

-
木薯(Manihot esculenta)是世界三大薯类作物之一,至少有4 000多年的栽培史和食用史[1]。19世纪20年代引进中国,主要种植在广东、广西、海南和福建等省区[2]。木薯块根富含淀粉,干样淀粉含量可达71.6%,还含有蛋白质、脂肪、维生素及钙、钾等微量元素[3]。木薯还是重要的生物能源作物,通过木薯的发酵和转化,可以生产出生物燃料如乙醇等[4]。然而,木薯在生长过程中会受到多种病害的侵袭,如细菌性枯萎病(cassava bacterial blight),木薯花叶病(cassava mosaic disease),褐斑病(brown speckle disease)等,这些病害不仅影响木薯的正常生长,还能导致产量下降和品质受损[5]。研究木薯抗病信号网络,对提高木薯抗病性起到关键作用。
植物在漫长的进化过程中演变出2大天然免疫系统,包括基础免疫反应和效应子触发的免疫反应,基础免疫反应是由病原物相关分子模式所诱发的植物免疫反应,这些分子模式是病原菌特有的、能够触发植物免疫反应的特征性信号[6]。病原菌释放出的分子模式会被模式识别受体识别,从而引发一系列的信号转导,包括诱导防御基因的表达、活性氧产生等[7]。这使得植物能够产生抗菌物质、增强细胞壁结构以及启动系统性免疫反应以抑制病原微生物的生长[8-9]。其中,模式识别受体由位于质膜上的类受体蛋白激酶或类受体蛋白组成。类受体蛋白激酶(receptor kinase,RLK)在生物体内,特别是在植物中,起着极其重要的作用。它们是一类存在于细胞膜上的单次跨膜蛋白,其具有一个存在于胞外用于接受外源信号的LRR结构域、一个作为信号传递工具的跨膜结构域及一个存在于细胞内通过磷酸化完成信号传递的激酶结构域,它们通过感知和响应外界信号,参与调控植物生长发育、免疫防御等多个过程[10-11]。在拟南芥(Arabidopsis thaliana)中有超过600个该类蛋白,其胞外受体结构域与信号分子结合后激活胞内激酶结构域,通过磷酸化作用将信号传递给下游,最终产生一系列生物效应[12 − 13]。类受体蛋白激酶可以根据胞外域的结构差异分为多个激酶家族,其中,富含亮氨酸的类受体蛋白激酶(Leucine-rich repeat receptor-like protein kinases,LRR-RLKs)是拟南芥中最大且研究最深入的RLK亚家族[14]。类受体蛋白激酶可以参与调控不同类型的免疫反应,与其他免疫组分(如共受体、防御蛋白等)相互作用形成复合物,这种互作不仅增强了免疫反应的强度和特异性,还使得植物能够更精准地应对不同类型的病原菌[15]。
周萌萌[16]通过分离鉴定得到RLK家族的MeRLK1,发现其参与木薯抗病。在其他植物中RLK1可以通过与蛋白互作来参与信号传导、免疫反应等过程,如拟南芥中类受体蛋白激酶(Wall-associated receptor-like kinase 4,WAKL4)与Cd转运蛋白(Natural resistance-associated macrophage protein 1,NRAMP1)相互作用并使其磷酸化,导致NRAMP1的泛素化和液泡依赖性降解增强,从而降低Cd摄取[17]。但MeRLK1在木薯体内的互作网络尚不清楚。为了探寻能够与MeRLK1蛋白协同调控木薯抗病的相关蛋白,本研究利用酵母双杂交技术,筛选MeRLK1的候选互作蛋白并初步验证候选互作蛋白与MeRLK1的相互作用,旨在进一步揭示木薯中RLK1的功能和作用机制,为木薯产业奠定抗病基础。
-
‘华南124号’木薯种植于海南省耐盐作物生物技术重点实验室;反转录酶(Cat K1622)、限制性内切酶BamH I (Cat FD0055)、EcoR I (Cat FD0275)购自Thermo Fisher公司;DNA聚合酶(Cat P112-03)购自Vazyme公司;质粒提取试剂盒(Cat DP103-03)、DNA纯化试剂盒(Cat DP214-03)购自天根生化公司;缺陷型培养基(Cat PM2220 PM2110)购自北京酷来搏科技有限公司;大肠杆菌DH5α (Cat DL1001S)购自上海唯地生物公司;酵母双杂交载体质粒pGBKT7、pGADT7由海南省耐盐作物生物技术重点实验室保存。
-
根据Tropical Crops Omics Database数据库(
https://ngdc.cncb.ac.cn/tcod/ )中MeRLK1的CDS序列信息[18],分别查询分析了MeRLK1在木薯不同组织部位中及叶片接种地毯草黄单胞菌木薯萎蔫致病变种(Xanthomonas axonopodis pv. manihotis,Xam)后1、3、5 d的表达量。 -
将pGBKT7-MeRLK1和pGADT7质粒共同转化到AH109酵母中,涂布于二缺(SD-T-L)和四缺(SD-T-L-H-A)培养基,检测MeRLK1蛋白是否存在自激活。
-
将pGBKT7-MeRLK1的质粒与木薯AD文库质粒共转入AH109中,并将产物分别涂在二缺和四缺缺陷培养基上倒置于28 ℃培养箱培养4 d,挑取单菌落于二缺液体培养基中培养2 d。提取酵母质粒,利用AD通用引物进行PCR扩增,琼脂糖凝胶电泳后切取单一条带纯化回收,并送华大公司测序,测序结果与Phytozome v13(
https://phytozome-next.jgi.doe.gov/ )中的木薯库进行序列比对,筛选出MeRLK1可能互作的蛋白并对其进行功能注释、亚细胞定位预测及聚类分析。通过Tropical Crops Omics Database数据库分析24个候选互作基因在不同组织部位及叶片接种Xam后的表达量,利用Tbtools软件制作表达量热图及相关性分析图。 -
以培养4周的木薯为材料,采用CTAB(cetyltrimethylammonium bromide)法提取总RNA,逆转录制备cDNA。以cDNA为模板,根据Phytozome v13数据库中MeBSK(brassinolide-signaling kinase) CDS序列设计特异性引物。利用cDNA扩增出MeBSK全长并进行胶回收,与中间载体pEASY-T3连接,转入DH5α后在氨苄固体培养基上培养12 h,进行菌落PCR验证,挑选明亮单一的单克隆送测序,比对测序结果并保留序列正确的菌株。
-
以测序正确的质粒为模板扩增MeBSK片段,使用BamH I和EcoR I对载体pGADT7进行双酶切使其线性化,纯化酶切产物后与MeBSK片段进行同源重组[19],经菌落PCR验证后选取条带大小正确的菌液提质粒并酶切验证。
-
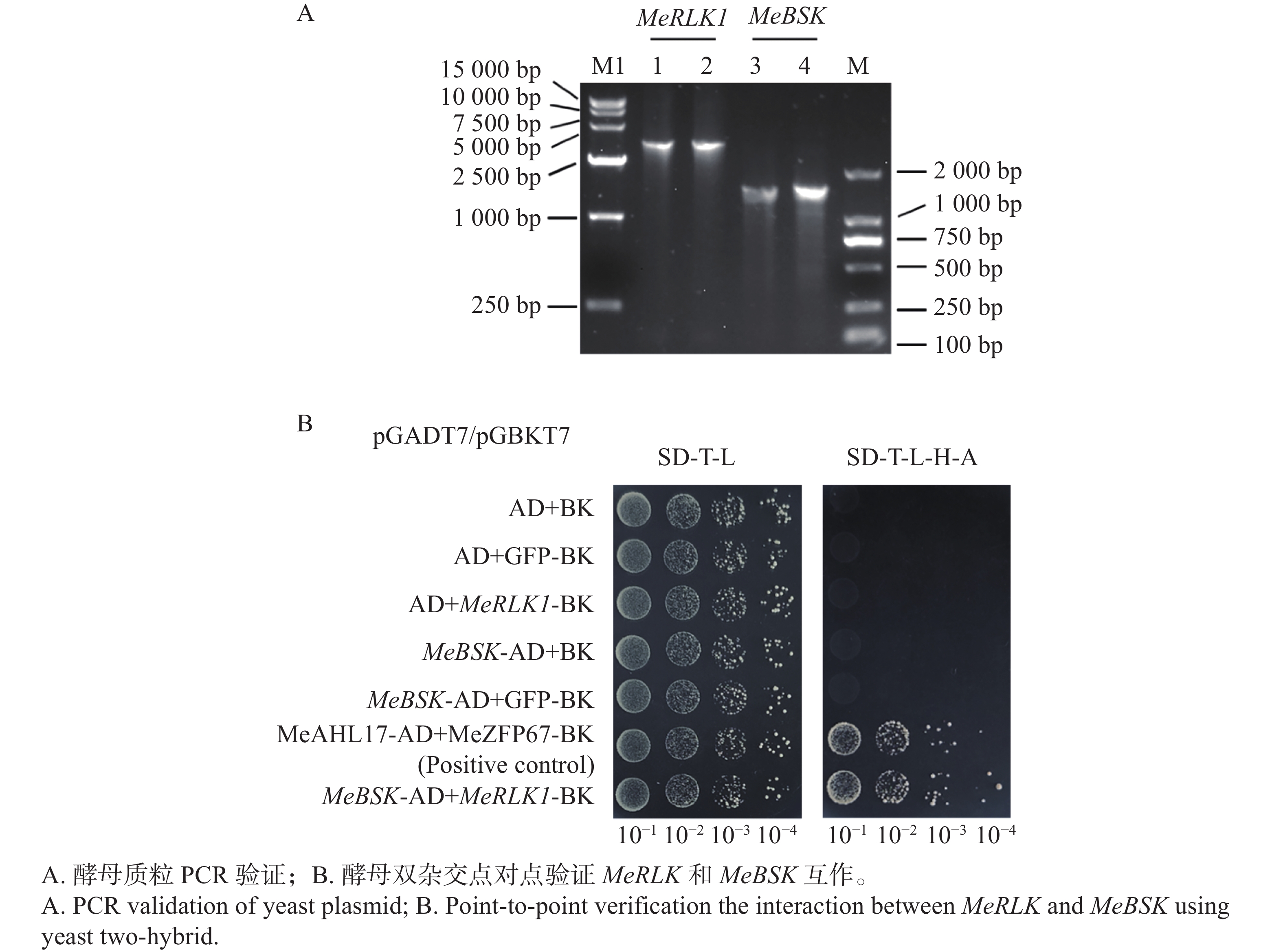
将pGBKT7-MeRLK1和pGADT7-MeBSK共转入AH109酵母感受态中,筛选出阳性克隆提取质粒,分别利用MeRLK1和MeBSK全长引物对酵母质粒进行PCR扩增。将验证正确的菌株及对照菌株按10−1、10−2、10−3、10−4进行梯度稀释,分别点滴于二缺和四缺培养基上,观察其生长情况。
-
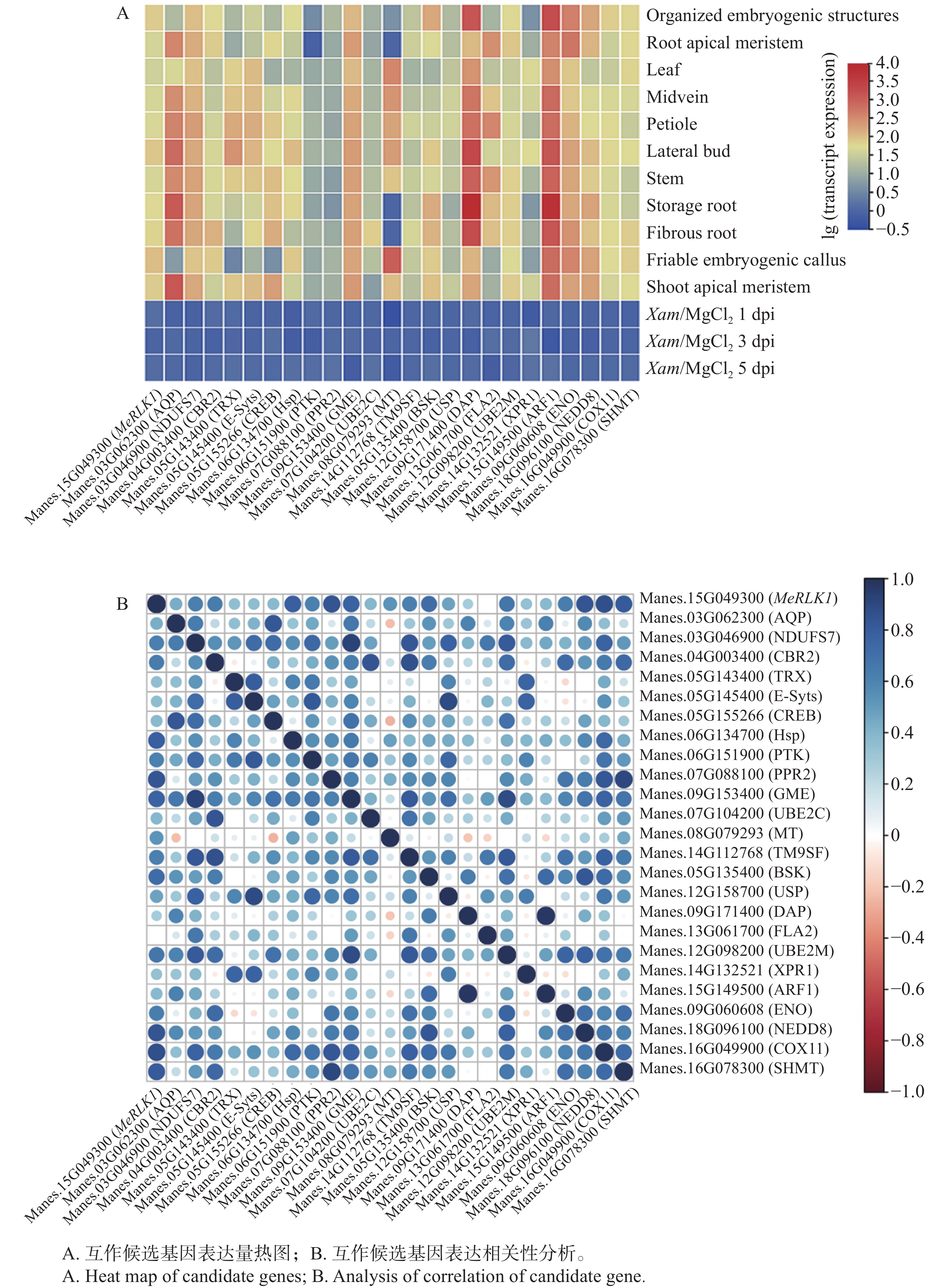
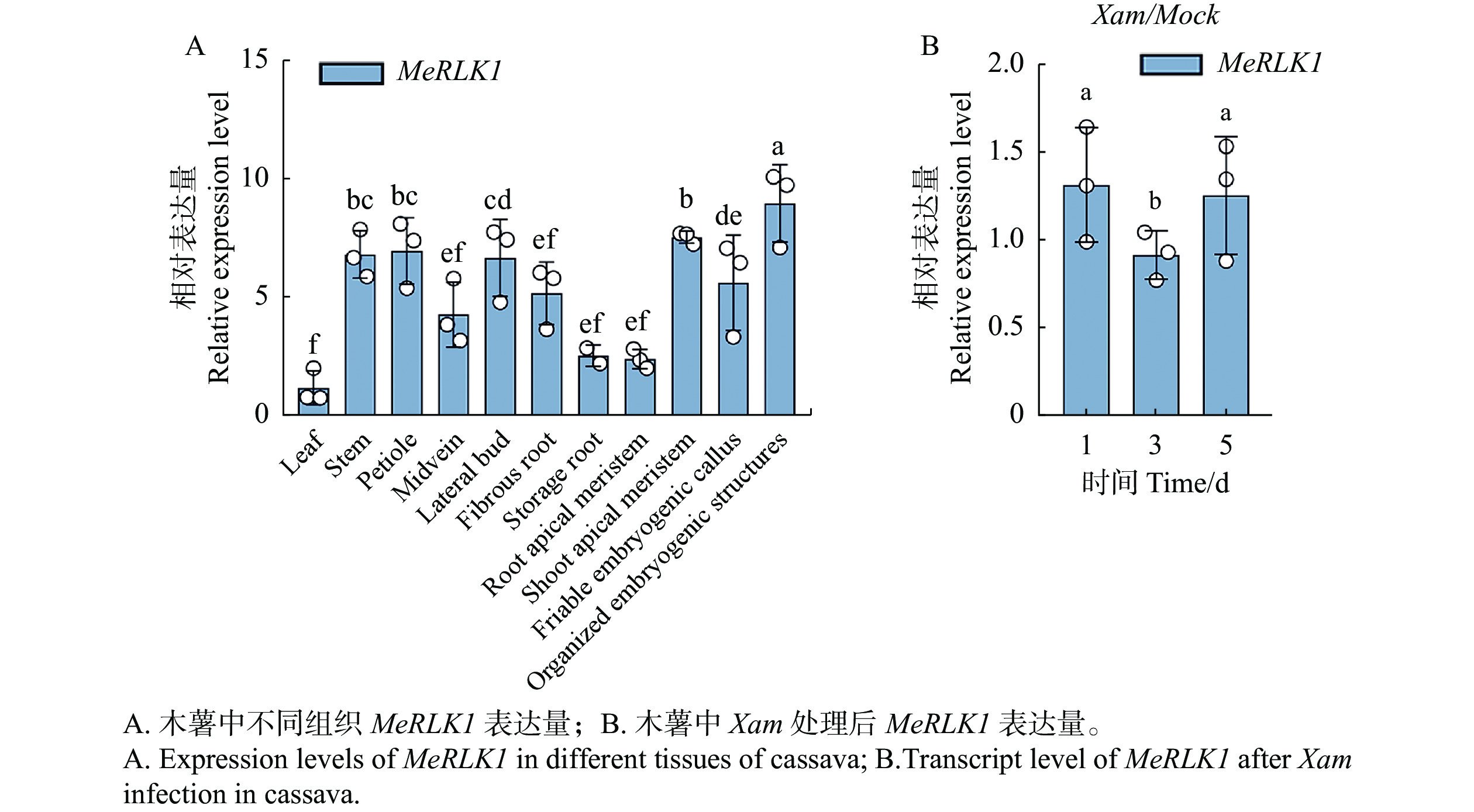
根据Tropical Crops Omics Database数据库分析了MeRLK1在木薯中叶、叶中脉、叶柄、侧芽、茎、贮藏根、须根、胚源性愈伤组织、有序胚胎发生结构、顶端分生组织、根尖分生组织等11个不同组织部位的基因表达量。结果发现,MeRLK1在木薯有序胚胎发生结构的表达量最高,其次是顶端分生组织、叶柄、茎、侧芽,有序胚胎发生结构、须根、叶中脉的表达量较低,在贮藏根、根尖分生组织、叶的表达量最低(图1-A)。同时还发现,木薯叶片在接种Xam的1、3、5 d后MeRLK1的相对表达量变化显著,表明MeRLK1可以响应Xam的侵染(图1-B)。
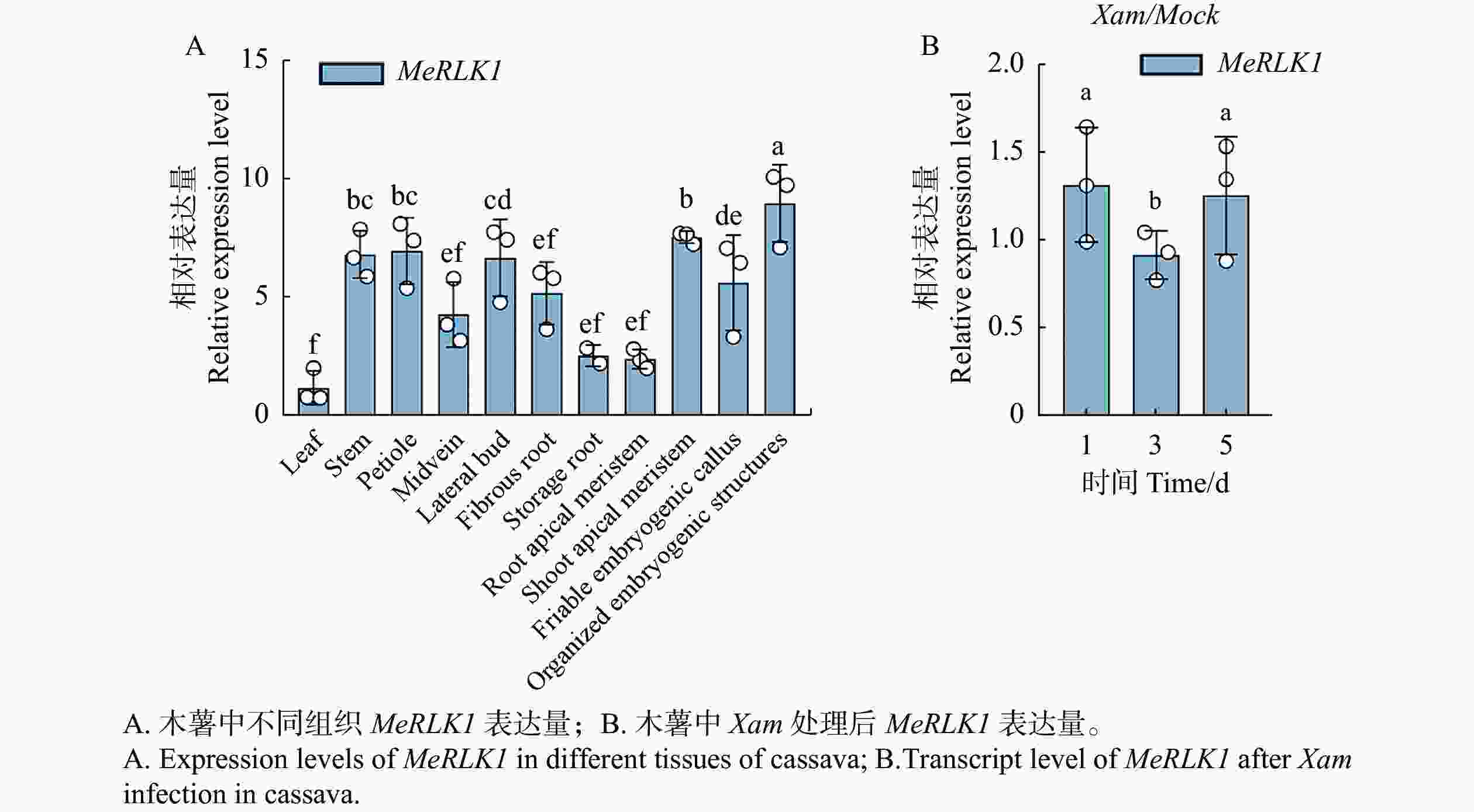
图 1 MeRLK1表达谱分析
Figure 1. Expression levels of MeRLK1
-
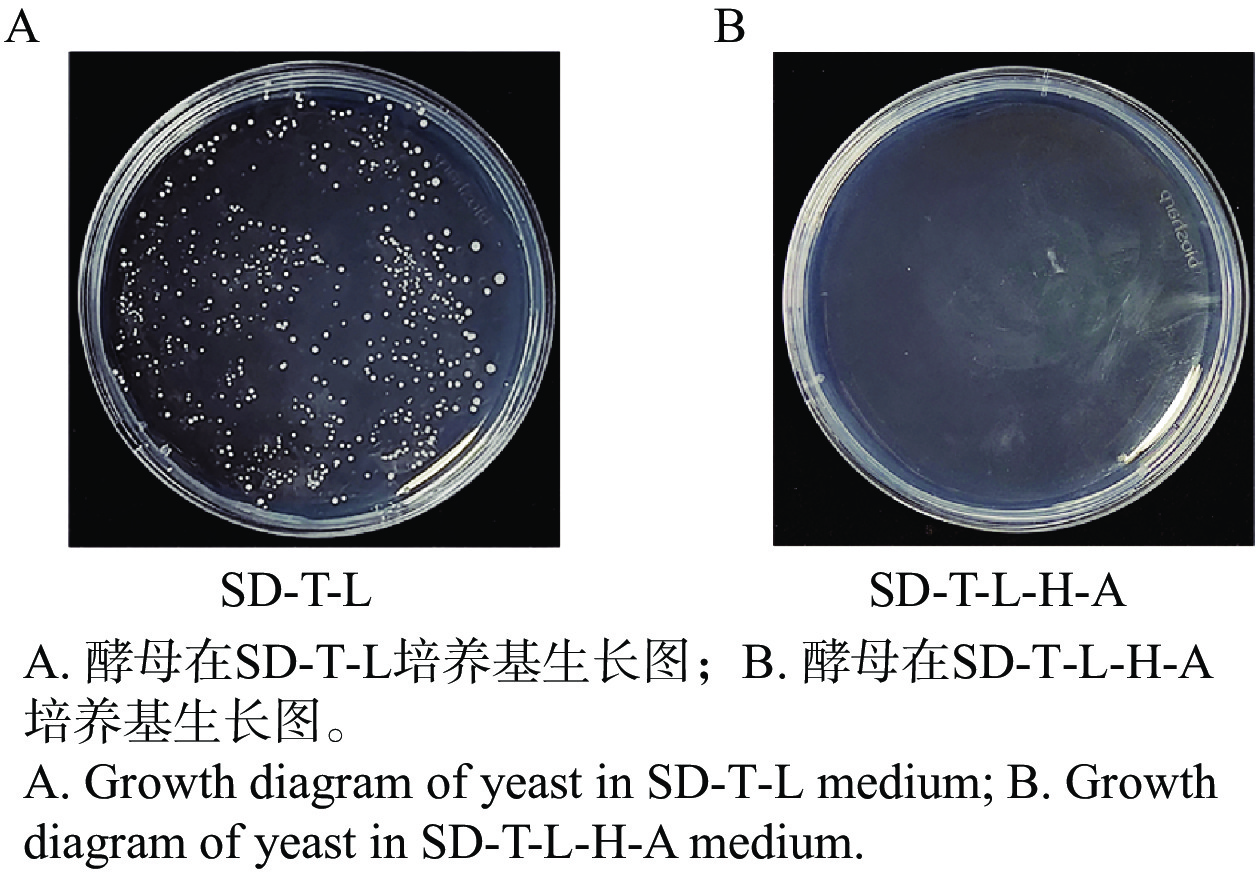
将pGBKT7-MeRLK1和pGADT7空载共转AH109进行自激活检测。实验结果发现,共转后的酵母菌在二缺培养基上的生长情况表明质粒成功转入酵母细胞,但在四缺培养基上无法正常生长(图2-A、图2-B),表明MeRLK1蛋白不具有自激活活性,可用于后续对木薯cDNA文库进行候选互作蛋白筛选。
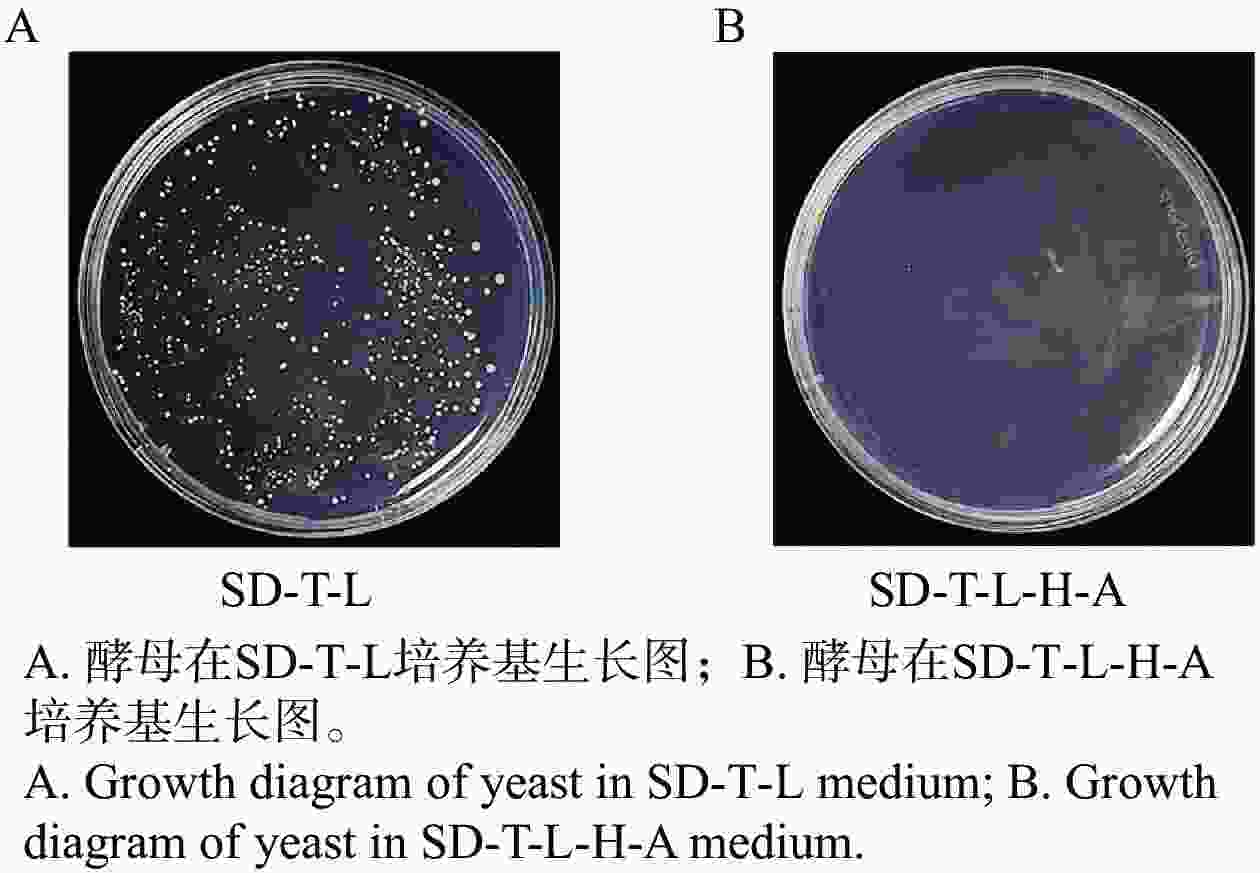
图 2 MeRLK1自激活验证
Figure 2. Verification of MeRLK1 auto-activation
-
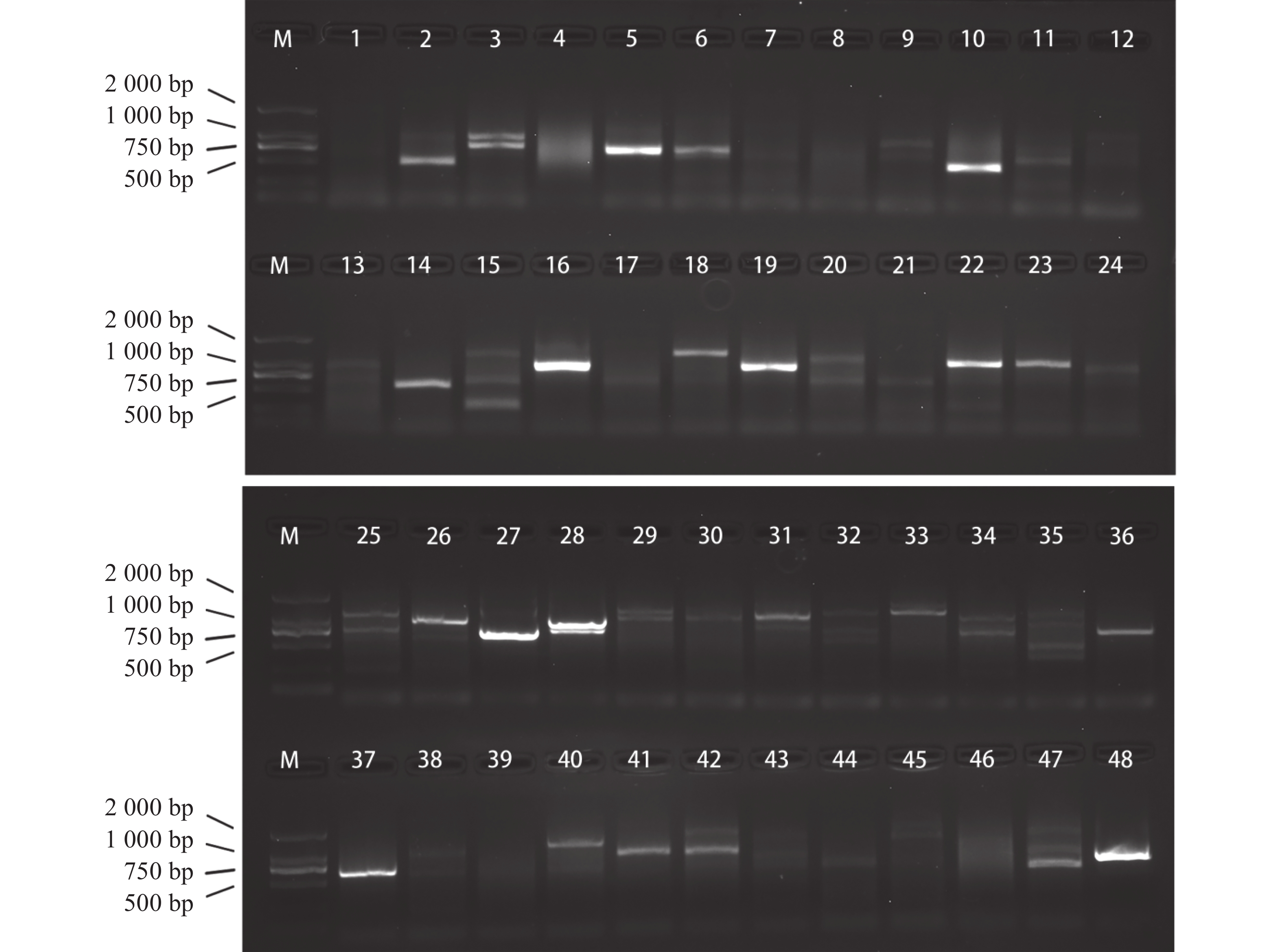
将pGADT7质粒文库与pGBKT7-MeRLK1质粒共转入AH109中,通过PCR鉴定,结果显示,PCR产物电泳条带大多在500~2 000 bp之间(图3),说明成功筛选到多个可能与MeRLK1互作的蛋白。送测后将测序结果与Phytozome v13数据库比对,去除移码和重复,初步获得24个与MeRLK1互作的候选互作蛋白,根据Phytozome v13数据库中的信息对其进行注释,并通过亚细胞定位预测网站(
https://wolfpsort.hgc.jp/ )对候选互作蛋白亚细胞定位进行预测(表1)。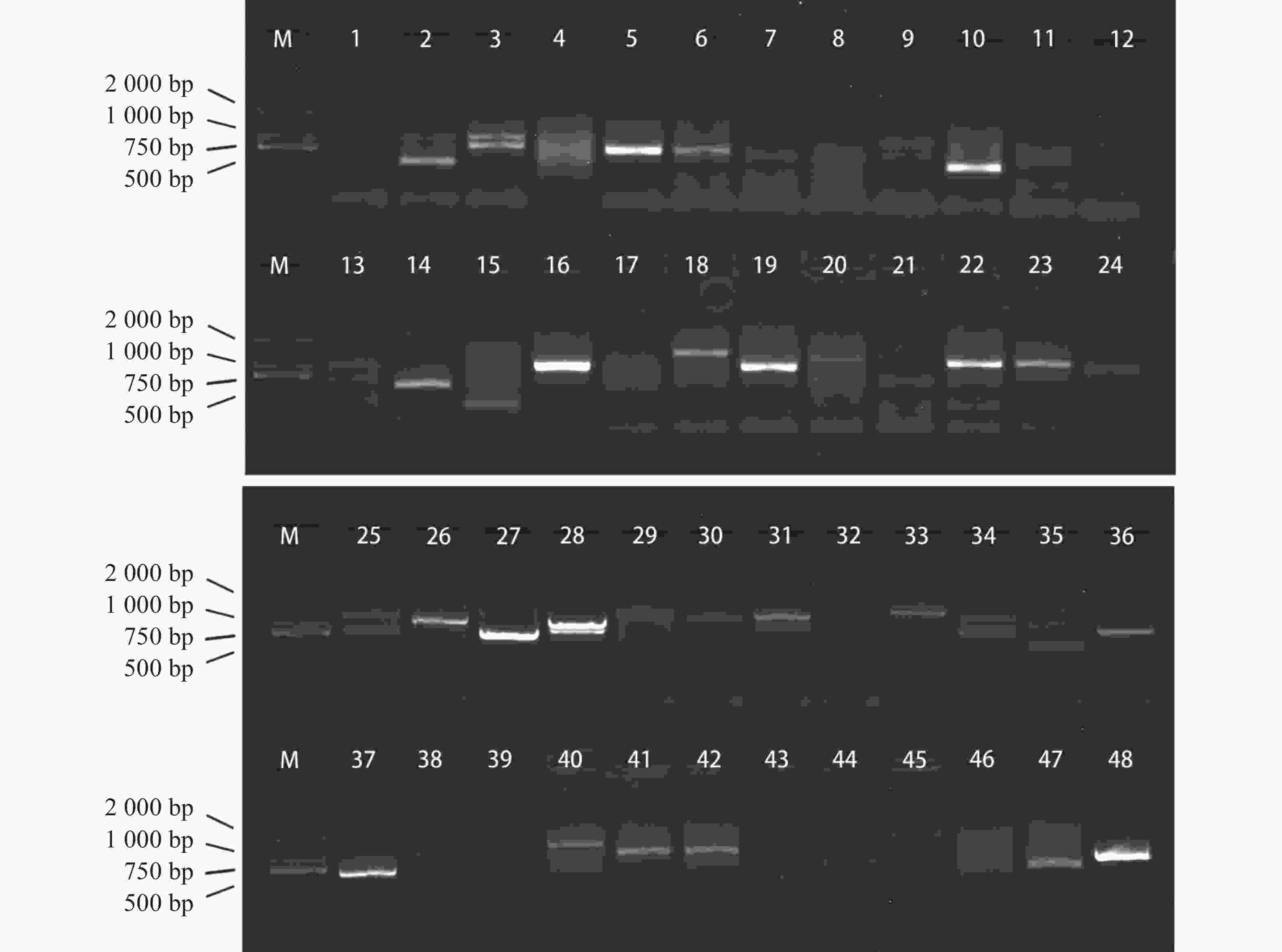
图 3 酵母双杂交筛选MeRLK1候选互作蛋白
Figure 3. Y2H screening for MeRLK1-interacting proteins
表 1 MeRLK1候选互作蛋白注释
Table 1. Annotation of candidate interacting proteins of MeRLK1
序号
Number基因号
Gene identifier蛋白名
Protein基因注释
Gene annotation亚细胞定位预测
Subcellular localization prediction1 Manes.03G062300 AQP Aquaporin transported Cell membrane 2 Manes.03G046900 NDUFS7 NADH dehydrogenase (ubiquinone) Fe-S protein 7 Mitochondrial matrix 3 Manes.04G003400 CBR2[20] NADH-cytochrome b5 reductase Chloroplast stroma 4 Manes.05G143400 TRX Thioredoxin Nucleus/Cytoplasm 5 Manes.05G145400 E-Syts[21] Extended synaptotagmin-related Cytoplasm/Endoplasmic reticulum 6 Manes.05G155266 CREB[21] cAMP-response element binding prorein related Nucleus 7 Manes.06G134700 Hsp Heat shock protein, mitochondrial-ralated Mitochondrial/Chloroplast 8 Manes.06G151900 PTK Protein tyrosine kinase Chloroplast 9 Manes.07G088100 PPR2 Pentatricopeptide repeat 2 Cytoplasm 10 Manes.09G153400 GME[22] GDP-D-mannose 3′, 5′-epimerase Cytoplasm 11 Manes.07G104200 UBE2C[23] Ubiquitin-conjugating enzyme E2C Nucleus 12 Manes.08G079293 MT Metallothionein Chloroplast 13 Manes.14G112768 TM9SF[24] Transmembrane 9 superfamily member Cell membrane 14 Manes.05G135400 BSK[25] BR-signaling kinase Chloroplast 15 Manes.12G158700 USP Universal stress proteins Chloroplast 16 Manes.09G171400 DAP Dormancy/auxin associated protein Nucleus 17 Manes.13G061700 FLA2 Fasciclin-like arabnogalactan protein 2 Cell membrane 18 Manes.12G098200 UBE2M[26] Ubiquitin-conjugating enzyme E2M Nucleus/Cytoplasm 19 Manes.14G132521 XPR1 Xenotropic and polytropic reterovirus receptor 1 Cytoplasm 20 Manes.15G149500 ARF1 ADP-ribosylation factor 1 Peroxisome 21 Manes.09G060608 ENO Enolase Cytoplasm 22 Manes.18G096100 NEDD8 Ubiquitin-like protein Nucleus/Cytoplasm 23 Manes.16G049900 COX11 Cytochrome c oxidase assembly protein subunit 11 Mitochondrial/Chloroplast 24 Manes.16G078300 SHMT[26] Glycine hydroxymethyltransferase Chloroplast 注:基因号从Phytozome v13数据库中获得。
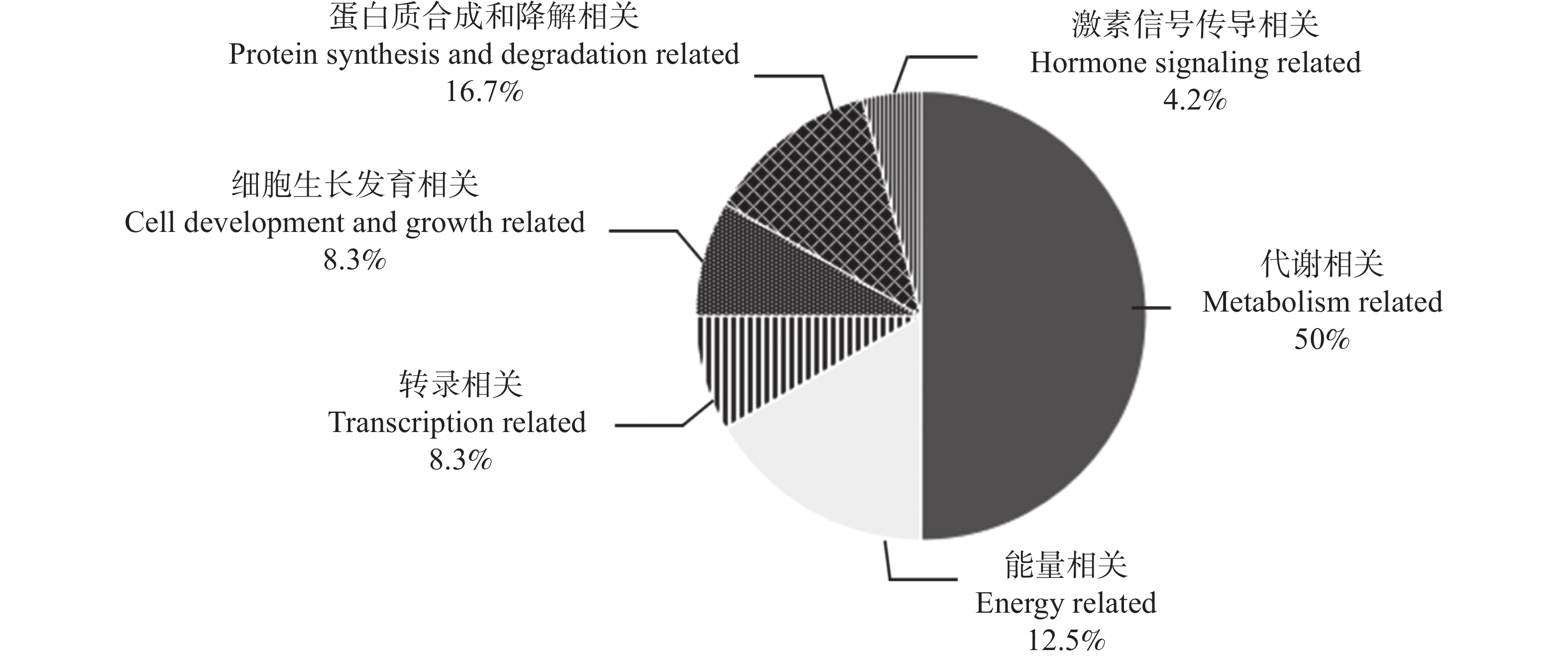
Note: The gene ID was obtained from Phytozome v13 database.依照蛋白功能对MeRLK1筛选获得的互作候选蛋白进行分类(图4),分类结果显示,互作候选蛋白中有50%与细胞代谢相关,12.5%参与能量代谢过程,8.3%与生长发育相关,16.7%与蛋白质合成与降解有关,8.3%与转录相关,4.2%参与激素信号传导即油菜素内酯信号激酶BSK。
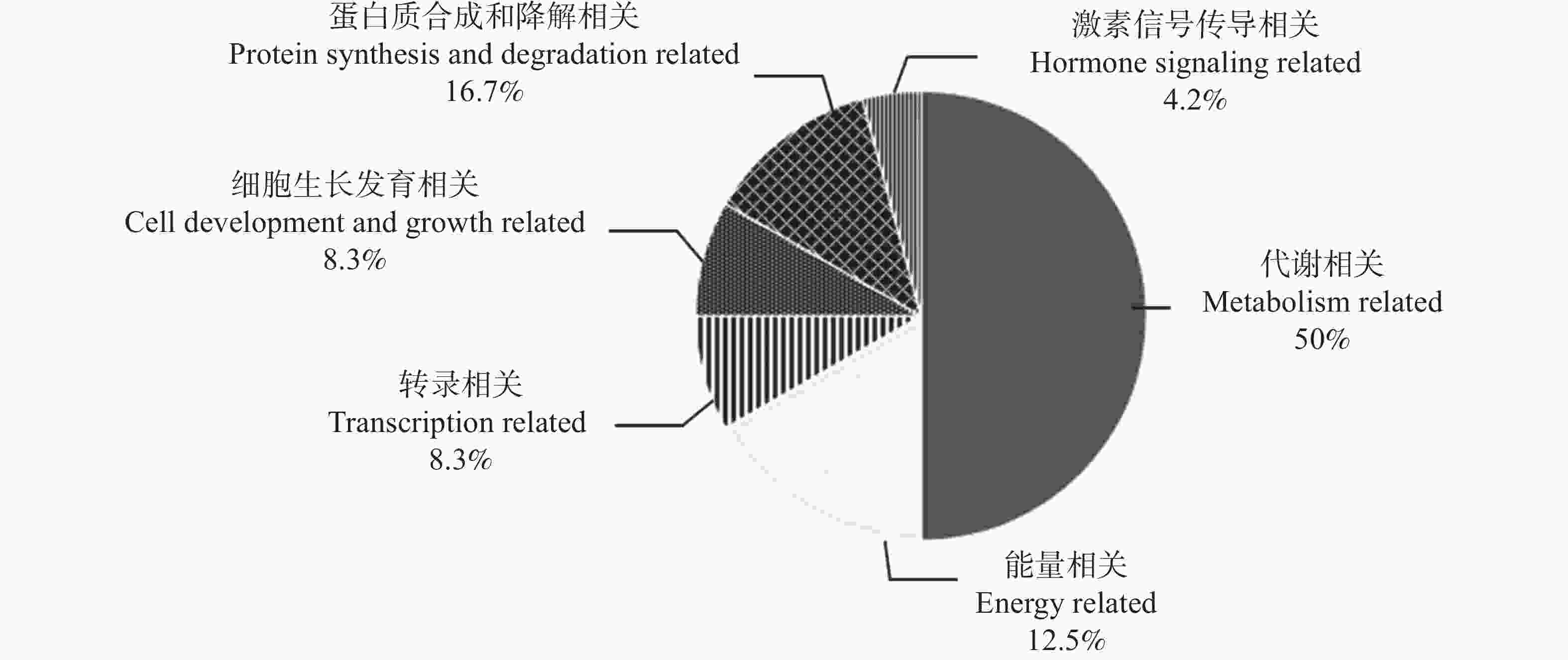
图 4 互作候选蛋白功能聚类分析图
Figure 4. Functional clustering analysis diagram of candidate interacting proteins
根据Tropical Crops Omics Database数据库分析了24个互作候选基因在木薯中11个不同组织部位的基因表达量及木薯叶片接种Xam后的表达量。应用Tbtools软件构建了MeRLK1与互作候选基因相关性分析热图,结果发现,与细胞生长发育相关(1个)、激素信号传导相关(1个)、能量相关(4个)以及代谢相关(11个)的基因都能在叶中脉、叶柄、侧芽、茎、储藏根、顶端分生组织中高度表达(图5-A),此外,通过对MeRLK1与其他24个基因的表达量进行了相关性分析,发现其与HSP、GME、COX11、SHMT、PPR2、BSK、NEDD8等相关系数较高(R>0.7)(图5-B),表明MeRLK1可能会与以上基因共同调控信号传导及能量代谢等途径,但各个途径之间是否存在交集还有待进一步研究。
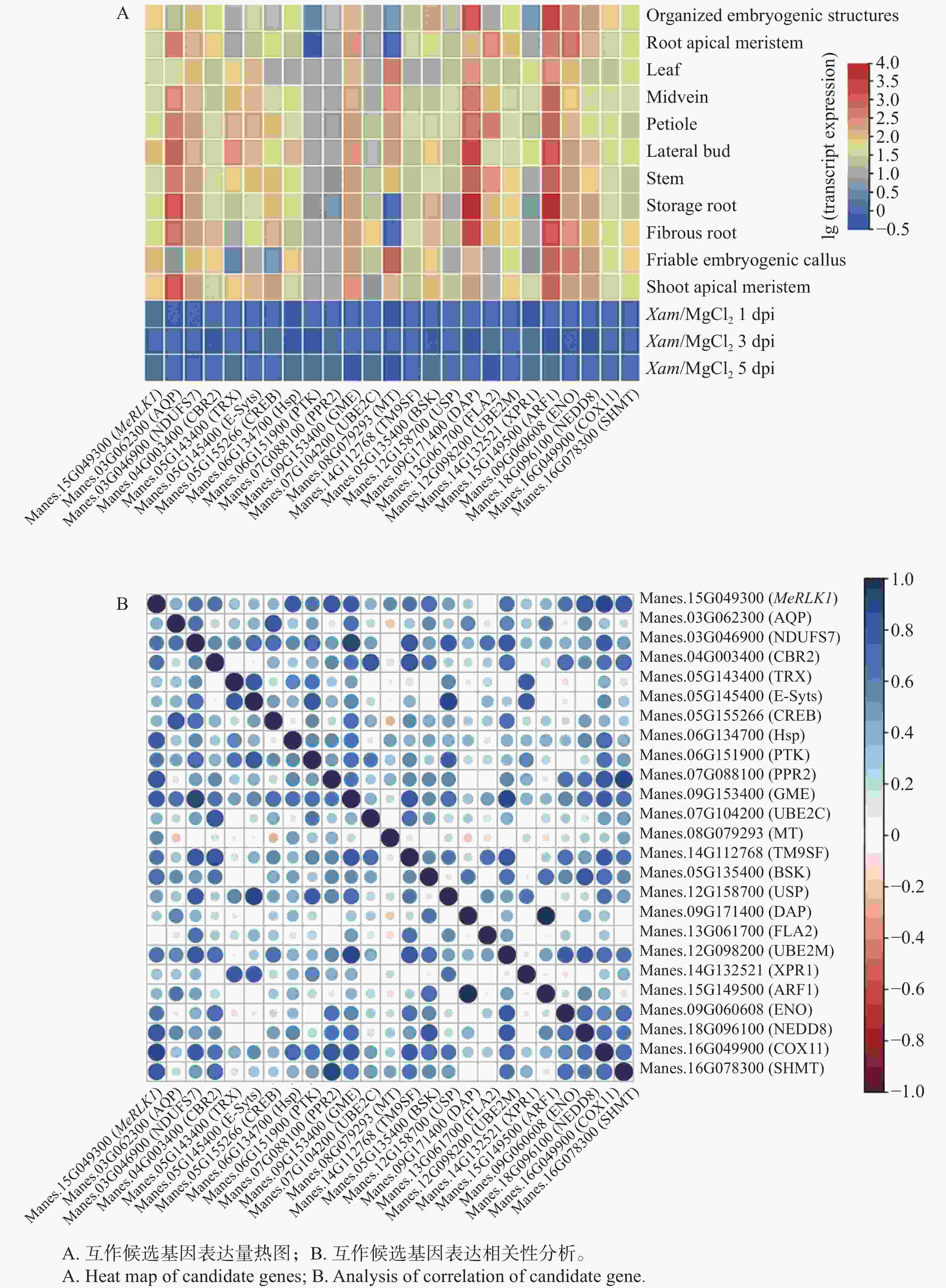
图 5 MeRLK1与互作候选基因表达相关性分析
Figure 5. Analysis of correlation between MeRLK and candidate-interaction protein gene
-
植物激素通过不同的信号传导途径参与植物的生理反应,在植物生长发育过程中起到重要的调控作用,而BSK作为互作候选蛋白中一个与植物激素信号传导相关的蛋白,参与着BR信号通路的传导过程。因此,选取BSK候选蛋白进一步验证相互作用。本研究以‘华南124’木薯品种的cDNA为模板,根据Phytozome v13中MeBSK的CDS序列,设计特异性引物扩增MeBSK的全长序列并构建pGADT7-MeBSK表达载体。利用MeRLK1和MeBSK全长引物对共转后的酵母质粒进行PCR验证,结果显示所提质粒可以分别扩增出MeRLK1和MeBSK全长片段大小相同的片段(图6-A)。将这些菌株及对照菌株按10−1、10−2、10−3、10−4进行梯度稀释,分别点滴于二缺和四缺培养基上,观察生长情况发现,实验组和对照组的酵母菌株均可以在二缺平板上生长,而在四缺平板上只有实验组pGBKT7-MeRLK1+pGADT7-MeBSK的菌株可以正常生长(图6-B)。通过酵母双杂交对MeRLK1和MeBSK的互作关系进行验证,但仅酵母点对点验证无法排除假阳性确定互作关系,后续将需要进一步在体内体外通过BiFC、Pull down等互作实验确认互作关系。
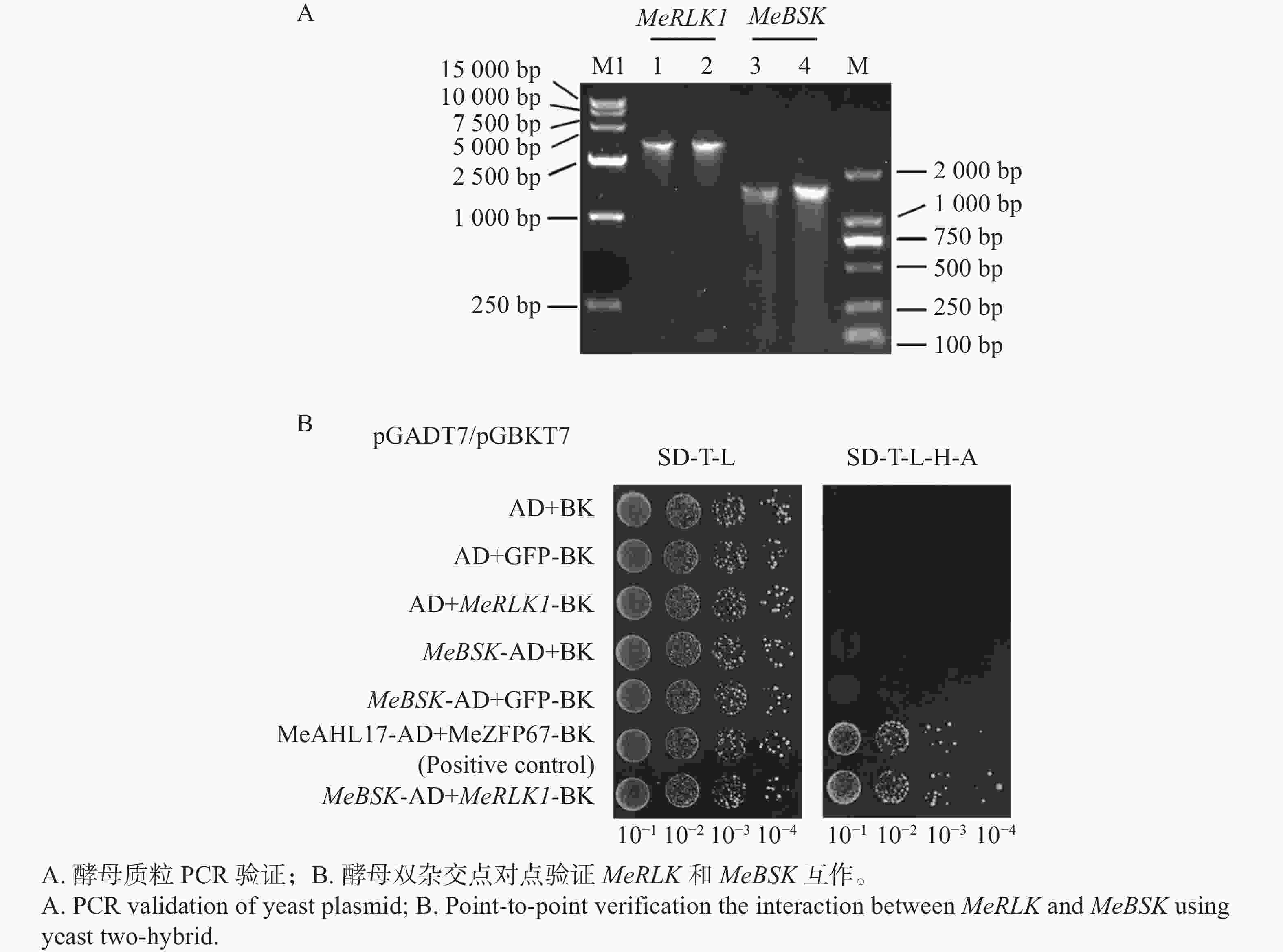
图 6 酵母双杂交验证
Figure 6. Yeast two hybrid validation
-
类受体蛋白激酶是一类在植物中不可或缺的跨膜蛋白,它们通过感知外部信号(如激素、病原体等),调控植物的生长发育,包括细胞分裂、分化和器官形成[27−28]。RLKs还参与植物的免疫防御,识别病原体并激活抗病反应。此外,它们可以启动防御机制并提高植物抗逆性,在非生物胁迫(如干旱、盐害)的适应中起关键作用[29]。
本研究利用酵母双杂交技术系统筛选得到24个MeRLK1的互作候选蛋白,分析相关系数发现,MeRLK1与候选基因存在时空表达的同步性。激素信号在植物中起着核心调控作用,它们通过复杂的信号网络协调植物的生长和适应能力。因此,本研究根据激素信号通路筛选出了1个BR信号转导途径下游的关键受体激酶BSK,分析后发现,该基因与MeRLK1存在较高的相关性,并通过酵母双杂点对点进一步验证了MeBSK与MeRLK1的互作关系。类受体蛋白激酶作为植物中最大的跨膜信号感知家族,通过其胞外结构域特异性识别配体(如多肽、激素或病原相关分子),并依赖胞内激酶结构域与下游蛋白质形成互作网络,以此实现对植物生长发育、免疫反应的调控。例如,类受体蛋白激酶FERONIA与RALF肽信号互作后可以调节钙离子通道使胞内Ca2+浓度升高,激活Ca2+依赖的蛋白激酶,进而磷酸化多种代谢相关的酶,如参与糖酵解和三羧酸循环的酶来调控能量代谢[30]。此外,类受体蛋白激酶TMK1与质膜H+-ATP酶相互作用,介导其磷酸化,导致质外体酸化从而调控植物根的生长[31]。基于上述文献及本研究筛选得到的CREB、GME等蛋白,推测MeRLK1可能通过与多种ATP酶(如质膜H+-ATP酶、线粒体ATP酶等)相互作用,调控其磷酸化状态或活性,从而影响ATP的合成与水解过程。亦或者MeRLK1还可能通过调节信号分子(如钙离子、活性氧等)的传递,间接调控能量代谢相关通路,最终影响植物的生长发育和逆境响应。但MeRLK1是否且如何直接或间接调控植物的能量代谢过程还有待进一步研究,这一推想为进一步研究MeRLK1在植物能量代谢中的具体作用机制提供了新的思路。在拟南芥中,LRR1与泛素连接酶PUB11互作介导蛋白降解从而负调控植物的干旱胁迫响应[32]。基于本研究的筛库结果发现,UBE2M与UBE2C同样参与泛素化修饰,因此,初步推测MeRLK1可能通过与UBE2M和UBE2C互作参与其他蛋白的泛素化修饰,但需要进一步通过体内体外酵母双杂、BiFC等验证互作,以及免疫沉淀结合质谱分析鉴定泛素化靶蛋白及修饰位点等实验深入研究泛素化修饰。但该筛选结果至少提供了下一步可供MeRLK1参与泛素化修饰的候选靶蛋白,为解析MeRLK1通过泛素化调控植物胁迫响应的分子通路奠定了基础。
本研究通过酵母筛选并初步验证了MeRLK1与BR信号关键组分MeBSK的互作,暗示着MeRLK1在BR信号通路中的潜在作用及二者介导免疫反应之间的潜在联系。在植物中,BR信号通路是一个调节植物生长发育的重要信号转导途径,其起始于激素信号与受体BRI1的结合[33]。BRI1是一种位于细胞膜上的富含亮氨酸重复序列的类受体蛋白激酶。当BR与BRI1结合后,会引起BRI1自身磷酸化,随后激活其下游的激酶。BSK是受体样细胞质激酶-XⅡ 亚家族的成员,其可作为接收来自BRI1信号的重要下游,并将信号进一步向下游传递[34]。除此之外,类受体蛋白BAK1通过与BRI1相互作用参与发育调节,并通过与FLS2相互作用参与对病原体的先天免疫[35]。MeRLK1与BRI1同属于类受体蛋白激酶家族,由MeRLK1与BSK的初步互作关系推测MeBSK可以接受来自MeRLK1传递的信号来调控BR信号网络,且MeRLK1可能与其他类受体蛋白如FlS2、BAK1等互作参与植物的免疫反应,但后续还需通过体内体外对MeRLK1与BSK的互作关系作进一步验证。这一推测为深入研究MeRLK1在植物生长发育和免疫调控中的多重功能提供了重要的理论依据,同时也为解析类受体蛋白激酶家族成员在植物信号网络中的功能多样性和协同作用机制开辟了新的研究方向。
作为BR信号系统关键激酶,本研究发现,MeRLK1在酵母系统中与MeBSK互相作用,但MeRLK1与MeBSK互作是否会影响MeRLK1的抗病性尚不清楚,并且MeRLK与MeBSK的互作是否影响BR信号通路的传导也有待进一步研究。本研究利用酵母双杂技术,鉴定到24个候选蛋白与MeRLK1可能存在相互作用,丰富了RLK家族的互作网络,为抗性基因的筛选提供新思路和想法,为木薯遗传改良奠定基础。
木薯RLK1候选互作蛋白筛选及其与BSK互作的酵母验证
DOI: 10.15886/j.cnki.rdswxb.20240187
 CSTR: 32425.14.j.cnki.rdswxb.20240187
CSTR: 32425.14.j.cnki.rdswxb.20240187
Screening of the candidate interacting protein of RLK1 and validation of its interacting relationship with BSK by yeast two-hybrid in cassava
-
摘要: 为了筛选出木薯(Manihot esculenta)中与MeRLK1 (receptor-like kinase 1)共同调控木薯抗病的相关蛋白,本研究通过酵母双杂交文库筛选,获得MeRLK1蛋白的24个候选互作蛋白,包括Heat shock protein (Hsp)、ADP-ribosylation factor 1 (ARF1)和Aquaporin transported protein (AQP)等。进一步分析MeRLK1与24个候选互作基因的时空表达相关性,结果发现,在不同组织部位和在胁迫条件下Pentatricopeptide repeat 2(PPR2)、BR-signaling kinase (BSK)等基因表达与MeRLK1存在较高的相关性(R>0.7)。在筛选的基础上,构建油菜素内酯(Brassinosteroid,BR)信号通路相关蛋白MeBSK全长序列载体,通过酵母双杂交实验发现,MeRLK1与MeBSK可能存在互作关系,后续需要进一步通过BiFC验证体内互作。Abstract: In order to screen proteins in cassava (Manihot esculenta) that jointly regulate cassava disease resistance with MeRLK1, 24 candidate interacting proteins of the MeRLK1 protein including heat shock protein (Hsp), ADP-ribosylation factor 1 (ARF1), aquaporin transported protein (AQP), etc were selected through screening from the yeast two-hybrid library. Further analysis of the spatiotemporal expression correlation between MeRLK1 and 24 candidate interacting genes revealed that genes including Pentatricopeptide repeat 2 (PPR2) and BR-signaling kinase (BSK) exhibited high correlations (R>0.7) with MeRLK1 expression across diverse tissues and under stress conditions. Based on the screening, the full-length sequence vector of MeBSK, a protein related to the brassinosteroid (BR) signaling pathway was constructed. Through the yeast two-hybrid experiment, it was found that there might be an interaction between MeRLK1 and MeBSK. Subsequently, bimolecular fluorescence complementation (BiFC) will be needed to further verify internal interactions.
-
Key words:
- Manihot esculenta /
- MeRLK1 /
- yeast two-hybrid /
- interaction protein
-
图 3 酵母双杂交筛选MeRLK1候选互作蛋白
注:候选蛋白重组质粒菌液PCR。M,DL2000 DNA Marker;1~48,阳性克隆。
Fig. 3 Y2H screening for MeRLK1-interacting proteins
Note: PCR of the bacterial liquid containing the recombinant plasmid of candidate proteins. M, DL2000 DNA Marker; 1−48, Positive colonies.
图 4 互作候选蛋白功能聚类分析图
Fig. 4 Functional clustering analysis diagram of candidate interacting proteins
图 5 MeRLK1与互作候选基因表达相关性分析
注:R,相关系数。
Fig. 5 Analysis of correlation between MeRLK and candidate-interaction protein gene
Note: R, Correlation coefficient.
图 6 酵母双杂交验证
注:1~4,以MeRLK1-BK+MeBSK-AD酵母质粒为模板扩增产物。
Fig. 6 Yeast two hybrid validation
Note: 1−4, Amplification product using the yeast plasmid MeRLK1-BK+MeBSK-ADas template.
表 1 MeRLK1候选互作蛋白注释
Table 1 Annotation of candidate interacting proteins of MeRLK1
序号
Number基因号
Gene identifier蛋白名
Protein基因注释
Gene annotation亚细胞定位预测
Subcellular localization prediction1 Manes.03G062300 AQP Aquaporin transported Cell membrane 2 Manes.03G046900 NDUFS7 NADH dehydrogenase (ubiquinone) Fe-S protein 7 Mitochondrial matrix 3 Manes.04G003400 CBR2[20] NADH-cytochrome b5 reductase Chloroplast stroma 4 Manes.05G143400 TRX Thioredoxin Nucleus/Cytoplasm 5 Manes.05G145400 E-Syts[21] Extended synaptotagmin-related Cytoplasm/Endoplasmic reticulum 6 Manes.05G155266 CREB[21] cAMP-response element binding prorein related Nucleus 7 Manes.06G134700 Hsp Heat shock protein, mitochondrial-ralated Mitochondrial/Chloroplast 8 Manes.06G151900 PTK Protein tyrosine kinase Chloroplast 9 Manes.07G088100 PPR2 Pentatricopeptide repeat 2 Cytoplasm 10 Manes.09G153400 GME[22] GDP-D-mannose 3′, 5′-epimerase Cytoplasm 11 Manes.07G104200 UBE2C[23] Ubiquitin-conjugating enzyme E2C Nucleus 12 Manes.08G079293 MT Metallothionein Chloroplast 13 Manes.14G112768 TM9SF[24] Transmembrane 9 superfamily member Cell membrane 14 Manes.05G135400 BSK[25] BR-signaling kinase Chloroplast 15 Manes.12G158700 USP Universal stress proteins Chloroplast 16 Manes.09G171400 DAP Dormancy/auxin associated protein Nucleus 17 Manes.13G061700 FLA2 Fasciclin-like arabnogalactan protein 2 Cell membrane 18 Manes.12G098200 UBE2M[26] Ubiquitin-conjugating enzyme E2M Nucleus/Cytoplasm 19 Manes.14G132521 XPR1 Xenotropic and polytropic reterovirus receptor 1 Cytoplasm 20 Manes.15G149500 ARF1 ADP-ribosylation factor 1 Peroxisome 21 Manes.09G060608 ENO Enolase Cytoplasm 22 Manes.18G096100 NEDD8 Ubiquitin-like protein Nucleus/Cytoplasm 23 Manes.16G049900 COX11 Cytochrome c oxidase assembly protein subunit 11 Mitochondrial/Chloroplast 24 Manes.16G078300 SHMT[26] Glycine hydroxymethyltransferase Chloroplast 注:基因号从Phytozome v13数据库中获得。
Note: The gene ID was obtained from Phytozome v13 database. 下载: 导出CSV
下载: 导出CSV
-
[1] 曹升, 陈江枫, 黄富宇, 等. 广西木薯产业现状分析及其发展建议[J]. 南方农业学报, 2021, 52(6): 1468−1476. https://doi.org/10.3969/j.issn.2095-1191.2021.06.005 doi: 10.3969/j.issn.2095-1191.2021.06.005 [2] 付海天, 郑华, 文峰, 等. 中国木薯研究及产业发展趋势 [J]. 农业研究与应用, 2022, 35(4): 9−22. [3] Chisenga S M, Workneh T S, Bultosa G, et al. Progress in research and applications of cassava flour and starch: a review [J]. Journal of Food Science and Technology, 2019, 56(6): 2799−2813. [4] Egbune E O, Ezedom T, Orororo O C, et al. Solid-state fermentation of cassava (Manihot esculenta Crantz): a review [J]. World Journal of Microbiology & Biotechnology, 2023, 39(10): 259. [5] 时涛, 李超萍, 王国芬, 等. 中国木薯病害研究进展与展望[J]. 热带作物学报, 2023, 4(12): 2355−2368. https://doi.org/10.3969/j.issn.1000-2561.2023.12.001 doi: 10.3969/j.issn.1000-2561.2023.12.001 [6] Cui Y, Lu X, Gou X. Receptor-like protein kinases in plant reproduction: Current understanding and future perspectives [J]. Plant Communications, 2022, 3(1): 100273. [7] Sellge G, Kufer T A. PRR-signaling pathways: Learning from microbial tactics [J]. Seminars in Immunology, 2015, 27(2): 75−84. https://doi.org/10.1016/j.smim.2015.03.009 doi: 10.1016/j.smim.2015.03.009 [8] Yuan M, Ngou B P M, Ding P, et al. PTI-ETI crosstalk: an integrative view of plant immunity [J]. Current Opinion in Plant Biology, 2021, 62: 102030. [9] Ngou B P M, Ding P, Jones J D G. Thirty years of resistance: Zig-zag through the plant immune system [J]. The Plant Cell, 2022, 34(5): 1447−1478. [10] Dievart A, Gottin C, Périn C, et al. Origin and diversity of plant receptor-like kinases [J]. Annual Review of Plant Biology, 2020, 71: 131−156. [11] Wei Z, Li J. Receptor-like protein kinases: key regulators controlling root hair development in Arabidopsis thaliana [J]. Journal of Integrative Plant Biology, 2018, 60(9): 841−850. [12] Shiu S H, Karlowski W M, Pan R, et al. Comparative analysis of the receptor-like kinase family in Arabidopsis and rice [J]. The Plant Cell, 2004, 16(5): 1220−1234. [13] Shiu S H, Bleecker A B. Receptor-like kinases from Arabidopsis form a monophyletic gene family related to animal receptor kinases [J]. Proceedings of the National Academy of Sciences of the United States of America, 2001, 98(19): 10763−10768. [14] Lehti-Shiu M D, Zou C, Hanada K, et al. Evolutionary history and stress regulation of plant receptor-like kinase/pelle genes [J]. Plant Physiology, 2009, 150(1): 12−26. https://doi.org/10.1104/pp.108.134353 doi: 10.1104/pp.108.134353 [15] Qu J, Dry I, Liu L, et al. Transcriptional profiling reveals multiple defense responses in downy mildew-resistant transgenic grapevine expressing a TIR-NBS-LRR gene located at the MrRUN1/MrRPV1 locus [J]. Horticulture Research, 2021, 8(1): 161. https://doi.org/10.1038/s41438-021-00597-w doi: 10.1038/s41438-021-00597-w [16] 周萌萌. MeAHL17转录因子调节木薯对细菌性枯萎病抗性分子机理研究[D]. 海口: 海南大学, 2023. [17] Yuan J J, Zhao Y N, Yu S H, et al. The Arabidopsis receptor-like kinase WAKL4 limits cadmium uptake via phosphorylation and degradation of NRAMP1 transporter [J]. Nature Communications, 2024, 15(1): 9537. [18] Kang H, Huang T, Duan G, et al. TCOD: an integrated resource for tropical crops [J]. Nucleic Acids Research, 2024, 52(D1): D1651−D1660. https://doi.org/10.1093/nar/gkad870 doi: 10.1093/nar/gkad870 [19] Eggleston A K, Kowalczykowski S C. An overview of homologous pairing and DNA strand exchange proteins [J]. Biochimie, 1991, 73(2/3): 163−176. [20] Chen J, Sun Y, Chen L, et al. NADH-Cytochrome B5 reductase 2 suppresses retinal vascular dysfunction through regulation of vascular endothelial growth factor A in diabetic retinopathy [J]. Experimental Eye Research, 2022, 222: 109186. https://doi.org/10.1016/j.exer.2022.109186 doi: 10.1016/j.exer.2022.109186 [21] Zheng W, Guo J, Lu X, et al. cAMP-response element binding protein mediates podocyte injury in diabetic nephropathy by targeting lncRNA DLX6-AS1 [J]. Metabolism, 2022, 129: 155155. https://doi.org/10.1016/j.metabol.2022.155155 doi: 10.1016/j.metabol.2022.155155 [22] Gilbert L, Alhagdow M, Nunes-Nesi A, et al. GDP-D-mannose 3,5-epimerase (GME) plays a key role at the intersection of ascorbate and non-cellulosic cell-wall biosynthesis in tomato [J]. The Plant Journal, 2009, 60(3): 499−508. https://doi.org/10.1111/j.1365-313X.2009.03972.x doi: 10.1111/j.1365-313X.2009.03972.x [23] Xie C, Powell C, Yao M, et al. Ubiquitin-conjugating enzyme E2C: a potential cancer biomarker [J]. The International Journal of Biochemistry & Cell Biology, 2014, 47: 113−117. [24] Yu Y, Song Y, Cheng L, et al. CircCEMIP promotes anoikis-resistance by enhancing protective autophagy in prostate cancer cells [J]. Journal of Experimental & Clinical Cancer Research, 2022, 41(1): 188. [25] Zhang B, Wang X, Zhao Z, et al. OsBRI1 activates BR signaling by preventing binding between the TPR and kinase domains of OsBSK3 via phosphorylation [J]. Plant Physiology, 2016, 170(2): 1149−1161. https://doi.org/10.1104/pp.15.01668 doi: 10.1104/pp.15.01668 [26] Lu X, Kong X, Wu H, et al. UBE2M-mediated neddylation of TRIM21 regulates obesity-induced inflammation and metabolic disorders [J]. Cell Metabolism, 2023, 35(8): 1390−1405. https://doi.org/10.1016/j.cmet.2023.05.011 doi: 10.1016/j.cmet.2023.05.011 [27] Richard M M S, Gratias A, Alvarez Diaz J C, et al. A common bean truncated CRINKLY4 kinase controls gene-for-gene resistance to the fungus Colletotrichum lindemuthianum [J]. Journal of Experimental Botany, 2021, 72(10): 3569−3581. https://doi.org/10.1093/jxb/erab082 doi: 10.1093/jxb/erab082 [28] Xun Q, Wu Y, Li H, et al. Two receptor-like protein kinases, MUSTACHES and MUSTACHES-LIKE, regulate lateral root development in Arabidopsis thaliana [J]. New Phytologist, 2020, 227(4): 1157−1173. https://doi.org/10.1111/nph.16599 doi: 10.1111/nph.16599 [29] Wang J, Wang J, Li J, et al. The RLK protein TaCRK10 activates wheat high-temperature seedling-plant resistance to stripe rust through interacting with TaH2A. 1 [J]. The Plant Journal, 2021, 108(5): 1241−1255. https://doi.org/10.1111/tpj.15513 doi: 10.1111/tpj.15513 [30] Haruta M, Sabat G, Stecker K, et al. A peptide hormone and its receptor protein kinase regulate plant cell expansion [J]. Science, 2014, 343(6169): 408−411. https://doi.org/10.1126/science.1244454 doi: 10.1126/science.1244454 [31] Li L, Verstraeten I, Roosjen M, et al. Cell surface and intracellular auxin signalling for H+ fluxes in root growth [J]. Nature, 2021, 599(7884): 273−277. https://doi.org/10.1038/s41586-021-04037-6 doi: 10.1038/s41586-021-04037-6 [32] Chen X, Wang T, Rehman A U, et al. Arabidopsis U-box E3 ubiquitin ligase PUB11 negatively regulates drought tolerance by degrading the receptor-like protein kinases LRR1 and KIN7 [J]. Journal of Integrative Plant Biology, 2021, 63(3): 494−509. https://doi.org/10.1111/jipb.13058 doi: 10.1111/jipb.13058 [33] Su B, Zhang X, Li L, et al. Dynamic spatial reorganization of BSK1 complexes in the plasma membrane underpins signal-specific activation for growth and immunity [J]. Molecular Plant, 2021, 14(4): 588−603. https://doi.org/10.1016/j.molp.2021.01.019 doi: 10.1016/j.molp.2021.01.019 [34] Zhang S, Hu X, Dong J, et al. Identification, evolution, and expression analysis of OsBSK gene family in Oryza sativa Japonica [J]. BMC Plant Biology, 2022, 22(1): 565. https://doi.org/10.1186/s12870-022-03905-1 doi: 10.1186/s12870-022-03905-1 [35] Zada A, Lv M, Li J. Molecular lesions in BRI1 and its orthologs in the plant Kingdom [J]. International Journal of Molecular Sciences, 2024, 25(15): 8111. https://doi.org/10.3390/ijms25158111 doi: 10.3390/ijms25158111 -

 点击查看大图
点击查看大图
计量
- 文章访问数: 1174
- HTML全文浏览量: 647
- PDF下载量: 79
- 被引次数: 0
