




-
在原核生物中,超过95%的DNA结合蛋白含有螺旋-转角-螺旋(Helix-turn-helix,HTH)基序(motif),其余的DNA结合蛋白可能具有螺旋−环−螺旋(Helix-loop-helix)、锌指(zinc-finger)和反向β折叠(anti-parallel β-sheet)等结构域。HTH基序大约由20个氨基酸组成,有2个α螺旋和1个连接型转角,第1个α螺旋起到稳定的作用,第2个α螺旋通过氢键和疏水作用与DNA结合[1-2]。HTH在基因表达调控中起到非常重要的作用,所有已知的LysR类转录调节因子和绝大多数的DNA结合型响应调节因子都含有HTH[3]。绝大多数HTH蛋白含有其他调节其活性的结构域,通过感应特殊的信号分子影响HTH结构以及与DNA的结合能力[4-5],同时,这些结构域还可能与其他蛋白互作进而影响其与DNA的结合效率[3-4]。在目前发现的调控蛋白中,只具有HTH结构的调控蛋白占比很少。只具有HTH结构的调控蛋白由于缺乏信号感应或蛋白互作的结构域,其调控机制仍需进一步研究。野油菜黄单胞菌(Xanthomonas campestris pv. campestris, Xcc)是十字花科植物重要的病原菌,也是研究病原细菌与宿主互作的重要模式菌株[6]。Xcc成功侵染宿主依赖于毒力相关基因的有序表达,并受到严格的调控[6]。只有细菌感应到宿主信号并启动侵染相关基因表达后,细菌才能进入植物体内、逃脱植物的防卫反应并成功繁殖。在该过程中,转录调节因子起着非常重要的作用。目前已在Xcc中分离鉴定了一系列与致病相关的转录调节因子,且大部分都具有HTH结构域[7-13]。如HrpX和HrpG调控III型分泌系统及效应因子的表达[10-12];Clp作为全局性的转录调节因子与细胞第二信使C-di-GMP结合调节细胞群体感应和致病力相关的基因表达[7-9];FleQ可以调节细菌鞭毛合成相关基因的表达,影响细菌的运动能力和生物膜(biofilm)的形成[13-14]。这些转录调节因子都具有HTH结构域和感应信号或蛋白互作的结构域。本研究分析了同一个基因簇中3个LysR类转录调节因子(VrhA, VrhB和VrhR)对Xcc致病力的影响及其可能调控的致病相关因子,发现VrhR可负调控Xcc的致病力,为进一步了解VrhR的功能及其调控通路奠定基础。 -
Xcc 8004由本实验室保存于−80 ℃冰箱;普通化学试剂购自广州化学试剂公司;PCR试剂购自诺唯赞生物科技有限公司;限制性内切酶购自New England Biolabs及Thermo Fisher Scientific公司;所用敲除载体为pK18mobSacB(本实验室保存);接种植物为文兴西兰花;基因预测及结构分析采用在线工具(KEGG,
https://www.kegg.jp/kegg-bin/show_organism?org=xcb 和SMART,http://smart.embl-heidelberg.de/ ),采用默认参数;同源性比对采用Clustal X2 (http://www.clustal.org/clustal2/ );基因组信息来源于NCBI (accession: CP000050.1)。 -
将vrhA,vrhR,vrhB上下游各约500 bp序列通过PCR扩增(引物见表1),分别用HindIII/BamHI和BamHI/EcoRI酶切后一起连接到经HindIII/EcoRI酶切的自杀性载体pK18mobSacB(KanR)上,用热激法进行转化,获得载体(pK18-vrhA, pK18-vrhR或pK18-vrhB),载体(pK18-vrhA, pK18-vrhR或pK18-vrhB)经测序正确后采用三亲接合的方法构建突变体[14]。具体方法:将供体菌(含敲除载体的菌)、协助菌(pRK2013)、受体菌(Xcc 8004)混合起来,28 ℃培养1 d,然后将供体菌涂布在含25 mg·L−1卡拉霉素的平板上,28 ℃培养3 d,长出的菌落在不含抗生素的液体培养基中振荡培养72h,200 r·min−1 ,涂布在含15%蔗糖,不含抗生素的平板上,经过此次负筛选后获得的相应菌株进行PCR验证。基因组提取采用SDS法,PCR反应条件:95 ℃ 4 min, 95 ℃ 30 s, 58 ℃ 30 s, 72 ℃ 2 min, 30循环数,最后72 ℃ 5 min。如果突变成功,则其PCR片段比野生型菌株小[14]。采取剪叶法,接种野生型菌株Xcc 8004和突变体ΔvrhB,ΔvrhR及ΔvrhA于生长40 d的西兰花叶片,15 d后量取从剪口处到枯斑内部边界的长度,统计病斑大小与数量[14]。
表 1 引物序列
Table 1. Primer sequences
名称 Primer 序列 Sequence, 5′−3′ 退火温度 Tm/℃ 产物长度 Product length/bp 用途 Uses vrhBqF AAGGATGTGACTGCAACCCG 60.61 147 vrhB qPCR vrhBqR GCTGGCAAAGCGTGCTATTT 60.11 vrhRqF ATGAAACCAGGACGCCCAAT 59.96 121 vrhR qPCR vrhRqR GGGTTGCCAGCTCTTCTTGA 60.25 vrhAqF CGTTCCTCTTCGATCACGGT 59.83 142 vrhA qPCR vrhAqR CTTTGTGGTTTTGGCCGAGG 59.97 rpoDqF GTCGGCTTCAACGACCTGAT 60.39 110 rpoD qPCR rpoDqR TCTTCGTCTGCGTCTTCGTC 60.11 vrhAddFF GACCAAGCTTCAGAACCGGGAGCTAGAACG 60.18 489 vrhA deletion vrhAddFR GACAGGATCCTGTCGTGCATCGCTGCTATC 60.87 vrhAddRF GACAGGATCCAACCGATGCCTAGATCGTAG 56.62 420 vrhA deletion vrhAddRR GACCGAATTCAGCTCTTCTTGAGATATGTC 51.81 vrhRddFF GACCAAGCTTGCATGCACGAATGGTTGCAG 61.07 443 vrhR deletion vrhRddFR GACAGGATCCCTCAGAAGGGTGAAGTGTTC 55.70 vrhRddRF GACAGGATCCCAATCGCTGGCTCAGCTTCA 64.86 437 vrhR deletion vrhRddRR GACCGAATTCGAACGAGCGGGCAAAGCGAC 62.91 vrhBddFF GACCAAGCTTGATGCAGAAGCGCACAATGA 59.55 364 vrhB deletion vrhBddFR GACAGGATCCTCTGTGGATCGCTTCACAAC 58.20 vrhBddRF GACAGGATCCCAGCTCCATCCGCAAGACTC 60.81 404 vrhB deletion vrhBddRR GACCGAATTCTGACTCATCGCATGTGTCAC 58.29 注:下划线表示酶切位点。
Note: Restriction sites are underlined. -
将Xcc 8004,ΔvrhA,ΔvrhR及ΔvrhB挑单克隆菌接种至5 mL NYG(Nutrition Yeast Glycerol)[1%胰化蛋白胨(Peptone),0.5%酵母提取物(Yeast extract),1%甘油(glycerol)]液体培养基中,28 ℃ 200 r·min−1振荡培养36 h(OD600=2.0)。取1 mL培养物,6 000 r·min−1室温离心1 min收集菌体,用10 mmol·L−1 MgCl2清洗菌体2次,重悬于100 μL MgCl2溶液中。取重悬于MgCl2的溶液1 μL接种到补充有纤维素(纤维素酶),淀粉(淀粉酶),脱脂奶粉(蛋白酶)的NYG固体培养基中,28 ℃培养48 h,每处理重复3次。如果菌株具有分泌相应酶的能力,则会在菌落周围形成一圈透明的水解圈。通过比较水解产物的直径(D)和菌落直径(d)的大小(D/d)以鉴定菌株产酶能力,D/d值越大,说明菌株产酶的能力越强。
-
菌培养及收集如1.3所述,取1 μL MgCl2菌悬液接种到含0.3%琼脂糖的NYG平板上,28 ℃培养48 h,测量菌落直径并拍照。
-
数据分析采用GraphPad Prism 5作图分析,所有实验至少设3个生物学重复,计算平均值±SD(标准差)。图像后期用Adobe Photoshop 8.0软件进行处理。
-
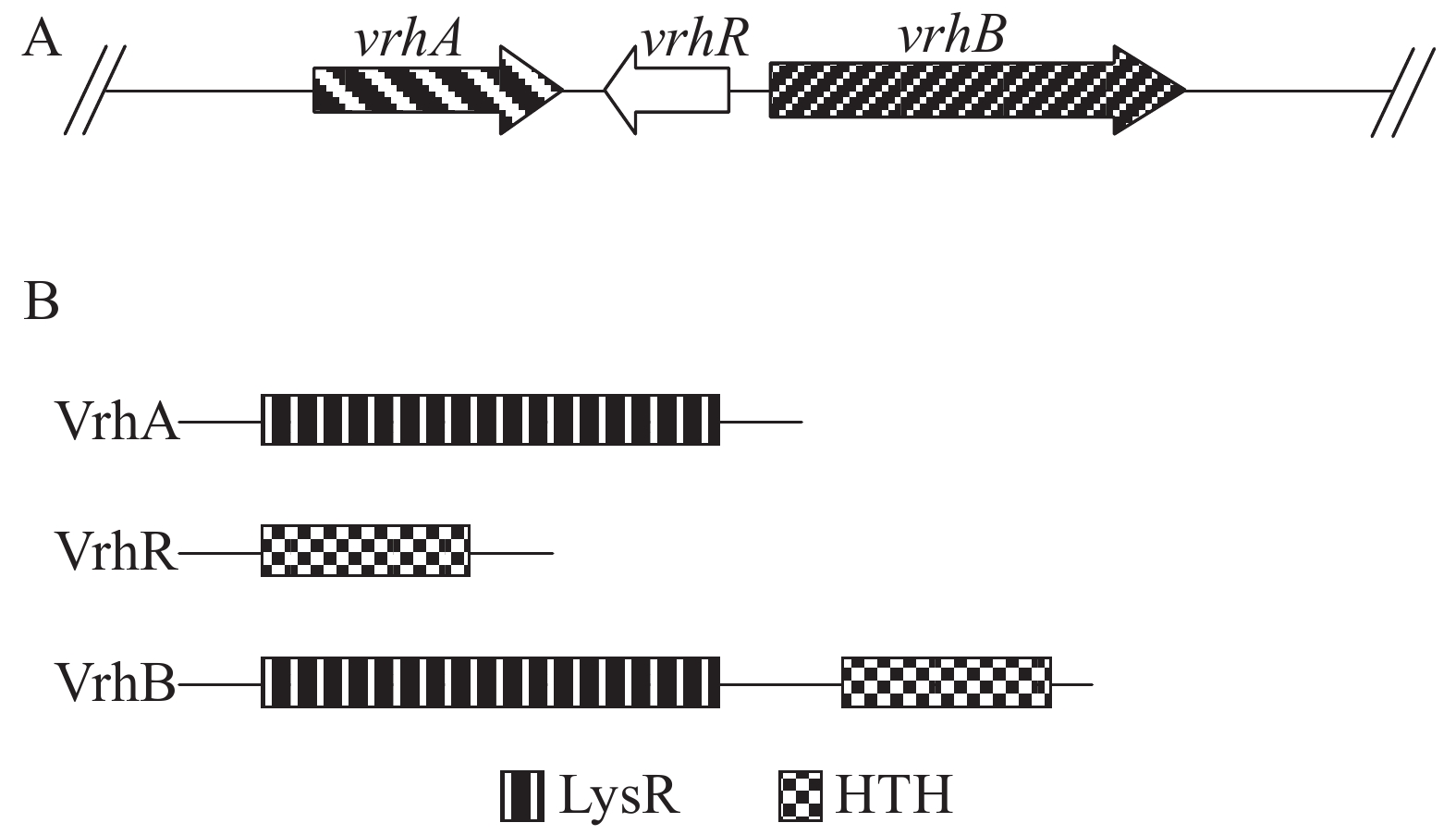
vrhR与另外2个基因vrhA和vrhB共同组成1个基因簇(图1A)。VrhR蛋白只有1个HTH结构域,没有其他结构域(图1B);VrhA只有LysR感应信号结构域(LysRS),可能参与信号感应作用,但没有输出结构域;VrhB为典型的转录调节因子,既有LysRS结构域,又有HTH结构域(图1B)。VrhA,VrhR和VrhB在细菌中,尤其是在植物病原菌中相对保守(图2),暗示VrhA,VrhR和VrhB对细菌的生存可能具有重要的功能,但VrhA的保守性较VrhR和VrhB低(图2),其功能的保守性可能低于VrhR和VrhB。
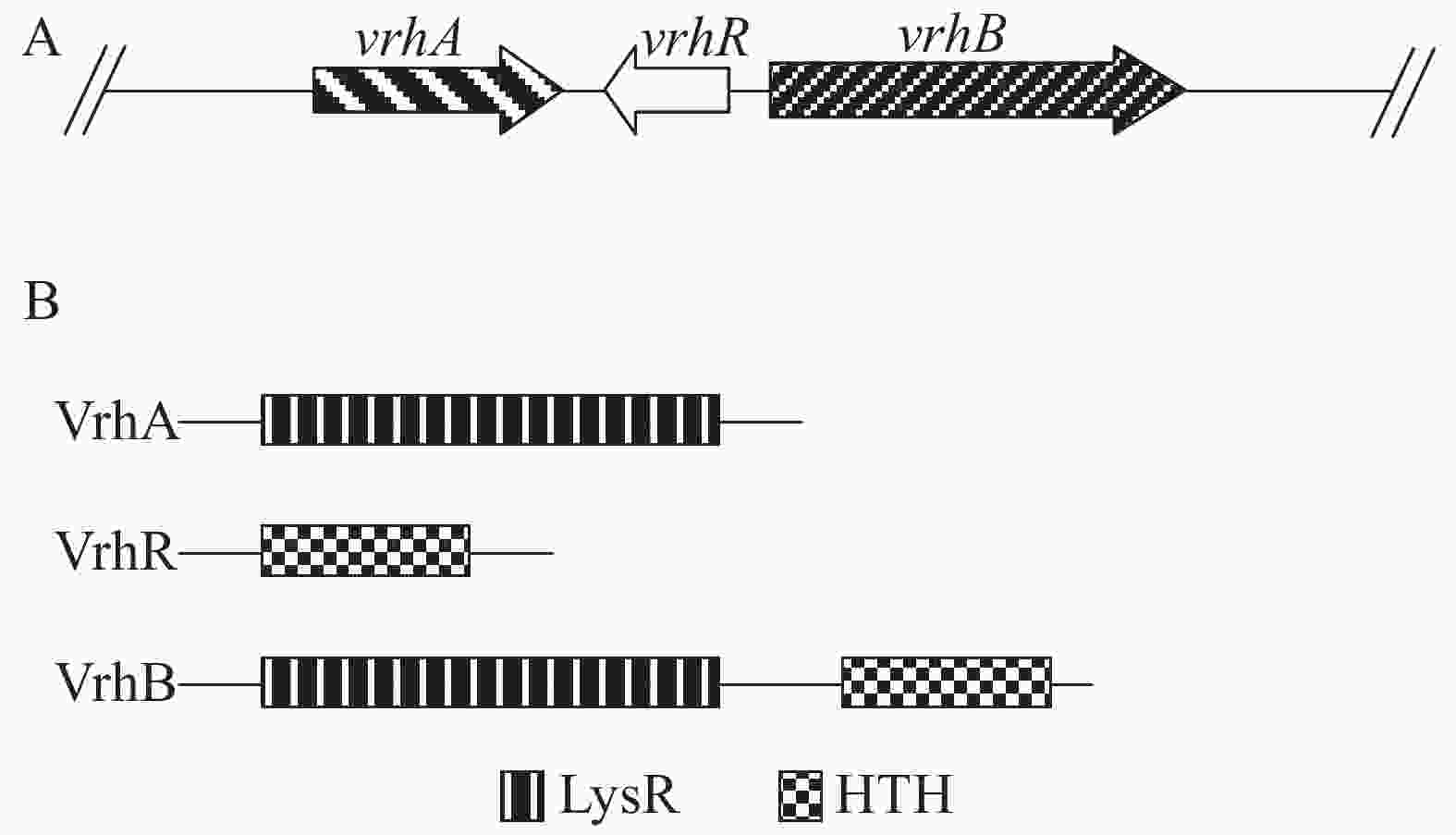
图 1 Xcc 8004中vrhA-vrhB基因簇(A)及其编码蛋白的结构特征(B)
Figure 1. The vrhA-vrhB gene cluster (A) and the domain organization (B) of VrhA, VrhB and VrhR

图 2 VrhA, VrhB和VrhR的同源序列比对
Figure 2. Alignments of
VrhA, VrhB and VrhR homologs -
通过PCR验证缺失突变体,在目的基因上下游约500 bp设计引物,以疑似突变体基因组为模板,以Xcc 8004基因组为对照。如果缺失成功,则扩增片段短于野生型菌株。ΔvrhB,ΔvrhR及ΔvrhA缺失型菌株的基因组PCR片段大约为1 000 bp(图3中1~3泳道验证基因为ΔvrhB;5~9泳道验证基因为ΔvrhR;11~15泳道验证基因为ΔvrhA),比野生型菌株(图3中4,10,16泳道)小,说明这些检测的菌株为突变体。
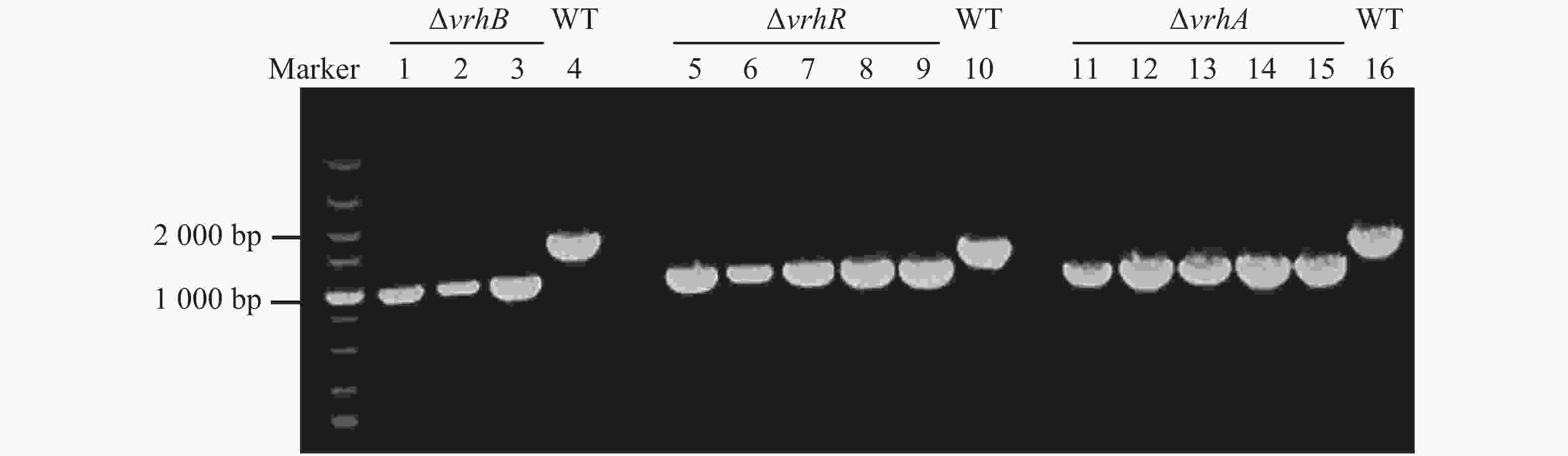
图 3 缺失突变体PCR凝胶电泳验证
Figure 3. PCR validation of the deletion mutants
-
为分析vrhA, vrhB和vrhR对Xcc致病力的影响,利用剪叶法接种野生型菌株Xcc 8004和突变体ΔvrhB,ΔvrhR,ΔvrhA于西兰花叶片,15 d后量取从剪口处到枯斑内部边界的长度,根据病斑长度获得致病力的变化。结果显示,剪接部位均出现不定形淡黄褐色坏死斑现象,ΔvrhR病斑比野生型增长大约1.85 cm,呈极显著性差异(**: P﹤0.01);ΔvrhA和ΔvrhB与比野生型相比,没有显著变化(图4),说明vrhR突变可增强细菌的致病力,而vrhA和vrhB突变对Xcc致病性没有显著影响。

图 4 Xcc 8004,ΔvrhB,ΔvrhR及ΔvrhA突变体的致病性分析
Figure 4. Pathogenicity analysis of Xcc 8004, ΔvrhB, ΔvrhR and ΔvrhA mutants
-
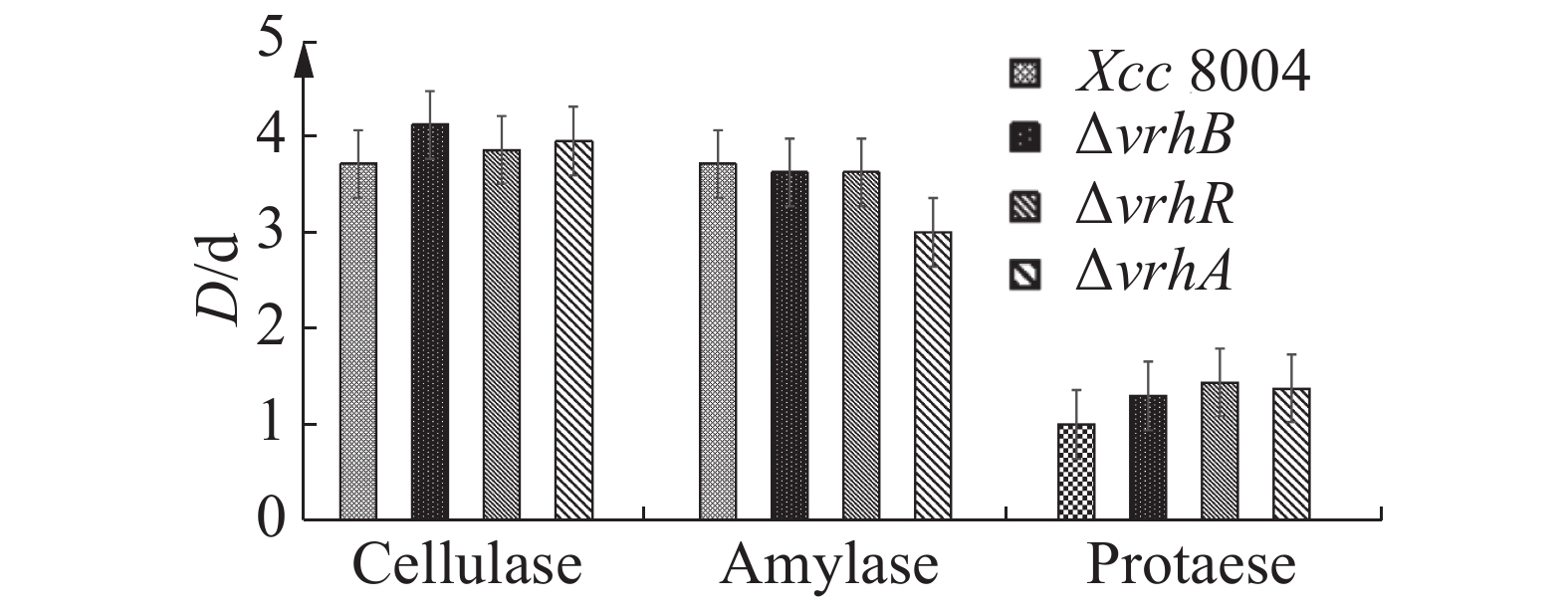
胞外酶是黄单胞属植物病原细菌的重要毒性因子,在病原菌侵染甘蓝的过程中发挥重要作用[6]。为分析ΔvrhR致病力变化是否与其胞外酶活性有关,笔者检测了ΔvrhB,ΔvrhR及ΔvrhA中的蛋白酶、纤维素和淀粉酶活性。结果显示,Xcc 8004,ΔvrhB,ΔvrhR及ΔvrhA菌落所形成的蛋白酶、纤维素酶和淀粉酶酶解圈都没有显著差异(图5),表明vrhR, vrhA和vrhB的突变并不影响胞外蛋白酶、纤维素酶和淀粉酶的活性,且与这些酶的合成或分泌无关。

图 5 Xcc 8004,ΔvrhB,ΔvrhR及ΔvrhA的胞外酶活性检测
Figure 5. Analysis of extracellular enzymes of Xcc 8004, ΔvrhB, ΔvrhR and ΔvrhA
-
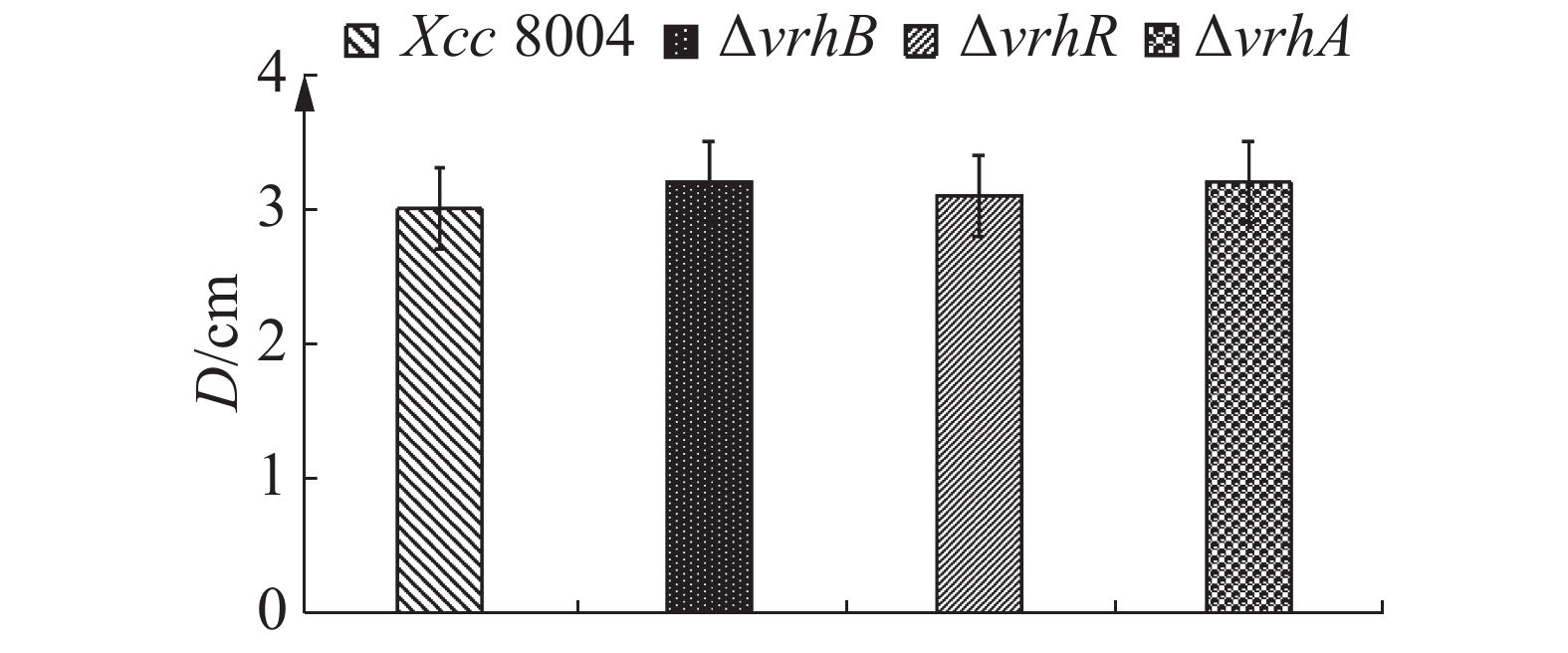
病原菌的致病力与细菌运动能力密切相关[15],因此,笔者采用半固体培养基平板法检测vrhR突变对菌体运动能力的影响。结果表明,在含0.3%琼脂糖的NYG平板上野生型菌株和ΔvrhB,ΔvrhR及ΔvrhA的泳动能力无明显差别(图6),说明vrhR, vrhA和vrhB突变对细菌的运动能力没有影响。
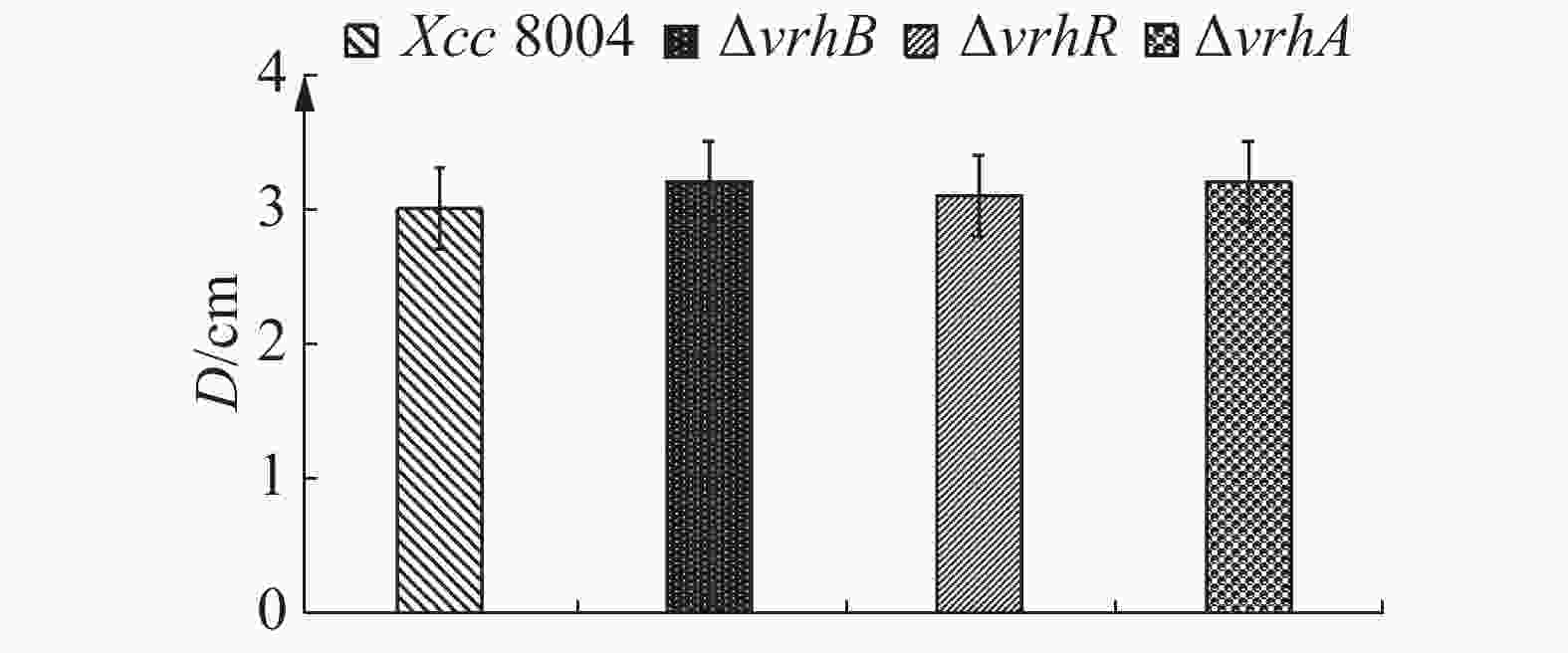
图 6 Xcc 8004,ΔvrhB,ΔvrhR及ΔvrhA游动力测定
Figure 6. Swimming ability of Xcc 8004, ΔvrhB, ΔvrhR and ΔvrhA
-
HTH类DNA结合蛋白是最广泛的转录调节因子,大约95%的转录因子属于HTH家族成员,调控着生物体内大部分基因的表达过程。一般情况下HTH结构域主要与DNA结合,而且活性调控需要其N末端信号输入结构域,此结构通过可感应信号分子或与其他蛋白相互作用来调控自身活性[3]。通过分析Xcc 8004基因组,绝大多数HTH类转录调节因子均匀分布在基因组上,没有HTH热点区域[6]。笔者发现,Xcc 8004只有1个基因簇包含的3个基因(vrhA, vrhR和vrhB)都编码LysR类转录调节因子,这3个基因的阅读框方向并不相同,表明其可能单独转录,各自构成1个转录单元。VrhA, VrhR和VrhB在细菌中广泛存在,保守性很高,表明VrhA, VrhR和VrhB对细菌的生存可能非常重要,功能上可能高度保守。由于Xcc属于高致病性植物病原菌,而这些LysR类转录调节因子可能调控其致病性,笔者在突变这些基因后发现vrhR突变体的致病力显著提高,说明VrhR负调控病原菌与植物的相互作用,但VrhA和VrhB并不参与致病力的调控;这些基因也都不参与胞外酶和游动性的调控;此外,在实验过程中笔者还发现这些突变体并不显著影响细菌的生长(数据未显示),因此,VrhR调控的生理生化途径及其对病原菌致病力的影响还需进一步探索。后续实验应关注VrhR调控的靶标基因,由于VrhR蛋白较小,只有HTH结构域,没有信号输入结构域,其确切的转录调控机制也有待进一步研究。
The Transcriptional Regulator VrhR Negatively Regulates the Virulence of Xanthomonas campestris pv. campestris
-
摘要: 以野油菜黄单胞菌(Xanthomonas campestris pv. campestris, Xcc)转录调节因子VrhR,VrhA和VrhB为研究对象,通过基因敲除和致病性分析确定它们在Xcc与宿主互作过程中的作用。结果表明:VrhR只含有HTH(helix-turn-helix)结构域;在基因组上vrhR位于其他2个lysR类转录调节因子(vrhA和vrhB)之间,但转录方向相反。vrhR突变会提高Xcc的致病能力,但vrhA和vrhB突变对病原菌的致病能力没有影响。同时, vrhR,vrhA和vrhB突变均不影响细菌的运动性及胞外酶(蛋白酶、纤维素酶和淀粉酶)的活性。Abstract: The roles of transcriptional regulators, VrhR, VrhA and VrhB, in the interaction between Xanthomonas campestris pv. campestris and plants were analyzed via gene mutation and virulence analysis. The results showed that VrhR contains only one HTH (helix-turn-helix) motif and is located between the other two LysR transcriptional regulators (vrhA and vrhB) in Xcc 8004 genome, but transcribed in an opposite direction. Mutation of vrhR increased the pathogenicity of Xcc, while mutation of vrhA and vrhB did not affect Xcc virulence. However, mutation of vrhR, vrhA and vrhB did not affect bacterial motility and extracellular enzyme (protease, cellulase and amylase) activities, suggesting that vrhR may regulate bacterial virulence by regulating other pathogenic factors.
-
图 2 VrhA, VrhB和VrhR的同源序列比对
Fig. 2 Alignments of
VrhA, VrhB and VrhR homologs xcb: Xanthomonas campestris pv. campestris 8004; xhr: Xanthomonas hortorum; xva: Xanthomonas vasicola; xga: Xanthomonas gardneri; sdf: Steroidobacter denitrificans; thk: Thauera sp. K11; eba: Aromatoleum aromaticum; tcl: Thauera chlorobenzoica; mela: Melaminivora sp. SC2-9; rme: Cupriavidus metallidurans; bxe: Paraburkholderia xenovorans LB400; axy: Achromobacter xylosoxidans A8; aes: Aeromonas sp. ASNIH5. 黑色:同源性高于90%;灰色:同源性60%~90%。Black: Sequence identity > 90%; Grey: Sequence identity 60%−90%.
图 3 缺失突变体PCR凝胶电泳验证
1~3,5~9,11~15,验证基因分别为ΔvrhB,ΔvrhR和ΔvrhA;4,10,16:野生型对照。
Fig. 3 PCR validation of the deletion mutants
Electrophoresis analysis of the PCR products of ΔvrhB (lanes 1−3),ΔvrhR (lanes 5−9), ΔvrhA (lanes 11−15) and Xcc 8004 (lanes 4, 10, 16).
图 4 Xcc 8004,ΔvrhB,ΔvrhR及ΔvrhA突变体的致病性分析
A:接种15 d后,Xcc 8004,ΔvrhB,ΔvrhR及ΔvrhA接种病斑表型,Xcc 8004为野生型菌株,ddH2O为空白对照。B:接种15 d后,西兰花叶片病斑长度(cm)统计分析(t检验,**, P<0.01)。
Fig. 4 Pathogenicity analysis of Xcc 8004, ΔvrhB, ΔvrhR and ΔvrhA mutants
A: 15 DPI morphology of lesions of broccoli inoculated with Xcc 8004, ΔvrhB, ΔvrhR andΔvrhA with ddH2O as blank control; B: Lesion length of Xcc 8004, ΔvrhB, ΔvrhR and ΔvrhA (t-Test, **: P<0.01).
图 5 Xcc 8004,ΔvrhB,ΔvrhR及ΔvrhA的胞外酶活性检测
Fig. 5 Analysis of extracellular enzymes of Xcc 8004, ΔvrhB, ΔvrhR and ΔvrhA
图 6 Xcc 8004,ΔvrhB,ΔvrhR及ΔvrhA游动力测定
Fig. 6 Swimming ability of Xcc 8004, ΔvrhB, ΔvrhR and ΔvrhA
表 1 引物序列
Table 1 Primer sequences
名称 Primer 序列 Sequence, 5′−3′ 退火温度 Tm/℃ 产物长度 Product length/bp 用途 Uses vrhBqF AAGGATGTGACTGCAACCCG 60.61 147 vrhB qPCR vrhBqR GCTGGCAAAGCGTGCTATTT 60.11 vrhRqF ATGAAACCAGGACGCCCAAT 59.96 121 vrhR qPCR vrhRqR GGGTTGCCAGCTCTTCTTGA 60.25 vrhAqF CGTTCCTCTTCGATCACGGT 59.83 142 vrhA qPCR vrhAqR CTTTGTGGTTTTGGCCGAGG 59.97 rpoDqF GTCGGCTTCAACGACCTGAT 60.39 110 rpoD qPCR rpoDqR TCTTCGTCTGCGTCTTCGTC 60.11 vrhAddFF GACCAAGCTTCAGAACCGGGAGCTAGAACG 60.18 489 vrhA deletion vrhAddFR GACAGGATCCTGTCGTGCATCGCTGCTATC 60.87 vrhAddRF GACAGGATCCAACCGATGCCTAGATCGTAG 56.62 420 vrhA deletion vrhAddRR GACCGAATTCAGCTCTTCTTGAGATATGTC 51.81 vrhRddFF GACCAAGCTTGCATGCACGAATGGTTGCAG 61.07 443 vrhR deletion vrhRddFR GACAGGATCCCTCAGAAGGGTGAAGTGTTC 55.70 vrhRddRF GACAGGATCCCAATCGCTGGCTCAGCTTCA 64.86 437 vrhR deletion vrhRddRR GACCGAATTCGAACGAGCGGGCAAAGCGAC 62.91 vrhBddFF GACCAAGCTTGATGCAGAAGCGCACAATGA 59.55 364 vrhB deletion vrhBddFR GACAGGATCCTCTGTGGATCGCTTCACAAC 58.20 vrhBddRF GACAGGATCCCAGCTCCATCCGCAAGACTC 60.81 404 vrhB deletion vrhBddRR GACCGAATTCTGACTCATCGCATGTGTCAC 58.29 注:下划线表示酶切位点。
Note: Restriction sites are underlined. 下载: 导出CSV
下载: 导出CSV
-
[1] PEREZ-RUEDA E, COLLADO-VIDES J. Common history at the origin of the position-function correlation in transcriptional regulators in archaea and bacteria [J]. Journal of Molecular Evolution, 2001, 53(3): 172 − 179. doi: 10.1007/s002390010207 [2] ARAVIND L, ANANTHARAMAN V, BALAJI S, et al. The many faces of the helix-turn-helix domain: transcription regulation and beyond [J]. FEMS Microbiology Reviews, 2005, 29(2): 231 − 262. doi: 10.1016/j.fmrre.2004.12.008 [3] MADDOCKS S E, OYSTON P C. Structure and function of the LysR-type transcriptional regulator (LTTR) family proteins [J]. Microbiology, 2008, 154(Pt12): 3609 − 3623. [4] DORMEYER M, LUBKE A L, MULLER P, et al. Hierarchical mutational events compensate for glutamate auxotrophy of a Bacillus subtilis gltC mutant [J]. Environmental Microbiology Reports, 2017, 9(3): 279 − 289. doi: 10.1111/1758-2229.12531 [5] DORMEYER M, LENTES S, RICHTS B, et al. Variants of the Bacillus subtilis LysR-Type regulator GltC with altered activator and repressor function [J]. Frontiers in Microbiology, 2019, 10: 2321. doi: 10.3389/fmicb.2019.02321 [6] QIAN W, JIA Y, REN S X, et al. Comparative and functional genomic analyses of the pathogenicity of phytopathogen Xanthomonas campestris pv campestris [J]. Genome Research, 2005, 15(6): 757 − 767. doi: 10.1101/gr.3378705 [7] LIU G F, SU H Z, SUN H Y, et al. Competitive control of endoglucanase gene engXCA expression in the plant pathogen Xanthomonas campestris by the global transcriptional regulators HpaR1 and Clp [J]. Molecular Plant Pathology, 2019, 20(1): 51 − 68. doi: 10.1111/mpp.12739 [8] LU X H, AN S Q, TANG D J, et al. RsmA regulates biofilm formation in Xanthomonas campestris through a regulatory network involving cyclic di-GMP and the Clp transcription factor [J]. PloS One, 2012, 7(12): e52646. doi: 10.1371/journal.pone.0052646 [9] TAO F, HE Y W, WU D H, et al. The cyclic nucleotide monophosphate domain of Xanthomonas campestris global regulator Clp defines a new class of cyclic di-GMP effectors [J]. Journal of Bacteriology, 2010, 192(4): 1020 − 1029. doi: 10.1128/JB.01253-09 [10] LI R F, LU G T, LI L, et al. Identification of a putative cognate sensor kinase for the two-component response regulator HrpG, a key regulator controlling the expression of the hrp genes in Xanthomonas campestris pv. campestris [J]. Environmental Microbiology, 2014, 16(7): 2053 − 2071. doi: 10.1111/1462-2920.12207 [11] WEI K, TANG D J, HE Y Q, et al. hpaR, a putative marR family transcriptional regulator, is positively controlled by HrpG and HrpX and involved in the pathogenesis, hypersensitive response, and extracellular protease production of Xanthomonas campestris pathovar campestris [J]. Journal of Bacteriology, 2007, 189(5): 2055 − 2062. doi: 10.1128/JB.01331-06 [12] WENGELNIK K, VAN DEN ACKERVEKEN G, BONAS U. HrpG, a key hrp regulatory protein of Xanthomonas campestris pv. vesicatoria is homologous to two-component response regulators [J]. Molecular Plant-microbe Interactions: MPMI, 1996, 9(8): 704 − 712. doi: 10.1094/MPMI-9-0704 [13] HU R M, YANG T C, YANG S H, et al. Deduction of upstream sequences of Xanthomonas campestris flagellar genes responding to transcription activation by FleQ [J]. Biochemical and Biophysical Research Communications, 2005, 335(4): 1035 − 1043. doi: 10.1016/j.bbrc.2005.07.171 [14] TAO J, HE C. Response regulator, VemR, positively regulates the virulence and adaptation of Xanthomonas campestris pv. campestris [J]. FEMS Microbiology Letters, 2010, 304(1): 20 − 28. doi: 10.1111/j.1574-6968.2009.01892.x [15] TAO J, LI C, LUO C, et al. RavA/RavR two-component system regulates Xanthomonas campestris pathogenesis and c-di-GMP turnover [J]. FEMS Microbiology Letters, 2014, 358(1): 81 − 90. doi: 10.1111/1574-6968.12529 -

 点击查看大图
点击查看大图
计量
- 文章访问数: 781
- HTML全文浏览量: 216
- PDF下载量: 57
- 被引次数: 0
