


-
木薯(Manihot esculenta)原产于巴西,系大戟科木薯属作物,在当今世界广泛栽培,是世界三大薯类作物之一。木薯因其独特的特性被称为“淀粉之王”,具有耐贫瘠、高产、四季可收的特点,并广泛应用于食品、饲料和能源等领域。植物激素作为植物体内微量而高效的小分子有机化合物,参与调控了植物的各种生理过程。生长素(Auxin)作为一种重要的植物激素,在植物的生长发育中起到重要作用。相关研究表明,生长素是通过其信号通路的各个组分来传递信号,进而使下游基因表达。而ARF(Auxin response factor)作为生长素信号通路的响应因子,在生长素信号通路中扮演转录因子的角色,可以与特定的启动子序列结合,从而激活下游基因的转录,因此ARF对于生长素信号通路具有重要意义,但是对于ARF的功能以及作用机理的研究尚不完善。ARF在植物的生长发育及逆境胁迫中发挥了非常重要的作用。相关研究表明,在拟南芥中,ARF17可以通过结合GPI-anchored lipid transfer protein4(LTPG4)基因启动子上的AuxRE(Auxin response element)元件,调控其表达进而影响花粉管的生长[1]。另外,拟南芥激活型生长素响应因子ARF6/8/7/19可以促进不定根的再生[2]。有研究表明,ARF还参与植物的向性运动。在拟南芥中,重力或光触发的生长素不对称分布诱导了ARF7和ARF19在下胚轴中对Small Auxin-Up RNA19(SAUR19)亚家族基因的不对称表达,从而导致了向地性和向光性反应过程中的弯曲生长[3];ARF7与PR-1 homolog(PRH1)基因的启动子结合进而调控PRH1的转录表达,从而影响拟南芥侧根的发育[4]。在番茄中,SlARF3可以通过调控苹果酸代谢过程相关基因的表达,促进果实中有机酸的积累,最终影响果实的风味品质[5]。OsARF18可以通过抑制水稻根的伸长,从而影响水稻的株高[6]。OsARF4通过生长素和油菜素类固醇途径调节水稻叶片倾斜,进而影响其光合效率和谷物产量[7]。此外,miR167d-ARF模块可以调控花朵开放和柱头大小[8]。GhARF2在棉花纤维伸长过程中具有较高的表达模式,GhARF2b过表达可以抑制棉纤维的伸长[9]。ARF基因广泛参与了对植物生长发育的调控。ARF还参与了植物的逆境胁迫。在水稻中,OsARF17在植物对不同类型植物病毒的防御中起着至关重要的作用,这些病毒利用独立进化的病毒蛋白来靶向生长素信号传导的这一关键成分,并促进感染[10]。ARF还参与植物翻译后的调控过程从而参与植物免疫,在拟南芥中过表达microRNA167,调控其靶点ARF6和ARF8,进而调控水杨酸的含量,从而增强对丁香假单胞菌(Pseudomonas syringae pv. tomato DC3000,Pst DC3000)的防御[11]。此外,ZmARF4的过表达可促进磷从根部向叶片的再动员和转运,进而增强了玉米对磷胁迫的耐受性[12]。在大豆中,ARF16激活Myelocytomatosis protein2(MYC2),通过下调脯氨酸的生物合成来降低耐盐性[13]。
在本研究中,团队获得了1个MeARF基因(Manes.18G015400.1),并对该基因进行了生物信息学分析,以及验证了其在木薯不同组织中的相关表达量,同时还筛选了MeARF的相关互作蛋白,旨在为进一步解析MeARF的功能和作用机理奠定理论基础。
-
本研究使用的木薯品种为‘华南124’,培养方法为盆栽,固体培养基由蛭石和营养土3∶1的比例混合制成,截取2 ~ 3个芽点的木薯茎秆,将其插入在培养基中培养,置于温室中培养,培养温度为28 ℃,湿度为80%。
-
取约0.3 g的木薯侧芽放于加入液氮的研钵中进行迅速研磨,将研磨好的样品使用CTAB裂解细胞的方法提取RNA(注意使用无酶EP管,在冰上进行实验),之后用分光光度计测定RNA的浓度,将所得的RNA放置于−80 ℃保存。
-
使用反转录试剂盒(YEASEN,11151ES10/11151ES60)将RNA反转录为cDNA。配置反转录体系(表1)于1.5 mL无酶离心管,在37 ℃水浴条件下反应5 min用于cDNA合成及gDNA的消化,最后在85 ℃水浴的条件下放置30 s,由此来终止反应,将所得反应产物放置于−80 ℃保存。
表 1 反转录体系
Table 1. The reverse transcription system
组分
Component体积/μL
Volume /μLRNase-free Water 12 4×Hifair® AdvanceFast One-Step RT SuperMix 5 gDNA Remover Mix 1 mRNA 2 -
通过开放数据库Phytozome(
https://phytozome-next.jgi.doe.gov/ )查找木薯MeARF的相关信息,发现MeARF的CDS区域长度为2 861 bp;由于MeARF全长过大,本研究以cDNA为模板对MeARF进行了分段克隆并送测序后,合成全长大小为2 861 bp的目的片段,并连上pEASY-Blunt3载体(Transgen,CB301-01),然后以Blunt3-MeARF为模板进行片段的扩增。PCR反应体系20 μL,包括Mix(Vazyme,P112-03)10 μL,ddH2O 8 μL,正向引物0.5 μL,反向引物0.5 μL,模板1 μL(PCR反应程序如表2所示)。表 2 PCR反应程序设置
Table 2. Setup of PCR reaction program
名称
Name温度/℃
Temperature/℃时间/min
Time /min预变性(Initial denaturation) 95 3.0 变性(Degeneration) 95 0.5 退火(Annealing) 55 0.5 延伸(Extension) 72 3.0 循环次数(Number of cycles) 36 最终延伸(Final extension) 72 5.0 -
通过开放数据库Phytozome查找木薯MeARF的氨基酸序列,将该序列进行Blast比对,得到不同物种中与MeARF同源性较高的氨基酸序列,将这些序列输入MEGA11软件上绘制成进化树。
-
通过Uniprot在线数据库(
https://www.uniprot.org/id-mapping/ )和SWISS-MODEL网站(https://swissmodel.expasy.org/ )进行MeARF蛋白结构的预测分析。 -
使用Primer5软件进行定量引物的设计,设计了前端引物QMeARF-F和后端引物QMeARF-R。以cDNA(木薯的茎、叶柄、叶片、侧芽和须根)为模板,用荧光定量PCR仪进行扩增,PCR程序如表3所示。反应体系为:ddH2O 6.5 μL,cDNA 0.2 μL,前端引物0.4 μL,后端引物0.4 μL,SYBR荧光染料(Vazyme,Q111-02)7.5 μL。
表 3 荧光定量PCR程序
Table 3. The fluorescence quantitative fluorescence PCR procedure
名称
Name温度/℃
Temperature/℃时间/s
Time/s预变性(Initial denaturation) 95 30 变性(Degeneration) 95 15 退火(Annealing) 55 15 延伸(Extension) 72 20 循环次数(Number of cycles) 45 -
使用(
https://crm.vazyme.com/cetool/singlefragment.html )网站设计特异性引物MeARF-AD-F/R,酶切位点选择NdeⅠ和EcoRⅠ,以Blunt3-MeARF质粒为模板,进行PCR扩增,回收目的片段,酶切pGADT7载体并进行胶回收,通过同源连接的方法进行目的基因和线性载体的连接,转入大肠杆菌感受态细胞DH5α(Zomanbio,ZK206),在氨苄抗性的固体培养基培养10 h,挑取单菌落,并以挑取的菌落为模板进行PCR验证,将得到的阳性克隆对应的菌液加入到相应抗性的液体LB中,置于摇床中进行培养(37 ℃,150 r·min−1),提取质粒,并进行双酶切验证,送测序,将测序的结果与Phytozome网站上的MeARF序列进行校对,与目的片段的序列一致,则完成相关基因表达载体的构建。本研究引物合成均由华大基因公司完成(表4)。表 4 引物碱基序列
Table 4. Primer base sequences
引物名称 Primer 碱基序列(5′−3′) Base sequence(5′−3′) QMeARF-F ATGATGATGGGTTCTGTTG QMeARF-R GCTCCGAGTTTATTGTCTT MeARF-AD-F GTACCAGATTACGCTCATATGATGCATTTTAAACAGAAGCACATGA MeARF-AD-R ATGCCCACCCGGGTGGAATTCTCAAGCACGGCTACCTTCTGT pGBKT7-F GTAATACGACTCACTATAGGGCGA pGBKT7-R TTTTCGTTTTAAAACCTAAGAGTC -
取−80 ℃保存的AH109菌株,划线于YPDA固体平板上,置于28 ℃培养2 ~ 3 d;挑取平板上生长的酵母单菌落,置于5 mL的YPDA液体中,28 ℃摇床培养8 ~ 10 h;吸取一定量的AH109菌液,置于50 mL的YPDA溶液中进行转摇,调节初始OD600值为0.2 ~ 0.3,之后置于28 ℃培养3 ~ 5 h,使OD600值到达0.6左右;将菌液分装在50 mL离心管中,4 000 r·min−1,低转速离心10 min,去上清;每管加入20 mL 的ddH2O重悬,4 000 r·min−1,低转速离心10 min,去上清;加入2 mL的1×TE/LiAC重悬,完成酵母感受态细胞的制备。
-
将ssDNA,木薯pGBKT7文库质粒,pGADT7-MeARF质粒各10 μL混合于2 mL离心管中,做8 ~ 10组;每管加入200 μL的酵母感受态细胞;加600 μL的PEG/LiAC,上下颠倒混匀,在28 ℃摇床培养1 h;加入70 μL的DMSO(Solarbio,D8370−100),42 ℃水浴热激30 min;12 000 r·min−1离心30 s,去上清;沉淀用300 uL的1×TE重悬后涂布于SD-Trp-Leu(Coolaber,PM2220)和SD-Trp-Leu-Ade-His(Coolaber,PM2110)固体培养基上,放置于28 ℃的培养箱中进行培养。
-
将转化后的酵母在28 ℃培养箱培养3 d,挑取在SD-Trp-Leu-Ade-His固体培养基上生长的酵母单菌落于5 mL的SD-Trp-Leu-Ade-His液体培养基中,在28 ℃的摇床中培养;3 d后,提取酵母质粒,分别使用MeARF-AD-F/R和pGBKT7-F/R引物对质粒进行扩增,将阳性克隆对应的目的片段进行回收,把回收的片段送测序。使用开放数据库Phytozome对测序的结果进行比对分析,最终得到与MeARF相互作用的候选蛋白。
-
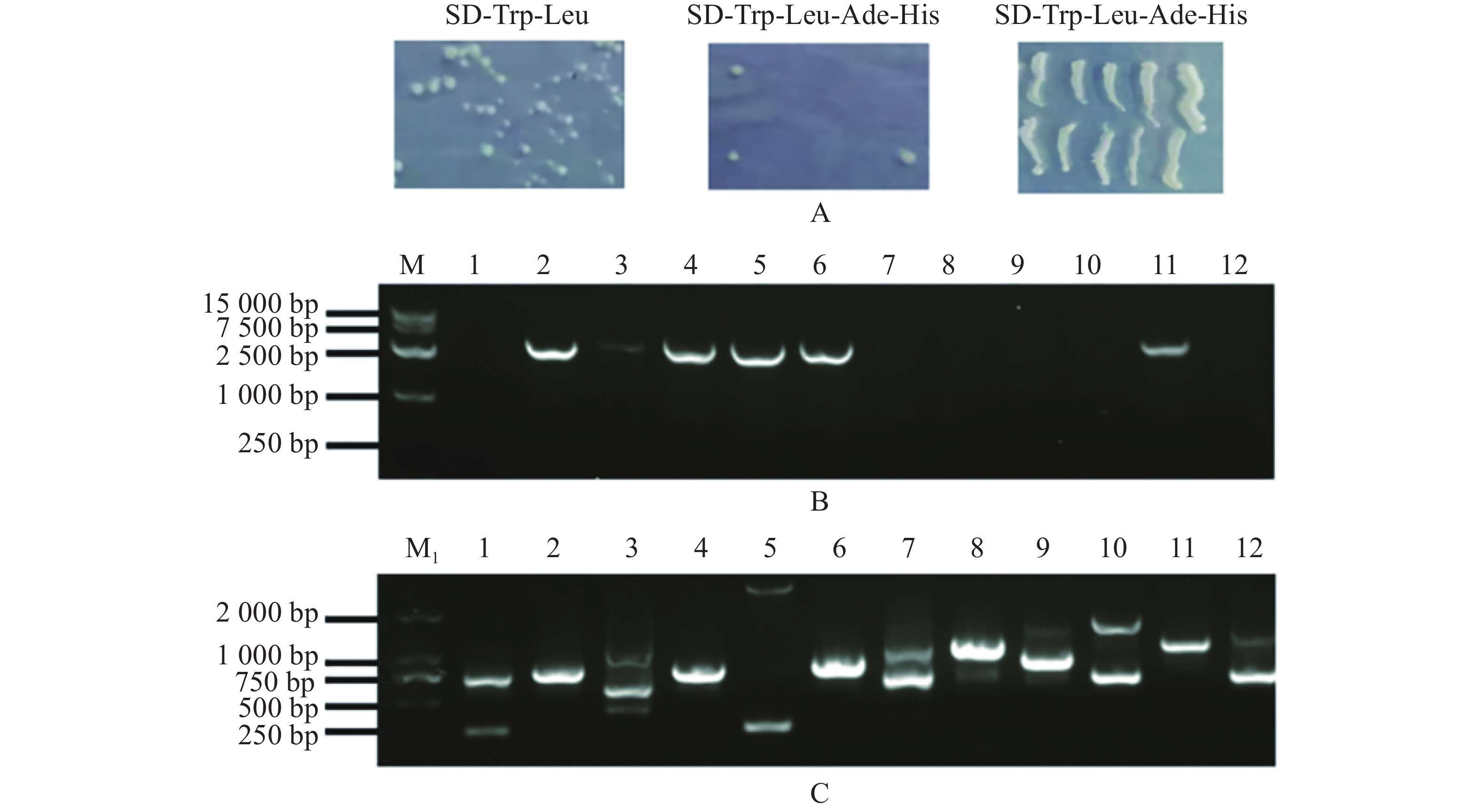
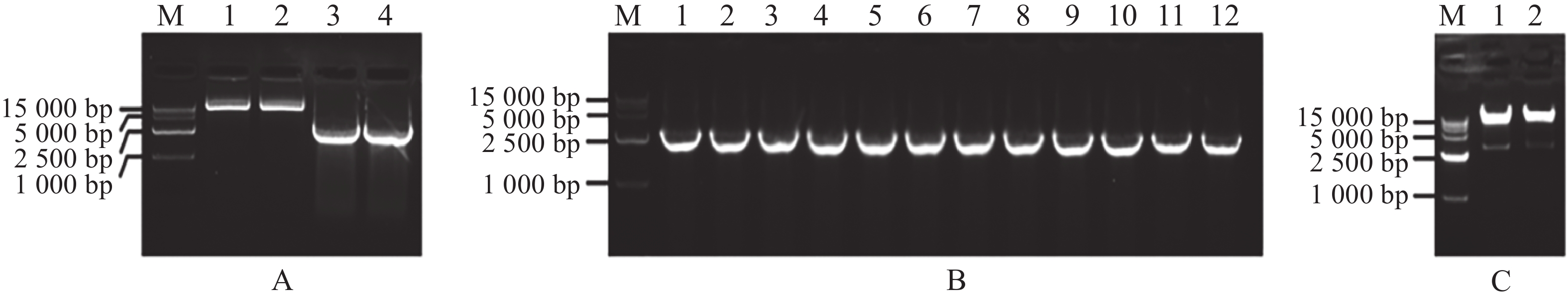
扩增MeARF片段,同时将pGADT7载体质粒用Nde Ⅰ和EcoR Ⅰ进行酶切,电泳条带大小符合预期实验结果(图1−A);挑取单菌落进行菌落PCR,电泳结果表明存在条带清晰单一的阳性克隆(图1−B);提取质粒进行双酶切验证,酶切位点选择Nde Ⅰ和EcoR Ⅰ,结果表明,酶切后的条带大小约为2 800 bp,符合目的片段的大小(图1−C)。将质粒送序列检测后结果一致,最终完成相关表达载体的构建。
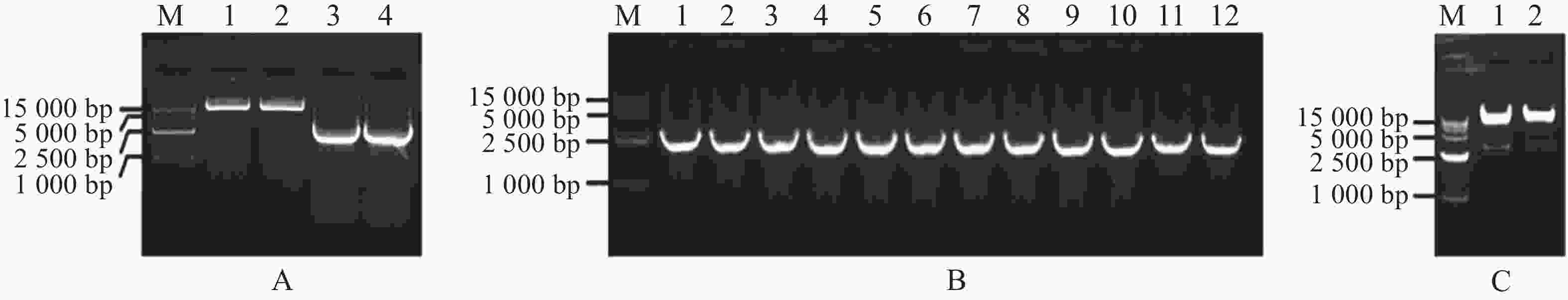
图 1 MeARF片段扩增以及构建基因表达载体pGADT7-MeARF
Figure 1. Amplification of the MeARF fragments and constructing the expression vector of pGADT7-MeARF
-
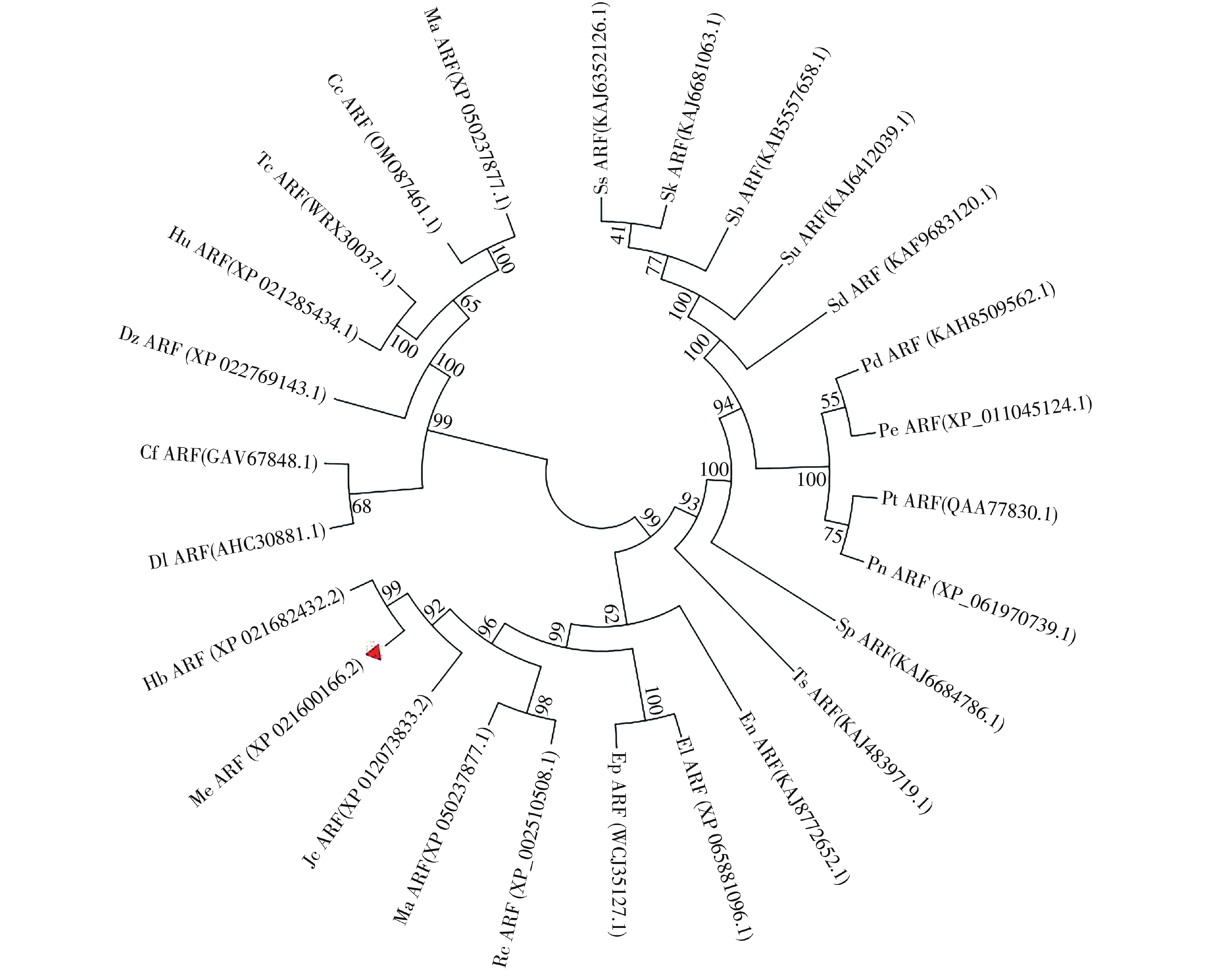
选取26个不同物种与MeARF同源性较高的氨基酸序列,将这些序列与MeARF的氨基酸序列一起输入到MEGA软件上绘制成进化树(图2)。结果表明,大戟科橡胶树属的橡胶树中的HbARF与MeARF的亲缘关系较近,用DNAMAN软件比对二者的氨基酸序列,置信度为86.73%。
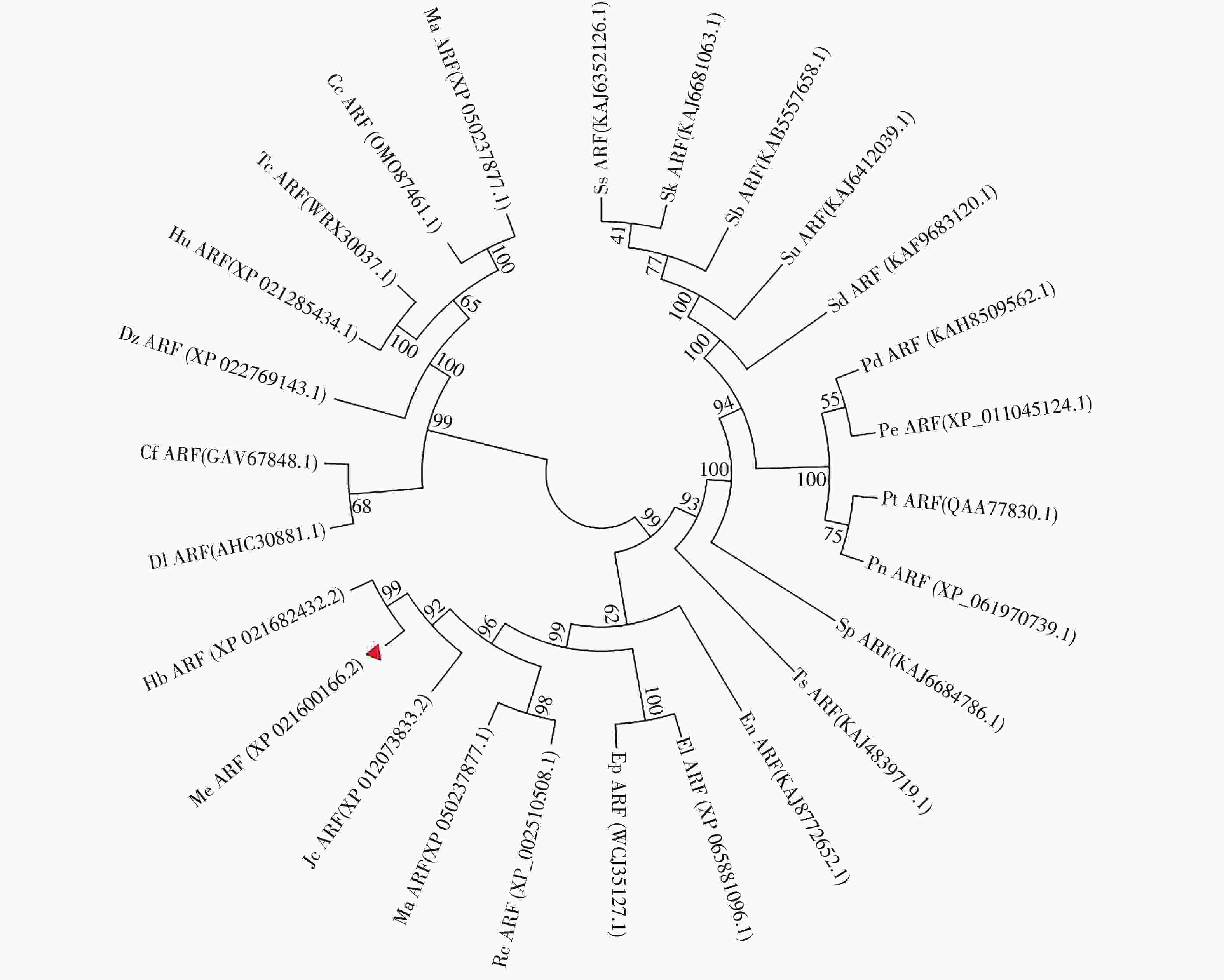
图 2 构建ARF进化树
Figure 2. Construction of the evolutionary tree of ARF
-
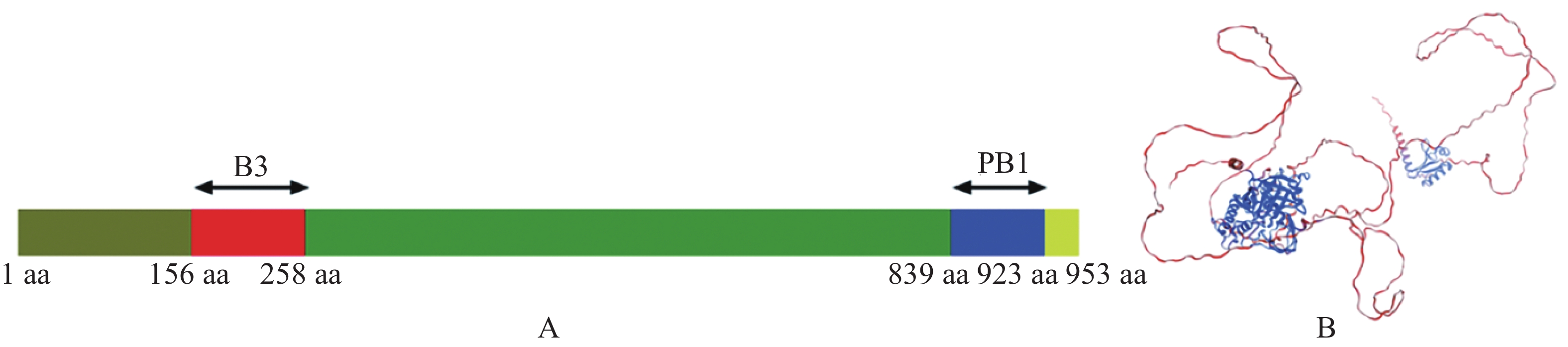
为了解析MeARF的功能,本研究首先分析了MeARF的蛋白结构。发现MeARF的结构域包含一个位于N端156~258 aa的B3结构域(B3 DNA binding domain,B3),该区域与结合元件AuxRE(Auxin-responsive element,AuxRE)有关。该蛋白的C端含有一个位于839~923 aa处的PB1结构域(Phox and Bem1p,PB1),能够与Aux/IAA(Auxin/indoleacetic acid,Aux/IAA)互作,稳定ARF的二聚体并促进其结合DNA(图3)。
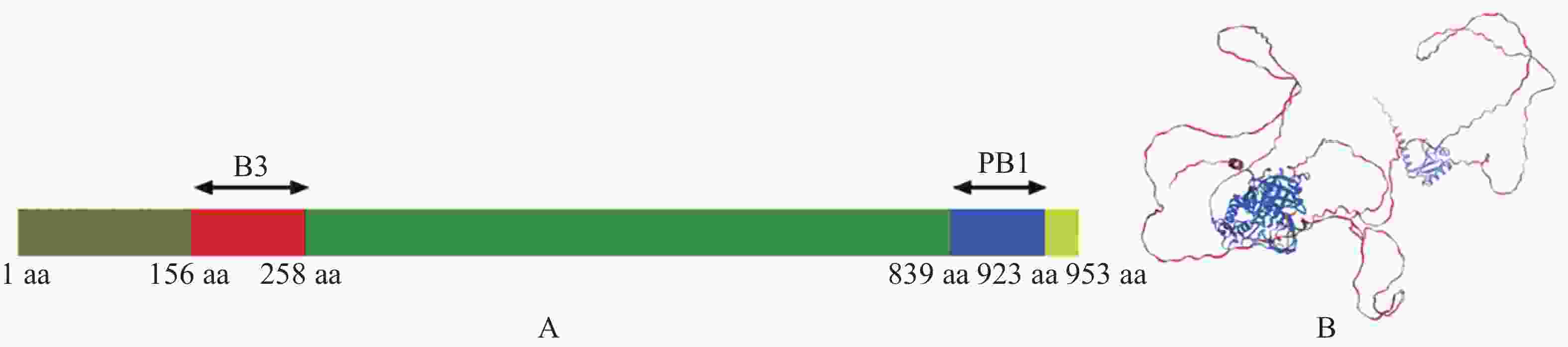
图 3 MeARF蛋白结构分析
Figure 3. Structural analysis of the MeARF protein
-
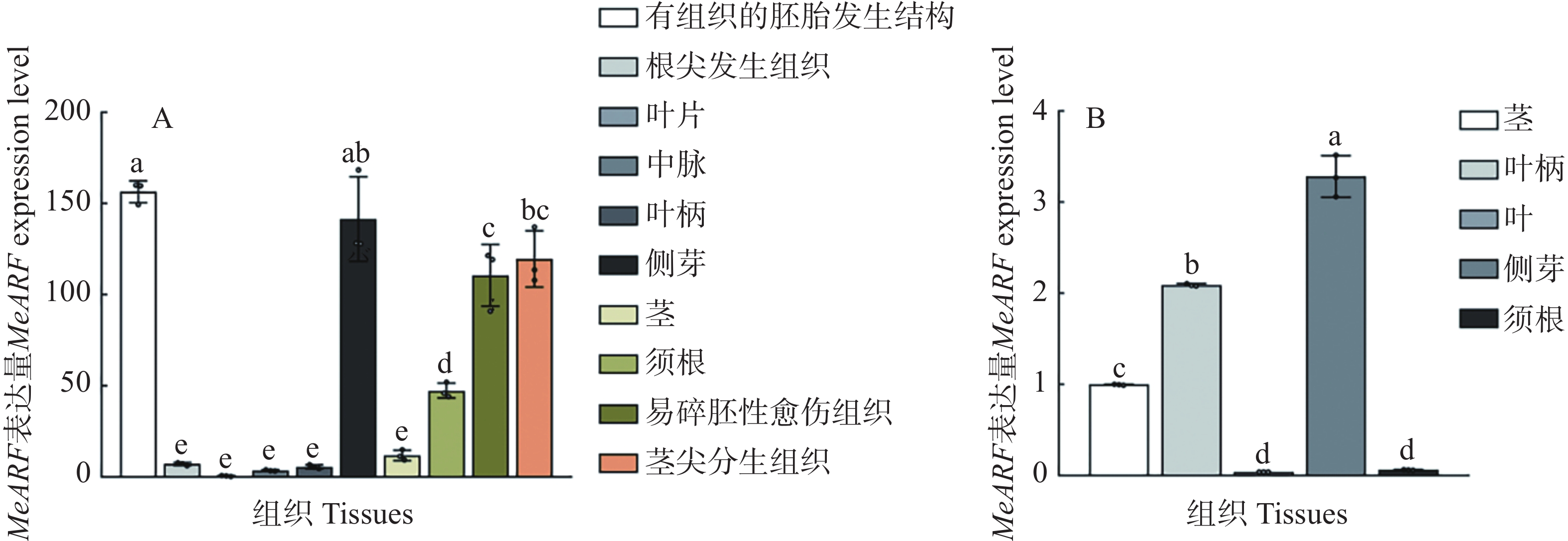
为了分析MeARF在木薯不同组织中的表达情况,本研究利用在线数据绘制图4−A。结果发现,MeARF在有组织的胚胎发生结构和侧芽的表达量较高;并且相关研究表明生长素可以调控植物胚胎的发育并促进侧芽的萌发[14 − 15],这些结果进一步说明了MeARF与生长素之间的联系。
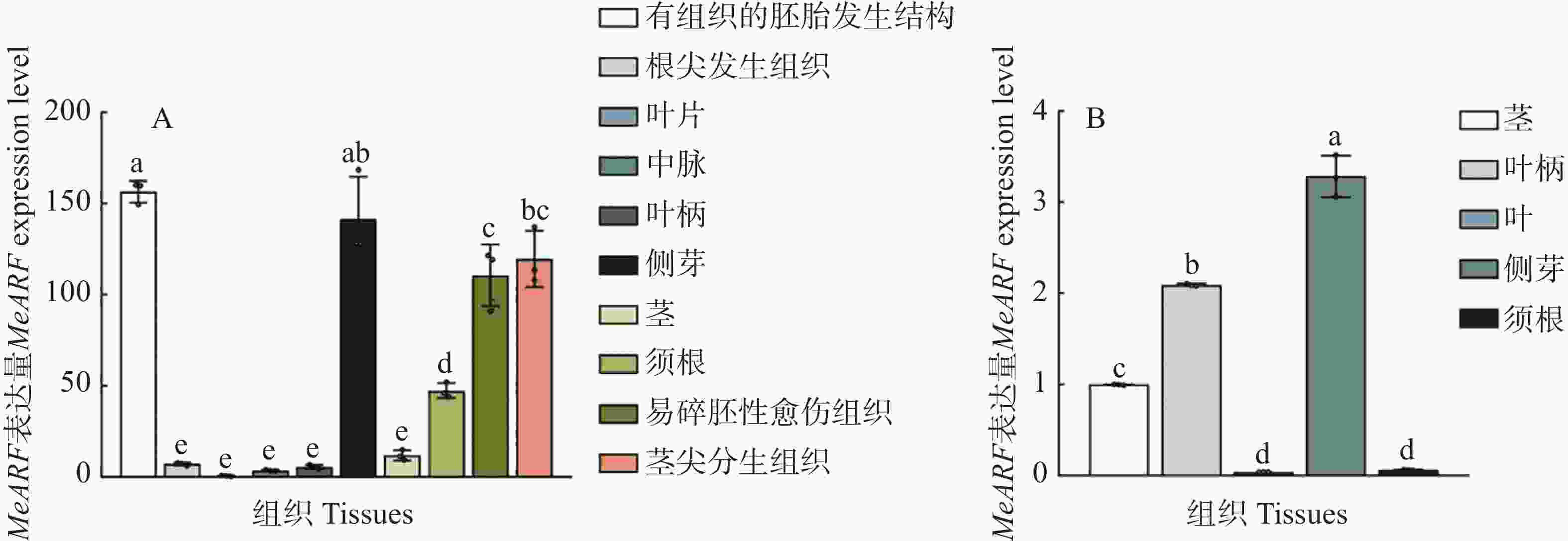
图 4 MeARF在木薯不同组织的表达量
Figure 4. Expression profiling of MeARF in different tissues of cassava
为了验证MeARF在木薯不同组织中的表达情况,本研究测定了MeARF在木薯茎、叶柄、叶片、侧芽和须根的表达量,结果发现MeARF在木薯侧芽的表达量最高,在木薯叶片的表达量最低,这个结论与在线数据库的结果类似,进一步说明了MeARF在木薯的不同组织都存在表达(图4−B)。
-
为探讨pGBKT7-MeARF进行酵母双杂交筛选pGADT7文库的可能性,本研究首先进行了pGBKT7-MeARF的自激活实验,通过观察pGBKT7与pGBKT7-MeARF两种组合分别在SD-Trp和SD-Trp-Ade-His固体培养基的生长情况来判断pGBKT7-MeARF是否有自激活现象。实验采用了4种浓度梯度稀释接种在培养基上。实验结果表明,pGBKT7与pGBKT7-MeARF两种组合均在SD-Trp固体培养基上生长;而在SD-Trp-Ade-His固体培养基上,只有pGBKT7-MeARF组合的酵母生长(图5)。这些现象说明了pGBKT7-MeARF存在自激活现象,可以单独激活下游基因的表达。
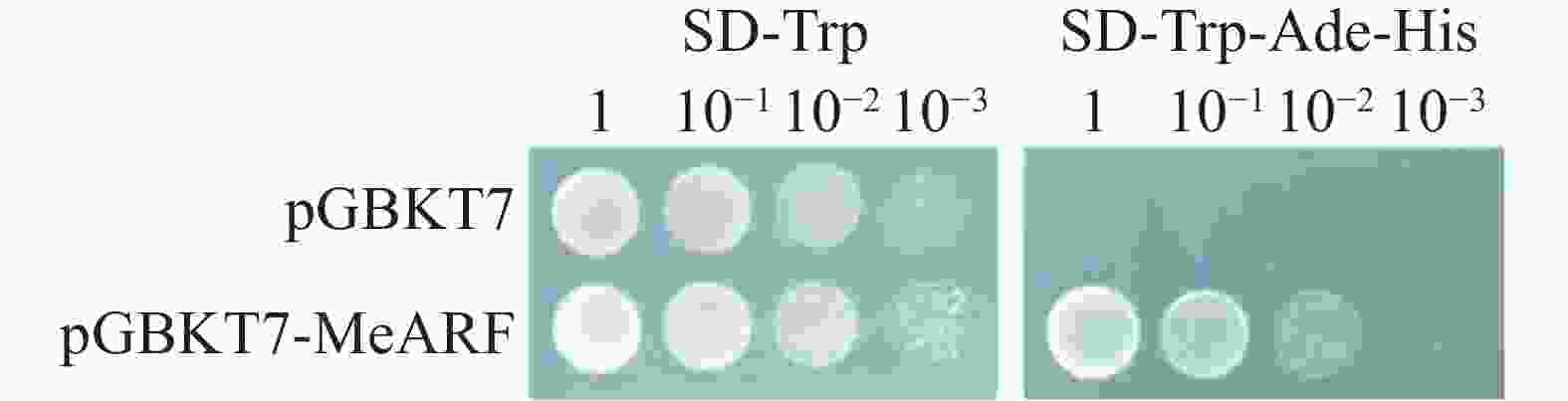
图 5 pGBKT7-MeARF自激活验证
Figure 5. The self-activation validation of pGBKT7-MeARF
由于pGBKT7-MeARF存在自激活现象,本实验采用pGADT7-MeARF作为诱饵蛋白对木薯的pGBKT7文库进行筛选,发现在SD-Trp-Leu固体培养基上均有酵母生长,在SD-Trp-Leu-Ade-His固体培养基上一共得到了12个单菌落,把这些单菌落在SD-Trp-Leu-Ade-His固体培养基上划线保存,发现其可以在SD-Trp-Leu-Ade-His固体培养基上正常生长(图6−A)。挑取这些单菌落,摇菌,提取酵母质粒,使用MeARF-AD-F/R引物进行PCR扩增,琼脂糖凝胶电泳结果表明存在6个阳性克隆(图6−B);另外,使用pGBKT7-F/R引物进行菌落进行扩增,共得到8个单一条带(孔道3含有3条单一条带)(图6−C)。选择对应的目的条带进行切胶回收,并将回收的片段送测序。
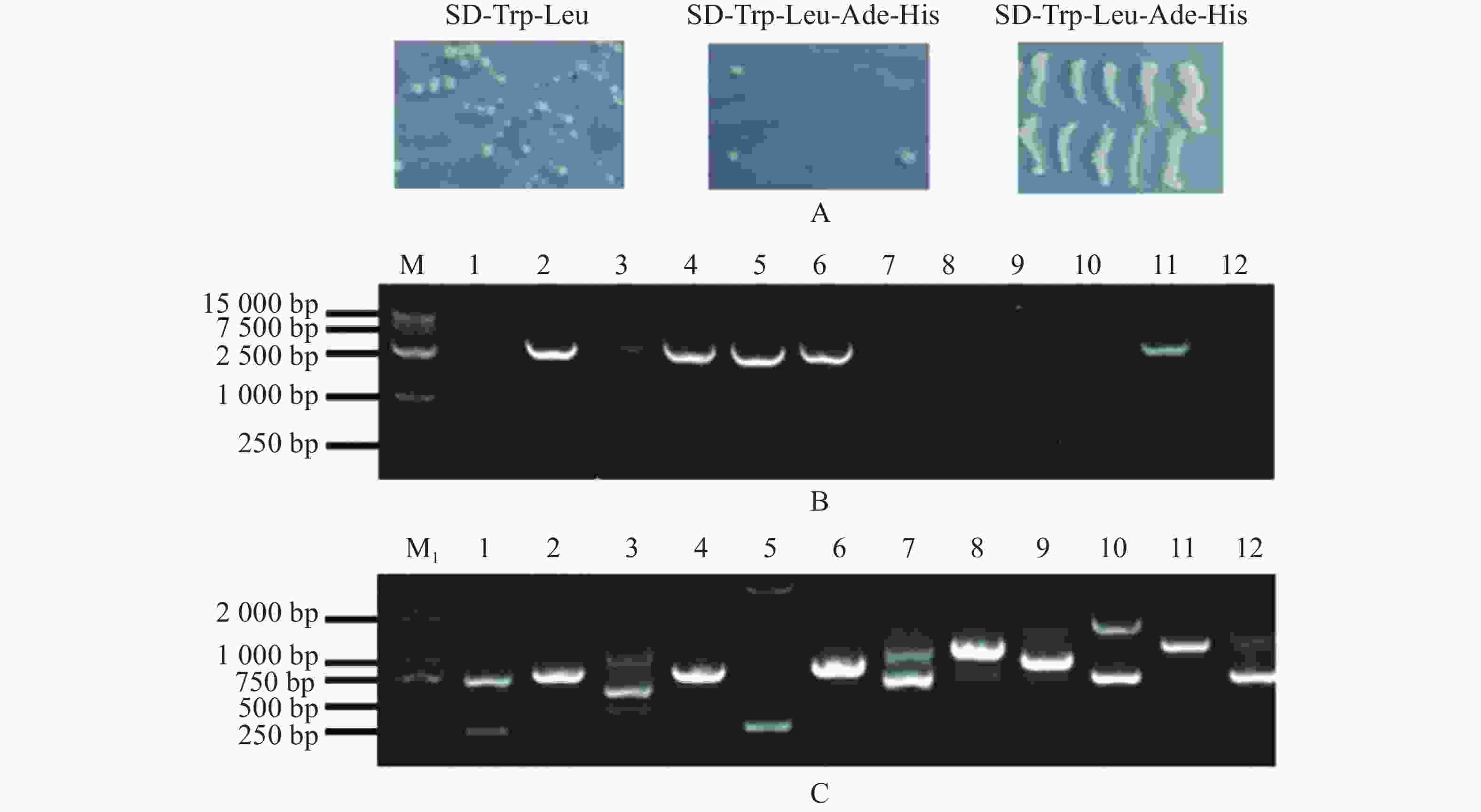
图 6 pGADT7-MeARF筛选木薯pGBKT7文库
Figure 6. Screening of the cassava pGBKT7 library by using pGADT7-MeARF
-
对测序的结果进行比对,最终得到了7个候选蛋白(表5),分别是线粒体相关的NADH脱氢酶[泛醌]-铁硫蛋白8-a(Manes.15G163400.1)、依赖钠的磷酸盐转运蛋白(Manes.14G035900.1)、多聚泛素4(Manes.14G035900.1)、GTP酶激活蛋白REN1(Manes.02G146100.1)、木薯生长素应答蛋白IAA8(Manes.05G187100.1)、溶质载体家族35(Manes.17G049400.1)、富含亮氨酸的重复蛋白,一些含有F-box的蛋白质(Manes.05G050000.1)。使用数据库中的E值和置信度来反映比对结果的可靠性,结果表明这7个测序结果与候选蛋白序列比对的置信度都在90%以上,E值接近或者等于0,这些结果表明了序列比对的准确性。
表 5 序列比对结果
Table 5. Results of the sequence alignment
基因ID
Gene ID基因注释
Gene annotationE值
E value置信度/%
Confidence level/%Manes.15G163400.1 NADH DEHYDROGENASE [UBIQUINONE] IRON-SULFUR PROTEIN 8-A, MITOCHONDRIAL-RELATED 0 99.77 Manes.14G035900.1 SODIUM-DEPENDENT PHOSPHATE TRANSPORTERS 1.01E-20 100.00 Manes.04G111000.1 POLYUBIQUITIN 4 0 98.23 Manes.02G146100.1 RHO GTPASE-ACTIVATING PROTEIN REN1 2.64E-01 96.00 Manes.05G187100.1 AUXIN-RESPONSIVE PROTEIN IAA8 0 99.54 Manes.17G049400.1 SOLUTE CARRIER FAMILY 35 0 100.00 Manes.05G050000.1 LEUCINE RICH REPEAT PROTEINS, SOME PROTEINS CONTAIN
F-BOX2.65E-02 93.00 -
ARF作为生长素信号通路的重要组成部分,在植物的生长发育以及逆境胁迫中发挥着重要作用[2, 10]。已经有27个ARF基因在木薯中被鉴定[16],但其在木薯发育及逆境胁迫的研究较少。本研究通过对其蛋白结构以及在木薯中各组织中的相对表达量的分析,并筛选到相关互作蛋白,为这一基因的功能研究提供研究基础。
为了探究MeARF的功能,本研究首先通过构建进化树的方法,发现大戟科橡胶树属的橡胶树中的HbARF与MeARF的亲缘关系较近。ARF结构的多样性,影响了其功能的差异性,进而影响生长素的作用[17 − 19],因此深入对ARF的蛋白结构进行研究,有利于更好解析生长素在植物中的作用。当前的研究普遍认为,ARF蛋白主要包括3个结构域:N端的DNA结合域(DNA binding domain,DBD)、中间区域(Middle region,MR)和C端PB1结构域[17]。DNA结合域主要负责与生长素响应元件结合,其与顺式作用元件的相互作用是通过植物特有的B3亚结构域发生的[18]。MR结构域作为ARF功能的决定因素,其参与了生长素感知、蛋白质可用性控制和DNA结合调节的过程中,还具有作为相互作用枢纽的潜在作用[19]。PB1结构域具有稳定ARF与DNA的结合和与Aux/IAA相互作用的双重功能[20]。本研究通过对MeARF蛋白的三维结构进行分析,发现了MeARF蛋白含有B3和PB1两个结构域,但是对于MeARF的B3结构域是否可以与下游的生长素响应元件结合还尚不清楚。通过在线数据库预测,发现MeARF在木薯的很多部位都存在表达,其中,MeARF在有组织的胚胎发生结构和侧芽的表达量较高,说明MeARF可能参与木薯胚胎和侧芽的发育,这为这一基因的功能研究提供了潜在的基础。
ARF作为转录因子,通过与一系列蛋白互作在植物中发挥作用。相关研究发现,生长素响应因子ARF与Ethylene response factor108(ERF108)相互作用,介导棉花纤维次生细胞壁的生物合成[21]。还有研究表明,植物激素油菜素内酯、生长素和赤霉素通过Brassinazole resistant(BZR)-ARF-Phytochrome-interacting factor(PIF)/DELLA的协同作用调控拟南芥中光形态建成相关的下胚轴伸长[22]。另外,ARF2与Ethylene-insensitive3-like1(EIL1)之间的相互作用可增强EIL1的活性,从而促进木瓜成熟过程中相关基因的表达[23]。在拟南芥中,ARF4能够通过竞争ARF5与IAA12结合来调控芽再生过程[24]。MeARF作为一个转录因子,其可能存在于一个巨大的互作网络中,本研究通过酵母双杂交实验获得了7个互作蛋白,其中一部分是与植物生长发育相关的蛋白,包括了线粒体相关的NADH脱氢酶[泛醌]-铁硫蛋白8-a、GTP酶激活蛋白REN1、木薯生长素应答蛋白IAA8和富含亮氨酸的重复蛋白,一些含有F-box的蛋白质。其中,对于模式植物拟南芥,其线粒体中的NADH脱氢酶亚基可以挽救slow growth3(slo3)突变体的生长缺陷以及种子的发育缺陷[25]。另外,Rho GTP酶激活蛋白REN1可以通过抑制拟南芥花粉管中的Formin homology5(FH5)来控制顶端肌动蛋白聚合[26]。在杨树中,ARF7.2的MR和PB1结构域分别负责其与DELLA或Aux/IAA蛋白的相互作用,进而调节杨树形成层的发育[27]。而MeARF也包含了PB1结构域,因此推测MeARF蛋白可能通过PB1结构域与AUX/IAA8的相互作用。大豆中存在一种具有C端leucine-rich repeat(LRR)结构域的F-BOX蛋白(F-BOX proteins with C-terminal LRR domains,FBXL),其可以显著提高大豆的种子产量、豆荚数、每株种子数、株高[28]。另外,本研究还筛选到了与植物胁迫相关的蛋白,比如多聚泛素4。之前的研究表明,在不同的胁迫下(渗透胁迫、生理盐水胁迫和冷冻胁迫),水稻中的多聚泛素启动子基因存在差异表达[29]。除此之外,筛库还获得了一些目前在植物中研究较少的蛋白,如依赖钠的磷酸盐转运蛋白、溶质载体家族35,它们在植物中的功能以及作用机理还有待进一步研究。
综上所述,本研究通过分析MeARF的蛋白结构域(DBD、MR和PB1结构域)及其在木薯不同组织中的相对表达量,并获得了7个与MeARF相互作用的候选蛋白,可为后续进一步解析MeARF在木薯中的功能和作用机理奠定了理论基础。
木薯生长素响应因子MeARF的克隆及互作蛋白的筛选
DOI: 10.15886/j.cnki.rdswxb.20240148
 CSTR: 32425.14.j.cnki.rdswxb.20240148
CSTR: 32425.14.j.cnki.rdswxb.20240148
Cloning of MeARF and screening of interacting proteins in cassava
-
摘要: 为了解析木薯(Manihot esculenta)中生长素响应因子(Auxin response factor,ARF)的功能和作用机理,本研究首先克隆了一个MeARF基因,然后通过生物信息学分析,构建了进化树明确了其亲缘关系。蛋白结构分析发现,该蛋白含有保守的B3结构域(B3 DNA binding domain,B3)和Phox and Bem1p(PB1)结构域。通过分析MeARF在不同组织的表达量,发现MeARF在木薯的侧芽表达量较高。本研究还构建了pGADT7-MeARF的基因表达载体,然后通过酵母双杂交实验筛选木薯的cDNA文库,最终筛选到了与植物生长发育和免疫相关的候选蛋白。结果表明,MeARF可能参与了木薯的生长发育和免疫。Abstract: An attempt was made to analyze the function and mechanism of auxin response factor (ARF) in cassava. A MeARF gene was cloned from different tissues of cassava and analyzed by using bioinformatics, and an evolutionary tree was constructed to illustrate its homologous relationship. The analysis of the protein structure of MeARF found that the protein contained the conserved B3 domain (B3 DNA binding domain, B3) and PB1 domain (Phox and Bem1p, PB1). The analysis of the expression levels of MeARF in different tissues showed that the expression level of MeARF was relatively high in the lateral buds of cassava. Furthermore, the gene expression vector of pGADT7-MeARF was constructed, and the cDNA library of cassava was screened by yeast two-hybrid experiments. The candidate proteins related to plant growth, development and immunity were screened. The results indicate that MeARF might be involved in growth, development and immunity of cassava.
-
Key words:
- auxin response factor /
- cassava /
- protein structure /
- yeast two-hybrid /
- candidate protein
-
图 2 构建ARF进化树
选取的物种为麻风树、橡胶树、蓖麻、续随子、一年生山靛、美洲黑杨、毛白杨、胡杨、黑杨、银白杨、白时钟花、簸箕柳、乌登柳、尖叶紫柳、小垫柳、长梗柳、紫红柳、哥伦比亚锦葵、黄麻、可可、南欧大戟、龙眼、哥伦比亚古柯、长蒴黄麻、土瓶草、榴莲。
Fig. 2 Construction of the evolutionary tree of ARF
The selected species were Jatropha curcas,Hevea brasiliensis,Ricinus communis,Euphorbia lathyris,Mercurialis annua,Populus deltoids,Populus tomentosa,Populus euphratica,Populus nigra,Populus alba,Turnera subulata,Salix suchowensis,Salix udensis,Salix koriyanagi,Salix brachista,Salix dunnii,Salix purpurea,Herrania umbratica,Corchorus capsularis,Theobroma cacao,Euphorbia peplus,Dimocarpus longan,Erythroxylum novogranatense,Corchorus olitorius,Cephalotus follicularis,Durio zibethinus.
图 3 MeARF蛋白结构分析
A. MeARF保守蛋白结构域分析; B. MeARF蛋白三维结构预测。
Fig. 3 Structural analysis of the MeARF protein
A. Domain analysis of MeARF conserved protein; B.Predicting the 3D structure of MeARF protein.
图 4 MeARF在木薯不同组织的表达量
A. TCOD expression在线数据库中,MeARF在有组织的胚胎发生结构、根尖分生组织、叶片、中脉、叶柄、侧芽、茎、须根、易碎胚性愈伤组织和茎尖分生组织中的表达量; B. MeARF在木薯茎、叶柄、叶片、侧芽和须根中的表达情况; 3个生物学重复,不同的字母表示显著性差异(P < 0.05)。
Fig. 4 Expression profiling of MeARF in different tissues of cassava
A. TCOD expression Online database for the relative expression of MeARF in organized embryogenic structures,root apical meristem,leaf、midvein,petiole,lateral bud,stem,fibrous root,friable embryogenic callus and shoot apical meristem; B. The relative expression of MeARF in stem,petiole,leaf,lateral bud and fibrous root. The figure shows three biological replicates; and different letters indicate significant differences (P < 0.05).
图 5 pGBKT7-MeARF自激活验证
pGBKT7与pGBKT7-MeARF两种组合的酵母在SD-Trp和SD-Trp-Ade-His固体培养基的生长情况,实验采用了4种浓度梯度(1、10−1、10−2、10−3)来进行稀释。
Fig. 5 The self-activation validation of pGBKT7-MeARF
The growth of yeast strains pGBKT7 and pGBKT7 MeARF in SD-Trp and SD-Trp-Ade-His solid media at four concentration gradients (1,10−1,10−2,10−3) for dilution was observed.
图 6 pGADT7-MeARF筛选木薯pGBKT7文库
M1. DL 2 000 DNA Marker; M. DL 15 000 DNA Marker; A. 酵母在SD-Trp-Leu和SD-Trp-Leu-Ade-His固体培养基上的生长情况,以及将SD-Trp-Leu-Ade-His固体培养基上生长的单菌落进行划线保存; B. 通过提取酵母单菌落的质粒,并以酵母质粒为模板,用MeARF的特异性引物扩增得到的阳性克隆; C. 用pGBKT7通用引物扩增得到候选互作基因的条带。
Fig. 6 Screening of the cassava pGBKT7 library by using pGADT7-MeARF
M1. DL 2 000 DNA Marker; M. DL 15 000 DNA Marker; A. Yeast growth on SD-Trp-Leu and SD-Trp-Leu-Ade-His solid media,and single colonies grown on SD-Trp-Leu-Ade-His solid medium; B. Positive clone amplified by MeARF specific primers using the yeast plasmid as a template; C. Bands of candidate interacting genes with pGBKT7 universal primers.
表 1 反转录体系
Table 1 The reverse transcription system
组分
Component体积/μL
Volume /μLRNase-free Water 12 4×Hifair® AdvanceFast One-Step RT SuperMix 5 gDNA Remover Mix 1 mRNA 2  下载: 导出CSV
下载: 导出CSV
表 2 PCR反应程序设置
Table 2 Setup of PCR reaction program
名称
Name温度/℃
Temperature/℃时间/min
Time /min预变性(Initial denaturation) 95 3.0 变性(Degeneration) 95 0.5 退火(Annealing) 55 0.5 延伸(Extension) 72 3.0 循环次数(Number of cycles) 36 最终延伸(Final extension) 72 5.0
下载: 导出CSV
表 3 荧光定量PCR程序
Table 3 The fluorescence quantitative fluorescence PCR procedure
名称
Name温度/℃
Temperature/℃时间/s
Time/s预变性(Initial denaturation) 95 30 变性(Degeneration) 95 15 退火(Annealing) 55 15 延伸(Extension) 72 20 循环次数(Number of cycles) 45
下载: 导出CSV
表 4 引物碱基序列
Table 4 Primer base sequences
引物名称 Primer 碱基序列(5′−3′) Base sequence(5′−3′) QMeARF-F ATGATGATGGGTTCTGTTG QMeARF-R GCTCCGAGTTTATTGTCTT MeARF-AD-F GTACCAGATTACGCTCATATGATGCATTTTAAACAGAAGCACATGA MeARF-AD-R ATGCCCACCCGGGTGGAATTCTCAAGCACGGCTACCTTCTGT pGBKT7-F GTAATACGACTCACTATAGGGCGA pGBKT7-R TTTTCGTTTTAAAACCTAAGAGTC
下载: 导出CSV
表 5 序列比对结果
Table 5 Results of the sequence alignment
基因ID
Gene ID基因注释
Gene annotationE值
E value置信度/%
Confidence level/%Manes.15G163400.1 NADH DEHYDROGENASE [UBIQUINONE] IRON-SULFUR PROTEIN 8-A, MITOCHONDRIAL-RELATED 0 99.77 Manes.14G035900.1 SODIUM-DEPENDENT PHOSPHATE TRANSPORTERS 1.01E-20 100.00 Manes.04G111000.1 POLYUBIQUITIN 4 0 98.23 Manes.02G146100.1 RHO GTPASE-ACTIVATING PROTEIN REN1 2.64E-01 96.00 Manes.05G187100.1 AUXIN-RESPONSIVE PROTEIN IAA8 0 99.54 Manes.17G049400.1 SOLUTE CARRIER FAMILY 35 0 100.00 Manes.05G050000.1 LEUCINE RICH REPEAT PROTEINS, SOME PROTEINS CONTAIN
F-BOX2.65E-02 93.00
下载: 导出CSV
-
[1] 钱双杰. 拟南芥生长素响应因子ARF17调控LTPG4对花粉管伸长影响[D]. 上海: 上海师范大学, 2024. doi: 10.27312/d.cnki.gshsu.2024.001030. [2] 温绍廷. 生长素调控拟南芥不定根再生的分子机制研究[D]. 上海: 上海师范大学, 2021. doi: 10.27312/d.cnki.gshsu.2021.000501. [3] WANG X Y, YU R B, WANG J J, et al. The asymmetric expression of SAUR genes mediated by ARF7/19 promotes the gravitropism and phototropism of plant hypocotyls[J]. Cell Reports, 2020, 31(3): 107529. doi: 10.1016/j.celrep.2020.107529 [4] ZHANG F, TAO W Q, SUN R Q, et al. Correction: PRH1 mediates ARF7-LBD dependent auxin signaling to regulate lateral root development in Arabidopsis thaliana[J]. PLoS Genetics, 2022, 18(3): e1010125. doi: 10.1371/journal.pgen.1010125 [5] 臧鹏跃. 番茄SlARF3对果实大小、可溶性糖以及有机酸影响的初步研究[D]. 沈阳: 沈阳农业大学, 2021. doi: 10.27327/d.cnki.gshnu.2021.000095. [6] 徐富贵. OsARF18调控水稻根系发育和磷素吸收的机制研究[D]. 郑州: 河南农业大学, 2021. doi: 10.27117/d.cnki.ghenu.2021.000173. [7] QIAO J Y, ZHANG Y J, HAN S Q L, et al. OsARF4 regulates leaf inclination via auxin and brassinosteroid pathways in rice[J]. Frontiers in Plant Science, 2022, 13: 979033. doi: 10.3389/fpls.2022.979033 [8] ZHAO Z X, YIN X X, LI S, et al. miR167d-ARFs module regulates flower opening and stigma size in rice[J]. Rice, 2022, 15(1): 40. doi: 10.1186/s12284-022-00587-z [9] ZHANG X F, CAO J F, HUANG C C, et al. Characterization of cotton ARF factors and the role of GhARF2b in fiber development[J]. BMC Genomics, 2021, 22(1): 202. doi: 10.1186/s12864-021-07504-6 [10] ZHANG H H, LI L L, HE Y Q, et al. Distinct modes of manipulation of rice auxin response factor OsARF17 by different plant RNA viruses for infection[J]. Proceedings of the National Academy of Sciences of the United States of America, 2020, 117(16): 9112 − 9121. doi: 10.1073/pnas.1918254117 [11] CARUANA J C, DHAR N, RAINA R. Overexpression of Arabidopsis microRNA167 induces salicylic acid-dependent defense against Pseudomonas syringae through the regulation of its targets ARF6 and ARF8[J]. Plant Direct, 2020, 4(9): e00270. doi: 10.1002/pld3.270 [12] LI J, WU F K, HE Y F, et al. Maize transcription factor ZmARF4 confers phosphorus tolerance by promoting root morphological development[J]. International Journal of Molecular Sciences, 2022, 23(4): 2361. doi: 10.3390/ijms23042361 [13] WANG C F, LI X M, ZHUANG Y B, et al. A novel miR160a-GmARF16-GmMYC2 module determines soybean salt tolerance and adaptation[J]. New Phytologist, 2024, 241(5): 2176 − 2192. doi: 10.1111/nph.19503 [14] 马俊杰, 郭凤丹, 王兴军, 等. 生长素合成、运输和信号转导调控植物胚胎早期发育[J]. 植物生理学报, 2019, 55(5): 547 − 557. doi: 10.13592/j.cnki.ppj.2018.0539 [15] 吴元彩, 王东登, 郑旭阳, 等. 激素和蔗糖对番茄子叶节位侧芽萌发与生长的影响[J]. 南方农业学报, 2024, 55(2): 509 − 519. doi: 10.3969/j.issn.2095-1191.2024.02.021 [16] 庞祥宇. 木薯IAA/ARF基因家族预测、表达及功能分析[D]. 南宁: 广西大学, 2019. [17] LI Y H, HAN S Q L, QI Y. Advances in structure and function of auxin response factor in plants[J]. Journal of Integrative Plant Biology, 2023, 65(3): 617 − 632. doi: 10.1111/jipb.13392 [18] SWAMINATHAN K, PETERSON K, JACK T. The plant B3 superfamily[J]. Trends in Plant Science, 2008, 13(12): 647 − 655. doi: 10.1016/j.tplants.2008.09.006 [19] CANCÉ C, MARTIN-AREVALILLO R, BOUBEKEUR K, et al. Auxin response factors are keys to the many auxin doors[J]. New Phytologist, 2022, 235(2): 402 − 419. doi: 10.1111/nph.18159 [20] FONTANA M, ROOSJEN M, CRESPO GARCÍA I, et al. Cooperative action of separate interaction domains promotes high-affinity DNA binding of Arabidopsis thaliana ARF transcription factors[J]. Proceedings of the National Academy of Sciences of the United States of America, 2023, 120(11): e2219916120. doi: 10.1073/PNAS.2219916120 [21] WANG Y, LI Y, HE S P, et al. The transcription factor ERF108 interacts with AUXIN RESPONSE FACTORs to mediate cotton fiber secondary cell wall biosynthesis[J]. The Plant Cell, 2023, 35(11): 4133 − 4154. doi: 10.1093/plcell/koad214 [22] LIU K, LI Y H, CHEN X N, et al. ERF72 interacts with ARF6 and BZR1 to regulate hypocotyl elongation in Arabidopsis[J]. Journal of Experimental Botany, 2018, 69(16): 3933 − 3947. doi: 10.1093/jxb/ery220 [23] ZHANG T, LI W J, XIE R X, et al. CpARF2 and CpEIL1 interact to mediate auxin-ethylene interaction and regulate fruit ripening in papaya[J]. The Plant Journal, 2020, 103(4): 1318 − 1337. doi: 10.1111/tpj.14803 [24] 李秉贞. 生长素响应因子ARF4通过与IAA12互作调控拟南芥芽再生[D]. 泰安: 山东农业大学, 2020. doi: 10.27277/d.cnki.gsdnu.2020.000946. [25] HSIEH W Y, LIN S C, HSIEH M H. Transformation of nad7 into the nuclear genome rescues the slow growth3 mutant in Arabidopsis[J]. RNA Biology, 2018, 15(11): 1385 − 1391. doi: 10.1080/15476286.2018.1546528 [26] XU Y N, SHEN J F, RUAN H Q, et al. A RhoGAP controls apical actin polymerization by inhibiting formin in Arabidopsis pollen tubes[J]. Current Biology, 2024, 34(21): 5040 − 5053. doi: 10.1016/j.cub.2024.09.053 [27] HU J, SU H L, CAO H, et al. AUXIN RESPONSE FACTOR7 integrates gibberellin and auxin signaling via interactions between DELLA and AUX/IAA proteins to regulate cambial activity in poplar[J]. The Plant Cell, 2022, 34(7): 2688 − 2707. doi: 10.1093/plcell/koac107 [28] HINA A, KHAN N, KONG K K, et al. Exploring the role of FBXL gene family in Soybean: implications for plant height and seed size regulation[J]. Physiologia Plantarum, 2024, 176(1): e14191. doi: 10.1111/ppl.14191 [29] PERALES L, PEÑARRUBIA L, CORNEJO M J. Induction of a polyubiquitin gene promoter by dehydration stresses in transformed rice cells[J]. Journal of Plant Physiology, 2008, 165(2): 159 − 171. doi: 10.1016/j.jplph.2006.12.012 -

 点击查看大图
点击查看大图
计量
- 文章访问数: 1013
- HTML全文浏览量: 362
- PDF下载量: 47
- 被引次数: 0
