


-
天然橡胶是重要的工业原料和战略物资,主要来源于巴西橡胶树(Hevea brasiliensis),其产量受到橡胶树病害的影响。胶孢炭疽菌(Colletotrichum gloeosporioides)是引起橡胶树胶孢炭疽病的主要病原菌[1 − 2],对小苗、幼树及成年开割胶树均能造成危害,导致橡胶树嫩芽回枯、叶片脱落和割胶时间延迟,给橡胶产业造成巨大的经济损失[3 − 5]。胶孢炭疽菌是典型的半活体营养寄生真菌,其对寄主植物的侵染包括入侵、定植的活体寄生营养阶段和破坏、分解植物组织的坏死寄生营养阶段 [6]。在活体寄生阶段的入侵过程中,分生孢子的萌发、附着胞和入侵菌丝的形成起关键作用[7]。
CFEM(common in fungal extracellular membrane)结构域是真菌所特有的基序,约由60个氨基酸残基组成,其中含有呈规律性间隔分布的8个保守的半胱氨酸残基(Cx3Cx10–12Cx6–7CxCx11–14Cx4Cx15–16C)[8 − 10]。根据是否存在跨膜结构域,真菌CFEM蛋白被分为两种类型:Pth11-like型和Non-Pth11-like型 [6, 11]。Non-Pth11-like型CFEM蛋白除了CFEM结构域外缺乏跨膜结构域,只含有信号肽序列或含有信号肽和GPI锚定位点,被认为是病原真菌的重要致病因子——效应蛋白[12 − 13]。Pth11-like型CFEM蛋白除了保守的CFEM结构域外还含多个跨膜结构域,可能作为信号受体在寄主–病原菌相互作用中发挥重要作用[12 − 14]。
在植物病原真菌中,Pth11-like型CFEM蛋白以稻瘟病菌(Magnaporthe grisea)Pth11蛋白为典型代表,该蛋白含有7个跨膜结构域,可作为G蛋白偶联受体发挥作用,调控稻瘟菌附着胞形成及致病性[11, 14]。稻瘟菌中另外1个结构相似的CFEM蛋白WISH也是1个典型的Pth11-like型CFEM蛋白,同样含有7个跨膜结构域,不仅与病菌的附着胞形成和致病性有关,还参与调控稻瘟菌对疏水表面的识别和细胞壁的完整性[9]。在灰霉菌(Botrytis cinerea)中,鉴定了3个Pth11-like型CFEM蛋白,分别是Bcin05g02420、Bcin15g02580和BCIN 07g03260,其中Bcin05g02420 和Bcin15g02580分别含有6个和8个跨膜结构域,BCIN 07g03260含有1个跨膜结构域,而BCIN 07g03260的缺失导致灰霉病菌的分生孢子萌发率和芽管伸长率降低约20%~30%,对番茄的致病力显著降低,但是不影响灰霉病菌的生长和分生孢子产量[15]。此外,在禾谷镰刀菌(Fusarium graminearum)、杨树黑斑病菌(Marssonina brunnea)、四川新小滴孢腔菌(Neostagonosporella sichuanensis)、粗糙脉孢菌(Neurospora crassa)、深绿木霉(Trichoderma atroviride)、里氏木霉(T. reesei)和绿色木霉(T. viride)中鉴定到了多个Pth11-like型CFEM蛋白,但对其功能缺乏研究[13, 16 − 20]。
在炭疽菌中,Non-Pth11-like型CFEM蛋白的功能研究较多[6, 21 − 25],但关于Pth11-like型CFEM蛋白的功能研究较少。已有的研究表明,杨树胶孢炭疽菌中Pth11的同源蛋白CgPth11参与疏水表面附着胞形成的调控,但并不是致病性所必需[26]。在希金斯炭疽菌(Colletotrichum higginsamum)中也鉴定到了6个Pth11-like型CFEM蛋白,但其功能未知[27]。在前期研究中,本研究团队对橡胶树胶孢炭疽菌(C. gloeosporioides)进行了转录组序列测定,并从中搜索到了1个编码Pth11-like 型CFEM蛋白的基因,将其命名为CgCFEM17。本研究团队对该基因进行了克隆和分析,通过构建该基因的敲除突变体△CgCFEM17和回补突变体△CgCFEM17-C,分析△CgCFEM17和△CgCFEM17-C 在生长发育和致病性方面的表型变化,解析了CgCFEM17在胶胞炭疽菌致病过程中的功能,为橡胶树炭疽病防控和新型农药设计开发提供新靶点提供理论依据。
-
胶孢炭疽菌(C. gloeosporioides)野生型菌株(WT)由本实验分离、鉴定并保存。巴西橡胶树品种为‘Reyan 7-33-97’。大肠杆菌(Escherichia coli)TOP10菌株(CC96105)购于上海吐露港生物科技有限公司。
-
马铃薯葡萄糖琼脂培养基(potato dextrose agar, PDA):称12.5 g PDA粉末,200 mL ddH2O溶解。完全培养基(complete medium, CM):蛋白胨1 g,酵母提取物0.5 g,微量元素0.5 mL,酪蛋白水解物0.5 g,葡萄糖5 g,维生素0.5 mL,硝酸盐25 mL,400 mL蒸馏水溶解,调pH至6.5,定容到500 mL。0.5%(v/v)ME:称0.5 g MALT EXTRACT粉末,ddH2O溶解,定容到100 mL。2%(v/v) YCS液体培养基:称酵母提取物0.1 g,酸水解酪蛋白0.1 g,蔗糖2 g,ddH2O溶解并定容到100 mL。
-
使用DNAMAN软件对CgCFEM17进行翻译;SignalP6.0(
https://services.healthtech.dtu.dk/services/SignalP-6.0/ )进行信号肽预测;SMART(https://smart.embl.de/smart/ )进行CFEM结构域预测;TMHMM2.0(TMHMM 2.0-DTU Health Tech-Bioinformatic Services)进行跨膜结构域预测;MEGA6.0软件进行系统发育树的构建。 -
将不同的菌株分别接种在马铃薯葡萄糖培养基(PDA)上,28 ℃恒温培养箱培养4~6 d,将培养好的菌株用手术刀切适量菌块于20 mL的CM液体培养基中,28 ℃摇床培养2 d,用滤膜过滤,5 000 r·min−1离心1 min,得到分生孢子。
-
根据同源重组的敲除原理,使用表1中的引物对CFEM17-5F/CFEM17-M5R、CFEM17-M3F/CFEM17-3R、CFEM17-M5F/CFEM17-M3R分别扩增CgCFEM17基因的上游同源臂、下游同源臂和抗性基因片段。使用引物对CFEM17-5F/SUR-SLR、SUR-SLF/CFEM17-3R将上下游同源臂与抗性基因片段分别融合得到融合片段,利用PEG介导的转化法将融合片段转入胶孢炭疽菌野生型原生质体中[28],用氯嘧磺隆进行筛选。使用引物对CFEM17-JC5F/SUR-JC5R和SUR-JC3F/CFEM17-JC3分别鉴定基因上下游,获得阳性转化子并对其进行单孢分离。使用引物对CFEM17-F(XbaI)/CFEM17-R(SmaI)检测CgCFEM17是否被敲除,从而获得CgCFEM17基因的敲除突变体。
表 1 引物信息
Table 1. Primer information
引物名称Primer 引物序列 Primer sequence(5′-3′) CFEM17-5F CGACGTGTAGAGCTATGACC CFEM17-M5F CTTGTGCGTTTCAAGAAGTGCCAACGCCACAGTG CFEM17-M5R CACTGTGGCGTTGGCACTTCTTGAAACGCACAAG CFEM17-M3F GAATTGCATGCTCTCACGCGTTTGAGTTTTGTTT CFEM17-M3R AAACAAAACTCAAACGCGTGAGAGCATGCAATTC CFEM17-3R TTTGTCATTCTGAAGTCCC SUR-SLR ATGTTGGCATAAGCCGAACCGT SUR-SLF CCTCTGATATTGGAAGCGACGC CFEM17-JC5F GCCGCTCGCATTTCATTCC SUR-JC5R GCGTTTGTAACTCTGCCTGTTTG SUR-JC3F ACGAGGACCGCTACTCACATAC CFEM17-JC3R ACACCCCAAAAGGTCCCAC CFEM17-F(XbaI) TCTAGAATGTTCACCAACGCCAGAAA CFEM17-R(SmaI) CCCGGGTACTTTATTTGAATGGCCAA CFEM17-OF(XbaI) TCTAGACATAACTGATGCGACACCT 使用引物对CFEM17-OF(XbaI)/CFEM17-R(SmaI)扩增CFEM17基因及其启动子,通过酶切连接的方式构建由CgCFEM17基因自身启动子驱动的表达载体,将构建好的表达载体线性化后转入CgCFEM17基因敲除突变菌株的原生质体中,使用潮霉素筛选出阳性转化子进行单孢分离。使用引物对CFEM17-F(XbaI)/CFEM17-R(SmaI)检测CgCFEM17是否回补成功,获得CgCFEM17基因的敲除回补菌株。
-
取待测菌株直径为6 mm的菌块,分别接种于PDA培养基上,28 ℃培养5 d,用十字交叉法测量菌落直径并拍照。实验独立重复3次,每次4个重复。
-
按照1.2.2中的方法得到不同菌株的分生孢子,用0.5%(v/v)ME液体培养基重悬孢子,使用血球计数板在显微镜下计数,将孢子悬浮液浓度调整到2×105 个·mL−1,取5 μL接种于嫩绿期的橡胶树叶片,密封保湿,于28 ℃培养4 d后,测量病斑直径并拍照记录。实验设置30个重复。
-
按照1.2.2中的方法得到不同菌株的分生孢子,用2%(v/v)的YCS液体培养基重悬孢子,调整孢子液浓度到5×105 个·mL−1,取20 μL接种于载玻片,28 ℃保湿培养,分别在0、1.0、1.5和2 h时于显微镜下观察并统计分生孢子的萌发率。实验独立重复3次,每次重复不少于100个孢子。
-
按照1.2.2中的方法得到不同菌株的分生孢子,调整孢子液为5×105 个·mL−1,取5 μL接种于疏水平板上,28 ℃保湿培养,分别在0、2、4、8 h于显微镜下观察附着胞的形成过程并统计附着胞形成率。实验独立重复3次,每次重复不少于100个孢子。
-
按照1.2.2中的方法得到不同菌株的分生孢子,分生孢子用无菌水洗涤两次,稀释孢子液到3×105 个·mL−1,将洋葱内表皮揭下置于培养皿中,将稀释好的孢子液取5 μL接种于洋葱表皮上,28 ℃保湿培养,分别于8、12 h在显微镜下观察入侵菌丝的形态并统计入侵结构的形成率。实验独立重复3次。
-
使用Graphpad Prism 6.0软件进行数据统计及显著性分析,通过单因素方差分析具有单个变量的数据,并通过 Duncan氏多范围检验进行均值分离。P < 0.05 时差异被认为是显著的。
-
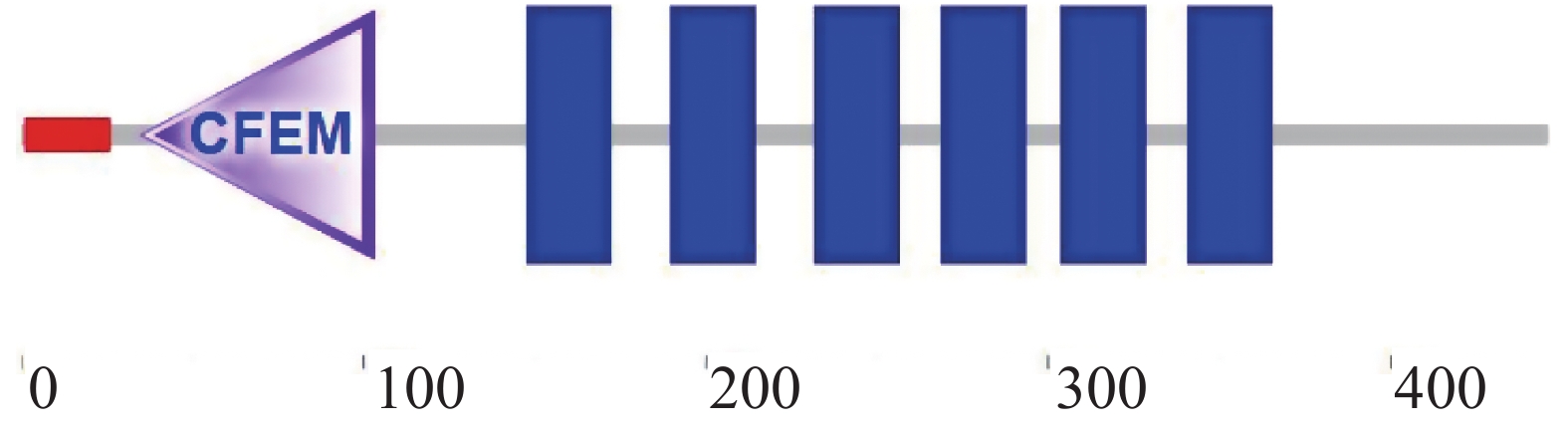
以橡胶树胶孢炭疽菌的cDNA为模板,通过RT-PCR的方法对CgCFEM17基因的编码区进行了扩增,测序结果显示,该基因的编码区包含1 341 bp,编码446 aa的肽链。对CgCFEM17蛋白的结构进行分析发现,该蛋白的N端含有1个信号肽序列和(1~26 aa)1个CFEM保守结构域(35~103 aa),C端含有6个跨膜结构域(149~171 aa,191~213 aa,233~255 aa,270~292 aa,305~327 aa,342~364 aa)(图1),符合Pth11-like 型CFEM蛋白的结构特征。利用MEGA 6.0软件对CgCFEM17与其他CFEM类蛋白的氨基酸序列进行系统进化树的构建(图2),结果显示,CgCFEM17与其他已知功能的Pth11-like型CFEM蛋白聚在一起,与稻瘟菌Pth11和杨树的CgPth11的亲缘关系较近。
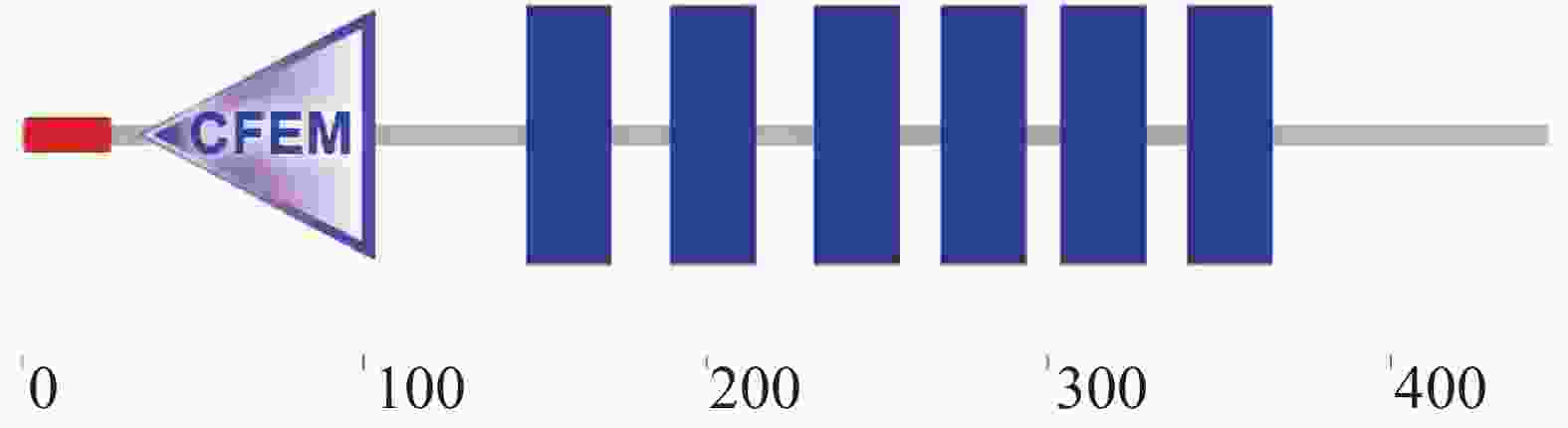
图 1 胶孢炭疽菌CgCFEM17结构分析
Figure 1. The structure analysis of CgCFEM17 of Colletotrichum gloeosporioides

图 2 胶孢炭疽菌CgCFEM17与其他病原真菌的CFEM类蛋白的氨基酸序列的系统发育关系
Figure 2. Phylogenetic relationship of the amino acid sequence between CgCFEM17 andCFEM protein of other pathogenic fungi
-
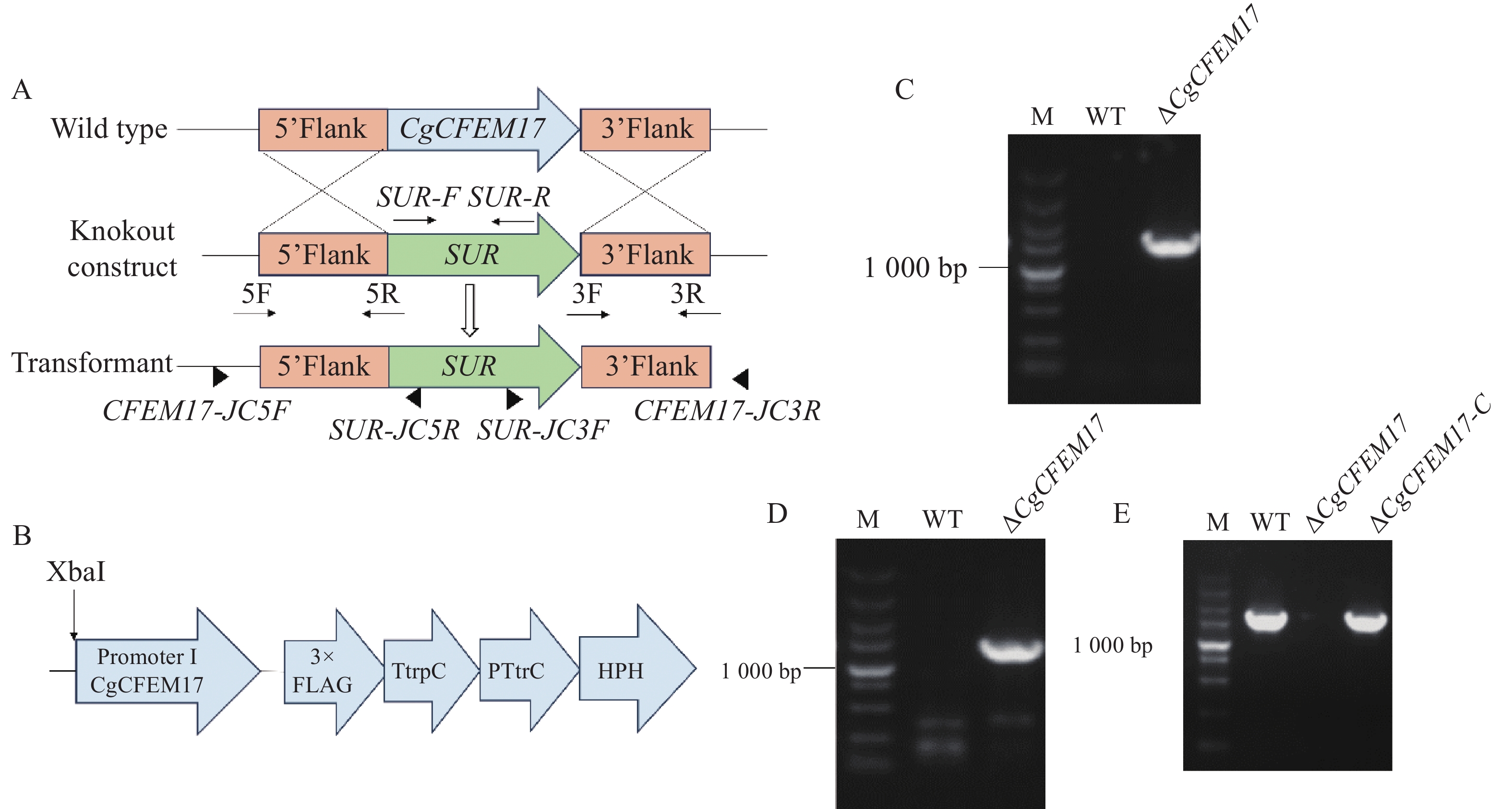
为了研究CgCFEM17的生物学功能,根据同源重组的原理构建CgCFEM17的基因敲除突变菌株(图3−A)。利用PCR技术分别扩增了CgCFEM17的上游同源臂(857 bp)、下游同源臂(851 bp)和氯嘧磺隆抗性基因 (SUR)片段(2 807 bp),用融合PCR的方式得到带有CgCFEM17上游同源臂-SUR-CgCFEM17下游同源臂的重组DNA片段,使用PEG介导法将该重组DNA片段转入胶孢炭疽菌野生型的原生质体中,利用氯嘧磺隆筛选出阳性转化子进行单孢分离,获得纯合株系。通过PCR方法在纯合株系的基因组中使用引物对CFEM17-JC5F/SUR-JC5R可检测到上游片段(图3−C),使用引物对SUR-JC3F/CFEM17-JC3R可检测到下游片段(图3−D),并对上下游片段进行测序与序列比对,结果表明测序序列与预测序列一致,而使用引物对CFEM17-F(XbaI)/CFEM17-R(SmaI)检测不到基因的存在(图3−E),表明CgCFEM17基因已经成功被敲除,并将该基因敲除菌株命名为ΔCgCFEM17。接着构建了由自身启动子驱动的CgCFEM17的回补表达载体(图3−B),将其转化到敲除突变体ΔCgCFEM17后获得抗性转化子进行回补。对抗性转化子的PCR检测结果显示,与野生型相似,在回补菌株中可扩增到与CgCFEM17基因大小一致的片段,进行测序与序列比对分析发现测序序列与预测序列一致,而在ΔCgCFEM17中则扩增不到(图3−E),表明ΔCgCFEM17的回补菌株构建成功,将其命名为ΔCgCFEM17-C。
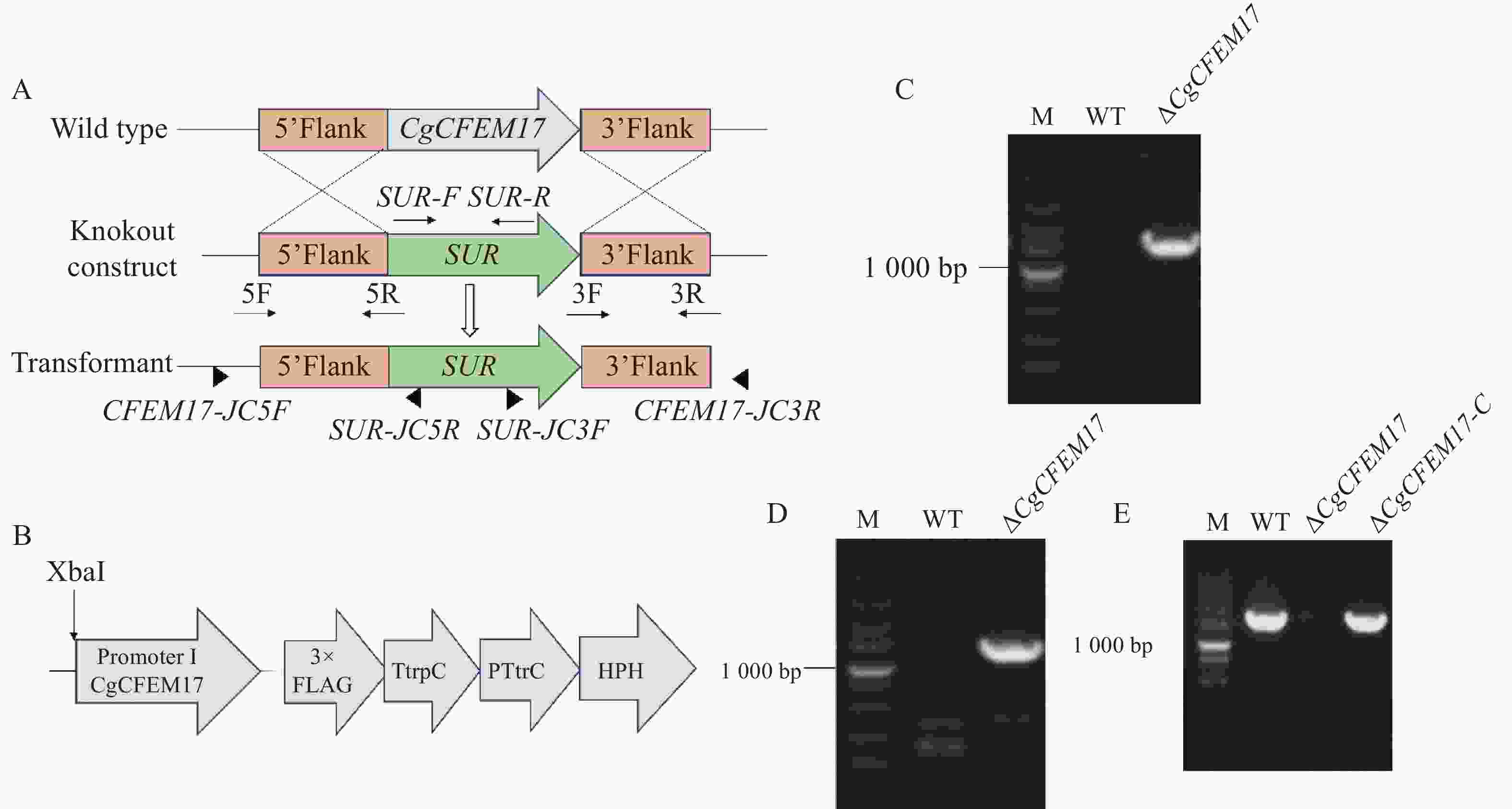
图 3 CgCFEM17敲除突变体的构建与分子鉴定
Figure 3. Construction and molecular characterization of CgCFEM17 knockout mutant strain
-
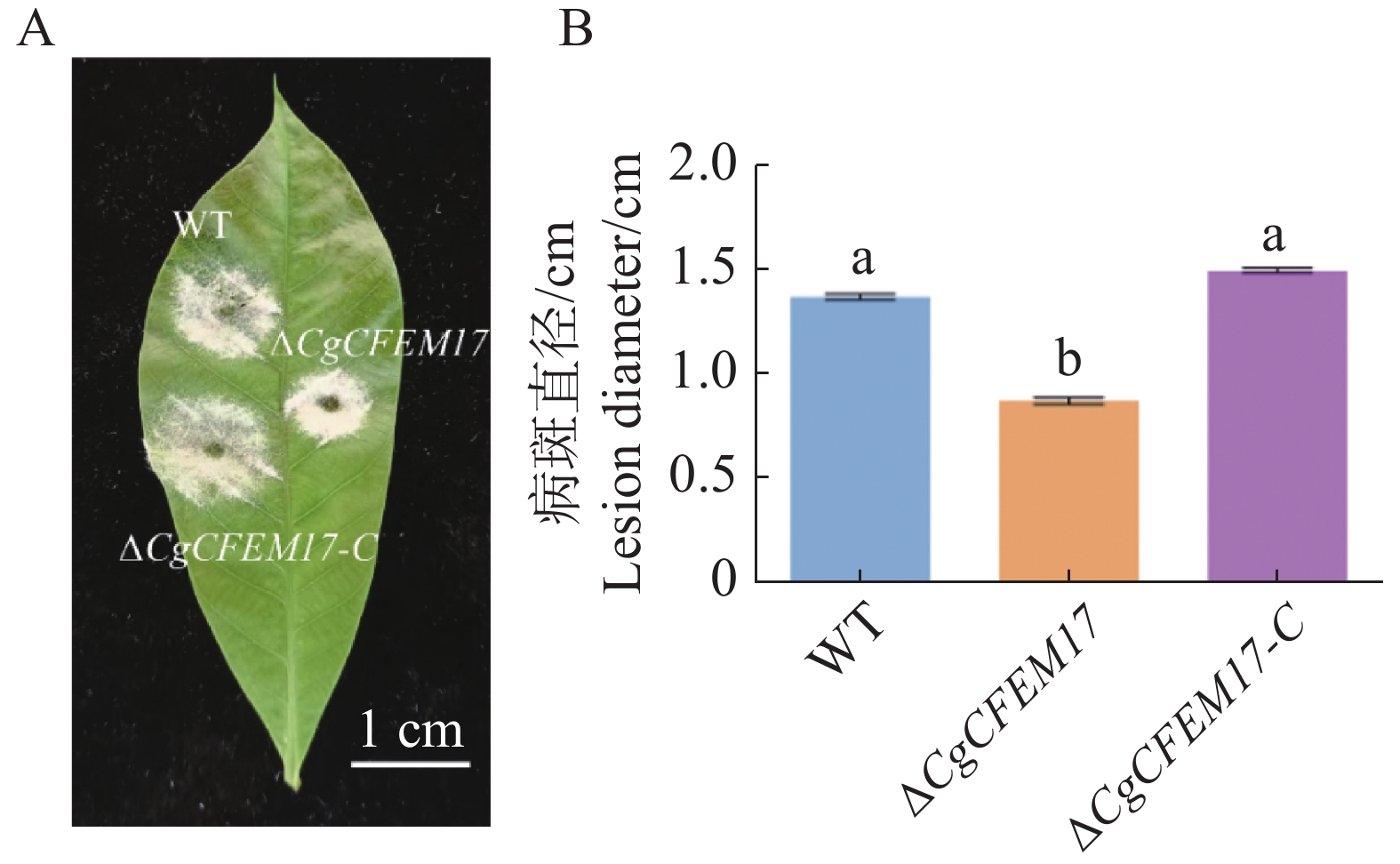
为了探究CgCFEM17对胶孢炭疽菌致病力的影响,利用WT、ΔCgCFEM17、ΔCgCFEM17-C的分生孢子悬浮液分别接种在离体的橡胶树叶片上,3 d后观察到WT、ΔCgCFEM17、ΔCgCFEM17-C在橡胶树叶片上都能够引起典型的病斑(图4−A),对病斑直径进行测量和统计分析的结果显示,ΔCgCFEM17引起的病斑直径显著小于WT和ΔCgCFEM17-C引起的病斑直径(图4−B),结果表明CgCFEM17的缺失导致胶孢炭疽菌对橡胶叶片的致病能力降低,说明CgCFEM17参与调节胶孢炭疽菌对橡胶树的致病力。
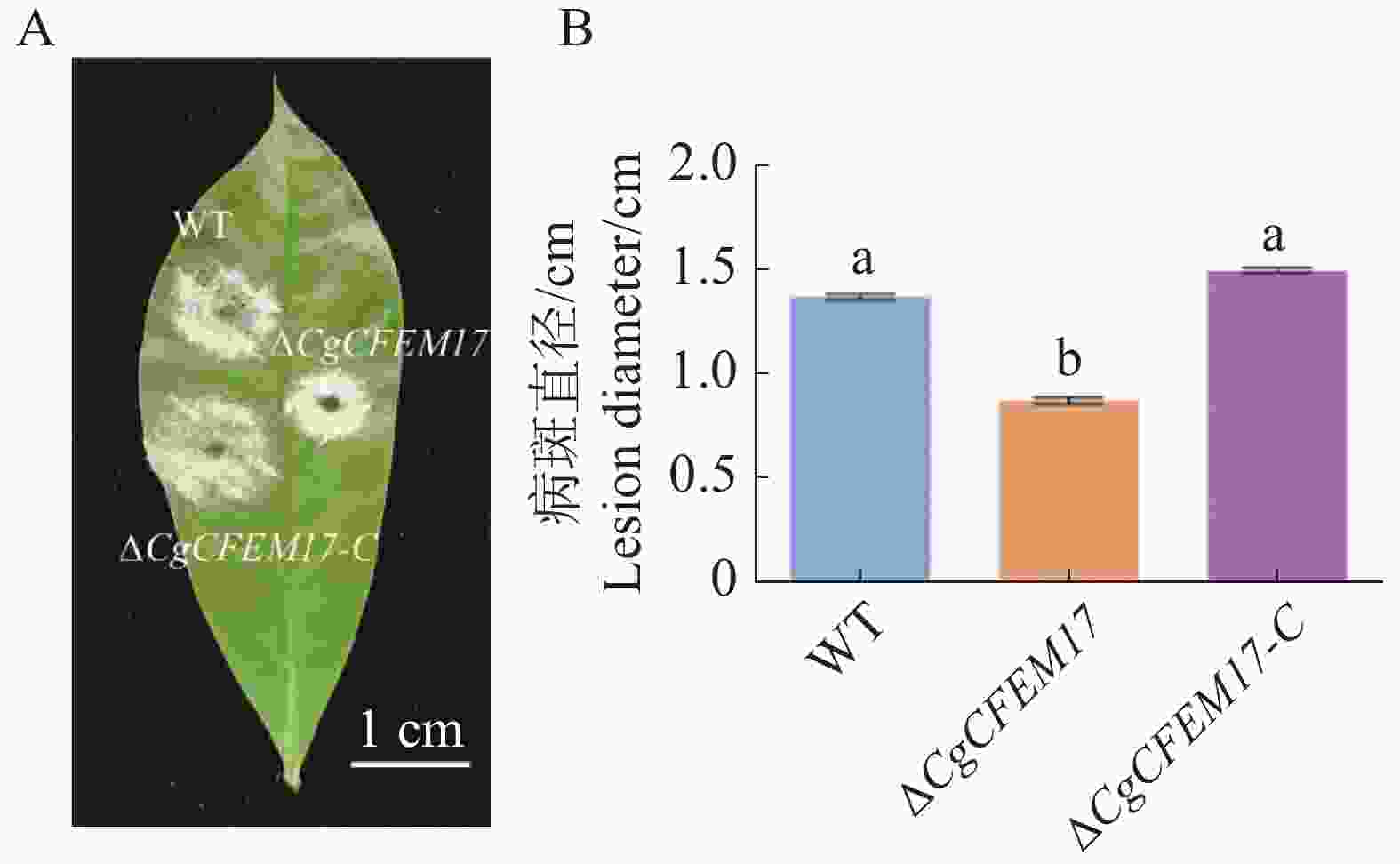
图 4 CgCFEM17对橡胶树叶片的致病性分析
Figure 4. Pathogenicity analysis of CgCFEM17 on rubber tree leaves
-
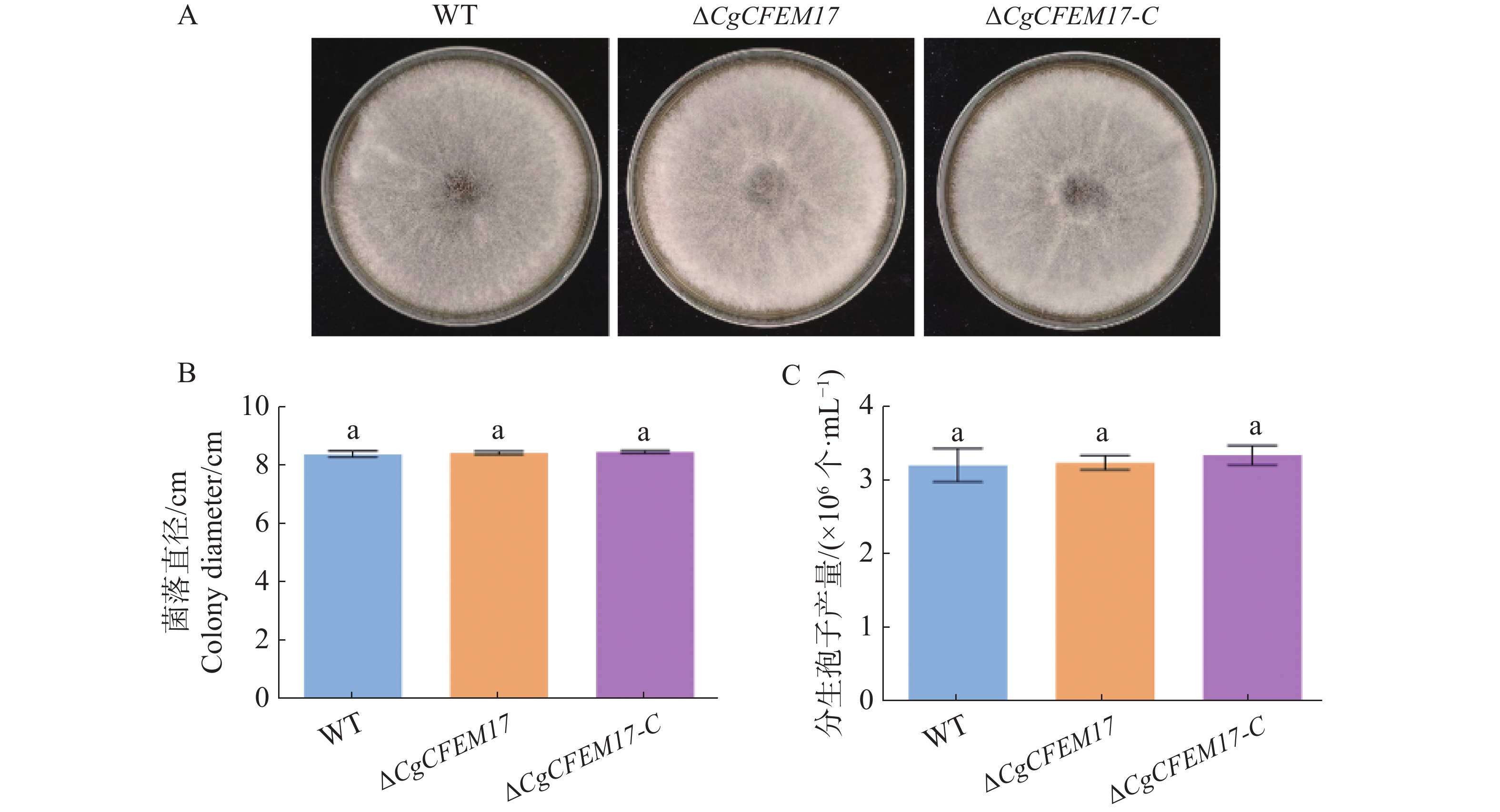
为了探究CgCFEM17是否影响胶孢炭疽菌菌落的生长情况,将WT、ΔCgCFEM17和ΔCgCFEM17-C分别接种于PDA培养基上,5 d后观察到WT、ΔCgCFEM17和ΔCgCFEM17-C的菌落形态没有明显差异(图5−A)。菌落生长直径的统计分析结果显示,WT、ΔCgCFEM17和ΔCgCFEM17-C的菌落直径无显著差异(图5−B)。孢子产量的分析结果显示,WT、ΔCgCFEM17和ΔCgCFEM17-C之间的孢子产量无明显差异(图5−C)。以上结果表明CgCFEM17不参与调控胶孢炭疽菌的菌落生长和产孢能力。
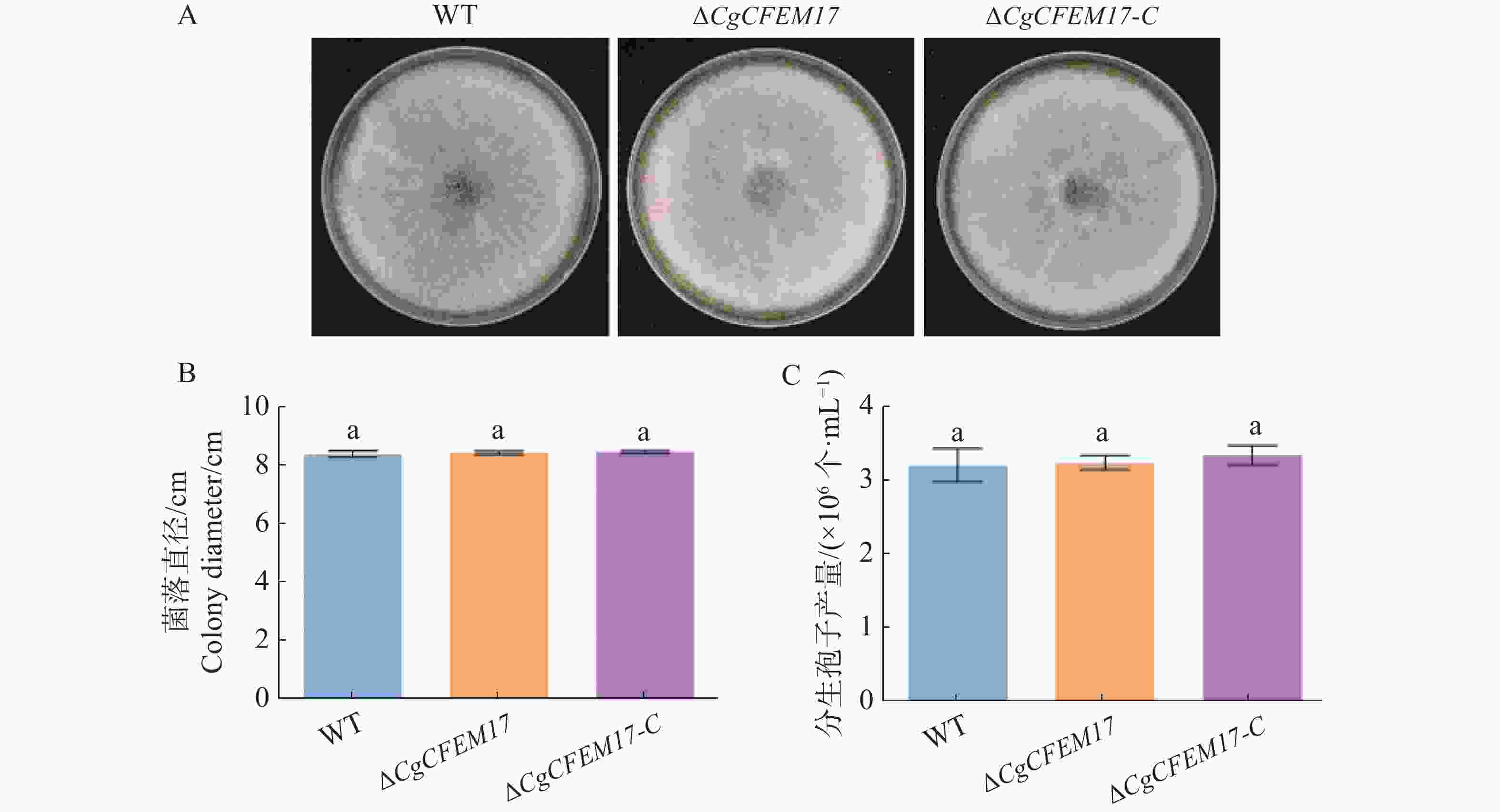
图 5 ∆CgCFEM17的菌落生长及产孢能力分析
Figure 5. Colony growth and conidia production analysis of ∆CgCFEM17
-
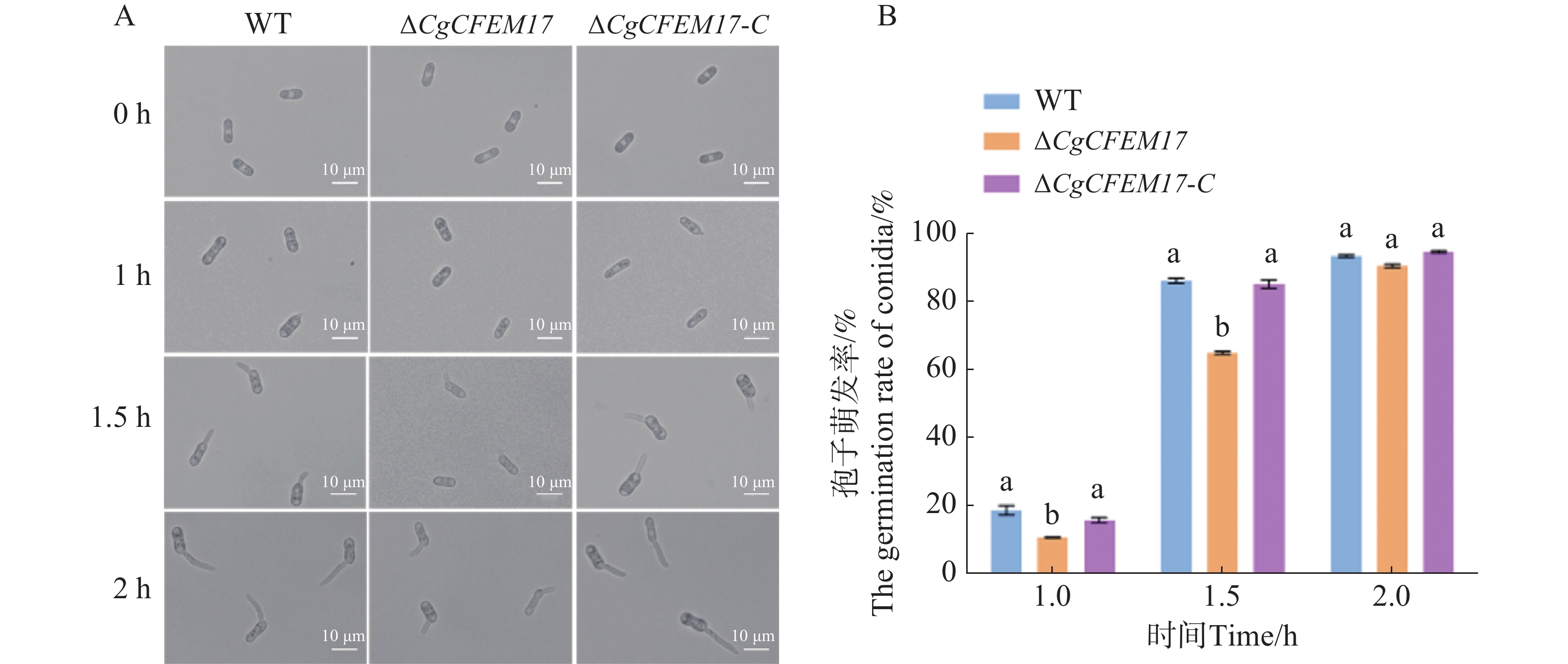
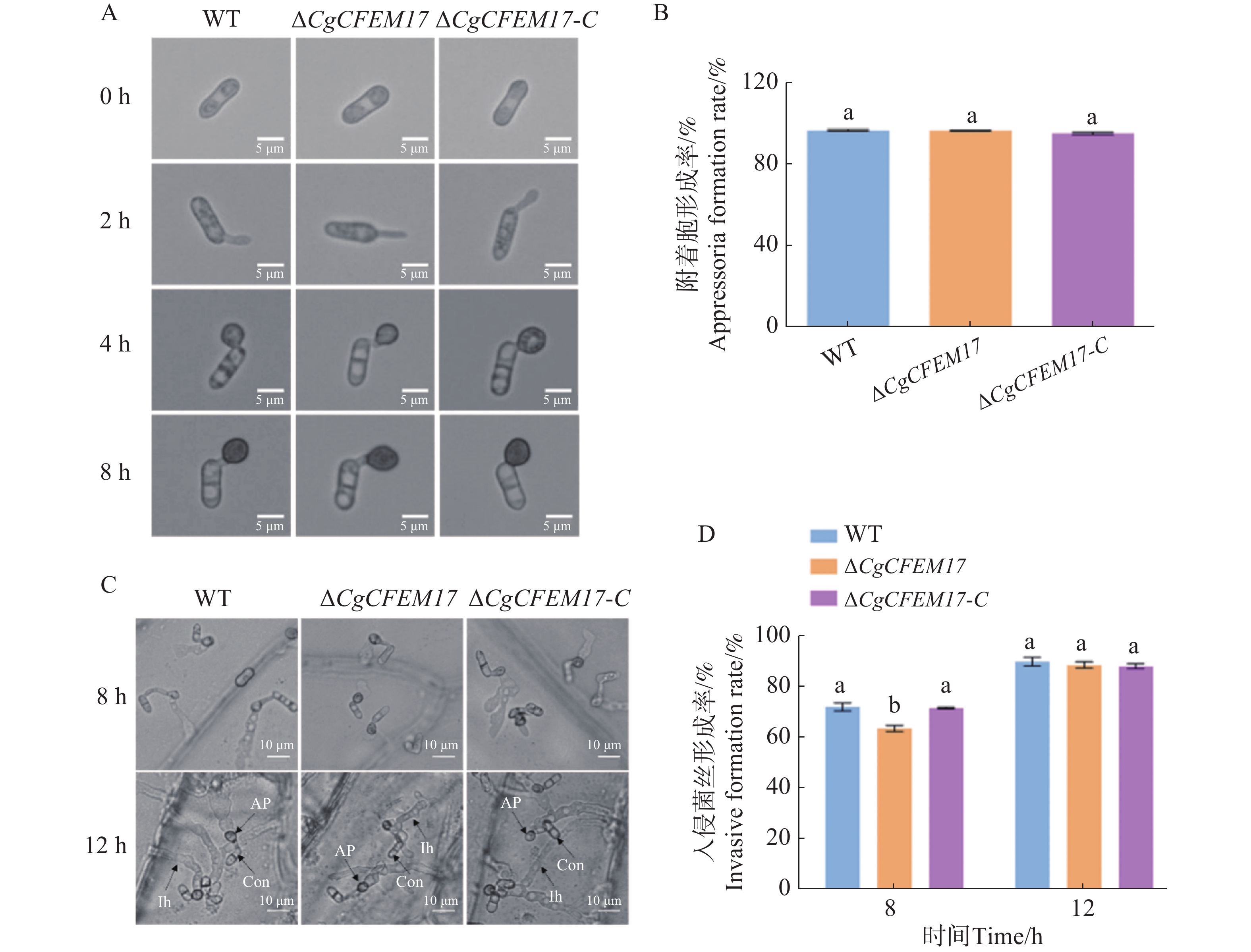
为了探究CgCFEM17是否影响了胶孢炭疽菌分生孢子的萌发,本研究团队分析了WT、ΔCgCFEM17和∆CgCFEM17-C的分生孢子的萌发过程(图6−A)。结果显示,在孢子萌发诱导1和1.5 h时,ΔCgCFEM17的分生孢子萌发率显著低于WT和∆CgCFEM17-C(图6−B);当萌发诱导2 h时,尽管ΔCgCFEM17的芽管长度显著小于WT和∆CgCFEM17-C(图6−A),但ΔCgCFEM17的分生孢子萌发率与WT和∆CgCFEM17-C没有明显差异(图6−B)。以上结果表明,CgCFEM17的缺失导致胶孢炭疽菌分生孢子的萌发迟缓,CgCFEM17参与胶孢炭疽菌的萌发进程。
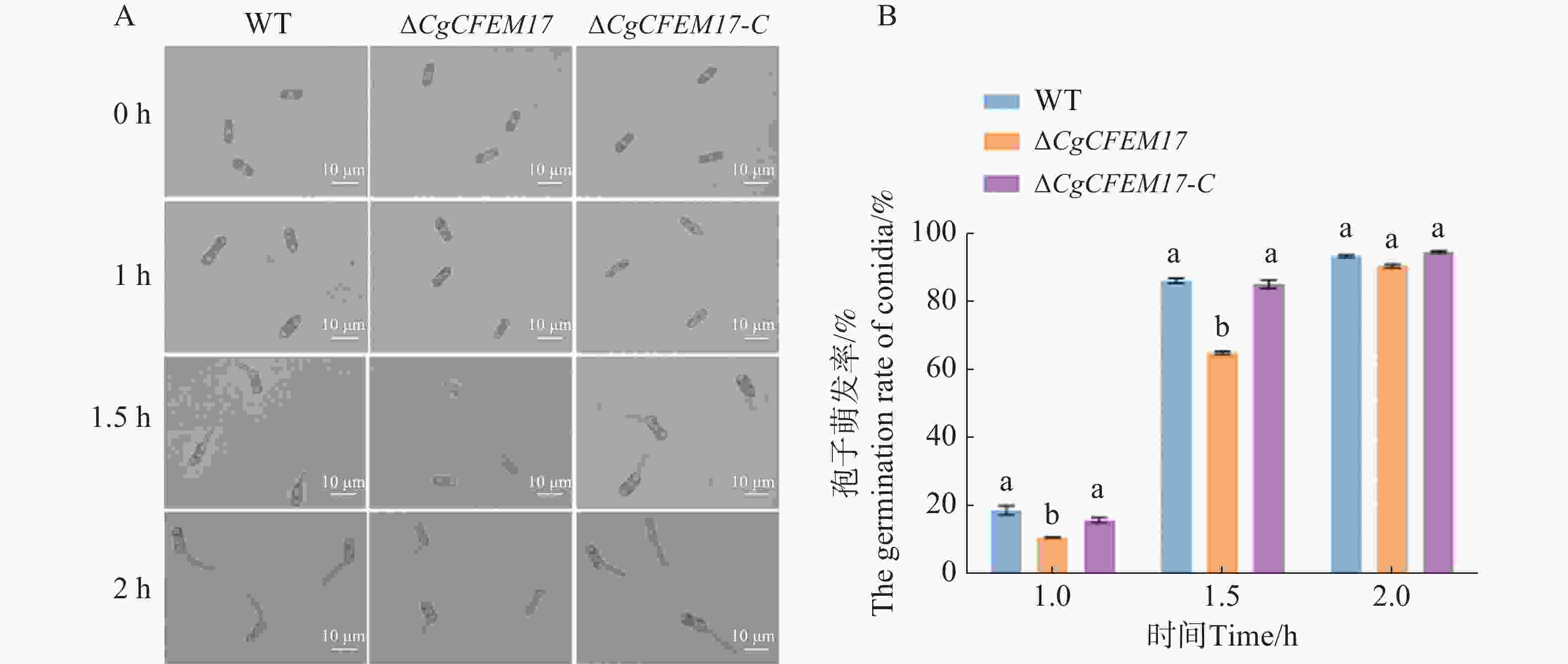
图 6 ∆CgCFEM17的孢子萌发能力分析
Figure 6. The nutritional germination analysis of ∆CgCFEM17
-
为了进一步探究CgCFEM17是否影响胶孢炭疽菌侵染结构的形成,观察并分析分生孢子在疏水表面上附着胞的形成过程和对洋葱表皮的侵染过程,结果表明,在疏水表面上WT、ΔCgCFEM17和ΔCgCFEM17-C的附着胞形态(图7−A)及形成率均没有明显差异(图7−B)。洋葱表皮接种实验结果显示,在侵染过程中ΔCgCFEM17的分生孢子能正常形成附着胞及入侵菌丝(图7−C),但早期入侵菌丝的形成率低于WT和ΔCgCFEM17-C(图7−D)。以上结果表明,CgCFEM17不影响附着胞的形成,但延缓了入侵结构的形成。
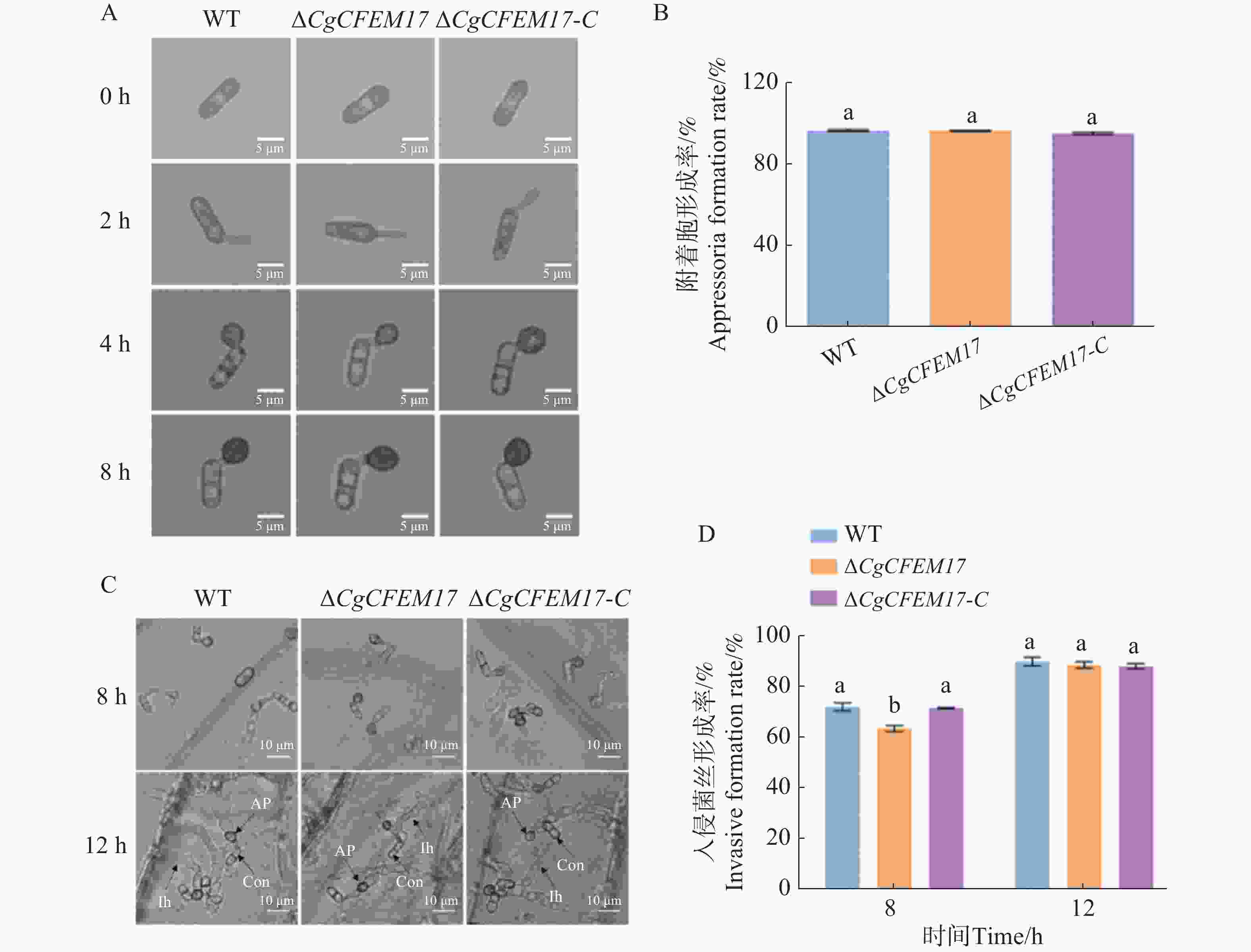
图 7 ∆CgCFEM17的附着胞形成率分析和对洋葱表皮的入侵分析
Figure 7. Analysis of appressorium formation rate of ∆CgCFEM17 and invasion onion epidermis
-
CFEM蛋白是真菌特有的一类蛋白,一般都含有1个信号肽、1个或多个CFEM结构域,有的还含有GPI锚定位点或跨膜结构域[29],其中含有跨膜结构域的CFEM蛋白被定义为Pth11-like类CFEM蛋白[11, 14]。在本研究中,从橡胶树胶孢炭疽菌中扩增到1个编码区长度为1 341bp的CgCFEM17基因,编码的蛋白除了含有N端信号肽和1个保守的CFEM结构域外,还包含 6个跨膜结构域,符合Pth11-like类CFEM蛋白的结构特征。系统进化树分析结果显示,CgCFEM17与稻瘟菌的Pth11、WISH和灰霉菌的CFEM-Bcin 07g03260等蛋白聚在一起,这些蛋白都属于已知功能的Pth11-like型CFEM蛋白[9, 14 − 15],结果说明CgCFEM17属于胶孢炭疽菌的1个Pth11-like型CFEM蛋白。
在植物病原真菌中,Pth11-like型CFEM蛋白参与调控病原真菌的生长和侵染结构的发育。在稻瘟菌中,Pth11的缺失导致附着胞形成率显著降低,但附着胞的形态没有发生变化,说明Pth11参与调控附着胞的分化而不是形态建成[14]。进一步的研究结果证明其中的CFEM 结构域是附着胞正常发育和致病性所必需[11],而另一个Pth11-like型CFEM蛋白WISH除了影响附着胞形态发生外还影响细胞壁的完整性。在灰霉菌中,Pth11-like型CFEM蛋白BCIN 07g03260则影响病菌分生孢子萌发率和芽管伸长[15]。在杨树炭疽病菌中,Pth11同源蛋白CgPth11参与附着胞形成的调控,但并不是病菌致病力所必需[26]。本研究结果显示,橡胶树胶孢炭疽菌CgCFEM17的缺失引起胶孢炭疽菌对橡胶树致病力降低,进一步研究发现CgCFEM17的缺失延缓了孢子萌发和入侵结构的形成,但对于病菌的菌落生长速率和产孢能力,以及附着胞的形成与形态发生没有影响,暗示CgCFEM17通过调控分生孢子萌发时间进而影响侵染结构形成时间来影响胶孢炭疽菌对橡胶树的致病力。以上研究结果说明不同植物病原真菌的Pth11-like型CFEM蛋白的功能各不相同,通过不同的作用方式和途径在病菌的致病过程中发挥作用。
Pth11-like类CFEM蛋白大多数是丝状子囊菌特有的G蛋白偶联受体(G protein-coupled receptors, GPCRs),含有典型的7个跨膜结构域,在对外界信号识别中扮演重要的角色。分生孢子萌发和附着胞形成是病原菌成功入侵寄主的关键,受营养条件和疏水表面的调节 [30]。稻瘟菌中的研究发现,可溶性植物角质单体和疏水表面是附着胞形成所必需[31]。作为典型的GPCR,Pth11被认为是环磷酸腺苷(cAMP)途径上游的一个表面传感器,该基因的缺失导致病菌在角质素单体和疏水表面条件下的附着胞形成受损[14]。稻瘟菌的另外一个GPCR蛋白WISH也参与了寄主表面信号识别,能够感知多种细胞外信号,调控附着胞的分化[32]。本研究中,CgCFEM17基因缺失突变体的分生孢子萌发和侵染结构形成与野生型菌株相比均表现出延迟发生,但附着胞的形成在疏水表面上不受影响,这种表现是否与植物表面的信号识别有关,还需要进一步的研究。
胶孢炭疽菌Pth11-like型CFEM蛋白CgCFEM17的功能分析
DOI: 10.15886/j.cnki.rdswxb.20250009
 CSTR: 32425.14.j.cnki.rdswxb.20250009
CSTR: 32425.14.j.cnki.rdswxb.20250009
Functional analysis of the Pth11-like CFEM protein CgCFEM17 in Colletotrichum gloeosporioides from rubber tree
-
摘要: 胶孢炭疽菌(Colletotrichum gloeosporioides)是引起橡胶树炭疽病的主要病原。CFEM(common in fungal extracellular membrane)结构域是真菌特有的基序,含有CFEM和跨膜结构域的蛋白属于Pth11-like型CFEM蛋白,在调控病原真菌生长发育和致病力中发挥重要作用。为了研究橡胶树胶孢炭疽菌的致病机制,从胶孢炭疽菌转录组中筛选出1个编码Pth11-like型CFEM蛋白的基因,将其命名为CgCFEM17。该基因的编码区全长1 341 bp,编码446个氨基酸。生物信息学分析显示,CgCFEM17蛋白N端具有长度为26个氨基酸的信号肽和1个保守的CFEM结构域,C端包含6个跨膜结构域。为了分析CgCFEM17基因的功能,构建了该基因的敲除突变体(ΔCgCFEM17)及回补突变体(ΔCgCFEM17-C),并对其生长发育和致病性表型进行了分析。结果表明,与野生型和回补突变体相比,ΔCgCFEM17的分生孢子萌发和入侵结构的形成延迟,致病力减弱,但菌落生长速率、产孢能力及附着胞的形成率与形态发生没有变化。CgCFEM17可能通过调控分生孢子萌发进程,延缓侵染结构形成,影响胶孢炭疽菌对橡胶树的致病力,这为探究胶孢炭疽菌的致病机制及建立新的橡胶树炭疽病防治策略奠定了一定的理论基础。Abstract: Colletotrichum gloeosporioides is the main pathogen causing anthracnose of rubber tree (Hevea brasiliensis). The common in fungal extracellular membrane (CFEM) domain is a unique motif of fungi, and the proteins containing CFEM and transmembrane domains are defined as Pth11-like CFEM protein, which plays important roles in regulating the development and pathogenicity of pathogenic fungi. In order to study the pathogenesis of C. gloeosporiorides, a gene encoding Pth11-like CFEM protein was screened from the transcriptome of C. gloeospriorides and named as CgCFEM17. The encoding region of the gene CgCFEM17 is 1341bp in length and encodes a peptide with 446 amino acids. Bioinformatics analysis showed that the N-terminus of CgCFEM17 has a signal peptide with 26 amino acids and a conserved CFEM domain, and the C-terminus contained six transmembrane domains. The knockout mutant strain (∆CgCFEM17) and complementary strain (∆CgCFEM17-C) of the gene CgCFEM17 were constructed to analyze the function of CgCFEM17. The results showed that the conidium germination and invasion structure of ∆CgCFEM17 were delayed compared with the wild type and complementary strain, but the colony growth rate, conidiation, appressorium formation rate, and morphogenesis were not changed. These results indicated that the gene CgCFEM17 could affect the pathogenicity of C. gloeosporioides on rubber tree by regulating the germination process of conidia and delaying the formation of infection structure, which lays a theoretical foundation for exploring of the pathogenic mechanism of C. gloeosporioides and establishing of a new control strategy for anthracnose of rubber trees.
-
Key words:
- rubber tree /
- Colletotrichum gloeosporioides /
- CgCFEM17 /
- functional analysis
-
图 2 胶孢炭疽菌CgCFEM17与其他病原真菌的CFEM类蛋白的氨基酸序列的系统发育关系
Fig. 2 Phylogenetic relationship of the amino acid sequence between CgCFEM17 andCFEM protein of other pathogenic fungi
图 3 CgCFEM17敲除突变体的构建与分子鉴定
A.CgCFEM17的基因重组敲除原理;B.ΔCgCFEM17-C的回补原理;C.上游片段的PCR检测;D.下游片段PCR检测;E. CgCFEM17基因的PCR检测;M.DL 5 000 bp DNA Maker。
Fig. 3 Construction and molecular characterization of CgCFEM17 knockout mutant strain
A.The principle of genetic recombination knockout of CgCFEM17; B.The complement principle of ∆CgCFEM17-C; C.The PCR detection of the upstream fragment; D.The PCR detection of the downstream fragment downstream; E.The PCR detection of CgCFEM17 gene; M.DL 5 000 bp DNA Maker.
图 4 CgCFEM17对橡胶树叶片的致病性分析
A.病斑症状图,标尺为1 cm;B.病斑直径统计分析图。不同的小写字母表示差异显著(P<0.05)。
Fig. 4 Pathogenicity analysis of CgCFEM17 on rubber tree leaves
A.Disease symptom, scale bar=1 cm; B.Statistical analysis of the disease lesion. Different letters indicate significant differences(P<0.05)
图 5 ∆CgCFEM17的菌落生长及产孢能力分析
A.WT、∆CgCFEM17及∆CgCFEM17-C在葡萄糖琼脂(PDA)培养基上培养6 d的菌落形态;B.对WT、∆CgCFEM17及∆CgCFEM17-C 的菌落直径进行统计分析;C.WT、∆CgCFEM17及∆CgCFEM17-C 的孢子产量分析。不同的小写字母表示差异显著(P<0.05)。
Fig. 5 Colony growth and conidia production analysis of ∆CgCFEM17
A.Colony growth of ∆CgCFEM17, ∆CgCFEM17 and ∆CgCFEM17-C on PDA for 6 days; B.Statistical analysis of colony diameter of WT, ∆CgCFEM17 and ∆CgCFEM17-C; C.The analysis of conidia productivity of WT, ∆CgCFEM17 and ∆CgCFEM17-C. Different letters indicate significant differences(P<0.05).
图 6 ∆CgCFEM17的孢子萌发能力分析
A.WT、∆CgCFEM17及的形态观察;B.对WT、∆CgCFEM17及∆CgCFEM17-C 的萌发率进行统计分析。不同的小写字母表示差异显著(P<0.05)。
Fig. 6 The nutritional germination analysis of ∆CgCFEM17
A.Morphological observation of ∆CgCFEM17, ∆CgCFEM17 and∆CgCFEM17-C; B.Statistical analysis of germination rate of WT, ∆CgCFEM17 and ∆CgCFEM17-C; C.Statistical analysis of germ tube length of WT, ∆CgCFEM17 and ∆CgCFEM17-C at 2 hours. Different letters indicate significant differences(P<0.05).
图 7 ∆CgCFEM17的附着胞形成率分析和对洋葱表皮的入侵分析
A.WT、∆CgCFEM17及∆CgCFEM17-C在疏水表面附着胞形成过程(标尺为5 μm);B.对WT、∆CgCFEM17及∆CgCFEM17-C 的附着胞形成率进行统计分析;C.WT、ΔCgCFEM17 和∆CgCFEM17-C入侵菌丝形态观察;D.WT、ΔCgCFEM17和∆CgCFEM17-C的入侵菌丝形成率统计分析(标尺为20 μm)。Con.孢子; AP.附着胞;Ih.入侵菌丝。不同的小写字母表示差异显著(P<0.05)。
Fig. 7 Analysis of appressorium formation rate of ∆CgCFEM17 and invasion onion epidermis
A.The process of appressorium formation of WT, ΔCgCFEM17 and ∆CgCFEM17-C on hydrophobic surfaces(scale bar=5 μm); B.Statistical analysis of germination rates of WT, ∆CgCFEM17 and ∆CgCFEM17-C; C.Statistical analysis of appressorium formation rates of WT, ∆CgCFEM17 and ∆CgCFEM17-C; D.Observation of the hyphal morphology of WT, ∆CgCFEM17 and ∆CgCFEM17-C(scale bar=20 μm); E.Statistical analysis of invasive hyphae formation of WT, ∆CgCFEM17 and ∆CgCFEM17-C. Con.Conidium; AP.Appressorium; Ih.invasive hyphae. Different letters indicate significant differences (P<0.05).
表 1 引物信息
Table 1 Primer information
引物名称Primer 引物序列 Primer sequence(5′-3′) CFEM17-5F CGACGTGTAGAGCTATGACC CFEM17-M5F CTTGTGCGTTTCAAGAAGTGCCAACGCCACAGTG CFEM17-M5R CACTGTGGCGTTGGCACTTCTTGAAACGCACAAG CFEM17-M3F GAATTGCATGCTCTCACGCGTTTGAGTTTTGTTT CFEM17-M3R AAACAAAACTCAAACGCGTGAGAGCATGCAATTC CFEM17-3R TTTGTCATTCTGAAGTCCC SUR-SLR ATGTTGGCATAAGCCGAACCGT SUR-SLF CCTCTGATATTGGAAGCGACGC CFEM17-JC5F GCCGCTCGCATTTCATTCC SUR-JC5R GCGTTTGTAACTCTGCCTGTTTG SUR-JC3F ACGAGGACCGCTACTCACATAC CFEM17-JC3R ACACCCCAAAAGGTCCCAC CFEM17-F(XbaI) TCTAGAATGTTCACCAACGCCAGAAA CFEM17-R(SmaI) CCCGGGTACTTTATTTGAATGGCCAA CFEM17-OF(XbaI) TCTAGACATAACTGATGCGACACCT  下载: 导出CSV
下载: 导出CSV
-
[1] 张春霞, 何明霞, 李加智, 等. 云南西双版纳地区橡胶炭疽病病原鉴定[J]. 植物保护, 2008, 34(1): 103 − 106. doi: 10.3969/j.issn.0529-1542.2008.01.024 [2] 赵晓宇, 王记圆, 廖小淼, 等. 橡胶树炭疽菌蛋白激酶A催化亚基基因的克隆及表达[J]. 基因组学与应用生物学, 2019, 38(8): 3622 − 3628. doi: 10.13417/j.gab.038.003622 [3] 林春花, 孙董董, 韩丹, 等. 中国橡胶树苗圃2种炭疽病菌分子鉴定及分布分析[J]. 热带作物学报, 2014, 35(9): 1802 − 1808. doi: 10.3969/j.issn.1000-2561.2014.09.024 [4] 李文杨, 郑春耀, 李超萍, 等. 中国橡胶树主栽品系和部分种质对尖孢炭疽病的室内抗性评价[J]. 热带农业工程, 2009, 33(5): 31 − 36. [5] 郭云峰, 安邦. 橡胶树胶孢炭疽菌NADPH氧化酶功能研究[J]. 生物技术通报, 2018, 34(10): 165 − 171. doi: 10.13560/j.cnki.biotech.bull.1985.2018-0344 [6] FENG L P, DONG M X, HUANG Z R, et al. CgCFEM1 is required for the full virulence of Colletotrichum gloeosporioides[J]. International Journal of Molecular Sciences, 2024, 25(5): 2937. doi: 10.3390/ijms25052937 [7] 李娴, 陈思思, 谢剑波. 杨树响应胶孢炭疽菌侵染的转录组学研究[J]. 北京林业大学学报, 2024, 46(4): 91 − 102. doi: 10.12171/j.1000-1522.20210481 [8] ZHANG Z N, WU Q Y, ZHANG G Z, et al. Systematic analyses reveal uniqueness and origin of the CFEM domain in fungi[J]. Scientific Reports, 2015, 5: 13032. doi: 10.1038/srep13032 [9] KULKARNI R D, THON M R, PAN H Q, et al. Novel G-protein-coupled receptor-like proteins in the plant pathogenic fungus Magnaporthe grisea[J]. Genome Biology, 2005, 6(3): R24. doi: 10.1186/gb-2005-6-3-r24 [10] 谢薇. 稻瘟病菌诱导植物细胞死亡基因MoCFEM的功能分析[D]. 福州: 福建农林大学, 2015. [11] KOU Y J, TAN Y H, RAMANUJAM R, et al. Structure-function analyses of the Pth11 receptor reveal an important role for CFEM motif and redox regulation in rice blast[J]. New Phytologist, 2017, 214(1): 330 − 342. doi: 10.1111/nph.14347 [12] CAI N, LIU R, YAN D Z, et al. Bioinformatics analysis and functional characterization of the CFEM proteins of Metarhizium anisopliae[J]. Journal of Fungi, 2022, 8(7): 661. doi: 10.3390/jof8070661 [13] ZUO N, BAI W Z, WEI W Q, et al. Fungal CFEM effectors negatively regulate a maize wall-associated kinase by interacting with its alternatively spliced variant to dampen resistance[J]. Cell Reports, 2022, 41(13): 111877. doi: 10.1016/j.celrep.2022.111877 [14] DEZWAAN T M, CARROLL A M, VALENT B, et al. Magnaporthe grisea pth11p is a novel plasma membrane protein that mediates appressorium differentiation in response to inductive substrate cues[J]. The Plant Cell, 1999, 11(10): 2013 − 2030. doi: 10.1105/tpc.11.10.2013 [15] ARYA G C, SRIVASTAVA D A, PANDARANAYAKA E P J, et al. Characterization of the role of a non-GPCR membrane-bound CFEM protein in the pathogenicity and germination of Botrytis cinerea[J]. Microorganisms, 2020, 8(7): 1043. doi: 10.3390/microorganisms8071043 [16] CHEN L Q, WANG H Y, YANG J H, et al. Bioinformatics and transcriptome analysis of CFEM proteins in Fusarium graminearum[J]. Journal of Fungi, 2021, 7(10): 871. doi: 10.3390/jof7100871 [17] QIAN Y L, ZHENG X Y, WANG X Y, et al. Systematic identification and functional characterization of the CFEM proteins in poplar fungus Marssonina brunnea[J]. Frontiers in Cellular and Infection Microbiology, 2022, 12: 1045615. doi: 10.3389/fcimb.2022.1045615 [18] LIANG F, LIU L J, LI C S, et al. Systematic identification and functional characterization of the CFEM proteins in fishscale bamboo rhombic-spot pathogen Neostagonosporella sichuanensis[J]. Frontiers in Plant Science, 2024, 15: 1396273. doi: 10.3389/fpls.2024.1396273 [19] GRUBER S, OMANN M, ZEILINGER S. Comparative analysis of the repertoire of G protein-coupled receptors of three species of the fungal genus Trichoderma[J]. BMC Microbiology, 2013, 13: 108. doi: 10.1186/1471-2180-13-108 [20] DILKS T, HALSEY K, DE VOS R P, et al. Non-canonical fungal G-protein coupled receptors promote Fusarium head blight on wheat[J]. PLoS Pathogens, 2019, 15(4): e1007666. doi: 10.1371/journal.ppat.1007666 [21] WANG J X, LONG F, ZHU H, et al. Bioinformatic analysis and functional characterization of CFEM proteins in Setosphaeria turcica[J]. Journal of Integrative Agriculture, 2021, 20(9): 2438 − 2449. doi: 10.1016/S2095-3119(20)63342-1 [22] SHANG S P, LIU G L, ZHANG S, et al. A fungal CFEM-containing effector targets NPR1 regulator NIMIN2 to suppress plant immunity[J]. Plant Biotechnology Journal, 2024, 22(1): 82 − 97. doi: 10.1111/pbi.14166 [23] 刘思珍, 欧阳超, 满益龙, 等. 辣椒胶孢炭疽菌CFEM效应因子鉴定及转录组分析[J]. 南京农业大学学报, 2021, 44(6): 1074 − 1082. doi: 10.7685/jnau.202102013 [24] 张丽勍, 段可, 邹小花, 等. 草莓胶孢炭疽菌CFEM候选效应子的生物信息学鉴定及其侵染过程中的转录分析[J]. 植物保护, 2017, 43(5): 43 − 51. doi: 10.3969/j.issn.0529-1542.2017.05.006 [25] 井忠英. 玉米炭疽病菌CFEM效应子的系统鉴定与功能分析[D]. 北京: 中国农业科学院, 2015. [26] WANG X L, LU D X, TIAN C M. Mucin Msb2 cooperates with the transmembrane protein Sho1 in various plant surface signal sensing and pathogenic processes in the poplar anthracnose fungus Colletotrichum gloeosporioides[J]. Molecular Plant Pathology, 2021, 22(12): 1553 − 1573. doi: 10.1111/mpp.13126 [27] XU X H, LI G P, LI L, et al. Genome-wide comparative analysis of putative Pth11-related G protein-coupled receptors in fungi belonging to Pezizomycotina[J]. BMC Microbiology, 2017, 17(1): 166. doi: 10.1186/s12866-017-1076-5 [28] WANG Q N, AN B, HOU X R, et al. Dicer-like proteins regulate the growth, conidiation, and pathogenicity of Colletotrichum gloeosporioides from Hevea brasiliensis[J]. Frontiers in Microbiology, 2018, 8: 2621. doi: 10.3389/fmicb.2017.02621 [29] PENG J B, WU L N, ZHANG W, et al. Systemic identification and functional characterization of common in fungal extracellular membrane proteins in Lasiodiplodia theobromae[J]. Frontiers in Plant Science, 2021, 12: 804696. doi: 10.3389/fpls.2021.804696 [30] SCHAMBER A, LEROCH M, DIWO J, et al. The role of mitogen-activated protein (MAP) kinase signalling components and the Ste12 transcription factor in germination and pathogenicity of Botrytis cinerea[J]. Molecular Plant Pathology, 2010, 11(1): 105 − 119. doi: 10.1111/j.1364-3703.2009.00579.x [31] LIU W D, ZHOU X Y, LI G T, et al. Multiple plant surface signals are sensed by different mechanisms in the rice blast fungus for appressorium formation[J]. PLoS Pathogens, 2011, 7(1): e1001261. doi: 10.1371/journal.ppat.1001261 [32] SABNAM N, ROY BARMAN S. WISH, a novel CFEM GPCR is indispensable for surface sensing, asexual and pathogenic differentiation in rice blast fungus[J]. Fungal Genetics and Biology, 2017, 105: 37 − 51. doi: 10.1016/j.fgb.2017.05.006 -

 点击查看大图
点击查看大图
计量
- 文章访问数: 1255
- HTML全文浏览量: 653
- PDF下载量: 22
- 被引次数: 0
