

-
植物病原真菌炭疽菌属(Colletotrichum)具有寄主范围广和全球性分布的特点,是农业生产中极具威胁的病原菌类群[1]。胶孢炭疽菌(Colletotrichum gloeosporioides))能够侵染橡胶树(Hevea brasiliensis),引发橡胶树炭疽病,严重时可导致橡胶产量锐减和胶乳品质下降,对国家天然橡胶安全构成严峻挑战[2−3]。因此,对胶孢炭疽菌致病相关基因进行深入挖掘与鉴定,系统解析其生物学功能,并在分子层面揭示其复杂的调控网络与作用靶点,不仅具有重要的理论价值,更能为开发新型高效靶向杀菌剂及培育抗病种质提供关键的理论基础。胶孢炭疽菌的侵染是典型的“半活体营养型”(hemibiotrophic)生活史,分为活体营养和死体营养两个典型的侵染阶段[4]。在侵染初期,胶孢炭疽菌的分生孢子(conidia)会先在橡胶树叶片表面附着。在适宜的条件下孢子开始萌发并形成芽管(germ tube),随后芽管的顶端分化形成一种特化的侵染结构附着胞(appressorium)。成熟的附着胞会在其附着叶片的一侧形成侵染钉,并通过其侵入叶片表皮,进而形成入侵菌丝。因此,附着胞的形成是能否成功侵染寄主植物的关键[5]。经过一段潜伏期后,进入死体营养阶段,入侵菌丝生长并分泌毒素,导致细胞坏死出现典型的炭疽病病斑[6]。另一方面,植物为了应对微生物病原菌的侵染,已经进化出复杂的免疫系统来抵御病原菌的侵染[7−8]。当病原菌侵染植物时,植物免疫受体能识别保守的病原体相关分子模式(pathogen-associated molecular patterns,PAMPs),并迅速启动被称为模式触发免疫(Pattern-Triggered Immunity,PTI)的基础免疫反应[9]。为了应对植物的基础免疫反应,病原菌会分泌被称为效应蛋白的毒性蛋白来干扰植物免疫[10]。植物也进化出识别这些效应蛋白的识别蛋白,识别后会快速激活以过敏反应(Hypersensitive Response,HR)为特征的效应子触发免疫(Effector-Triggered Immunity,ETI)[11−12]。
胶孢炭疽菌附着胞的形成是能否侵染成功的关键。在稻瘟菌(Magnaporthe oryzae)中,假定效应蛋白MGG_03436.5在附着胞时期表达量显著上调[13];在苹果炭疽菌(C. gloeosporioides)中,通过转录组分析筛选到39个候选效应蛋白基因在附着胞时期表达量上调,其中效应蛋白Sntf2能够抑制植物免疫反应[14];在稻瘟菌(M. oryzae)中,发现多个假定效应蛋白基因(MoLLEs)在附着胞形成时期表达上调[15],因此,推测候选效应蛋白在附着胞时期表达量上调可能与胶孢炭疽菌的侵染结构形成有关。本研究团队在前期研究中预测到Cg694和Cg2346为胶孢炭疽菌的潜在效应蛋白,并通过对胶孢炭疽菌不同发育阶段的转录组数据进行分析,发现其在胶孢炭疽菌成熟附着胞中高水平表达,因此推测,Cg694和Cg2346可能与胶孢炭疽菌的侵染结构形成及致病力密切相关。通过信息学分析,Cg694被预测属于主要协助转运蛋白超家族(major facilitator superfamily transporter,MFS),而Cg2346是功能未知的蛋白。MFS转运蛋白是最大的次级膜转运蛋白家族,广泛存在于细菌和真菌中[16],可以分为共转运体、反转运体和促进转运体,介导如糖类和代谢物等多种底物的跨膜转运[17]。在交链格孢菌(Alternaria alternata)中,由AaMFS19基因编码一种MFS蛋白,该MFS蛋白是交链格孢菌抗氧化应激和抗杀菌剂所必需的[18];在白色念珠菌(Candida albicans)中,名为FLU1的MFS蛋白与细胞对氟康唑(Fluconazole)的耐药性相关[19];在胶孢炭疽菌中,鉴定到的一个MFS蛋白CgMFS1被证明与真菌的糖转运、抗氧化应激和致病性相关[20]。本研究通过构建基因缺失突变体并对其生理表型以及其在淡绿期橡胶树叶片的致病能力进行了分析,初步探究了Cg694和Cg2346在橡胶树胶孢炭疽菌中的功能,旨在为炭疽病害的防治提供理论依据。
-
供试的野生型菌株材料由本实验室提供。本实验室已分离和鉴定橡胶树胶孢炭疽菌野生型菌株(C. gloeosporioides Wild Type,WT),并完成该菌株的基因组测序和数据库的建立。
本实验使用的PCR引物由铂尚生物公司和睿博兴科公司负责合成(表1)。DNA测序工作由睿博兴科公司负责完成。大肠杆菌(Escherichia coli)感受态细胞TOP10购于上海吐露港生物科技有限公司。Spark HiFi Seamless Cloning Kit(单片段)试剂盒购于山东思科捷公司。GV3101农杆菌感受态购于Takara Bio宝日医生物技术(北京)有限公司。RNA prep Pure Plant Kit试剂盒购于北京天根生物技术有限公司。SPARKscript Ⅱ All-in-one RT SuperMix for qPCR(With gDNA Eraser)试剂盒购于山东思科捷公司。pEGAD-GFP及pBS-SUR载体由本实验室改造。本氏烟草(Nicotiana bentha miana)材料由本实验室储存种子和种植培养,在气候室培养6 周(25℃,每天光照10 h)。
表 1 本实验所用引物
Table 1. Primers used in this experiment
引物名称Primer 序列(5′−3′)Sequence 用途Usage Cg694-5F CTATTGCTTGTCTTGTTCCT 同源臂扩增
Homologous arm amplificationCg694-5R AGGCAGCGTATACCGACCTG 同源臂扩增
Homologous arm amplificationCg694-MF1 GACGCAATCTTCTACAGAACGTGCCAACGCCACAGTGCCC 同源臂扩增
Homologous arm amplificationCg694-MR1 GGGCACTGTGGCGTTGGCACGTTCTGTAGAAGATTGCGTC 同源臂扩增
Homologous arm amplificationCg694-MF2 CGGGAATTGCATGCTCTCAC AATTTTGTGAAGCGCAAAGC 同源臂扩增
Homologous arm amplificationCg694-MR2 GCTTTGCGCTTCACAAAATT GTGAGAGCATGCAATTCCCG 同源臂扩增
Homologous arm amplificationCg2346-5F AGGCCACCTGCAGCGCTTGAAC 同源臂扩增
Homologous arm amplificationCg2346-5R GGAAAATTGGTGCTCTTGCTGC 同源臂扩增
Homologous arm amplificationCg2346-MF1 ATCCGGCGTCTTTTCCATCACCGTGCCAACGCCACAGTGCCC 同源臂扩增
Homologous arm amplificationCg2346-MR1 GGGCACTGTGGCGTTGGCACGGTGATGGAAAAGACGCCGGAT 同源臂扩增
Homologous arm amplificationCg2346-MF2 CGGGAATTGCATGCTCTCACACATCCGTTGGGGTTGTGTTGA 同源臂扩增
Homologous arm amplificationCg2346-MR2 TCAACACAACCCCAACGGATGTGTGAGAGCATGCAATTCCCG 同源臂扩增
Homologous arm amplificationSur-SLR ATGTTGGCATAAGCCGAACCGT 同源臂扩增
Homologous arm amplificationSur-SLF CCTCTGATATTGGAAGCGACGC 同源臂扩增
Homologous arm amplificationCg694-JC5F CTCTAATCCGCCAAATCCTGAG 突变体鉴定
Mutant Strain IdentificationCg694-JC3R GATGGTTCGTGAACATCCCTTG 突变体鉴定
Mutant Strain IdentificationCg694-JCOF CCGAGCGTAGTGGTCTCATCAA 突变体鉴定
Mutant Strain IdentificationCg694-JCOR AGGCTTCAGCCGCAATTACATC 突变体鉴定
Mutant Strain IdentificationCg694-JCGF ATGCCGGCCGACTACTCCGCTC 突变体鉴定
Mutant Strain IdentificationCg694-JCGR AACATCATGGCCGTCGGAACCG 突变体鉴定
Mutant Strain IdentificationCg2346-JC5F GTAATGAGACAACAGTCGGGCA 突变体鉴定
Mutant Strain IdentificationCg2346-JC3R ACAGAGACGAAAAGACAAGGCG 突变体鉴定
Mutant Strain IdentificationCg2346-JCOF AGGACCGCTACTCACATACACA 突变体鉴定
Mutant Strain IdentificationCg2346-JCOR TTAAAGTTCGAGGGATGACACA 突变体鉴定
Mutant Strain IdentificationCg2346-JCGR AAGAAGAACAACAACGGAAAGA 突变体鉴定
Mutant Strain IdentificationCg2346-JCGF ATCAGAGTGGAGTGACAAGAGT 突变体鉴定
Mutant Strain IdentificationCg694-OF(EcoRI) CTGCGGCAGCGGCCGAATTCATGCCGGCCGACTACTCCGC 同源臂扩增
Homologous arm amplificationCg694-OR(HindIII) GATCCGGTGGATCCAAGCTTAACATCATGGCCGTCGGAAC 同源臂扩增
Homologous arm amplificationCg2346-OF(EcoRI) CTGCGGCAGCGGCCGAATTCATGAAGGCAGTTTCCGTTCT 同源臂扩增
Homologous arm amplificationCg2346-OR(HindIII) GATCCGGTGGATCCAAGCTTGAACAGGCGAGCAGCGGCAG 同源臂扩增
Homologous arm amplificationCgActin-qF GATTGGTATGGGCCAGAAGG 相对表达量检测
Relative expression level detectionCgActin-qR GGAGCTCGTTGTAGAAGGTG 相对表达量检测
Relative expression level detectionCg694-QF TCAGAAATTCAGCCCCCGTC 相对表达量检测
Relative expression level detectionCg694-QR AACTCAGCGGTAAGCTCGTC 相对表达量检测
Relative expression level detectionCg2346-QF AGGAAACAACAATGGCACCG 相对表达量检测
Relative expression level detectionCg694-QR CAGCAGCAGAGCCACTAGAA 相对表达量检测
Relative expression level detection -
首先使用总RNA提取试剂盒,采用液氮研磨的方法提取胶孢炭疽菌野生型菌株的菌丝(mycelia)、孢子(conidia)、萌发的芽管(germ tubes)、附着胞(appressoria)等不同发育时期以及胶孢炭疽菌野生型菌株侵染橡胶树叶片(Infection of rubber tree leaves by C. gloeosporioides Wild Type)不同时间段的总RNA,使用反转录试剂盒将总RNA反转录为cDNA。随后以cDNA为模板,用荧光定量PCR仪进行实时荧光定量(Quantitative Real-time polymerase chain reaction,qRT-PCR)检测。
-
使用以同源重组原理为基础的Spark HiFi Seamless Cloning Kit(单片段)无缝克隆试剂盒进行pEGAD载体的构建:首先用总RNA提取试剂盒提取孢炭疽菌野生型菌株菌丝时期的总RNA,使用反转录试剂盒进行cDNA合成,以菌丝时期cDNA为模板,扩增获得Cg694和Cg2346基因的CDS片段,使用无缝克隆试剂盒构建pEGAD-Cg694-GFP和pEGAD-Cg2346-GFP载体,并通过农杆菌转化法转入GV3101农杆菌。随后,将分别携带有 INF1(INF1 elicitin)、BAX(BCL2-associated X protein)、pEGAD-GFP、pEGAD-Cg694-GFP 和 pEGAD-Cg2346-GFP 的农杆菌菌液,使用含 0.1% 卡那霉素和 0.1% 利福平的 LB 液体培养基,在 28℃ 下振荡培养 24 小时,使其生长至对数期,离心后重悬于浸润缓冲液(10 mmol·L−1 MES pH5.6、10 mmol·L−1 MgCl2、10 μmol·L−1 AS)中,使用分光光度计将菌液调整浓度至OD600=0.6~0.7。采用注射器侵染本氏烟草叶片,并使用黑色记号笔将烟草叶片的浸润部分进行标记。侵染后将烟草置于25℃培养室中遮光培养24 h,再放置于光照条件下培养48~96 h后观察叶片情况。
-
本研究使用以同源重组原理为基础的Split-Marker基因敲除技术分别构建胶胞炭疽菌Cg694和Cg2346基因的敲除突变体:首先以野生型菌株基因组为模板,使用高保真扩增酶分别扩增Cg694和Cg2346的上下游同源臂和氯嘧磺隆抗性基因(sulfonylurea resistance gene,SUR)的上下两部分。然后,利用融合引物,将上游同源臂与抗性基因的上部分、下游同源臂与抗性基因的下部分进行融合,获得敲除转化片段。橡胶树胶孢炭疽菌原生质体的制备与转化方法参照郭云峰等[21]的实验体系。使用CM(complete medium)培养基在28℃,150 r·min−1的条件下培养胶孢炭疽菌野生型以获得孢子,通过滤膜过滤后将孢子液注入100 mL的1% ME(malt extract)液体培养基中,于28℃,150 r·min−1的条件下培养24 h以获得菌丝。将菌丝过滤后加入到含有蜗牛酶与R10融壁酶的缓冲溶液中,立即在28℃,110 r·min−1摇床中酶解5 h,过滤获得原生质体,并通过PEG4000介导法将片段转入原生质体。随后,将含有原生质体的转化液均匀分布于15 cm培养皿中,加入YCS固体培养基(酵母提取物、酸水解酪蛋白、蔗糖、琼脂)均匀铺满培养皿后放置于28℃倒置培养16 h。配置含有100 μg·mL−1氯嘧磺隆的YCS固体培养基倒入培养皿中,进行筛选培养,并将能够在筛选培养基上生长的菌落接入马铃薯葡萄糖琼脂培养基(potato dextrose agar,PDA)上进行培养和检测。
-
将经过初步抗性筛选的菌株提取基因组进行鉴定,阴性对照为野生型基因组,利用上、下游同源臂外侧引物Cg694-JC5F/Cg694-JC3R、Cg694-JO5F/Cg694-JO3R与Cg2346-JC5F/Cg2346-JC3R、Cg2346-JO5F/Cg2346-JO3R进行检测。使用含100 μg·mL−1氯嘧磺隆的抗性培养基将阳性菌株进行单胞,通过上下游检测引物与基因检测引物Cg694-JCGF/Cg694-JCGR,Cg2346-JCGF/Cg2346-JCGR进行鉴定。
-
将WT、ΔCg694和ΔCg2346生长正常的菌株使用打孔器取相同尺寸的菌块接种于PDA固体培养基中心,28℃培养箱中倒置培养,5 d后使用十字交叉法测量菌落直径并拍照记录,对各菌株的生长速率进行统计分析。
-
将PDA培养基上正常生长的WT、ΔCg694以及ΔCg2346菌株取部分切碎,使用CM液体培养基在28℃,150 r·min−1条件下培养2 d,将培养好的菌液过滤得到孢子液。将孢子液充分震荡均匀后使用血球计数板统计孢子浓度,用无菌水将孢子液稀释至1×103个·mL−1,然后吸取等量孢子液均匀混合到装有30 mL CM液体培养基的小瓶中,28℃,150 r·min−1培养3 d后,将全部菌液过滤并使用血球计数板统计孢子浓度。
-
制备致病力所需的孢子液与1.6.2方法相同。将过滤得到的孢子液
5000 rpm,1 min离心,使用0.5% ME(malt extract broth)液体培养基重悬,使用血球计数板统计孢子浓度并用0.5% ME液体培养基稀释孢子浓度至5×105个·mL−1。选择幼嫩时期的橡胶树叶片作为接种材料,选择叶片合适位置接种5 μL菌液,在28℃保湿条件下培养3 d后,测量统计橡胶树叶片发病情况和病斑大小。 -
萌发实验所需孢子液的培养方法与1.6.2方法相同。将过滤得到的孢子液5 000 r·min−1,1 min离心,使用2% YCS液体培养基将孢子液稀释至5×105个·mL−1,并使用移液枪吸取20 μL置于干净透明的玻璃载玻片上,在28℃保湿条件下培养2、4和6 h。使用显微镜观察并统计孢子萌发情况,拍照记录。
-
附着胞离体诱导形成率实验所需孢子液的培养方法与1.6.2方法相同。将过滤得到的孢子液5 000 r·min−1,1 min离心,使用无菌水进行洗涤后,离心收集孢子,重复3次,确保充分将残留的培养基营养洗净。随后,用无菌水将孢子液稀释至5×105个·mL−1并使用移液枪吸取5 μL接种于疏水表面,在28℃保湿条件下培养12 h与24 h,使用显微镜观察并统计附着胞的形成情况,拍照记录。
-
洋葱内表皮入侵菌丝观察实验所需孢子液的培养方法和稀释方法与1.8.2相同。选取新鲜的洋葱的内表皮作为接种载体,将洋葱内表皮疏水面向上贴于含有0.05 g·mL−1 NaCl的1.5%固体水琼脂表面。将稀释完成的孢子液使用移液枪吸取5 μL接种于洋葱内表皮疏水面,在28℃保湿条件下培养12 h与24 h,使用显微镜观察并统计入侵菌丝的形成情况,拍照记录。
-
本实验使用ImageJ软件进行数据统计和显著性分析,通过单因素方差分析单个变量的数据,使用Duncan氏多范围检验进行均值分离。P<0.05时,认为存在显著性差异。实验单独重复3次。
-
通过UniProt网站查询,Cg694被预测属于主要协助转运蛋白超家族(MFS),MFS转运蛋白通常具有保守的结构域,由12个跨膜螺旋组成,具有对称的N端和C端结构域[22]。胶胞炭疽菌Cg694的基因序列包含一个长度为684 bp的开放阅读框(open reading frame,ORF),没有内含子,编码区(coding sequence,CDS)总长为684 bp,编码一个具有227个氨基酸的多肽链,分子质量为25.47 kDa。使用SMART对结构域进行预测并分析,Cg694蛋白具有低复杂度区域和跨膜螺旋结构域(图1-A)。
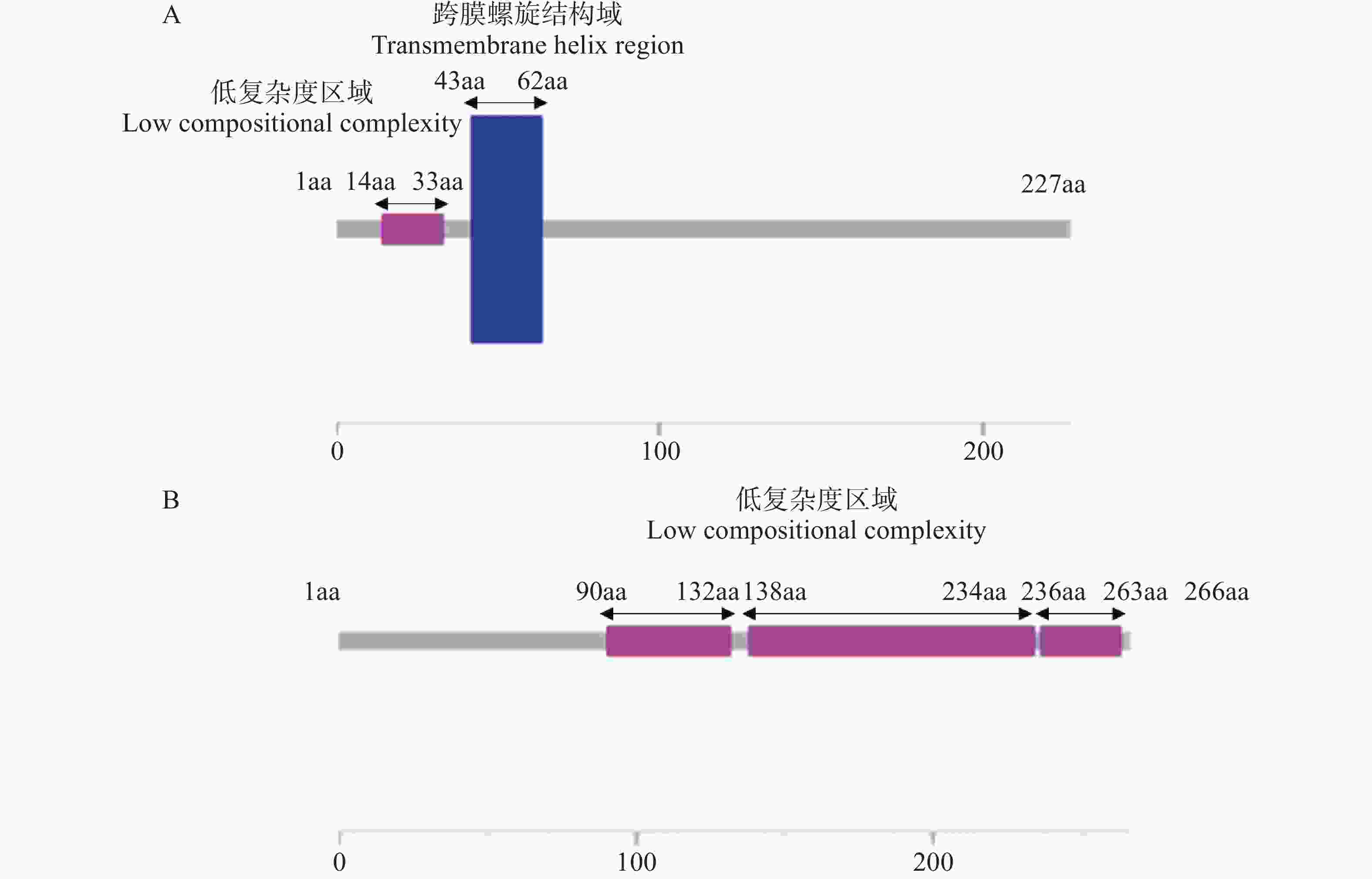
图 1 胶胞炭疽菌Cg694和Cg2346蛋白氨基酸序列结构分析
Figure 1. Analysis of amino acid sequence structure of Cg694 and Cg2346 proteins in C. gloeosporioides
目前,Cg2346鲜见报道,是一个功能未知的蛋白。胶胞炭疽菌Cg2346的基因序列包含一个长度为864 bp的开放阅读框,含有一个内含子,编码区总长为801 bp,编码一个具有266个氨基酸的多肽链,分子质量为27.40 kDa。使用SMART对结构域进行预测并分析,发现Cg2346蛋白有多个低复杂度区域(图1-B)。
-
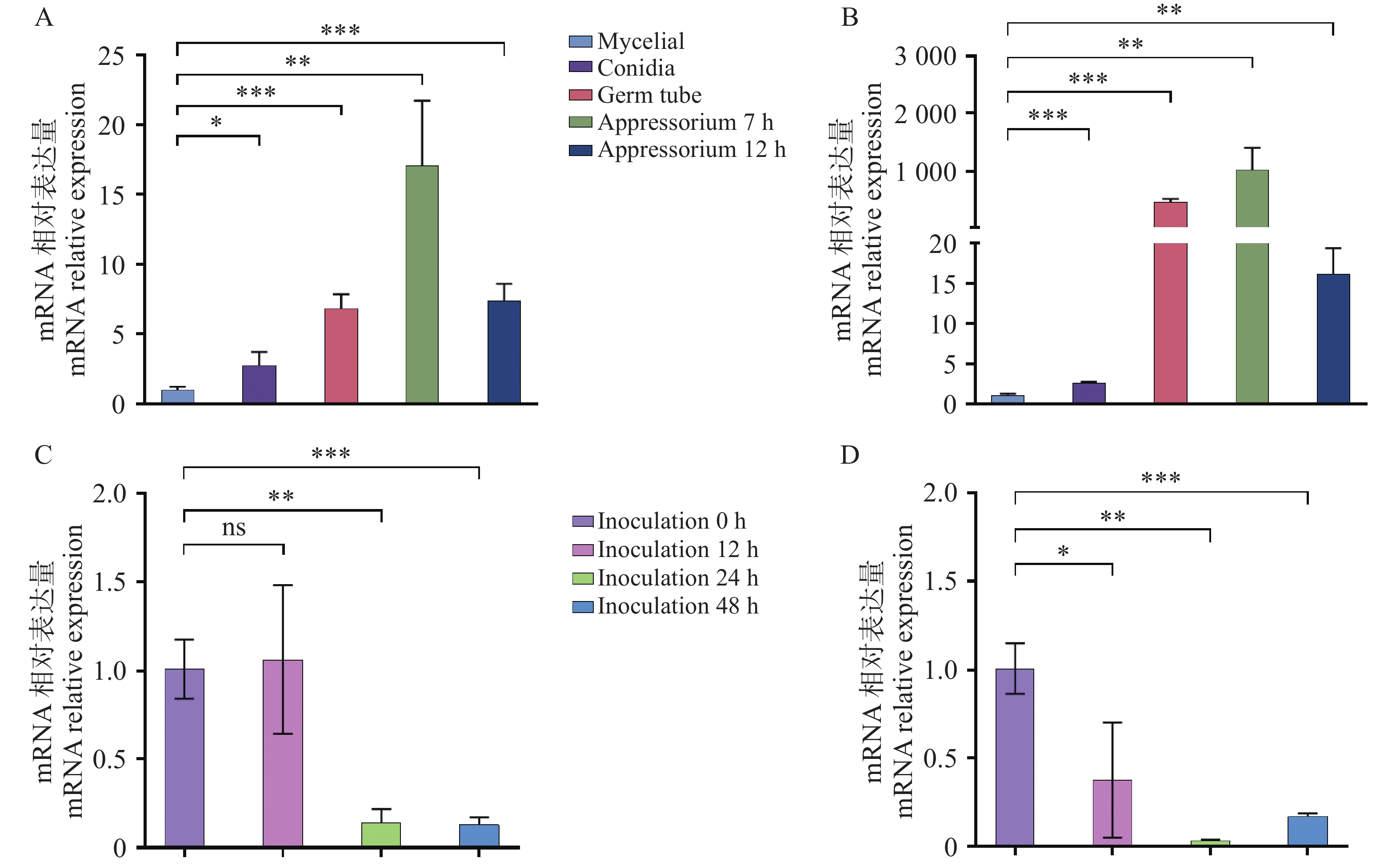
本研究分析了Cg694和Cg2346基因在胶孢炭疽菌不同发育阶段(营养菌丝mycelial、孢子conidia、萌发的芽管germ tubes、附着胞appressoria)和胶孢炭疽菌侵染橡胶树叶片(Infection of rubber tree leaves by C. gloeosporioides Wild Type)等不同时期的相对表达量差异。结果表明:与营养菌丝时期的表达量相比,Cg694和Cg2346在孢子、萌发及附着胞发育阶段的表达量显著上调(图2-A—B);与胶孢炭疽菌侵染橡胶树叶片0 h时期相比,Cg694和Cg2346在侵染后的橡胶树组织中显著下调(图2-C—D)。Cg694和Cg2346在胶孢炭疽菌孢子、萌发和附着胞等发育阶段表达量显著增加,推测这2个蛋白可能参与调控胶孢炭疽菌孢子的萌发和附着胞结构的形成。
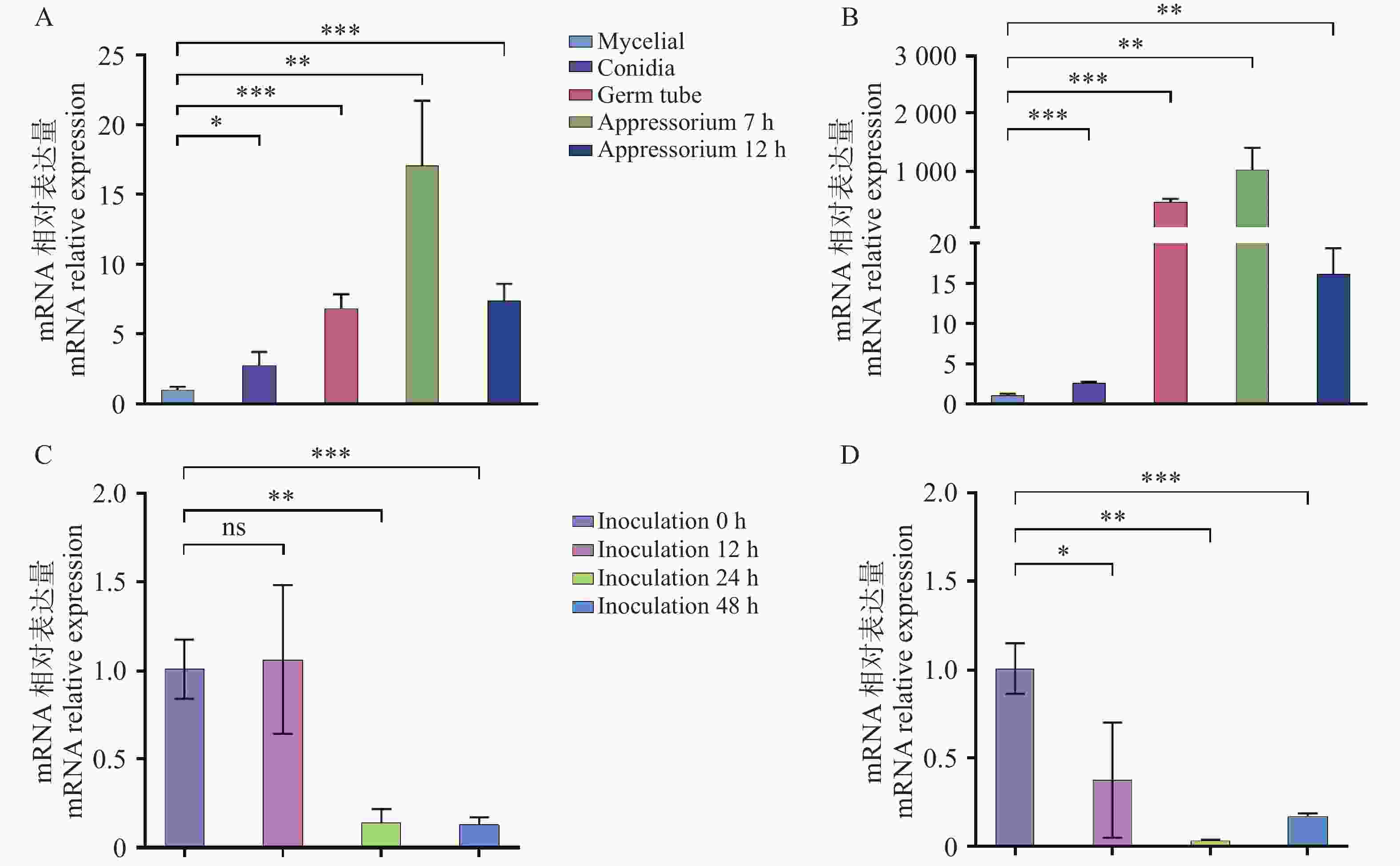
图 2 Cg694和Cg2346基因在胶孢炭疽菌不同时期的相对表达量检测
Figure 2. Detection of relative expression levels of genes Cg694 and Cg2346 in C. gloeosporioides at different developmental stages
-
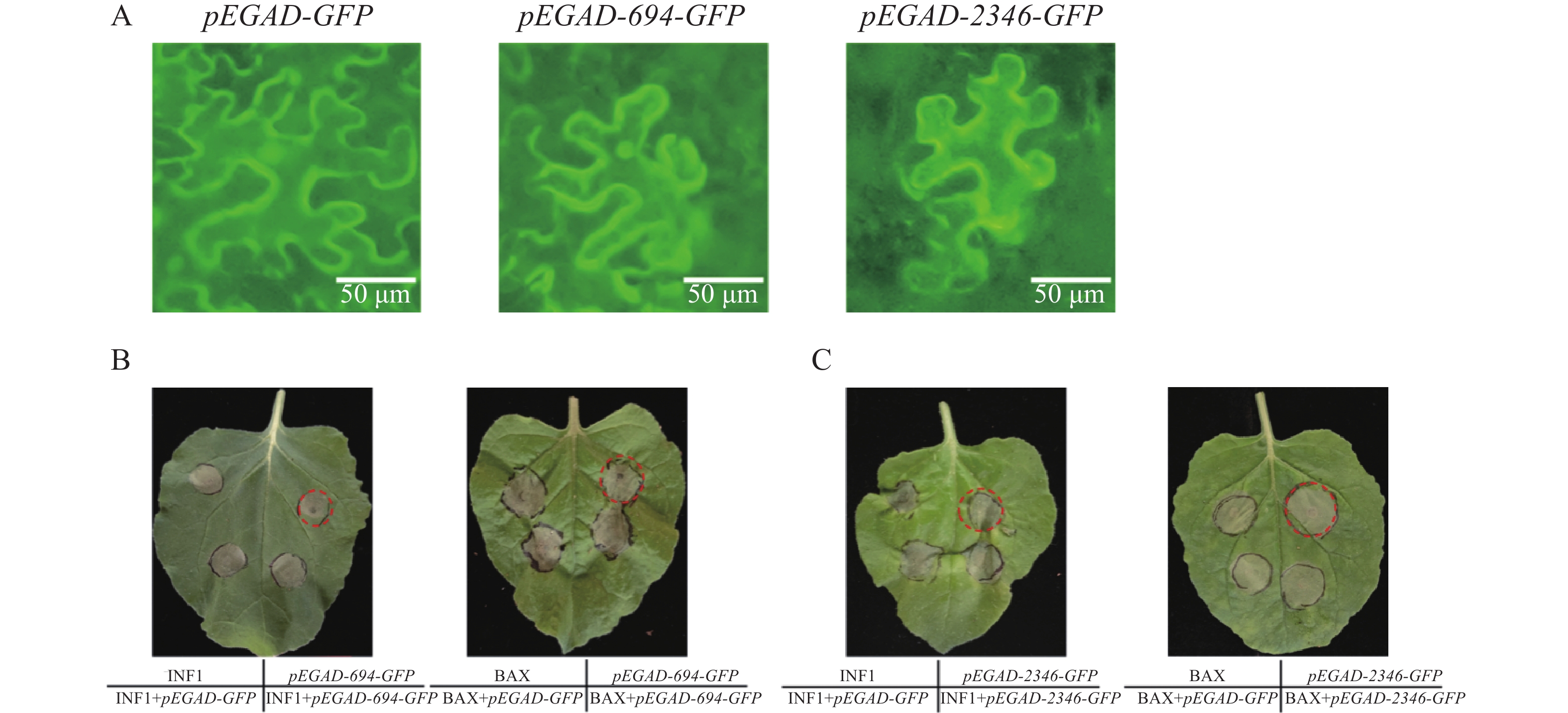
本研究在本氏烟草叶片上分别注射了pEGAD-GFP,pEGAD-Cg694-GFP和pEGAD-Cg2346-GFP。侵染48 h后在荧光显微镜下观察烟草叶片的亚细胞定位,发现空载GFP定位于细胞膜、细胞质和细胞核;而Cg694-GFP和Cg2346-GFP均定位在细胞膜和细胞核(图3-A)。继续放置72 h后,观察到注射pEGAD-Cg694-GFP和pEGAD-Cg2346-GFP的叶片区域出现了明显的坏死现象,推测其可能会影响烟草的免疫系统。随后,在烟草叶片上分别注射INF1/BAX、pEGAD-GFP,pEGAD-Cg694-GFP/pEGAD-Cg2346-GFP,以及将INF1/BAX分别与pEGAD-Cg694-GFP/pEGAD-Cg2346-GFP共注射。放置5 d后,观察烟草叶片发现,单独注射INF1/BAX和单独注射pEGAD-Cg694-GFP和pEGAD-Cg2346-GFP的叶片区域均出现明显的坏死,且INF1/BAX与pEGAD-Cg694-GFP/pEGAD-Cg2346-GFP共同表达的叶片区域也出现明显坏死(图3-B—C)。说明Cg694和Cg2346会触发烟草细胞的免疫反应,诱导烟草叶片出现坏死,但它们不能抑制由INF1或BAX引起的烟草叶片坏死。
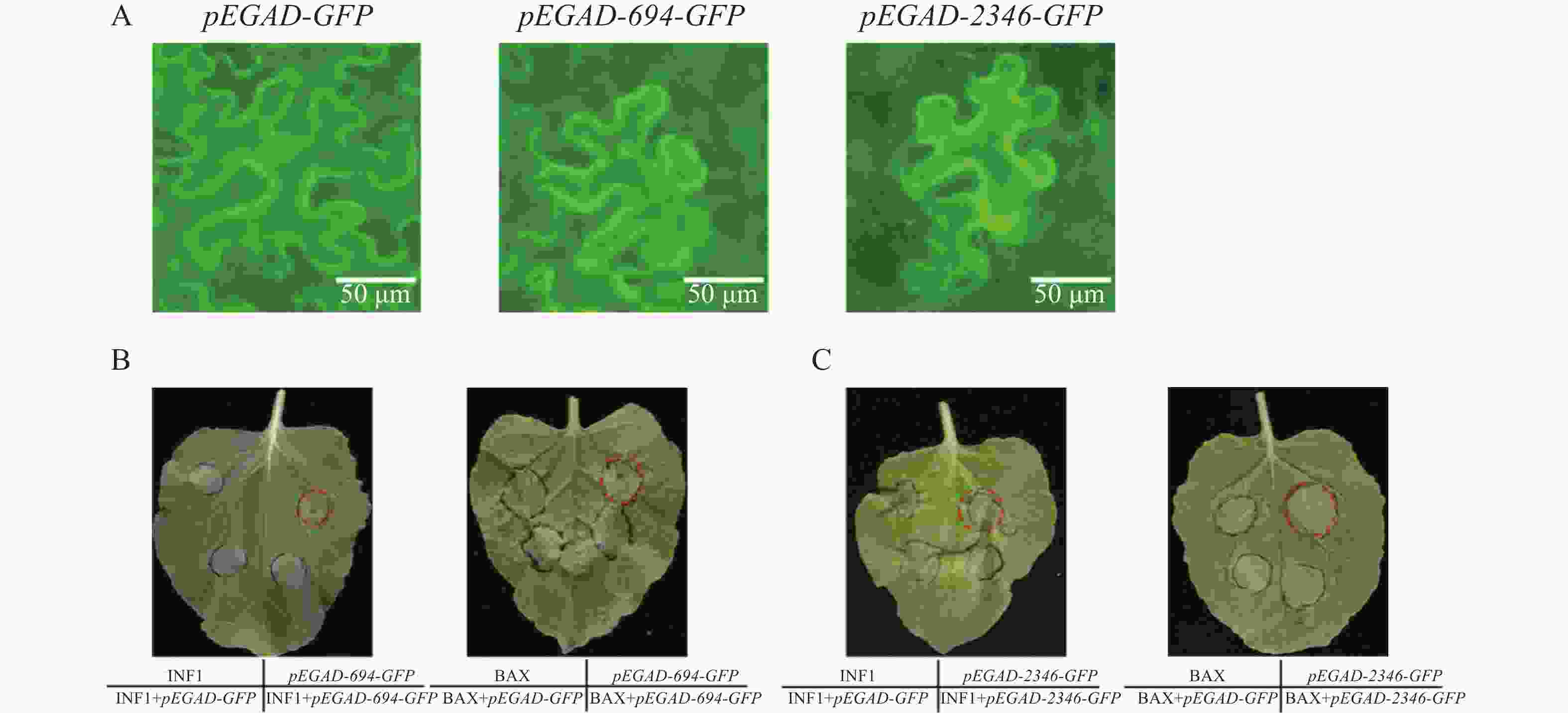
图 3 烟草叶片瞬时表达Cg694和Cg2346基因
Figure 3. Transient expression of genes Cg694 and Cg2346 in N. benthamiana leaves
-
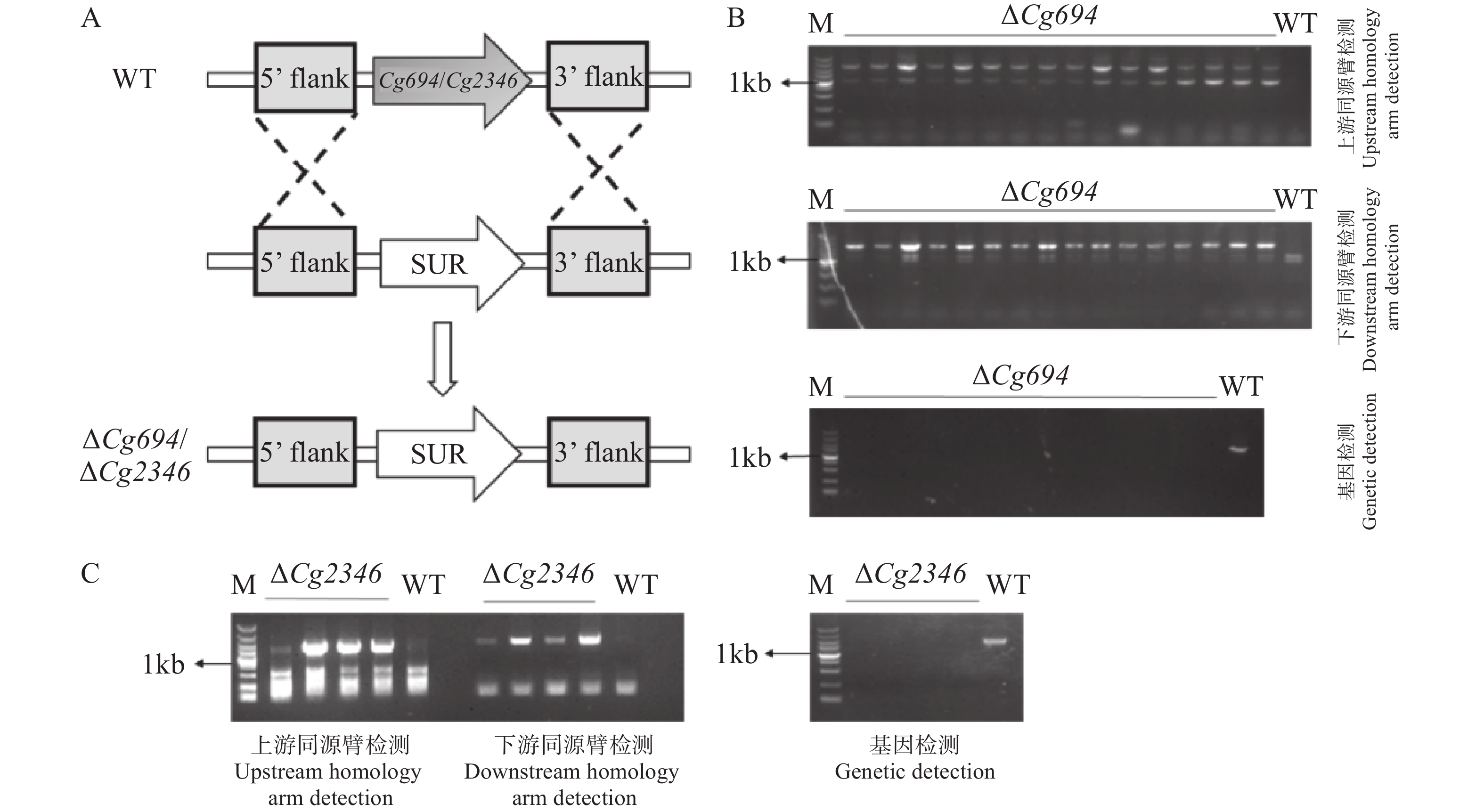
本研究分别对Cg694和Cg2346基因构建基因敲除突变体(图4-A)。先将在筛选培养基上获得的转化子进行上下游片段检测,将检测正确的阳性转化子进行单孢,获得纯合的转化子。再进行上下游片段检测和基因检测,转化子均能扩增出上下游片段且未检测出基因条带(图4-B—C)。检测结果表明,ΔCg694和ΔCg2346突变体已经构建成功。
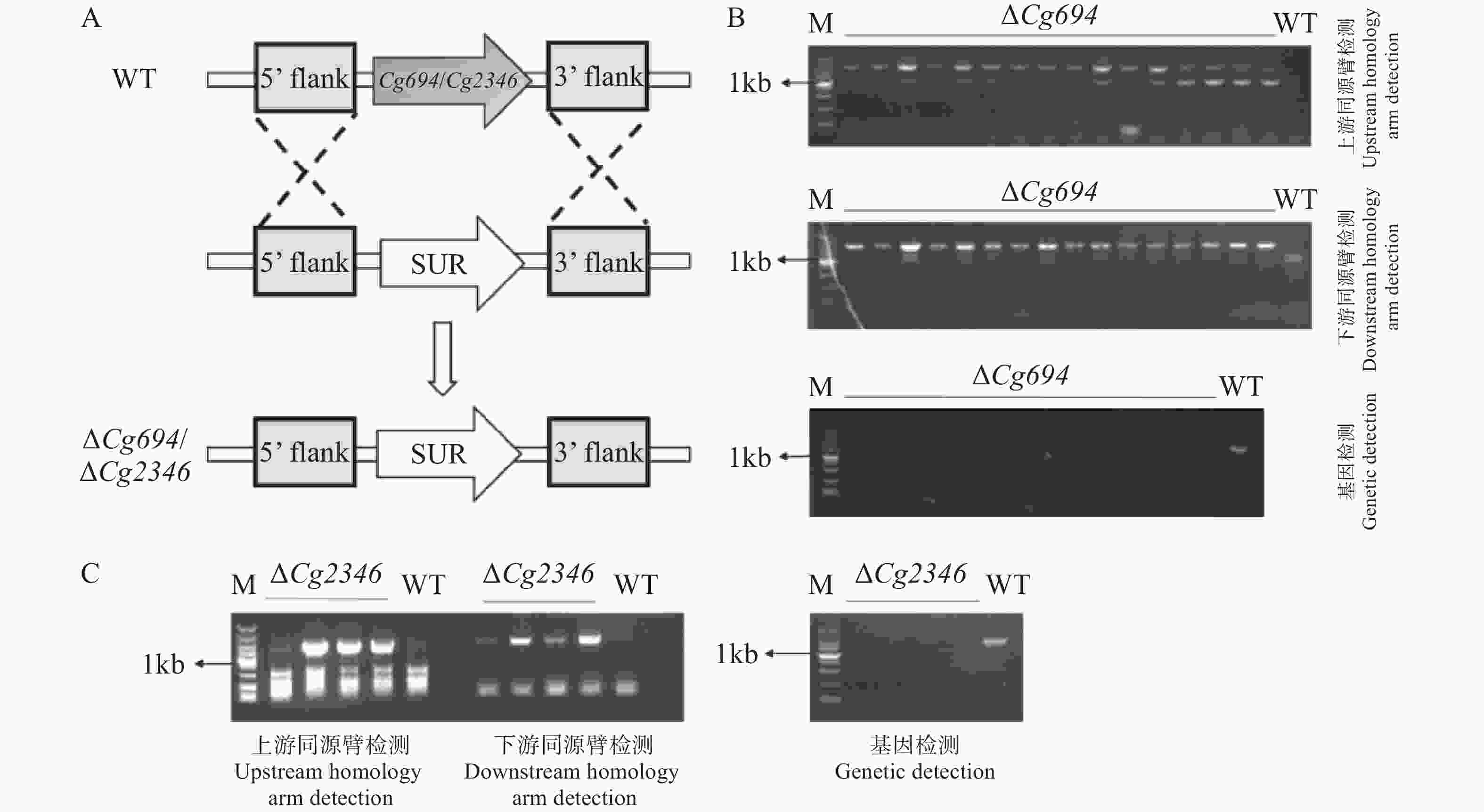
图 4 Split-Marker基因敲除原理以及基因敲除突变体的鉴定
Figure 4. Principle of Split-Marker gene knockout and identification of knockout mutants
-
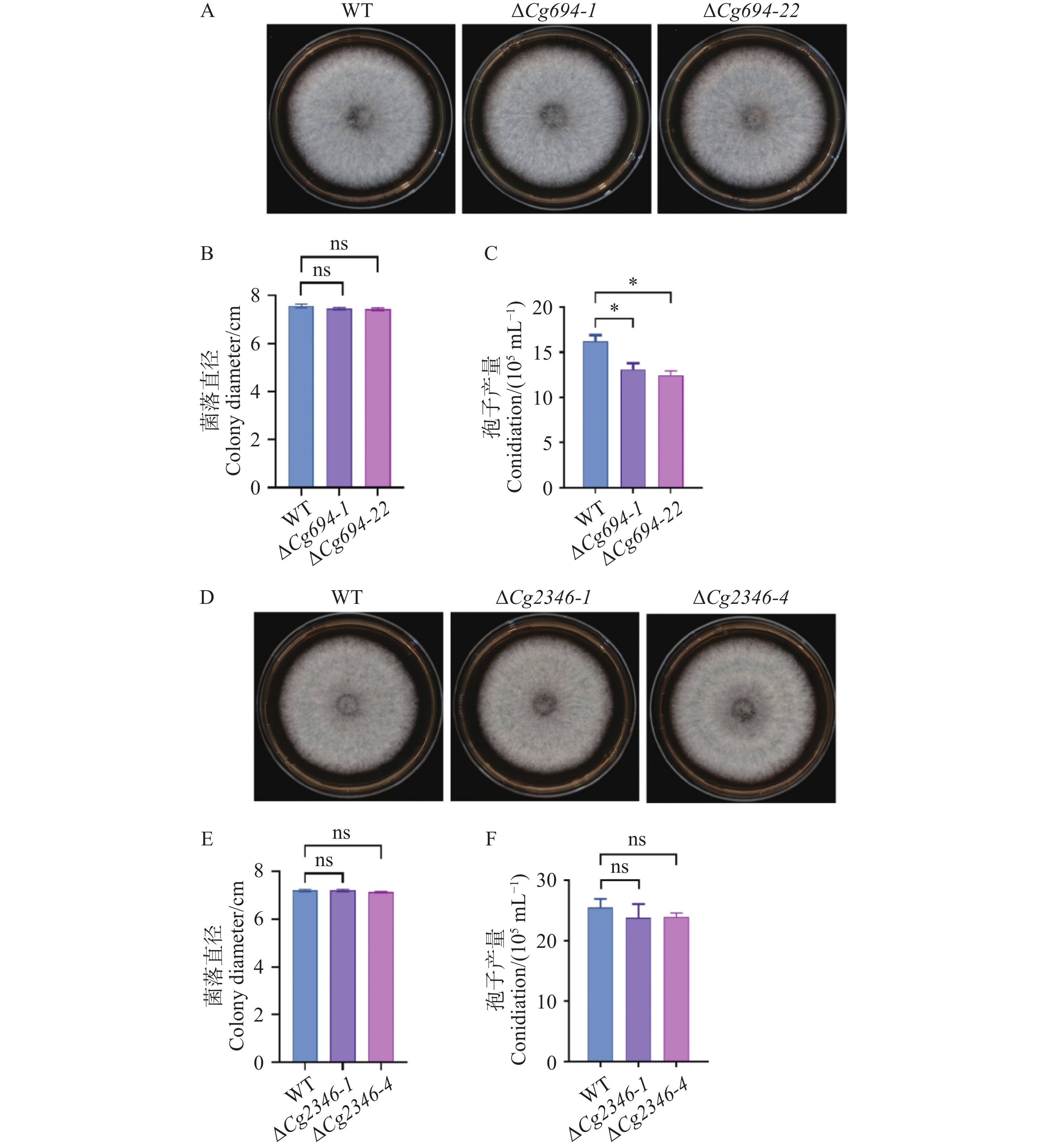
在ΔCg694突变体生长实验中,WT的菌株平均直径为7.5 cm;ΔCg694突变体的菌株平均直径为7.45 cm(图5-A—B);在ΔCg2346突变体生长实验中,WT的菌株平均直径为7.2 cm,ΔCg2346突变体的菌株平均直径为7.2 cm(图5-D—E)。结果表明ΔCg694和ΔCg2346突变体菌落生长速率与野生型相比均没有显著差异。在ΔCg694突变体产孢实验中,WT的孢子产量为16.2×105个·mL−1,ΔCg694突变体的的孢子产量为12.75×105个·mL−1(图5-C);在ΔCg2346突变体产孢实验中,WT的孢子产量为25.5×105个·mL−1,ΔCg694突变体的的孢子产量为23.85×105个·mL−1(图5-F)。结果表明,Cg694和Cg2346不影响菌株的营养生长,Cg2346对孢子产量没有影响,但Cg694参与对孢子产量的调控。
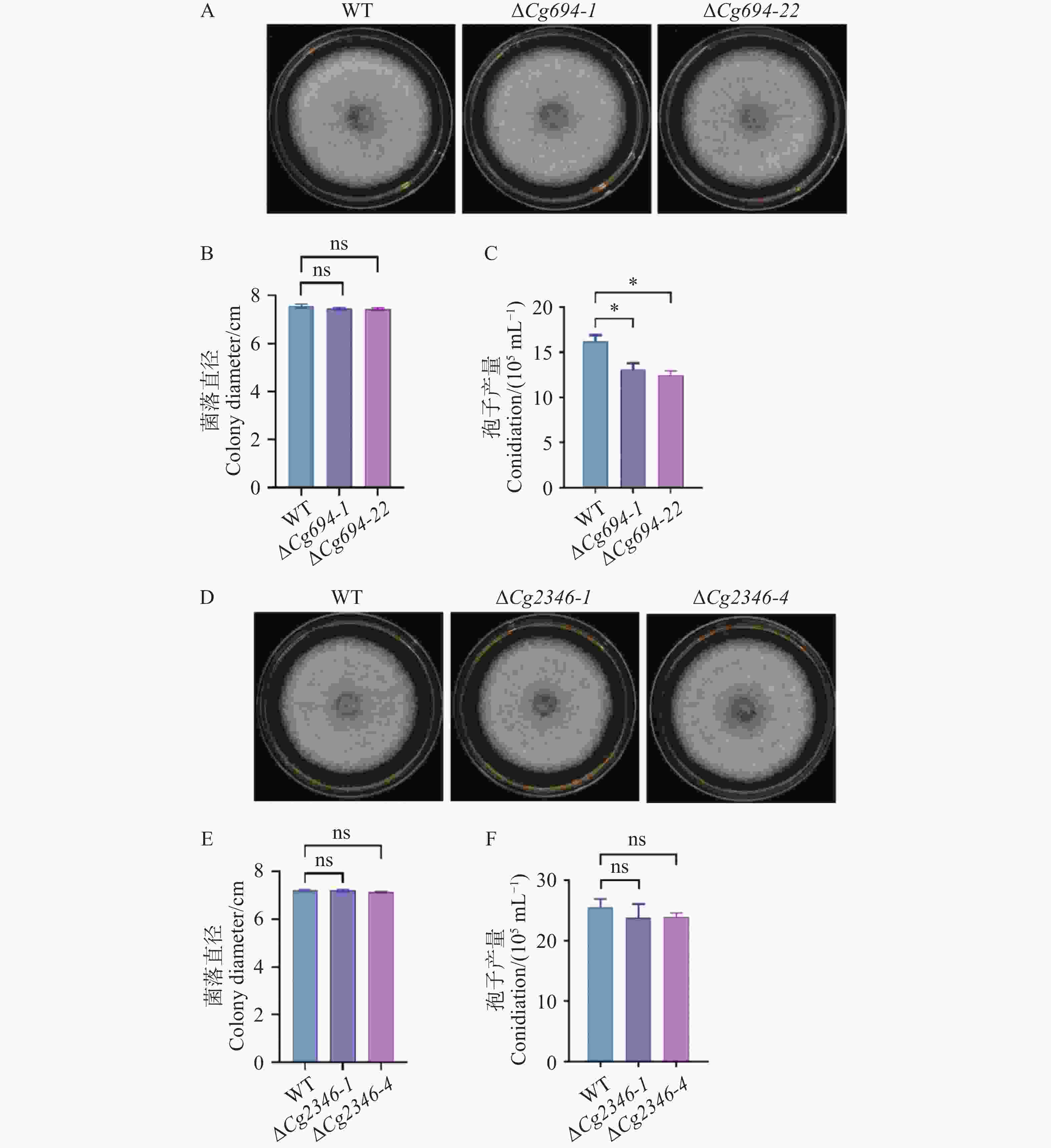
图 5 ΔCg694和ΔCg2346突变体生长速率与产孢量统计
Figure 5. Growth rate and sporulation quantification of ΔCg694 and ΔCg2346 mutant strains
-
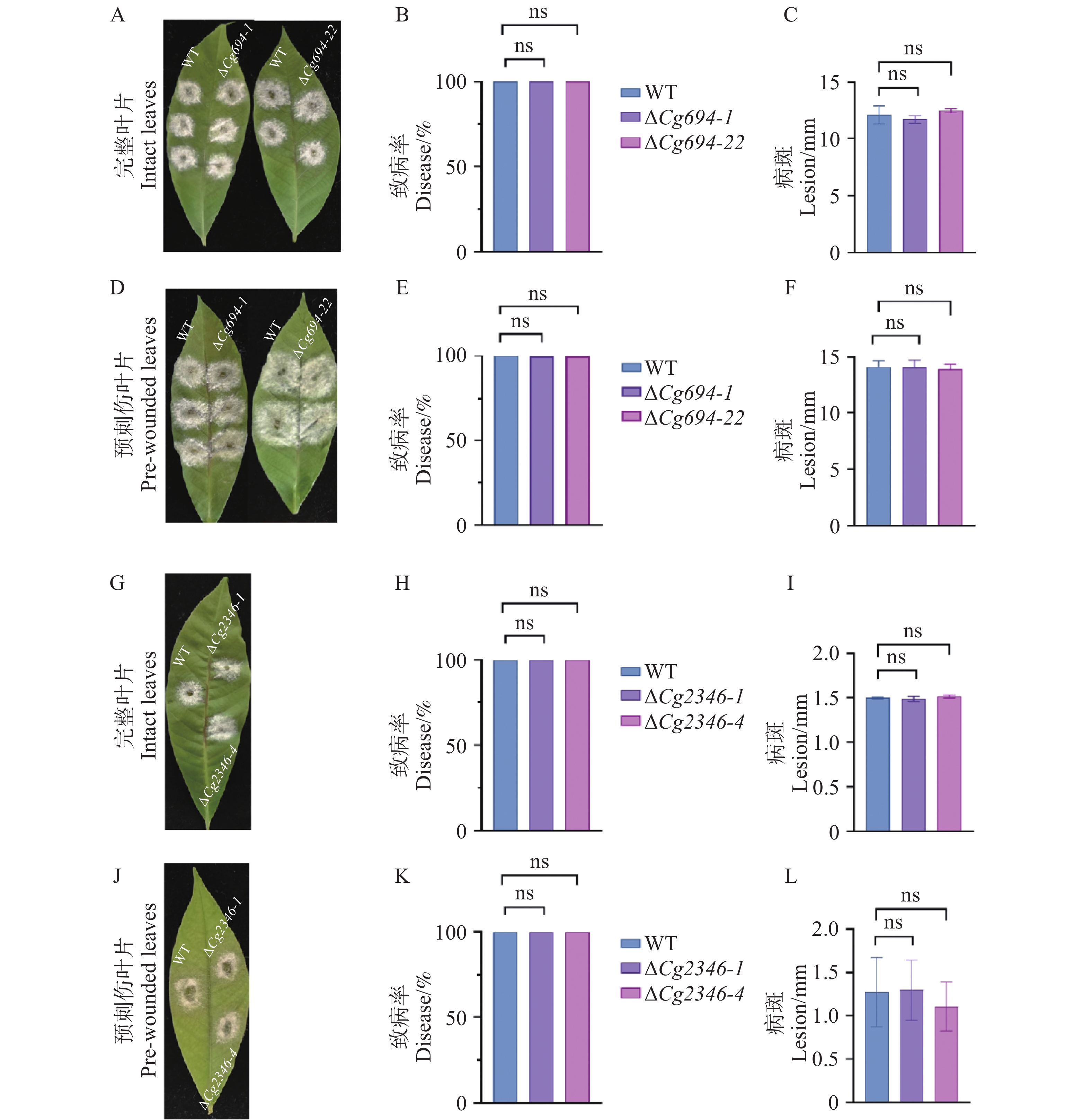
胶孢炭疽菌能够侵染橡胶树叶片,其侵染病斑的大小可以直接反映病原菌致病的能力。从图6可知,分别将WT、ΔCg694和ΔCg2346突变体接种于完整和预刺伤处理的淡绿期橡胶树叶片3 d后,各菌株的发病率和病斑大小基本相同。以上实验结果说明,Cg694和Cg2346基因的缺失没有显著影响到胶胞炭疽菌对于橡胶树叶片的致病力。
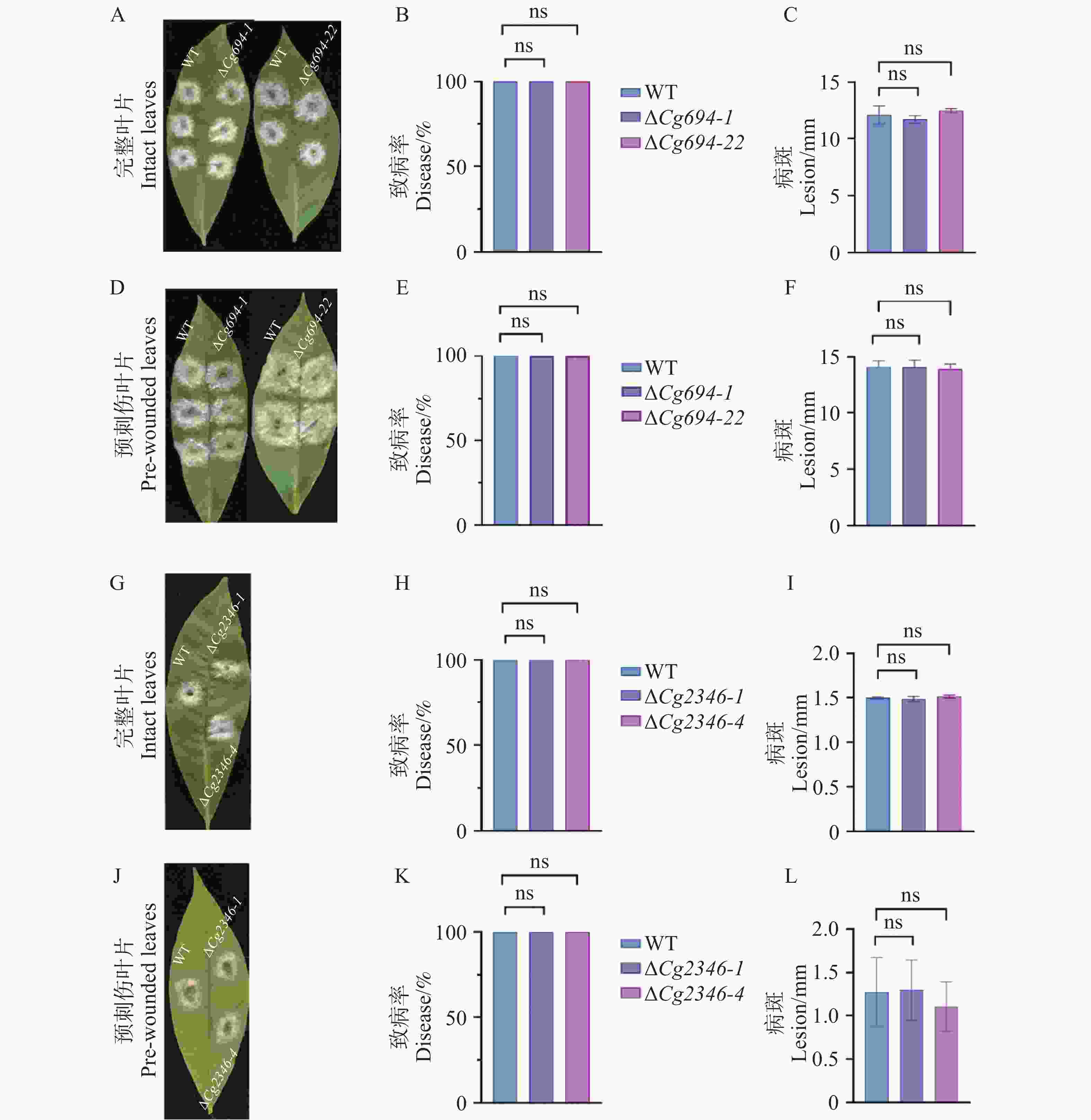
图 6 ΔCg694和ΔCg2346突变体致病力分析
Figure 6. Pathogenicity analysis of ΔCg694 and ΔCg2346 strains
-
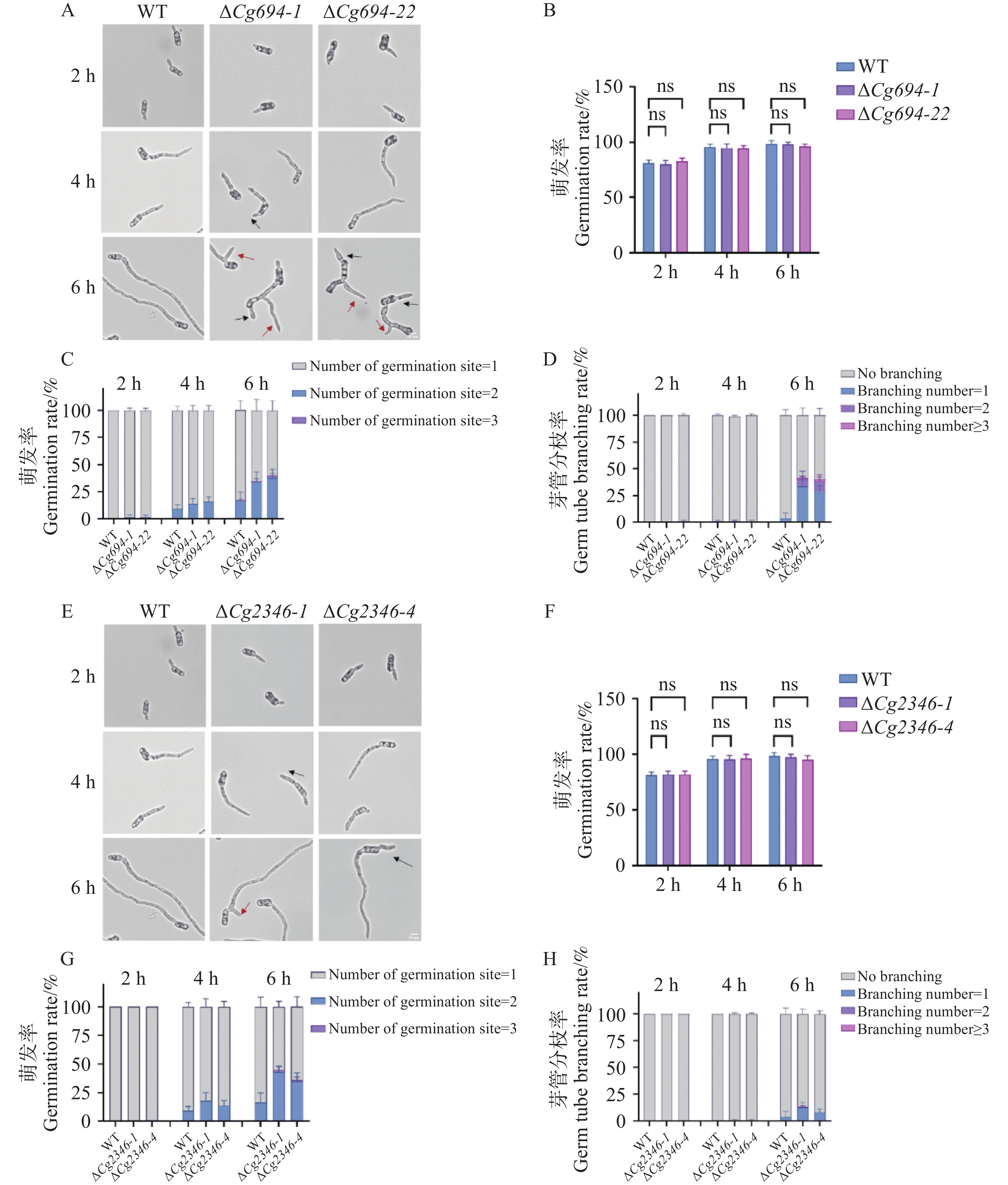
通过显微镜观察发现,ΔCg694突变体的孢子萌发率与WT的孢子萌发率相比无显著差异。但在萌发6 h后,39.2%的ΔCg694突变体孢子呈现多端萌发现象,53.9%的ΔCg694突变体孢子呈现芽管分枝情况,比例明显高于野生型(16.3%和3.7%)(图7-A—D)。与之相似的是,ΔCg2346突变体的孢子萌发率与WT的孢子萌发率相比不存在显著差异,但是在萌发6 h后,44.3%的ΔCg2346突变体孢子呈现多端萌发现象,10.8%的ΔCg2346突变体孢子呈现芽管分枝情况,比例明显高于野生型(16.3%和3.7%)(图7-E—H)。以上结果表明,Cg694和Cg2346基因的缺失对孢子的萌发能力没有影响,Cg694和Cg2346参与调控芽管的极性生长。
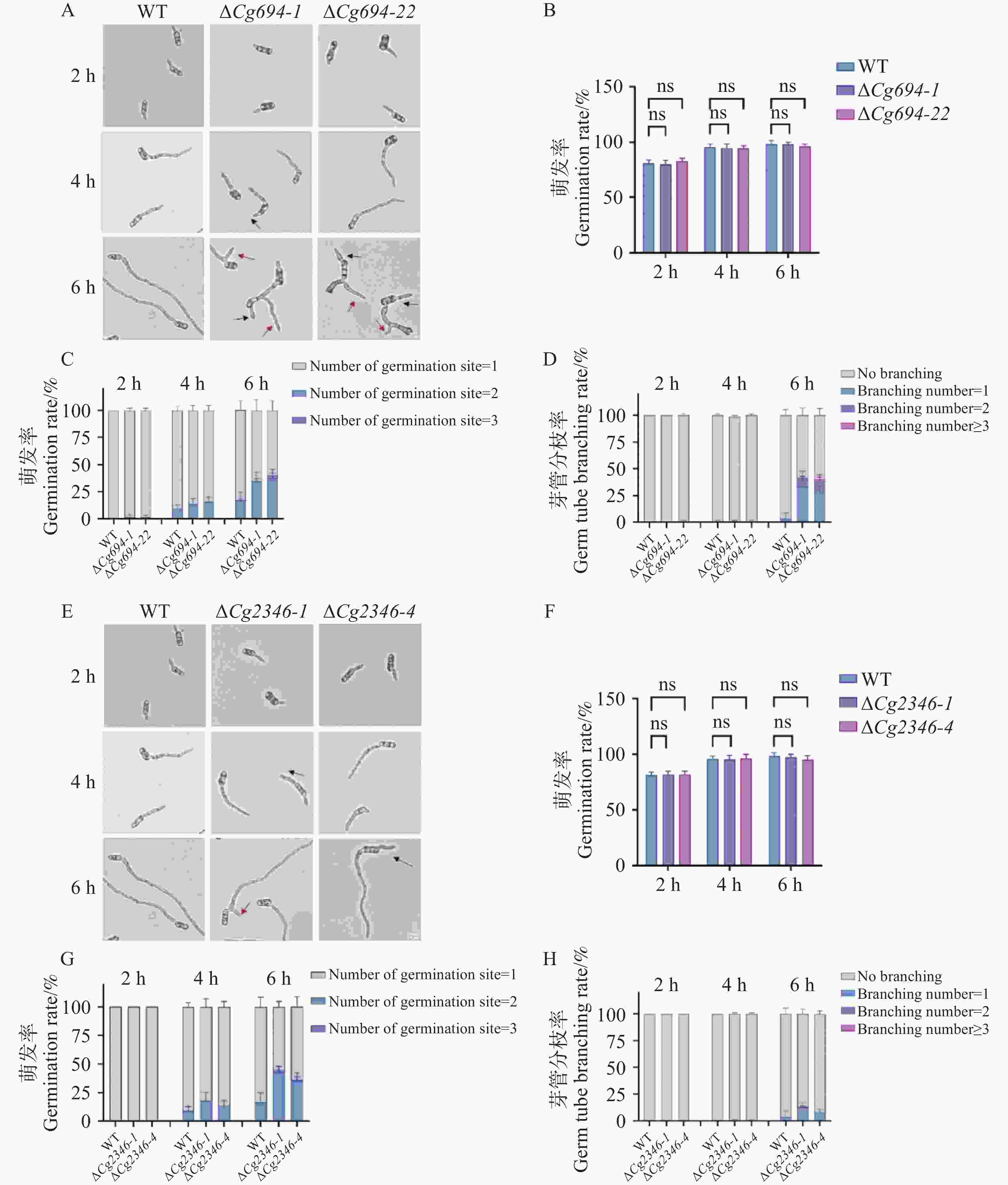
图 7 ΔCg694和ΔCg2346突变体孢子萌发情况分析
Figure 7. Analysis of spore germination characterization in ΔCg694和ΔCg2346 strains
-
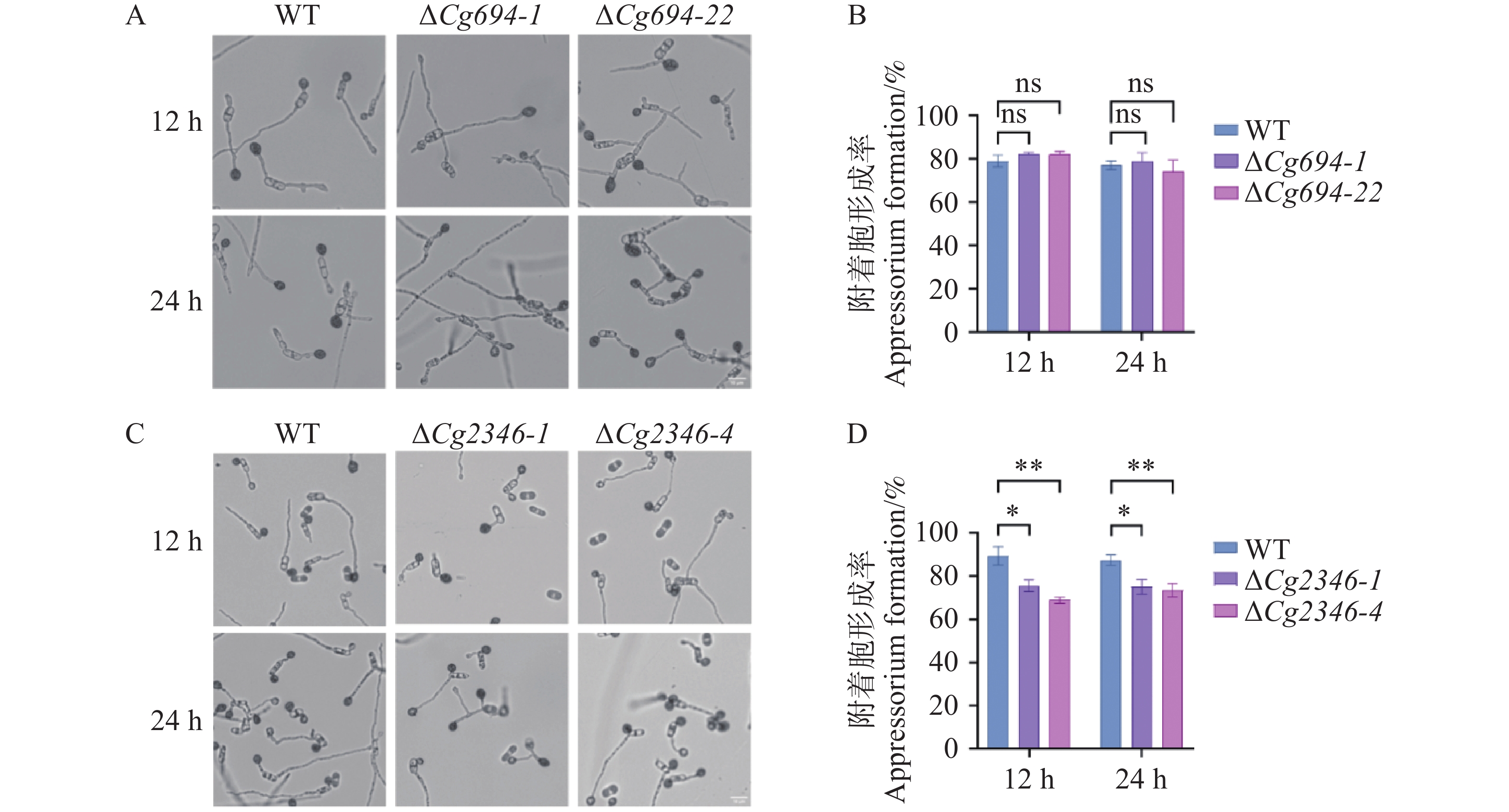
12 h和24 h,ΔCg694突变体附着胞形成率与WT的附着胞率相比没有显著差异(图8-A—B)。观察ΔCg2346突变体附着胞形成情况发现,12 h时WT的附着胞率形成率(附着胞数量/分生孢子总数)平均为89.4%,ΔCg2346突变体附着胞形成率约为75%;24 h时,WT的附着胞率形成率平均为88%,ΔCg2346突变体附着胞形成率约为75.5%,存在显著差异(图8-C—D)。以上结果说明,Cg694不参与调控胶胞炭疽菌附着胞的形成,Cg2346参与调控胶胞炭疽菌附着胞的形成。
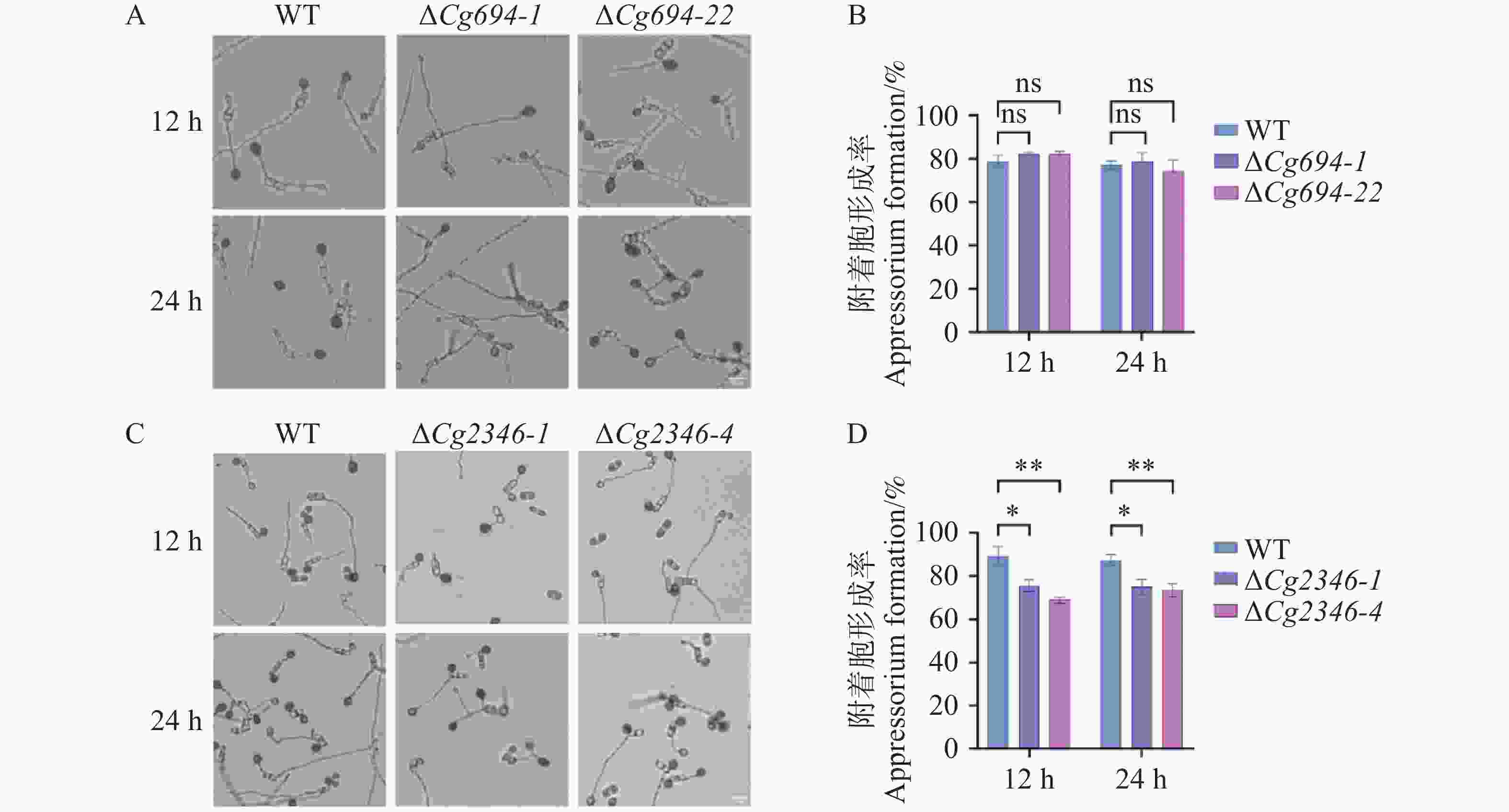
图 8 ΔCg694和ΔCg2346突变体附着胞形成率分析
Figure 8. Analysis of appressorium formation rate in ΔCg694和ΔCg2346 strains
-
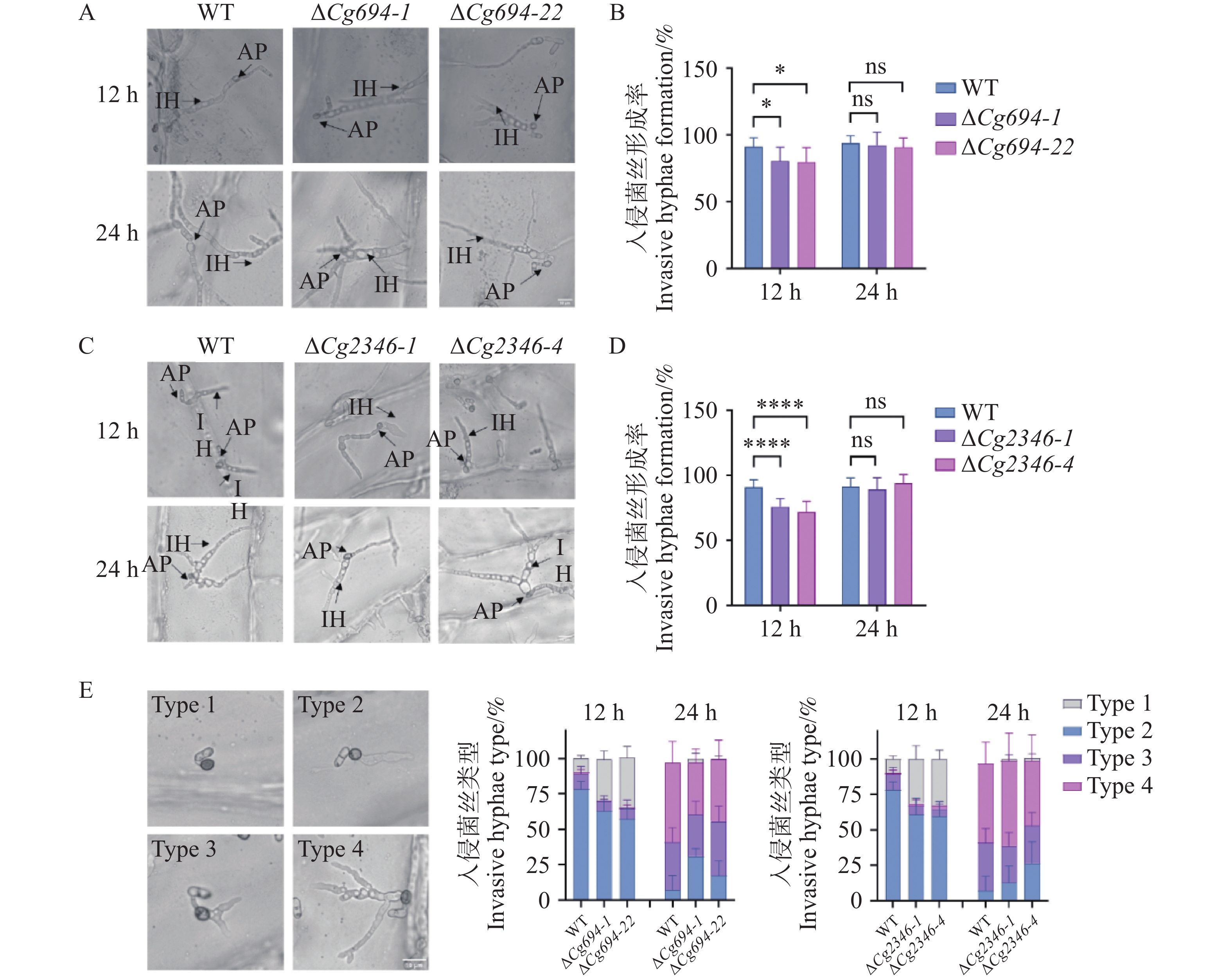
在12 h时,ΔCg694和ΔCg2346突变体在洋葱内表皮上的入侵菌丝的平均形成率(形成入侵菌丝的孢子数/分生孢子总数)分别为78.7%和74.1%,显著低于野生型为90.3%的平均形成率;24 h时,ΔCg694和ΔCg2346突变体在洋葱内表皮上的入侵菌丝的平均形成率分别为91.3%和91.7%,与野生型91.1%的平均形成率没有显著差异(图9-A—D)。接下来,对入侵菌丝的不同形成情况进行了统计,结果表明,在12 h时,与野生型相比ΔCg694和ΔCg2346突变体在洋葱内表皮上的入侵菌丝的形成率差异主要集中于只形成附着胞而不形成入侵菌丝(类型1),在24 h时,与野生型相比没有显著差异(图9-E)。由此可知,Cg694和Cg2346在胶孢炭疽菌对寄主的入侵前期有一定的调控作用。
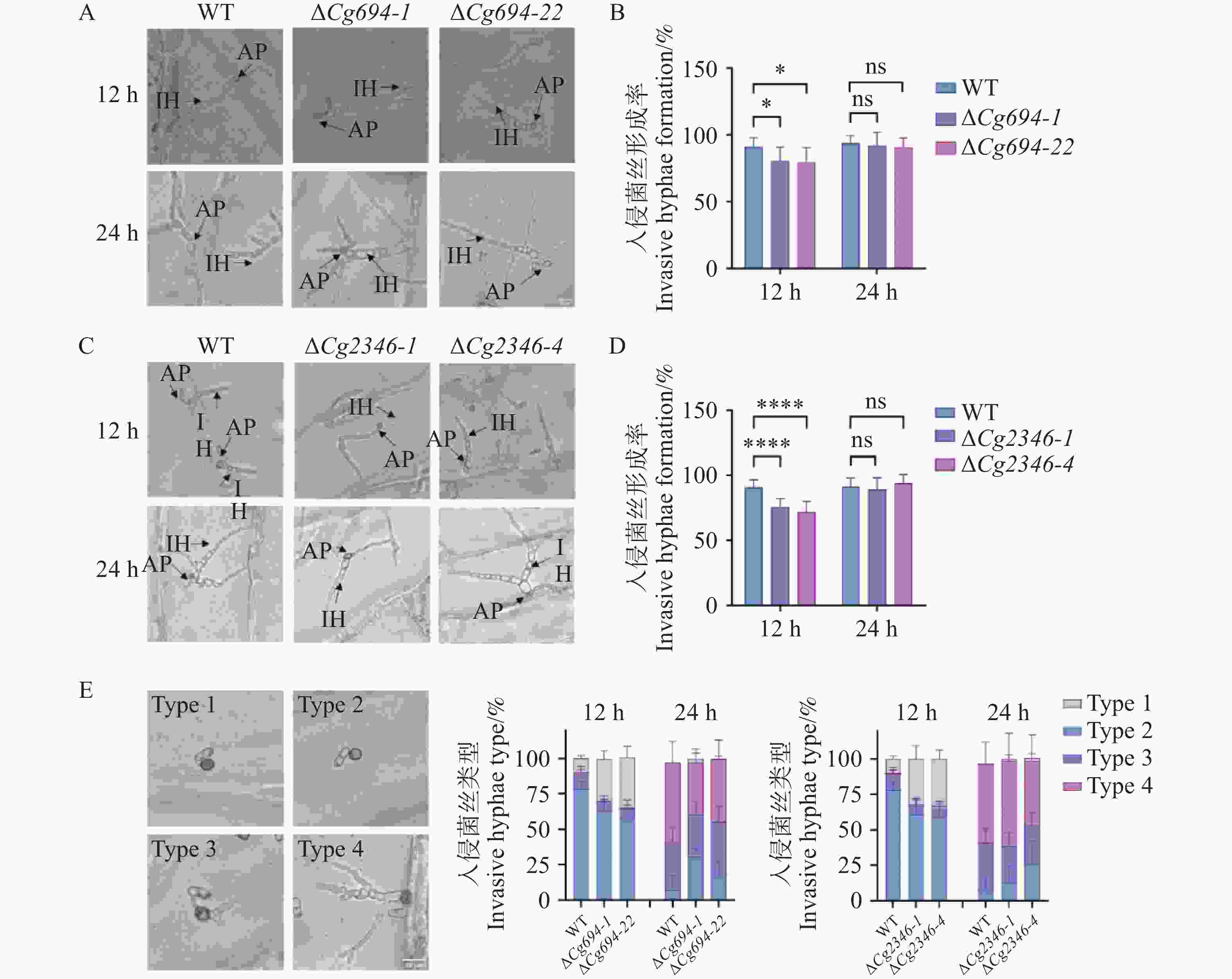
图 9 ΔCg694和ΔCg2346突变体入侵菌丝形成率分析
Figure 9. Analysis of invasive hypha formation rate in ΔCg694和ΔCg2346 strains
-
效应蛋白是病原真菌在侵染寄主过程所分泌的一类蛋白,能够影响植物的免疫系统和寄主细胞的环境[23]。效应蛋白在植物与微生物互作过程中发挥了重要调控作用,是当下植物与微生物互作研究领域的热点[24]。病原菌中的显性抗性(avirulence,Avr)基因与寄主的抗性(resistance,R)基因相对应并发生相互作用,导致如超敏感反应(HR)等寄主防御反应的发生,从而抑制病原菌的侵染[25]。多年来,研究人员们一直致力于寻找更多的Avr和R基因以增加人类对植物−微生物互作机制的了解[26]。植物的免疫系统存在2条主动防御途径:一是基于PAMP识别受体(pattern recognition receptors,PRRs)对保守的病原菌相关分子模式(PAMPs)的识别,从而激活PAMP触发的免疫(PTI)[27];二是植物通过R蛋白效应子感知后而激活的效应触发免疫(ETI),激活植物中包括过敏反应(HR)等防御反应。目前,真菌中已经有多个关于效应蛋白的研究:在稻瘟菌(M.Oryzae)中,研究证明其效应蛋白AVR-Pia与RGA5的相互作用是使水稻触发ETI的关键因素[28];玉米黑粉菌(Ustilago maydis)中,效应蛋白Pep1直接抑制寄主过氧化物酶POX12,从而抑制由过氧化物酶驱动的氧化爆发[29];在灰霉菌(Botrytis cinerea)中,效应蛋白BcPTP1基因在侵染的晚期阶段高表达,并且与灰霉的致病力密切相关[30]。因此,发掘和研究橡胶树胶孢炭疽菌潜在的效应蛋白对阐明其侵染橡胶树的分子机制和发现新的植物抗病靶标具有重要意义。
由于真菌效应蛋白一般缺乏序列相似性和保守的结构域,因此,具有预测上的困难性[31]。效应蛋白通常具有分泌性、分子质量小(通常小于300个氨基酸)[32]、富含半胱氨酸[33]等特点。真菌效应蛋白的功能验证可以利用农杆菌浸润法在本氏烟叶片中共表达植物抗性蛋白(如INF1或BAX)和效应蛋白来实现,通过评估过敏反应以及细胞器靶向等功能来验证[34],如在山田胶锈菌(Gymnosporangium yamadae)中GyS-2、GyS-3等5个候选效应蛋白能够引起本氏烟草叶片坏死[35];在杨树溃疡菌(Cytospora chrysosperma)中,通过农杆菌侵染本氏烟草叶片观察亚细胞定位,确认效应蛋白CcCAP1的免疫抑制活性与其核定位有关[36]。本实验在前期研究工作中预测,Cg694和Cg2346是潜在的胶孢炭疽菌效应蛋白,并通过在烟草叶片上瞬时表达Cg694和Cg2346基因来观察亚细胞定位。研究发现,Cg694和Cg2346定位于细胞核和细胞膜,且瞬时表达Cg694和Cg2346基因会诱导烟草叶片出现坏死,这表明Cg694和Cg2346会引起植物免疫反应。在茶树炭疽菌(C. camelliae)中,发现预测的多个候选效应蛋白在附着胞时期表达量较高[37]。因此,本研究推测候选效应蛋白在附着胞时期表达量上调可能与胶孢炭疽菌的侵染结构形成有关。Cg694和Cg2346基因的表达谱分析表明,相比在菌丝时期的表达,它们在胶孢炭疽菌孢子、萌发和附着胞等发育阶段的表达量明显上调,这表明在胶孢炭疽菌的发育阶段,Cg694和Cg2346可能参与调控胶孢炭疽菌孢子的萌发和附着胞结构的形成。同时,在侵染橡胶树组织时期,Cg694和Cg2346基因的表达量显著下调,推测可能是受到植物防御机制的影响。在杨树溃疡菌(C. chrysosperma)中,效应蛋白CcSge1基因的缺失会显著降低分生孢子产量以及影响毒力相关基因的表达[38];在辣椒炭疽菌(C. gloeosporioides)中,效应蛋白Cghn13741基因的缺失影响了菌丝的极性生长并导致产孢量下降[39]。为了进一步探究Cg694和Cg2346对胶孢炭疽菌生理表型的调控作用,本研究通过分别构建Cg694和Cg2346基因敲除突变体分析其表型差异。研究结果表明,Cg694和Cg2346不参与调控橡胶树胶孢炭疽菌的生长表型;但Cg694参与对孢子产量的调控。本研究通过接种完整和预刺伤处理的淡绿期橡胶树叶片发现,Cg694和Cg2346基因的缺失没有引起胶胞炭疽菌的致病力的变化,这说明Cg694和Cg2346并不是胶孢炭疽菌的关键毒性因子。接下来,本研究了橡胶树胶孢炭疽菌侵染发展的不同阶段,对突变体的萌发率、附着胞形成率以及入侵菌丝形成率进行了观察和研究。发现 Cg694和Cg2346参与调控芽管的极性生长,Cg694不参与调控附着胞的形成,而Cg2346参与调控附着胞的形成。Cg694和Cg2346基因的缺失都在侵染的前期导致了入侵菌丝的形成率降低,说明Cg694和Cg2346可能在入侵的前期参与调控橡胶树胶孢炭疽菌入侵菌丝的形成。因此,结合对突变体的萌发率、附着胞形成率以及入侵菌丝形成率进行分析,本研究推测,Cg694和Cg2346基因的缺失没有引起胶胞炭疽菌的致病力变化的原因可能有2点:1)Cg694和Cg2346本身不具有致病的能力,使得敲除后致病力没有受到影响;2)它们虽然参与调控芽管的极性生长和前期入侵结构的形成,但是并没有降低孢子的萌发能力以及影响后续入侵结构的形成,使得整体的侵染能力没有出现明显降低,导致致病力没有发生变化。因此本研究推测,Cg694和Cg2346参与调控芽管极性生长和侵染前期入侵结构的形成,但其靶标蛋白以及调控机制有待进一步深入验证。
-
研究Cg694和Cg2346基因的表达谱发现,Cg694和Cg2346基因在胶孢炭疽菌孢子、萌发和附着胞发育阶段的相对表达量较高。本研究通过农杆菌浸润法在本氏烟草叶片上表达了Cg694和Cg2346,发现其定位于细胞核和细胞膜,且其瞬时过表达会诱导烟草叶片出现坏死。针对基因敲除突变体的生理表型与致病能力进行分析发现,Cg694和Cg2346参与橡胶树胶孢炭疽菌的孢子极性生长、早期入侵结构形成的调控,Cg694对孢子产量有调控作用,但是Cg694和Cg2346基因的缺失没有引起突变体致病力的改变。Cg694和Cg2346影响胶孢炭疽菌生理表型的分子机制有待深入研究。
Functional analysis of candidate effector proteins Cg694 and Cg2346 of Colletotrichum gloeosporioides from Hevea brasiliensis
-
摘要: 胶胞炭疽菌(Colletotrichum gloeosporioides)在对橡胶树的侵染过程中会分泌许多蛋白效应子,但是目前关于它们在克服植物免疫中发挥的作用仍知之甚少。开展橡胶树胶孢炭疽菌潜在效应蛋白的研究可以为炭疽病害的防治提供理论基础。本研究在橡胶树胶孢炭疽菌中预测到2个潜在效应蛋白Cg694和Cg2346,实时荧光定量分析显示,与胶孢炭疽菌营养菌丝阶段相比,Cg694和Cg2346基因在孢子、萌发和附着胞发育阶段表达量显著上调。通过烟草(Nicotiana benthamiana)表皮细胞中瞬时表达体系(绿色荧光蛋白Green fluorescent protein)对其亚细胞定位情况进行了观察,结果发现,Cg694和Cg2346基因均定位在细胞核和细胞膜,且它们的瞬时表达会诱导烟草叶片出现坏死。随后,分别构建了这2个基因的敲除突变体ΔCg694和ΔCg2346,并对其营养生长以及致病相关表型进行了分析。研究发现,ΔCg694和ΔCg2346基因敲除突变体的萌发芽管极性生长的能力增强、在洋葱(Allium cepa)表皮细胞中的前期的入侵菌丝形成率降低,其中,ΔCg2346突变体的附着胞形成能力降低,但ΔCg694和ΔCg2346突变体对橡胶树叶片上的致病力均没有出现显著变化。实验结果显示,Cg694和Cg2346会引起胶胞炭疽菌的芽管极性生长能力增强和前期入侵能力减弱等表型的改变,但其潜在的功能有待进一步研究。Abstract: Natural rubber is a critical strategic resource in China, and anthracnose of rubber trees caused by Colletotrichum gloeosporioides directly threatens the healthy development of the rubber industry. During infection, C. gloeosporioides secretes numerous protein effectors, and their roles in overcoming plant immunity remain poorly understood. Investigating potential effector proteins in C. gloeosporioides can provide a theoretical basis for controlling anthracnose. In this context, two candidate effector proteins, Cg694 and Cg2346, were predicted in C. gloeosporioides. Quantitative real-time PCR revealed that the transcriptional expression of the genes Cg694 and Cg2346 was significantly upregulated during the development stages of spore, germination, and appressorium compared with the vegetative hyphal stage. Subcellular localization assays via transient expression (Green fluorescent protein) in Nicotiana benthamiana epidermal cells showed that both Cg694 and Cg2346 genes were localized to the nucleus and cell membrane, and their transient overexpression induced necrosis in tobacco leaves. Furthermore, gene knockout mutants ΔCg694 and ΔCg2346 were generated. Phenotypic analysis indicated that ΔCg694 and ΔCg2346 exhibit enhanced germ tube polar growth and reduced early invasive hyphal formation rates in onion epidermal cells. Notably, the ΔCg2346 mutant also show a decreased ability to form appressoria. However, neither ΔCg694 nor ΔCg2346 mutants display significant changes in pathogenicity on rubber tree leaves. These results demonstrate that Cg694 and Cg2346 are involved in modulating the polar growth and early invasion capabilities of C. gloeosporioides, though their precise functions require further investigation.
-
Key words:
- C. gloeosporioides /
- effector protein /
- appressorium /
- pathogenicity
-
图 2 Cg694和Cg2346基因在胶孢炭疽菌不同时期的相对表达量检测
注: A. Cg694在胶孢炭疽菌不同发育阶段的相对表达量检测; B. Cg2346在胶孢炭疽菌不同发育阶段的相对表达量检测;C. Cg694在胶孢炭疽菌侵染橡胶树叶片的不同时期相对表达量检测;D. Cg2346在胶孢炭疽菌侵染橡胶树叶片的不同时期相对表达量检测。误差线表示样品平均值±标准差SD; 显著差异分析:ns: P>0.05;*: P<0.05;**: P<0.01;***: P<0.001,下同。
Fig. 2 Detection of relative expression levels of genes Cg694 and Cg2346 in C. gloeosporioides at different developmental stages
Note: A. Relative expression levels of Cg694 at different developmental stages of C. gloeosporioides; B. Relative expression levels of Cg2346 at different developmental stages of C. gloeosporioides ; C. Relative expression levels of Cg694 at different time points during C. gloeosporioides infection of rubber tree leaves; D. Relative expression levels of Cg2346 at different time points during C. gloeosporioides infection of rubber tree leaves. Error bars represent the mean ± standard deviation (SD)of the samples. Significant difference analysis: ns. P>0.05; *. P<0.05; **. P<0.01; ***. P<0.001;****. P<0.0001, similarly hereinafter.
图 3 烟草叶片瞬时表达Cg694和Cg2346基因
注: A. pEGAD-GFP,pEGAD-694-GFP,pEGAD-2346-GFP 烟草亚细胞定位观察B. Cg694基因在烟草叶片上瞬时表达;C. Cg2346基因在烟草叶片上瞬时表达。Bars=50 μm。
Fig. 3 Transient expression of genes Cg694 and Cg2346 in N. benthamiana leaves
Note: A. Subcellular localization observation of pEGAD-GFP, pEGAD-694-GFP, and pEGAD-2346-GFP in tobacco; B. Transient expression of the Cg694 gene in tobacco leaves; C. Transient expression of the Cg2346 gene in tobacco leaves. Bars=50 μm.
图 4 Split-Marker基因敲除原理以及基因敲除突变体的鉴定
注:A. Split-Marker基因敲除原理示意图;B. Cg694基因敲除突变体上、下游同源臂和基因检测(顺序由上到下);C. Cg2346基因敲除突变体上、下游同源臂和基因检测(顺序由左到右)。
Fig. 4 Principle of Split-Marker gene knockout and identification of knockout mutants
Note: A. Schematic diagram of the Split-Marker gene knockout principle; B. Detection of upstream and downstream homologous arms and the gene in Cg694 gene knockout mutants (order from top to bottom); C. Detection of upstream and downstream homologous arms and the gene in Cg2346 gene knockout mutants (order from left to right).
图 5 ΔCg694和ΔCg2346突变体生长速率与产孢量统计
注: A. WT和ΔCg694突变体在PDA培养基上生长5 d后的菌落形态;B. WT和ΔCg694突变体5 d 后菌落直径统计;C. WT和ΔCg694突变体产孢量统计;D. WT和ΔCg2346突变体在PDA培养基上生长5 d后的菌落形态;E. WT和ΔCg2346突变体5 d后菌落直径统计;F. WT和ΔCg2346突变体产孢量统计。
Fig. 5 Growth rate and sporulation quantification of ΔCg694 and ΔCg2346 mutant strains
Note: A. Colony morphology of WT and ΔCg694 mutant after 5 days of growth on PDA medium; B. Statistics of colony diameter for WT and ΔCg694 mutant after 5 days; C. Statistics of sporulation for WT and ΔCg694 mutant; D. Colony morphology of WT and ΔCg2346 mutant after 5 days of growth on PDA medium; E. Statistics of colony diameter for WT and ΔCg2346 mutant after 5 days; F. Statistics of sporulation for WT and ΔCg2346 mutant.
图 6 ΔCg694和ΔCg2346突变体致病力分析
注: A. ΔCg694突变体接种完整橡胶树叶片3 d发病情况; B. ΔCg694突变体接种完整橡胶树叶片3 d发病率; C. ΔCg694突变体接种完整橡胶树叶片3 d病斑大小差异分析; D. ΔCg694突变体接种预刺伤处理橡胶树叶片3 d发病情况; E. ΔCg694突变体接种预刺伤橡胶树叶片3 d发病率;F. ΔCg694突变体接种橡胶树预刺伤叶片3 d病斑大小差异分析;G. ΔCg2346突变体接种完整橡胶树叶片3 d发病情况; H. ΔCg2346突变体接种完整橡胶树叶片3 d发病率;I. ΔCg2346突变体接种完整橡胶树叶片3 d病斑大小差异分析; J. ΔCg2346突变体接种预刺伤处理橡胶树叶片3 d发病情况;K. ΔCg2346突变体接种预刺伤橡胶树叶片3 d发病率;L. ΔCg2346突变体接种预刺伤橡胶树叶片3 d病斑大小差异分析。
Fig. 6 Pathogenicity analysis of ΔCg694 and ΔCg2346 strains
Note: A. Disease symptoms of ΔCg694 mutant inoculated on intact rubber tree leaves at 3 days post-inoculation (dpi); B. Disease incidence of ΔCg694 mutant inoculated on intact rubber tree leaves at 3 dpi; C. Analysis of lesion size differences caused by ΔCg694 mutant on intact rubber tree leaves at 3 dpi; D. Disease symptoms of ΔCg694 mutant inoculated on pre-wounded rubber tree leaves at 3 dpi; E. Disease incidence of ΔCg694 mutant inoculated on pre-wounded rubber tree leaves at 3 dpi; F. Analysis of lesion size differences caused by ΔCg694 mutant on pre-wounded rubber tree leaves at 3 dpi; G. Disease symptoms of ΔCg2346 mutant inoculated on intact rubber tree leaves at 3 dpi; H. Disease incidence of ΔCg2346 mutant inoculated on intact rubber tree leaves at 3 dpi; I: Analysis of lesion size differences caused by ΔCg2346 mutant on intact rubber tree leaves at 3 dpi; J: Disease symptoms of ΔCg2346 mutant inoculated on pre-wounded rubber tree leaves at 3 dpi; K. Disease incidence of ΔCg2346 mutant inoculated on pre-wounded rubber tree leaves at 3 dpi; L: Analysis of lesion size differences caused by ΔCg2346 mutant on pre-wounded rubber tree leaves at 3 dpi.
图 7 ΔCg694和ΔCg2346突变体孢子萌发情况分析
注: A. ΔCg694萌发2 h、3 h和6 h情况观察; B. ΔCg694培养2 h、3 h和6 h的萌发率统计;C. ΔCg694萌发2 h、3 h和6 h多极萌发率统计;D. ΔCg694萌发2 h、3 h和6 h芽管分枝率统计;E. ΔCg2346萌发2 h、3 h和6 h情况观察;F. ΔCg2346培养2 h、3 h和6 h的萌发率统计;G. ΔCg2346萌发2 h、3 h和6 h多极萌发率统计;H. ΔCg2346萌发2 h、3 h和6 h芽管分枝率统计;黑色箭头表示多端萌发,红色箭头表示芽管分枝,每组样品至少包含100个分生孢子。Bars=10 μm。
Fig. 7 Analysis of spore germination characterization in ΔCg694和ΔCg2346 strains
Note: A. Observation of ΔCg694 germination at 2 h, 3 h, and 6 h; B. Germination rate statistics of ΔCg694 cultured for 2 h, 3 h, and 6 h; C. Statistics of multipolar germination rate for ΔCg694 at 2 h, 3 h, and 6 h; D. Statistics of germ tube branching rate for ΔCg694 at 2 h, 3 h, and 6 h. E. Observation of ΔCg2346 germination at 2 h, 3 h, and 6 h; F. Germination rate statistics of ΔCg2346 cultured for 2 h, 3 h, and 6 h; G. Statistics of multipolar germination rate for ΔCg2346 at 2 h, 3 h, and 6 h; H. Statistics of germ tube branching rate for ΔCg2346 at 2 h, 3 h, and 6 h. Black arrows indicate multipolar germination, and red arrows indicate germ tube branching. At least 100 conidia were included for each sample. Bars=10 μm.
图 8 ΔCg694和ΔCg2346突变体附着胞形成率分析
注: A. 疏水平板ΔCg694附着胞形成情况; B. ΔCg694附着胞形成率统计; C. 疏水平板ΔCg2346附着胞形成情况; D. ΔCg2346附着胞形成率统计。Bars=10 μm。
Fig. 8 Analysis of appressorium formation rate in ΔCg694和ΔCg2346 strains
Note: A. Appressoria formation of ΔCg694 on hydrophobic surface; B. Appressoria formation rate of ΔCg694; C. Appressoria formation of ΔCg2346 on hydrophobic surface; D. Appressoria formation rate of ΔCg2346. Bars=10 μm.
图 9 ΔCg694和ΔCg2346突变体入侵菌丝形成率分析
注: A. ΔCg694接种洋葱内表皮入侵菌丝形成情况;B; ΔCg694入侵菌丝形成率统计;C. ΔCg2346接种洋葱内表皮入侵菌丝形成情况;D; ΔCg2346入侵菌丝形成率统计;E.不同类型侵染菌丝在洋葱细胞中的百分比。1型:只有附着胞;2型:只有一个入侵菌丝(IH);3型:单个侵染结构的入侵菌丝有2个分支;类型4:单个侵染结构的入侵菌丝有有2个以上分支。每组样品至少包含100个分生孢子。 误差线表示样品平均值±标准差SD; ns: P>0.05; *: P<0.05; ****: P<0.0001; Bar=10 μm。
Fig. 9 Analysis of invasive hypha formation rate in ΔCg694和ΔCg2346 strains
Note: A.Formation of invasive hyphae by ΔCg694 inoculated on onion epidermal cells; B. Statistical analysis of invasive hypha formation rate in ΔCg694; C. Formation of invasive hyphae by ΔCg694 inoculated on onion epidermal cells; D. Statistical analysis of invasive hypha formation rate in ΔCg694; E. Percentage of different types of infection hyphae within onion cells. Type 1. Only appressorium; Type 2. Only one invasive hypha (IH); Type 3. Two branches from a single infection structure; Type 4. More than two branches from a single infection structure. Bar=10 μm.
表 1 本实验所用引物
Table 1 Primers used in this experiment
引物名称Primer 序列(5′−3′)Sequence 用途Usage Cg694-5F CTATTGCTTGTCTTGTTCCT 同源臂扩增
Homologous arm amplificationCg694-5R AGGCAGCGTATACCGACCTG 同源臂扩增
Homologous arm amplificationCg694-MF1 GACGCAATCTTCTACAGAACGTGCCAACGCCACAGTGCCC 同源臂扩增
Homologous arm amplificationCg694-MR1 GGGCACTGTGGCGTTGGCACGTTCTGTAGAAGATTGCGTC 同源臂扩增
Homologous arm amplificationCg694-MF2 CGGGAATTGCATGCTCTCAC AATTTTGTGAAGCGCAAAGC 同源臂扩增
Homologous arm amplificationCg694-MR2 GCTTTGCGCTTCACAAAATT GTGAGAGCATGCAATTCCCG 同源臂扩增
Homologous arm amplificationCg2346-5F AGGCCACCTGCAGCGCTTGAAC 同源臂扩增
Homologous arm amplificationCg2346-5R GGAAAATTGGTGCTCTTGCTGC 同源臂扩增
Homologous arm amplificationCg2346-MF1 ATCCGGCGTCTTTTCCATCACCGTGCCAACGCCACAGTGCCC 同源臂扩增
Homologous arm amplificationCg2346-MR1 GGGCACTGTGGCGTTGGCACGGTGATGGAAAAGACGCCGGAT 同源臂扩增
Homologous arm amplificationCg2346-MF2 CGGGAATTGCATGCTCTCACACATCCGTTGGGGTTGTGTTGA 同源臂扩增
Homologous arm amplificationCg2346-MR2 TCAACACAACCCCAACGGATGTGTGAGAGCATGCAATTCCCG 同源臂扩增
Homologous arm amplificationSur-SLR ATGTTGGCATAAGCCGAACCGT 同源臂扩增
Homologous arm amplificationSur-SLF CCTCTGATATTGGAAGCGACGC 同源臂扩增
Homologous arm amplificationCg694-JC5F CTCTAATCCGCCAAATCCTGAG 突变体鉴定
Mutant Strain IdentificationCg694-JC3R GATGGTTCGTGAACATCCCTTG 突变体鉴定
Mutant Strain IdentificationCg694-JCOF CCGAGCGTAGTGGTCTCATCAA 突变体鉴定
Mutant Strain IdentificationCg694-JCOR AGGCTTCAGCCGCAATTACATC 突变体鉴定
Mutant Strain IdentificationCg694-JCGF ATGCCGGCCGACTACTCCGCTC 突变体鉴定
Mutant Strain IdentificationCg694-JCGR AACATCATGGCCGTCGGAACCG 突变体鉴定
Mutant Strain IdentificationCg2346-JC5F GTAATGAGACAACAGTCGGGCA 突变体鉴定
Mutant Strain IdentificationCg2346-JC3R ACAGAGACGAAAAGACAAGGCG 突变体鉴定
Mutant Strain IdentificationCg2346-JCOF AGGACCGCTACTCACATACACA 突变体鉴定
Mutant Strain IdentificationCg2346-JCOR TTAAAGTTCGAGGGATGACACA 突变体鉴定
Mutant Strain IdentificationCg2346-JCGR AAGAAGAACAACAACGGAAAGA 突变体鉴定
Mutant Strain IdentificationCg2346-JCGF ATCAGAGTGGAGTGACAAGAGT 突变体鉴定
Mutant Strain IdentificationCg694-OF(EcoRI) CTGCGGCAGCGGCCGAATTCATGCCGGCCGACTACTCCGC 同源臂扩增
Homologous arm amplificationCg694-OR(HindIII) GATCCGGTGGATCCAAGCTTAACATCATGGCCGTCGGAAC 同源臂扩增
Homologous arm amplificationCg2346-OF(EcoRI) CTGCGGCAGCGGCCGAATTCATGAAGGCAGTTTCCGTTCT 同源臂扩增
Homologous arm amplificationCg2346-OR(HindIII) GATCCGGTGGATCCAAGCTTGAACAGGCGAGCAGCGGCAG 同源臂扩增
Homologous arm amplificationCgActin-qF GATTGGTATGGGCCAGAAGG 相对表达量检测
Relative expression level detectionCgActin-qR GGAGCTCGTTGTAGAAGGTG 相对表达量检测
Relative expression level detectionCg694-QF TCAGAAATTCAGCCCCCGTC 相对表达量检测
Relative expression level detectionCg694-QR AACTCAGCGGTAAGCTCGTC 相对表达量检测
Relative expression level detectionCg2346-QF AGGAAACAACAATGGCACCG 相对表达量检测
Relative expression level detectionCg694-QR CAGCAGCAGAGCCACTAGAA 相对表达量检测
Relative expression level detection 下载: 导出CSV
下载: 导出CSV
-
[1] Cai Z Y, Li G H, Lin C H, et al. Identifying pathogenicity genes in the rubber tree anthracnose fungus Colletotrichum gloeosporioides through random insertional mutagenesis [J]. Microbiological Research, 2013, 168(6): 340−350. https://doi.org/10.1016/j.micres.2013.01.005 doi: 10.1016/j.micres.2013.01.005 [2] 冯淑芬, 刘秀娟, 郑服丛, 等. 橡胶树炭疽菌生物学和侵染特征研究[J]. 热带作物学报, 1998, 19(2): 7−14. [3] Liu L P, Zhang L, Qiu P L, et al. Leaf spot of Polygonatum odoratum caused by Colletotrichum spaethianum [J]. Journal of General Plant Pathology, 2020, 86(2): 157−161. https://doi.org/10.1007/s10327-019-00903-4 doi: 10.1007/s10327-019-00903-4 [4] 李博勋, 刘先宝, 陈丽琼, 等. 橡胶树主要病害研究现状与展望[J]. 中国科学: 生命科学, 2024, 54(10): 1798−1813. https://doi.org/10.1360/SSV-2024-0194 doi: 10.1360/SSV-2024-0194 [5] 韩长志. 胶孢炭疽菌侵染过程相关基因研究进展[J]. 广东农业科学, 2014, 41(9): 165−169. https://doi.org/10.16768/j.issn.1004-874x.2014.09.020 doi: 10.16768/j.issn.1004-874x.2014.09.020 [6] Perfect S E, Hughes H B, O'connell R J, et al. Colletotrichum: a model genus for studies on pathology and fungal–plant interactions [J]. Fungal Genetics and Biology, 1999, 27(2/3): 186−198. https://doi.org/10.1006/fgbi.1999.1143 doi: 10.1006/fgbi.1999.1143 [7] Chisholm S T, Coaker G, Day B, et al. Host-microbe interactions: shaping the evolution of the plant immune response [J]. Cell, 2006, 124(4): 803−814. https://doi.org/10.1016/j.cell.2006.02.008 doi: 10.1016/j.cell.2006.02.008 [8] Jones J D G, Dangl J L. The plant immune system [J]. Nature, 2006, 444(7117): 323−329. https://doi.org/10.1038/nature05286 doi: 10.1038/nature05286 [9] Boller T, He S Y. Innate immunity in plants: an arms race between pattern recognition receptors in plants and effectors in microbial pathogens [J]. Science, 2009, 324(5928): 742−744. https://doi.org/10.1126/science.1171647 doi: 10.1126/science.1171647 [10] Dangl J L, Horvath D M, Staskawicz B J. Pivoting the plant immune system from dissection to deployment [J]. Science, 2013, 341(6147): 746−751. https://doi.org/10.1126/science.1236011 doi: 10.1126/science.1236011 [11] Cui H T, Tsuda K, Parker J E. Effector-triggered immunity: from pathogen perception to robust defense [J]. Annual Review of Plant Biology, 2015, 66: 487−511. https://doi.org/10.1146/annurev-arplant-050213-040012 doi: 10.1146/annurev-arplant-050213-040012 [12] Wu L, Chen H, Curtis C, et al. Go in for the kill: how plants deploy effector-triggered immunity to combat pathogens [J]. Virulence, 2014, 5(7): 710−721. https://doi.org/10.4161/viru.29755 doi: 10.4161/viru.29755 [13] Oh Y, Donofrio N, Pan H Q, et al. Transcriptome analysis reveals new insight into appressorium formation and function in the rice blast fungus Magnaporthe oryzae [J]. Genome Biology, 2008, 9(5): R85. https://doi.org/10.1186/gb-2008-9-5-r85 doi: 10.1186/gb-2008-9-5-r85 [14] 王美玉. 苹果炭疽叶枯病菌效应蛋白Sntf2抑制植物免疫的分子机制研究[D]. 北京: 中国农业科学院, 2022. doi: 10.27630/d.cnki.gznky.2022.000109 [15] 韩艺娟, 钟振晖, 吴剑英, 等. 稻瘟病菌类LxAR家族基因的预测及表达分析[J]. 热带作物学报, 2018, 39(5): 979−986. https://doi.org/10.3969/j.issn.1000-2561.2018.05.023 doi: 10.3969/j.issn.1000-2561.2018.05.023 [16] Saier M H, Jr, Tran C V, Barabote R D. TCDB: the Transporter Classification Database for membrane transport protein analyses and information [J]. Nucleic Acids Research, 2006, 34(S1): D181−D186. https://doi.org/10.1093/nar/gkj001 doi: 10.1093/nar/gkj001 [17] Yan N E. Structural biology of the major facilitator superfamily transporters [J]. Annual Review of Biophysics, 2015, 44: 257−283. https://doi.org/10.1146/annurev-biophys-060414-033901 doi: 10.1146/annurev-biophys-060414-033901 [18] Chen L H, Tsai H C, Yu P L, et al. A major facilitator superfamily transporter-mediated resistance to oxidative stress and fungicides requires Yap1, Skn7, and MAP kinases in the citrus fungal pathogen Alternaria alternata [J]. PLoS One, 2017, 12(1): e0169103. https://doi.org/10.1371/journal.pone.0169103 doi: 10.1371/journal.pone.0169103 [19] Calabrese D, Bille J, Sanglard D. A novel multidrug efflux transporter gene of the major facilitator superfamily from Candida albicans (FLU1) conferring resistance to fluconazole [J]. Microbiology, 2000, 146(11): 2743−2754. https://doi.org/10.1099/00221287-146-11-2743 doi: 10.1099/00221287-146-11-2743 [20] Liu N, Wang Q N, He C Z, et al. CgMFS1, a major facilitator superfamily transporter, is required for sugar transport, oxidative stress resistance, and pathogenicity of Colletotrichum gloeosporioides from Hevea brasiliensis [J]. Current Issues in Molecular Biology, 2021, 43(3): 1548−1557. https://doi.org/10.3390/cimb43030109 doi: 10.3390/cimb43030109 [21] 郭云峰, 安邦. 橡胶树胶孢炭疽菌NADPH氧化酶功能研究[J]. 生物技术通报, 2018, 34(10): 165−171. https://doi.org/10.13560/j.cnki.biotech.bull.1985.2018-0344 doi: 10.13560/j.cnki.biotech.bull.1985.2018-0344 [22] Drew D, North R A, Nagarathinam K, et al. Structures and general transport mechanisms by the Major Facilitator Superfamily (MFS) [J]. Chemical Reviews, 2021, 121(9): 5289−5335. https://doi.org/10.1021/acs.chemrev.0c00983 doi: 10.1021/acs.chemrev.0c00983 [23] Zhang S Y, Li C, Si J P, et al. Action mechanisms of effectors in plant-pathogen interaction [J]. International Journal of Molecular Sciences, 2022, 23(12): 6758. https://doi.org/10.3390/ijms23126758 doi: 10.3390/ijms23126758 [24] Langin G, Gouguet P, Üstün S. Microbial effector proteins-a journey through the proteolytic landscape [J]. Trends in Microbiology, 2020, 28(7): 523−535. https://doi.org/10.1016/j.tim.2020.02.010 doi: 10.1016/j.tim.2020.02.010 [25] Flor H H. Inheritance of pathogenicity in Melampsora lini [J]. Phytopathology® , 2025, 115(8V): 653−669. https://doi.org/10.1094/Phyto-32-653 doi: 10.1094/Phyto-32-653 [26] De Wit P J G M, Mehrabi R, Van Den Burg H A, et al. Fungal effector proteins: past, present and future [J]. Molecular Plant Pathology, 2009, 10(6): 735−747. https://doi.org/10.1111/j.1364-3703.2009.00591.x doi: 10.1111/j.1364-3703.2009.00591.x [27] De Wit P J G M. How plants recognize pathogens and defend themselves [J]. Cellular and Molecular Life Sciences, 2007, 64(21): 2726−2732. https://doi.org/10.1007/s00018-007-7284-7 doi: 10.1007/s00018-007-7284-7 [28] Ortiz D, De Guillen K, Cesari S, et al. Recognition of the Magnaporthe oryzae effector AVR-Pia by the decoy domain of the rice NLR immune receptor RGA5 [J]. The Plant Cell, 2017, 29(1): 156−168. https://doi.org/10.1105/tpc.16.00435 doi: 10.1105/tpc.16.00435 [29] Hemetsberger C, Herrberger C, Zechmann B, et al. The Ustilago maydis effector Pep1 suppresses plant immunity by inhibition of host peroxidase activity [J]. PLoS Pathogens, 2012, 8(5): e1002684. https://doi.org/10.1371/journal.ppat.1002684 doi: 10.1371/journal.ppat.1002684 [30] Zhu W J, Yu M X, Xu R, et al. Botrytis cinerea BcSSP2 protein is a late infection phase, cytotoxic effector [J]. Environmental Microbiology, 2022, 24(8): 3420−3435. https://doi.org/10.1111/1462-2920.15919 doi: 10.1111/1462-2920.15919 [31] Sperschneider J, Dodds P N, Gardiner D M, et al. Advances and challenges in computational prediction of effectors from plant pathogenic fungi [J]. PLoS Pathogens, 2015, 11(5): e1004806. https://doi.org/10.1371/journal.ppat.1004806 doi: 10.1371/journal.ppat.1004806 [32] Thatcher L F, Williams A H, Garg G, et al. Transcriptome analysis of the fungal pathogen Fusarium oxysporum f. sp. medicaginis during colonisation of resistant and susceptible Medicago truncatula hosts identifies differential pathogenicity profiles and novel candidate effectors [J]. BMC Genomics, 2016, 17(1): 860. https://doi.org/10.1186/s12864-016-3192-2 doi: 10.1186/s12864-016-3192-2 [33] Liu T L, Song T Q, Zhang X, et al. Unconventionally secreted effectors of two filamentous pathogens target plant salicylate biosynthesis [J]. Nature Communications, 2014, 5: 4686. https://doi.org/10.1038/ncomms5686 doi: 10.1038/ncomms5686 [34] Lovelace A H, Dorhmi S, Hulin M T, et al. Effector identification in plant pathogens [J]. Phytopathology® , 2023, 113(4): 637−650. https://doi.org/10.1094/PHYTO-09-22-0337-KD doi: 10.1094/PHYTO-09-22-0337-KD [35] 金之鑫, 张晓漫, 梁英梅. 山田胶锈菌性、锈孢子阶段效应蛋白的预测和筛选[J]. 菌物学报, 2025, 44(6): 240341. https://doi.org/10.13346/j.mycosystema.240341 doi: 10.13346/j.mycosystema.240341 [36] Han Z, Xiong D G, Xu Z Y, et al. The Cytospora chrysosperma virulence effector CcCAP1 mainly localizes to the plant nucleus to suppress plant immune responses [J]. mSphere, 2021, 6(1): e00883−20. https://doi.org/10.1128/mSphere.00883-20 doi: 10.1128/mSphere.00883-20 [37] 熊飞. 基于转录组测序的茶树炭疽菌致病机理初探[D]. 南京: 南京农业大学, 2020. https://doi.org/10.27244/d.cnki.gnjnu.2020.001851 [38] Han Z, Yu R, Xiong D G, et al. A Sge1 homolog in Cytospora chrysosperma governs conidiation, virulence and the expression of putative effectors [J]. Gene, 2021, 778: 145474. https://doi.org/10.1016/j.gene.2021.145474 doi: 10.1016/j.gene.2021.145474 [39] 盛家伟. 辣椒胶孢炭疽病菌基因Cghn01197和Cghn13741对生物学功能的影响[D]. 长沙: 湖南农业大学, 2022. https://doi.org/10.27136/d.cnki.ghunu.2022.001061 -

 点击查看大图
点击查看大图
计量
- 文章访问数: 337
- HTML全文浏览量: 186
- 被引次数: 0
