

-
水稻(Oryza sativa L.)作为全球重要的粮食作物之一,具有种植历史悠久、栽培范围广的特点[1]。水稻栽培过程中受非生物胁迫和生物胁迫等许多因素的干扰,其中病害是导致水稻严重减产的重要因素之一[2 − 3]。由稻属黄单胞菌水稻致病变种(Xanthomonas oryzae pv. oryzae,Xoo)引起的水稻白叶枯病是中国水稻生产中的三大病害之一[4 − 5]。水稻白叶枯病侵染范围极为广泛,会造成高达20%至30%的大量减产,严重时减产可能达到50%,甚至颗粒无收[6 − 8]。
分泌系统对于黄单胞菌侵染植物至关重要,众多效应蛋白和致病因子通过分泌系统传送至宿主体内,达到侵染的目的。目前已发现的分泌系统共有9类,包括Ⅰ型至Ⅸ型,其中Ⅲ型分泌系统(Type Ⅲ Secretion System,T3SS)的相关研究最为深入[9]。HrpX和HrpG是T3SS表达的关键因子,HrpG激活HrpX的表达,然后HrpX结合到PIP-box位点,进而调控效应蛋白基因的转录。T3SEs由T3SS分泌到宿主植物中,影响细胞信息传递和调控网络,改变宿主细胞的生理生化过程[10 − 12]。例如,XopD在植物体内展现出类泛素化修饰蛋白蛋白酶活性抑制宿主转录,参与调控蛋白质的稳定性[13];XopL具有E3泛素连接酶活性,可以通过抑制植物细胞死亡及防卫基因表达从而抑制PTI (PRR-Triggered Immunity)过程[14 − 15];在本氏烟草中XopQ的表达会引发依赖于识别XopQ1 (ROQ1抗原体)的效应子触发ETI (Effector-Triggered Immunity)免疫反应,并伴随质体在细胞核周围的积累以及叶绿体嵴突的形成[16 − 18]。因此,解析T3SEs的功能及其调控途径对于研究病原菌与植物相互作用至关重要。
xopG基因全长468 bp,编码155个氨基酸。XopG可抑制XopQ-XopX诱导的水稻免疫应答,Deb等研究表明XopG与XopQ、XopX均存在相互作用[19 − 20]。但到目前为止,XopG在水稻中的互作靶标蛋白仍然不清楚。
本研究对XopG进行了生物信息学分析,同时通过启动子活力测定对该基因的表达进行评估,对水稻应用剪叶法进行接种实验分析xopG突变体的致病力。实验结果初步说明xopG表达受HrpX正调控,且缺失xopG后Xoo致病力显著降低,为解析XopG与水稻互作机制提供了重要的理论基础。
-
供试水稻TP309、本氏烟草(Nicotiana benthamiana)由本实验室保存,本实验所用的菌株和质粒如表1所示。
表 1 实验所用菌株与质粒
Table 1. Experimental strains and plasmids
菌株/质粒 Strain/plasmid 相关特征 Relevant characteristics 来源 Source 大肠杆菌 Escherichia coli DH5α F-φ80lacZΔM15 Δ(lacZYA-argF)u169 recA1 endA1
hsdR17(rk-, mk+)phoA supE44 λ- thi-1 gyrA96 relA1上海唯地生物
Shanghai Weidi Biotechnology农杆菌 Agrobacterium GV3101 C58(rifR)TipMP90(pTiC58DT-DNA)
(gentR)Nopaline上海唯地生物
Shanghai Weidi Biotechnology黄单胞菌 Xanthomonas oryzae pv. oryzae T7174 Xoo strain 本实验室
This StudyΔxopG Full length deletion mutant of xopG 本实验室
This StudyC-ΔxopG The complementation strain of ΔxopG 本实验室
This StudyΔhrpX Full length deletion mutant of hrpX(PXO_01953) 本实验室
This Study质粒 Plasmids pK18mobsacB Suicide plasmid in Xanthomonas, KanR Schäfe et al., 1994[21] pK18-xopG pK18mobsacB based plasmid for xopG deletion 本实验室
This StudypHM1 broad-host range vector, SpR Huynh et al., 1989[22] pHM1-xopG Complementary vector containing
full length xopG gene, SpR本实验室
This StudypRTVCGFP Transient over-expression, Ubi promoter,
N terminal EGFP Tag, KanRHe et al., 2018[23] pRTVC-xopG-GFP Subcellular localization in rice 本实验室 This Study pCambia-1300-eGFP Stable over-expression, Hygromycin, 35S promoter,
C terminal GFP tag, KanR本实验室 This Study pCambia-1300-xopG-eGFP Subcellular localization in tobacco 本实验室 This Study 注:KanR、rifR和SpR分别表示对卡那霉素、利福平和壮观霉素的抗性。 Note: KanR,rifR,and SpR respectively represent resistance to kanamycin,rifampicin,and spectinomycin. -
从KEGG数据库下载xopG基因CDs序列,使用Primer 5.0分别设计引物。本实验引物见表2。
表 2 实验所用引物
Table 2. Primers used in the experiment
引物 Primer 核苷酸序列 Nucleotide sequences(5′-3′) 用途 Usage xopGddFF GCTCAAGCTTCTGCACGGCAAGGAAGACAG xopG敲除
xopG deletionxopGddFR ACAAGGATCCCTATCTCTGCTATGGTCACT xopGddRF ACAAGGATCCCAGGTGAGGCTGCATATCGA xopGddRR GCTCGAATTCTGGGTCAACGTGGTGTTGTC xopGhmF GACCAAGCTTGGTGACCATAGCAGAGATAGGC xopG回补
xopG complementxopGhmR GACCGGTACCACCTGCCGTGAGGCTTATATTT hrpXddFF CAGCAAGCTTTCACGCTTGCGAACGCTTCT hrpX敲除
hrpX deletionhrpXddFR CAGCGGATCCCCTCACTCTGTTCTCAAACG hrpXddRF CAGCGGATCCACCTTGCAACGGTAATCTCT hrpXddRR CAGCGAATTCGATTTCCTGCTGGGTCAGTC pCambia-1300-xopG-eGFPF GCAATGTCTTCACTGTTGATAATGACCATAGCA
GAGATAGGCXopG烟草亚细胞定位
Subcellular localization of XopG in tobaccopCambia-1300-xopG-eGFPR CTCCTCGCCCTTGCTCACCATCCTGCCGTGAG
GCTTATATTTpK18gusA-xopGF TGTAAAACGACGGCCAGTGCCCTGCACGGCA
AGGAAGACAGxopG启动子活力测定
Analysis of xopG promoter activitypK18gusA-xopGR CATAAGGGACTGACCACCCGGGCCTATCTCTG
CTATGGTCApRTVc-xopG-GFPF GCGCGGATCCGTGACCATAGCAGAGATAGGC XopG水稻亚细胞定位
Subcellular mapping of XopG in ricepRTVc-xopG-GFPR CCCAAGCTTCCTGCCGTGAGGCTTATATTT 注:下划线表示酶切位点。 Note: The underline indicates the enzyme digestion site. -
根据KEGG (
https://www.genome.jp/kegg/ )网站查询到的XopG的基因序列,采用SMART在线网站(https://smart.embl.de/ )进行XopG蛋白结构域分析;应用PSIPRED (http://bioinf.cs.ucl.ac.uk/psipred/ )在线网站预测XopG蛋白二级结构;利用TMHMM (https://services.healthtech.dtu.dk/services/TMHMM-2.0/ )在线网站分析XopG有无跨膜结构域,基于SignalP (https://services.healthtech.dtu.dk/services/SignalP-6.0/ )在线网站预测XopG有无信号肽;通过在线网站NCBI (https://www.ncbi.nlm.nih.gov/ )和黄单胞菌数据库(https://www.xanthomonas.org/t3e. )获取10个XopG同源蛋白,采用MEGA11.0软件的NJ法(Neighbor-Joining)构建系统发育树,通过DNAMAN 8.0进行蛋白多序列比对;通过比对基因上游500 bp的核酸序列,预测xopG启动子区是否具有PIP-box特征区。 -
应用酶切连接法构建pK18gusA-xopG表达载体[26 − 27],将pK18gusA-xopG分别转化野生型菌株T7174及ΔhrpX后,挑取单克隆进行GUS染色和GUS活力测定,以分析xopG表达情况,实验步骤如参考文献[28 − 29]。
-
应用剪叶法[30 − 31]进行水稻叶片接种实验,将T7174、xopG和C-ΔxopG菌株分别划线于PSA和PSSP (PSA+100 mg·L-1 Sp)固体培养基上,28 ℃培养2 d后,用ddH2O洗脱菌体,调至OD600约为0.6。接种生长60 d左右的TP309水稻,每天观察水稻发病情况,14 d后记录病斑长度。
-
选择培养12 d左右的水稻黄化苗,取其茎秆切碎,参考酶解分离法进行水稻原生质体分离[32 − 33],获得水稻原生质体。运用PEG转化法[34]将构建好的重组载体pRTVc-xopG-GFP转化到水稻原生质体中,培养12 h后于共聚焦显微镜下观察荧光。
-
将构建好的烟草定位载体pCambia-1300-xopG-eGFP转入农杆菌感受态GV3101中,于LB (含kana,Rif)液体培养基28 ℃培养16~18 h。培养好后,4 000 r·min−1离心10 min,弃培养液,向管中加入2 mL农杆菌重悬Buffer,调至OD600约为0.2,注射烟草叶片背面[35],保湿培养36 h于共聚焦显微镜下观察荧光。
-
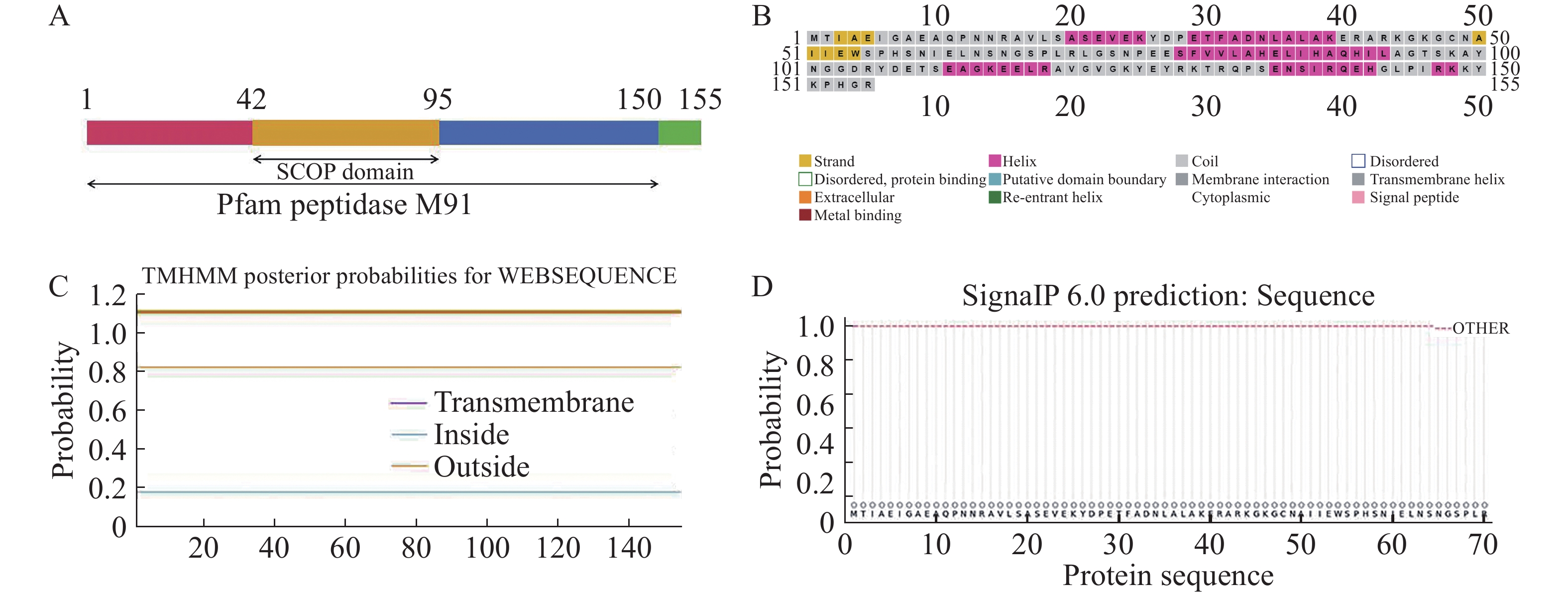
xopG基因全长468 bp,预测编码155个氨基酸。采用SMART在线网站进行XopG蛋白结构域分析,发现XopG蛋白1~150位氨基酸存在典型的锌金属肽酶结构域,42~95位氨基酸存在SCOP结构域(图1−A),预测为典型锌金属肽酶。应用PSIPRED在线网站预测XopG蛋白二级结构,发现其有6处α−螺旋结构,2处β−折叠结构 (图1−B)。应用TMHMM在线网站分析发现XopG蛋白无跨膜结构(图1−C),可能为非膜定位蛋白。SignalP在线网站预测结果表明,XopG蛋白无信号肽(图1−D)。
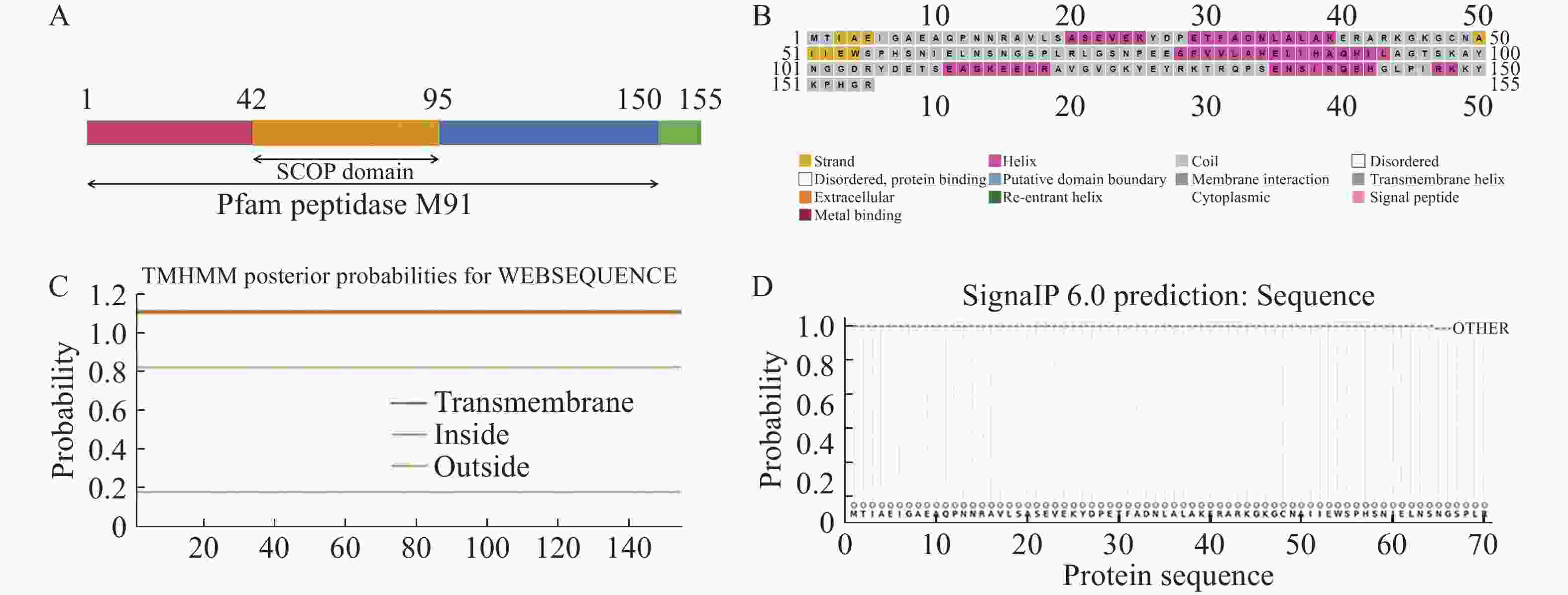
图 1 XopG的生物信息学分析
Figure 1. Bioinformatics analysis of XopG
-
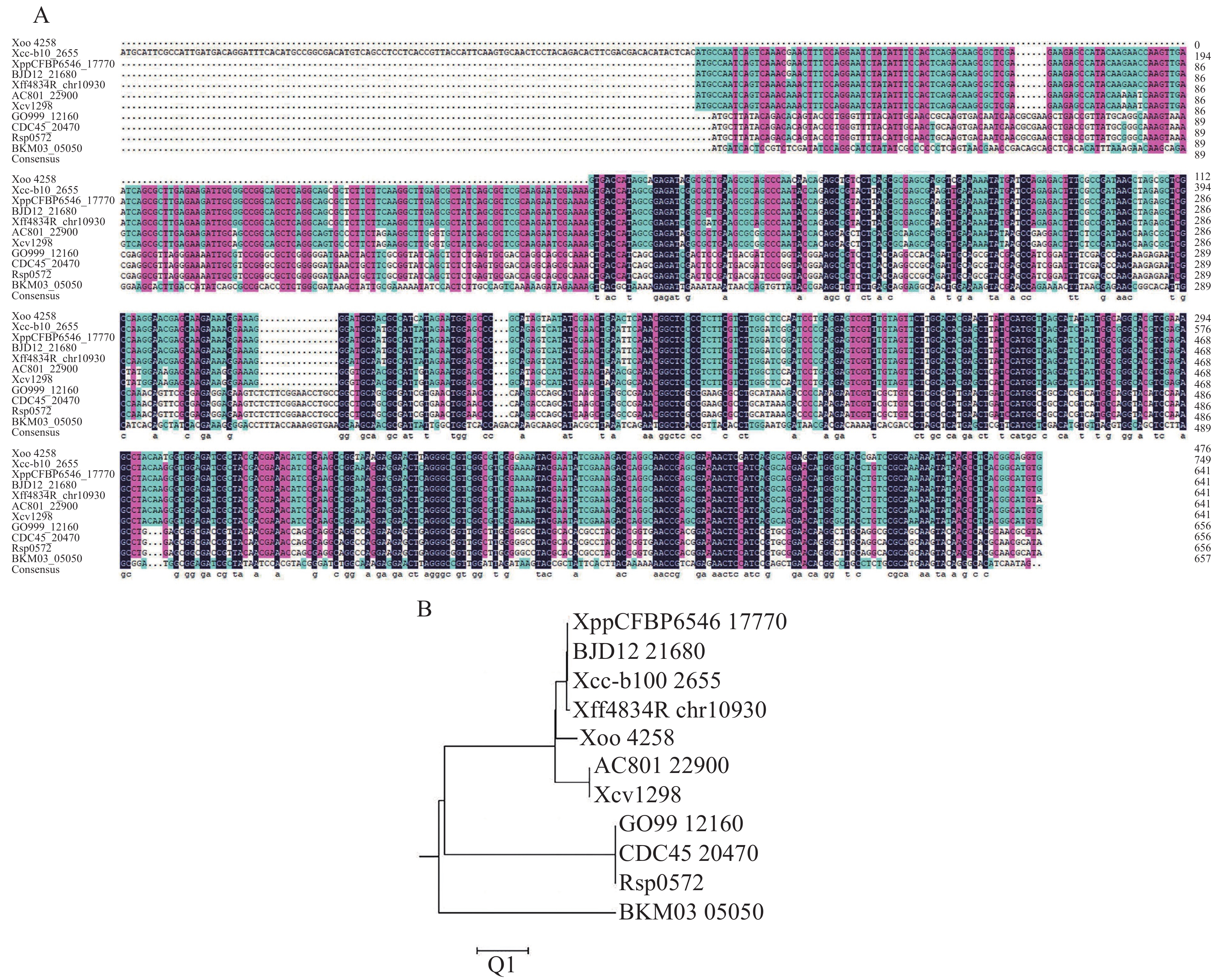
为了研究XopG,从NCBI网站和黄单胞菌数据库中获得了与XopG (Xoo
4258 )蛋白同源性较高的10条同源蛋白的序列数据(表3),并使用DNAMEN8.0软件对其同源蛋白进行多序列比对图2−A),发现XopG蛋白在不同细菌中具有较高的序列保守性,其中与柑橘黄单胞菌柑橘致病变种同源性最高。应用MEGA11.0软件的NJ法对这11个蛋白进行系统进化树的构建(图2−B),发现XopG共有6个分支,Xoo中XopG与柑橘黄单胞菌柑橘致病变种同源性最高,与野油菜致病变种、菜豆黄单胞菌和番茄黄单胞菌同源性也较高。表 3 本研究中用于XopG进化分析的11个菌原
Table 3. Eleven bacterial strains used in this study for the evolutionary analysis of XopG
病原菌Pathogens 简称 Abbreviation Xanthomonas citri pv. fuscans 柑橘黄单胞菌柑橘致病变种 Xff4834R chr10930 Xanthomonas campestris pv. campestris B100 野油菜黄单胞菌油菜致病变种 Xcc-b100 2655 Xanthomonas phaseoli 菜豆黄单胞菌 XppCFBP6546 17770 Xanthomonas vesicatoria 番茄黄单胞菌 BJD12 21680 Xanthomonas campestris pv. vesicatoria 野油菜黄单胞菌疮痂致病变种 Xcv1298 Xanthomonas sp. ISO98C4 黄单胞菌ISO98C4 AC801 22900 Pseudomonas avellanae 黄单胞菌铜绿假单胞菌 BKM03-05050 Ralstonia pseudosolanacearum RS 青枯菌RS G0999 12160 Ralstonia pseudosolanacearum GMI1000 青枯菌GMI1000 Rsp0572 Ralstonia pseudosolanacearum RS 476 青枯菌RS 476 CDC45 20470 Xanthomonas oryzae pv. oryzae 4258 黄单胞菌水稻致病变种 Xoo 4258 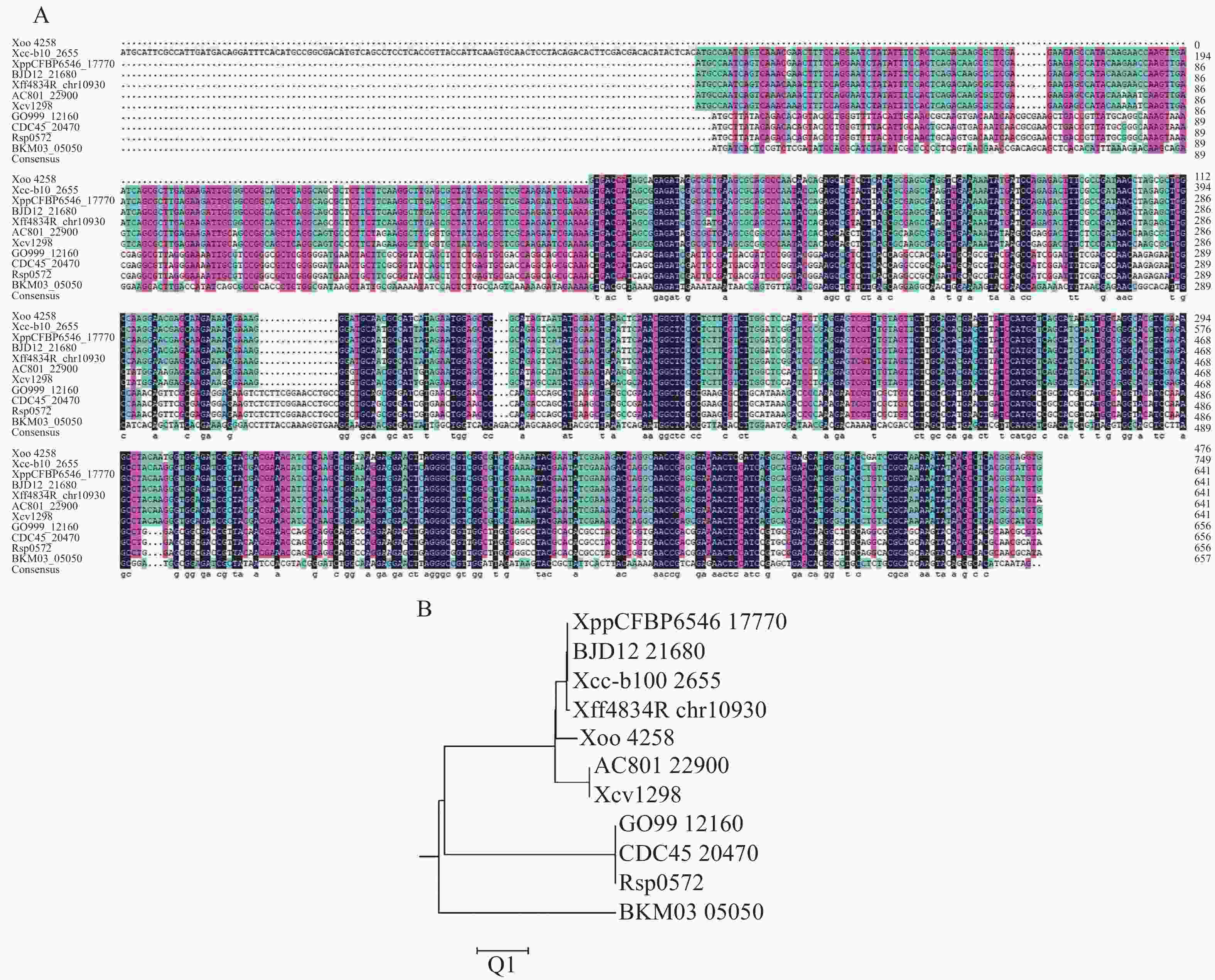
图 2 不同病原菌中XopG蛋白多序列比对以及系统发育树分析
Figure 2. Multi-sequence alignment and phylogenetic tree analysis of XopG protein in different pathogenic bacteria
-
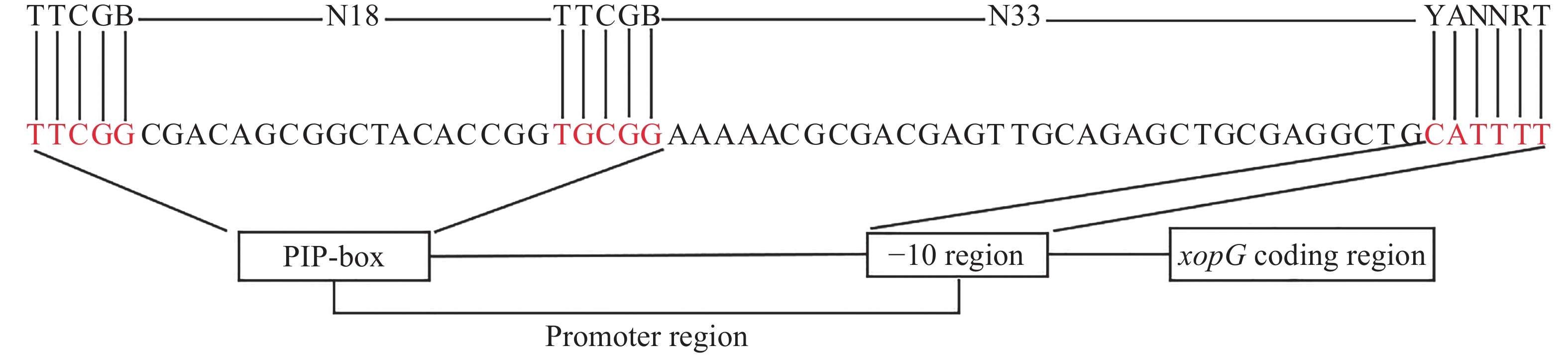
T3SEs的表达依赖于两个重要的调控因子HrpX和HrpG。HrpG调控hrpX表达,而HrpX可以直接结合到T3SEs和T3SS基因启动子的PIP-box基序上,促进效应因子基因及T3SS结构基因的转录。PIP-box的保守结合序列是TTCGB-N15-TTCGB-N32-YANNRT(B和Y代表C、G、T;R代表A、G、T)。但不是所有的T3SEs编码基因都具有典型的PIP-box保守结构域,部分PIP-box的TTCGB中个别碱基可以发生变化,称之为非典型的PIP-box[36]。通过序列比对,发现xopG启动子区域的PIP保守结合序列与典型的PIP-box的保守结合序列不完全一致。如图3所示,xopG启动子区域TTCGB与TTCGB之间含有18个碱基,TTCGB与YANNRT之间含有33个碱基,与PIP-box结构相比,其第25位的T突变为G,因此xopG启动子区域含有非典型的PIP-box结构,表明xopG表达受HrpX调控。
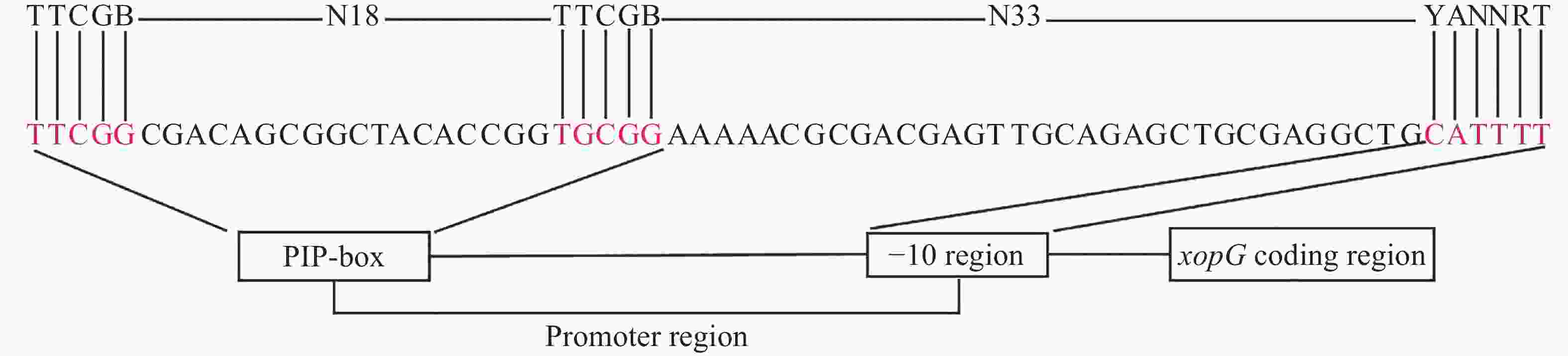
图 3 xopG上游启动子区PIP-box分析
Figure 3. Analysis of the PIP-box in the upstream promoter region of xopG
-
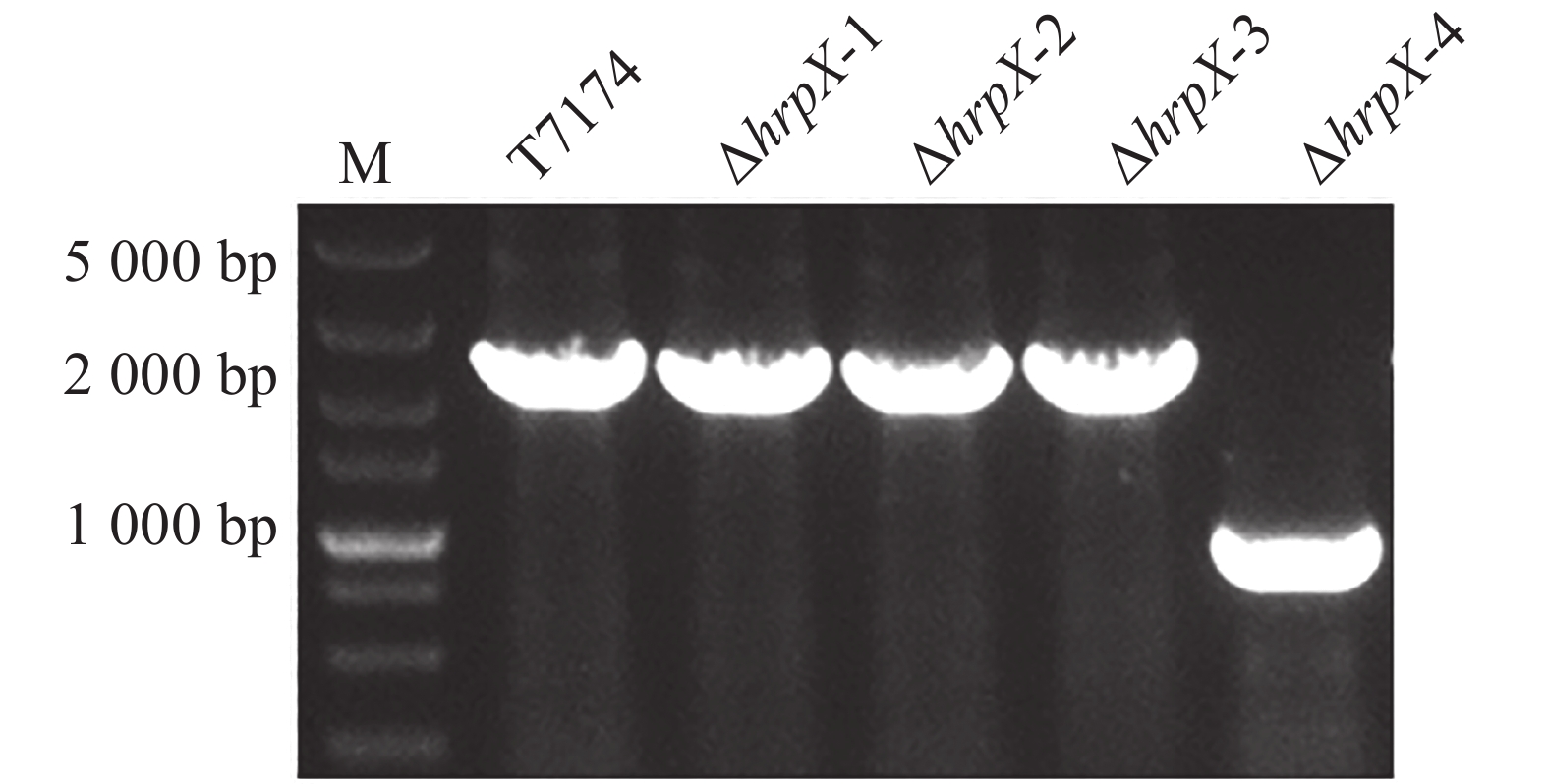
为了验证xopG表达是否确实受HrpX调控,构建了pK18-hrpX载体用于hrpX的敲除,将其通过电击转化法转入T7174中,使用载体引物进行菌落PCR验证。结果如图4所示,野生型条带大小应为2 000 bp左右,而ΔhrpX的扩增条带为1 000 bp左右,表明hrpX基因发生了缺失突变(ΔhrpX)。其中1-3个克隆为未突变菌株,而第4个克隆为突变菌株。
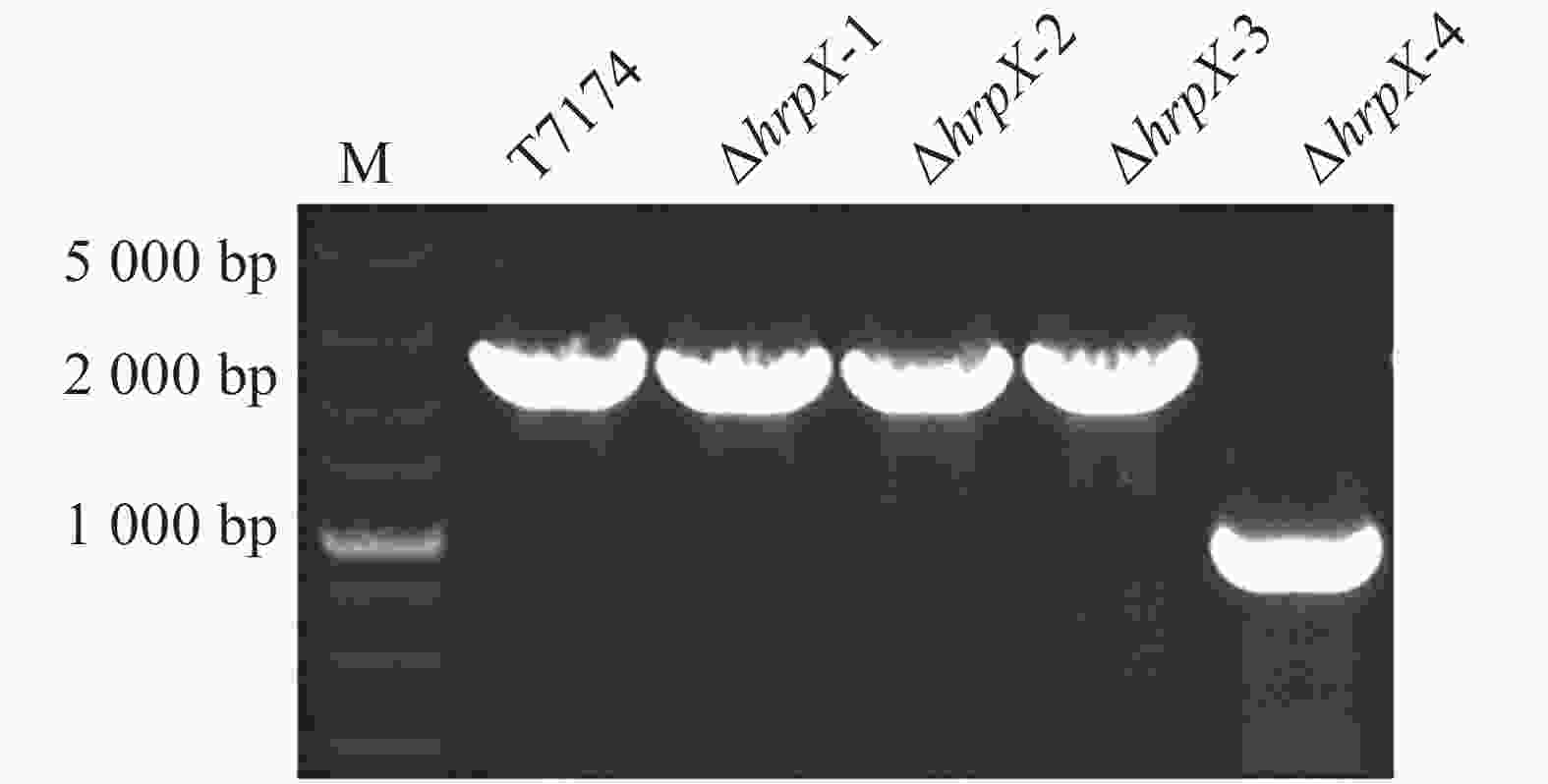
图 4 突变体hrpX的PCR鉴定
Figure 4. PCR identification of mutant hrpX
-
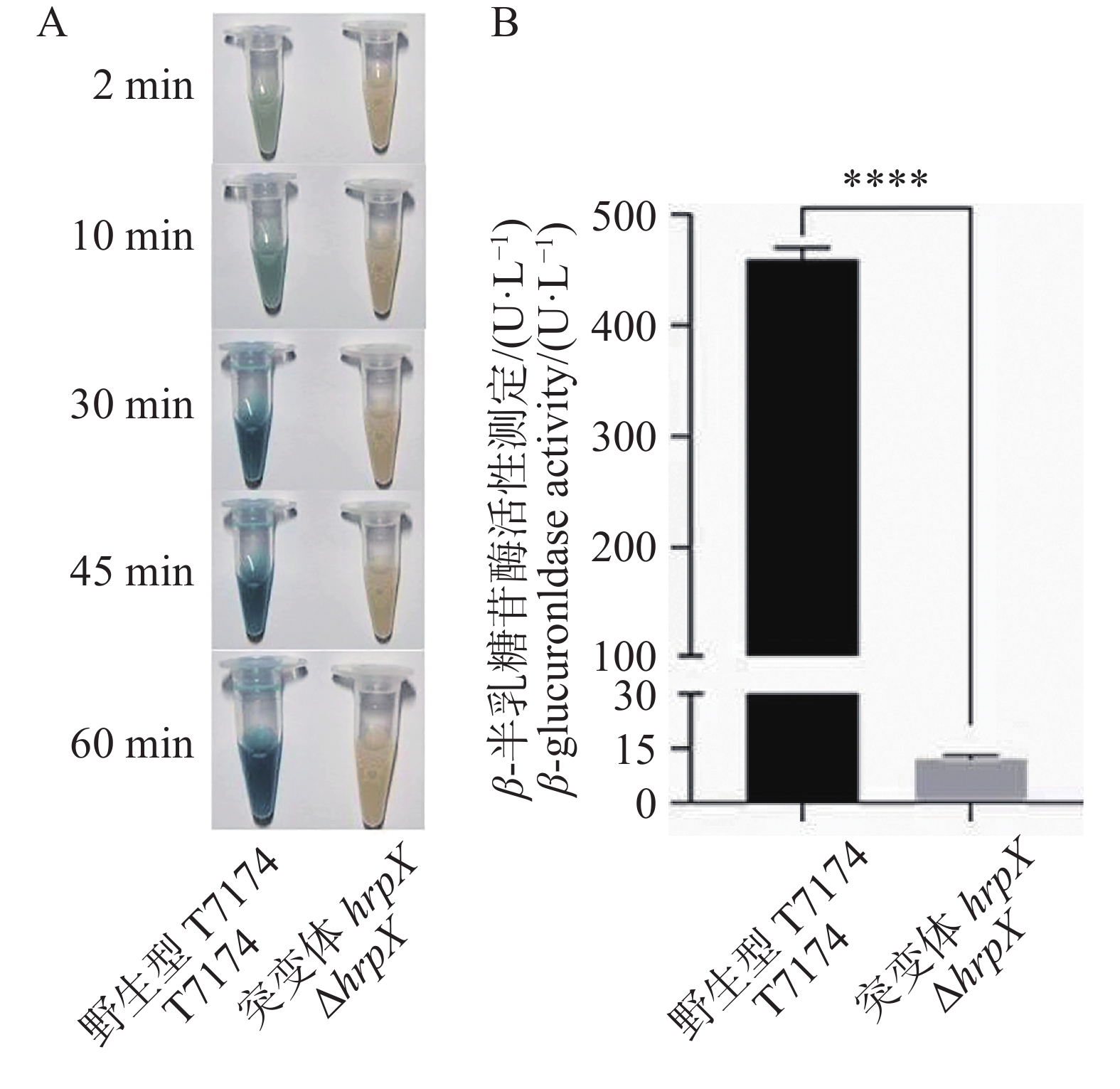
为了评估xopG的表达,将pK18gusA-xopG表达载体分别转化野生型菌株T7174、突变体菌株ΔhrpX后进行GUS染色及活力测定。结果发现在T7174的转化株中,加入GUS底物2 min后出现蓝色且随着染色时间的延长蓝色变深,而在ΔhrpX转化株中,加入GUS底物60 min还没有呈现蓝色(图5−A)。本研究团队也对gusA表达水平进行了定量检测。以ΔhrpX作为实验组,以T7174作为正对照,应用荧光法对其进行启动子活性测定。结果发现hrpX基因敲除后,GUS活性极显著低于T7174菌株(图5−B)。
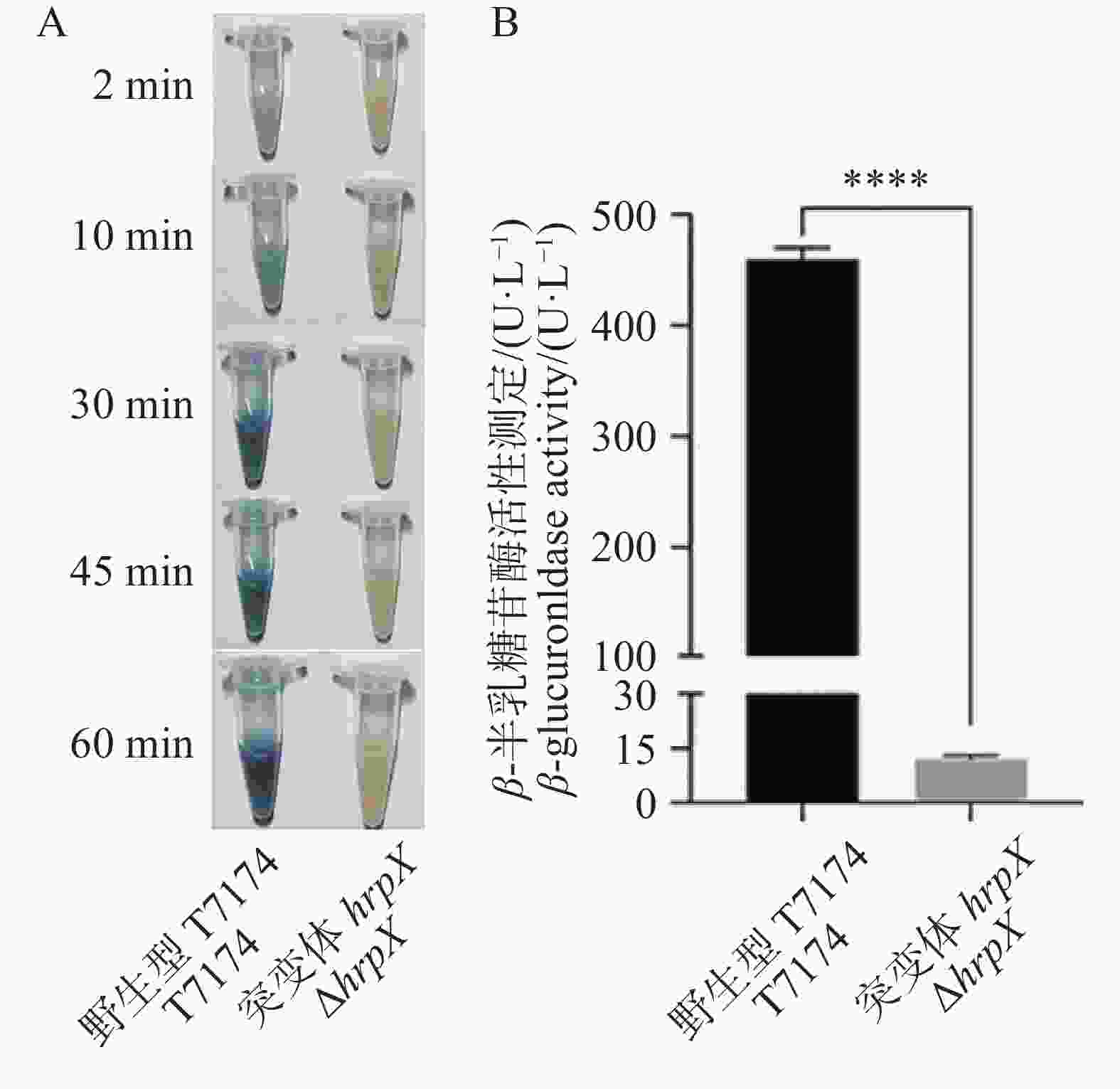
图 5 xopG表达受HrpX正调控
Figure 5. The expression of xopG is positively regulated by HrpX
-
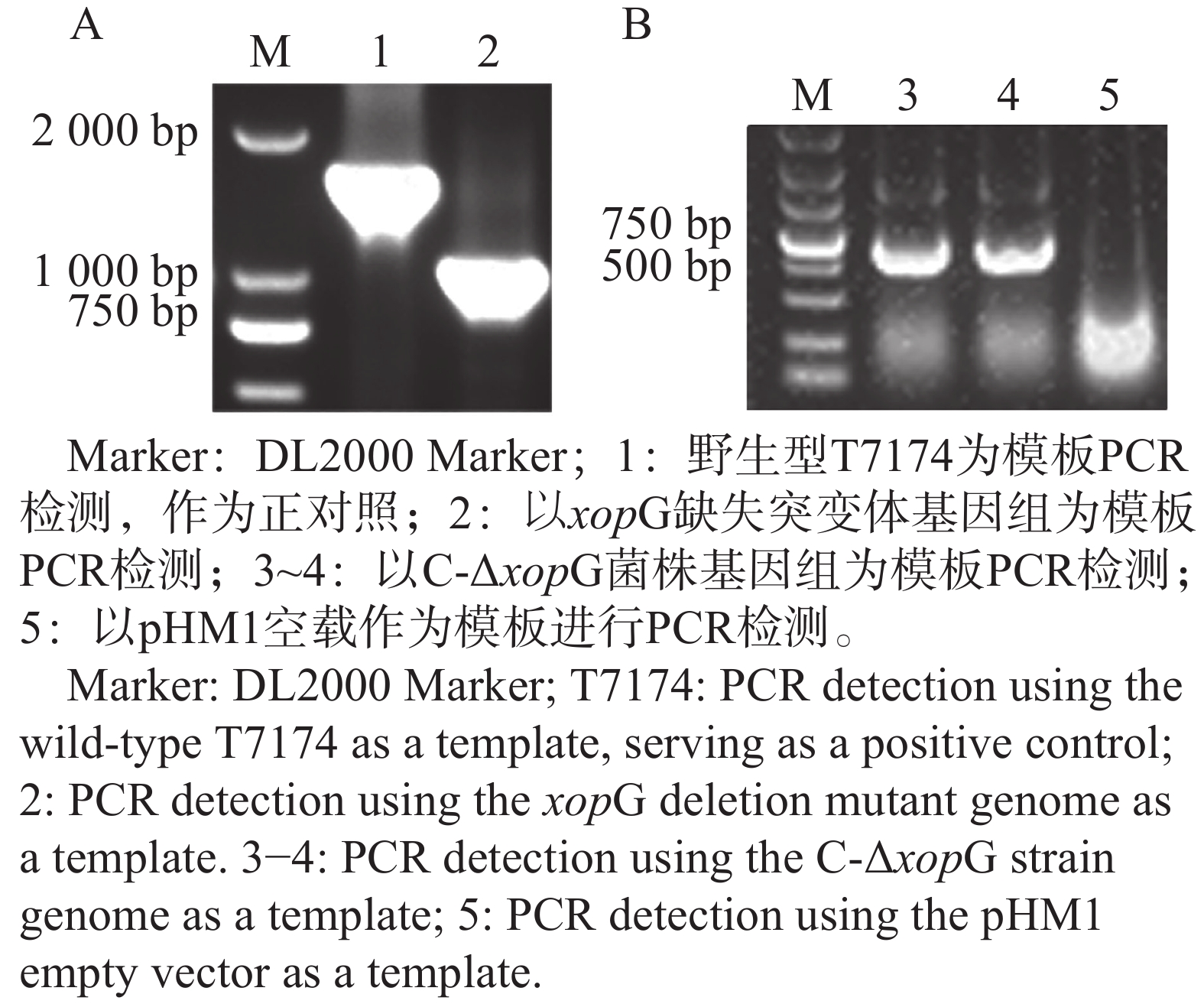
为了检测XopG是否调控Xoo的致病力,构建了xopG突变体菌株,野生型条带大小约为1 500 bp,突变体条带大小约为1 000 bp,二者片段大小之差正好符合预期敲除基因大小,表明突变体构建成功(图6−A);将回补载体pHM1-xopG应用电击转化法转入ΔxopG,使用pHM1载体引物进行PCR检测,以pHM1空载为模板作为实验负对照,结果如图6−B所示,1~3号泳道均能扩增出条带,1~2号泳道条带大小为500 bp至750 bp之间符合预期,3号泳道为负对照组,结果显示回补菌株C-ΔxopG构建成功。
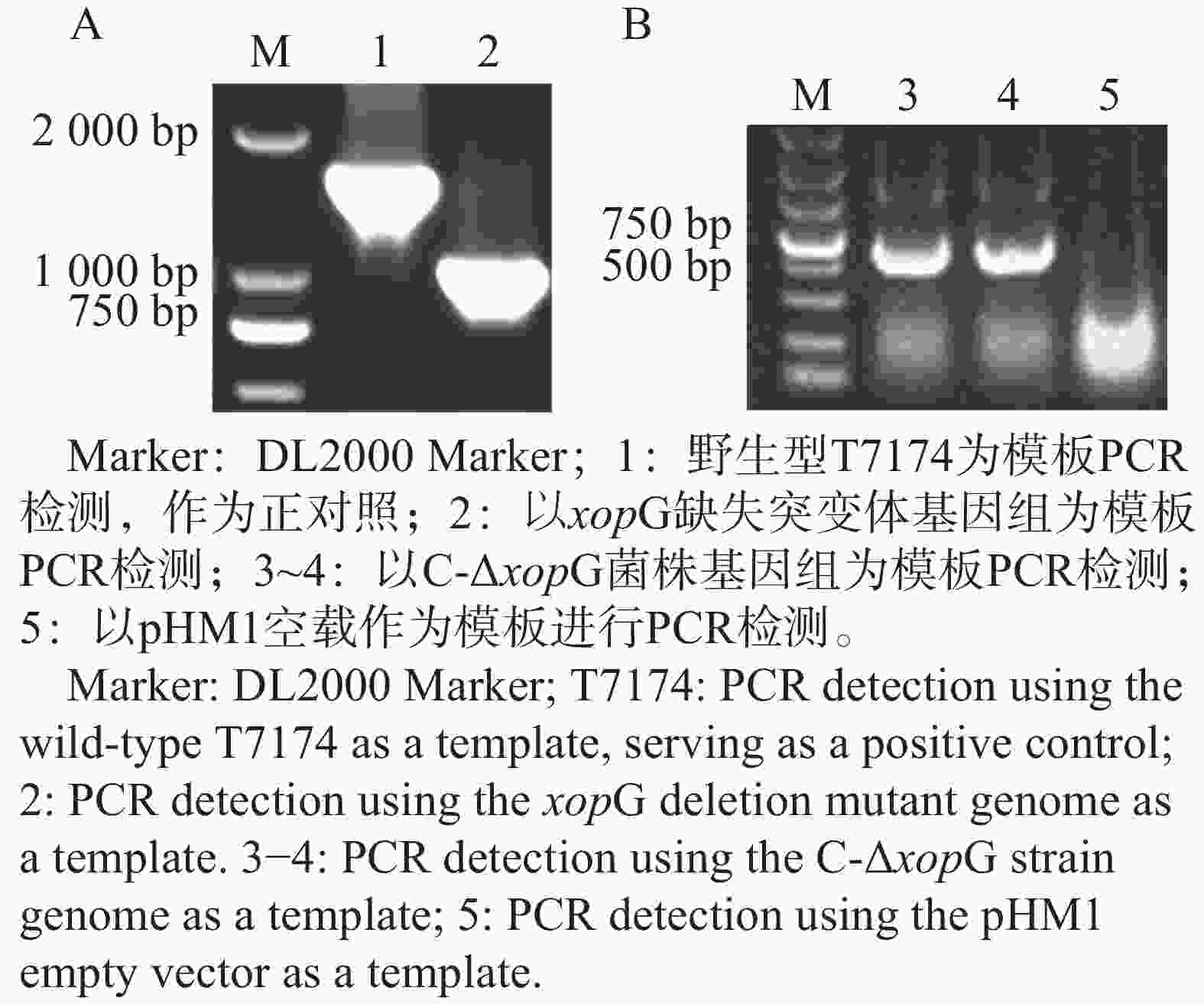
图 6 xopG缺失突变体以及C-ΔxopG菌株构建
Figure 6. Construction of xopG deletion mutant and C-ΔxopG strain
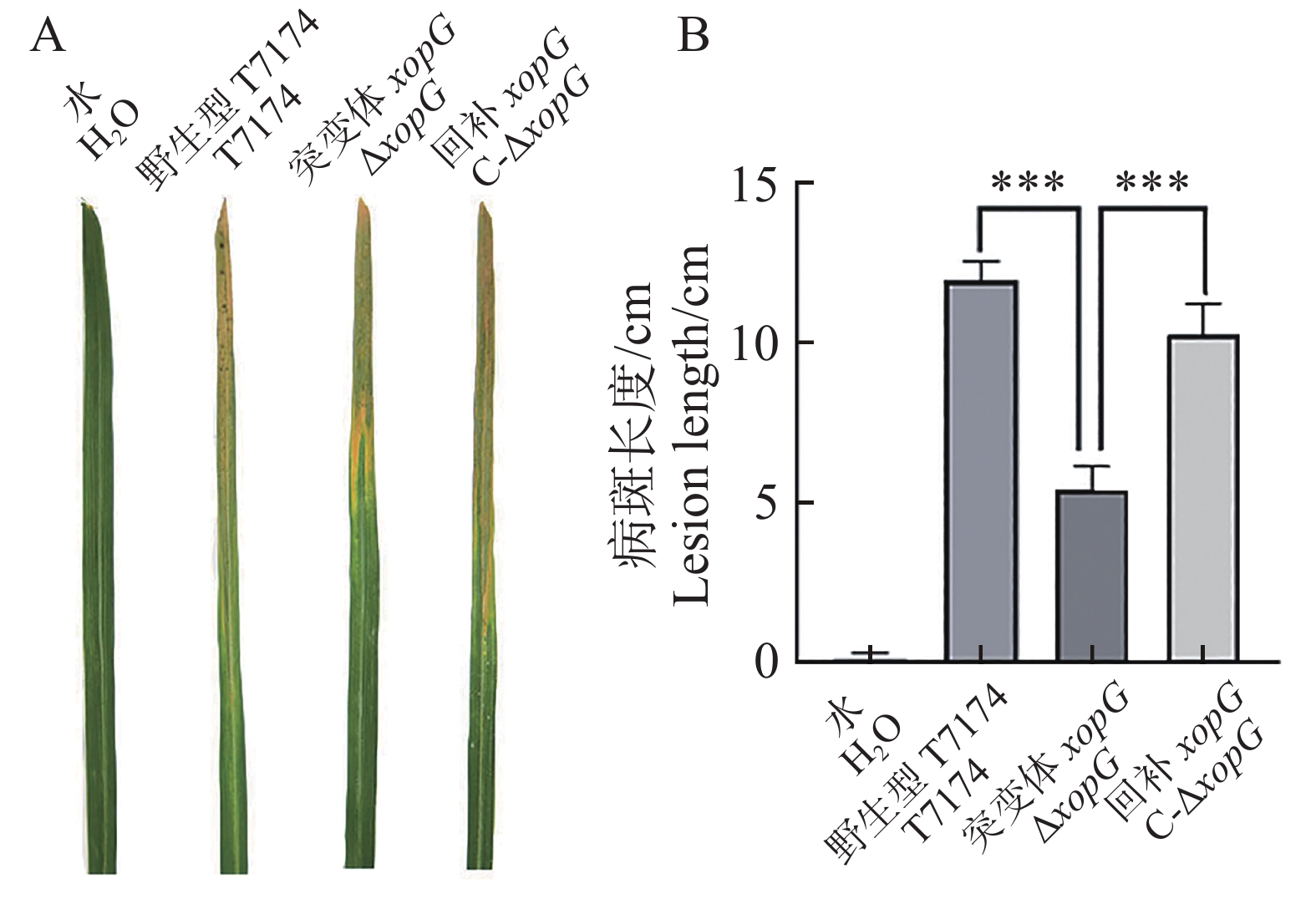
将构建的突变体菌株、回补菌株及野生型菌株应用剪叶法对水稻叶片进行接种实验,接种14 d调查病斑长度并拍照记录(图7−A)。T7174野生型菌株病斑长度为12 cm左右、突变体菌株ΔxopG病斑长度为6 cm左右、回补菌株C-ΔxopG病斑长度为10 cm左右,突变体ΔxopG的病斑长度显著低于野生型T7174及回补菌株C-ΔxopG,野生型T7174及回补菌株C-ΔxopG病斑长度没有显著差异(图7−B)。以上结果表明XopG正调控Xoo致病力。
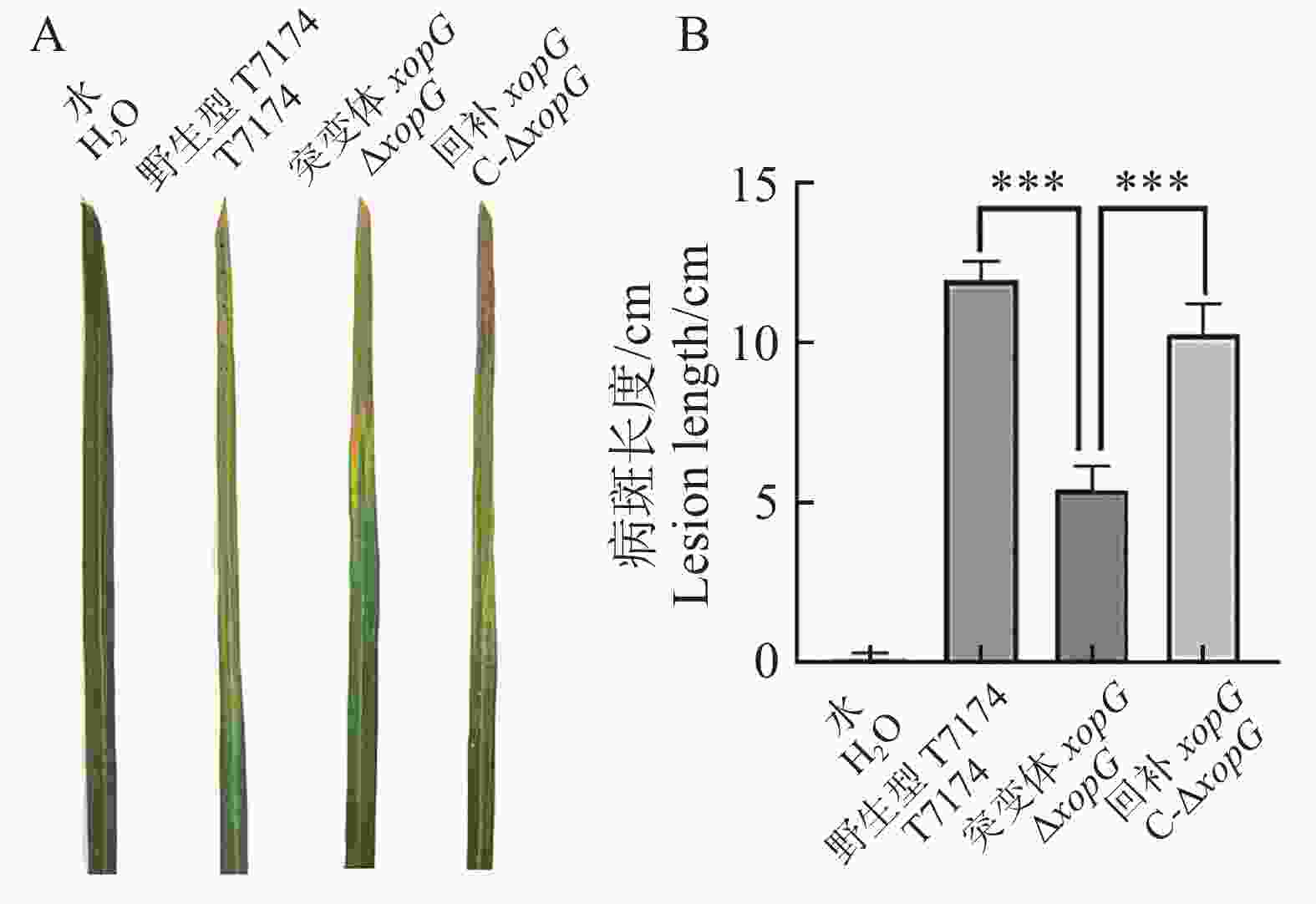
图 7 XopG正调控Xoo在水稻上的致病力
Figure 7. XopG positively regulates the pathogenicity of Xoo on rice
-
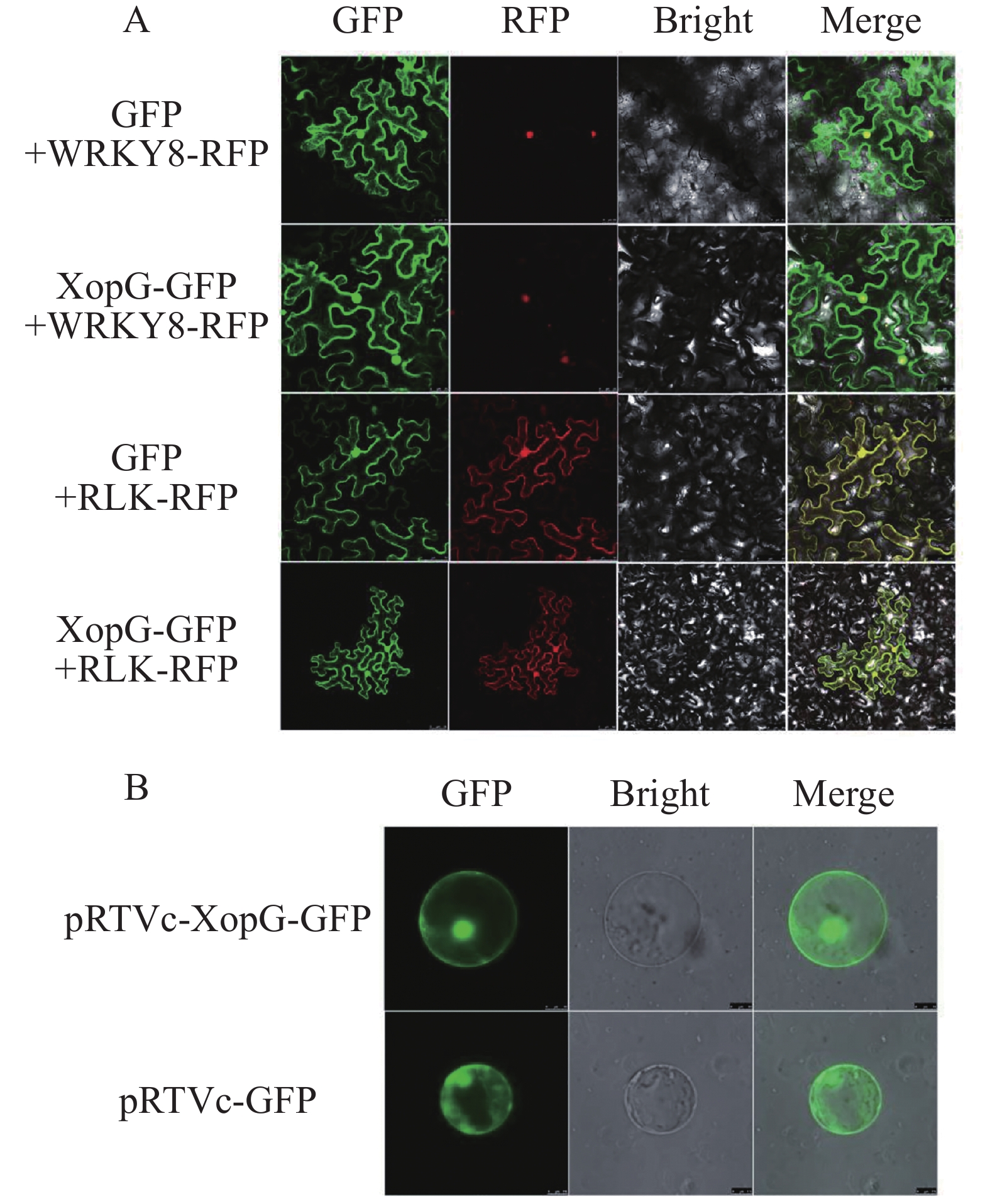
将pCambia-1300-xopG-eGFP重组载体和pCambia-1300-eGFP空质粒分别转入农杆菌,所得农杆菌注射烟草下表皮,避光保湿培养24 h后,继续保湿培养16 h,制成玻片在激光共聚焦显微镜下观察绿色荧光。结果显示:注射携带pCambia-1300-xopG-eGFP重组载体的烟草叶片细胞在细胞核、细胞质、细胞膜中均具有绿色荧光。由此,可以得出XopG蛋白可能定位于烟草细胞细胞核、细胞质和细胞膜(图8−A)。为明确XopG的亚细胞定位情况,将pRTVc-xopG-GFP及pRTV-cGFP空载体分别转化水稻原生质体,12 h后在共聚焦显微镜下观察水稻原生质体的荧光发光情况。如图8−B所示,转化pRTV-cGFP和pRTVc-xopG-GFP的水稻细胞在细胞核、细胞质、细胞膜中均具有绿色荧光,表明XopG在水稻细胞中也定位于细胞核、细胞质、细胞膜,与在烟草细胞中的定位相同。
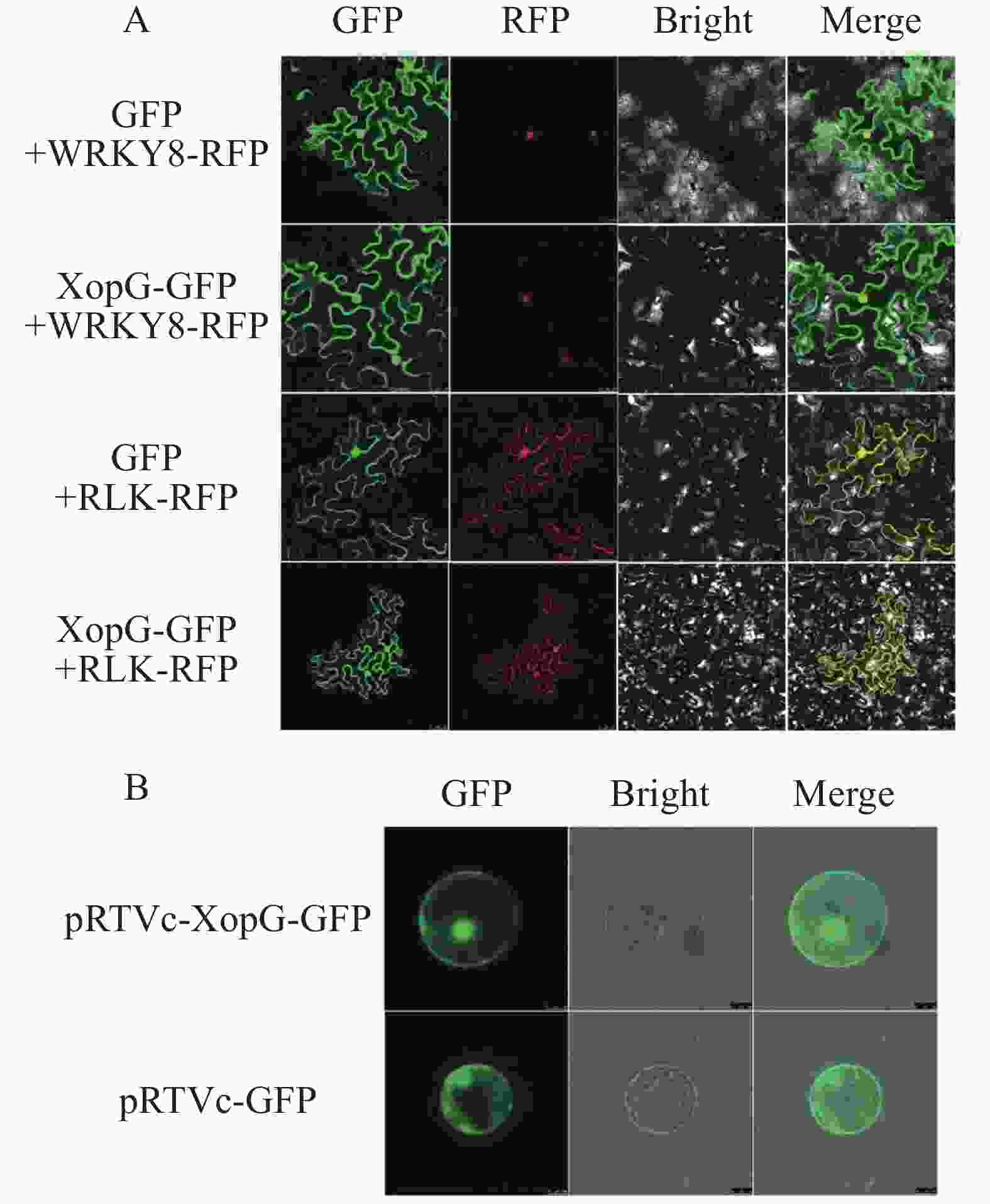
图 8 XopG在烟草叶肉细胞和水稻原生质体中的亚细胞定位
Figure 8. Subcellular localization of XopG in tobacco leaf cells and rice protoplasts
-
在革兰氏阴性菌中,T3SS对病原菌与宿主的相互作用起着重要的调控作用。T3SEs的表达依赖于两个重要的调控因子HrpX和HrpG。HrpG调控hrpX表达,而HrpX可以直接结合到T3SEs和T3SS基因启动子的PIP-box基序上,促进效应因子基因的转录。其中,HrpX通过结合到植物诱导启动子盒PIP-box上促进相关基因转录[9 − 12]。本研究预测xopG基因启动子区含有一个非典型的PIP-box结构,通过启动子活力测定实验发现xopG表达受HrpX调控,因而推测XopG为Ⅲ型效应蛋白,后续将继续进行XopG蛋白外排实验,进一步验证XopG确为Ⅲ型效应蛋白。
通过XopG在不同病原菌中直系同源基因多序列比对及系统进化树分析,推测XopG在稻属黄单胞菌水稻致病变种和柑橘黄单胞菌柑橘致病变种中可能执行相似的功能。Deb等[16 − 17]将Xoo BXO43菌株对易感型水稻TN-1进行接种实验发现,xopG突变体的病斑长度显著短于野生型菌株的病斑长度。本研究在稻属黄单胞菌水稻致病变种T7174小种中敲除xopG基因,接种水稻14 d后发现野生型菌株病斑长度约为12 cm、突变体菌株ΔxopG病斑长度约为6 cm而C-ΔxopG回补菌株病斑长度约为10 cm,突变体菌株病斑长度显著短于野生型菌株,与Deb等实验结果一致,说明XopG正调控Xoo致病力。有研究人员发现XopU、XopV、XopP、XopG和AvrBs2蛋白可以抑制XopQ-XopX介导的水稻免疫应答[18 − 19]。XopQ-XopX相互作用显示发生在细胞核中。XopG与XopQ或XopX的相互作用发生在细胞质中,表明XopG将XopQ和XopX蛋白隔离在细胞质中,从而阻止了它们在细胞核中的共定位和相互作用。同时,研究人员在黄单胞菌中发现五种效应子(XopK、XopG、XopQ、XopL和XopX)抑制flg22诱导基因WRKY33的表达[20]。但至今为止XopG在水稻细胞中的互相作用靶标及其调控机制仍不清楚。
XopG没有可预测的跨膜结构和信号肽,在水稻原生质体的细胞核、细胞膜以及细胞质中均有XopG-GFP荧光信号,在烟草细胞中也定位于细胞核、细胞质、细胞膜。亚细胞定位结果显示XopG没有一个明显的定位情况,在细胞的核、膜及质中均存在,推测其定位可能受到其水稻中靶标蛋白的影响。
综上,本研究验证了xopG的表达受HrpX调控,明确了XopG正调控Xoo的致病力及在植物细胞中的亚细胞定位情况,为后续筛选XopG在水稻内的互作靶标蛋白,进一步揭示在Xoo侵染过程中XopG通过何种途径影响水稻的抗病性从而调控植物免疫的方式提供了实验支撑,为水稻抵抗白叶枯病提供新策略。
-
本研究通过对XopG蛋白结构的生物信息学分析、基因启动子活力测定,证明xopG受HrpX的正调控;通过病原菌致病力检测实验,发现xopG缺失导致Xoo侵染所致的水稻病斑长度显著低于野生型菌株T7174,C-ΔxopG菌株接种水稻的病斑长度与野生型菌株接种水稻的病斑长度无显著关系,这表明XopG正调控Xoo致病力;水稻原生质体及烟草亚细胞定位实验结果表明XopG在植物细胞内定位于细胞核、细胞膜和细胞质。
Ⅲ型效应蛋白XopG调控水稻白叶枯病菌的致病力及其在植物细胞中的定位分析
DOI: 10.15886/j.cnki.rdswxb.20240058
 CSTR: 32425.14.j.cnki.rdswxb.20240058
CSTR: 32425.14.j.cnki.rdswxb.20240058
The virulence of Xanthomonas oryzae pv. oryzae regulated by the type Ⅲ effector XopG and the localization of XopG in plant cells
-
摘要: 稻属黄单胞菌水稻致病变种(Xanthomonas oryzae pv. oryzae,Xoo)是引起水稻白叶枯病的病原菌。Ⅲ型效应蛋白(Type Ⅲ Secretion Effectors,T3SEs)在Xoo致病过程中发挥重要作用。本研究以Xoo T7174中预测的Ⅲ型效应蛋白XopG为研究对象,通过生物信息学分析、基因敲除和表达调控分析、致病力分析及蛋白亚细胞定位分析,初步明确XopG对Xoo致病力的影响及XopG在植物细胞中的定位。结果显示,xopG启动子序列含有非典型的植物诱导启动子盒PIP-box,预测xopG表达受HrpX调控;启动子活力测定结果显示xopG在hrpX突变体中的表达水平显著低于野生型菌株T7174,表明HrpX正调控xopG的表达;缺失xopG会显著降低T7174对水稻的致病力,表明XopG是Xoo侵染水稻所必需的细胞组分;亚细胞定位显示,XopG定位于植物的细胞核、细胞膜和细胞质中。本研究为探讨XopG调控水稻白叶枯病菌致病力的分子机制奠定了基础。Abstract: Xanthomonas oryzae pv. oryzae is the pathogen that causes bacterial leaf blight in rice. Type Ⅲ secretion effectors (T3SEs) play an important role in the virulence of Xoo. To study the function of XopG in Xoo virulence and the localization in plant cells, we carried out the informatics analysis, gene knockout, expression analysis, subcellular localization of XopG, and analyzed the effect of xopG mutation on Xoo virulence. The results showed that the xopG promoter sequence contains an atypical plant-induced motif (PIP-box), indicating that xopG expression is regulated by HrpX. The promoter activity of xopG in hrpX deletion mutant was significantly lower than that in the wild type strain T7174, suggesting that HrpX positively regulates the expression of xopG. Compared to the wild type T7174, the virulence of xopG mutant was significantly reduced, indicating that XopG is a key effector for Xoo to infect rice. Subcellular localization showed that XopG locates in the nucleus, cell membrane and cytoplasm in plant cells. This study laid a foundation for exploring the molecular mechanism of XopG-Xoo interaction.
-
Key words:
- Xoo /
- type Ⅲ effector /
- XopG /
- virulence
-
图 2 不同病原菌中XopG蛋白多序列比对以及系统发育树分析
注: A.不同病原菌中XopG蛋白的多序列比对;B.不同病原菌中XopG的系统发育树分析。
Fig. 2 Multi-sequence alignment and phylogenetic tree analysis of XopG protein in different pathogenic bacteria
Note: A. Multiple sequence alignment of XopG protein in different pathogenic bacteria; B. Phylogenetic tree analysis of XopG in different pathogenic bacteria.
图 3 xopG上游启动子区PIP-box分析
Fig. 3 Analysis of the PIP-box in the upstream promoter region of xopG
图 4 突变体hrpX的PCR鉴定
Marker:DL5000 Marker;ΔhrpX-4为阳性突变体,ΔhrpX-1、ΔhrpX-2和ΔhrpX-3为阴性突变体。
Fig. 4 PCR identification of mutant hrpX
Marker: DL5000 Marker; ΔhrpX-4 is a positive mutant,while ΔhrpX-1,ΔhrpX-2,andΔhrpX-3 are negative mutants.
图 5 xopG表达受HrpX正调控
注: A. 组织染色法分析启动子活力;B. 荧光定量法分析启动子活力。****:P < 0.001。xopG的启动子连上gusA报告基因,分别转入ΔhrpX和T7174,以ΔhrpX为实验组,T7174作为对照。
Fig. 5 The expression of xopG is positively regulated by HrpX
Note: A. Analysis of promoter activity using tissue staining method; B. Analysis of promoter activity using fluorescence quantitative method. ****: P < 0.001. The promoter of xopG was fused with the gusA reporter gene and then transformed into ΔhrpX and T7174 separately,with ΔhrpX as the experimental group and T7174 as the control group.
图 6 xopG缺失突变体以及C-ΔxopG菌株构建
注: A. xopG缺失突变体菌株构建;B. xopG回补菌株C-ΔxopG构建。
Fig. 6 Construction of xopG deletion mutant and C-ΔxopG strain
Note: A. Construction of the xopG deletion mutant strain;B. Construction of the xopG complemented strain C-ΔxopG.
图 7 XopG正调控Xoo在水稻上的致病力
注: A. 剪叶法接种14 d后发病情况(OD600=1.0);B. 接种14 d后病斑长度。P < 0.01(t检验),表示突变体与野生型之间的致病力存在极显著性差异,***:P < 0.001。
Fig. 7 XopG positively regulates the pathogenicity of Xoo on rice
Note: A. The disease incidence after 14 days of leaf clipping inoculation (OD600=1.0);B. The lesion length after 14 days of inoculation,P < 0.01 (t-test),indicating a significant difference in pathogenicity between the mutant and the wild type,***: P < 0.001.
图 8 XopG在烟草叶肉细胞和水稻原生质体中的亚细胞定位
注: A. XopG在烟草中的亚细胞定位(bar=50 μm)。WRKY8-RFP为细胞核共定位Marker,以GFP与WRKY8-RFP共同注射作为对照,PLK-RFP为细胞核膜质共定位Marker,以GFP与PLK-RFP共同注射作为对照;B. XopG在水稻原生质体中的定位(bar=50 μm)。pRTVc-GFP空载质粒为对照组,pRTVc-xopG-GFP为实验组。
Fig. 8 Subcellular localization of XopG in tobacco leaf cells and rice protoplasts
Note:A. Tobacco subcellular colocalization (bar=50 μm),using WRKY8-RFP as the nuclear colocalization marker,with GFP and WRKY8-RFP co-injection as the control group,and PLK-RFP as the nuclear membrane colocalization marker,with GFP and PLK-RFP co-injection as the control group.B. Rice protoplast subcellular localization (bar=50 μm),using pRTVc-GFP empty plasmid as the control group,and pRTVc-xopG-GFP as the experimental group.
表 1 实验所用菌株与质粒
Table 1 Experimental strains and plasmids
菌株/质粒 Strain/plasmid 相关特征 Relevant characteristics 来源 Source 大肠杆菌 Escherichia coli DH5α F-φ80lacZΔM15 Δ(lacZYA-argF)u169 recA1 endA1
hsdR17(rk-, mk+)phoA supE44 λ- thi-1 gyrA96 relA1上海唯地生物
Shanghai Weidi Biotechnology农杆菌 Agrobacterium GV3101 C58(rifR)TipMP90(pTiC58DT-DNA)
(gentR)Nopaline上海唯地生物
Shanghai Weidi Biotechnology黄单胞菌 Xanthomonas oryzae pv. oryzae T7174 Xoo strain 本实验室
This StudyΔxopG Full length deletion mutant of xopG 本实验室
This StudyC-ΔxopG The complementation strain of ΔxopG 本实验室
This StudyΔhrpX Full length deletion mutant of hrpX(PXO_01953) 本实验室
This Study质粒 Plasmids pK18mobsacB Suicide plasmid in Xanthomonas, KanR Schäfe et al., 1994[21] pK18-xopG pK18mobsacB based plasmid for xopG deletion 本实验室
This StudypHM1 broad-host range vector, SpR Huynh et al., 1989[22] pHM1-xopG Complementary vector containing
full length xopG gene, SpR本实验室
This StudypRTVCGFP Transient over-expression, Ubi promoter,
N terminal EGFP Tag, KanRHe et al., 2018[23] pRTVC-xopG-GFP Subcellular localization in rice 本实验室 This Study pCambia-1300-eGFP Stable over-expression, Hygromycin, 35S promoter,
C terminal GFP tag, KanR本实验室 This Study pCambia-1300-xopG-eGFP Subcellular localization in tobacco 本实验室 This Study 注:KanR、rifR和SpR分别表示对卡那霉素、利福平和壮观霉素的抗性。 Note: KanR,rifR,and SpR respectively represent resistance to kanamycin,rifampicin,and spectinomycin.  下载: 导出CSV
下载: 导出CSV
表 2 实验所用引物
Table 2 Primers used in the experiment
引物 Primer 核苷酸序列 Nucleotide sequences(5′-3′) 用途 Usage xopGddFF GCTCAAGCTTCTGCACGGCAAGGAAGACAG xopG敲除
xopG deletionxopGddFR ACAAGGATCCCTATCTCTGCTATGGTCACT xopGddRF ACAAGGATCCCAGGTGAGGCTGCATATCGA xopGddRR GCTCGAATTCTGGGTCAACGTGGTGTTGTC xopGhmF GACCAAGCTTGGTGACCATAGCAGAGATAGGC xopG回补
xopG complementxopGhmR GACCGGTACCACCTGCCGTGAGGCTTATATTT hrpXddFF CAGCAAGCTTTCACGCTTGCGAACGCTTCT hrpX敲除
hrpX deletionhrpXddFR CAGCGGATCCCCTCACTCTGTTCTCAAACG hrpXddRF CAGCGGATCCACCTTGCAACGGTAATCTCT hrpXddRR CAGCGAATTCGATTTCCTGCTGGGTCAGTC pCambia-1300-xopG-eGFPF GCAATGTCTTCACTGTTGATAATGACCATAGCA
GAGATAGGCXopG烟草亚细胞定位
Subcellular localization of XopG in tobaccopCambia-1300-xopG-eGFPR CTCCTCGCCCTTGCTCACCATCCTGCCGTGAG
GCTTATATTTpK18gusA-xopGF TGTAAAACGACGGCCAGTGCCCTGCACGGCA
AGGAAGACAGxopG启动子活力测定
Analysis of xopG promoter activitypK18gusA-xopGR CATAAGGGACTGACCACCCGGGCCTATCTCTG
CTATGGTCApRTVc-xopG-GFPF GCGCGGATCCGTGACCATAGCAGAGATAGGC XopG水稻亚细胞定位
Subcellular mapping of XopG in ricepRTVc-xopG-GFPR CCCAAGCTTCCTGCCGTGAGGCTTATATTT 注:下划线表示酶切位点。 Note: The underline indicates the enzyme digestion site.
下载: 导出CSV
表 3 本研究中用于XopG进化分析的11个菌原
Table 3 Eleven bacterial strains used in this study for the evolutionary analysis of XopG
病原菌Pathogens 简称 Abbreviation Xanthomonas citri pv. fuscans 柑橘黄单胞菌柑橘致病变种 Xff4834R chr10930 Xanthomonas campestris pv. campestris B100 野油菜黄单胞菌油菜致病变种 Xcc-b100 2655 Xanthomonas phaseoli 菜豆黄单胞菌 XppCFBP6546 17770 Xanthomonas vesicatoria 番茄黄单胞菌 BJD12 21680 Xanthomonas campestris pv. vesicatoria 野油菜黄单胞菌疮痂致病变种 Xcv1298 Xanthomonas sp. ISO98C4 黄单胞菌ISO98C4 AC801 22900 Pseudomonas avellanae 黄单胞菌铜绿假单胞菌 BKM03-05050 Ralstonia pseudosolanacearum RS 青枯菌RS G0999 12160 Ralstonia pseudosolanacearum GMI1000 青枯菌GMI1000 Rsp0572 Ralstonia pseudosolanacearum RS 476 青枯菌RS 476 CDC45 20470 Xanthomonas oryzae pv. oryzae 4258 黄单胞菌水稻致病变种 Xoo 4258
下载: 导出CSV
-
[1] 朱荣松, 邓九胜, 李勇. 水稻产量与氮肥利用率对不同配方肥料的响应[J]. 现代农业, 2008(10): 17 − 21. doi: 10.3969/j.issn.1008-0708.2008.10.016 [2] 李初军, 刘建萍, 贾丽颖, 等. 我国水稻育种的现状与展望[J]. 中国种业, 2007(1): 11 − 12. doi: 10.3969/j.issn.1671-895X.2007.01.004 [3] 高明君, 何祖华. 水稻免疫机制研究进展[J]. 中国科学: 生命科学, 2013, 43(12): 1016 − 1029. doi: 10.1360/052013-311 [4] TENG K P, LIU Q, ZHANG M, et al. Design and enantioselective synthesis of chiral pyranone fused indole derivatives with antibacterial activities against Xanthomonas oryzae pv oryzae for protection of rice[J]. Journal of Agricultural and Food Chemistry, 2024, 72(9): 4622 − 4629. doi: 10.1021/acs.jafc.3c07491 [5] AHAMMAD I, JAMAL T B, LAMISA A B, et al. Subtractive genomics study of Xanthomonas oryzae pv. oryzae reveals repurposable drug candidate for the treatment of bacterial leaf blight in rice[J]. Journal of Genetic Engineering and Biotechnology, 2024, 22(1): 100353. doi: 10.1016/j.jgeb.2024.100353 [6] JAITHON T, ATICHAKARO T, PHONPHOEM W, et al. Potential usage of biosynthesized zinc oxide nanoparticles from mangosteen peel ethanol extract to inhibit Xanthomonas oryzae and promote rice growth[J]. Heliyon, 2024, 10(1): e24076. doi: 10.1016/j.heliyon.2024.e24076 [7] YANG Z X, ZHU Z, GUO Y L, et al. OsMKK1 is a novel element that positively regulates the Xa21-mediated resistance response to Xanthomonas oryzae pv. oryzae in rice[J]. Plant Cell Reports, 2024, 43(2): 31. doi: 10.1007/s00299-023-03085-8 [8] TAYI L, NATHAWAT R, KUMAR S, et al. Mutational analysis of predicted active site residues of an exoglucanase secreted by Xanthomonas oryzae pv. oryzae to determine their role in catalysis and in virulence on rice[J]. Enzyme and Microbial Technology, 2024, 174: 110372. doi: 10.1016/j.enzmictec.2023.110372 [9] 徐荣旗. 野油菜黄单胞菌野油菜致病变种新的依赖于Ⅲ型分泌系统的效应物的鉴定[D]. 南宁: 广西大学, 2006. [10] 姜珊. 黄单胞菌Ⅲ型分泌系统抑制剂的筛选及其作用机制研究[D]. 广州: 华南农业大学, 2020. doi: 10.27152/d.cnki.ghanu.2020.000324. [11] 刘红霞. 三种Ⅲ型蛋白影响黄单胞菌—水稻互作与植物抗病性研究[D]. 南京: 南京农业大学, 2006. [12] 易杰祥, 景晓辉, 吴伦英. 黄单胞菌Ⅲ型分泌系统效应蛋白的研究进展[J]. 热带农业科学, 2014, 34(8): 74 − 79. doi: 10.3969/j.issn.1009-2196.2014.08.015 [13] KIM J G, TAYLOR K W, HOTSON A, et al. XopD SUMO protease affects host transcription, promotes pathogen growth, and delays symptom development in xanthomonas-infected tomato leaves[J]. The Plant Cell, 2008, 20(7): 1915 − 1929. doi: 10.1105/tpc.108.058529 [14] ORTMANN S, MARX J, LAMPE C, et al. A conserved microtubule-binding region in Xanthomonas XopL is indispensable for induced plant cell death reactions[J]. PLoS Pathogens, 2023, 19(8): e1011263. doi: 10.1371/journal.ppat.1011263 [15] MA W X, XU X M, CAI L L, et al. A Xanthomonas oryzae type Ⅲ effector XopL causes cell death through mediating ferredoxin degradation in Nicotiana benthamiana[J]. Phytopathology Research, 2020, 2(1): 16. doi: 10.1186/s42483-020-00055-w [16] DEB S, GOKULAN C G, NATHAWAT R, et al. Suppression of XopQ-XopX-induced immune responses of rice by the type Ⅲ effector XopG[J]. Molecular Plant Pathology, 2022, 23(5): 634 − 648. doi: 10.1111/mpp.13184 [17] DEB S, GUPTA M K, PATEL H K, et al. Xanthomonas oryzae pv. oryzae XopQ protein suppresses rice immune responses through interaction with two 14-3-3 proteins but its phospho-null mutant induces rice immune responses and interacts with another 14-3-3 protein[J]. Molecular Plant Pathology, 2019, 20(7): 976 − 989. doi: 10.1111/mpp.12807 [18] DEB S, GHOSH P, PATEL H K, et al. Interaction of the Xanthomonas effectors XopQ and XopX results in induction of rice immune responses[J]. The Plant Journal, 2020, 104(2): 332 − 350. doi: 10.1111/tpj.14924 [19] STORK W, KIM J G, MUDGETT M B. Functional analysis of plant defense suppression and activation by the Xanthomonas core type Ⅲ effector XopX[J]. Molecular Plant-Microbe Interactions, 2015, 28(2): 180 − 194. doi: 10.1094/MPMI-09-14-0263-R [20] PRAUTSCH J, LEE ERICKSON J L, ÖZYÜREK S, et al. Effector XopQ-induced stromule formation in Nicotiana benthamiana depends on ETI signaling components ADR1 and NRG1[J]. Plant Physiology, 2023, 191(1): 161 − 176. doi: 10.1093/plphys/kiac481 [21] SCHÄFER A, TAUCH A, JÄGER W, et al. Small mobilizable multi-purpose cloning vectors derived from the Escherichia coli plasmids pK18 and pK19: selection of defined deletions in the chromosome of Corynebacterium glutamicum[J]. Gene, 1994, 145(1): 69 − 73. doi: 10.1016/0378-1119(94)90324-7 [22] HUYNH T V, DAHLBECK D, STASKAWICZ B J. Bacterial blight of soybean: regulation of a pathogen gene determining host cultivar specificity[J]. Science, 1989, 245(4924): 1374 − 1377. doi: 10.1126/science.2781284 [23] STAVRINIDES J, MCCANN H C, GUTTMAN D S. Host-pathogen interplay and the evolution of bacterial effectors[J]. Cellular Microbiology, 2008, 10(2): 285 − 292. doi: 10.1111/j.1462-5822.2007.01078.x [24] 李开怀, 刘凤权. 植物病原细菌基因敲除技术在本科开放实验中的设计与实践[J]. 科教导刊, 2024(1): 67 − 69. doi: 10.16400/j.cnki.kjdk.2024.1.022 [25] 严霞. 野油菜黄单胞菌8004效应蛋白在调节植物先天免疫中的功能研究[D]. 海口: 海南大学, 2016. [26] 于浩泉. 水稻黄单胞菌新型植物致病相关因子的发掘及作用机制探究[D]. 武汉: 华中农业大学, 2015. doi: 10.27158/d.cnki.ghznu.2015.000184. [27] 景晓辉. 野油菜黄单胞菌效应蛋白XopDxcc8004与NAC转录因子ATAF2互作干扰寄主植物的免疫反应[D]. 海口: 海南大学, 2014. [28] 米多. 水稻白叶枯病原菌Xoo中RaxM与MutL互作对raxX表达调控的影响[D]. 海口: 海南大学, 2022. [29] 冯锋. 黄单胞菌效应蛋白AvrAC调节植物先天免疫的分子机制[D]. 北京: 清华大学, 2012. [30] JOSEPH C A. 水稻基于产量的抗旱、耐冷和白叶枯病抗性遗传重叠的数量解析[D]. 北京: 中国农业科学院, 2015. [31] 于燕燕, 夏影影, 吴可建, 等. XppI调控水稻白叶枯病菌致病力的潜在机制[J]. 热带生物学报, 2023, 14(6): 642 − 650. doi: 10.15886/j.cnki.rdswxb.20230024 [32] 马超越, 彭世清, 郭冬, 等. 植物原生质体分离及其瞬时转化的应用[J]. 热带生物学报, 2024, 15(2): 241 − 250. doi: 10.15886/j.cnki.rdswxb.20230026 [33] 李悦, 宋慧云, 王志, 等. 植物原生质体分离与瞬时表达体系研究进展[J]. 植物生理学报, 2023, 59(1): 21 − 32. doi: 10.13592/j.cnki.ppj.300049 [34] 李婧瑶, 刘龙飚, 丁兵, 等. 植物原生质体分离及培养研究进展[J]. 分子植物育种, 2023, 21(2): 620 − 632. doi: 10.13271/j.mpb.021.000620 [35] 郭圣婷. NbRAF2互作蛋白筛选及功能分析[D]. 沈阳: 沈阳农业大学, 2023. doi: 10.27327/d.cnki.gshnu.2023.001338. [36] FENSELAU S. Sequence and expression analysis of the hrpB Pathogenicity operon of Xanthomonas campestris pv. vesicatoria which encodes eight proteins with similarity to components of the hrp, ysc, spa, and fli secretion systems[J]. Molecular Plant-Microbe Interactions, 1995, 8(6): 845 − 854. doi: 10.1094/mpmi-8-0845 -

 点击查看大图
点击查看大图
计量
- 文章访问数: 1547
- HTML全文浏览量: 786
- PDF下载量: 45
- 被引次数: 0
