

-
油菜素内酯(brassinosteroid,BR)是一种植物生长调节剂,在植物中广泛分布,可参与多种细胞信号传导过程[1]。BR最早在油菜(Brassica napus)花粉中被分离并于1979年鉴定出分子结构和命名[2-3],BR及其衍生物能调节植物生长和发育活动,包括细胞伸长和分裂、光形态发生、木质部分化、繁殖以及对逆境胁迫的反应[4]。越来越多研究证明,油菜素内酯在调控植物抗逆性方面具有重要的积极作用,BR可以促进植物在非生物胁迫下的种子发芽率,外源施加BR提升了高粱种子在渗透胁迫下的种子发芽率和幼苗存活率[5],在盐胁迫下,外源施加BR能提高黄瓜(Cucumis sativus)和水稻(Oryza sativa)种子的发芽率[6-7]。BR还可以促使植物的根系生长,在盐胁迫下BR可以促进马铃薯(Solanum tuberosum)根的生长[8]。在干旱胁迫下,BR可以正向调节烟草(Nicotiana tabacum)幼苗根系生长和发育[9]。因此,BR是否也可以提高橡胶草(Taraxacum kok-saghyz)抗逆能力?BR提高抗逆能力的分子机制有哪些?这些问题的深入研究对于完善橡胶树(Hevea brasiliensis)的抗逆应答研究具有重要的参考价值。当BR被胞膜上的受体激酶(brassinosteroid insensitive1,BRI1)的外部结构域识别后,会促使下游发生各种信号传递。这种结合使BRI1被激活,随后触发了信号转导级联反应,最终导致下游基因的转录调控[10]。 BRI1 激活涉及共受体激酶 BRI1 相关激酶 1(BAK1)的募集,BRI1 与 BAK1 在胞外的结合导致 BRI1 激酶抑制剂 1(BKI1)从细胞质膜解离,进一步通过相互磷酸化使 BRI1 和 BAK1 的激酶结构域激活[11-12]。激活后 BRI1 通过磷酸化激活 BR 信号激酶(BSK)和组成型差异生长1(CDG1)家族[13]。然后,BSK 和 CDG1 激活PP1型磷酸酶Brisuppressor1(BSU1),随后,使Brassinosteroid insensitive2(BIN2)去磷酸化并失活[14]。BSU1 能使失活的BIN2去磷酸化,进 而消除BIN2对 Brassinazole-resistant1(BZR1)和Bri1-ems-suppressor1(BES1)的磷酸化抑制 [15-16]。
BZR/BES是植物特异性转录因子和介导BR信号传导的关键调控因子[11],BZR/BES成员N端均有1个bHLH结构域, 它们能够直接与靶基因启动子区的E-box或BRRE元件结合从而调控这些基因的表达[17-18]。这2个转录因子在氨基酸水平上有88%的同源性,特别是N端结构域上,同源性高达91%。此外,BZRs有可以被各种激酶磷酸化的富集位点[15-16]。BZR1和BES1在功能上存在一些差别。BZR1与BR的响应元件(CGTGT/CG 基序)结合,从而抑制了有关BR生物合成基因的表达,而BES1能够与 E-box(CANNTG 序列)结合,从而激活BR诱导的基因表达[16]。迄今为止, 已经在模式植物拟南芥( Arabidopsis thaliana)和主要粮食作物水稻中完成了BZR1/BES1基因家族的全面鉴定,这些基因家族的鉴定和克隆工作也在逐步进行中[7, 17]。
橡胶草是一种多年生草本植物,主要分布在中国西北(新疆)和华东等地区[19]。因其产胶性质和质量与橡胶树相当,成为研究产胶机制的模式植物[20]。橡胶草根部可以合成高分子质量的天然橡胶和其他化学物质,并因其具有生命力强、适应环境变化广泛、转基因遗传转化体系较成熟等优势, 被认为是一种有潜力替代传统橡胶生产的经济作物,成为产胶研究的模式植物[21]。但是由于各种植地环境因素影响,橡胶草的种植和生产常受到干旱、高温、盐渍等胁迫的影响,严重阻碍了橡胶草的生长发育,限制了橡胶草产业的发展。但目前关于橡胶草抗逆分子机制的相关报道还较少,因此,对橡胶草抗逆基因的鉴定和功能研究具有重要意义[22]。为了探究BZR/BES基因在植物中特别是橡胶草的生长发育和抵抗逆境中的重要作用,本研究鉴定了橡胶草中BZR/BES家族基因成员,构建系统进化树并系统地分析了其在染色体上的分布、结构域特征及表达模式。从橡胶草中成功克隆到表达丰度最高的1个TkBZR基因TkBZR2,并在大肠杆菌(Escherichia coli )BL21 (DE3)中成功表达TkBZR2重组蛋白。经过对TkBZR2基因启动子区域的分析,发现了激素响应元件和胁迫响应元件。这一发现不仅为后续TkBZR基因家族的深入研究打下了基础,而且为橡胶草的抗逆性研究提供了理论支持。
-
本研究所使用的橡胶草采自新疆伊宁野外(42°51′36″ N, 80°43′48″ E),取植株叶片,经消毒后在无菌环境下,将其切割成小块组织,置于含有吲哚乙酸和细胞分裂素的MS培养基上,通过组织培养扩繁得到无菌植株,移栽至温度约24 ℃、湿度约57%的培养间,光照周期为每天光照16 h加黑暗8 h,按常规栽培措施进行管理。
本研究所使用的化学试剂、RNA提取试剂盒和质粒提取试剂盒均购于北京天根生物有限公司,构建载体使用的Ready-to-Use Seamless Cloning Kit购于BBI生命科学有限公司。PrimeSTAR Max DNA聚合酶、pMD-19T载体购买于Takara生物公司,pET28a载体大肠杆菌DH5α、DE3感受态细胞为本实验室留存。
-
在基因组数据库(GWH;
http://bigd.big.ac.cn/gwh/ )中下载橡胶草的全基因组文件PRJCA000437,并使用BioEdit软件建立本地库。从TAIR网站(http: //www.arabidopsis.org/ )获取拟南芥已知的AtBZR氨基酸序列,通过本地BLAST将P<1e−10的序列筛选出BZR/BES基因家族候选成员。将候选基因序列利用InterproScan(https://www.ebi.ac.uk/interpro/search/sequence/ )和SMART(https://smart.embl.de/ )检测是否具有BES1_N结构域,从而得到橡胶草BZR基因家族成员。将橡胶草BZR基因家族成员的氨基酸序列上传至 ExPASy数据库(http://web.expasy.org/protparam/ )计算蛋白质的物理参数并预测蛋白的分子质量、等电点和亲疏水性。还利用在线网站Plant-mPLoc(http://www.csbio.sjtu.edu.cn/bioinf/plant-multi/ )对TkBZRs蛋白的亚细胞定位进行预测。 -
通过NCBI数据库(
https://www.ncbi.nlm.nih.gov/ )下载巴西橡胶树、拟南芥和莴苣(Lactuca sativa)中已被鉴定的BZR家族基因序列。通过 MEGA v6.0 的邻接法(Neighbor-Joining)将Bootstrap 值设定为1 000,创建橡胶草BZR蛋白序列的系统发育树,并利用AtBZR、LsBZR、HbBZR和NtBZR构建多物种进化树。 -
根据基因组注释文件将橡胶草BZR/BES基因家族成员在染色体上的物理位置利用在线工具MapGene2Choromosome v2.0(
http://mg2c.iask.in/mg2c_v2.0/ )进行可视化展示。橡胶草BZR蛋白的保守基序利用MEME(http://meme-suite.org/tools/meme )网站进行分析,并使用TBtools软件分析橡胶草BZR基因家族基因结构(内含子–外显子)。 -
将橡胶草全生育期划分为S1~S9[23],将无性繁殖而来的生根至3~5 cm的橡胶草移入组培瓶中生长30 d为S1,组培60 d为S2,组培90 d为S3, 然后移栽土培,土培30 d为S4,土培60 d为S5,土培90 d为S6,土培120 d为S7,土培150 d为S8, 土培180 d为S9。其中,S5 时期进行30 d的春化处理,S6 时期为盛花期,S7 时期为种子成熟期,S8时期为衰老期,S9 时期为休眠期。课题组基于前期获得的转录组数据,对橡胶草根、叶、胶乳、花及花莛等5个组织中的TkBZR成员表达水平(FPKM值)进行系统分析,同时整合全生育期S1~S9阶段叶、根、胶乳组织的转录组数据。通过TBtools软件对上述FPKM值进行log2标准化转换,最终构建了TkBZRs在组织特异性和发育时序维度上的差异表达热图。
-
收集约100 mg的橡胶草叶片组织用液氮低温研磨后,按照RNA提取试剂盒说明书进行RNA的提取,得到的RNA经微量分光光度计检测其浓度后,使用反转录试剂盒,根据其说明书进行反转录,得到橡胶草cDNA后进行后续实验。
通过对橡胶草转录组数据分析后得出BZR/BES家族差异表达最显著的TkBZR2序列作为参考序列,利用Primer v6.0软件设计特异性引物并以橡胶草cDNA为模板扩增TkBZR2。并连接pMD19-T Vector并转化大肠杆菌感受态,挑取抗性筛选培养基上的阳性单克隆进行PCR检测后交由擎科生物公司测序。
-
利用Primer v6.0软件设计基因引物,根据TkBZR2基因设计引物(表1)进行PCR片段扩增。PCR产物回收纯化后,将TkBZR2通过同源重组法构建到pET28a(+)载体上(总体积为10 μL、2×Seamless Cloning Master Mix 5 μL、线性化载体1 μL、带有同源臂的基因片段4 μL,PCR仪中50 ℃反应20 min)并转入大肠杆菌DE3感受态细胞,加入不同浓度的IPTG(0.01、0.10、0.50、1.00 mmol·L−1),分别在28、37 ℃诱导0、2、4、6、8 h。
表 1 引物序列
Table 1. Primer sequences
引物名称 Primer 序列 Primer sequence(5′−3′) TkBZR2-F ATGACCGGCGCCGGTTCT TkBZR2-R TTAAATTACACTTTTCCC pET28a-TkBZR2-F caaatgggtcgcggaATGACCGGCGCC pET28a-TkBZR2-R gagctcgaattcggagaAATTACACTTT
TCCCACTACCAAG -
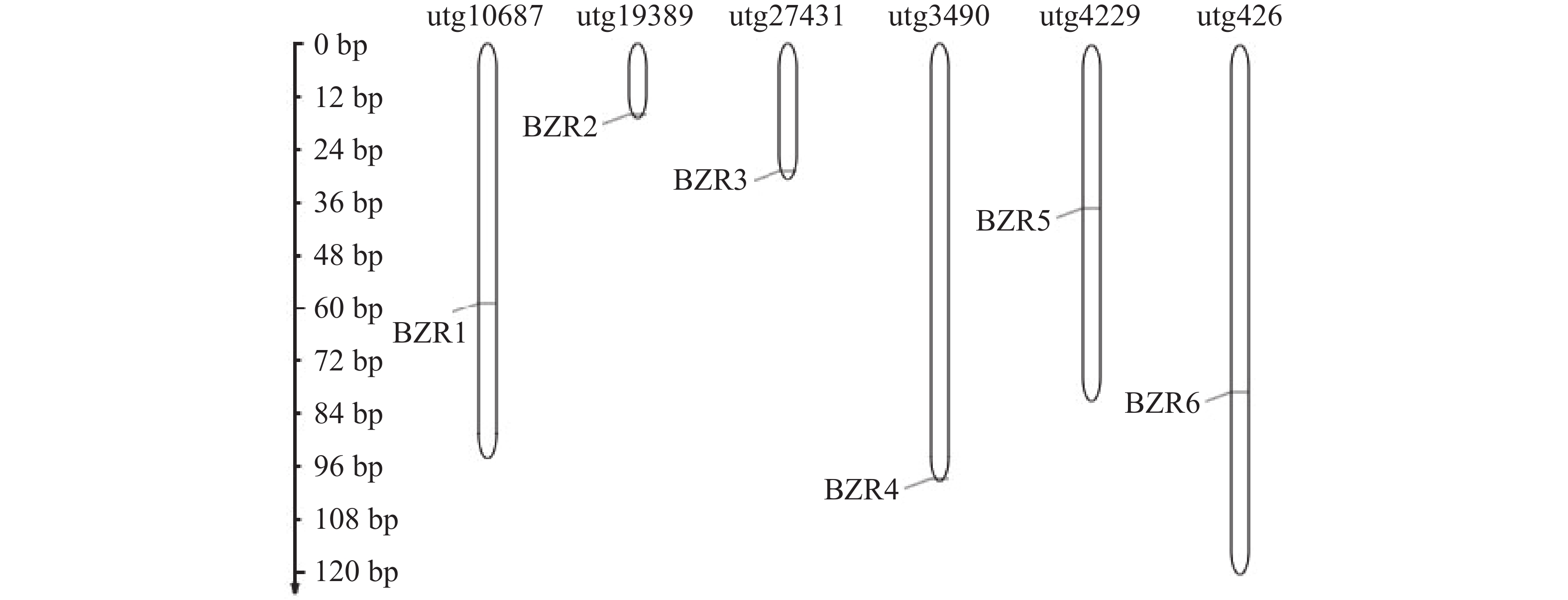
为了了解TkBZR/BES家族成员的情况,本研究利用软件HMMER v3.0的hmmersearch功能将BZR的保守结构域在橡胶草基因组文件PRJCA000437进行搜索,将阈值P<1e−10的序列作为TkBZR可能存在的成员。通过对候选成员进行结构域分析,初步获得9个TkBZR/BES家族基因,使用InterproScan和SMART数据库进一步分析候选基因,去除不完整序列。最终鉴定出6个TkBZR/BES家族基因,并根据其分子质量大小命名TkBZR1~TkBZR6。从图1可知,这6个基因分布于6个独立的基因组片段(scaffold)上。TkBZR/BES家族的氨基酸长度为151~316 aa,理论等电点最低为7.63,最高为9.38,表明绝大多数TkBZR/BES家族蛋白是碱性蛋白,亲水性系数均小于0,这说明TkBZR/BES家族蛋白均为亲水性蛋白(表2)。
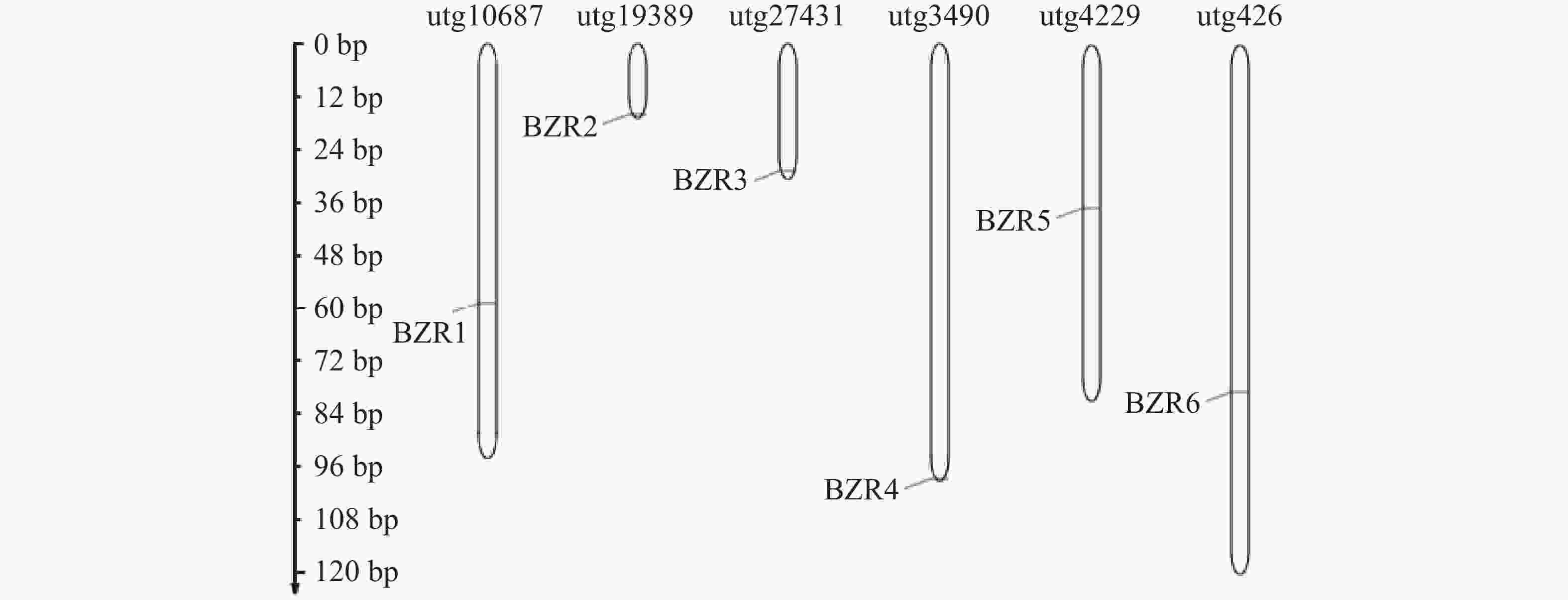
图 1 TkBZR基因家族在橡胶草染色体上的基因分布
Figure 1. Gene distribution of the TkBZR gene family on Taraxacum kok-saghyz chromosomes
表 2 橡胶草基因组BZR/BES基因家族信息
Table 2. The information of BZR/BES gene family in Taraxacum kok-saghyz genome
基因名称
Gene基因编号
Gene ID染色体定位
Position氨基酸数目/aa
Length of
amino acid分子质量/kDa
Molecular
weight等电点
pI亲水性系数
Hydropathy
Index亚细胞定位
Subcellular
locationBZR1 GWHPAAAA046609 utg9916 316 34.35 9.38 −0.705 细胞核Nucleus BZR2 GWHPAAAA015365 utg19389 307 33.30 8.92 −0.589 细胞核Nucleus BZR3 GWHPAAAA030593 utg3490 302 32.99 8.45 −0.753 细胞核Nucleus BZR4 GWHPAAAA024338 utg27431 294 32.13 7.63 −0.711 细胞质Cytoplasm BZR5 GWHPAAAA045865 utg950 263 28.39 8.66 −0.572 细胞质与细胞核
Cytoplasm and NucleusBZR6 GWHPAAAA001420 utg10687 151 17.13 9.13 −0.909 细胞核Nucleus -
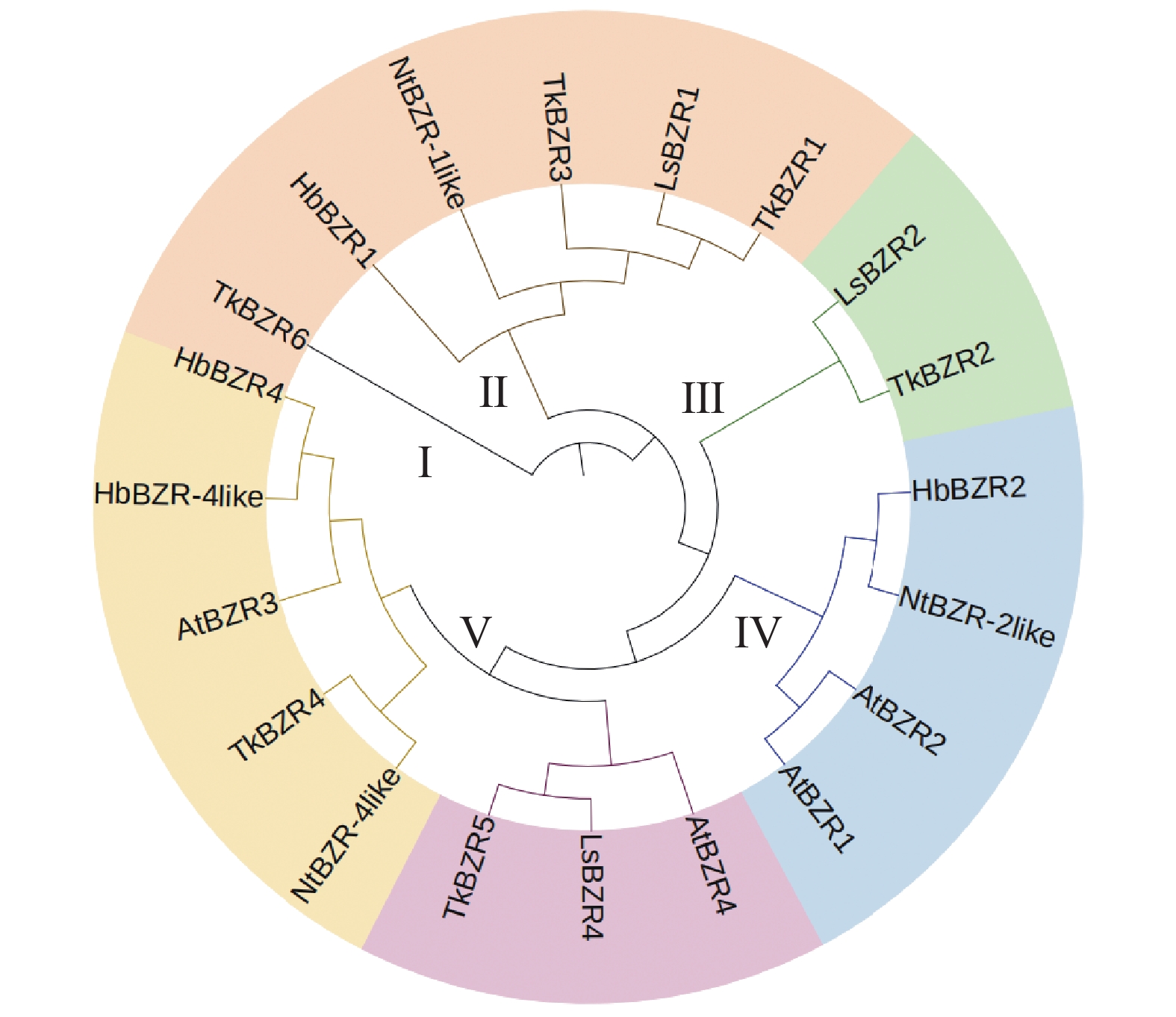
基于邻接法对橡胶草、橡胶树、莴苣、烟草和拟南芥的20个TkBZR家族成员进行1 000次bootstrap统计学检验得到其蛋白系统发育关系。如图2所示,BZR蛋白被分成了5个(Ⅰ~Ⅴ)亚组。其中,橡胶草TkBZR/BES家族基因分布在Ⅰ、Ⅱ、Ⅲ、Ⅴ等4个亚组,与莴苣具有相似的分布模式;橡胶草与莴苣的LsBZR家族基因聚类在同一亚组的成员更多,这表明橡胶草蛋白与莴苣的系统发育关系较为密切。TkBZR1大部分位于Ⅴ和Ⅱ亚组,其他4个物种的TkBZR/BES基因在这2个亚组也有分布。
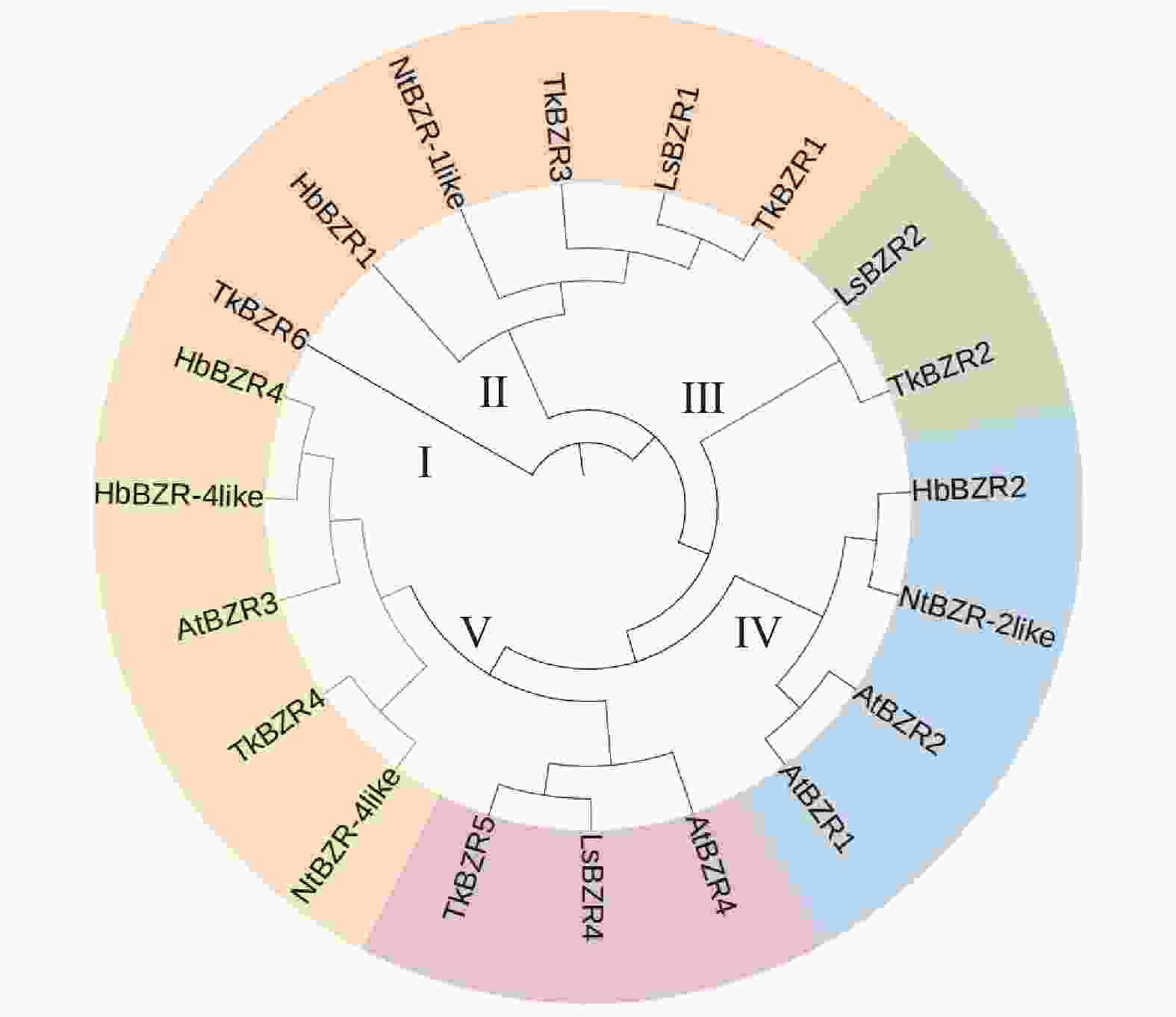
图 2 橡胶草和其他植物BZR基因家族进化树
Figure 2. Phylogenetic tree of BZR gene families in Taraxacum kok-saghyz and other plants
通过上述分析,相同类型的BZR蛋白在理化性质方面相似程度较高,且系统发育关系较为密切的蛋白之间理化性质更为相似。对进化树进行比较后发现,在Ⅱ、Ⅲ、Ⅴ亚组中莴苣和橡胶草BZR基因处于同一分支,说明莴苣和橡胶草的BZR基因家族在进化过程中有较强的亲缘关系。
-
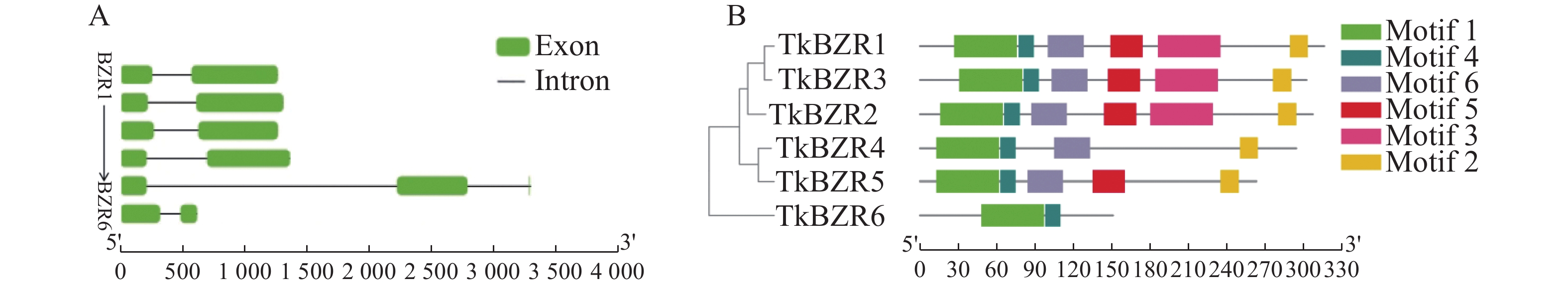
TkBZR家族基因成员除TkBZR5外,均含有2个外显子,不同的TkBZR之间在保守结构基序和基因结构上的差异可能是影响其亚细胞定位和功能的重要原因。将TkBZR家族蛋白序列输入到MEME网站上进行在线分析,并利用TBtools将其与进化树结合和美化。根据保守基序分析,TkBZR蛋白中鉴定出6个保守基序,TkBZR1、TkBZR2和TkBZR3含有全部的motif,而TkBZR6只含有motif 1和motif 2,motif1和motif2与TkBZR家族的BES1_N保守结构域所在位置一致(表3),以上结果说明TkBZR家族结构高度相似,它们在基因进化过程和功能模式上可能具有保守性(图3)。
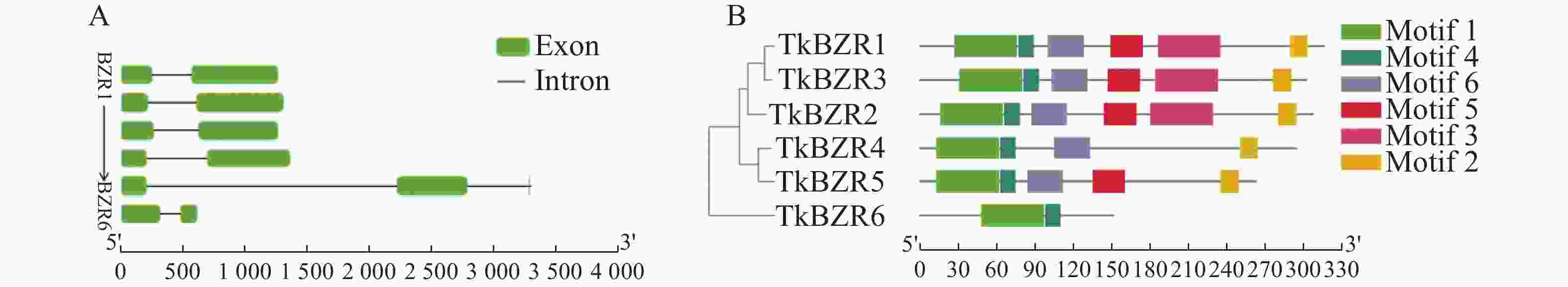
图 3 TkBZR家族的基因结构和基序分析
Figure 3. Gene structure and motif analysis of TkBZR family
表 3 Motif序列信息
Table 3. Sequence information of motifs
名称 Motif 序列 Motif sequence Motif1 YAGLRAQGNYKLPKHCDNNEVLKALCEEAGWVV Motif2 KAWEGERIHEVGVDD Motif3 FTKQSIPSFNLPFFASSAPTSPTRRHRFTPATIPECDESDCSTIDSCQWV Motif4 EPDGTTYRKGCKP Motif5 NPSSLPPLRISNSAPVTPPLSSPTSK Motif6 STNTSPCSSQKPSPISSSFPSPSPSDQAS -
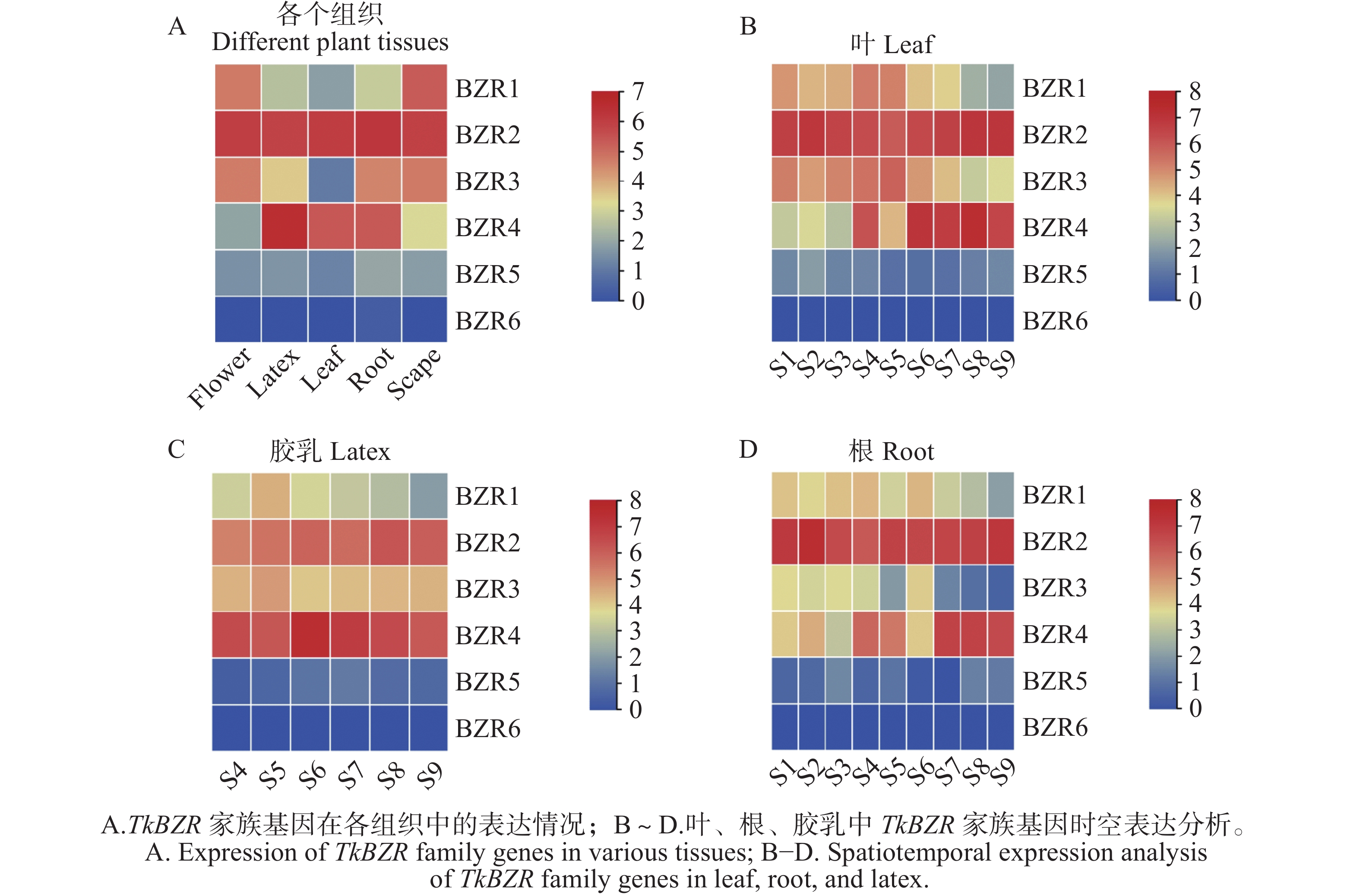
通过对橡胶草不同组织的转录组数据进行分析,推测TkBZRs家族基因在根、叶、茎、胶乳等不同组织中的表达模式。如图4所示,TkBZRs具有组织特异性,除TkBZR6几乎在每个组织中都不表达外,TkBZRs在不同组织中表达量均有显著差异。TkBZR4在橡胶草S6生育期的胶乳组织中表达量达到最高。TkBZR1~TkBZR5在根、茎、叶中均呈现先上升后下降的趋势,但达到峰值的时期并不相同,这暗示TkBZR基因家族成员行使相似的功能,但由于蛋白质序列的内部差异使TkBZRs基因在功能上既有相似性又有差异性。
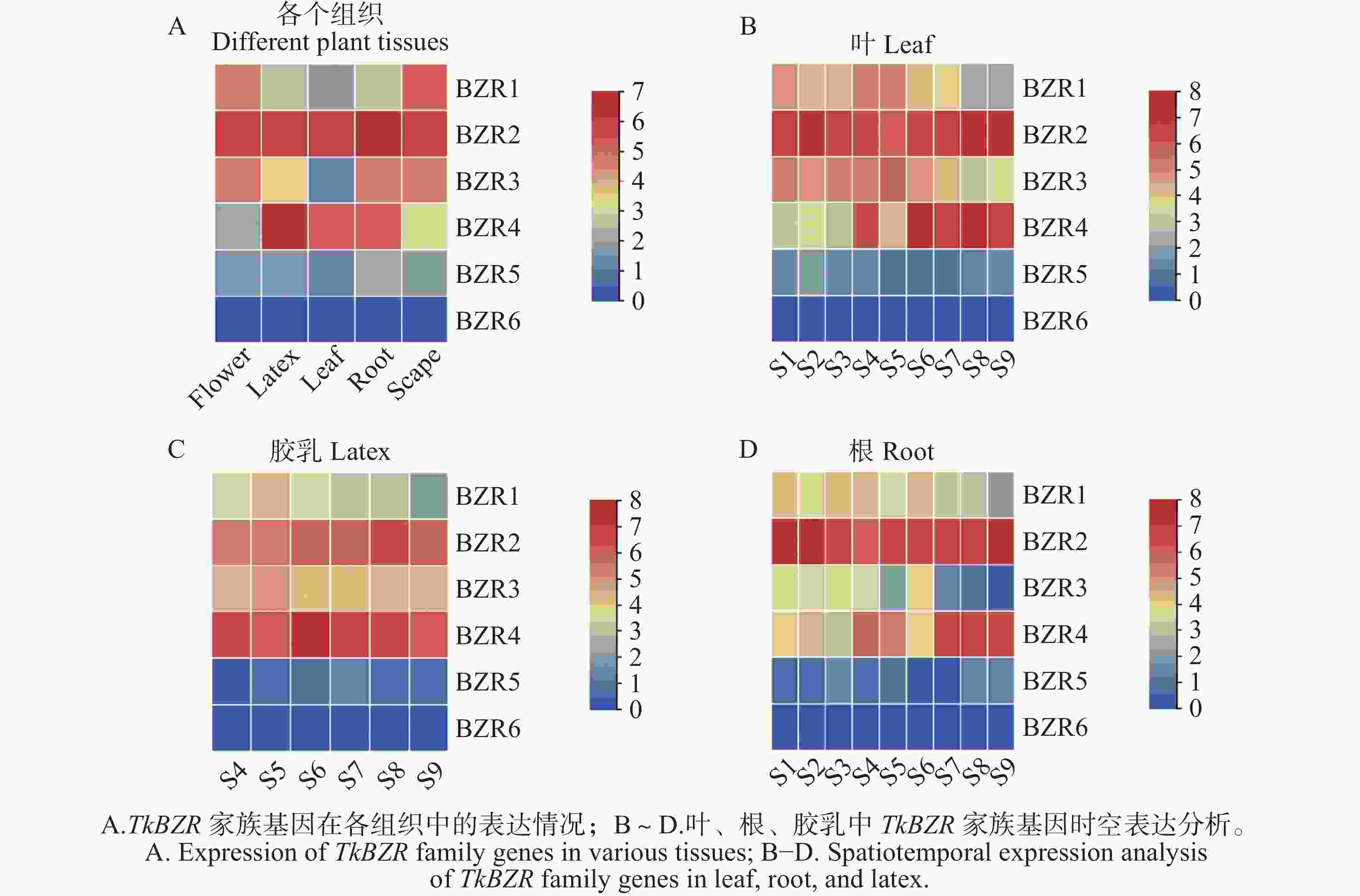
图 4 TkBZR表达模式分析
Figure 4. Analysis of TkBZR expression patterns
-
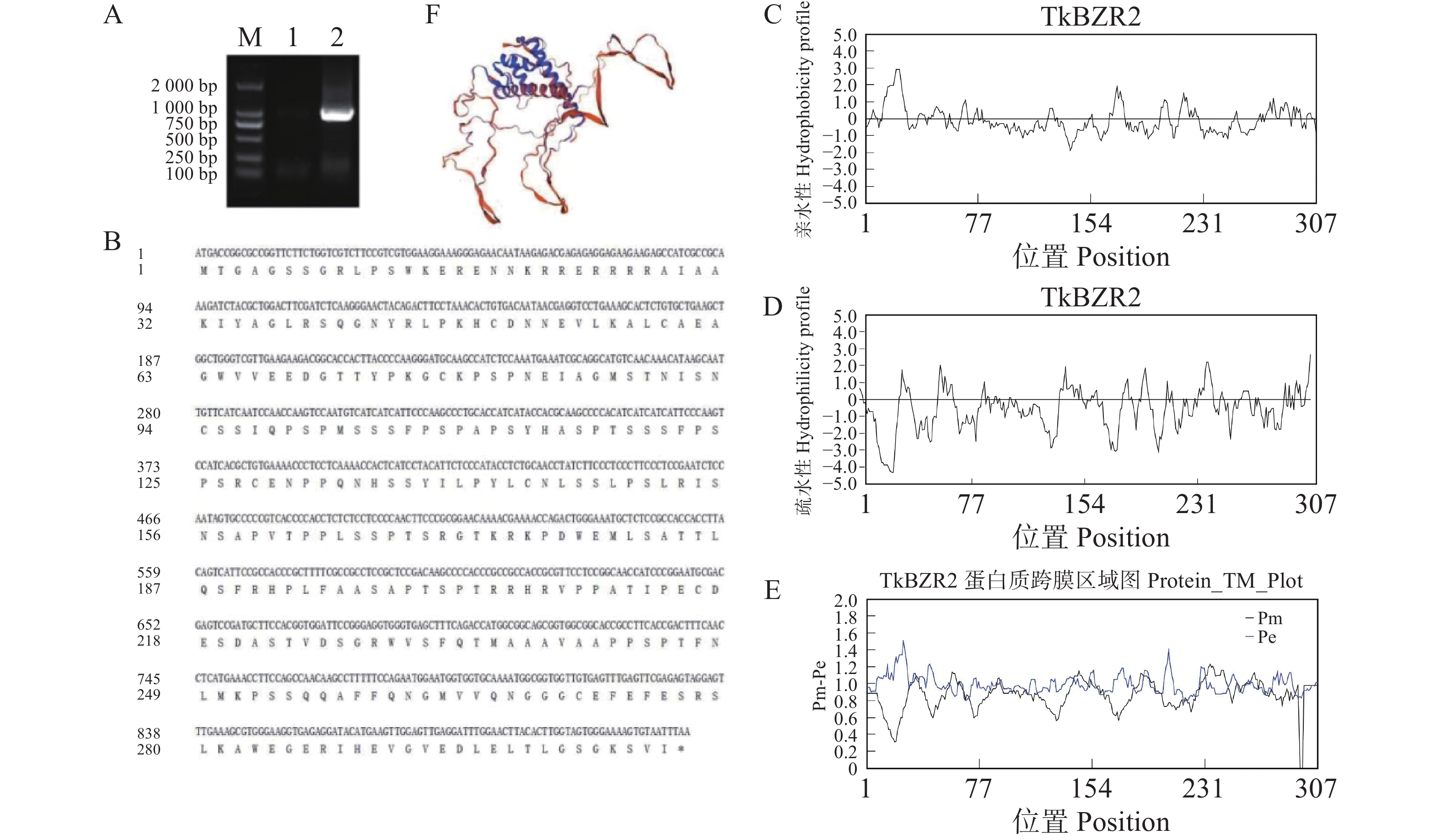
为了进一步探究TkBZR家族基因的功能,本研究克隆了在全生育期表达丰度最高的TkBZR2,利用软件Primer v6.0设计引物,以橡胶草叶片cDNA为模板克隆得到了1条长为924 bp的片段,且与目的条带大小相符。将克隆得到的基因片段纯化回收后重组到pMD-19T载体上,挑取单克隆进行菌落PCR然后进行测序。然后在NCBI网站上运用BLAST工具对测序结果进行比对,结果发现,所得基因片段与预期的目标基因序列完全一致。使用DNAMAN将TkBZR2氨基酸序列与其他物种中的BZR氨基酸序列进行比,并预测其蛋白质的亲疏水性(图5)。
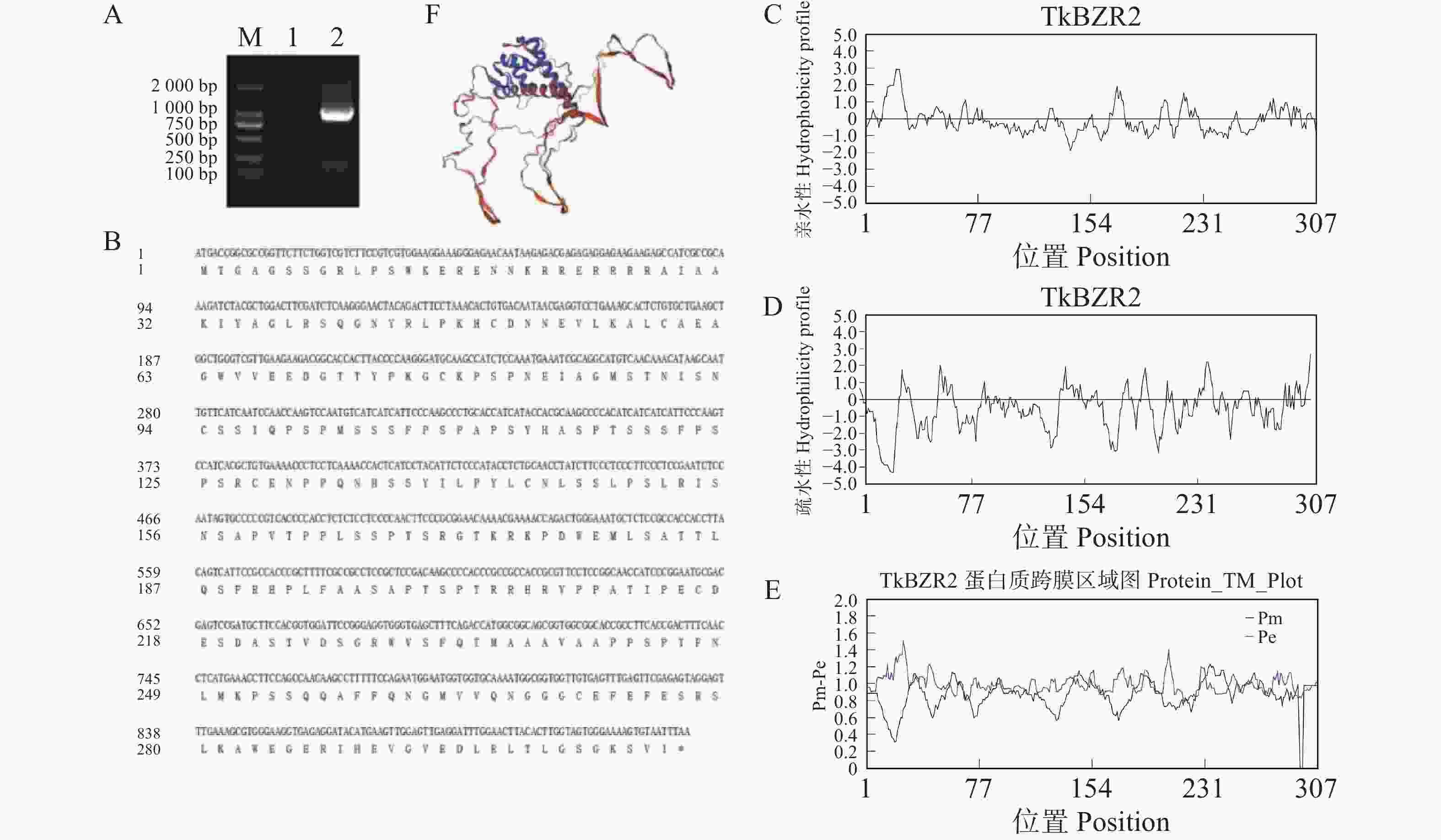
图 5 TkBZR2基因的扩增与分析
Figure 5. Amplification and analysis of TkBZR2 gene
-
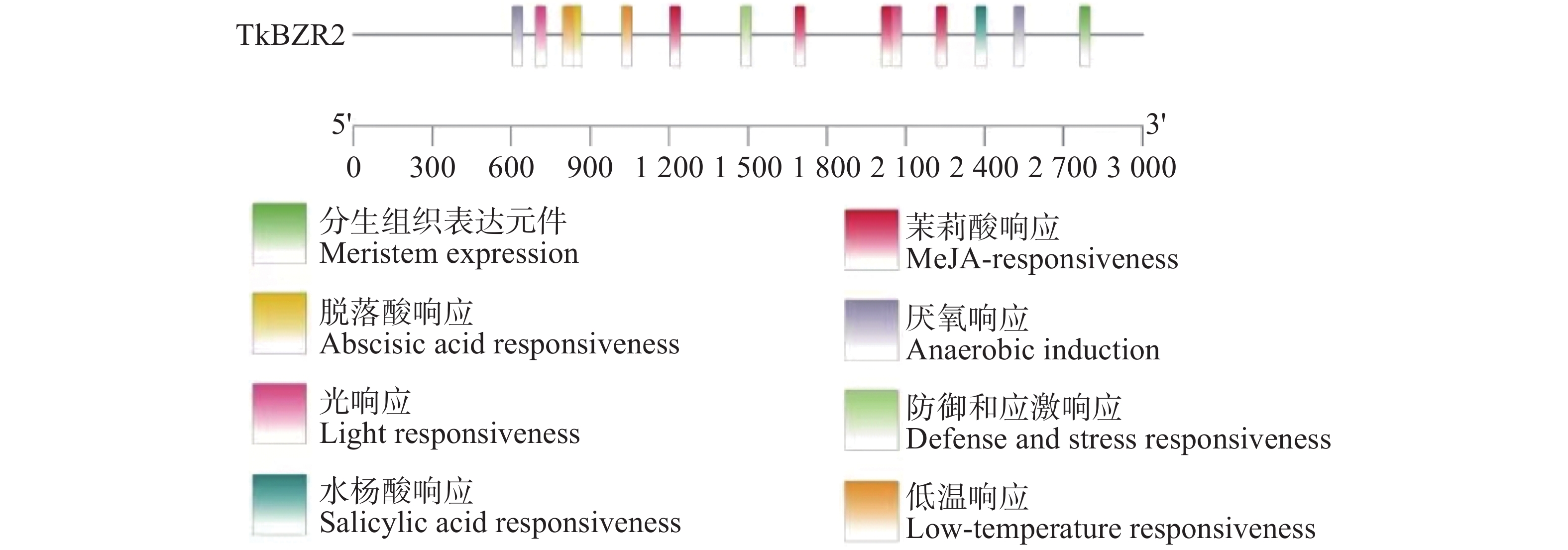
启动子顺式元件与基因功能密切相关,为了进一步探究TkBZR2的调控机制,找到能够调控TkBZR2的上游基因。如图6所示,本研究提取TkBZR2起始密码子前2 000 bp作为TkBZR2的启动子进行分析,将数据上传至PlantCARE网站进行顺式作用元件进行可视化分析,结果如图6所示。从中可知,TkBZR2基因的启动子区域含有多种激素响应元件、非生物胁迫响应元件等,这表明TkBZR2基因可能受到多种激素的调控,并能够对多种胁迫条件做出响应。
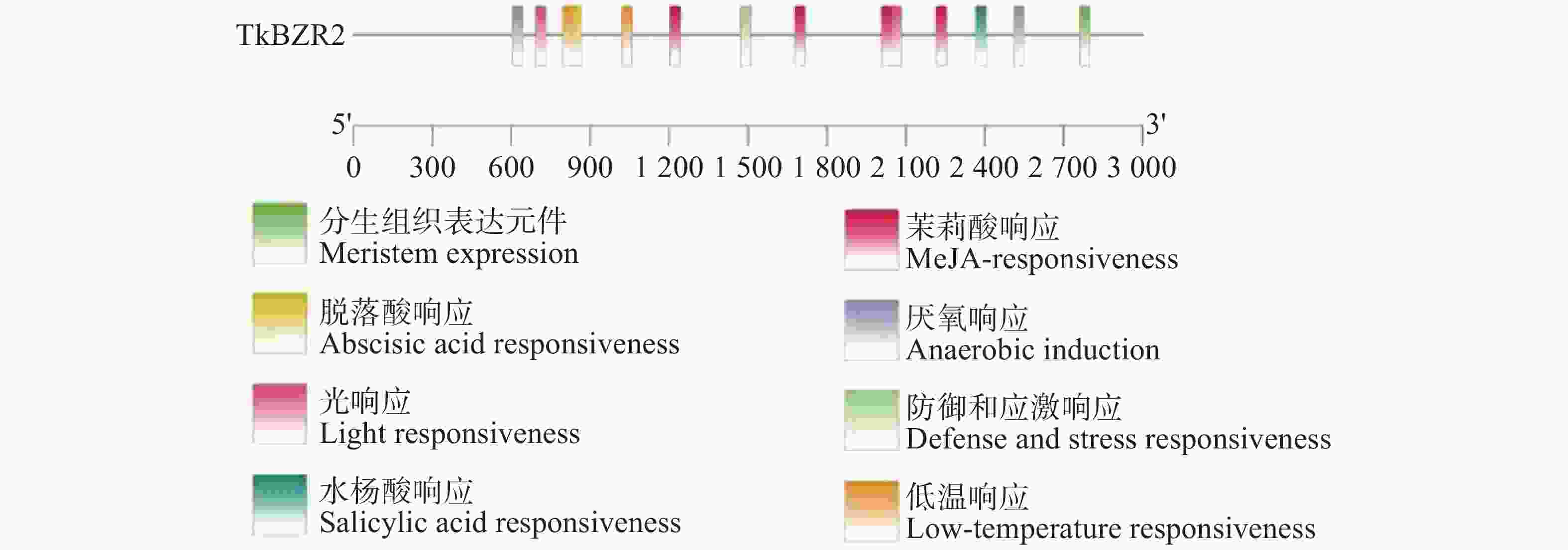
图 6 橡胶草BZR2启动子顺式作用元件分析
Figure 6. Analysis of cis-acting elements of BZR2 promoter of Taraxacum kok-saghyz
-
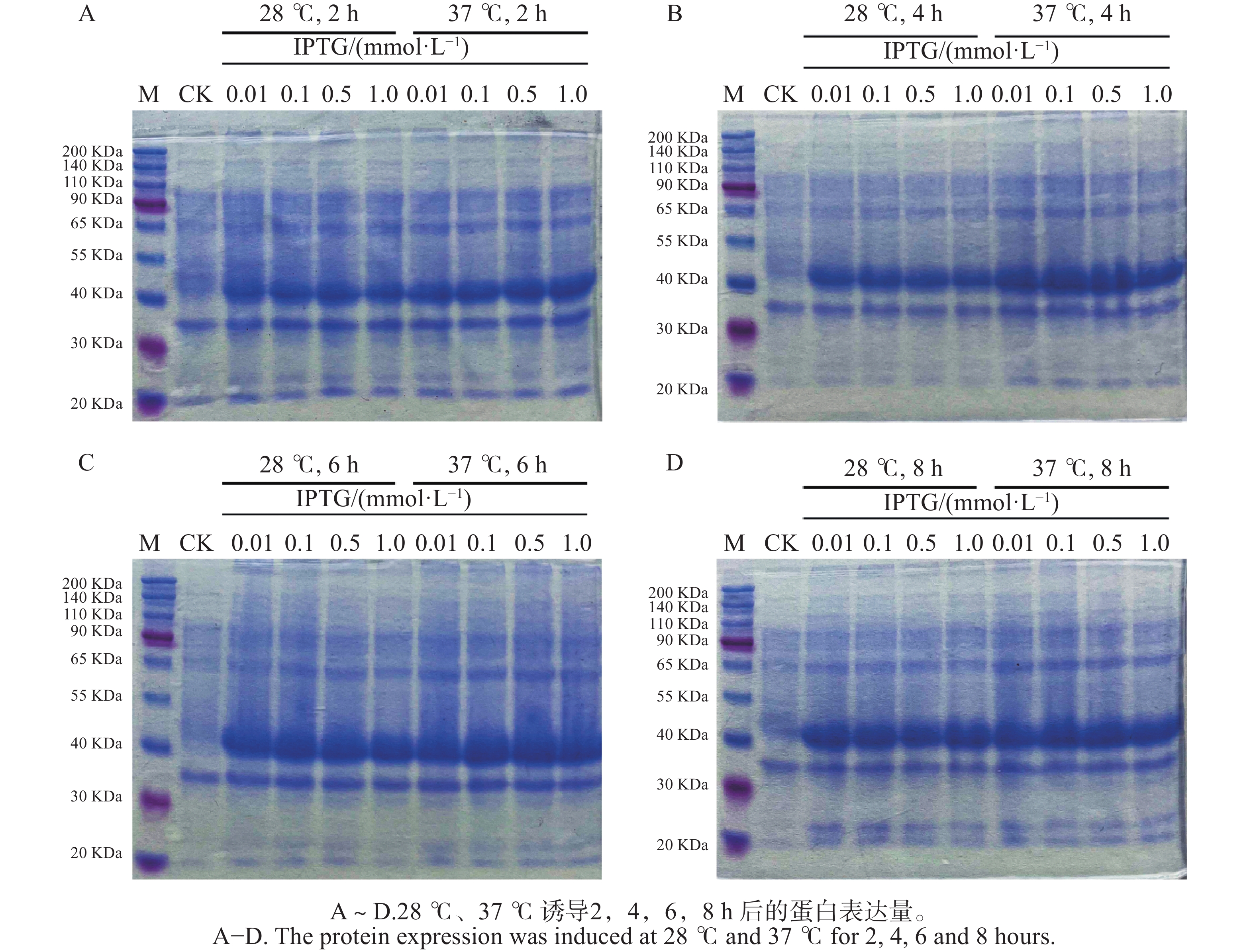
BZRs是BR信号通路中重要的转录因子,验证其是否具有转录因子的功能,往往需要表达出目标转录因子蛋白来验证其功能,表达出的TkBZR2蛋白可以为进一步探究其功能打下基础,也为后续实验提供了材料。 通过设置不同的诱导温度(28 ℃和37 ℃)和IPTG浓度(0.01、0.10、0.50、1.00 mmol·L−1)对TkBZR2蛋白进行原核表达。利用SDS-PAGE电泳检测pET28a-TkBZR2的原核表达条件情况,以找到最优的表达浓度、时间、温度。结果如图7所示,所设的4个IPTG浓度诱导在28 ℃和37 ℃下诱导2、4、6、8 h均能诱导表达出条带大小约为40 kDa的TkBZR2-His融合蛋白,且诱导8 h时蛋白表达量最大。所以选择28 ℃ 0.01 mmol·L−1 IPTG作为表达 TkBZR2蛋白的条件。后续纯化得到的蛋白经过Western-blot检验可以用于EMSA、ChIP等实验来进行DNA结合活性分析,从而筛选和鉴定TkBZR2下游靶基因。
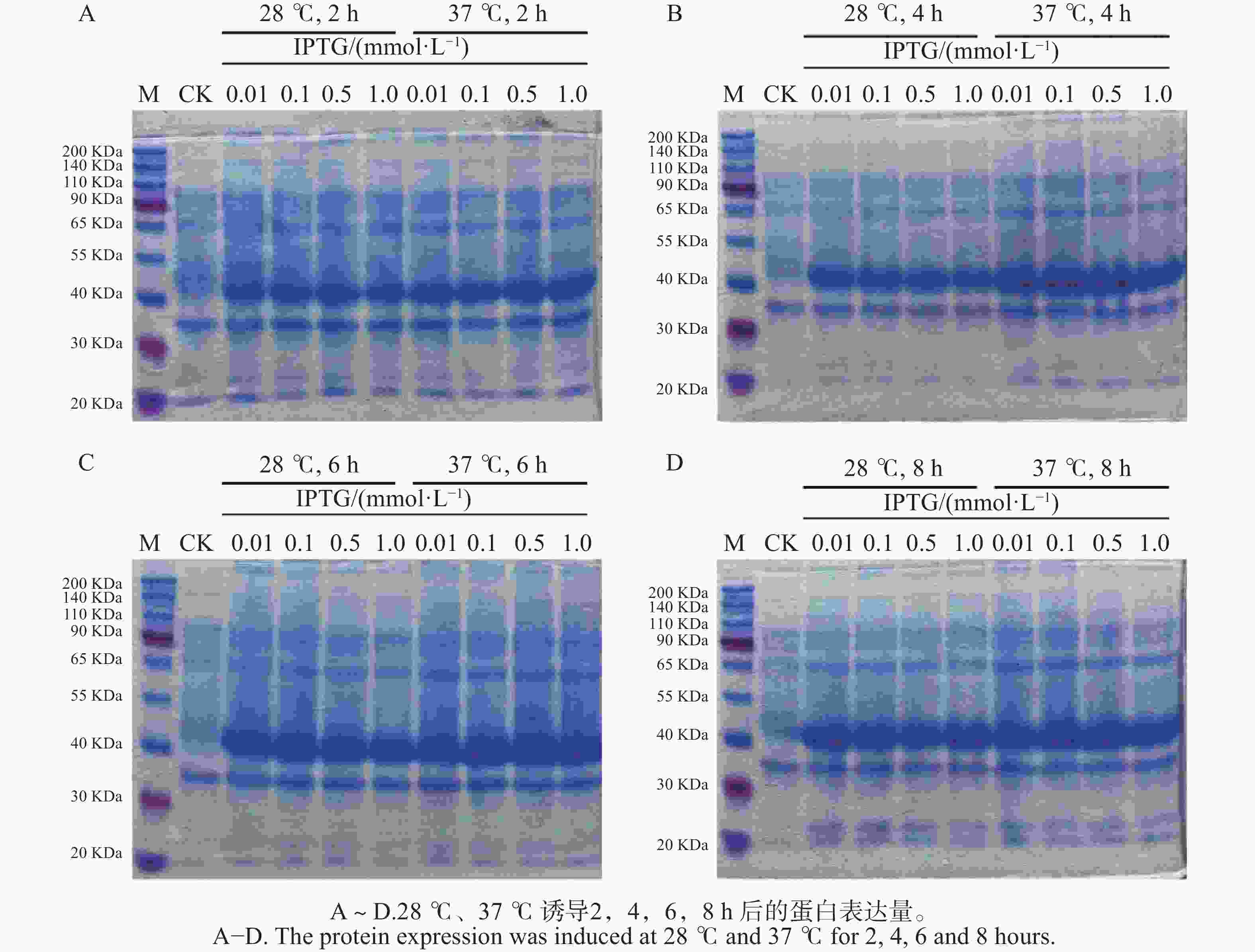
图 7 TkBZR2的原核表达
Figure 7. Prokaryotic expression of TkBZR2
-
在植物基因家族研究不断深入的进程中,BZR家族基因备受关注。当前,众多物种中BZR家族基因的鉴定与分析工作正逐步推进,本研究在橡胶草中鉴定出 6 个BZR/BES家族基因成员,这与拟南芥[18]、黄瓜[6]、甜瓜[24]、葡萄[25]、枣[26]等植物中BZR/BES家族基因成员个数相同,与水稻(4个)[7]、大麦(7个)[27]、番茄(9个)[28]等作物中的家族基因成员各不相同,暗示不同物种中需要不同个数的BZR成员行使功能。对不同物种中的BZR家族成员的系统发育进化树分析结果显示,莴苣和橡胶草BZR基因家族在进化过程中有较强的亲缘关系。橡胶草BZR含有典型的BZR/BES保守结构基序,表明BZR/BES基因家族在植物演化进程中具备高度的保守性。这意味着在漫长的植物进化历程中,BZR/BES基因家族发挥着不可或缺且相对稳定的生物学功能,可能参与了植物生长发育的基本调控过程,为植物适应环境变化提供了重要的分子基础。
橡胶草TkBZR基因家族成员展现出独特的理化性质,其平均等电点大多高于7.5,且多数成员主要定位于细胞核内。这一现象背后可能涉及多个关键因素,N端的核定位信号序列可能引导这些成员向细胞核运输,使其在细胞核内发挥转录调控等重要功能;高度保守的DNA结合结构域使它们能够精准识别并结合特定的DNA序列,从而调控下游基因的表达;磷酸化位点的存在则为其功能调节提供了动态机制,通过磷酸化与去磷酸化过程,响应细胞内各种信号通路的变化,进而调控基因表达活性;PEST序列可能参与了蛋白质的降解过程,确保细胞内TkBZR蛋白水平的精确调控;C端结构域的特性也可能对其亚细胞定位及功能发挥产生影响,例如参与蛋白质间的相互作用等。这些结构域和序列共同协作,使得TkBZR基因家族成员在细胞内发挥着复杂而有序的生物学功能[15-16]。
TkBZR基因家族的基因结构相对简洁,除TkBZR5包含3个外显子和2个内含子外,其余成员均由2个外显子和1个内含子构成。值得注意的是,橡胶草TkBZR基因与烟草[29]、南瓜[30]和棉花[31]BZR基因家族的基因结构呈现出相似性,多数成员也都只有2个外显子和1个内含子,极少数成员有3个外显子和2个内含子。这种相似的基因结构暗示着这些BZR/BES基因家族成员在功能上可能具有一定的保守性,它们可能通过相似的转录调控机制,影响植物的生长发育、激素响应以及逆境适应等生理过程。例如,在响应外界环境信号时,这些基因可能通过类似的剪接方式产生不同的转录本,进而调控不同的下游基因表达,以适应环境变化对植物生长发育的需求。
在对TkBZR2启动子序列的深入剖析中,预测到了一系列参与植物生长发育以及响应生物和非生物胁迫的结合元件。这一发现表明,TkBZR2在植物应对各种胁迫时,能够利用不同的信号通路精细调控植物的生理应答过程,进而对植物生长发育及胁迫响应产生深远影响。过往研究已明确,BZR/BES作为油菜素内酯(BR)信号传导途径中的核心转录因子,在植物激素信号转导网络中占据关键地位。例如,在 bzr1-1D突变体中,BZR1能够与ABA响应元件结合,抑制ABI5转录因子的转录激活,从而降低该突变体对 ABA信号的敏感性[32-34]。此外,BES1通过限制ABI3基因的转录活性,阻断ABI3对ABI5的激活作用,从而干扰ABA信号的传递过程。这一系列复杂的调控机制表明,BZR/BES家族基因在植物激素互作网络中扮演着重要角色,它们与ABA等激素信号通路相互交织,共同调控植物的生长发育和逆境适应能力[35-36]。
综上所述,本研究在橡胶草BZR基因家族研究方面取得了一系列重要成果。通过全基因组鉴定,本研究成功发现了 6 个橡胶草BZR基因家族的新成员,丰富了对橡胶草基因资源的认识。对橡胶草BZR基因在不同组织部位表达特异性的分析,揭示了其表达具有组织特异性和时空表达特异性。同时,探索该基因在原核系统中的最佳表达条件,为进一步研究其功能提供了有力工具。这些研究结果能为橡胶草BZR/BES家族基因的功能研究提供重要参考,有助于深入解析调控橡胶草生长发育与抗逆响应的分子机制,对橡胶草的遗传改良和品种选育具有重要的理论指导意义,也为其他植物中BZR/BES家族基因的研究提供了有益借鉴。今后,可进一步深入研究TkBZR基因家族成员在橡胶草中的具体生物学功能,明确其上下游调控因子,揭示其在橡胶草生长发育及抗逆过程中的精细调控网络。
橡胶草TkBZR家族基因的鉴定与原核表达分析
DOI: 10.15886/j.cnki.rdswxb.20250029
 CSTR: 32425.14.j.cnki.rdswxb.20250029
CSTR: 32425.14.j.cnki.rdswxb.20250029
Genome-wide identification and prokaryotic expression analysis of TkBZR family genes in rubber dandelion (Taraxacum kok-saghyz)
-
摘要: 为了进一步了解TkBZRs基因在橡胶草(Taraxacum kok-saghyz)生长发育和抵抗逆境中的作用,本研究通过分析橡胶草的全基因组测序数据,对TkBZR/BES家族基因进行鉴定分析,得到了6个TkBZR/BES家族成员,分别命名为TkBZR1至TkBZR6,并探讨其在染色体上的分布、结构域特征、组织特异性和时空表达特征。结果显示,6个TkBZR/BES家族成员分布在6条独立的骨架上。系统发育树显示,TkBZR/BES基因家族成员在进化上保守,橡胶草与莴苣的基因聚类在同一亚组,表明两者之间可能存在较近的亲缘关系。基因结构和保守结构域分析结果显示,除了TkBZR5外,其余TkBZR基因都包含2个外显子和1个内含子,所有成员都展示了高度保守的BES1_N结构域。表达模式分析发现,有5个成员在所有5个组织中表达,1个成员几乎不表达。此外,成功克隆获得表达丰度最高的TkBZR2基因,该基因编码307个氨基酸。将TkBZR2同源重组至原核表达载体上,重组成功后转入大肠杆菌(Escherichia coli) BL21(DE3)中并表达TkBZR2重组蛋白。Abstract: To further understand the role of the TkBZR family genes in the growth, development, and stress resistance of rubber dandelion (Taraxacum kok-saghyz), the TkBZR/BES family genes were identified and analyzed by examining the whole genome sequencing data of T. kok-saghyz. A total of 6 members of the TkBZR family were identified, named from TkBZR1 to TkBZR6, and their distribution on chromosomes, domain characteristics, tissue expression profiles and the spatial-temporal gene expression were analyzed. The results revealed that TkBZR/BES gene family members were distributed on 6 independent scaffolds. Phylogenetic trees indicate that TkBZR/BES gene family members are evolutionarily conserved, with rubber dandelion and lettuce genes clustering in the same subgroup, suggesting a possible close evolutionary relationship between the two. Gene structure and conserved domain analysis show that except for TkBZR5, all the other TkBZR genes contain two exons and one intron, and that all the family members exhibit a highly conserved BES1_N domain. Expression pattern analysis reveals that 5 members are expressed in all five tissues, while one member is almost unexpressed. Additionally, the TkBZR2 gene with the highest expression abundance was successfully cloned, encoding 307 amino acids. Homologous recombination of TkBZR2 was made into prokaryotic expression vector, and the recombinant plasmid was transferred into E. coli BL21 (DE3) .The recombinant protein TkBZR2 was expressed in E. coli BL21 (DE3).
-
Key words:
- Taraxacum kok-saghyz /
- BZR/BES /
- gene family /
- gene structure analysis /
- prokaryotic expression
-
图 2 橡胶草和其他植物BZR基因家族进化树
注:At. 拟南芥;Hb. 巴西橡胶树;Tk. 橡胶草;Ls. 莴苣;Nt. 烟草。
Fig. 2 Phylogenetic tree of BZR gene families in Taraxacum kok-saghyz and other plants
Note: At. Arabidopsis thaliana; Hb. Hevea brasiliensis; Tk. Taraxacum koksaghyz; Ls. Lactuca sativa; Nt. Nicotiana tabacum.
图 3 TkBZR家族的基因结构和基序分析
A.TkBZR家族成员序列信息(方格表示外显子,线条表示内含子);B.TkBZR家族成员进化树分析和结构域信息(方块表示基序结构)。
Fig. 3 Gene structure and motif analysis of TkBZR family
A. Information of TkBZR family member sequence (the square indicates exons and the line represents introns); B. Phylogenetic analysis and domain information of TkBZR family genes (block represents motif structure).
图 4 TkBZR表达模式分析
注:S4~S9表示不同的植物组织,从左至右分别为花、胶乳、叶片、根、茎;S1~S9表达不同的橡胶草不同的生育期;纵坐标表示基因表达的丰度值。
Fig. 4 Analysis of TkBZR expression patterns
Note: S4−S9 represents different plant tissues, from left to right: flower, latex, leaf, root, and scape; S1−S9 represents different growth stages of Taraxacum koksaghyz; the y-axis indicates the gene expression abundance values.
图 5 TkBZR2基因的扩增与分析
A.电泳检测TkBZR2;B.TkBZR2与其他物种中BZR的氨基酸序列比对;C~D.TkBZR2的亲水性和疏水性分析;E.TkBZR2跨膜结构域预测;F.TkBZR2二级结构预测。
Fig. 5 Amplification and analysis of TkBZR2 gene
A. Electrophoresis detection of TkBZR2; B. Amino acid sequence comparison between TkBZR2 and BZR in other species; C−D. Hydrophilic and hydrophobic analysis of TkBZR2; E. Prediction of transmembrane domain of TkBZR2; F. Prediction of secondary structure of TkBZR2.
图 6 橡胶草BZR2启动子顺式作用元件分析
注:不同颜色的方块代表不同的响应元件,黑色线条表示启动子长度。
Fig. 6 Analysis of cis-acting elements of BZR2 promoter of Taraxacum kok-saghyz
Note: Different colored blocks represent distinct response elements, the black lines indicate the length of the promoter region.
图 7 TkBZR2的原核表达
注:M,Protein marker;CK,诱导前的全蛋白;3~6,28 ℃,0.01、0.10、0.50、1.00 mmol·L−1 IPTG诱导后的全蛋白;7~10,37 ℃, 0.01、0.10、0.50、1.00 mmol·L−1 IPTG诱导后的全蛋白。
Fig. 7 Prokaryotic expression of TkBZR2
Note: M, Protein marker; CK, Whole protein before induction; 3−6, At 28 ℃, 0.01, 0.10, 0.50, 1.00 mmol·L−1 IPTG, induced whole protein; 7−10, At 37 ℃, 0.01, 0.10, 0.50, 1.00 mmol·L−1 IPTG, induced whole protein.
表 1 引物序列
Table 1 Primer sequences
引物名称 Primer 序列 Primer sequence(5′−3′) TkBZR2-F ATGACCGGCGCCGGTTCT TkBZR2-R TTAAATTACACTTTTCCC pET28a-TkBZR2-F caaatgggtcgcggaATGACCGGCGCC pET28a-TkBZR2-R gagctcgaattcggagaAATTACACTTT
TCCCACTACCAAG 下载: 导出CSV
下载: 导出CSV
表 2 橡胶草基因组BZR/BES基因家族信息
Table 2 The information of BZR/BES gene family in Taraxacum kok-saghyz genome
基因名称
Gene基因编号
Gene ID染色体定位
Position氨基酸数目/aa
Length of
amino acid分子质量/kDa
Molecular
weight等电点
pI亲水性系数
Hydropathy
Index亚细胞定位
Subcellular
locationBZR1 GWHPAAAA046609 utg9916 316 34.35 9.38 −0.705 细胞核Nucleus BZR2 GWHPAAAA015365 utg19389 307 33.30 8.92 −0.589 细胞核Nucleus BZR3 GWHPAAAA030593 utg3490 302 32.99 8.45 −0.753 细胞核Nucleus BZR4 GWHPAAAA024338 utg27431 294 32.13 7.63 −0.711 细胞质Cytoplasm BZR5 GWHPAAAA045865 utg950 263 28.39 8.66 −0.572 细胞质与细胞核
Cytoplasm and NucleusBZR6 GWHPAAAA001420 utg10687 151 17.13 9.13 −0.909 细胞核Nucleus
下载: 导出CSV
表 3 Motif序列信息
Table 3 Sequence information of motifs
名称 Motif 序列 Motif sequence Motif1 YAGLRAQGNYKLPKHCDNNEVLKALCEEAGWVV Motif2 KAWEGERIHEVGVDD Motif3 FTKQSIPSFNLPFFASSAPTSPTRRHRFTPATIPECDESDCSTIDSCQWV Motif4 EPDGTTYRKGCKP Motif5 NPSSLPPLRISNSAPVTPPLSSPTSK Motif6 STNTSPCSSQKPSPISSSFPSPSPSDQAS
下载: 导出CSV
-
[1] 陈秋惠, 杨玉双, 覃碧, 等. 橡胶草育种研究进展[J/OL]. 分子植物育种, 1−15(2023-12-16)[2025-02-07]. http://kns.cnki.net/kcms/detail/46.1068.S.20231215.0914.002.html. [2] 张在宝, 王涵, 罗天, 等. 橡胶草育种及橡胶提取工艺研究进展[J/OL]. 宁夏大学学报(自然科学版), 1−8(2023-05-06)[2025-02-07]. http://kns.cnki.net/kcms/detail/64.1006.N.20230504.1519.022.html. [3] 贺俊亚, 杨雪, 邢剑锋, 等. 橡胶草TCTP基因家族的鉴定及表达分析[J/OL]. 分子植物育种, 1−18(2023-11-15)[2025-02-07]. http://kns.cnki.net/kcms/detail/46.1068.S.20231114.1714.022.html. [4] 刘星, 杨雪, 张芳, 等. 橡胶草乳管特异性高表达基因TkREF的启动子区段鉴定[J]. 分子植物育种, 2025, 23(4): 1154−1163. [5] 李闯, 王肖肖, 杨玉双, 等. 橡胶草Tk-bZIP11转录因子基因的克隆及其表达模式分析[J]. 分子植物育种, 2024, 22(5): 1444−1453. [6] Ma Y, Xu J, Qi J, et al. Crosstalk among plant hormone regulates the root development [J]. Plant Signaling & Behavior, 2024, 19(1): 2404807. [7] Mitchell J W, Mandava N, Worley J F, et al. Brassins: a new family of plant hormones from rape pollen [J]. Nature, 1970, 225(5237): 1065−1066. https://doi.org/10.1038/2251065a0 doi: 10.1038/2251065a0 [8] Grove M D, Spencer G F, Rohwedder W K, et al. Brassinolide, a plant growth-promoting steroid isolated from Brassica napus pollen [J]. Nature, 1979, 281(5728): 216−217. https://doi.org/10.1038/281216a0 doi: 10.1038/281216a0 [9] Chaudhuri A, Halder K, Abdin M Z, et al. Abiotic stress tolerance in plants: brassinosteroids navigate competently [J]. International Journal of Molecular Sciences, 2022, 23(23): 14577. https://doi.org/10.3390/ijms232314577 doi: 10.3390/ijms232314577 [10] 杜巧丽, 刘均霞, 陈美晴, 等. 高粱BR信号转录因子BZR1基因家族的鉴定及激素应答分析[J]. 植物保护学报, 2022, 49(3): 848−856. [11] Luo S, Zhang G, Zhang Z, et al. Genome-wide identification and expression analysis of BZR gene family and associated responses to abiotic stresses in cucumber (Cucumis sativus L.) [J]. BMC Plant Biology, 2023, 23(1): 214. https://doi.org/10.1186/s12870-023-04216-9 doi: 10.1186/s12870-023-04216-9 [12] Bai M Y, Zhang L Y, Gampala S S, et al. Functions of OsBZR1 and 14-3-3 proteins in brassinosteroid signaling in rice [J]. Proceedings of the National Academy of Sciences of the United States of America, 2007, 104(34): 13839−13844. [13] 李明, 颉嘉丽, 石铭福, 等. 外源2, 4-表油菜素内酯对碱性盐胁迫下马铃薯根系生长、生理特性及土壤酶活性的影响[J]. 江苏农业学报, 2024, 40(3): 394−402. https://doi.org/10.3969/j.issn.1000-4440.2024.03.002 doi: 10.3969/j.issn.1000-4440.2024.03.002 [14] 丁丹阳, 张璐翔, 朱智威, 等. 叶面喷施2, 4-表油菜素内酯对烟草抗旱性的影响[J]. 中国烟草科学, 2018, 39(4): 50−57. [15] Li J, Chory J. A putative leucine-rich repeat receptor kinase involved in brassinosteroid signal transduction [J]. Cell, 1997, 90(5): 929−938. https://doi.org/10.1016/S0092-8674(00)80357-8 doi: 10.1016/S0092-8674(00)80357-8 [16] Yin Y, Vafeados D, Tao Y, et al. A new class of transcription factors mediates brassinosteroid-regulated gene expression in Arabidopsis [J]. Cell, 2005, 120(2): 249−259. https://doi.org/10.1016/j.cell.2004.11.044 doi: 10.1016/j.cell.2004.11.044 [17] Belkhadir Y, Jaillais Y. The molecular circuitry of brassinosteroid signaling [J]. The New Phytologist, 2015, 206(2): 522−540. https://doi.org/10.1111/nph.13269 doi: 10.1111/nph.13269 [18] Clouse S D. Brassinosteroid signal transduction: from receptor kinase activation to transcriptional networks regulating plant development [J]. The Plant Cell, 2011, 23(4): 1219−1230. https://doi.org/10.1105/tpc.111.084475 doi: 10.1105/tpc.111.084475 [19] Kim T W, Guan S, Burlingame A L, et al. The CDG1 kinase mediates brassinosteroid signal transduction from BRI1 receptor kinase to BSU1 phosphatase and GSK3-like kinase BIN2 [J]. Molecular Cell, 2011, 43(4): 561−571. https://doi.org/10.1016/j.molcel.2011.05.037 doi: 10.1016/j.molcel.2011.05.037 [20] Wang Z Y, Nakaano T, Gendron J, et al. Nuclear-localized BZR1 mediates brassinosteroid-induced growth and feedback suppression of brassinosteroid biosynthesis [J]. Developmental Cell, 2002, 2(4): 505−513. https://doi.org/10.1016/S1534-5807(02)00153-3 doi: 10.1016/S1534-5807(02)00153-3 [21] 赵同,张盛敏,曹杰,等. 橡胶草bZIP基因家族鉴定及胶乳高表达基因功能分析[J]. 南方农业学报, 2025, 56(9): 2723−2735. https://doi.org/10.3969/j.issn.2095-1191.2025.09.003 doi: 10.3969/j.issn.2095-1191.2025.09.003 [22] 郑立鹏, 吐汗姑丽·托合提, 高强, 等. 橡胶草萌发期耐盐碱性鉴定评价及优异种质挖掘 [J]. 南方农业学报, 2025, 56(9): 2736−2747. https://doi.org/10.3969/j.issn.2095-1191.2025.09.004 doi: 10.3969/j.issn.2095-1191.2025.09.004 [23] He J X, Gendron J M, Sun Y, et al. BZR1 is a transcriptional repressor with dual roles in brassinosteroid homeostasis and growth responses [J]. Science, 2005, 307(5715): 1634−1638. https://doi.org/10.1126/science.1107580 doi: 10.1126/science.1107580 [24] Xu M, Zhang Y, Yang X, et al. Genome-wide analysis of the SWEET genes in Taraxacum kok-saghyz Rodin: an insight into two latex-abundant isoforms [J]. Plant Physiology and Biochemistry, 2023, 194: 440−448. https://doi.org/10.1016/j.plaphy.2022.11.014 doi: 10.1016/j.plaphy.2022.11.014 [25] 臧文蕊, 马明, 砗根, 等. 甜瓜BZR转录因子家族基因的全基因组鉴定及表达模式分析[J]. 生物技术通报, 2024, 40(7): 163−171. [26] 江倩倩, 王雨婷, 惠竹梅. 葡萄BZR基因家族的鉴定及表达分析[J]. 植物生理学报, 2021, 57(6): 1218−1228. [27] 尹魁林, 闫雪晴, 王玖瑞, 等. 枣EIN3/EIL基因家族鉴定及其在枣果实发育中的表达分析[J/OL]. 分子植物育种, 1−14(2024-08-08)[2025-02-08]. http://kns.cnki.net/kcms/detail/46.1068.S.20240808.1201.004.html. [28] 薛正刚, 王树杰, 杨永乾, 等. 大麦GRF家族的基因组鉴定及生物信息学分析[J]. 分子植物育种, 2021, 19(6): 1750−1757. [29] 陈旭, 沈春洋, 莫福磊, 等. 番茄BZR基因家族鉴定及非生物胁迫下表达模式分析[J]. 东北农业大学学报, 2021, 52(11): 9−17. https://doi.org/10.3969/j.issn.1005-9369.2021.11.002 doi: 10.3969/j.issn.1005-9369.2021.11.002 [30] Chen X, Wu X, Qiu S, et al. Genome-wide identification and expression profiling of the BZR transcription factor gene family in Nicotiana benthamiana [J]. International Journal of Molecular Sciences, 2021, 22(19): 10379. https://doi.org/10.3390/ijms221910379 doi: 10.3390/ijms221910379 [31] 李春, 刘小俊, 蔡鹏, 等. 中国南瓜BZR基因家族的全基因组鉴定及生物信息学分析[J]. 分子植物育种, 2022, 20(19): 6324−6330. [32] 郭新磊, 路普, 王园园, 等. 棉花BZR基因家族的全基因组鉴定及表达分析[J]. 棉花学报, 2017, 29(5): 415−427. https://doi.org/10.11963/1002-7807.gxllf.20170830 doi: 10.11963/1002-7807.gxllf.20170830 [33] Liu Z Q, Yan L, Wu Z, et al. Cooperation of three WRKY-domain transcription factors WRKY18, WRKY40, and WRKY60 in repressing two ABA-responsive genes ABI4 and ABI5 in Arabidopsis [J]. Journal of Experimental Botany, 2012, 63(18): 6371−6392. https://doi.org/10.1093/jxb/ers293 doi: 10.1093/jxb/ers293 [34] Cai R, Zhao Y, Wang Y, et al. Overexpression of a maize WRKY58 gene enhances drought and salt tolerance in transgenic rice [J]. Plant Cell, Tissue and Organ Culture (PCTOC), 2014, 119(3): 565−577. https://doi.org/10.1007/s11240-014-0556-7 doi: 10.1007/s11240-014-0556-7 [35] Fahrendorf T, Dixon R A. Stress responses in alfalfa (Medicago sativa L.). XVIII: Molecular cloning and expression of the elicitor-inducible cinnamic acid 4-hydroxylase cytochrome P450 [J]. Archives of biochemistry and biophysics, 1993, 305(2): 509−515. https://doi.org/10.1006/abbi.1993.1454 doi: 10.1006/abbi.1993.1454 [36] Ren X, Chen Z, Liu Y, et al. ABO3, a WRKY transcription factor, mediates plant responses to abscisic acid and drought tolerance in Arabidopsis [J]. The Plant journal: for cell and molecular biology, 2010, 63(3): 417−429. -

 点击查看大图
点击查看大图
计量
- 文章访问数: 1052
- HTML全文浏览量: 496
- PDF下载量: 84
- 被引次数: 0
