



-
木薯(Manihot esculenta)含有丰富的淀粉、矿物质和少量的蛋白质[1],是世界第六大粮食作物[2]。木薯耐贫瘠,但经常遭受各种病害的侵袭[3],如细菌性枯萎病、花叶病、褐斑病等。其中,木薯细菌性枯萎病由地毯草黄单胞菌属木薯萎蔫致病变种(Xanthomonas axonopodis pv. manihotis,Xam)引起,是木薯最严重的世界性病害之一,对木薯产量能造成50%~75%的损失[4]。因此,挖掘木薯候选抗病基因,培育优良木薯抗病品种,是当前需要解决的问题。
为了抵抗病原菌的侵染,植物通过一系列的抗病基因和抗病信号分子产生复杂的免疫防卫反应[5]。例如,植物通过模式触发免疫(pattern-triggered immunity,PTI)[6]及效应触发免疫(effector-triggered immunity,ETI)[7]来识别和对抗发病机制,PTI会触发快速而短暂的活性氧(reactive oxygen species,ROS)爆发,而PTI和ETI的共同激活则会导致二次持续的ROS产生[8]。ROS不仅可以对病原物直接造成损伤,还可以作为信号分子传递信号诱导植物细胞的程序性死亡从而限制病原物[9]。
DAWDLE(DDL)是一类含有保守叉头相关结构域(forkhead-associated domain,FHA)的蛋白,该类蛋白在植物发育和免疫中发挥了重要作用。Feng等[10]利用筛选拟南芥免疫缺陷型突变体的手段以及生化实验方法,鉴定到拟南芥(Arabidopsis thaliana)中DAWDLE蛋白能够通过多聚ADP-核糖化(poly ADP-ribosylation,PARylation)修饰来启动对丁香假单胞菌(Pseudomonas syringae pv. tomato DC3000,Pst DC3000)的免疫反应,且在DDL突变体中,防御基因表达和胼胝质沉积减少,表明DDL在晚期免疫反应中起作用[11]。在拟南芥中DAWDLE能够结合DCL1(Dicer-like 1)和DCL3(Dicer-like 3)后调节DCL酶的活性,从而调节内源small RNA生成[12]。有研究发现在过表达miR394(microRNAs)后拟南芥会对灰孢杆菌(Botrytis cinerea)感染表现出易感性,而在侵染灰孢杆菌前miR394过表达株中DDL的表达水平呈上升状态,但在拟南芥受到灰孢杆菌侵染后DDL的表达水平则受到抑制[13]。DAWDLE除了参与植物免疫防御外还影响其生长发育,前人研究发现在拟南芥中DAWDLE在根和茎的分生组织中表达,DDL突变体导致植株发育迟缓、结实率降低、生长周期延长等多效性生长缺陷[14]。此外,2022年Xiong等[15]发现DAWDLE可以与bud site selection protein 13 (BUD13)、growth development and splicing 1 (GDS1)组成retention and splicing (RES)复合体,通过影响前体RNA剪接调控拟南芥根与胚发育。现有研究表明DAWDLE在模式植物中积极参与植物的生长发育和免疫调节过程,然而其在木薯中抗病功能仍不清晰。
为探究MeDAWDLE蛋白是否参与木薯抗胁迫反应,本研究通过分离鉴定MeDAWDLE,并对其进行了生物信息学分析,通过公共转录组数据分析得到了MeDAWDLE在不同组织中的表达及对木薯细菌性枯萎病致病菌的响应情况,并完成MeDAWDLE-pET28a载体构建和蛋白诱导及部分功能验证,为木薯抗病基因挖掘以及精准分子育种提供一定的理论基础。
-
本研究的试验材料为木薯品种‘华南124号’ (South China 124,SC124),种植于本实验室的种质基地。使用的试剂材料有:2×Taq Master MIX(Dye Plus)(Cat P112-03,诺唯赞生物科技)、2×ChamQ Universal SYBR qPCR Master Mix(Cat Q711-02,诺唯赞生物科技)、西班牙琼脂粉(Cat
162135 ,Biowest)、通用型DNA纯化回收试剂盒(Cat DP214-03,天根生化科技)、RNA提取酚试剂(Cat W0250,Solarbio)、反转录试剂盒(Cat K1622,赛默飞)、β-巯基还原剂(Cat M8210,Solarbio)、异丙基-β-D-硫代吡喃半乳糖苷(IPTG)(Cat I8070-5g,Solarbio)、脱脂奶粉(Cat P0216-300g,碧云天)、His-tag抗体(Cat AH367,碧云天)、生物素标记山羊抗小鼠lgG(H+L)(Cat A0216,碧云天)、DH5α(Cat DL1001S,唯地生物)和BL21 (DE3)感受态细胞(Cat ZC121-2,唯地生物)等。 -
将适量的叶片放入到研钵中,利用液氮将其研磨成粉末后,用CTAB提取液提取RNA,并使用反转录试剂盒将其反转录成cDNA,放入−80 ℃冰箱保存[16]。
-
根据在线数据库中木薯的数据,查找到MeDAWDLE(Phytozome数据库编号:Manes.03G127500)的编码序列(coding sequence,CDS),使用Primer5软件,设计全长引物MeDAWDLE-F (5′-ATGGCCCCTTCCTTGCTCG-3′)和MeDAWDLE-R (5′-TCAGCCTGCTGAATTCTCGTG-3′)。以cDNA为模板,进行MeDAWDLE片段的扩取(表1),扩增程序为:95 ℃ 3 min;34个循环,95 ℃ 30 s,55 ℃ 30 s,72 ℃ 90 s;72 ℃ 10 min。将目的片段进行胶回收并与pEASY-T3连接,随后通过菌落PCR进行验证,选择单一明亮的单克隆,进行测序比对,将测序结果与目的基因序列相同的菌株保存到−40 ℃冰箱。
表 1 PCR体系
Table 1. The volume of PCR reaction
试剂
Reagent体系/μL
System/μL2×Taq Master MIX(Dye Plus) 25.00 dd H2O 20.00 MeDAWDLE-F 1.25 MeDAWDLE-R 1.25 cDNA 2.50 -
从Phytozome数据库(
https://phytozome.jgi.doe.gov/pz/portal.html )中获得的MeDAWDLE蛋白序列和陆地棉(Gossypium hirsutum)、拟南芥、水稻(Oryza sativa)和苹果(Malus pumila)中高同源性的蛋白序列比对,通过MEGA 11.0绘图软件绘制成完整的系统进化树。在ExPASy-ProtParam tool (https://web.expasy.org/protparam/ )网站进行蛋白大小和等电点预测;利用NetPhos 3.1在线预测MeDAWDLE蛋白的磷酸化位点;在TMHMM 2.0分析MeDAWDLE蛋白的跨膜结构;在Signal 5.0对MeDAWDLE蛋白的信号肽进行预测;使用Phytozome中的数据分析其结构域[17]。 -
通过分析TCOD gene search(
https://ngdc.cncb.ac.cn/tcod/genes? )中公共转录组数据得到了MeDAWDLE在木薯不同部位的表达量;通过在线网站(https://vigs.solgenomics.net/ )选取MeDAWDLE特异序列,并根据此序列设计定量引物qMeDAWDLE-F (5′-TTCCTTGCTCGCTGAG-3′)和qMeDAWDLE-R(5′-ATGGGAGATTGTGAATGACT-3′),用Xam侵染多株长势相同的木薯,对照株注射10 mmol·L−1 MgCl2,并于受侵染后0、1、3、6、12、24 h进行叶片取样后,提取RNA并使用反转录试剂盒(Cat K1622,赛默飞)反转录成cDNA,用实时荧光定量PCR(Quantitative Real-time PCR,RT-qPCR)(表2)分析木薯受侵染后MeDAWDLE的表达水平变化,每个样品进行3次重复。表 2 荧光实时定量PCR反应体系
Table 2. The volume of quantitative real-time PCR reaction
试剂
Reagent体系/μL
System/μL2×ChamQ Universal SYBR qPCR Master MIX 7.5 dd H2O 6.4 qMeDAWDLE-F 0.3 qMeDAWDLE-R 0.3 cDNA 0.5 -
挑选构建成功的MeDAWDLE-pEASY-T3菌株质粒作为模板,用PCR扩取(引物:MeDAWDLE-pET28a-F (5′-TTGGGGATGGGTCGCGGATCCATGGCCCCTTCCTTGCTCG-3′)和MeDAWDLE-pET28a-R (5′-ACGGAGCTCGAATTCGGATCCGCCTGCTGAATTCTCGTGCA-3′))目的片段(表1),扩增程序为:95 ℃ 3 min;34个循环,95 ℃ 30 s,55 ℃ 30 s,72 ℃ 90 s;72 ℃ 10 min。采用同源连接将MeDAWDLE片段连接到pET28a载体上,并进行大肠杆菌转化,通过菌落PCR验证,以及后续提质粒,酶切验证[18]。将成功构建的重组质粒转入DE3菌株中,37 ℃摇床进行培养,待OD600值达到0.6,按照体积1∶
1 000 的比例加入1 mol·L−1 IPTG,并于0、3、6 h时取样菌体各2 mL,低温保存,超声破碎后4 ℃离心,取上清进行检测。 -
将37 ℃条件下诱导的菌体超声破碎,离心吸取上清100 μL,向其加入10 μL上样缓冲液后涡旋混匀,沸水浴5 min后,将每个样品等量上样至蛋白胶孔中,然后开始电泳。电泳结束后,将其中一块蛋白胶进行考马斯亮蓝染色,另一块胶放置于PVDF (polyvinylidene fluoride)膜上进行转膜,用1×TBS (tris buffered saline)溶液清洗膜,先倒入牛奶进行孵育后,加入10 mL的一抗(His 抗体)孵育2 h后,更换二抗进行孵育1 h,最后用A、B显影液进行显影拍照[19]。
-
取长势一致的木薯,分别用30、300 μg·mL−1质量浓度的pET28a和MeDAWDLE-pET28a蛋白,并添加体积分数0.5%的表面活性剂(Silwet L-77)外源喷施处理木薯叶片3 d,随后使用圆形打孔器取叶圆,并将背面划伤后朝下放置在水面上过夜;次日将叶圆分别放置在黑色96孔酶标板中,配置反应液[1%的Flg22+2%的鲁米诺+2%的辣根过氧化物酶(horseradish peroxidase,HRP)]。将反应液加入放有叶片的酶标板后迅速放入酶标仪中自动检测数据,检测45次,持续时间30 min[20]。
-
外源蛋白喷施木薯叶片3 d后,于第4 天对木薯叶片背面进行接种实验。事先将实验室保存的Xam菌体在无抗LB平板上划线活化,挑取单菌落加入到无抗LB培养基中培养,至OD600值为0.6时离心,
4000 r·min−1,10 min。并用1 mol·L−1MgCl2重悬,测OD600值为1.0时,用1 mL注射器将菌液侵染至木薯叶片背面。 -
在完成Xam侵染后,对木薯叶片侵染部位进行0 d取样,每个样品取4个接种位置(面积约1 cm2),样品使用体积分数75%的乙醇消杀5 min,随后用灭菌水清洗两次后,将叶片充分研磨,用灭菌水进行10倍浓度的梯度稀释,将样品滴定到无抗LB平板上,每个样品至少重复3次。封板后于28 ℃摇床中倒置培养24 h。并统计菌落数以确保病原菌接种程序一致,后续于接种后4 d取样,方法同上。
-
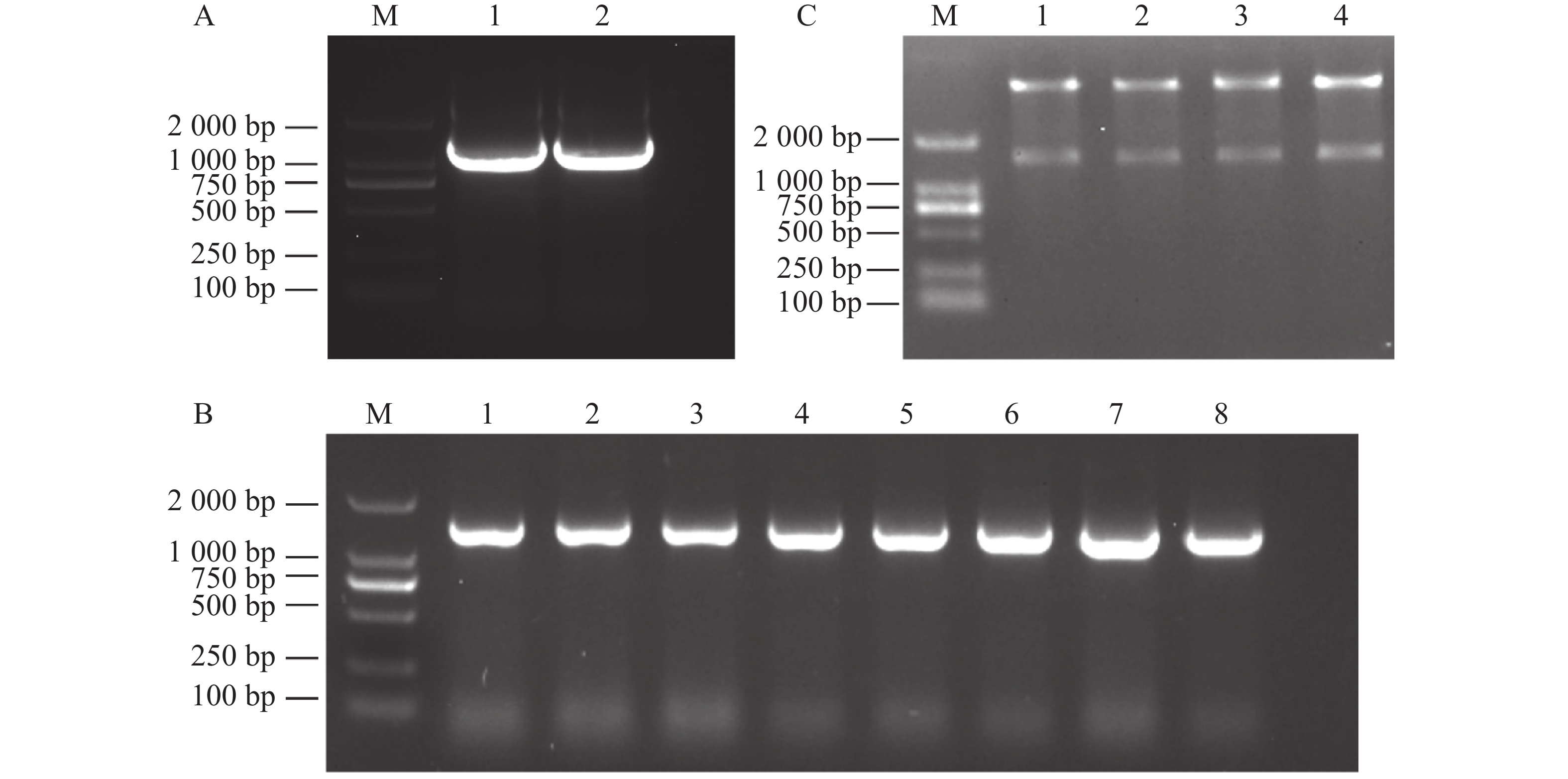
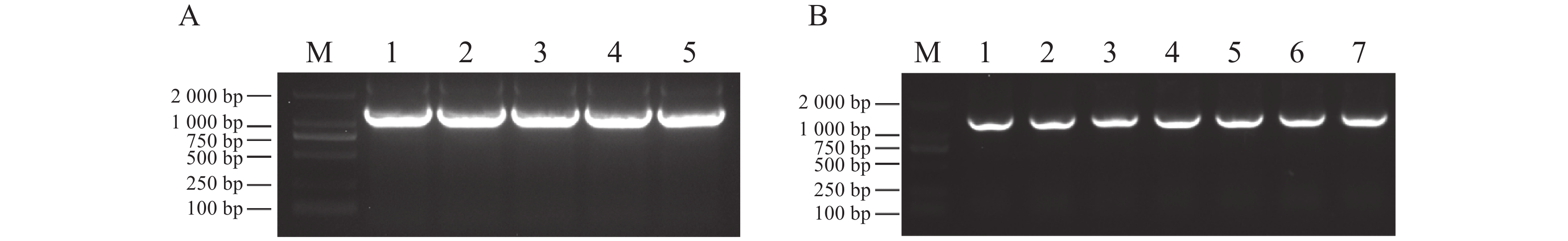
提取木薯叶片RNA并反转录,通过PCR扩增获得目的片段(图1−A)。然后将回收的片段连接到pEASY-T3载体后,转入DH5α菌株中,涂板后挑选单菌落进行PCR验证,得到了7个阳性克隆(图1−B),摇菌提取质粒后送测序。
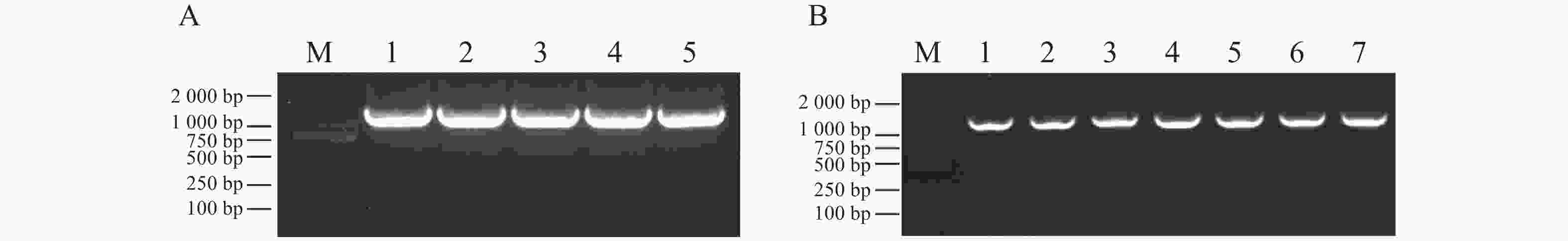
图 1 MeDAWDLE克隆
Figure 1. MeDAWDLE clone
-
将所得测序结果输入NCBI中比对发现该基因为木薯中的DAWDLE,CDS序列为
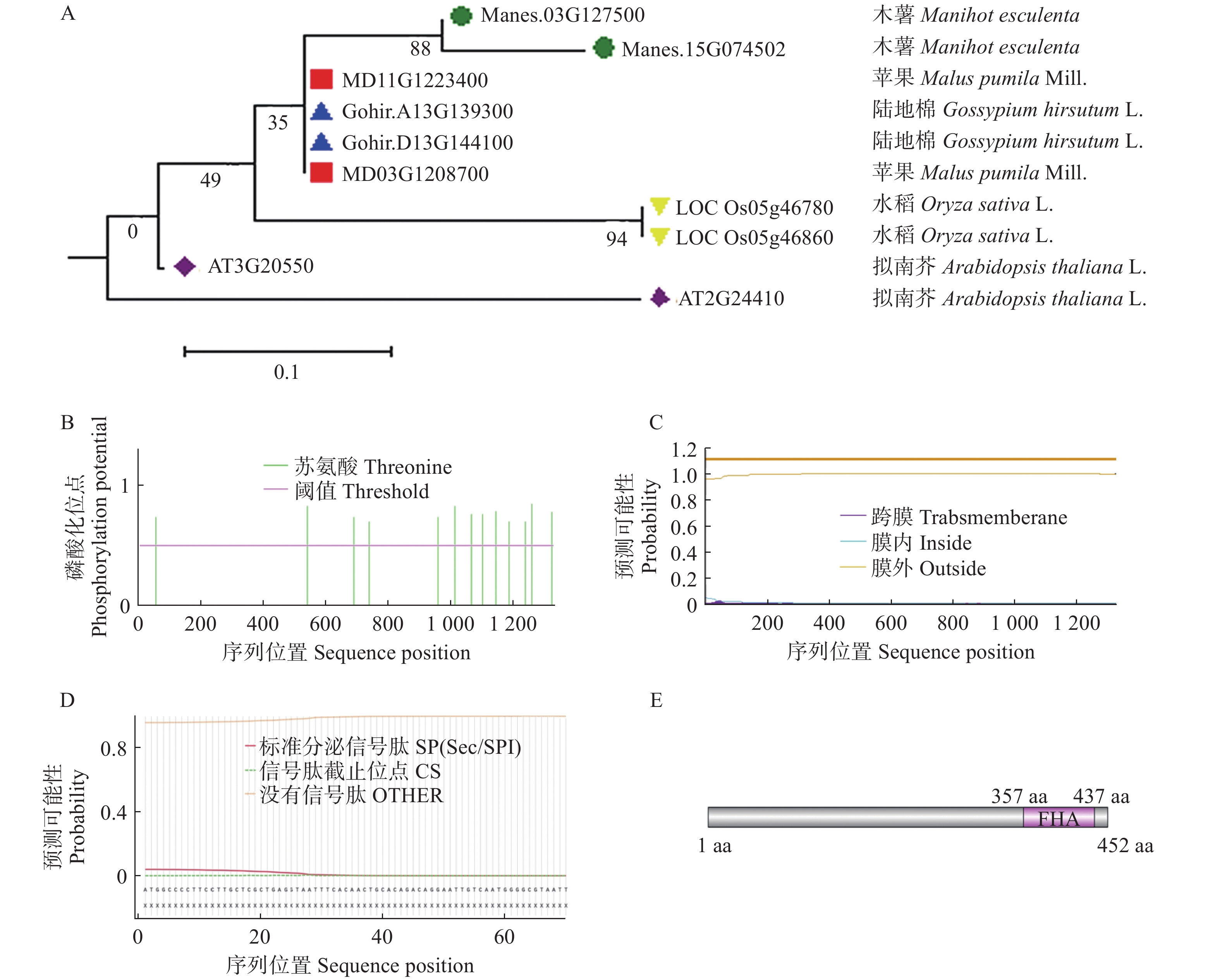
1359 bp,将目标蛋白序列进行同源blast比对,并通过MEGA 11.0构建了木薯、陆地棉、拟南芥、水稻和苹果中的DAWDLE进化树,结果显示木薯中的DAWDLE与苹果中的DAWDLE蛋白同源性较高,同源率为76.3% (图2−A)。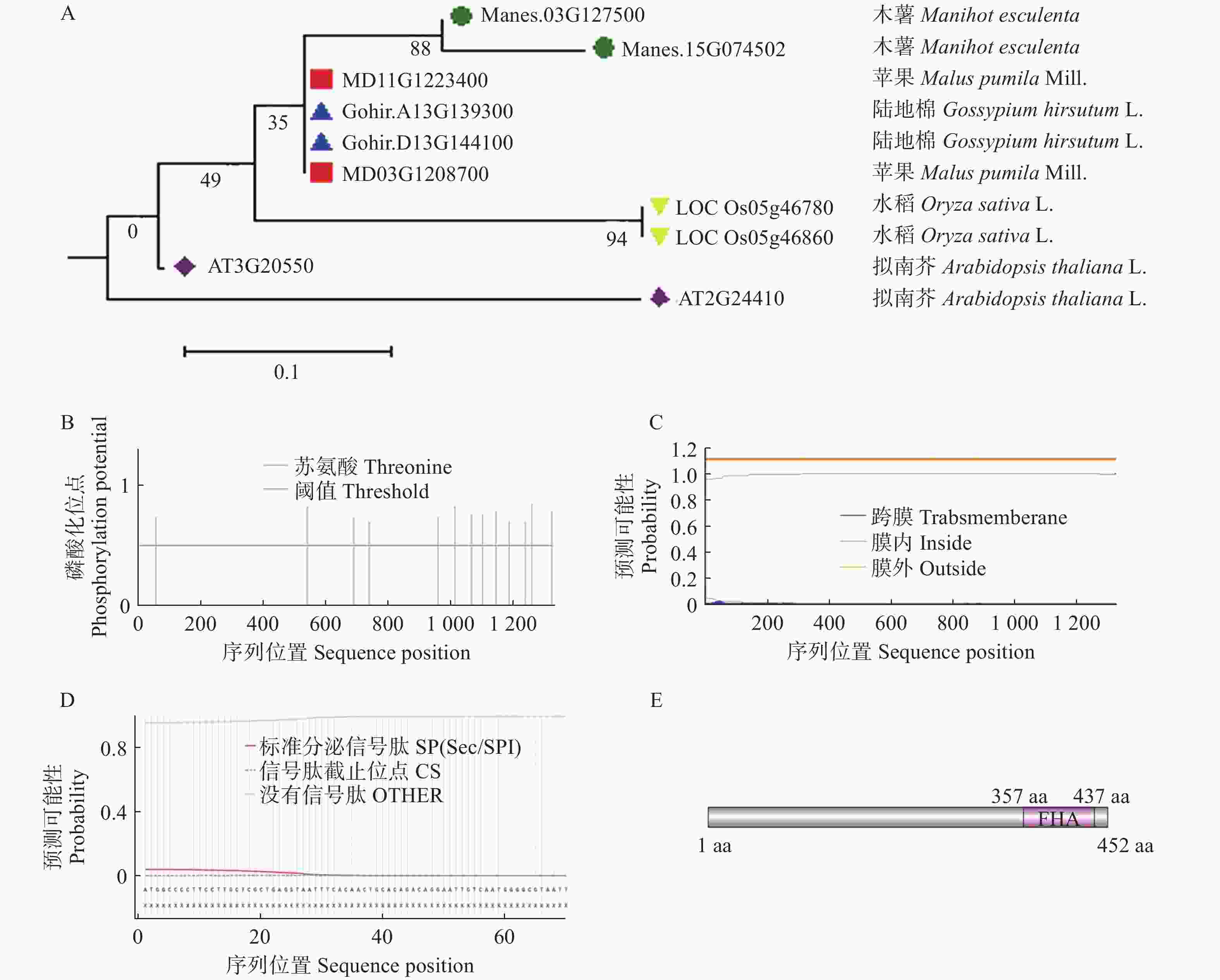
图 2 MeDAWDLE生物信息学分析
Figure 2. Bioinformatics analysis ofMeDAWDLE
-
通过ProParam在线预测了MeDAWDLE的蛋白分子式为C4009H6661N1359O1647S259,蛋白分子质量为52 kDa,理论等电点(isoelectric point, PI)为5.04,不稳定系数为39.86,为稳定蛋白,亲水总平均数为0.831,表明该蛋白属于疏水蛋白。用NetPhos网站在线预测,发现MeDAWDLE的蛋白可能存在磷酸化位点(图2−B);使用TMHMM网站发现,其不存在跨膜结构域(图2−C);通过SignalIP在线网站预测其无信号肽(图2−D);使用Phytozome对其结构进行分析,发现其氨基酸序列在357~437 aa存在FHA结构域(图2−E),FHA是叉头相关结构域,其在细胞内发挥着重要的调控作用[21]。
-
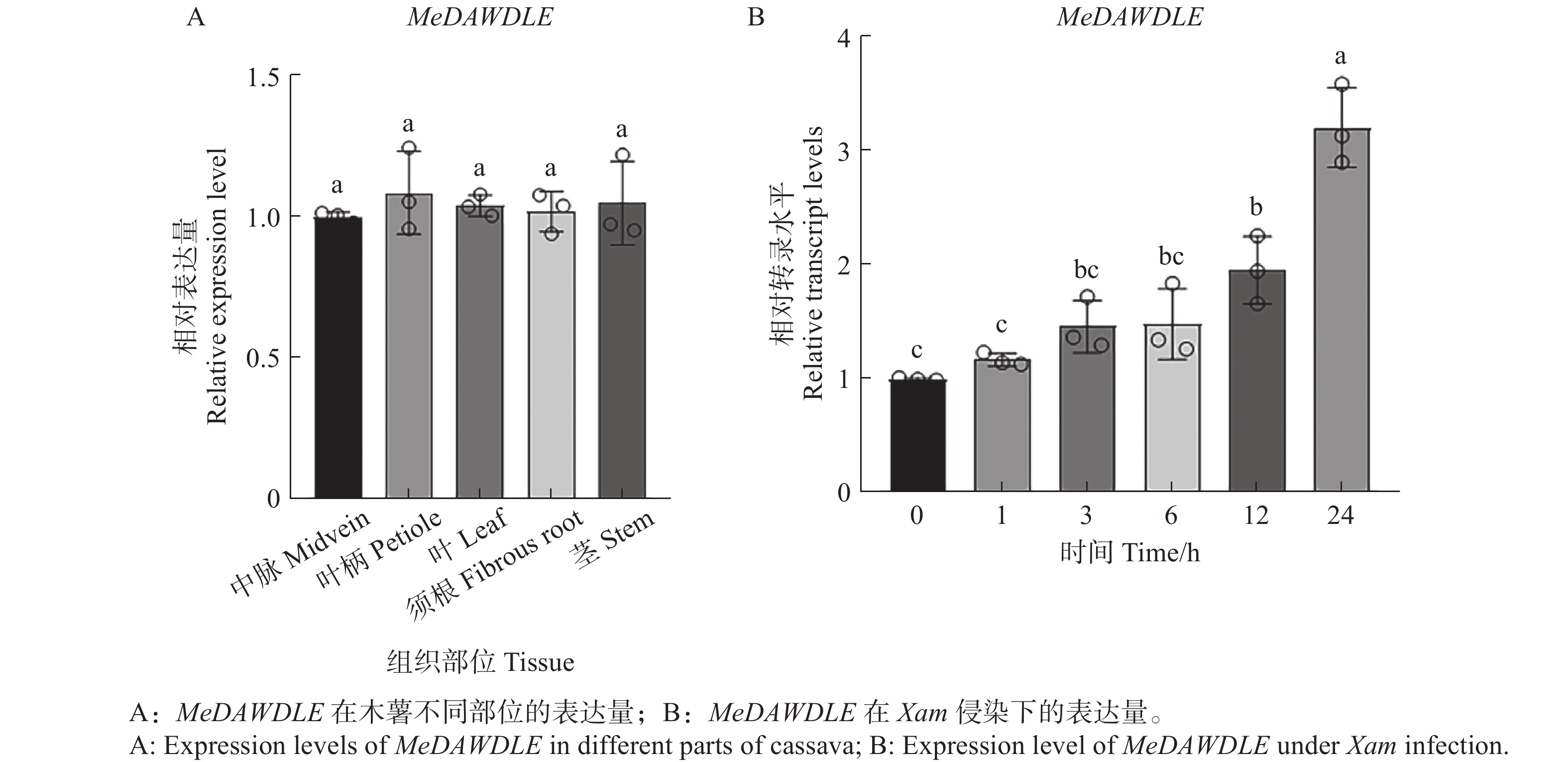
通过分析TCOD gene search中公共转录组数据得到其在木薯不同部位的表达量以及Xam侵染木薯后基因的表达数据,发现MeDAWDLE在木薯不同组织中均有分布(图3−A);为了探究MeDAWDLE在木薯叶片被Xam侵染后表达量是否发生变化,分别选取接种后0、1、3、6、12、24 h的木薯叶片,通过RT-qPCR检测MeDAWDLE的表达量变化。结果表明MeDAWDLE的表达量呈上升趋势,即MeDAWDLE可能在木薯抗病过程中发挥一定的作用(图3−B)。
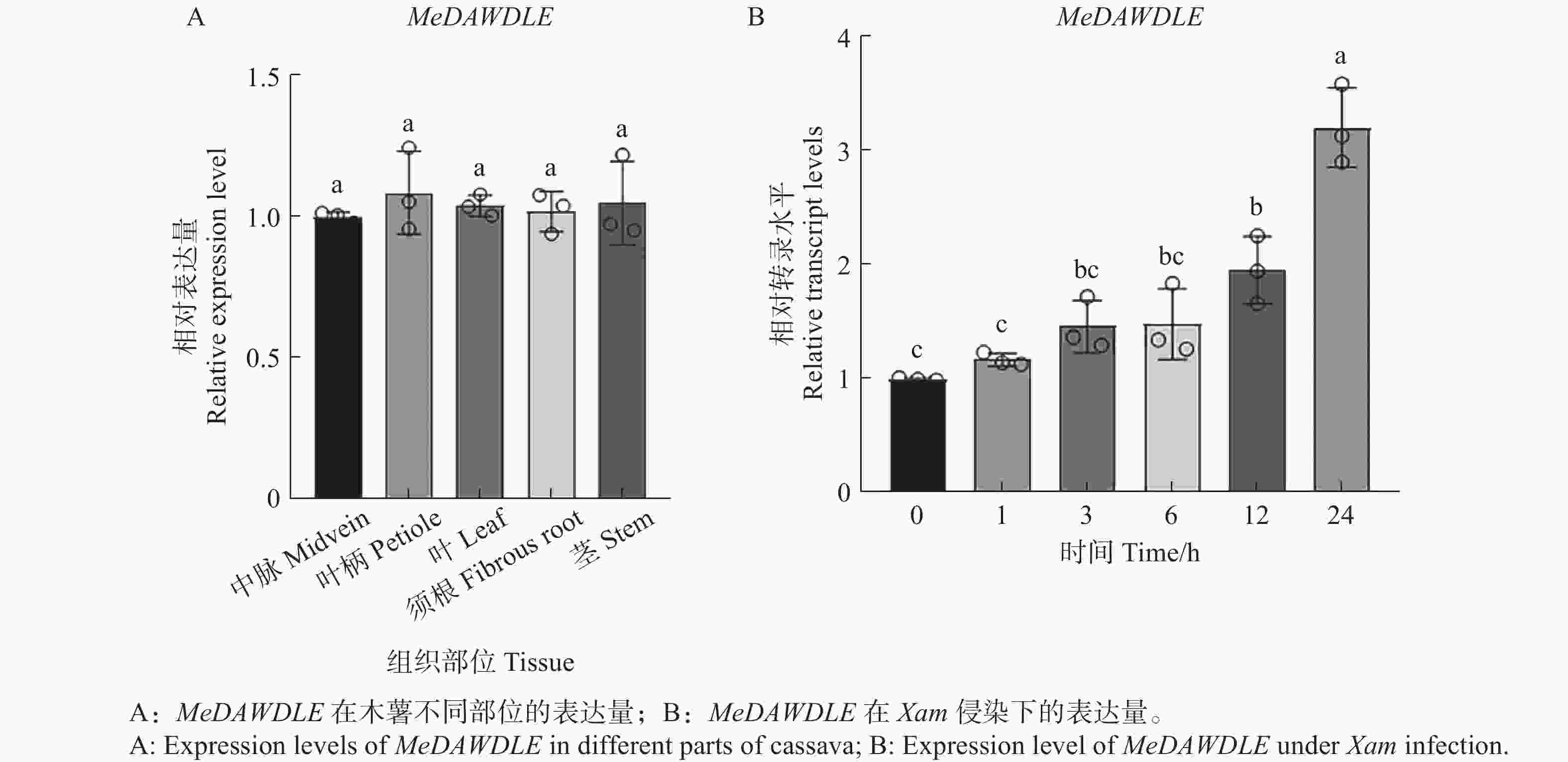
图 3 MeDAWDLE的表达模式分析
Figure 3. Expression levels of MeDAWDLE in different parts of cassava
-
挑选构建成功的MeDAWDLE-pEASY-T3菌株质粒作为模板,通过PCR扩增获得目的片段(图4−A)。使用同源连接将MeDAWDLE片段连接到pET28a载体上,涂板后挑选单菌落进行PCR验证(图4−B),挑选阳性克隆摇菌提质粒。用BamHⅠ酶切验证,在酶切预测大小(
1359 bp)出现目的条带,其中4个克隆均能切出目的条带(图4−C),得到4个原核表达载体的单克隆。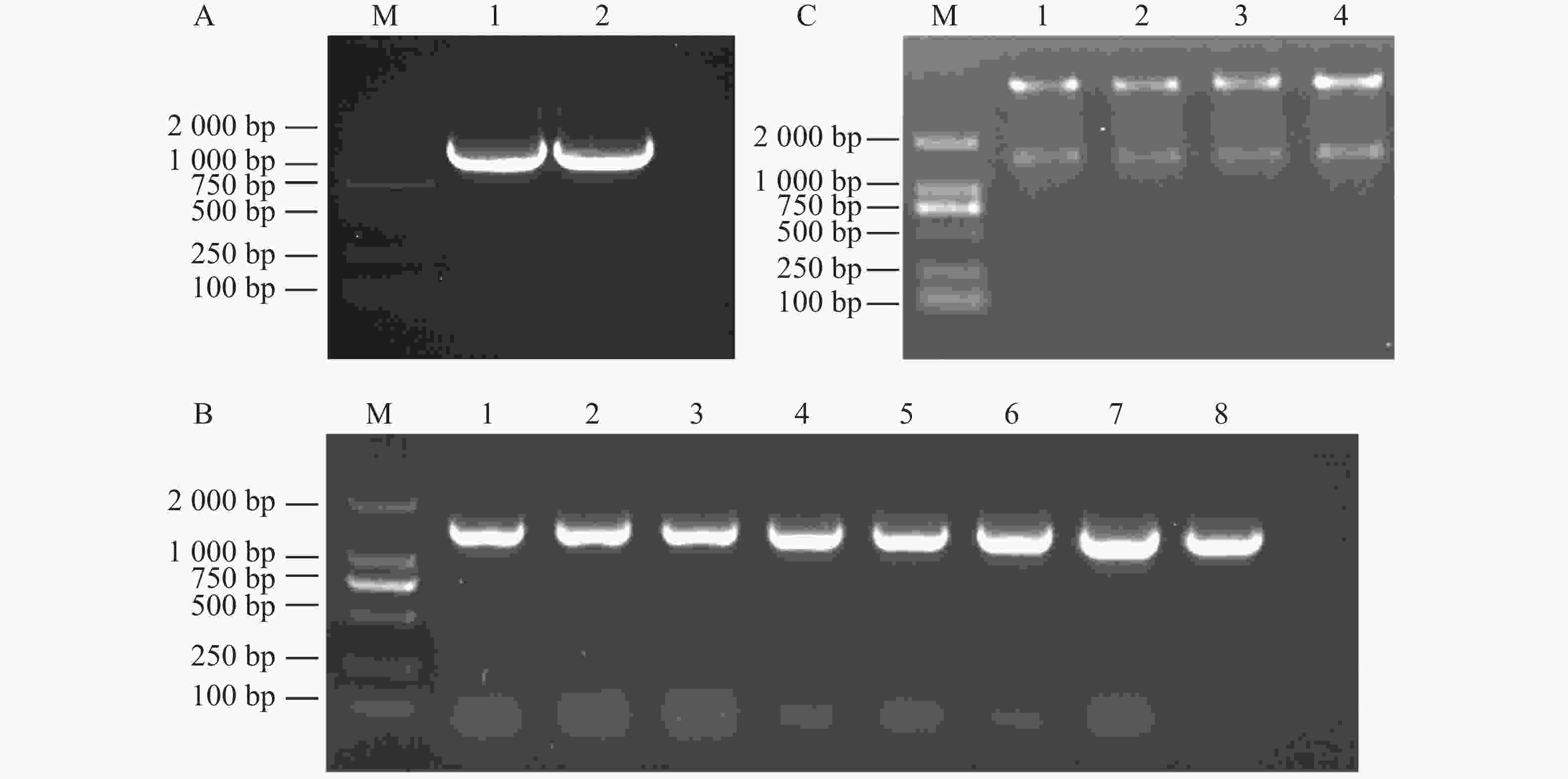
图 4 MeDAWDLE-pET28a载体构建
Figure 4. Construction ofMeDAWDLE-pET28a vector
-
将MeDAWDLE-pET28a重组质粒转入到DE3细胞感受态后进行蛋白诱导。先将菌液37 ℃培养直至OD600值达到0.6,然后按照体积1∶
1 000 的比例加入1 mol·L−1 IPTG,37 ℃诱导,并将诱导0、3、6 h所得的蛋白取样检测,其中一部分用于考马斯亮蓝染色(图5−A),另一部分用于Western blot验证(图5−B)。结果显示,随着时间的推移MeDAWDLE的蛋白诱导量越来越高,在6 h的蛋白浓度为2.2 mg·mL−1,根据考马斯亮蓝染色(图5−A)和Western blot(图5−B)结果可以确定蛋白诱导成功,且依据蛋白大小预测结果,再加上载体上融合的氨基酸序列,可确定上面条带是目的条带。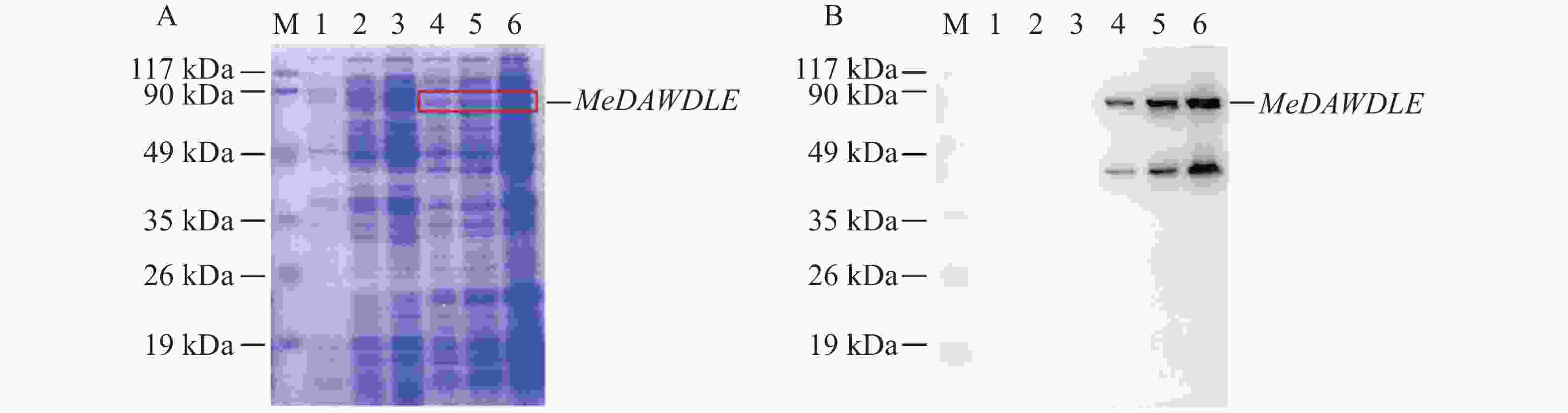
图 5 MeDAWDLE蛋白诱导表达
Figure 5. Induced expression of the MeDAWDLE protein
-
为了明确MeDAWDLE蛋白的功能,本研究使用两个浓度梯度:30 和300 μg·mL−1的MeDAWDLE-pET28a,加上pET28a蛋白,喷施木薯叶片3 d后进行ROS检测。试验结果显示,与对照相比,喷施MeDAWDLE-pET28a蛋白后,木薯植株由flg22诱导的ROS爆发更为剧烈,且喷施300 μg·mL−1蛋白处理组的ROS爆发水平显著高于喷施30 μg·mL−1的处理组(图6−A);此外,在完成外源蛋白喷施处理后,在木薯叶片上侵染Xam,在对侵染后0 d和4 d的叶片细菌数进行定量分析发现,与对照相比喷施MeDAWDLE-pET28a蛋白后的木薯细菌数更少,且300 μg·mL−1的蛋白浓度比30 μg·mL−1处理后抗病效果更好(图6−B),结果说明MeDAWDLE可能正调控了木薯对细菌性枯萎病的抗性。
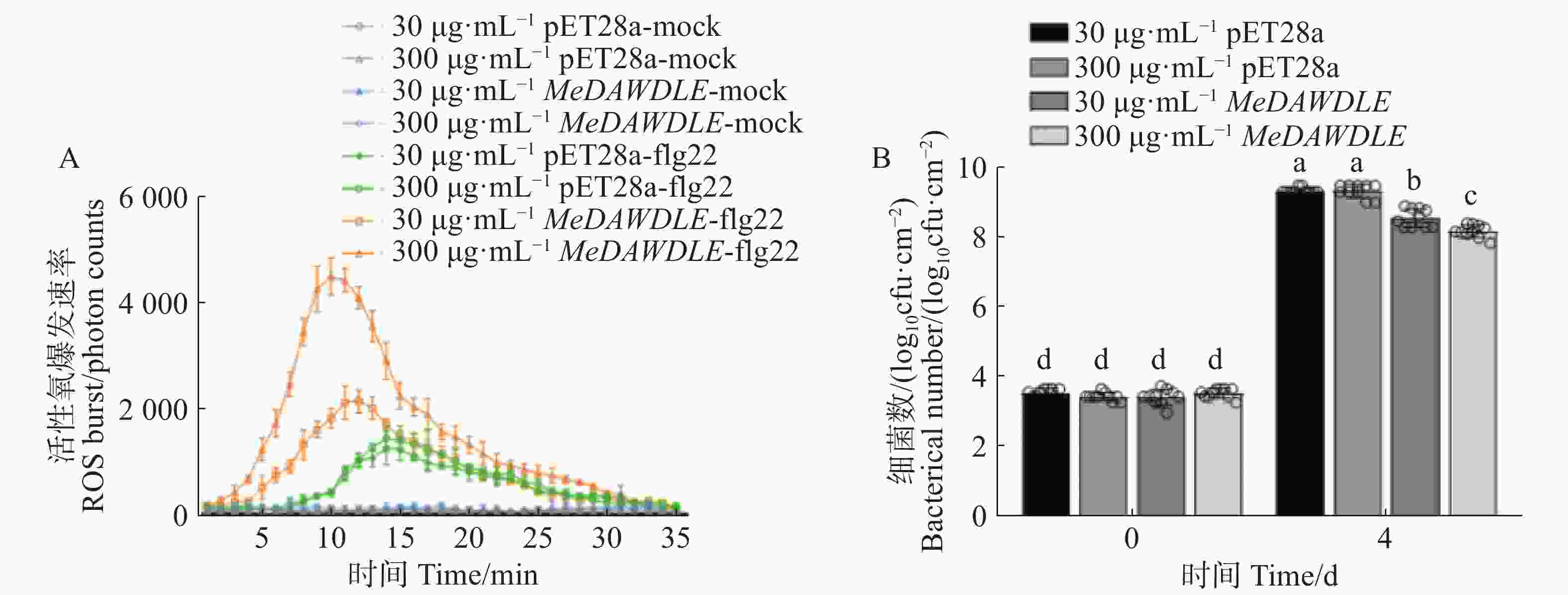
图 6 MeDAWDLE蛋白功能验证
Figure 6. Functional validation ofMeDAWDLE protein
-
DAWDLE 是一类含有保守FHA结构域的蛋白,在植物中FHA 结构域作为一种磷酸肽识别结构域,其结构蛋白通过独特的结构域和相互作用模式在细胞内发挥着重要的调控作用,确保细胞的正常生理功能和对外界环境的适应能力[22]。在拟南芥中FHA结构域蛋白激酶相关蛋白磷酸酶(kinase-associated protein phosphatase,KAPP)通过FHA结构域与多种受体样激酶(receptor-like kinases,RLKs)相互作用,如:BRI1(brassinosteroid insensitive 1)、BAK1(BRI1-associated kinase 1)和FLS2(Flagellin sens 2),使RLKs去磷酸化,导致RLKs激酶的活性降低和信号减弱,从而调控植物生长和响应病原体等多种信号通路[23];在拟南芥中DDL蛋白通过FHA结构域结合DCL1和DCL3后调节两者酶活性来影响拟南芥体内的small RNA生成[12],在木薯中,通过结构与分析发现MeDAWDLE氨基酸序列在357 ~ 437 aa的位置上含有FHA结构域,且与苹果中的DAWDLE蛋白同源性为76.3%,由此推测,FHA可能介导DDL与其他蛋白的互作。此外还有研究报道发现DAWDLE蛋白可以与AtBUD13、GDS1组成一个RES复合体,调控拟南芥根与胚的发育[15];并且其蛋白的多聚ADP-核糖化修饰能够调控拟南芥对Pst DC3000的免疫反应[11]。但在木薯中DDL是否通过FHA结构域与其他蛋白互作以及是否存在类似的生物学功能还有待进一步研究。
本研究通过分离鉴定了MeDAWDLE并明确了其在Xam侵染下的表达差异。通过木薯在胁迫下一些基因的差异性表达,进而可知这些基因的表达差异对于理解其适应环境和抵抗胁迫的分子机制是很重要的。如木薯中有多个CC类谷氧还蛋白(glutaredoxins,GRXs)在叶片中的表达受干旱胁迫的诱导,其中MeGRXC3通过调控过氧化氢酶的活性促使木薯应对干旱胁迫[24];并且在木薯中验证了SR(arginine/serine-rich)蛋白在植物应对盐胁迫反应中的关键作用,且其中MeRS40的转录水平在盐胁迫下被明显诱导[25];除此之外,在木薯受到Xam侵染后,MebHLH149在侵染早期表达水平显著上调,通过参与关键信号级联增强了植物对病原体的反应能力[26]。即MeDAWDLE在木薯受Xam侵染后会被显著诱导表达,表明其可能参与到木薯的免疫调控反应过程。
本研究诱导表达MeDAWDLE-pET28a蛋白,并对其蛋白功能进行初步验证,发现MeDAWDLE可能正调控了木薯对细菌性枯萎病的抗性。ROS是植物激活免疫反应的关键信号[27],在小麦研究中发现来源于黄花叶病毒(wheat yellow mosaicvirus,WYMV)的VsiRNA(virus-derivedsmall interference RNA)通过调控ROS信号激活植物免疫反应[28];在拟南芥中研究发现PRR( pattern recognition receptor)激活的蛋白激酶BIK1(botrytis-induced kinase 1)和MPK4(mitogen-activated protein kinase 4)对脂质激酶DGK5(diacylglycerol kinase 5)的差异磷酸化反向调节磷脂酸(phosphatidic acid,PA)爆发,从而调节ROS的产生影响植物免疫[29]。本研究发现喷施MeDAWDLE蛋白能诱导ROS的爆发,进而表明MeDAWDLE可能参与调控木薯的抗逆性。小麦条锈菌 (Puccinia striiformis f. sp. tritici) 效应蛋白Hasp98能够与小麦抗病正调控因子TaMAPK4(mitogen-activated protein kinase 4)互作,并抑制TaMAPK4激酶活性,调控寄主免疫促进条锈菌侵染[30]。本研究发现对于喷施MeDAWDLE蛋白后再侵染Xam的木薯,其细菌数明显低于对照组,这说明外源喷施MeDAWDLE蛋白增强了木薯的抗病性。
综上所述,本研究通过部分生物信息学分析、构建表达载体完成蛋白诱导以及后续的蛋白功能验证,揭示了MeDAWDLE对木薯活性氧水平的积极调控并初步确定其参与木薯免疫应答过程中,能够为木薯的抗逆性基因研究提供新的参考。
MeDAWDLE基因的克隆和蛋白功能验证
DOI: 10.15886/j.cnki.rdswxb.20240146
 CSTR: 32425.14.j.cnki.rdswxb.20240146
CSTR: 32425.14.j.cnki.rdswxb.20240146
Cloning and protein functional validation of MeDAWDLE gene
-
摘要: 为探究MeDAWDLE蛋白功能,以木薯‘华南124号’为材料,通过PCR扩增对MeDAWDLE进行分离和鉴定。分析进化树发现,MeDAWDLE蛋白与苹果中的DAWDLE蛋白具有76.3%的同源性。蛋白序列的理化性质分析显示,MeDAWDLE蛋白大小为52 kDa,理论等电点为5.04,表达分析发现MeDAWDLE在木薯不同组织器官中均有表达,并在胁迫条件下表达上调。通过构建表达载体完成蛋白诱导,并在进一步的蛋白功能分析中发现,MeDAWDLE能显著提升木薯的活性氧(reactive oxygen species,ROS)水平及抗病能力,表明MeDAWDLE可能在木薯的胁迫响应中发挥着重要作用。Abstract: In order to investigate the protein function of MeDAWDLE, Cassava SC 124 was used as a variety of cassava to isolate and identify its MeDAWDLE through PCR amplification. The evolutionary tree revealed that MeDAWDLE shares high homology with the DAWDLE protein in apples at a homology rate of 76.3%. Analysis of the physicochemical properties of MeDAWDLE sequence revealed that the molecular weight of MeDAWDLE is 52 kDa and that the theoretical isoelectric point of MeDAWDLE is 5.04. Expression analysis showed that MeDAWDLE is distributed in different tissues and organs of cassava, and the transcript of MeDAWDLE is upregulated under stress conditions. To explore the protein function of MeDAWDLE, the expression vector was constructed and the protein was induced. Further protein function analysis revealed that MeDAWDLE can significantly increase the reactive oxygen species (ROS) levels and the disease resistance of cassava. All these results indicated that MeDAWDLEmight play an important role in cassava stress response.
-
图 2 MeDAWDLE生物信息学分析
A:DAWDLE进化树分析(目的基因与陆地棉、拟南芥、水稻和苹果的进化树分析);B:磷酸化位点预测; C:跨膜结构域预测; D:信号肽预测; E:蛋白保守结构域分析。
Fig. 2 Bioinformatics analysis ofMeDAWDLE
A: The evolutionary tree analysis of DAWDLE( Evolutionary tree analysis of the target gene (in green) and those of Gossypium hirsutum (in blue), Arabidopsis thaliana (in purple), Oryza sativa (in yellow), and Malus pumila (in red)); B: Phosphorylation site prediction; C: Prediction of transmembrane domains; D: Signal peptide prediction; E: Analysis of conserved protein domains.
图 3 MeDAWDLE的表达模式分析
不同小写字母表示差异显著(P<0.05,SPSS 26邓肯单因素多样本差异显著性方差分析)。
Fig. 3 Expression levels of MeDAWDLE in different parts of cassava
Different letters indicate significant differences (P<0.05, SPSS 26 Duncan one-way multiple sample analysis of variance).
图 4 MeDAWDLE-pET28a载体构建
M:DL2000 DNA Marker;A:MeDAWDLE扩增,1~2是以T载作为模板扩增的MeDAWDLE;B:同源重组后菌落PCR,1~8表示PCR检验;C:同源重组后验证,1~4表示酶切4个MeDAWDLE-pET28a载体。
Fig. 4 Construction ofMeDAWDLE-pET28a vector
M:DL2000 DNA Marker; A:MeDAWDLE amplification, 1-2:MeDAWDLE amplified using T-carrier as a template; B:Colony PCR after homologous recombination,1-8: PCR testing; C:Verification after homologous recombination,1-4: Enzyme digestion of 4 MeDAWDLE-pET28a vectors.
图 5 MeDAWDLE蛋白诱导表达
M:预染蛋白质分子质量标准(19~117 kDa); A:考马斯亮蓝染色检验,从左到右依次为对照蛋白的0、3、6 h 的上清液,MeDAWDLE-pET28a蛋白0、3、6 h的上清液;B:Westren blot检验,从左到右依次为对照蛋白的0、3、6 h的上清,MeDAWDLE-pET28a蛋白0、3、6 h的上清液。
Fig. 5 Induced expression of the MeDAWDLE protein
M:Pre-stained protein markers (19-117 kDa); A:Coomassie brilliant blue staining test, From left to right, the supernatants of control protein at 0, 3, 6 h, and the supernatants and precipitation ofMeDAWDLE-pET28a protein at 0, 3, 6 h; B:Western blot test, From left to right, the supernatants of control protein at 0, 3, 6 h, and the supernatants and precipitation of MeDAWDLE-pET28a protein at 0, 3, 6 h.
图 6 MeDAWDLE蛋白功能验证
A:外源喷施MeDAWDLE蛋白对木薯ROS水平的影响;B:细菌数。
Fig. 6 Functional validation ofMeDAWDLE protein
A:The effect of exogenous MeDAWDLE protein sprayed on ROS levels; B:Bacterial count.
表 1 PCR体系
Table 1 The volume of PCR reaction
试剂
Reagent体系/μL
System/μL2×Taq Master MIX(Dye Plus) 25.00 dd H2O 20.00 MeDAWDLE-F 1.25 MeDAWDLE-R 1.25 cDNA 2.50  下载: 导出CSV
下载: 导出CSV
表 2 荧光实时定量PCR反应体系
Table 2 The volume of quantitative real-time PCR reaction
试剂
Reagent体系/μL
System/μL2×ChamQ Universal SYBR qPCR Master MIX 7.5 dd H2O 6.4 qMeDAWDLE-F 0.3 qMeDAWDLE-R 0.3 cDNA 0.5
下载: 导出CSV
-
[1] LIN Z J D, TAYLOR N J, BART R. Engineering disease-resistant cassava[J]. Cold Spring Harbor Perspectives in Biology, 2019, 11(11): a034595. doi: 10.1101/cshperspect.a034595 [2] ZÁRATE-CHAVES C A, DE LA CRUZ D G, VERDIER V, et al. Cassava diseases caused by Xanthomonas phaseoli pv. manihotis and Xanthomonas cassavae[J]. Molecular Plant Pathology, 2021, 22(12): 1520 − 1537. doi: 10.1111/mpp.13094 [3] 时涛, 李超萍, 王国芬, 等. 中国木薯病害研究进展与展望[J]. 热带作物学报, 2023, 44(12): 2355 − 2368. doi: 10.3969/j.issn.1000-2561.2023.12.001 [4] YOODEE S, KOBAYASHI Y, SONGNUAN W, et al. Phytohormone priming elevates the accumulation of defense-related gene transcripts and enhances bacterial blight disease resistance in cassava[J]. Plant Physiology and Biochemistry, 2018, 122: 65 − 77. doi: 10.1016/j.plaphy.2017.11.016 [5] 崔亚宁, 钱虹萍, 赵艳霞, 等. 模式识别受体的胞内转运及其在植物免疫中的作用[J]. 植物学报, 2020, 55(3): 329 − 339. doi: 10.11983/CBB19139 [6] CHANG M, CHEN H, LIU F Q, et al. PTI and ETI: convergent pathways with diverse elicitors[J]. Trends in Plant Science, 2022, 27(2): 113 − 115. doi: 10.1016/j.tplants.2021.11.013 [7] HERMANS D, VAN BEERS L, BROUX B. Nectin family ligands trigger immune effector functions in health and autoimmunity[J]. Biology, 2023, 12(3): 452. doi: 10.3390/biology12030452 [8] WANG D, YUAN M H, ZHUANG Y M, et al. DGK5-mediated phosphatidic acid homeostasis interplays with reactive oxygen species in plant immune signaling[J]. Journal of Integrative Plant Biology, 2024, 66(7): 1263 − 1265. doi: 10.1111/jipb.13683 [9] YU X Q, NIU H Q, LIU C, et al. PTI-ETI synergistic signal mechanisms in plant immunity[J]. Plant Biotechnology Journal, 2024, 22(8): 2113 − 2128. doi: 10.1111/pbi.14332 [10] FENG B M, MA S S, CHEN S X, et al. PARylation of the forkhead-associated domain protein DAWDLE regulates plant immunity[J]. EMBO Reports, 2016, 17(12): 1799 − 1813. doi: 10.15252/embr.201642486 [11] BIGEARD J, COLCOMBET J, HIRT H. Signaling mechanisms in pattern-triggered immunity (PTI)[J]. Molecular Plant, 2015, 8(4): 521 − 539. doi: 10.1016/j.molp.2014.12.022 [12] ZHANG S X, DOU Y C, LI S J, et al. DAWDLE interacts with DICER-LIKE proteins to mediate small RNA biogenesis[J]. Plant Physiology, 2018, 177(3): 1142 − 1151. doi: 10.1104/pp.18.00354 [13] TIAN X, SONG L P, WANG Y, et al. miR394 Acts as a negative regulator of Arabidopsis resistance to B. cinerea infection by targeting LCR[J]. Frontiers in Plant Science, 2018, 9: 903. doi: 10.3389/fpls.2018.00903 [14] MORRIS E R, CHEVALIER D, WALKER J C. DAWDLE, a forkhead-associated domain gene, regulates multiple aspects of plant development[J]. Plant Physiology, 2006, 141(3): 932 − 941. doi: 10.1104/pp.106.076893 [15] XIONG F, REN J J, WANG Y Y, et al. An Arabidopsis retention and splicing complex regulates root and embryo development through pre-mRNA splicing[J]. Plant Physiology, 2022, 190(1): 621 − 639. doi: 10.1093/plphys/kiac256 [16] PARK S J, CHOI S W, KIM G M, et al. Light-stabilized FHA2 suppresses miRNA biogenesis through interactions with DCL1 and HYL1[J]. Molecular Plant, 2021, 14(4): 647 − 663. doi: 10.1016/j.molp.2021.01.020 [17] BROOKS III L, HEIMSATH E G, JR, LORING G L, et al. FHA-RING ubiquitin ligases in cell division cycle control[J]. Cellular and Molecular Life Sciences, 2008, 65(21): 3458 − 3466. doi: 10.1007/s00018-008-8220-1 [18] WANG Q L. The role of forkhead-associated (FHA)-domain proteins in plant biology[J]. Plant Molecular Biology, 2023, 111(6): 455 − 472. doi: 10.1007/s11103-023-01338-4 [19] 昌燕李, 韦运谢. 木薯MeCAMTA基因的克隆与原核表达[J]. 分子植物育种, 2020, 18(3): 744 − 750. doi: 10.13271/j.mpb.018.000744 [20] HERB M, SCHRAMM M. Functions of ROS in macrophages and antimicrobial immunity[J]. Antioxidants, 2021, 10(2): 313. doi: 10.3390/antiox10020313 [21] MACHIDA S, YUAN Y A. Crystal structure of Arabidopsis thaliana dawdle forkhead-associated domain reveals a conserved phospho-threonine recognition cleft for dicer-like 1 binding[J]. Molecular Plant, 2013, 6(4): 1290 − 1300. doi: 10.1093/mp/sst007 [22] NARAYANAN L A, MUKHERJEE D, ZHANG S X, et al. Mutational analyses of a fork head associated domain protein, DAWDLE, in Arabidopsis thaliana[J]. American Journal of Plant Sciences, 2014, 5(18): 2811 − 2822. doi: 10.4236/ajps.2014.518297 [23] DING Z F, WANG H C, LIANG X Y, et al. Phosphoprotein and phosphopeptide interactions with the FHA domain from Arabidopsis kinase-associated protein phosphatase[J]. Biochemistry, 2007, 46(10): 2684 − 2696. doi: 10.1021/bi061763n [24] GUO X, YU X L, XU Z Y, et al. CC-type glutaredoxin, MeGRXC3, associates with catalases and negatively regulates drought tolerance in cassava (Manihot esculenta Crantz)[J]. Plant Biotechnology Journal, 2022, 20(12): 2389 − 2405. doi: 10.1111/pbi.13920 [25] MA X W, MA Q X, MA M Q, et al. Cassava MeRS40 is required for the regulation of plant salt tolerance[J]. Journal of Integrative Agriculture, 2023, 22(5): 1396 − 1411. doi: 10.1016/j.jia.2023.04.003 [26] CUI M, AN F F, CHEN S B, et al. Expression pattern and functional analysis of MebHLH149 gene in response to cassava bacterial blight[J]. Plants, 2024, 13(17): 2422. doi: 10.3390/plants13172422 [27] SIES H, BELOUSOV V V, CHANDEL N S, et al. Defining roles of specific reactive oxygen species (ROS) in cell biology and physiology[J]. Nature Reviews Molecular Cell Biology, 2022, 23(7): 499 − 515. doi: 10.1038/s41580-022-00456-z [28] LIU P, ZHANG X X, ZHANG F, et al. A virus-derived siRNA activates plant immunity by interfering with ROS scavenging[J]. Molecular Plant, 2021, 14(7): 1088 − 1103. doi: 10.1016/j.molp.2021.03.022 [29] KONG L, MA X Y, ZHANG C, et al. Dual phosphorylation of DGK5-mediated PA burst regulates ROS in plant immunity[J]. Cell, 2024, 187(3): 609 − 623.e21. doi: 10.1016/j.cell.2023.12.030 [30] WEI J, WANG X, HU Z, et al. The Puccinia striiformis effector Hasp98 facilitates pathogenicity by blocking the kinase activity of wheat TaMAPK4[J]. Journal of Integrative Plant Biology, 2022, 65(1): 249 − 264. doi: 10.1111/jipb.13374 -

 点击查看大图
点击查看大图
计量
- 文章访问数: 747
- HTML全文浏览量: 499
- PDF下载量: 11
- 被引次数: 0
