验室前期分离、鉴定所得,目前已完成该菌株的基因组测序、数据总结及建库。PCR引物由北京擎科生物公司与睿博兴科公司提供(表1)。DNA测序由北京擎科生物公司负责完成。中长链脂肪酸靶向代谢物(脂质)含量测定由上海派森诺生物科技股份有限公司完成。pBS-SUR由本实验室改造、保存。Spark HiFi Seamless Cloning Kit(单片段)和SPARKscript Ⅱ All-in-one RT SuperMix for qPCR(With gDNA Eraser)均购于山东思科捷公司。RNA prep Pure Plant Kit试剂盒购于北京天根生物技术有限公司。
表 1 实验用引物
Table 1. Primers used in the present experiment
引物名称 Primer 序列(5′—3′)Sequence 用途Usage CgElo3-5F GCGGCGTGCTTGACATAC 同源臂扩增Homologous arm amplification CgElo3-5R CTTTCCAAAGTCTGTGCC 同源臂扩增 Homologous arm amplification CgElo3-MF1 TTGGTCAATCCCCCACCGTGCCAACGCCACAGTG 同源臂扩增 Homologous arm amplification CgElo3-MR1 CACTGTGGCGTTGGCACGGTGGGGGATTGACCAA 同源臂扩增 Homologous arm amplification CgElo3-MF2 GAATTGCATGCTCTCACTTGTCTTGTGCGAAGTG 同源臂扩增 Homologous arm amplification CgElo3-MR2 CACTTCGCACAAGACAAGTGAGAGCATGCAATTC 同源臂扩增 Homologous arm amplification Sur-SLF CCTCTGATATTGGAAGCGACGC 同源臂扩增 Homologous arm amplification Sur-SLR ATGTTGGCATAAGCCGAACCGT 同源臂扩增 Homologous arm amplification Sur-JC5F GCGTTTGTAACTCTGCCTGTTTG 突变体鉴定 Mutant Strain Identification Sur-JC3R ACGAGGACCGCTACTCACATAC 突变体鉴定 Mutant Strain Identification CgElo3-JC5F CCCTTAGTTTGGACTGATGC 突变体鉴定 Mutant Strain Identification CgElo3-JC3R CGGGAACAAGGGAATAAAGAT 突变体鉴定 Mutant Strain Identification CgActin-qF GATTGGTATGGGCCAGAAGG 相对表达量检测 Relative expression level detection CgActin-qR GGAGCTCGTTGTAGAAGGTG 相对表达量检测 Relative expression level detection CgElo3-QF CTTCGCCTTTGTCGTTCTGC 相对表达量检测 Relative expression level detection CgElo3-QR TACCGTTGAGCTGGCAGAAG 相对表达量检测 Relative expression level detection 菌株培养所用培养基(以配制200 mL为例):PDA马铃薯葡萄糖固体培养基(12.5 g马铃薯葡萄糖琼脂粉末);CM完全培养基(2 g葡萄糖、5 mL硝酸盐溶液、0.2 mL维生素、0.2 mL微量元素、0.2 g酸水解酪蛋白、0.4 g蛋白胨、0.2 g酵母提取物);0.5% ME麦芽浸膏培养基(0.1 g麦芽提取物)、YCS培养基(64.4 g蔗糖、0.2 g酵母提取物、0.2 g酸水解酪蛋白)。
-
使用uniprot在线网站(
https://www.uniprot.org/ )查找不同真菌中Elo蛋白序列,利用MEGA6.0软件制作Elo同源蛋白进化树。 -
通过液氮研磨方法提取胶孢炭疽菌野生型在不同发育时期及胶孢炭疽菌野生型侵染橡胶树叶片不同时间段的总RNA,反转录总RNA可获得检测所需的cDNA,以此cDNA为模板,进行实时荧光定量RT-qPCR检测(以CgActin基因的表达量作为内参)。
-
本研究以同源重组原理为基础的Split-Marker基因敲除技术构建CgElo3基因缺失突变株。以WT基因组为模板扩增CgElo3基因上、下游同源片段并测序,再扩增由实验室保存的氯嘧磺隆(Sulfonylurea resistance gene,SUR)抗性基因,融合同源片段与抗性基因以获得敲除突变株构建的转化片段。将PDA平板上正常生长的WT菌株切成小块,在CM液体培养基中培养2 d(培养条件:28℃摇床,150 r·min−1),再取1 mL过滤后的菌液在ME液体培养基内培养1 d,培养条件同上。参考郭云峰等[26]的实验方法,过滤出菌丝置于蜗牛酶(Snailase)与纤维素酶R-10(Cellulase R10)的混合液体中酶解4 h以获得野生型原生质体,通过PEG介导将敲除所用的转化片段转入原生质体中。采用不含有筛选抗性的YCS培养基过夜培养,第二天再向该培养基内加入含有氯嘧磺隆SUR抗性的YCS培养基。将YCS培养基上长出的转化子转接到PDA平板上培养3 d,提取转化子基因组并检测CgElo3上、下游同源臂;选取检测结果为阳性的转化子在含有氯嘧磺隆SUR抗性的ME平板上的单胞,2 d后将纯合转化子转接到PDA平板上,3 d后提取纯合转化子基因组检测CgElo3上、下游同源臂和基因,将检测阳性的纯合转化子命名为ΔCgElo3开展后续实验。
-
使用打孔器(孔径:6.0 mm)取相同大小的WT和ΔCgElo3菌块,转接到CM平板上培养5 d(温度:28℃)。用十字交叉法测量菌落直径并拍照记录形态,统计分析WT和ΔCgElo3菌落生长速率。
-
将PDA平板上正常生长的WT菌株切成小块,在CM液体培养基中培养2 d(培养条件:28℃摇床,150 r·min−1),使用血球计数板统计孢子浓度,在无菌条件下过滤出孢子液并稀释至1×103个·mL−1,然后吸取1 mL孢子液在30 mL CM液体培养基中培养,培养条件同上。3 d后滤出全部菌液并记录孢子数量,统计分析WT和ΔCgElo3孢子产量。
-
按照1.4.2方法培养致病力所需的菌液,将过滤得到的孢子液以5 500 r·min−1转速离心1 min,使用0.5% ME液体培养基重悬并稀释至2.5×105个·mL−1。在橡胶树叶片合适位置上接种5 μL菌液,于28℃培养箱内培养3 d。用十字交叉法测量病斑直径大小并拍照记录,统计分析WT和ΔCgElo3菌株致病力。
-
将玻璃纸裁剪成圆形(直径约6 cm),灭菌后用无菌水浸湿并覆盖在PDA固体培养基表面,使用打孔器取相同大小菌块接种于表面覆有玻璃纸的PDA中心,于28℃培养箱内倒置培养3 d并拍照留存,3 d后揭下玻璃纸继续培养,3 d后拍照记录。
-
按照1.4.2方法培养孢子萌发所需的菌液。将过滤得到的孢子液以5 500 r·min−1转速离心1 min,使用2% YCS液体培养基将孢子液稀释至5×105个·mL−1,吸取20 μL置于干净透明的玻璃载玻片上,于28℃保湿条件下培养2、4 h,使用显微镜观察并拍照记录。
-
按照1.4.2方法获得离体诱导附着胞所需的菌液。将过滤得到的孢子液以5 500 r·min−1转速离心1 min,使用无菌水将孢子液充分洗净后再至3.5×105个·mL−1,吸取5 μL接种于疏水表面,在28℃保湿条件下连续培养12、24 h,使用显微镜观察并拍照记录。
-
按照1.6.2方法获得洋葱内表皮入侵菌丝观察所需的菌液。将洋葱内表皮疏水面向上置于1.5%水琼脂表面。吸取5 μL接种于洋葱内表皮疏水面,在28℃保湿条件下连续培养12、24 h,使用显微镜观察并拍照记录。。
-
将PDA平板上正常生长的WT菌株切成小块,在CM液体培养基中培养2 d(培养条件:28℃摇床,150 r·min−1),过滤得到孢子液,取1 mL孢子液于置于100 mL CM液体培养基中培养1 d,培养条件同上。过滤出菌丝经液氮冷冻后送至上海派森诺生物科技股份有限公司测定中长链脂肪酸靶向代谢物(脂质组)含量。
-
本研究实验均进行3次重复,使用ImageJ软件统计萌发、附着胞和入侵菌丝数量和裁剪原图,SPSS软件进行正态分布和方差齐性验证, GraphPad Prism软件分析数据和作图,采用Duncan多重比较检验组间差距,P<0.05时,认为存在显著性差异。
-
分析胶孢炭疽菌(C. gloeosporioides)基因组发现,除本实验室前期已鉴定到的2个编码Elo的基因(命名为CgElo1和CgElo2)还鉴定出一个注释为Elo的基因,将其命名为CgElo3。进化树分析结果(图1−A)显示,在酿酒酵母(Saccharomyces cerevisiae)、稻瘟菌(M. oryzae)、刺孢炭疽菌 (Colletotrichum. spinosum)、禾谷镰刀菌 (F. graminearum)、大丽轮枝菌(Verticillium dahliae)等植物病原菌中均存在CgElo3的同源蛋白,且同源性较高,说明CgElo3在真菌中有较高的保守性。本实验室前期的研究中发现,CgElo3在胶孢炭疽菌入侵阶段表达量显著升高(图1−B)。为验证CgElo3在不同发育和侵染时期的表达情况,本研究通过RT-qPCR检测了CgElo3在营养菌丝(Hypha,HY)、孢子(Spore)、萌发芽管(Germ Tube ,GT 3h)、附着胞(Appressorium,AP 16 h)及活体侵染(0~48 h)时间段的相对表达量。结果发现,与孢子时期的表达量相比,CgElo3在萌发、附着胞发育阶段的表达量显著上调(图1−C);与侵染橡胶树叶片0 h相比,CgElo3在侵染12 h时的表达量显著上调,然而在侵染24、48 h时的表达量显著下调(图1−D)。以上结果表明,CgElo3可能参与调控胶孢炭疽菌对橡胶树叶片入侵的早期阶段。
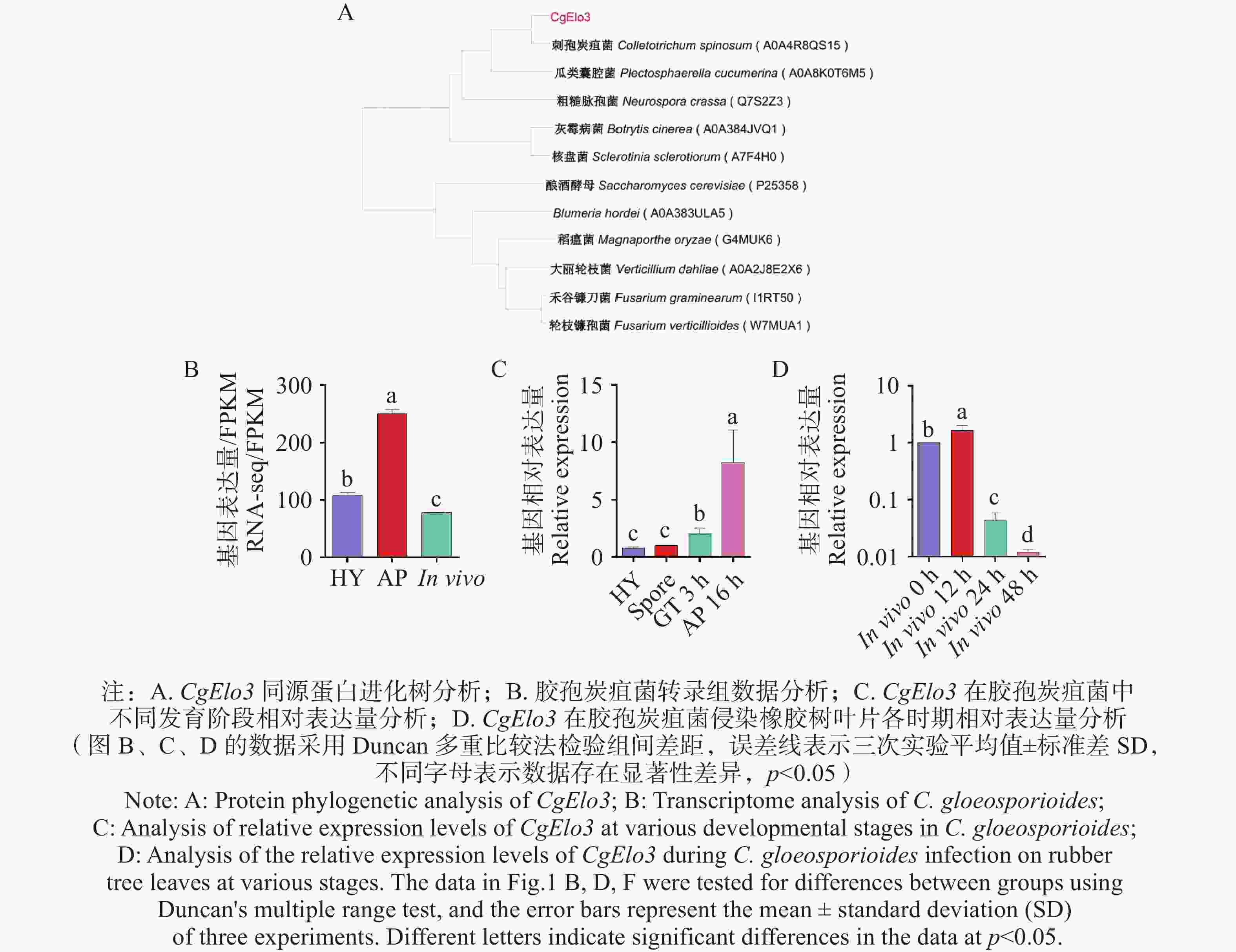
图 1 胶胞炭疽菌CgElo3蛋白进化树分析和CgElo3在不同阶段相对表达量分析
Figure 1. Phylogenetic analysis of CgElo3 proteins and relative expression analysis of CgElo3 at different stages
-
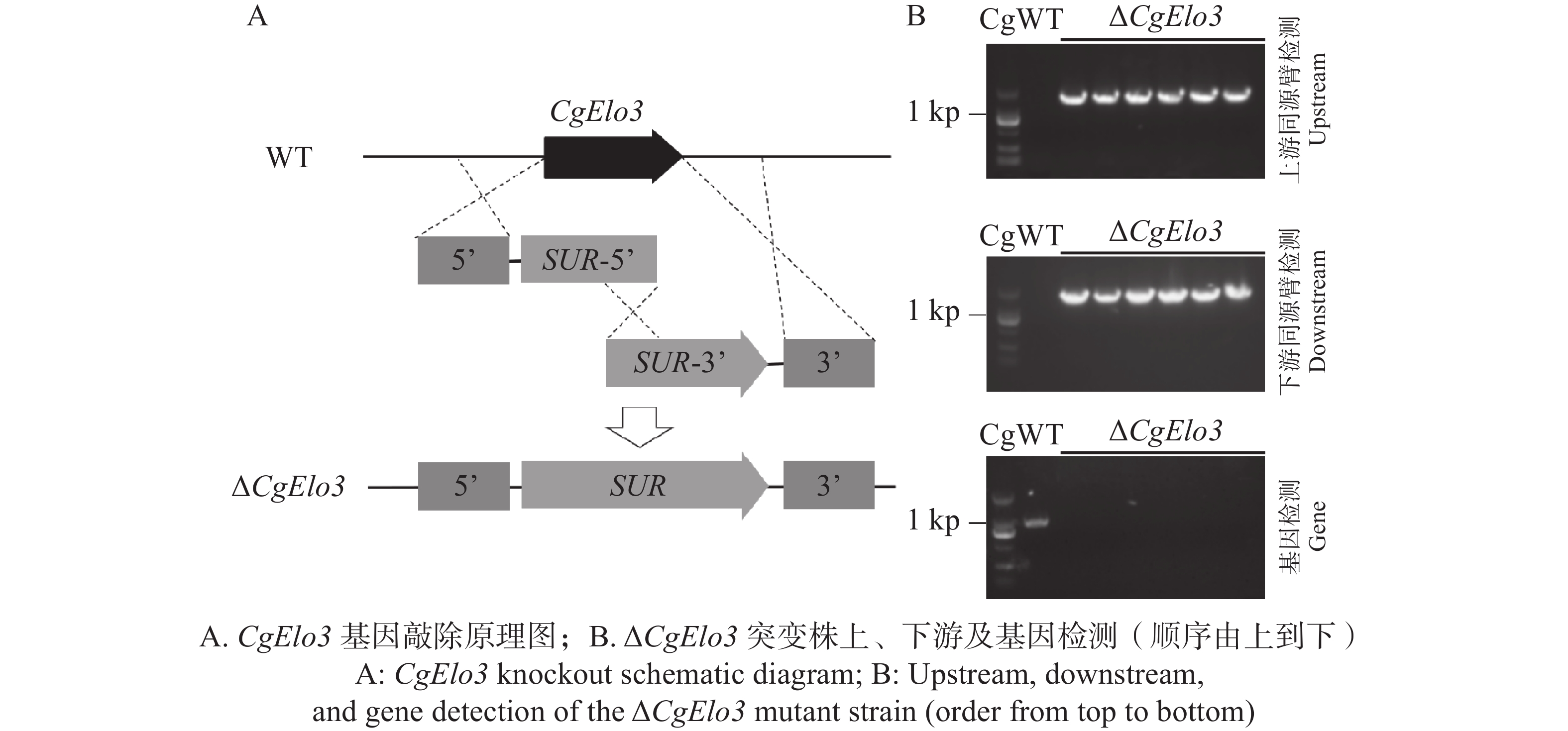
本研究通过Split-Marker同源重组的原理(图2−A)构建CgElo3基因敲除突变体。首先对筛选抗性培养基上所获得的转化子进行上、下游同源片段PCR检测;将检测结果为阳性的转化子单孢分离以获得纯合转化子;再进行上、下游同源臂段和基因PCR检测。ΔCgElo3突变株能扩增出单一的上、下同源臂并且无目的基因阳性条带(图2−B)。PCR检测结果表明成功构建ΔCgElo3敲除突变株。
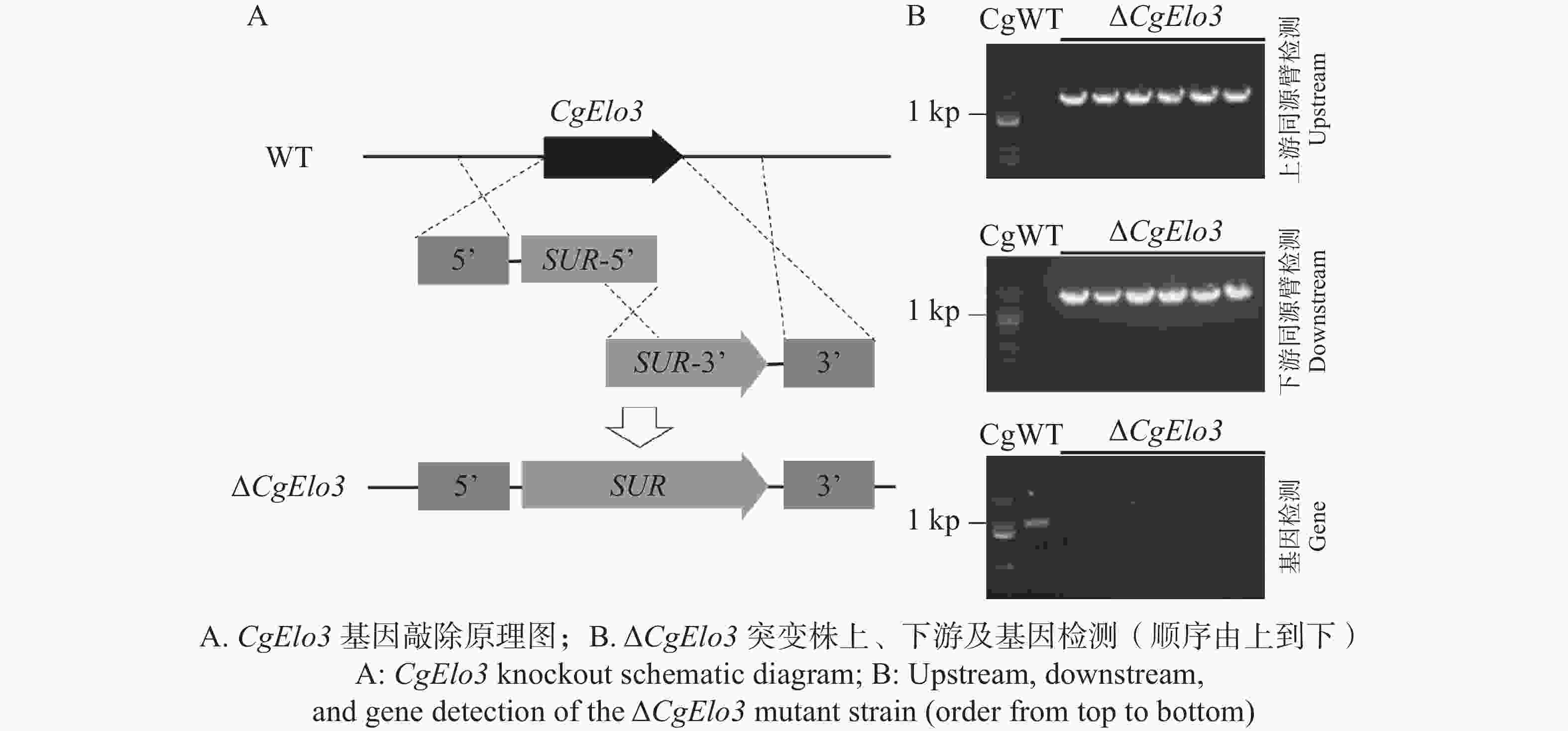
图 2 Split-Marker基因敲除原理及鉴定ΔCgElo3突变株
Figure 2. Principle of split-marker gene knockout and identification of ΔCgElo3 mutant strain
-
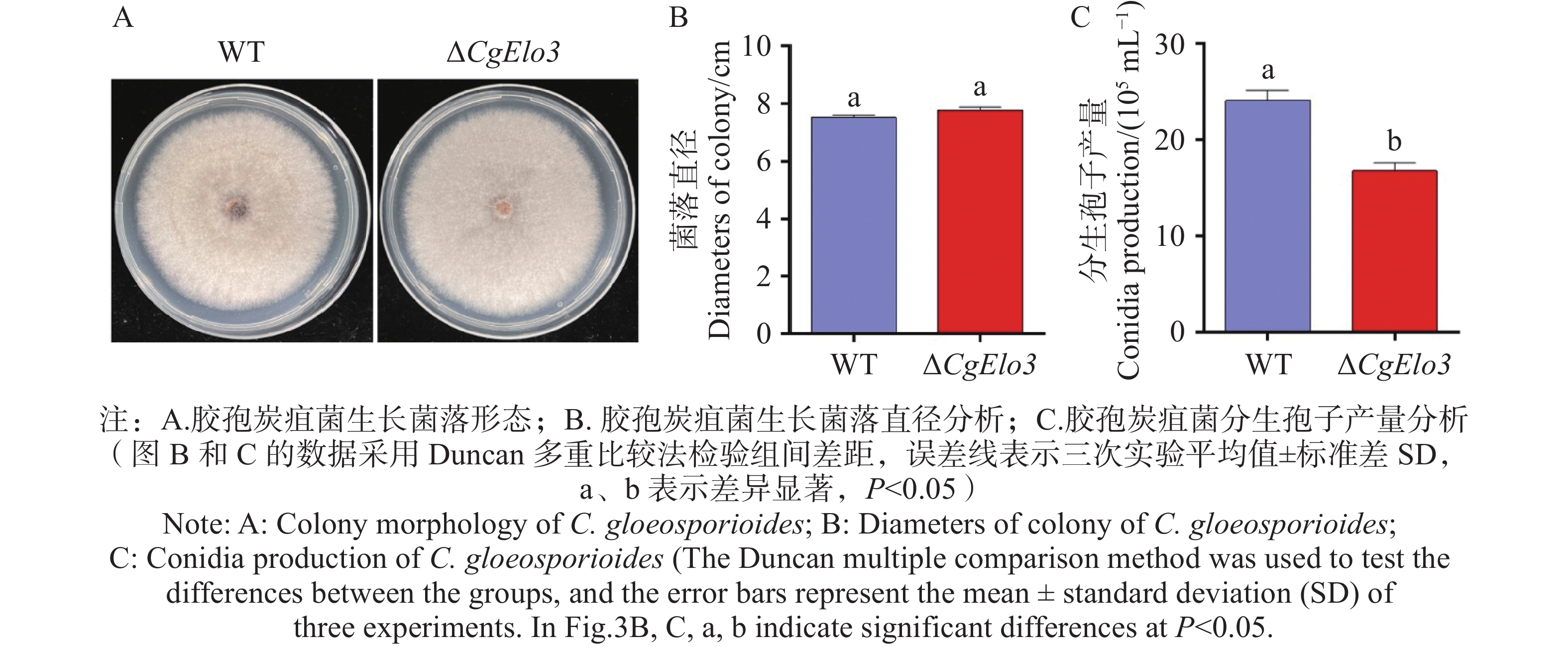
在CM固体培养基上接种WT和ΔCgElo3,5 d后观察胶孢炭疽菌菌落形态并记录生长直径。结果发现WT和ΔCgElo3的菌落形态及直径均不存在显著性差异(图3−A,B)。进一步探究产孢量发现,ΔCgElo3的分生孢子产量平均值为16.8×105个·mL−1,WT的分生孢子产量平均值为24.1×105个·mL−1(图3−C)。以上结果表明,CgElo3不影响胶孢炭疽菌的菌落形态和生长速率,但参与调控胶孢炭疽菌产孢。
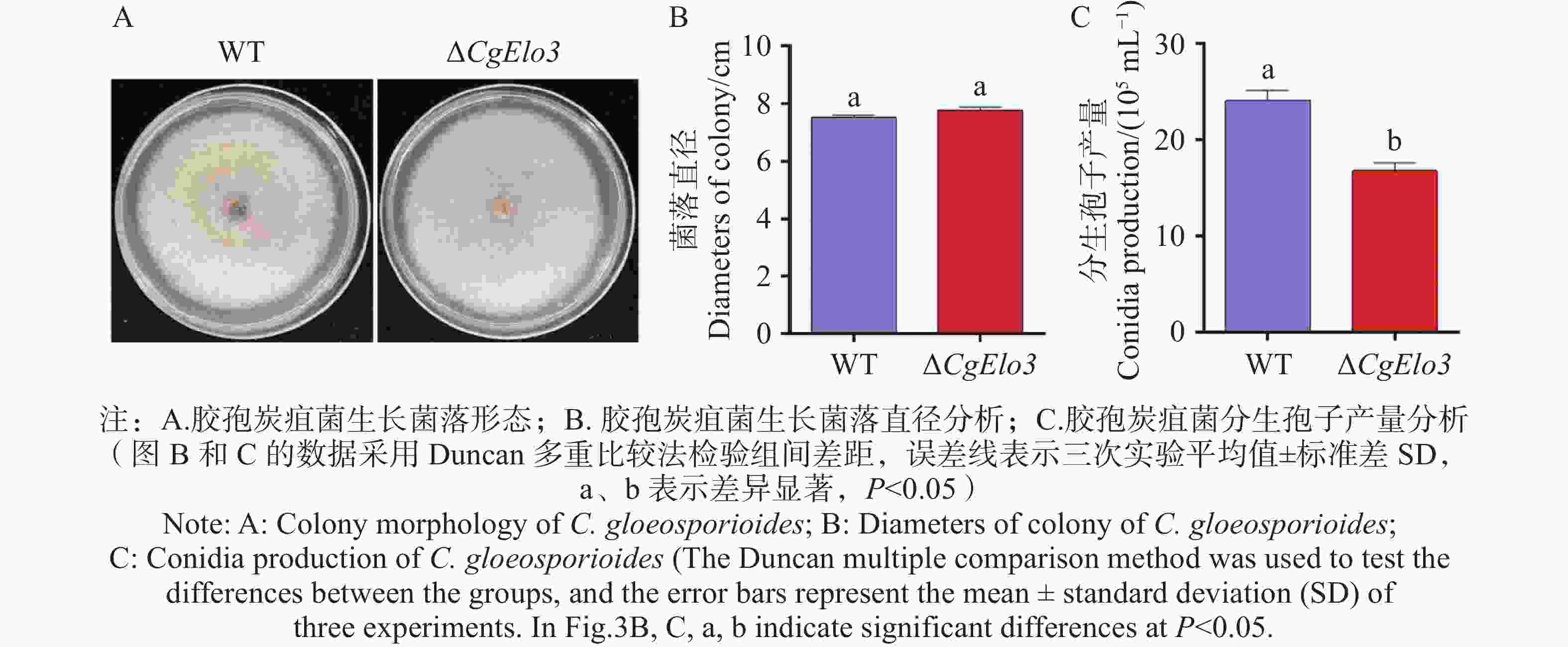
图 3 ΔCgElo3突变株生长速率与产孢量统计
Figure 3. Growth rate and conidia production statistics of the ΔCgElo3 mutant strain
-
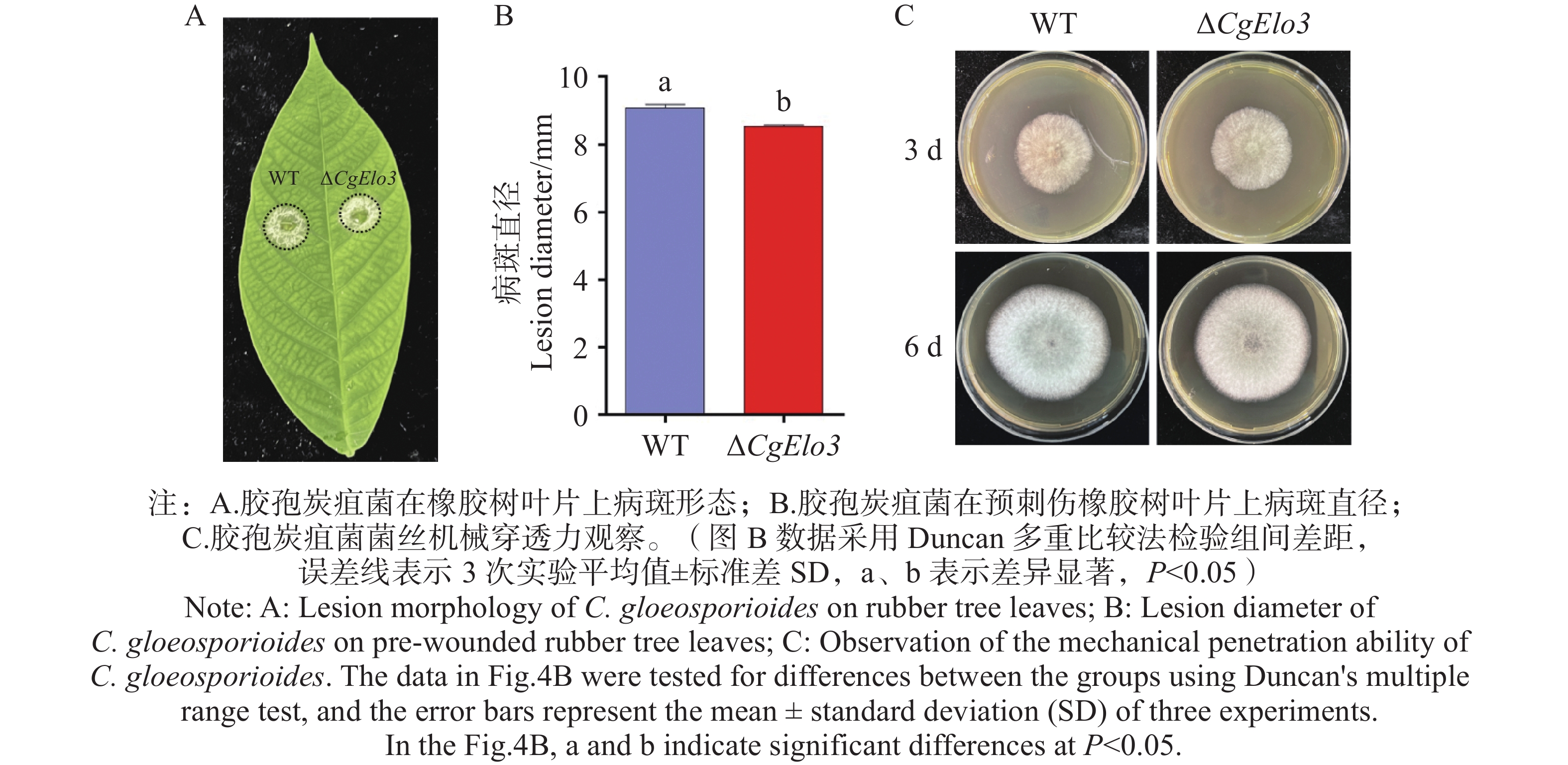
胶孢炭疽菌致病力结果显示,接种橡胶树叶片3 d后,WT引起的病斑平均直径为9.1 mm,而ΔCgElo3引起的病斑平均直径为8.5 mm,显著小于WT的病斑(图4−A,B)。在覆有玻璃纸的PDA培养基上进行机械穿透力实验结果发现,CgElo3的缺失并不影响胶孢炭疽菌的机械穿透力(图4−C)。这表明,CgElo3参与调控胶孢炭疽菌的致病力,但并非通过影响机械穿透力实现。
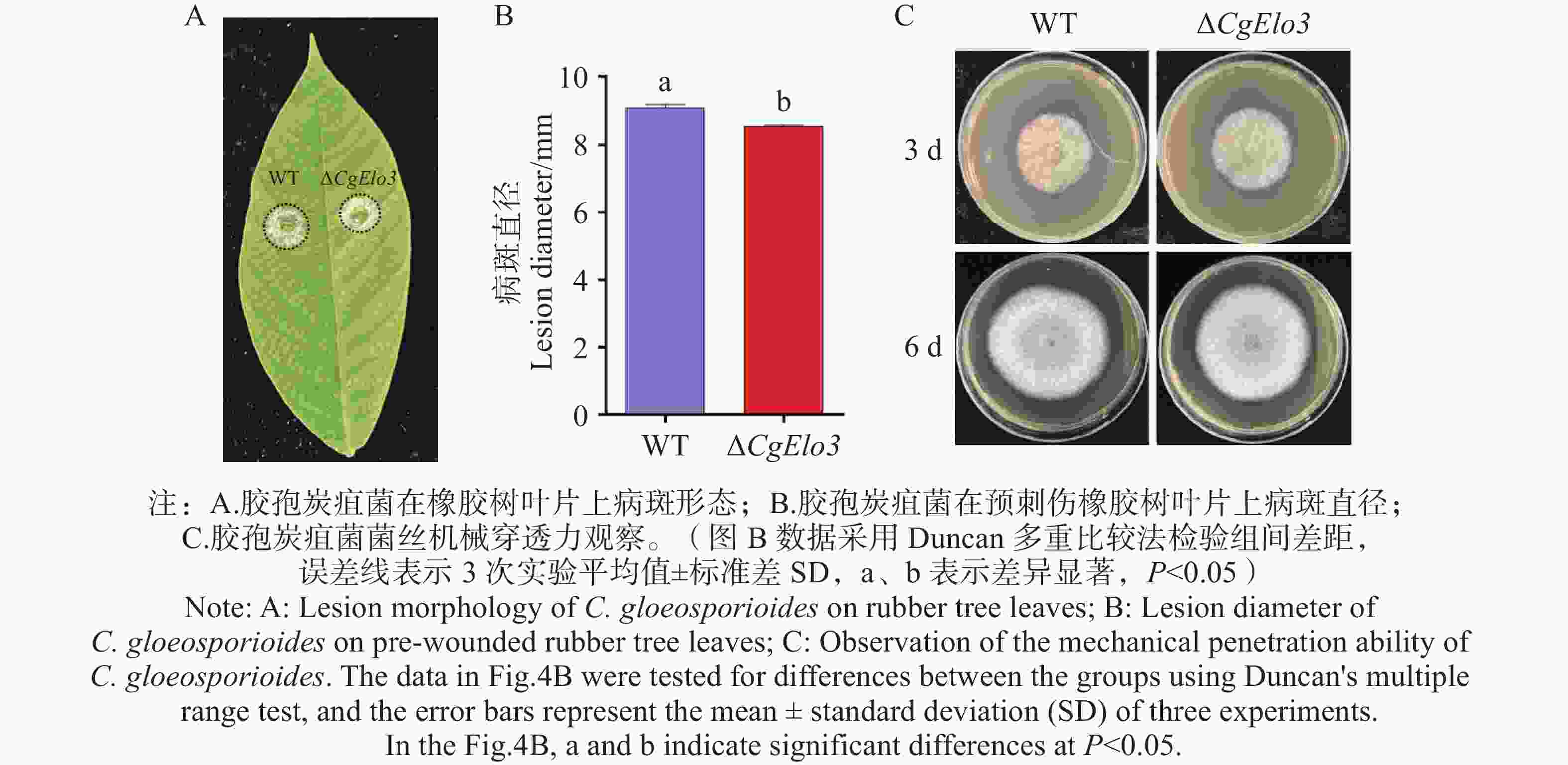
图 4 ΔCgElo3突变体致病力分析和机械穿透力观察
Figure 4. Pathogenicity analysis and mechanical penetration observation of the ΔCgElo3 mutant strain
-
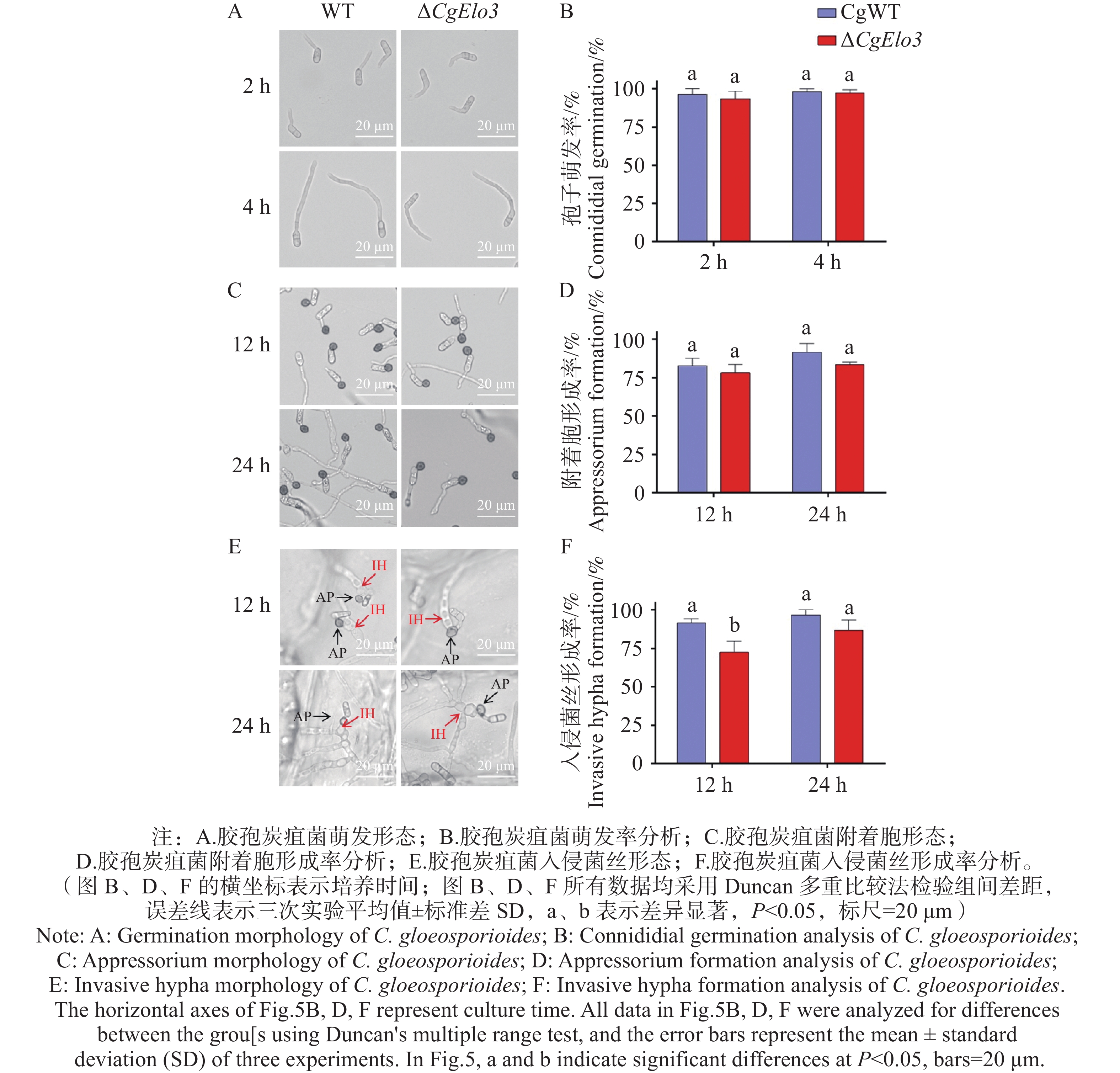
胶孢炭疽菌孢子萌发、附着胞及入侵菌丝形成结果(图5-A—D))发现,ΔCgElo3 的孢子萌发率以及其在疏水表面的附着胞形成率与WT相比均无显著性差异,说明CgElo3不影响胶孢炭疽菌萌发和附着胞的形成。然而,在洋葱内表皮侵染结果(图5−E,F)中,培养12 h后,WT入侵菌丝形成率为91.6%,ΔCgElo3 仅为72.5%,这说明CgElo3参与调控胶孢炭疽菌在侵染早期对寄主细胞的入侵。
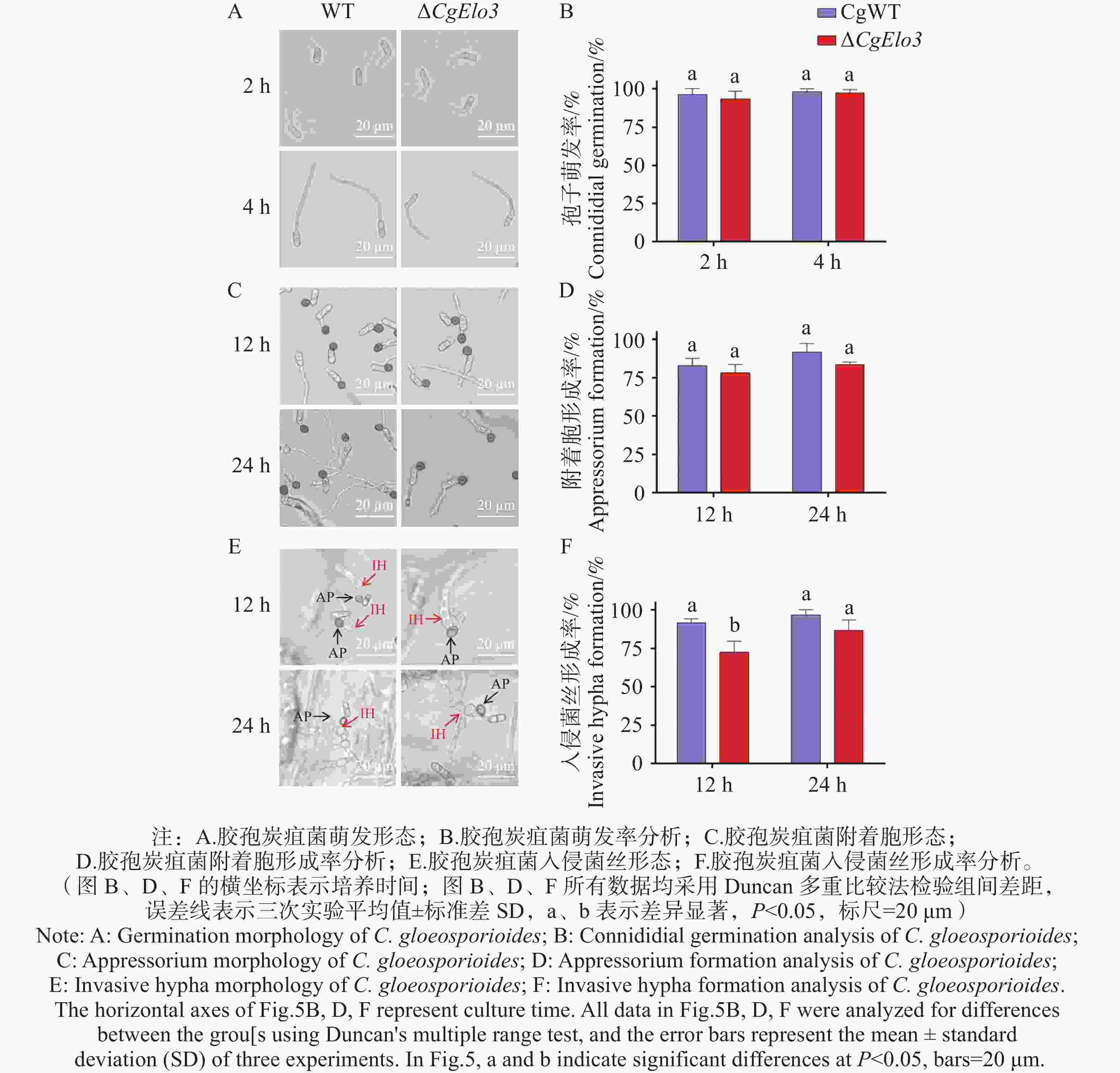
图 5 ΔCgElo3突变体萌发与入侵结构形成率分析
Figure 5. Analysis of germination and invasive structure formation rates of ΔCgElo3 mutant strain
-
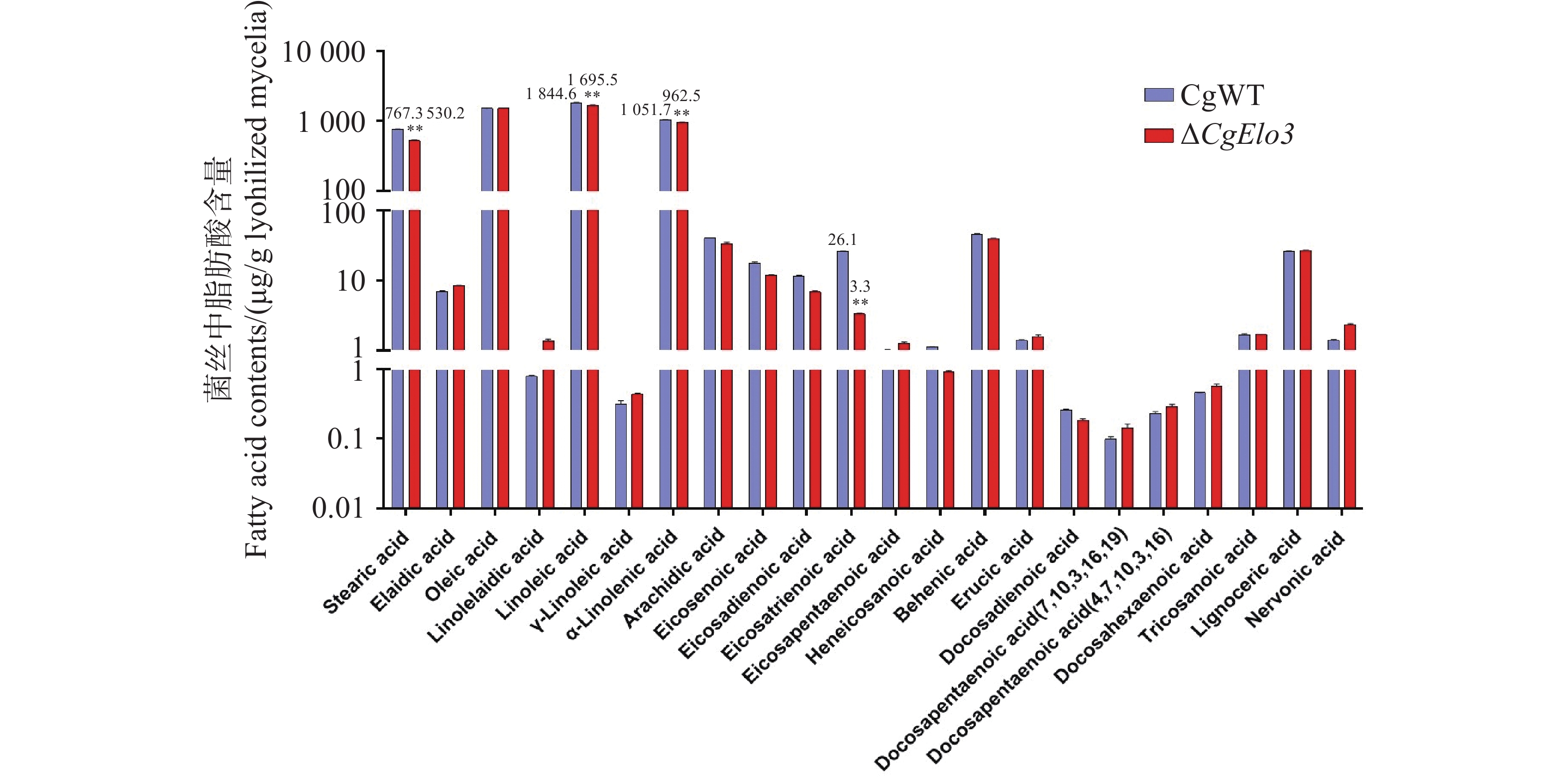
通过脂质组学分析WT与ΔCgElo3菌株中VLCFAs的相对含量,结果发现,ΔCgElo3中硬脂酸(Stearic acid)、亚油酸(Linoleic acid)、亚麻酸(α-Linolenic acid)及二十碳三烯酸(Eicosatrienoic acid)的相对含量显著低于WT(图6)。这证明了CgElo3在调控胶孢炭疽菌VLCFAs生物合成中的重要作用。
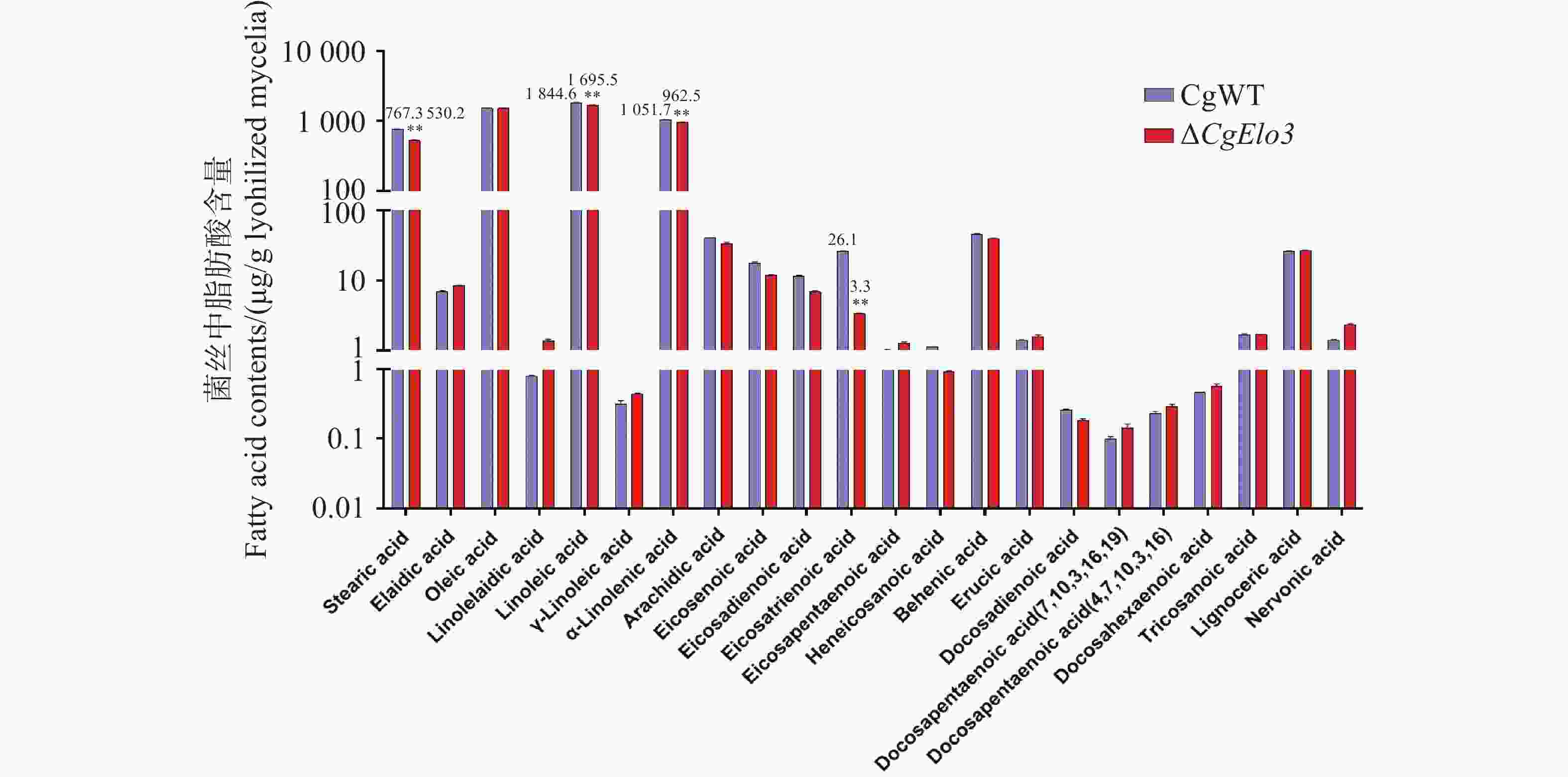
图 6 胶孢炭疽菌超长链脂肪酸相对含量的分析
Figure 6. Analysis of the relative content of Very Long-Chain Fatty Acids in C. gloeosporioides.
-
胶孢炭疽菌是当前引发橡胶树炭疽病的主要病原。胶孢炭疽菌的侵染过程可分为多个连续阶段:1)分生孢子(Conidia)产生粘性物质附着在橡胶树叶片表面并萌发出芽管(Germ Tube)[27-28];2)芽管尖端膨大形成的圆顶状结构-附着胞(Appressorium)[29];3)附着胞进一步发育形成侵染钉(Penetration peg)[30],积累黑色素等代谢物并产生膨压,膨压推动侵染钉以机械穿透力穿透叶片表面[31];4)胶孢炭疽菌分化形成入侵菌丝(Infection hyphae)吸取叶片营养[32],同时炭疽菌分泌效应蛋白(Effectors)干扰植物免疫促进菌丝在叶片内进一步扩展延伸[33]。本研究聚焦于胶孢炭疽菌中一个注释为Elo的基因,将其命名为CgElo3。分析转录组数据发现CgElo3在附着胞时期高水平表达,为进一步验证其在不同发育时期及侵染阶段的表达水平,采用荧光定量 RT-qPCR进行检测,发现与孢子时期相比,CgElo3在萌发和附着胞发育阶段的表达量显著升高;在胶孢炭疽菌侵染橡胶树过程中,CgElo3的表达量在侵染12 h时显著上调,但24 h至48 h的表达量持续降低。为探究CgElo3在胶孢炭疽菌中的功能,本实验构建了ΔCgElo3突变株并对其生理表型、致病力及VLCFAs含量等进行了系统分析。表型结果显示,ΔCgElo3分生孢子产量显著低于野生型,在接种预刺伤的橡胶树叶片上ΔCgElo3病斑偏小,并且不是机械穿透力丧失导致的。为进一步明确致病力下降的原因,本研究观察了胶孢炭疽菌萌发和入侵结构发育过程,发现CgElo3并不影响胶孢炭疽菌的正常萌发和形成附着胞,但在侵染早期洋葱表皮细胞内的入侵菌丝形成率显著低于WT。类似现象在稻瘟菌的研究中也有报道,其本质是缺失Elo1导致VLCFAs减少,从而干扰附着胞内Septin环的组装,最终阻断附着胞介导的植物侵染[14]。
VLCFAs是真核细胞膜磷脂、鞘脂以及表面蜡质层的核心构成单元,在调控细胞稳态[9-10],维持膜动态平衡[10]、雄性生育[19]、哺乳动物视力[34]及植物病原真菌附着胞膨压和机械穿透[14]等方面具有重要作用。Elo是调控VLCFAs合成的关键酶,参与调控细胞内VLCFAs的合成与积累,维持细胞膜上脂肪酸的平衡[11]、介导信号传导[18]、调控生长发育[11]及病原菌对逆境胁迫的响应[25]等。研究人员发现沉默PcElo7可显著降低柑橘全爪螨(Panonychus citri)体内的游离脂肪酸含量,从而促进杀虫剂的渗透[35]。类似地,在埃及伊蚊(Aedes aegypti)中,AeElo9与伊蚊调控卵正常发育和渗透压平衡密切相关[36]。Elo2和Elo3参与调控枯氏锥虫(Trypanosoma cruzi)维持脂质稳态[37]。酿酒酵母(S. cerevisiae)中有3个Elo基因,ScElo1与ScElo3的功能相似,主要催化14碳脂肪酸延伸至16碳[38-40],ScElo2参与合成24个碳原子以下的VLCFAs[40]。在植物病原真菌的研究证实Elo能够参与调控真菌的侵染过程及致病力。例如,研究人员在探究Elo的功能时发现,Elo2能够调控禾谷镰刀菌(F. graminearum)生长、对戊唑醇敏感性和致病力等[25]。Elo1缺失阻碍了稻瘟菌(M. oryzae)附着胞发育及对水稻的侵染[14]。这些结果说明,Elo不仅在脂质代谢中发挥重要作用,还可能通过调控特定代谢产物影响病原真菌的侵染结构形成、抗药性及对环境的适应性。本研究通过分析脂质组数据发现ΔCgElo3中硬脂酸(Stearic acid)、亚油酸(Linoleic acid)、亚麻酸(α-Linolenic acid)及二十碳三烯酸(Eicosatrienoic acid)的含量显著低于野生型。其中,硬脂酸(Stearic acid)是构成了脂质的核心骨架,为三酰甘油和甘油磷脂提供疏水尾部结构,从而影响脂滴和细胞膜形成、维持膜结构完整性等[41-42]。在酿酒酵母(S. cerevisiae)中,硬脂酸含量下降会导致细胞极性重塑及细胞内运输功能失调[43]。外源添加亚油酸可促进莱氏绿僵菌(Metarhizium rileyi)孢子萌发,影响角质层侵染进程,并改变莱氏绿僵菌致病力和逆境适应能力[44]。亚麻酸(α-Linolenic acid)不仅能够抑制细胞增殖、诱导细胞凋亡,还具有抗癌和抗氧化作用[45]。在植物中,亚麻酸(α-Linolenic acid)分布叶绿体基粒膜上,是多种多不饱和脂肪酸合成前体物质[46],不仅与植物抗氧化剂及茉莉酸合成密切相关[47],还在植物应对寒冷、干旱等非生物胁迫的过程中发挥重要作用[48-49]。二十碳三烯酸(Eicosatrienoic acid)在真核生物中可进一步代谢为花生四烯酸,生成前列腺素、血栓烷、白三烯等生物活性介质,参与免疫与细胞信号调控[50]。值得注意的是,亚油酸(Linoleic acid)、亚麻酸(α-Linolenic acid)及二十碳三烯酸(Eicosatrienoic acid)都属于多不饱和脂肪酸(Polyunsaturated fatty acids,PUFAs),PUFAs可通过环氧合酶、脂氧合酶或细胞色素P450途径代谢成氧化脂质[51]。一些氧化脂质能够诱导白色念珠菌(Monilia albican)酵母菌相与菌丝相的转化,调控烟曲霉(Aspergillus fumigatus)和黄曲霉(Aspergillus flavus)侧枝分化,并促进灰梨孢(Magnaporthe grisea)附着胞的形成[52-53]。此外,脂肪酸的合成受到多种因素的影响,包括不同脂肪酸合成调控方式不同[54],底物含量会影响酶反应效率[55]等。如油酸(Oleic acid)是一种常见的单不饱和脂肪酸,通常由硬脂酸经硬脂酰辅酶A去饱和酶(Stearoyl-CoA Desaturase, SCD)催化生成[54]。本研究发现ΔCgElo3中硬脂酸含量显著降低,但油酸水平与WT相比并未明显下降。这一现象可能与底物浓度变化影响酶反应效率有关,即硬脂酸减少可能影响了SCD的催化效率,从而导致ΔCgElo3与WT中油酸的含量无明显差异。同时,本实验室前期研究已证实,胶孢炭疽菌中存在另外两个Elo,其中CgElo2主要参与调控C16至C26超长链脂肪酸的延伸[8]。因此,胶孢炭疽菌缺失CgElo3后,虽然硬脂酸、亚麻酸等VLCFAs的含量显著减少,但由于底物减少可能调控下游去饱和酶的效率及CgElo2仍能正常发挥作用,所以油酸(Oleic acid)、二十碳烯酸(Eicosenoic acid)、神经酸(Nervonic acid)等VLCFAs的合成并未受到抑制。这解释了为何ΔCgElo3中部分VLCFAs的含量与WT相比无明显差异。
综上所述,亚麻酸(α-Linolenic acid)等VLCFAs作为氧化脂质的关键前体物质在真菌生长发育及侧枝分化中发挥重要调控作用,VLCFAs的含量可能直接影响胶孢炭疽菌孢子形成、侵染结构发育及致病力表现。由此推测,在胶孢炭疽菌中CgElo3的缺失导致亚麻酸(α-Linolenic acid)等VLCFAs的含量降低,从而影响ΔCgElo3分生孢子产量、侵染早期入侵菌丝的形成率,最终表现为致病力下降。这一结果揭示了CgElo3在胶孢炭疽菌脂肪酸代谢中的核心作用,为深入理解CgElo3致病机制提供了可能。这一结果也为绿色、靶向性防控策略的设计提供了重要依据。虽然本研究证实了CgElo3在胶孢炭疽菌生长发育及致病过程中的关键作用,但表型与基因间更深层次的因果关系还有待回补株构建后进一步验证。
The function of CgElo3, a very long-chain fatty acid elongase of Colletotrichum gloeosporioides
-
摘要: 橡胶树(Hevea brasiliensis)是天然橡胶的主要来源,作为重要的经济作物支持中国热带农业经济。胶孢炭疽菌(Colletotrichum gloeosporioides)引发的炭疽病是天然橡胶减产的重要原因。超长链脂肪酸(Very Long-Chain Fatty Acids, VLCFAs)在维持细胞膜稳定性及病原菌附着胞的发育过程中发挥重要作用。超长链脂肪酸延伸酶(Very Long-Chain Fatty Acids Elongase, Elo)是参与调控VLCFAs合成的限速酶。然而,Elo在胶孢炭疽菌中的功能尚不清楚。本研究聚焦于胶孢炭疽菌超长链脂肪酸延伸酶(CgElo3),通过分析ΔCgElo3生理表型可知,缺失CgElo3不影响菌丝生长和附着胞的形成,但显著降低了胶孢炭疽菌的分生孢子产量、对橡胶树叶片的致病力及对洋葱表皮侵染早期入侵菌丝的形成率。脂质组数据证实ΔCgElo3中硬脂酸(Stearic acid)、亚油酸(Linoleic acid)、亚麻酸(α-Linolenic acid)及二十碳三烯酸(Eicosatrienoic acid)相对含量显著减少。以上结果表明,CgElo3通过调控VLCFAs的生物合成,影响胶孢炭疽菌产孢及侵染早期对寄主细胞的入侵,进而降低其致病力。Abstract: Hevea brasiliensis is the main source of natural rubber and serves as an important economic crop supporting the tropical agricultural economy in our country. Anthracnose caused by Colletotrichum gloeosporioides is an important reason for reduced natural rubber yield. Very long-chain fatty acids (VLCFAs) play an important role in maintaining cell membrane stability and in the development of pathogen appressorium structures. The very-long-chain fatty acid elongase (Elo) is a rate-limiting enzyme involved in regulating VLCFA synthesis. However, the function of Elo in C. gloeosporioides remains unclear. This study focuses on the very-long-chain fatty acid elongase CgElo3 of C. gloeosporioides. Analysis of physiological phenotype in ΔCgElo3 shows that deletion of CgElo3 does not affect hyphal growth or the formation of appressorium, but it significantly reduces the conidiation of C. gloeosporioides, its pathogenicity on rubber tree leaves, and the formation rate of invasive hyphae during the early infection of onion epidermis. Lipidomic data confirmed that the relative contents of stearic acid, linoleic acid, α-linolenic acid, and eicosatrienoic acid were significantly reduced in ΔCgElo3. The above results indicate that CgElo3 affects the production of conidia and early host cell invasion of C. gloeosporioides by regulating the biosynthesis of VLCFAs, thereby reducing its pathogenicity.
-
图 2 Split-Marker基因敲除原理及鉴定ΔCgElo3突变株
Fig. 2 Principle of split-marker gene knockout and identification of ΔCgElo3 mutant strain
图 3 ΔCgElo3突变株生长速率与产孢量统计
Fig. 3 Growth rate and conidia production statistics of the ΔCgElo3 mutant strain
图 4 ΔCgElo3突变体致病力分析和机械穿透力观察
Fig. 4 Pathogenicity analysis and mechanical penetration observation of the ΔCgElo3 mutant strain
图 5 ΔCgElo3突变体萌发与入侵结构形成率分析
Fig. 5 Analysis of germination and invasive structure formation rates of ΔCgElo3 mutant strain
图 6 胶孢炭疽菌超长链脂肪酸相对含量的分析
注:数据采用Duncan多重比较法检验组间差距,误差线表示三次实验平均值±标准差SD。图中**表示P<0. 01
Fig. 6 Analysis of the relative content of Very Long-Chain Fatty Acids in C. gloeosporioides.
Note: The data were analyzed using Duncan's multiple range test to examine differences between the groups, and the error bars represent the mean ± standard deviation (SD)of three experiments. **: P<0. 01
表 1 实验用引物
Table 1 Primers used in the present experiment
引物名称 Primer 序列(5′—3′)Sequence 用途Usage CgElo3-5F GCGGCGTGCTTGACATAC 同源臂扩增Homologous arm amplification CgElo3-5R CTTTCCAAAGTCTGTGCC 同源臂扩增 Homologous arm amplification CgElo3-MF1 TTGGTCAATCCCCCACCGTGCCAACGCCACAGTG 同源臂扩增 Homologous arm amplification CgElo3-MR1 CACTGTGGCGTTGGCACGGTGGGGGATTGACCAA 同源臂扩增 Homologous arm amplification CgElo3-MF2 GAATTGCATGCTCTCACTTGTCTTGTGCGAAGTG 同源臂扩增 Homologous arm amplification CgElo3-MR2 CACTTCGCACAAGACAAGTGAGAGCATGCAATTC 同源臂扩增 Homologous arm amplification Sur-SLF CCTCTGATATTGGAAGCGACGC 同源臂扩增 Homologous arm amplification Sur-SLR ATGTTGGCATAAGCCGAACCGT 同源臂扩增 Homologous arm amplification Sur-JC5F GCGTTTGTAACTCTGCCTGTTTG 突变体鉴定 Mutant Strain Identification Sur-JC3R ACGAGGACCGCTACTCACATAC 突变体鉴定 Mutant Strain Identification CgElo3-JC5F CCCTTAGTTTGGACTGATGC 突变体鉴定 Mutant Strain Identification CgElo3-JC3R CGGGAACAAGGGAATAAAGAT 突变体鉴定 Mutant Strain Identification CgActin-qF GATTGGTATGGGCCAGAAGG 相对表达量检测 Relative expression level detection CgActin-qR GGAGCTCGTTGTAGAAGGTG 相对表达量检测 Relative expression level detection CgElo3-QF CTTCGCCTTTGTCGTTCTGC 相对表达量检测 Relative expression level detection CgElo3-QR TACCGTTGAGCTGGCAGAAG 相对表达量检测 Relative expression level detection  下载: 导出CSV
下载: 导出CSV
-
[1] 曾霞, 李维国, 高新生, 等. 巴西橡胶树选育种研究现状、趋势及我国的研究策略[J]. 热带农业科学, 2013, 33(6): 30−36. https://doi.org/10.3969/j.issn.1009-2196.2013.06.008 doi: 10.3969/j.issn.1009-2196.2013.06.008 [2] 高静, 程汉. 橡胶树改变世界的150年[J]. 中国科学: 生命科学, 2024, 54(10): 1744−1751. https://doi.org/10.1360/SSV-2024-0225 doi: 10.1360/SSV-2024-0225 [3] 李达, 张绍文. 天然橡胶产业发展政策分析: 红线与红利[J]. 林业经济问题, 2020, 40(2): 208−215. https://doi.org/10.16832/j.cnki.1005-9709.2020.02.012 doi: 10.16832/j.cnki.1005-9709.2020.02.012 [4] 蔡志英, 黄贵修. 巴西橡胶树炭疽病研究进展[J]. 西南林业大学学报, 2011, 31(1): 89−93. https://doi.org/10.3969/j.issn.1003-7179.2011.01.020 doi: 10.3969/j.issn.1003-7179.2011.01.020 [5] 符兴义. 海南橡胶树主要病害的防治[J]. 特种经济动植物, 2021, 24(9): 37−38. https://doi.org/10.3969/j.issn.1001-4713.2021.09.016 doi: 10.3969/j.issn.1001-4713.2021.09.016 [6] 郑肖兰, 刘先宝, 郑行恺, 等. 海南橡胶树炭疽病调查及喀斯特炭疽病菌的鉴定[J]. 热带作物学报, 2023, 44(12): 2546−2557. https://doi.org/10.3969/j.issn.1000-2561.2023.12.021 doi: 10.3969/j.issn.1000-2561.2023.12.021 [7] 刘西莉, 苗建强, 张灿. 植物病原菌抗药性及其抗性治理策略[J]. 农药学学报, 2022, 24(5): 921−936. https://doi.org/10.16801/j.issn.1008-7303.2022.0109 doi: 10.16801/j.issn.1008-7303.2022.0109 [8] 张贝. 橡胶树胶孢炭疽菌附着胞侵染结构形成的分子机制研究[D]. 海南: 海南大学, 2022. https://doi.org/10.27073/d.cnki.ghadu.2022.001354 [9] Erdbrügger P, Fröhlich F. The role of very long chain fatty acids in yeast physiology and human diseases [J]. Biological Chemistry, 2020, 402(1): 25−38. https://doi.org/10.1515/hsz-2020-0234 doi: 10.1515/hsz-2020-0234 [10] Micoogullari Y, Basu S S, Ang J, et al. Dysregulation of very-long-chain fatty acid metabolism causes membrane saturation and induction of the unfolded protein response [J]. Molecular Biology of the Cell, 2020, 31(1): 7−17. https://doi.org/10.1091/mbc.E19-07-0392 doi: 10.1091/mbc.E19-07-0392 [11] Wang Q, Du X X, Ma K, et al. A critical role for very long-chain fatty acid elongases in oleic acid-mediated Saccharomyces cerevisiae cytotoxicity [J]. Microbiological Research, 2018, 207: 1−7. https://doi.org/10.1016/j.micres.2017.11.001 doi: 10.1016/j.micres.2017.11.001 [12] Zimmermann C, Santos A, Gable K, et al. TORC1 inhibits GSK3-mediated Elo2 phosphorylation to regulate very long chain fatty acid synthesis and autophagy [J]. Cell Reports, 2013, 5(4): 1036−1046. https://doi.org/10.1016/j.celrep.2013.10.024 doi: 10.1016/j.celrep.2013.10.024 [13] Su J, Xu Y P, Lei M L, et al. A fatty acid elongase complex regulates cell membrane integrity and septin-dependent host infection by the rice blast fungus [J]. Molecular Plant Pathology, 2024, 25(7): e13494. https://doi.org/10.1111/mpp.13494 doi: 10.1111/mpp.13494 [14] He M, Su J, Xu Y P, et al. Discovery of broad-spectrum fungicides that block septin-dependent infection processes of pathogenic fungi [J]. Nature Microbiology, 2020, 5(12): 1565−1575. https://doi.org/10.1038/s41564-020-00790-y doi: 10.1038/s41564-020-00790-y [15] Jakobsson A, Westerberg R, Jacobsson A. Fatty acid elonhases in mammals: their regulation and roles in metabolism [J]. Progress in Lipid Research, 2006, 45(3): 237−249. https://doi.org/10.1016/j.plipres.2006.01.004 doi: 10.1016/j.plipres.2006.01.004 [16] Guillou H, Zadravec D, Martin P G P, et al. The key roles of elongases and desaturases in mammalian fatty acid metabolism: insights from transgenic mice [J]. Progress in Lipid Research, 2010, 49(2): 186−199. https://doi.org/10.1016/j.plipres.2009.12.002 doi: 10.1016/j.plipres.2009.12.002 [17] Ohno Y, Suto S, Yamanaka M, et al. ELOVL1 production of C24 acyl-CoAs is linked to C24 sphingolipid synthesis [J]. Proceedings of the National Academy of Sciences of the United States of America, 2010, 107(43): 18439−18444. https://doi.org/10.1073/pnas.1005572107 doi: 10.1073/pnas.1005572107 [18] Zhang Y, Pang S J, Sun B, et al. ELOVLs Predict distinct prognosis value and immunotherapy efficacy in patients with hepatocellular carcinoma [J]. Frontiers in Oncology, 2022, 12: 884066. https://doi.org/10.3389/fonc.2022.884066 doi: 10.3389/fonc.2022.884066 [19] Zadravec D, Tvrdik P, Guillou H, et al. ELOVL2 controls the level of n-6 28: 5 and 30: 5 fatty acids in testis, a prerequisite for male fertility and sperm maturation in mice [J]. Journal of Lipid Research, 2011, 52(2): 245−255. https://doi.org/10.1194/jlr.M011346 doi: 10.1194/jlr.M011346 [20] Wang X Y, Yu H, Gao R, et al. A comprehensive review of the family of very-long-chain fatty acid elongases: structure, function, and implications in physiology and pathology [J]. European Journal of Medical Research, 2023, 28(1): 532. https://doi.org/10.1186/s40001-023-01523-7 doi: 10.1186/s40001-023-01523-7 [21] Ramkumar H L, Tuo J, Shen D F, et al. Nutrient supplementation with n3 polyunsaturated fatty acids, lutein, and zeaxanthin decrease A2E accumulation and VEGF expression in the retinas of Ccl2/Cx3cr1-deficient mice on Crb1rd8 background1-3 [J]. The Journal of Nutrition, 2013, 143(7): 1129−1135. https://doi.org/10.3945/jn.112.169649 doi: 10.3945/jn.112.169649 [22] Li W M, Sandhoff R, Kono M, et al. Depletion of ceramides with very long chain fatty acids causes defective skin permeability barrier function, and neonatal lethality in ELOVL4 deficient mice [J]. International Journal of Biological Sciences, 2007, 3(2): 120−128. https://doi.org/10.7150/ijbs.3.120 doi: 10.7150/ijbs.3.120 [23] Moon Y A, Hammer R E, Horton J D. Deletion of ELOVL5 leads to fatty liver through activation of SREBP-1c in mice [J]. Journal of Lipid Research, 2009, 50(3): 412−423. https://doi.org/10.1194/jlr.M800383-JLR200 doi: 10.1194/jlr.M800383-JLR200 [24] Kinugasa Y, Hirano Y, Sawai M, et al. The very-long-chain fatty acid elongase Elo2 rescues lethal defects associated with loss of the nuclear barrier function in fission yeast cells [J]. Journal of Cell Science, 2019, 132(10): jcs229021. https://doi.org/10.1242/jcs.229021 doi: 10.1242/jcs.229021 [25] Wang H X, Chen Y, Hou T J, et al. The very long-chain fatty acid elongase FgElo2 governs tebuconazole sensitivity and virulence in Fusarium graminearum [J]. Environmental Microbiology, 2022, 24(11): 5362−5377. https://doi.org/10.1111/1462-2920.16212 doi: 10.1111/1462-2920.16212 [26] 郭云峰, 安邦. 橡胶树胶孢炭疽菌NADPH氧化酶功能研究[J]. 生物技术通报, 2018, 34(10): 165−171. https://doi.org/10.13560/j.cnki.biotech.bull.1985.2018-0344 doi: 10.13560/j.cnki.biotech.bull.1985.2018-0344 [27] Breth B, Odenbach D, Yemelin A, et al. The role of the Tra1p transcription factor of Magnaporthe oryzae in spore adhesion and pathogenic development [J]. Fungal Genetics and Biology, 2013, 57: 11−22. https://doi.org/10.1016/j.fgb.2013.05.008 doi: 10.1016/j.fgb.2013.05.008 [28] Kershaw M J, Basiewicz M, Soanes D M, et al. Conidial morphogenesis and septin-mediated plant infection require Smo1, a Ras GTPase-activating protein in Magnaporthe oryzae [J]. Genetics, 2019, 211(1): 151−167. https://doi.org/10.1534/genetics.118.301490 doi: 10.1534/genetics.118.301490 [29] Hamer J E, Howard R J, Chumley F G, et al. A mechanism for surface attachment in spores of a plant pathogenic fungus [J]. Science, 1988, 239(4837): 288−290. https://doi.org/10.1126/science.239.4837.288 doi: 10.1126/science.239.4837.288 [30] 林春花, 牟保辉, 刘文波, 等. 疏水表面诱导橡胶树炭疽菌侵染结构的发育分化过程[J]. 植物保护学报, 2018, 45(3): 470−477. https://doi.org/10.13802/j.cnki.zwbhxb.2018.2016207 doi: 10.13802/j.cnki.zwbhxb.2018.2016207 [31] 张晓林, 张俊娥, 贺璞慧中, 等. 胶孢炭疽菌侵染杨树叶片的组织病理学研究[J]. 北京林业大学学报, 2018, 40(3): 101−109. https://doi.org/10.13332/j.1000-1522.20170385 doi: 10.13332/j.1000-1522.20170385 [32] Thomson D D, Wehmeier S, Byfield F J, et al. Contact-induced apical asymmetry drives the thigmotropic responses of Candida albicans hyphae [J]. Cellular Microbiology, 2015, 17(3): 342−354. https://doi.org/10.1111/cmi.12369 doi: 10.1111/cmi.12369 [33] Cannon P F, Damm U, Johnston P R, et al. Colletotrichum–current status and future directions [J]. Studies in Mycology, 2012, 73: 181−213. https://doi.org/10.3114/sim0014 doi: 10.3114/sim0014 [34] Barabas P, Liu A H, Xing W, et al. Role of ELOVL4 and very long-chain polyunsaturated fatty acids in mouse models of Stargardt type 3 retinal degeneration [J]. Proceedings of the National Academy of Sciences of the United States of America, 2013, 110(13): 5181−5186. https://doi.org/10.1073/pnas.1214707110 doi: 10.1073/pnas.1214707110 [35] Wang H Y, Xia B, Wang H F, et al. Fatty acid elongase gene PcELO7 is essential for lipid accumulation and fecundity of Panonychus citri (Acari: Tetranychidae) [J]. Journal of Agricultural and Food Chemistry, 2024, 72(4): 2100−2108. https://doi.org/10.1021/acs.jafc.3c07412 doi: 10.1021/acs.jafc.3c07412 [36] Chen J, Wu Y C, Chen J K, et al. Elongases of long-chain fatty acids ELO2 and ELO9 are involved in cuticle formation and function in fecundity in the yellow fever mosquito, Aedes aegypti [J]. Insects, 2023, 14(2): 189. https://doi.org/10.3390/insects14020189 doi: 10.3390/insects14020189 [37] Pagura L, Dumoulin P C, Ellis C C, et al. Fatty acid elongases 1-3 have distinct roles in mitochondrial function, growth, and lipid homeostasis in Trypanosoma cruzi [J]. Journal of Biological Chemistry, 2023, 299(6): 104715. https://doi.org/10.1016/j.jbc.2023.104715 doi: 10.1016/j.jbc.2023.104715 [38] Toke D A, Martin C E. Isolation and characterization of a gene affecting fatty acid elongation in Saccharomyces cerevisiae [J]. Journal of Biological Chemistry, 1996, 271(31): 18413−18422. https://doi.org/10.1074/jbc.271.31.18413 doi: 10.1074/jbc.271.31.18413 [39] Schneiter R, Tatzer V, Gogg G, et al. Elo1p-dependent carboxy-terminal elongation of C14: 1Δ9 to C16: 1Δ11 fatty acids in Saccharomyces cerevisiae [J]. Journal of Bacteriology, 2000, 182(13): 3655−3660. https://doi.org/10.1128/JB.182.13.3655-3660.2000 doi: 10.1128/JB.182.13.3655-3660.2000 [40] Oh C S, Toke D A, Mandala S, et al. ELO2 and ELO3, homologues of the Saccharomyces cerevisiae ELO1 gene, function in fatty acid elongation and are required for sphingolipid formation [J]. Journal of Biological Chemistry, 1997, 272(28): 17376−17384. https://doi.org/10.1074/jbc.272.28.17376 doi: 10.1074/jbc.272.28.17376 [41] Alvarado Cader A, Allan Butterfield D, Watkins B A, et al. Electron spin resonance studies of fatty acid-induced alterations in membrane fluidity in cultured endothelial cells [J]. The International Journal of Biochemistry & Cell Biology, 1995, 27(7): 665−673. https://doi.org/10.1016/1357-2725(95)00036-O doi: 10.1016/1357-2725(95)00036-O [42] Shiraishi K, Matsuzaki S, Itakura M, et al. Abnormality in membrane fatty acid compositions of cells measured on erythrocyte in alcoholic liver disease [J]. Alcoholism, Clinical and Experimental Research, 1996, 20(S1): 56a−59a. https://doi.org/10.1111/j.1530-0277.1996.tb01730.x doi: 10.1111/j.1530-0277.1996.tb01730.x [43] Doignon F, Laquel P, Testet E, et al. Requirement of phosphoinositides containing stearic acid to control cell polarity [J]. Molecular and Cellular Biology, 2016, 36(5): 765−780. https://doi.org/10.1128/MCB.00843-15 doi: 10.1128/MCB.00843-15 [44] Wang G, Zhang X, Du G Z, et al. Oleic acid and linoleic acid enhances the biocontrol potential of Metarhizium rileyi [J]. Journal of Fungi, 2024, 10(8): 521. https://doi.org/10.3390/jof10080521 doi: 10.3390/jof10080521 [45] Yan H R, Zhang S M, Yang L, et al. The antitumor effects of α-linolenic acid [J]. Journal of Personalized Medicine, 2024, 14(3): 260. https://doi.org/10.3390/jpm14030260 doi: 10.3390/jpm14030260 [46] Hageman J H J, Hooyenga P, Diersen-Schade D A, et al. The impact of dietary long-chain polyunsaturated fatty acids on respiratory illness in infants and children [J]. Current Allergy and Asthma Reports, 2012, 12(6): 564−573. https://doi.org/10.1007/s11882-012-0304-1 doi: 10.1007/s11882-012-0304-1 [47] 张红. 海岛棉α-亚麻酸代谢相关基因在NaCl胁迫下的功能鉴定[D]. 乌鲁木齐: 新疆农业大学, 2022. https://doi.org/10.27431/d.cnki.gxnyu.2022.000248 [48] Weber H. Fatty acid-derived signals in plants [J]. Trends in Plant Science, 2002, 7(5): 217−224. https://doi.org/10.1016/S1360-1385(02)02250-1 doi: 10.1016/S1360-1385(02)02250-1 [49] 李东, 甄春燕, 陈德富, 等. α-亚麻酸对干旱胁迫下水稻种子萌发的影响[J]. 广西植物, 2018, 38(8): 1025−1031. https://doi.org/10.11931/guihaia.gxzw201709004 doi: 10.11931/guihaia.gxzw201709004 [50] Wei C F, Wang M, Wang X J. Evolutionary conservation analysis of human arachidonic acid metabolism pathway genes [J]. Life Medicine, 2023, 2(2): lnad004. https://doi.org/10.1093/lifemedi/lnad004 doi: 10.1093/lifemedi/lnad004 [51] Eccles J A, Baldwin W S. Detoxification cytochrome P450s (CYPs) in families 1-3 produce functional oxylipins from polyunsaturated fatty acids [J]. Cells, 2022, 12(1): 82. https://doi.org/10.3390/cells12010082 doi: 10.3390/cells12010082 [52] Tsitsigiannis D I, Keller N P. Oxylipins as developmental and host–fungal communication signals [J]. Trends in Microbiology, 2007, 15(3): 109−118. https://doi.org/10.1016/j.tim.2007.01.005 doi: 10.1016/j.tim.2007.01.005 [53] Niu M Y, Steffan B N, Fischer G J, et al. Fungal oxylipins direct programmed developmental switches in filamentous fungi [J]. Nature Communications, 2020, 11(1): 5158. https://doi.org/10.1038/s41467-020-18999-0 doi: 10.1038/s41467-020-18999-0 [54] Kalish B T, Fallon E M, Puder M. A tutorial on fatty acid biology [J]. Journal of Parenteral and Enteral Nutrition, 2012, 36(4): 380−388. https://doi.org/10.1177/0148607112449650 doi: 10.1177/0148607112449650 [55] Cormier H, Rudkowska I, Lemieux S, et al. Effects of FADS and ELOVL polymorphisms on indexes of desaturase and elongase activities: results from a pre-post fish oil supplementation [J]. Genes & Nutrition, 2014, 9(6): 437. https://doi.org/10.1007/s12263-014-0437-z doi: 10.1007/s12263-014-0437-z -

 点击查看大图
点击查看大图
计量
- 文章访问数: 193
- HTML全文浏览量: 131
- 被引次数: 0
