

-
珊瑚礁生态系统具有全球最高生态多样性和初级生产力,被誉为海底热带雨林,具有生境功能、产出功能、调节功能和信息功能等[1]。受人类活动和全球气候变化的影响,全球珊瑚礁正在遭受众多危机,珊瑚白化等问题日渐严重,目前,大约有三分之一的珊瑚物种面临灭绝的风险[2]。在当前珊瑚生存面临严重威胁的背景下,相比于全球变暖、海洋酸化而言,盐度是影响珊瑚生长发育的重要环境因子,但其在珊瑚低盐胁迫响应中的研究仍较缺乏。主要的原因可能是海洋中盐度的变化通常较为稳定,其次珊瑚正常生长的盐度范围较为广泛,一般是27~40盐度,且具有一定的抗高盐特性[3]。但珊瑚在面对低盐环境时的表现却较为敏感[4 − 5],海水变暖期间暴露于低盐度的近岸珊瑚比离岸珊瑚经历了更多的白化[6],且珊瑚自身缺乏渗透压调节能力[7],因此,在低盐环境下,珊瑚白化的风险极高。除此之外,在热带地区,尤其是入海口区域,极端强降雨时沿岸陆地径流骤增可能会导致海水盐度降低超过10[8],部分珊瑚被迫生活在盐度低于15的环境中,珊瑚很容易白化甚至死亡[3,9]。
珊瑚白化是指珊瑚体内的虫黄藻(Effrenium voratum)与珊瑚的共生关系被破坏从而导致珊瑚变白的现象。虫黄藻在珊瑚的生长发育过程中扮演着极为重要的角色,虫黄藻的光合作用产物是珊瑚宿主的主要能量来源[10]。以往的研究将盐度的变化导致珊瑚白化归因于盐度对虫黄藻的影响,导致转移到珊瑚的能量减少[11 − 13]。低盐环境下的渗透压使虫黄藻细胞体积增加、膨胀,导致离子和有机溶质的流失以及膜和细胞器的损伤,最终可能导致酶系统受到抑制,破坏新陈代谢功能,甚至细胞破裂[14 − 15]。Downs等[16]的研究揭示了低盐胁迫会诱导虫黄藻产生氧化应激反应。而Sakami[17]的研究也证实,低盐条件下,会对离体虫黄藻光合作用带来直接的负面影响。然而,虫黄藻含有上百种不同的类群,不同类群的虫黄藻无论是在生活方式还是对珊瑚虫的相互作用方面,都有各自的生理特征。只有通过对不同类群的虫黄藻进行分离培养,收集与分析环境胁迫下虫黄藻的响应数据,才能更好地揭示虫黄藻在珊瑚-虫黄藻共生体关系构建、维持、破坏和重建中的作用[18]。目前,离体培养的虫黄藻对低盐胁迫的响应机制研究相对较少,一定程度限制了人们对当前虫黄藻−珊瑚共生关系的基本理解[19]。近期的研究者将共生虫黄藻主要分为Symbiodinium、Breviolum、Cladocopium、Durusdinium、Effrenium、Fugacium、Gerakladium、Clade H 和Clade I,其中,虫黄藻Effrenium属目前只包含Effrenium voratum[20]。E. voratum广泛分布于温带和亚热带水域,如太平洋、大西洋和地中海地区,得益于其在世界各地的各种海洋环境中的生存,近期研究发现,该物种具有宽温度范围、高光强度的耐受性及混养能力[21]。E. voratum虫黄藻不仅可以与珊瑚宿主共生,还可以在珊瑚礁生态系统中浮游生存。研究低盐胁迫下离体培养E. voratum的响应机制,有助于初步了解虫黄藻对盐度变化的生理耐受性。然而,离体状态与共生状态下的胁迫响应可能存在显著差异。在共生状态下,虫黄藻位于珊瑚细胞内,受到宿主的渗透调节和保护,而离体培养的虫黄藻直接暴露于低盐环境,可能表现出更强的敏感性。因此,未来研究需结合共生体系,进一步探究低盐胁迫对虫黄藻−珊瑚共生关系的影响。因此,本研究以低盐作为环境压力源,将E. voratum在不同盐度环境下进行培养,测定E. voratum的生理生化指标包括生长状况、单个虫黄藻Chl a浓度、超氧化物歧化酶(SOD)和过氧化氢酶(CAT)活力,研究在低盐环境下虫黄藻E. voratum的响应过程,旨在为虫黄藻种群的耐盐性研究提供了基础数据。
-
虫黄藻(E. voratum)从实验室已有的E. voratum母液中转接获取,在250 mL的装有L1-Si培养基的锥形瓶中保存,置于室内恒温光照培养箱中培养,温度为26 °C,光照强度为90 μmol·m–2·s–1,光暗周期为L(光)︰D(暗)=12 h︰12 h,盐度条件为35、25、15。每隔1周用显微镜(奥克巴斯−BX53)观察转接的虫黄藻生长情况,并对其进行形态学鉴定。
-
为了确定培养的共生虫黄藻的纯度,对转接后培养的虫黄藻进行了DNA提取,使用DNeasy®植物试剂盒(Qiagen, Hilden, Germany)按说明书步骤提取基因组DNA。然后,将提取的DNA样品送至上海美吉生物有限公司进行高通量测序。利用引物F: 5′−GAATTGCAGAACTCCGTG−3'和R: 5′−GGGATCCATATGCTTAAGTTCAGCGGGT−3′对虫黄藻rDNA的ITS2区进行了PCR扩增。使用AxyPrep DNA凝胶提取试剂盒(Axygen Biosciences)和QuantiFluorTM ST荧光定量系统(Promega)将最后的PCR产物纯化和定量在301~340 bp。纯化的扩增子以等摩尔量汇集,然后,根据制造商标准使用协议在Illumina Mi Seq测序平台进行测序。
-
本实验使用L1-Si培养基,利用虫黄藻细胞作为实验对象开展实验研究。设定3个不同的盐度条件,分别为正常的35盐度对照组和低盐的25、15盐度实验组。每个盐度条件设置至少3个独立的生物学重复,每个重复包含200 mL培养基,初始细胞密度为1 × 104 cells·mL−1。待细胞进入指数增长期后,重新接种于灭菌后盐度分别为35、25、15的L1-Si培养基中,所有培养瓶在恒温光照培养箱中随机排列,在26 ℃,L(光)︰D(暗)=12 h︰12 h光周期等其他培养条件相同的环境下胁迫48 h。在0、4、24、48、72 h收集样品,每个时间点的样品均进行3次重复测定,以测定细胞密度、单个虫黄藻Chl a含量、酶活性等生理指标。
-
用微量移液器取一式3份,每份100 μL均匀藻液于2 mL离心管内,加入900 μL无菌海水稀释,将稀释后的虫黄藻溶液用旋涡混匀器充分混合均匀。然后,用微量移液器移取100 μL稀释后的藻液到浮游生物计数框中(覆盖盖玻片),在显微镜(奥克巴斯−BX53)下进行虫黄藻的计数(n=10),最后将统计结果换算成总体积溶液内所含虫黄藻的数量,即虫黄藻密度(D,单位:cells·mL−1)。
在充分混合均匀锥形瓶中的藻液之后,从中取出一定量的溶液,每份为2 mL,共取3份。将这些溶液分别加入离心管中,使用台式高速冷冻离心机(Sigma−15)进行离心分离(设定4 000 r·min−1的速度,离心5 min)。之后,慢慢地倾倒去除上清液,留下的沉淀物再加入1.5 mL φ=100%的丙酮,在4 ℃条件下进行24 h的萃取。在萃取了24 h后,将溶液再次进行离心分离(设定4 000 r·min−1的速度,离心5 min),然后取其上清液(200 μL),用分光光度计分别测试波长750、664、647和630 nm处的吸光度,代入公式(1)计算Chl a质量浓度:
$$ \begin{split} \mathit{Ca} =&11.85\times ( \mathit{A} _{664} - \mathit{A} _{750} )-1.54\times ( \mathit{A} _{647} - \mathit{A} _{750} )-\\&0.08\times ( \mathit{A} _{ {630}} - \mathit{A} _{{750}} )\text{,} \end{split}$$ (1) 式中,Ca代表Chl a浓度(mg·L−1)。
再换算成实验总体积的藻液内Chl a质量浓度。最后,使用根据单位体检虫黄藻密度和Chl a质量浓度,用公式(2)计算出单个虫黄藻Chl a含量:
$$ \begin{split} \text{单个细胞的Chla含量}=&({\mathrm{Chla}}质量浓度\times 10^9)/\\&细胞密度\text{,} \\[-1pt] \end{split}$$ (2) 式中,单个虫黄藻Chl a含量的单位为pg·cell−1;Chl a质量浓度的单位为mg·mL−1;细胞密度的单位为cells·mL−1。
-
使用商用试剂盒(货号A001−3,A007−2−1)分别测定藻液中超氧化物歧化酶(SOD)与过氧化氢酶(CAT)总活性,所有试剂盒购买自南京建成生物工程研究所,使用方法参照测定试剂盒的操作方法,具体操作步骤见文献[22]。
所有数据均记录为至少3个独立生物重复实验的平均值 ± 标准偏差。使用非参数检验Kruskal-Wallis对实验数据作显著性分析,并将P<0.05视为具有显著差异,P>0.05视为差异不显著。对于同一盐度组内不同时间点的数据,不同盐度组在同一时间点的数据,均采用Kruskal-Wallis检验进行组间比较。所有结果的分析使用软件SPSS Statistics进行,可视化使用软件Origin2021进行。
-
本实验对离体培养的藻液样品进行高通量测序,以检测藻液的虫黄藻组成(原始数据上传至NCBI数据库
https://www.ncbi.nlm.nih.gov/ ,SRA:PRJNA1000185)。测序结果(表1)表明,本研究中离体培养的藻样中虫黄藻(E. voratum)的纯度达到(99.971±0.032)%,可以确定本次实验藻种为虫黄藻(E. voratum),这些虫黄藻可以用于开展后续实验。表 1 离体培养虫黄藻E. voratum藻液样品高通量测序结果
Table 1. High-throughput sequencing of the algal liquid of the dinoflagellate E. voratum cultured in vitro
% 虫黄藻物种
Taxon样本1相对丰度
Sample1
Relative abundance样本2相对丰度
Sample2
Relative abundance样本3相对丰度
Sample3
Relative abundance均值
MeanSD Effrenium voratum 99.988 99.934 99.991 99.971 0.032 Others 0.011 0.066 0.009 -
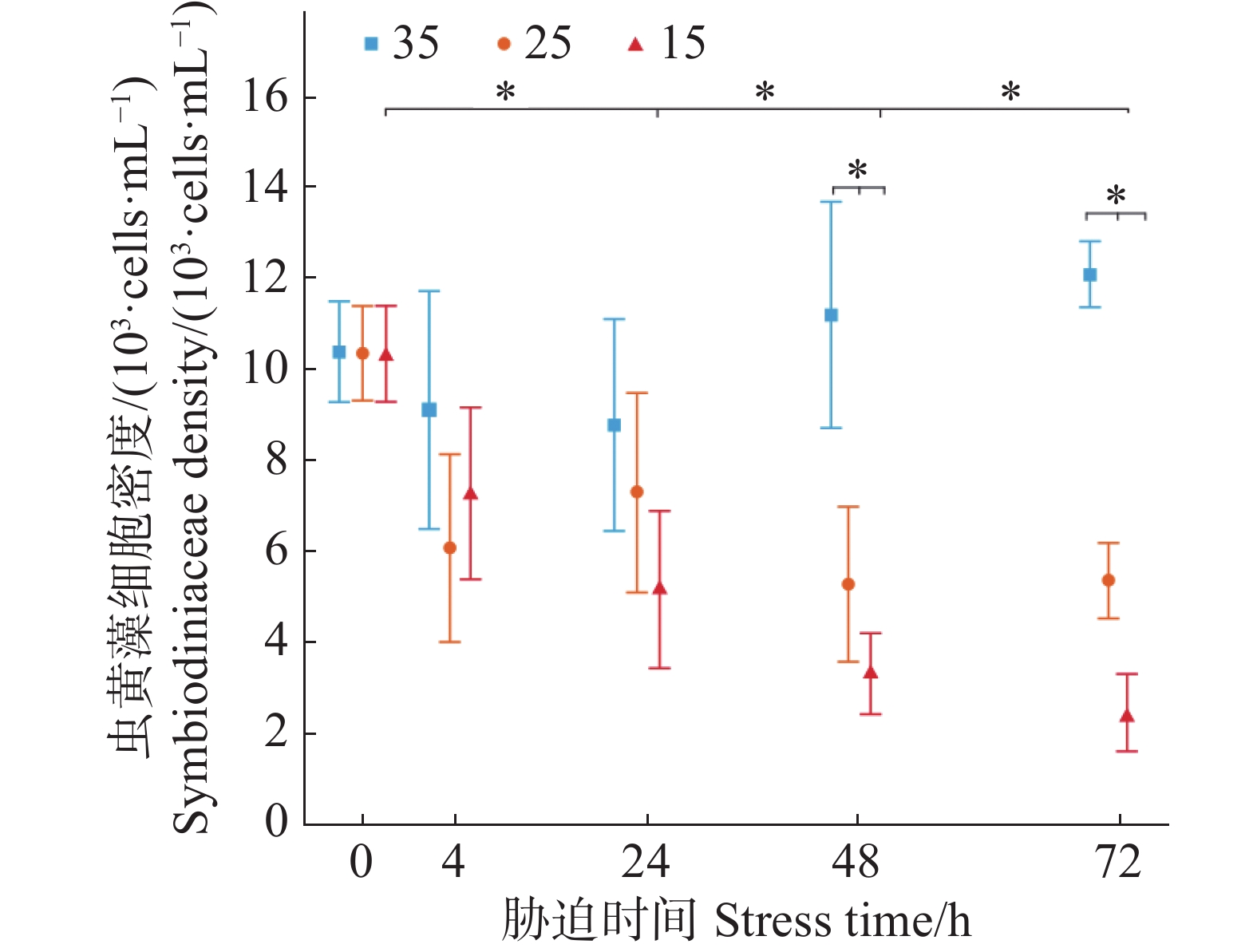
本实验将正常条件培养盐度为35的虫黄藻作为对照组,盐度条件改变为25、15培养的虫黄藻作为胁迫实验组。虫黄藻的初始理论接种浓度控制在1 × 104 cells·mL−1。从图1可知,盐度35正常条件培养下的虫黄藻密度在72 h内有较为稳定的上升趋势但不显著,25的低盐胁迫下在72 h内虫黄藻密度变化不显著,而15的低盐胁迫下虫黄藻密度变化呈现不断下降趋势且在第48 h已显著低于的初始阶段(P = 0.045)。在48 h时,25与15盐度下的虫黄藻密度均已显著低于35盐度下的虫黄藻密度(P < 0.05)。
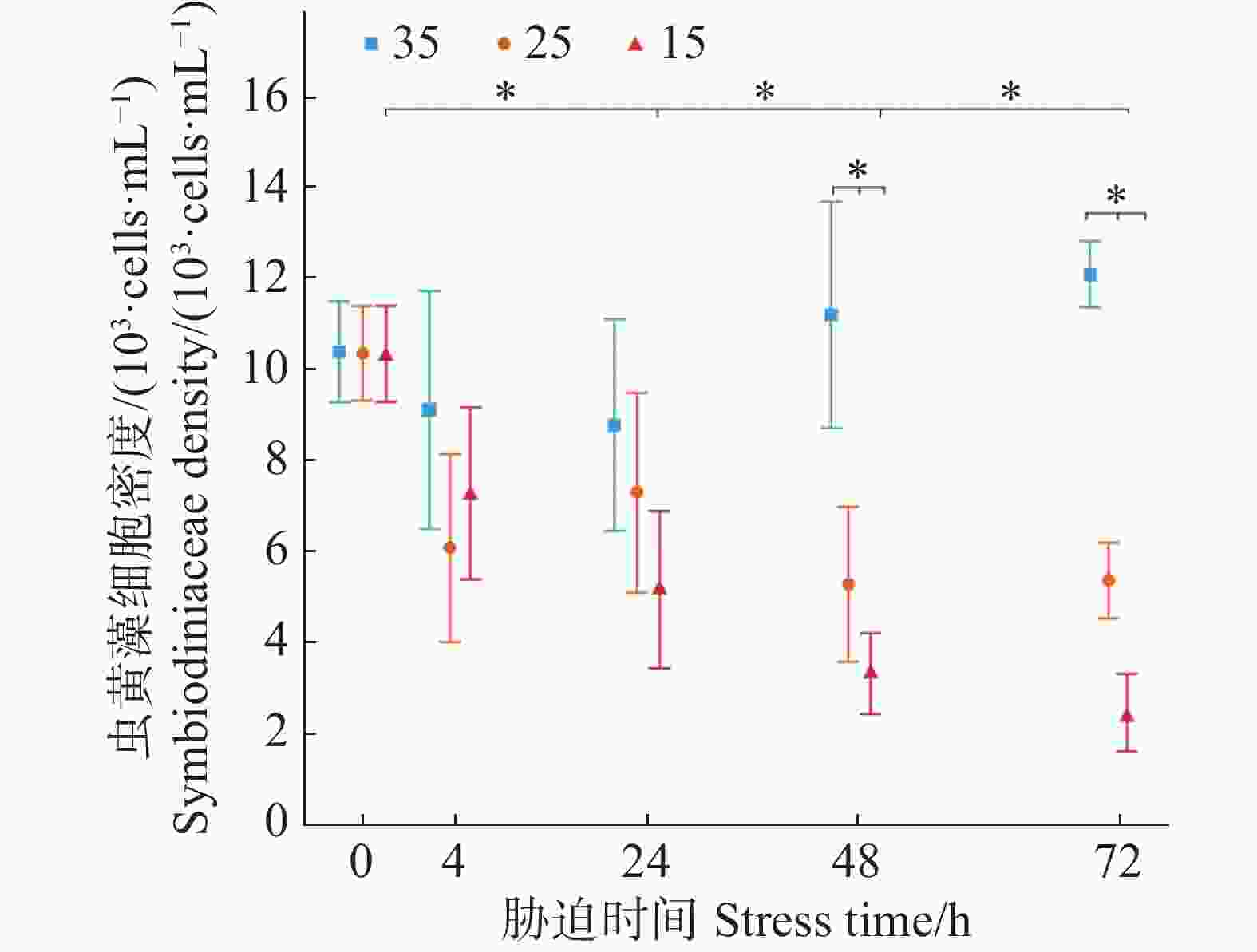
图 1 不同盐度条件下虫黄藻密度随胁迫时间的变化
Figure 1. Changes in Symbiodiniaceae density under different salinity conditions over stress time
-
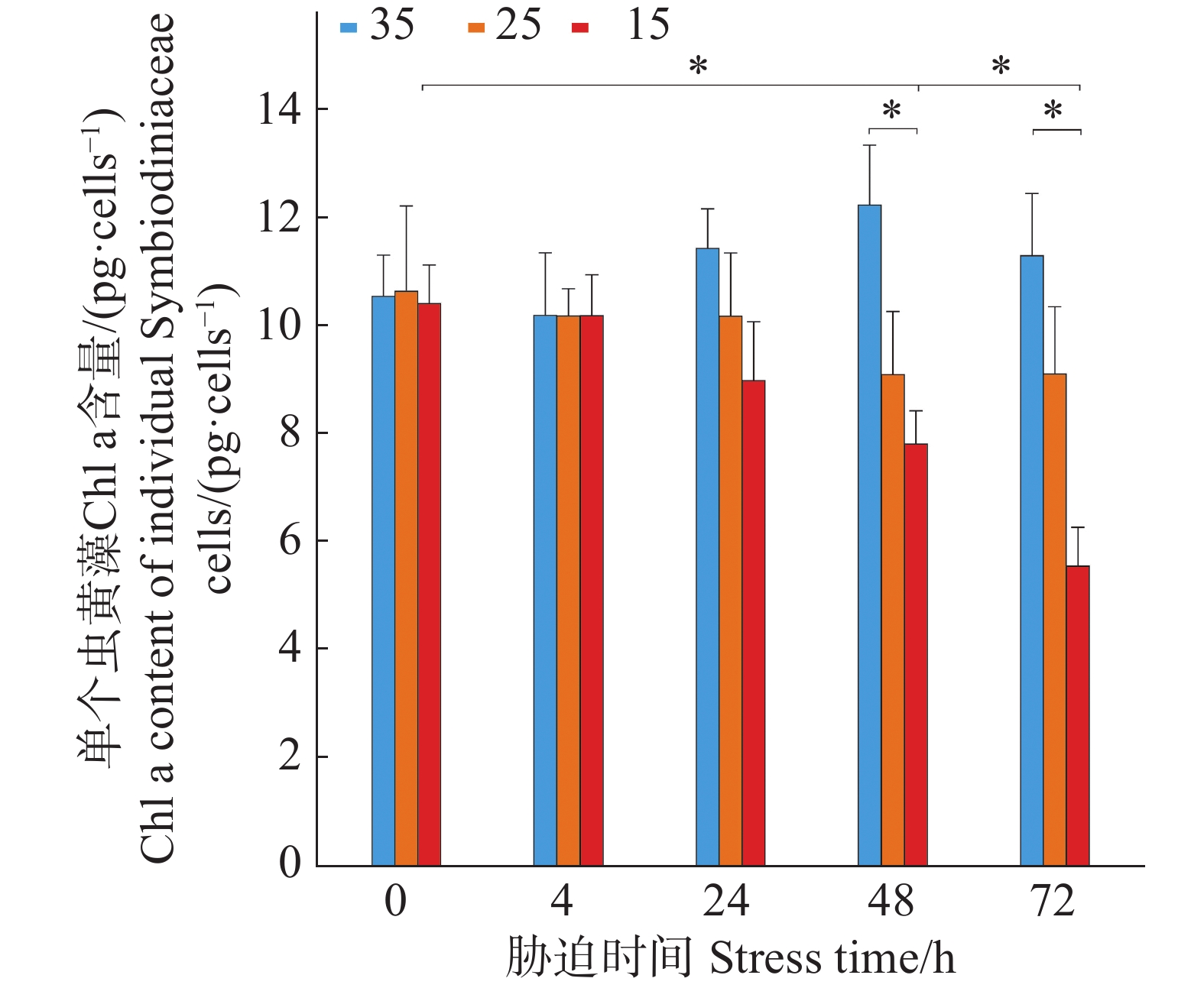
如图2所示,3个盐度梯度(35、25、15)下,单个虫黄藻细胞的Chl a初始含量分别为(10.55 ± 0.76)、(10.66 ± 1.57)和(10.43 ± 0.69)pg·cell−1,4 h时单个虫黄藻细胞的Chl a含量与初始含量无显著差异(P > 0.05)。在盐度35培养条件下,单个虫黄藻细胞的Chl a的含量在48 h内有上升趋势并在72 h后有所下降,但仍比初始含量高。在25盐度的培养条件下,单个虫黄藻细胞的Chl a的含量在24 h内保持稳定,在24 h后略有下降,但差异不显著(P > 0.05)。而15盐度培养下,从4 h开始的单个虫黄藻细胞的Chl a含量出现连续下降趋势,48、72 h时单个虫黄藻细胞的Chl a含量显著低于初始含量(P < 0.05)。
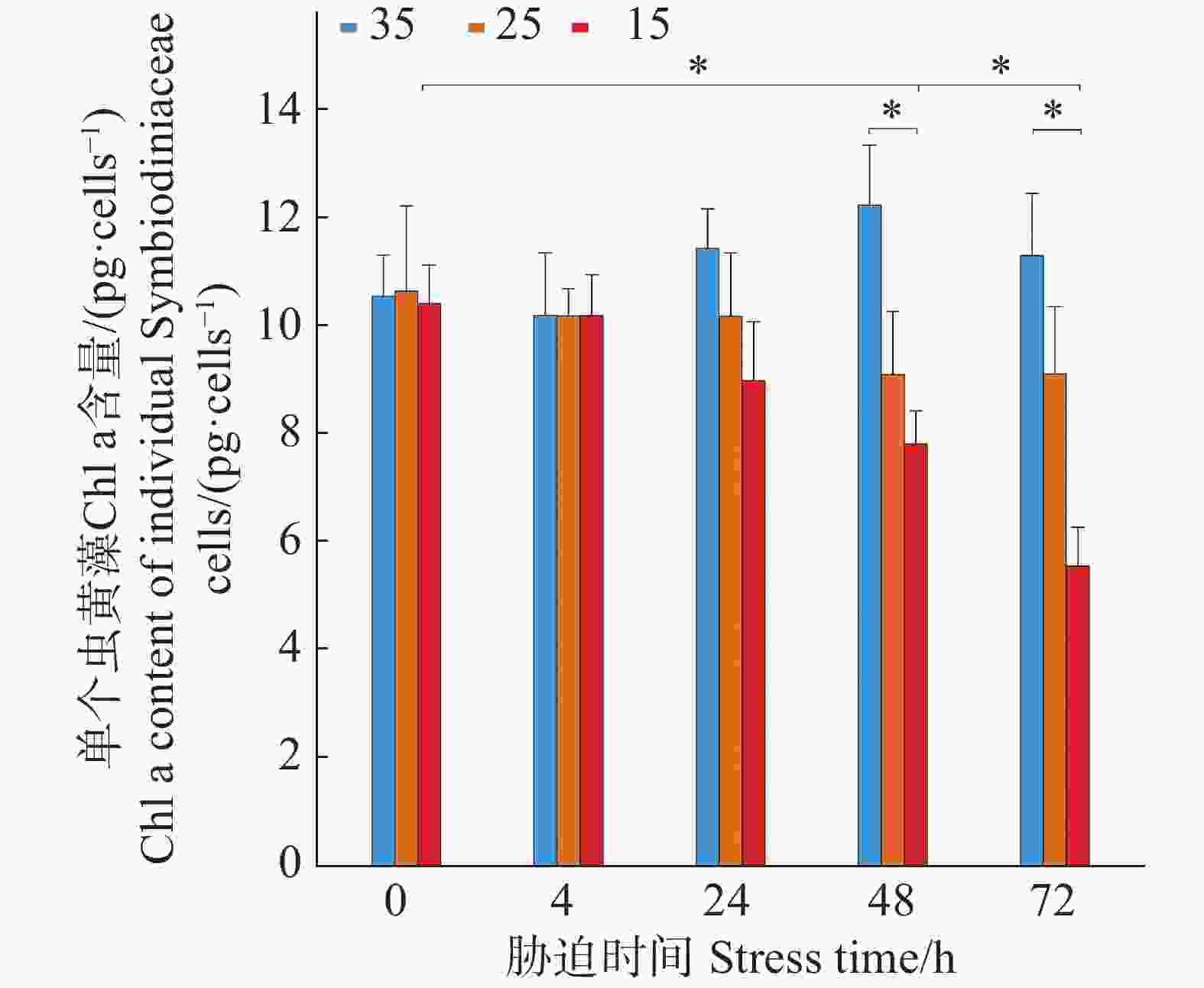
图 2 不同盐度条件下单个虫黄藻的Chl a含量随胁迫时间的变化
Figure 2. Changes in Chl a content of individual Symbiodiniaceae cells under different salinity conditions over stress time
-
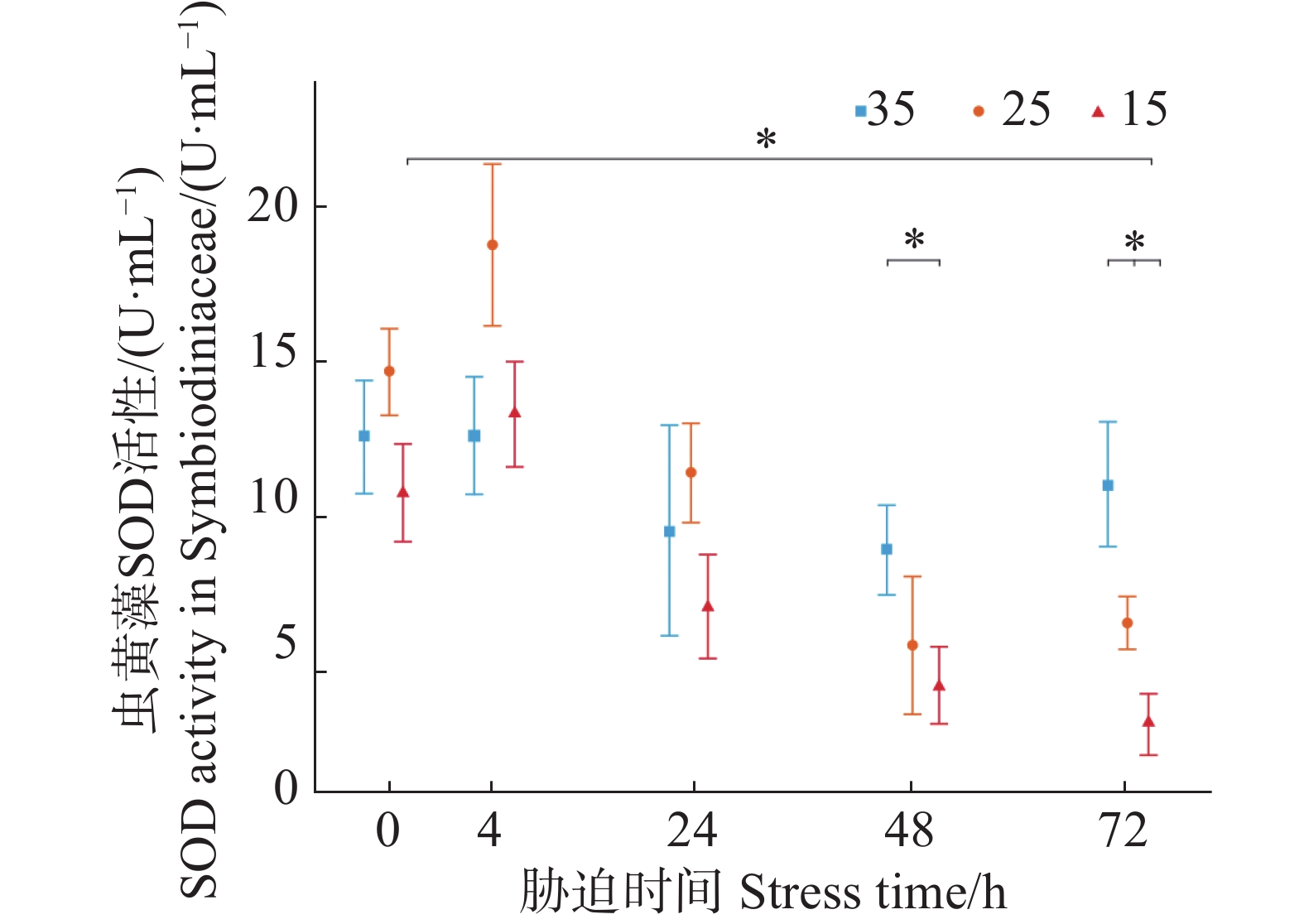
如图3所示,3个盐度梯度(35、25、15)的虫黄藻SOD初始活力分别为(12.57±1.83)、(14.68±1.39)、(10.74±1.59 )U·mL−1,低盐胁迫实验组的平均值在4 h时要高于对照组但两者的差异性并不显著。在随后的24 h与48 h,低盐胁迫实验组呈现不断下降趋势;盐度25实验组在72 h时与48 h持平,而盐度15实验组则仍在下降。在胁迫48、72 h时胁迫实验组中盐度25和15的SOD活力均小于初始阶段的虫黄藻的SOD活力(P < 0.05),且均小于对照组中盐度35的SOD在胁迫了48、72 h的虫黄藻SOD活力。
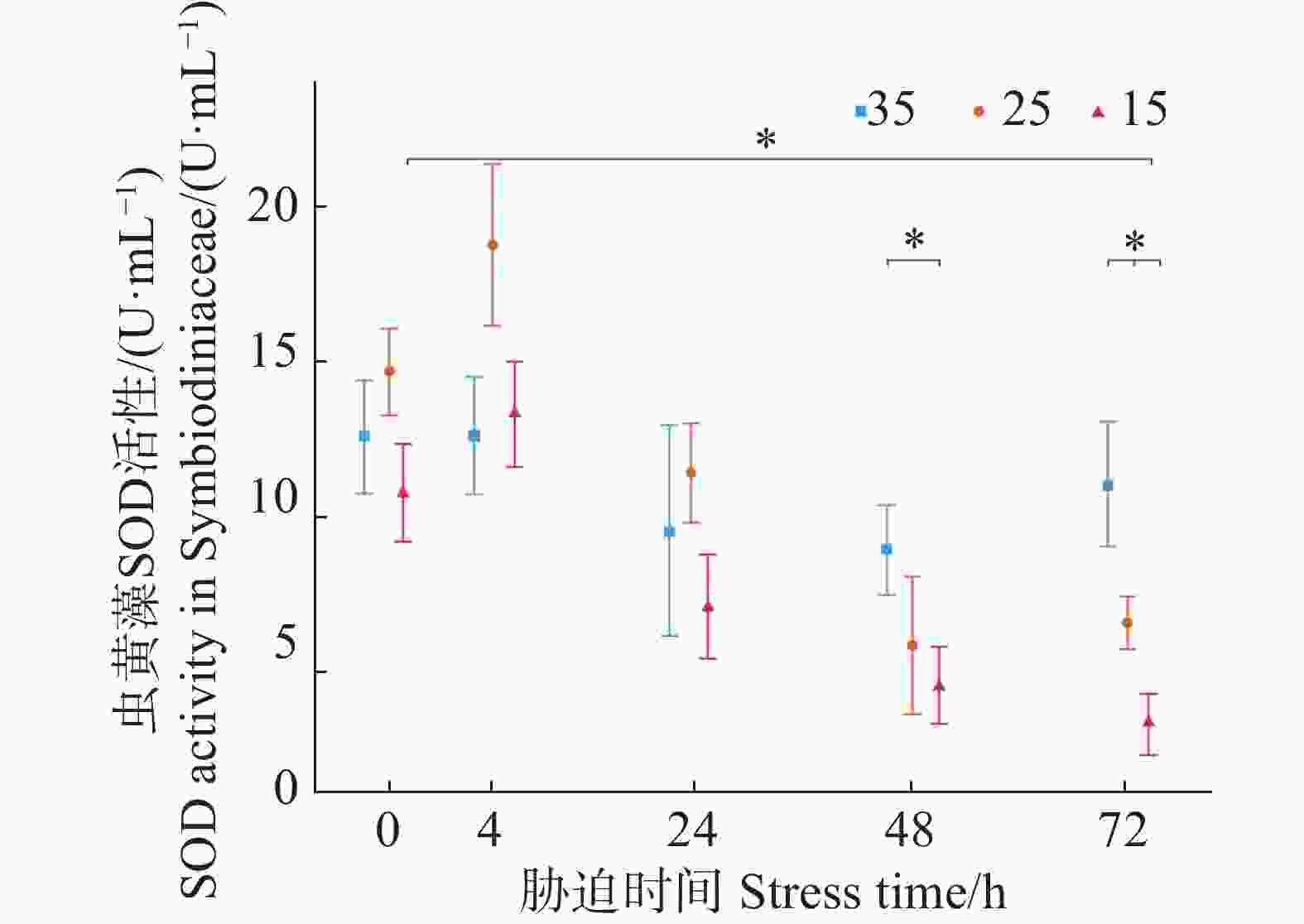
图 3 不同盐度条件下虫黄藻超氧化物歧化酶SOD活力随胁迫时间的变化
Figure 3. Changes in SOD activity in Symbiodiniaceae under different salinity conditions over stress time.
-
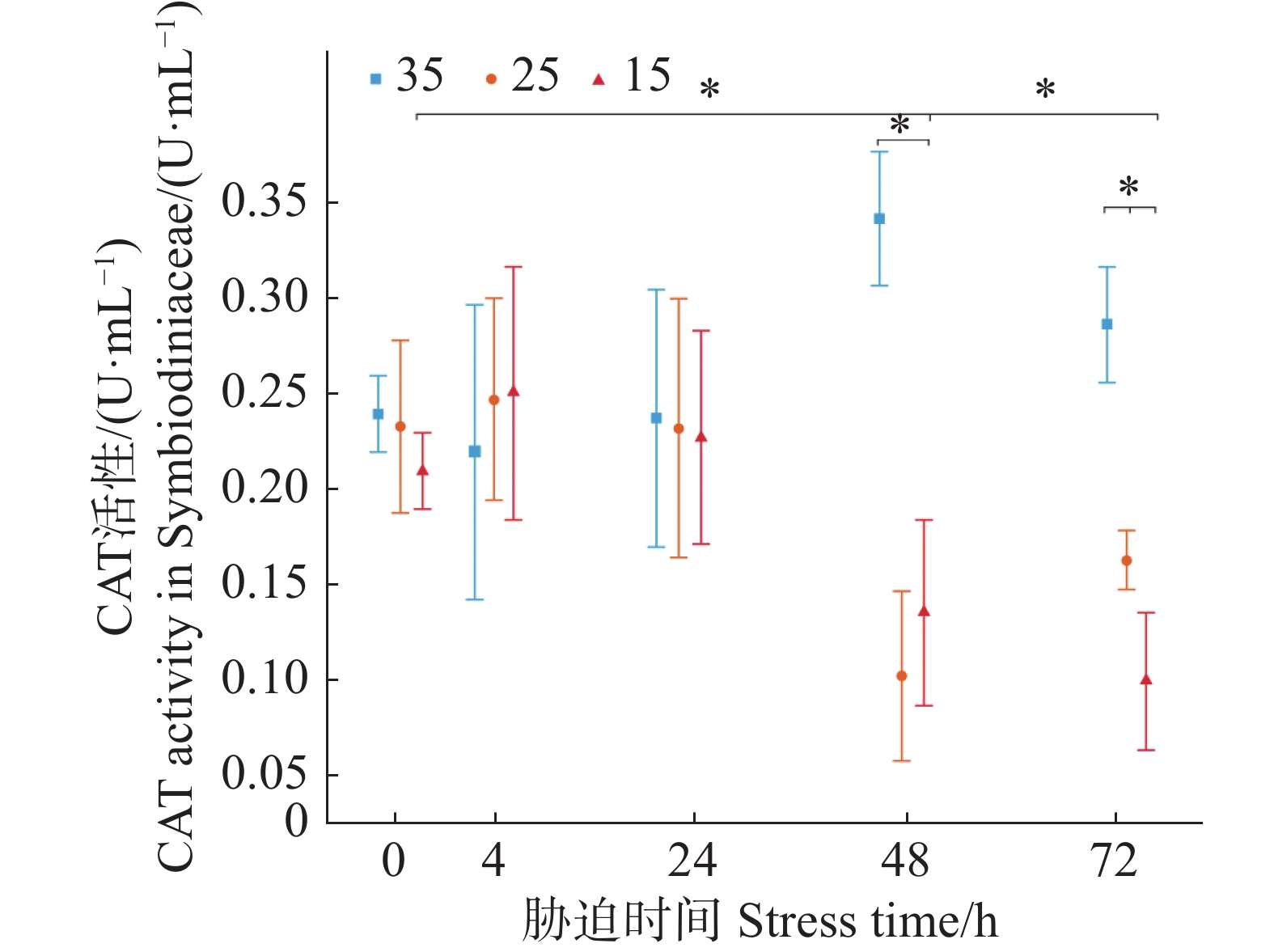
如图4所示,3个盐度梯度(35、25、15)的虫黄藻CAT初始活力分别为(0.24±0.02)、(0.23±0.04)、(0.21±0.02)U·mL−1,低盐胁迫实验组的平均值在4 h要高于对照组,但两者的差异不显著。在随后的24与48 h,低盐胁迫实验组则呈现不断下降趋势。在72 h时,25盐度实验组CAT活性略有上升,而盐度15实验组仍在下降。在胁迫48、72 h时,胁迫实验组中盐度25和15的CAT活力均显著小于正常培养盐度35条件下对照组虫黄藻的CAT活力(P < 0.05)。
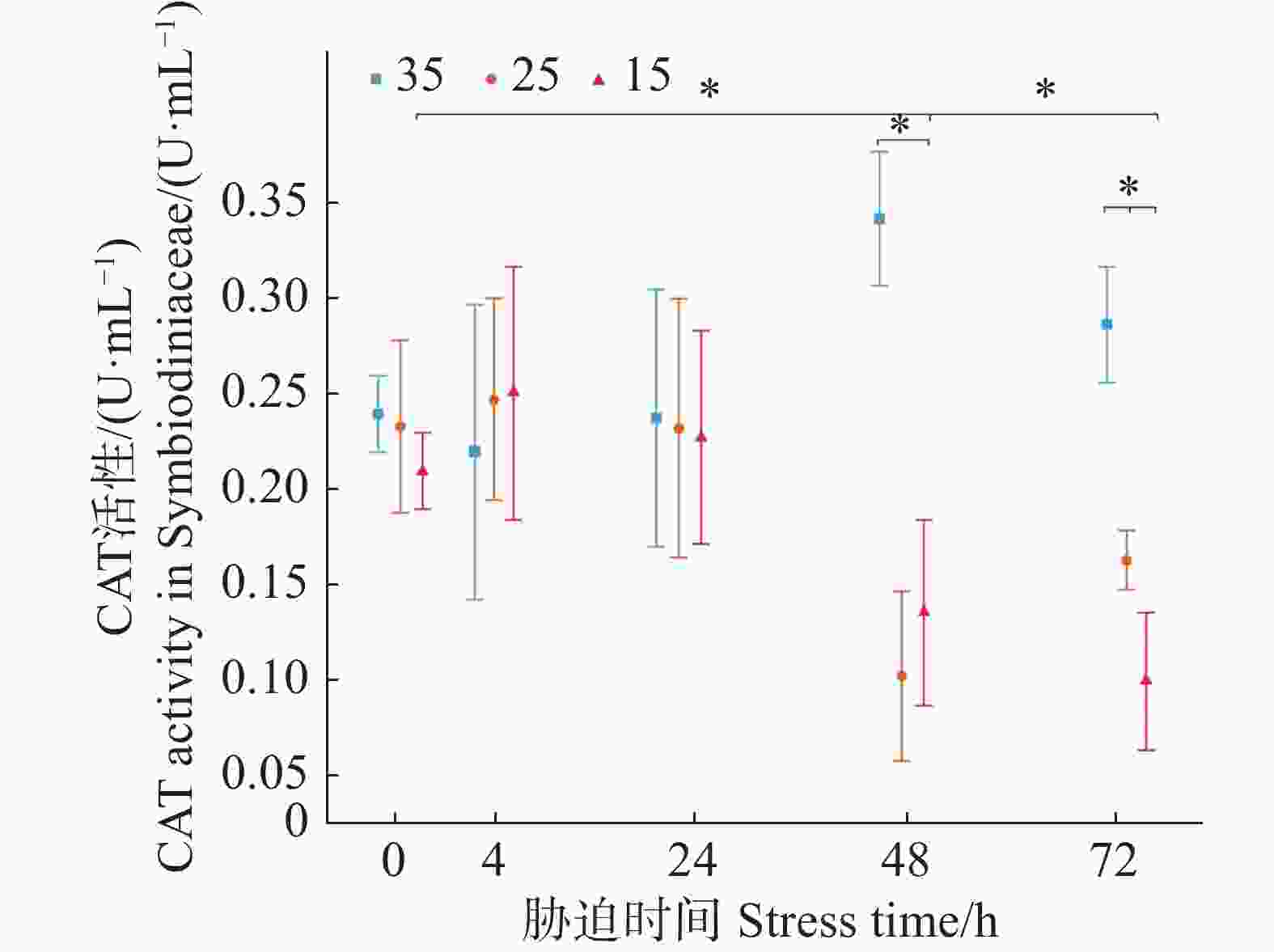
图 4 不同盐度条件下虫黄藻过氧化氢酶CAT活力随胁迫时间的变化
Figure 4. Changes in CAT activity of Symbiodiniaceae under varying salinity conditions over stress time
-
本研究结果表明,低盐胁迫显著抑制了虫黄藻的生长并导致其抗氧化系统崩溃,15的盐度环境已超过其耐受阈值。具体而言,在15盐度下,E. voratum的细胞密度在48 h后呈现显著负增长,同时伴随单个虫黄藻细胞的叶绿素a浓度持续下降,表明低盐环境严重抑制了其光合作用能力,这与Sakami[17]的研究一致,即在高温加低盐胁迫条件下会显著抑制虫黄藻的光合作用,结合本研究中单个虫黄藻细胞Chl a浓度的下降,推测其机制可能涉及光合系统Ⅱ的损伤。此外,Chl a的降解可能与ROS诱导的脂质过氧化有关[23],低盐胁迫下细胞膜透性增加,加速了叶绿体结构的破坏,这进一步解释了光合能力下降的机制。且SOD与CAT活性大幅降低(P < 0.05),说明低盐胁迫引发的渗透压失衡导致虫黄藻细胞内ROS过量积累,而抗氧化系统(如SOD和CAT)的活性下降进一步削弱了ROS清除能力,最终加剧细胞损伤。值得注意的是,在25盐度下,尽管SOD与CAT活性初期短暂升高(4 h),但随着胁迫时间延长(24 h后),酶活性仍持续下降,表明虫黄藻的短期应激机制不足以应对长期低盐压力。这些发现首次明确了E. voratum的盐度耐受下限为15,并提出了其抗氧化系统的“双阶段响应”模式,短期激活与长期失效的共存。这一机制不仅解释了低盐环境下珊瑚白化的关键驱动因素(虫黄藻功能崩溃先于宿主损伤),也暗示非优势虫黄藻类群(如E. voratum)可能在极端盐度波动区域中通过快速响应策略维持局部生态功能,为珊瑚礁适应性研究提供了新视角。
以往的研究将盐度胁迫下珊瑚产生白化主要归因于盐度对其体内的虫黄藻光合作用的影响[11 − 13]。在本研究中,观察到15盐度条件下单个虫黄藻细胞Chla的降低趋势且差异显著(P<0.05)。结合以往的研究,导致E. voratum的Chl a浓度下降的主要原因可以分为外在因素与内在因素。首先是外在因素,珊瑚受到盐度胁迫时,会发生水螅体回缩的行为,从而减少组织与周围海水接触的表面积,可能会导致到达虫黄藻的有效光合辐射大大减少,进而影响虫黄藻的Chl a浓度[11 − 12];其次是内在因素,可能是E. voratum受到压力,活性氧大量积累进而加速了色素降解[23 − 24],也可能是低盐环境导致细胞器产生损伤甚至细胞凋亡[14 − 15]。
虫黄藻在低盐压力胁迫下会产生氧化应激反应,加速产生具有破坏性的活性氧分子(ROS),比如过氧化氢(H2O2)、羟基自由基(OH−)、超氧阴离子自由基(
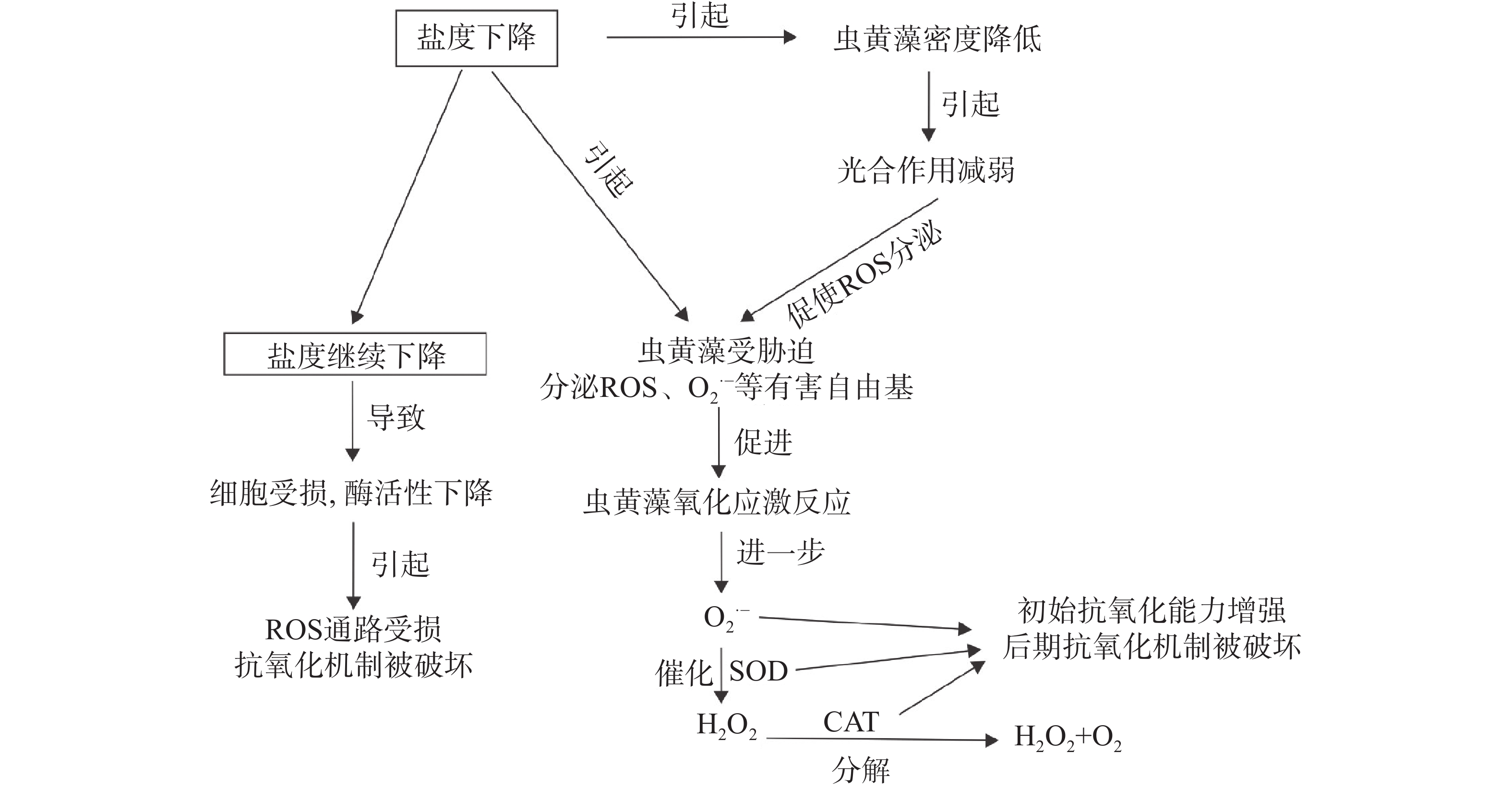
$ {{\mathrm{O}}_{2}}^{\bullet -} $ )等[16,25 − 27],其次是光合作用和细胞呼吸的常见副产物[28 − 29],但当细胞内的ROS浓度累积到超过阈值则会引起细胞氧化损伤和细胞死亡[27]。与此同时,为了控制细胞水平的毒性,保护细胞免受氧化损伤,细胞内的抗氧化机制也会被触发[30 − 31],包括酶与非酶的抗氧化剂,比如超氧化物歧化酶(SOD)、过氧化氢酶(CAT)、过氧化物酶(POD),以及一些小分子,如维生素C和硫醇类抗氧化剂[27,32 − 33],其中的SOD与CAT则作为其中的2种抗氧化剂在这一过程中起到重要作用。SOD通过催化$ {{\mathrm{O}}_{2}}^{\bullet-} $ 转化成水和H2O2提供了抗氧化的第一道防线[34],CAT则会将H2O2还原成水和氧气[35]。因此,SOD与CAT的活性作为氧化应激参数已被广泛用作低盐胁迫下细胞应激的生物学指标[36 − 37],通过对氧化酶活性的测定,能够反映取样时刻细胞内ROS和抗氧化机制之间的平衡状态。在本研究中,25与15的低盐度实验组E. voratum的SOD与CAT活性随着时间变化趋势一致,表现为初始阶段均高于35盐度对照组活性水平,并且随着时间的推移,活性均不断下降至低于对照组活性水平。初始阶段抗氧化酶的上升可能是由于渗透调节的需求导致代谢率升高进而导致虫黄藻体内ROS的增加所导致的[38],即盐度下降激活了E. voratum细胞的氧化应激反应。而随后的酶活下降可能是低盐条件下,细胞受损严重且活性丧失,细胞内产生的ROS通路机制严重损坏,从而导致ROS水平下降[27],或是在ROS上升期间,产生抗氧化酶的通路受损,又或是其他的抗氧化物可能承担了受损或者未受损通路产生的抗氧化酶的部分功能[32]。综合来看,25与15的盐度胁迫下的E. voratum氧化或者抗氧化机制可能已经被严重破坏。综上所述,面对低盐胁迫,E. voratum的响应模式如图5所示。低盐胁迫下,虫黄藻E. voratum的生长受到抑制,光合作用减弱,产生氧化应激反应,出现活性氧分子等有害自由基积累,抗氧化酶SOD与CAT活性初期升高,增强了虫黄藻的抗氧化能力,后期抗氧化机制被破坏。极端低盐环境下,细胞受损,酶活性下降,ROS通路受损,导致抗氧化机制被破坏。
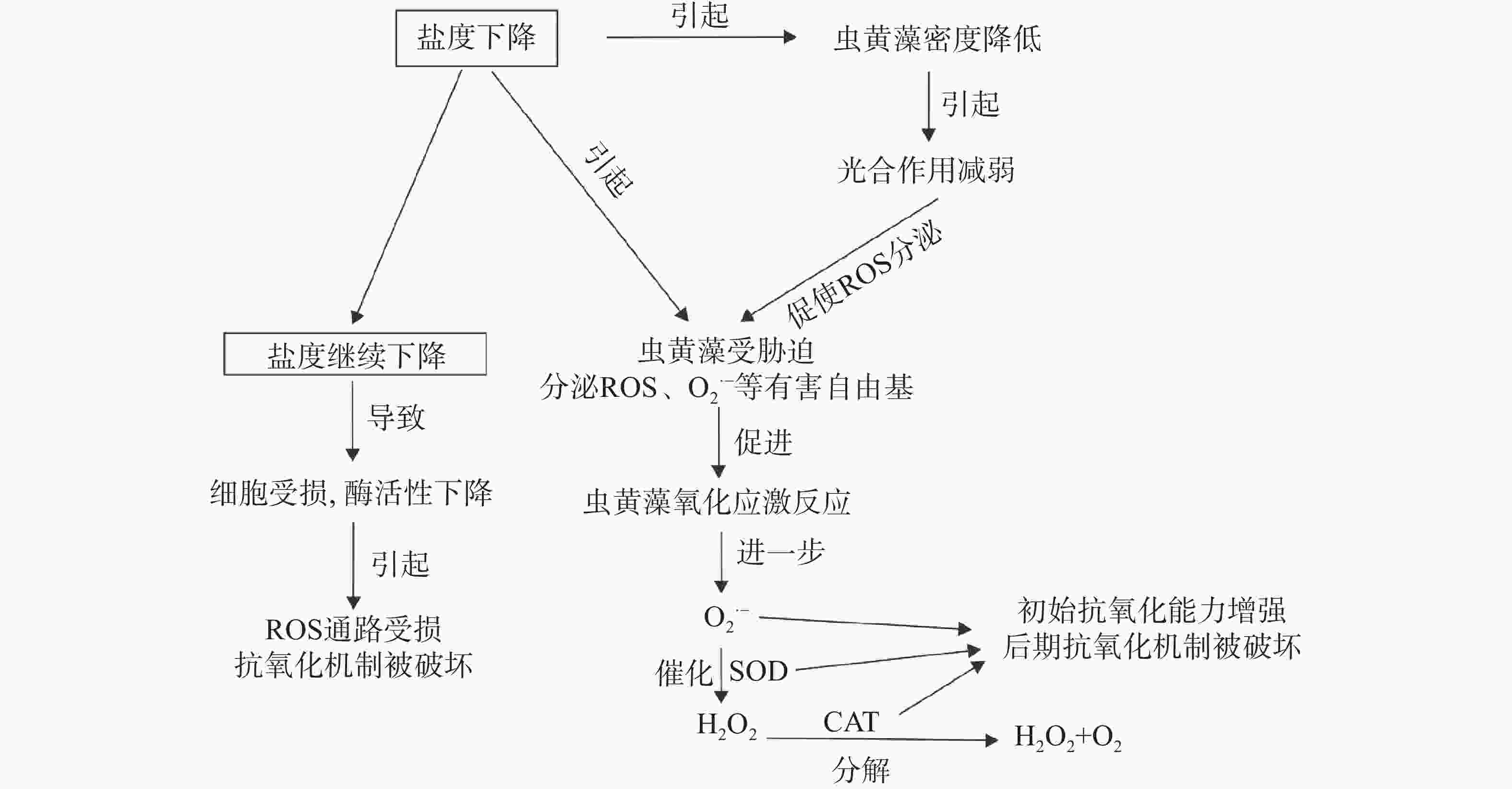
图 5 E. voratum对低盐胁迫的响应模式
Figure 5. Response patterns of E. voratum to low-salt stress
因此,低盐胁迫对离体虫黄藻造成的负面影响,包括虫黄藻密度降低、单个虫黄藻细胞Chl a浓度减少和关键抗氧化酶活性下降等,其他虫黄藻系群和共生藻类也可能会表现出类似的反应,因为盐度、Chl a浓度和抗氧化酶活性在虫黄藻生长中起着基本的生理作用。已有研究证明,低盐胁迫对紫色巴里沙软珊瑚共生虫黄藻的超氧化物歧化酶和过氧化氢酶具有抑制作用[39],但不同种类的虫黄藻对盐度变化可能表现出不同的耐受性,然而目前对虫黄藻低盐胁迫响应机制的研究较少,具体不同系群虫黄藻对低盐胁迫的响应机制有待进一步研究。本研究为理解虫黄藻种群对低盐胁迫的响应和适应机制提供了数据支持,有助于深入探究虫黄藻在低盐条件下的生理变化,为未来构建和培育适应低盐环境的虫黄藻物种提供了参考依据。本研究首次揭示了离体虫黄藻的低盐响应机制,但仍存在以下局限:1)离体与共生状态下虫黄藻的胁迫响应差异需通过共培养实验进一步验证;2)不同虫黄藻属(如Cladocopium、Durusdinium)对低盐的响应可能存在差异,需扩展比较研究以完善理论框架。希望下一步开展对Breviolum、Cladocopium、Durusdinium等不同属虫黄藻的低盐胁迫研究,完善低盐条件对具体虫黄藻系群的影响及其不同的响应机制。
-
本研究以低盐作为环境压力源,研究了低盐胁迫下虫黄藻(E. voratum)的生理生化反应。基于高通量测序,本研究进行实验的虫黄藻样品中E. voratum的相对丰度达到(99.971±0.032)%。盐度为25与15的低盐胁迫下,E. voratum均表现出生长受到抑制出现负增长趋势,抗氧化酶SOD和CAT活性的下降。15的盐度条件下已经限制了细胞分裂,甚至出现细胞死亡,表明这些盐度条件已经超过了E. voratum对盐度耐受的阈值。本研究通过离体培养实验,初步揭示了E. voratum对低盐胁迫的生理响应机制,为虫黄藻种群的耐盐性研究提供了基础数据。然而,离体状态与共生状态下的胁迫响应可能存在显著差异,未来研究需结合共生体系,进一步探究低盐胁迫对虫黄藻−珊瑚共生关系的综合影响,为珊瑚礁生态系统的保护与修复提供更全面的科学依据。
Physiological response of coral Symbiodiniaceae to low-salt stress
-
摘要: 为了探究虫黄藻(Effrenium voratum)在低盐胁迫条件下的生理变化,以离体培养的虫黄藻为对象,对虫黄藻进行高通量测序获得其相对丰度,开展低盐胁迫实验,研究低盐环境对E. voratum的生长状况、单个虫黄藻Chl a浓度、超氧化物歧化酶(SOD)和过氧化氢酶(CAT)活力的生理影响。结果显示:1)35盐度下,E. voratum的细胞密度稳定增长,表明其生长状态良好;25的低盐胁迫下,其生长受抑制但仍能维持生存。15的低盐胁迫下,其细胞密度在48 h后显著下降,呈负增长趋势,表明其生长受到严重抑制。2)15的低盐胁迫下,E. voratum的单个虫黄藻叶绿素a浓度在4 h后持续下降,表明其光合作用能力受到显著抑制,趋势明显。3)25和15的低盐胁迫下,E. voratum的SOD活性和CAT活性在4 h内有所上升,但在24 h后持续下降,表明其抗氧化能力初期有所增强,但随胁迫时间的延长,抗氧化能力逐渐减弱。Abstract: To investigate the physiological responses of Symbiodiniaceae under low-salinity stress, high-throughput sequencing was conducted on isolated culture of Effrenium voratum to obtain its relative abundance, and low-salinity stress experiments were performed on E. voratum for systematical analysis of the effects of low-salinity environments on the growth status, chlorophyll a (Chl a) concentration per symbiodiniaceae, superoxide dismutase (SOD) and catalase (CAT) activities of E. voratum. The results demonstrated that the cell density of E. voratum cultured under 35 salinity showed stable growth, indicating favorable growth at this level of salinity. At 25 salinity, E. voratum was inhibited in growth but could maintain survival. Severe growth inhibition occurred at 15 salinity, with cell density significantly declining after 48 hours, exhibiting a negative growth trend. Under 15 salinity stress, the Chl a concentration per symbiodiniaceae progressively decreased after 4 hours, suggesting significant suppression of photosynthetic capacity. Both SOD and CAT activities of E. voratum initially increased within 4 hours under 25 and 15 salinity stresses, but showed a continuous decline after 24 hours, revealing transient enhancement of antioxidant capacity at the early stages followed by progressive weakening with prolonged stress exposure.
-
Key words:
- Effrenium voratum /
- low-salt stress /
- physiological response /
- next-generation sequencing
-
图 2 不同盐度条件下单个虫黄藻的Chl a含量随胁迫时间的变化
Fig. 2 Changes in Chl a content of individual Symbiodiniaceae cells under different salinity conditions over stress time
图 3 不同盐度条件下虫黄藻超氧化物歧化酶SOD活力随胁迫时间的变化
Fig. 3 Changes in SOD activity in Symbiodiniaceae under different salinity conditions over stress time.
图 4 不同盐度条件下虫黄藻过氧化氢酶CAT活力随胁迫时间的变化
Fig. 4 Changes in CAT activity of Symbiodiniaceae under varying salinity conditions over stress time
表 1 离体培养虫黄藻E. voratum藻液样品高通量测序结果
Table 1 High-throughput sequencing of the algal liquid of the dinoflagellate E. voratum cultured in vitro
% 虫黄藻物种
Taxon样本1相对丰度
Sample1
Relative abundance样本2相对丰度
Sample2
Relative abundance样本3相对丰度
Sample3
Relative abundance均值
MeanSD Effrenium voratum 99.988 99.934 99.991 99.971 0.032 Others 0.011 0.066 0.009  下载: 导出CSV
下载: 导出CSV
-
[1] 李宇亮, 高宇, 郑新庆, 等. 珊瑚礁生态系统功能产生机制及评价方法[J]. 应用海洋学学报, 2022, 41(3): 475 − 482. doi: 10.3969/J.ISSN.2095-4972.2022.03.012 [2] CARPENTER K E, ABRAR M, AEBY G, et al. One-third of reef-building corals face elevated extinction risk from climate change and local impacts[J]. Science, 2008, 321(5888): 560 − 563. doi: 10.1126/science.1159196 [3] 陈燕, 李成才, 晁飞飞, 等. 环境因子对造礁石珊瑚白化影响的研究进展及思考[J]. 黑龙江科技信息, 2016(2): 129 − 130. doi: 10.3969/j.issn.1673-1328.2016.02.114 [4] MOBERG F, NYSTRÖM M, KAUTSKY N, et al. Effects of reduced salinity on the rates of photosynthesis and respiration in the hermatypic corals Porites lutea and Pocillopora damicornis[J]. Marine Ecology Progress Series, 1997, 157: 53 − 59. doi: 10.3354/meps157053 [5] HOEGH-GULDBERG O. Climate change, coral bleaching and the future of the world’s coral reefs[J]. Marine and Freshwater Research, 1999, 50(8): 839 − 866. doi: 10.1071/MF99078 [6] BERKELMANS R, OLIVER J K. Large-scale bleaching of corals on the great barrier reef[J]. Coral Reefs, 1999, 18(1): 55 − 60. doi: 10.1007/s003380050154 [7] HOEGH-GULDBERG O, SMITH G J. The effect of sudden changes in temperature, light and salinity on the population density and export of zooxanthellae from the reef corals Stylophora pistillata Esper and Seriatopora hystrix Dana[J]. Journal of Experimental Marine Biology and Ecology, 1989, 129(3): 279 − 303. doi: 10.1016/0022-0981(89)90109-3 [8] HAAPKYLÄ J, UNSWORTH R K F, FLAVELL M, et al. Seasonal rainfall and runoff promote coral disease on an inshore reef[J]. PLoS One, 2011, 6(2): e16893. doi: 10.1371/journal.pone.0016893 [9] 李泽鹏. 主要环境因子对滨珊瑚的胁迫作用研究[D]. 湛江: 广东海洋大学, 2012. doi: 10.7666/d.Y2215224 [10] FALKOWSKI P G, DUBINSKY Z, MUSCATINE L, et al. Light and the bioenergetics of a symbiotic coral[J]. BioScience, 1984, 34(11): 705 − 709. doi: 10.2307/1309663 [11] MUTHIGA N A, SZMANT A M. The effects of salinity stress on the rates of aerobic respiration and photosynthesis in the hermatypic coral Siderastrea siderea[J]. The Biological Bulletin, 1987, 173(3): 539 − 551. doi: 10.2307/1541699 [12] MANZELLO D, LIRMAN D. The photosynthetic resilience of Porites furcata to salinity disturbance[J]. Coral Reefs, 2003, 22(4): 537 − 540. doi: 10.1007/s00338-003-0327-0 [13] KUANUI P, CHAVANICH S, VIYAKARN V, et al. Effects of temperature and salinity on survival rate of cultured corals and photosynthetic efficiency of zooxanthellae in coral tissues[J]. Ocean Science Journal, 2015, 50(2): 263 − 268. doi: 10.1007/s12601-015-0023-3 [14] LOBBAN C S, HARRISON P J. Seaweed Ecology and Physiology[M]. Cambridge: Cambridge University Press, 1994: 210−240. [15] KERSWELL A P, JONES R J. Effects of hypo-osmosis on the coral Stylophora pistillata: nature and cause of ‘low-salinity bleaching’[J]. Marine Ecology Progress Series, 2003, 253: 145 − 154. doi: 10.3354/meps253145 [16] DOWNS C A, KRAMARSKY-WINTER E, WOODLEY C M, et al. Cellular pathology and histopathology of hypo-salinity exposure on the coral Stylophora pistillata[J]. Science of the Total Environment, 2009, 407(17): 4838 − 4851. doi: 10.1016/j.scitotenv.2009.05.015 [17] SAKAMI T. Effects of temperature, irradiance, salinity and inorganic nitrogen concentration on coral zooxanthellae in culture[J]. Fisheries Science, 2000, 66(6): 1006 − 1013. doi: 10.1046/j.1444-2906.2000.00162.x [18] WANG J, CHEN J Q, WANG S Y, et al. Monoclonal culture and characterization of Symbiodiniaceae C1 strain from the scleractinian coral Galaxea fascicularis[J]. Frontiers in Physiology, 2021, 11: 621111. doi: 10.3389/fphys.2020.621111 [19] 龙超, 罗肇河, 韦章良, 等. 海南三亚鹿回头虫黄藻(Effrenium voratum)的形态学和系统发育学研究[J]. 热带海洋学报, 2021, 40(4): 35 − 43. doi: 10.11978/2020102 [20] LAJEUNESSE T C, PARKINSON J E, GABRIELSON P W, et al. Systematic revision of Symbiodiniaceae highlights the antiquity and diversity of coral endosymbionts[J]. Current Biology, 2018, 28(16): 2570-2580. e6. doi: 10.1016/j.cub.2018.07.008 [21] KANG N S, KIM E S, LEE J A, et al. First report of the dinoflagellate genus Effrenium in the East Sea of Korea: morphological, genetic, and fatty acid characteristics[J]. Sustainability, 2020, 12(9): 3928. doi: 10.3390/su12093928 [22] 刘旭. 造礁石珊瑚对温度胁迫的响应机制研究[D]. 南宁: 广西大学, 2020. doi: 10.27034/d.cnki.ggxiu.2020.001544 [23] XU Z Z, JIANG Y L, ZHOU G S. Response and adaptation of photosynthesis, respiration, and antioxidant systems to elevated CO2 with environmental stress in plants[J]. Frontiers in Plant Science, 2015, 6: 701. doi: 10.3389/fpls.2015.00701 [24] MCCOY S J, KAMENOS N A. Coralline algae (Rhodophyta) in a changing world: integrating ecological, physiological, and geochemical responses to global change[J]. Journal of Phycology, 2015, 51(1): 6 − 24. doi: 10.1111/jpy.12262 [25] LESSER M P. OXIDATIVE STRESS IN MARINE ENVIRONMENTS: biochemistry and physiological ecology[J]. Annual Review of Physiology, 2006, 68: 253 − 278. doi: 10.1146/annurev.physiol.68.040104.110001 [26] DOWNS C A, FAUTH J E, HALAS J C, et al. Oxidative stress and seasonal coral bleaching[J]. Free Radical Biology and Medicine, 2002, 33(4): 533 − 543. doi: 10.1016/S0891-5849(02)00907-3 [27] 杨晓红. 高温及营养缺乏诱导虫黄藻氧化胁迫的初步研究[D]. 厦门: 厦门大学, 2020. doi: 10.27424/d.cnki.gxmdu.2020.000447 [28] ASADA K. THE WATER-WATER CYCLE IN CHLOROPLASTS: scavenging of active oxygens and dissipation of excess photons[J]. Annual Review of Plant Physiology and Plant Molecular Biology, 1999, 50: 601 − 639. doi: 10.1146/annurev.arplant.50.1.601 [29] BYCZKOWSKI J Z, GESSNER T. Biological role of superoxide ion-radical[J]. International Journal of Biochemistry, 1988, 20(6): 569 − 580. doi: 10.1016/0020-711X(88)90095-X [30] CZIESIELSKI M J, SCHMIDT-ROACH S, ARANDA M. The past, present, and future of coral heat stress studies[J]. Ecology and Evolution, 2019, 9(17): 10055 − 10066. doi: 10.1002/ece3.5576 [31] 蒙林庆, 黄雯, 阳恩广, 等. 高温白化事件可提高涠洲岛澄黄滨珊瑚(Porites lutea)的耐热性[J]. 海洋学报, 2022, 44(8): 87 − 96. doi: 10.12284/hyxb2022126 [32] MISHRA S, IMLAY J. Why do bacteria use so many enzymes to scavenge hydrogen peroxide?[J]. Archives of Biochemistry and Biophysics, 2012, 525(2): 145 − 160. doi: 10.1016/j.abb.2012.04.014 [33] VANAPORN M, WAND M, MICHELL S L, et al. Superoxide dismutase C is required for intracellular survival and virulence of Burkholderia pseudomallei[J]. Microbiology, 2011, 157(Pt 8): 2392 − 2400. doi: 10.1099/mic.0.050823-0 [34] FRIDOVICH I. Superoxide radical and superoxide dismutases[M]//GILBERT D L. Oxygen and living processes. New York, NY: Springer, 1981: 250 − 272. doi: 10.1007/978-1-4612-5890-2_13 [35] VENN A A, LORAM J E, DOUGLAS A E. Photosynthetic symbioses in animals[J]. Journal of Experimental Botany, 2008, 59(5): 1069 − 1080. doi: 10.1093/jxb/erm328 [36] MCGINTY E S, PIECZONKA J, MYDLARZ L D. Variations in reactive oxygen release and antioxidant activity in multiple Symbiodinium types in response to elevated temperature[J]. Microbial Ecology, 2012, 64(4): 1000 − 1007. doi: 10.1007/s00248-012-0085-z [37] DIAS M, MADEIRA C, JOGEE N, et al. Oxidative stress on scleractinian coral fragments following exposure to high temperature and low salinity[J]. Ecological Indicators, 2019, 107: 105586. doi: 10.1016/j.ecolind.2019.105586 [38] FREIRE C A, WELKER A F, STOREY J M, et al. Oxidative stress in estuarine and intertidal environments (temperate and tropical)[M]//ABELE D, VÁZQUEZ-MEDINA J P, ZENTENO-SAVIN T. Oxidative stress in aquatic ecosystems. Chichester: Wiley-Blackwell, 2011: 41-57. doi: 10.1002/9781444345988.ch3 [39] DING D S, WANG S H, SUN W T, et al. The effect of feeding on Briareum violacea growth, survival and larval development under temperature and salinity stress[J]. Biology, 2022, 11(3): 410. doi: 10.3390/biology11030410 -

 点击查看大图
点击查看大图
计量
- 文章访问数: 1168
- HTML全文浏览量: 549
- PDF下载量: 25
- 被引次数: 0




