

 下载:
下载:
-
白粉菌(Powdery mildew)是一种专性活体寄生真菌,可以侵染多种植物,造成巨大的经济损失[1-2]。白粉菌与寄主相互作用的过程中,首先刺入寄主细胞壁形成吸器(haustorium),从寄主细胞内获取水分和营养物质。同时,白粉菌通过在吸器转录,翻译形成大量的效应蛋白(Effector proteins),分泌到寄主细胞内,从而实现在寄主体内的增殖和侵染[3-4]。目前,利用基因组、转录组测序及蛋白质组分析,通过对大麦白粉菌(Bgh,Blumeria graminis f. sp. Hordei)、小麦白粉菌(Bgt,Blumeria graminis f. sp. tritici)、拟南芥白粉菌(Gor,Golovinomyces orontii)、葡萄白粉菌(En,Erysiphe necator)和葫芦白粉菌(Pxa,Podosphaera xanthii)等进行预测,发现了一些潜在的效应蛋白[5]。大麦和小麦白粉菌编码的效应蛋白数量最多,通常是一些小的蛋白质,并且与已知蛋白同源性很低[6]。另外,大麦白粉菌形成的效应蛋白N端通常含有一个保守的结构域:第一个氨基酸是芳香族氨基酸(酪氨酸,苯丙氨酸或色氨酸),最后一个氨基酸是半胱氨酸(Y/F/WXC)[7]。然而,双子叶植物白粉菌编码的效应蛋白并不是都含有该保守结构域[8-9]。白粉菌是专性活体寄生,无法进行遗传操作,因此,目前对白粉菌效应蛋白功能的了解还非常少。最近,通过寄主诱导的基因沉默技术(HIGS,host-induced gene silencing),发现大麦白粉菌的一些效应蛋白在刺入寄主植物和形成吸器的过程中发挥毒性功能[10]。橡胶树白粉病是由橡胶树白粉菌(Oidium heveae)引发的一种真菌病害,对天然橡胶的生产造成了严重的经济损失。该病于1918年在印尼爪哇首次发现[11-13],迄今已遍布全球各橡胶种植区。由于橡胶树白粉菌与橡胶树很难进行遗传操作,因此严重阻碍了二者相互作用机理的研究。最近研究发现,橡胶树白粉菌在野生型拟南芥Col-0激发依赖于EDS1(enhanced disease susceptibility 1)和PAD4(Phytoalexin Deficient 4)的抗病反应,推测拟南芥TIR-NB-LRR(Toll-Interleukin1 Receptor-nucleotide binding-leucine-rich repeat)类抗性基因参与了对橡胶树白粉菌的识别过程[14]。
通过基因组和转录组测序分析,预测出橡胶树白粉菌含有133个潜在的效应蛋白[8],然而,这些效应蛋白的功能还是未知的。笔者克隆了橡胶树白粉菌潜在的效应蛋白基因OhEF 2 (Oidium heveae effector protein 2),并对该效应蛋白结构进行分析,构建在Col-0背景下的OhEF 2基因过表达转基因植株。结果表明,过表达植株对橡胶树白粉菌敏感性增加,并且有效降低了橡胶树白粉菌在拟南芥上诱导的胼胝体沉积和致病相关基因的表达。
-
植物材料:拟南芥野生型Col-0,Hevea brasiliensis 73397;菌株:O. Heveae HN1106[14],假单胞菌DC 3000。
-
橡胶树叶片古铜期接种白粉菌,8 d后剪取叶片,利用快速通用植物RNA提取试剂盒3.0(北京华越洋公司,产品编号:0416-50)提取总RNA。反转录试剂盒(北京华越洋公司,产品编号:HYY871)进行反转录,获得cDNA。设计用于扩增OhEF 2基因的引物OhEF 2-F和OhEF 2-R,进行PCR扩增。引物序列:OhEF 2-F,5′-AAACTCGAGCATCCTGGTCAGAAACGAGA-3′;OhEF 2-R,5′-AAATTCGAAGTTTCCAATAGTTTGAGCAAT-3′(Gene_id: OH_02339)[8]。利用限制性内切酶Xho I和BstB I(NEB公司)分别对PCR扩增片段和pER8[15]载体进行酶切。T4(NEB公司)连接酶进行连接,测序正确后转入农杆菌GV3101。利用花粉管侵染方法,侵染拟南芥野生型Col-0。收取拟南芥种子,在含有潮霉素的MS培养基中筛选,挑取长根的拟南芥植株,移栽土中。
-
生长4周后,喷洒雌激素,剪取1个叶片,放入1.5 mL离心管中,加入100 μL蛋白提取液(0.15 mol·L−1 KCl,50 mmol·L−1 HEPES-KOH,pH 7.5,1 mmol·L−1 EDTA,1 mmol·L−1 DTT,0.2% Triton-X 100,cocktail蛋白酶抑制剂),研碎后,12 000 r·min−1,4 ℃,离心10 min,取上清,加入loading buffer,进行SDA-PAGE电泳[15]。
利用湿转法在转膜电泳槽(伯乐)中,将蛋白条带转移至PVDF膜上,转膜液(7.58 g Tris,36 g Glycine,800 mL甲醇,加水至4 L),电压100伏,电泳1 h。5%的脱脂奶粉,室温封闭2 h,加入FLAG抗体(sigma),室温孵育2 h,加入HRP标记的羊抗鼠2抗,室温作用0.5 h,在黑暗条件下显影、定影。
-
将长40 cm,宽40 cm,孔径为50 μm的尼龙膜接种箱置于生长5周的拟南芥植株上方,用毛笔刷将橡胶树白粉菌从橡胶树叶片刷到接种箱上,白粉菌透过接种箱,均匀洒落在拟南芥叶片上。橡胶树白粉菌为橡胶树古铜期叶片接种10 d后的白粉菌孢子。接种1 d后,剪取叶片,置于脱色液:体积比(乙醇︰苯酚︰水︰乳酸= 2︰1︰1︰1)中,脱色过夜。次日,于考马斯亮蓝染色液(G250,浓度为6 g·L−1乙醇溶液)中染色30 s。脱色后,显微镜下进行观察。计数橡胶树白粉菌产生1根芽管和2根芽管的白粉菌孢子数目。利用计算公式:2根芽管孢子数目/(1根芽管孢子数目+2根芽管孢子数目),计算细胞进入率[14]。
-
利用接种箱在拟南芥叶片接种橡胶树白粉菌,接种2 d后,剪取叶片,于脱色液中脱色过夜。考马斯亮蓝染色30 s,显微镜观察,利用MIE3.1软件测量菌丝长度[14]。
-
利用接种箱在拟南芥叶片接种橡胶树白粉菌,接种10 d后,观察叶片发病症状并进行拍照。剪取叶片置于脱色液中脱色过夜,考马斯亮蓝染色液中染色30 s,计数单个孢子产生的分生孢子数[14]。
-
利用接种箱在拟南芥叶片接种橡胶树白粉菌,接种6 d后。剪取叶片,置于遇冷的研钵中,加入液氮,研磨3次。最后加入1 mL Invitrogen Trizol (Lot No. 66223),提取总RNA。经DNA酶消化后,加水溶解。利用Invitrogen RNA反转录试剂盒Ⅲ (Lot No. 696045)对总RNA进行反转录。荧光定量PCR检测采用SYBR Premix ExTaq Ⅱ(Lot No. AK2702)试剂,7500 ABI Real-time PCR Detection System,20 μL反应体系。检测程序为:预变性95 ℃ 30 s;变性95 ℃ 5 s、退火延伸62 ℃ 40 s,40个循环。内参基因ACTIN引物序列为:5′-TGGTGGAAGCACAGAAGTTG-3′;5′-GATCCATGTTTGGCTCCTTC-3′;PR1:5′-TACGCAGAACAACTAAGAGG-3′;5′-TCGTTCACATAATTCCCACG-3′[14]。
-
利用接种箱在拟南芥叶片接种橡胶树白粉菌,接种6 d后。将叶片置于脱色液,V水︰V甘油︰V乳酸︰V水饱和酚︰V乙醇=1︰1︰1︰1︰8,抽真空30 min,65 ℃放置60 min,10 min摇1次。50%酒精和水漂洗各漂洗1次。将叶片置于染色液(0.1 g·L−1苯胺蓝,150 mmol·L−1 K2HPO4,pH9.5)中染色60 min。于荧光显微镜紫外激发光下照相。采用Image J软件(
http://www.uhnresearch.ca/wcif )对0.1 mm2叶片面积内的胼胝体进行计数,每个样品共计数6片叶片,通过Student’s t test进行统计分析,差异极显著P<0.01,差异显著P<0.05[14]。 -
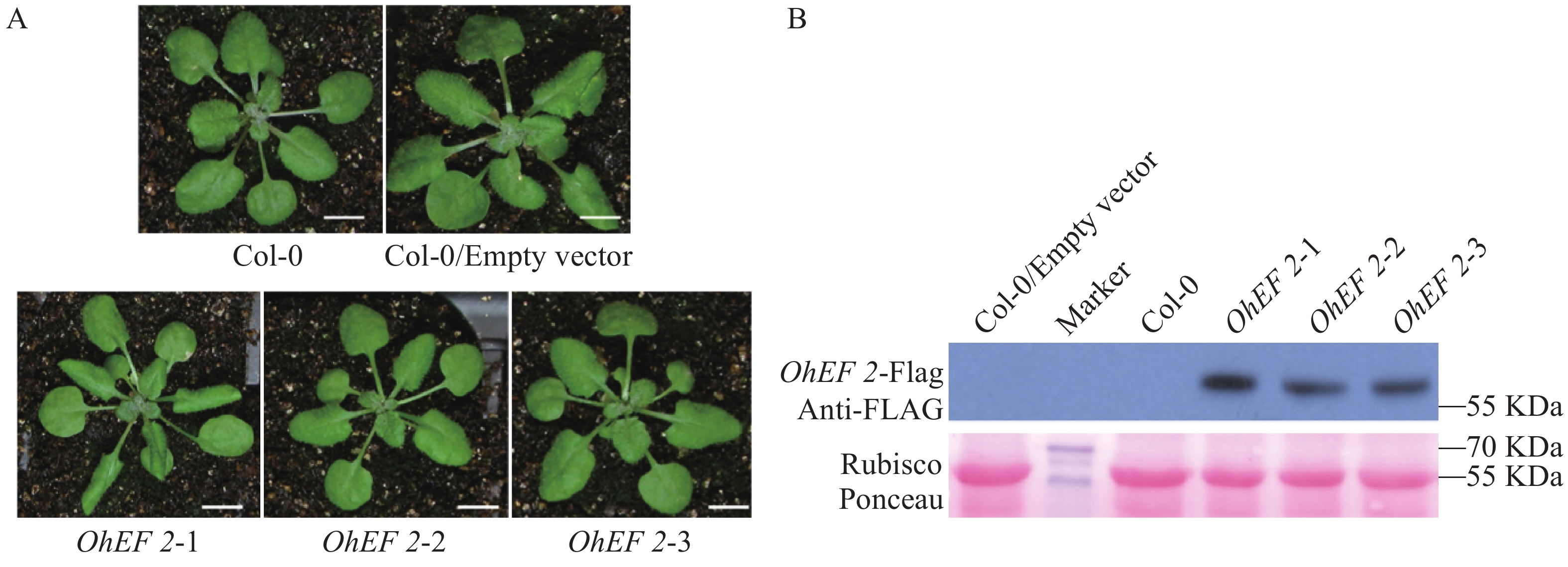
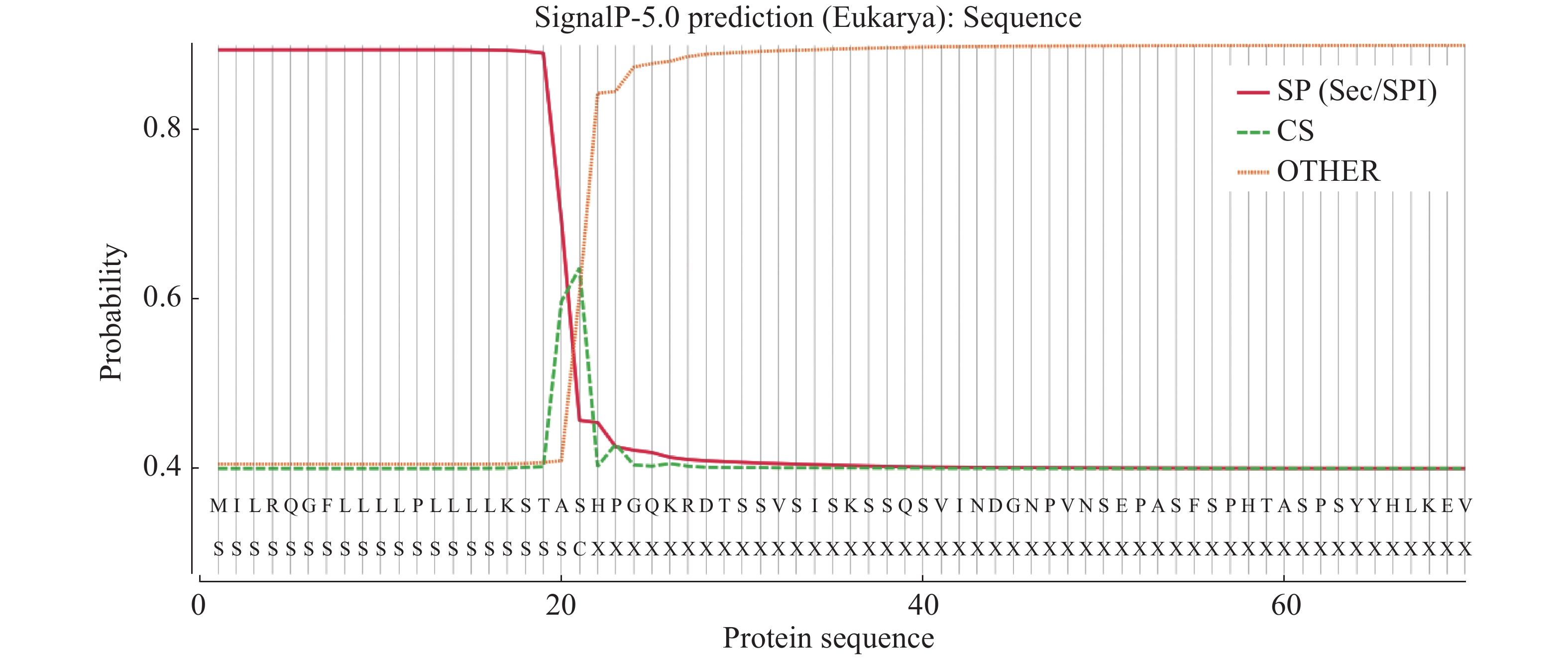
白粉菌通过分泌效应蛋白到植物体内,从而完成白粉菌的侵染过程。笔者克隆了其中的1个效应蛋白基因OhEF 2(Gene_id: OH_02339)[8],该蛋白编码543个氨基酸,信号肽分析发现1~21个氨基酸为其信号肽序列(图1)。为了研究该效应蛋白在植物体内的作用机理,设计引物,去除信号肽序列,经过PCR扩增、酶切后连入雌激素诱导的转基因载体pER8。利用潮霉素筛选,通过雌激素诱导和蛋白质印迹分析后,获得了3个在拟南芥Col-0背景下独立的OhEF 2转基因植株,分别命名为OhEF 2-1,OhEF 2-2和OhEF 2-3(图2A,B)。雌激素诱导6 d后,与野生型植物Col-0和空载体转基因植株对比,并未发现效应蛋白OhEF 2可以激发拟南芥细胞坏死和发黄的表型(图2B)。
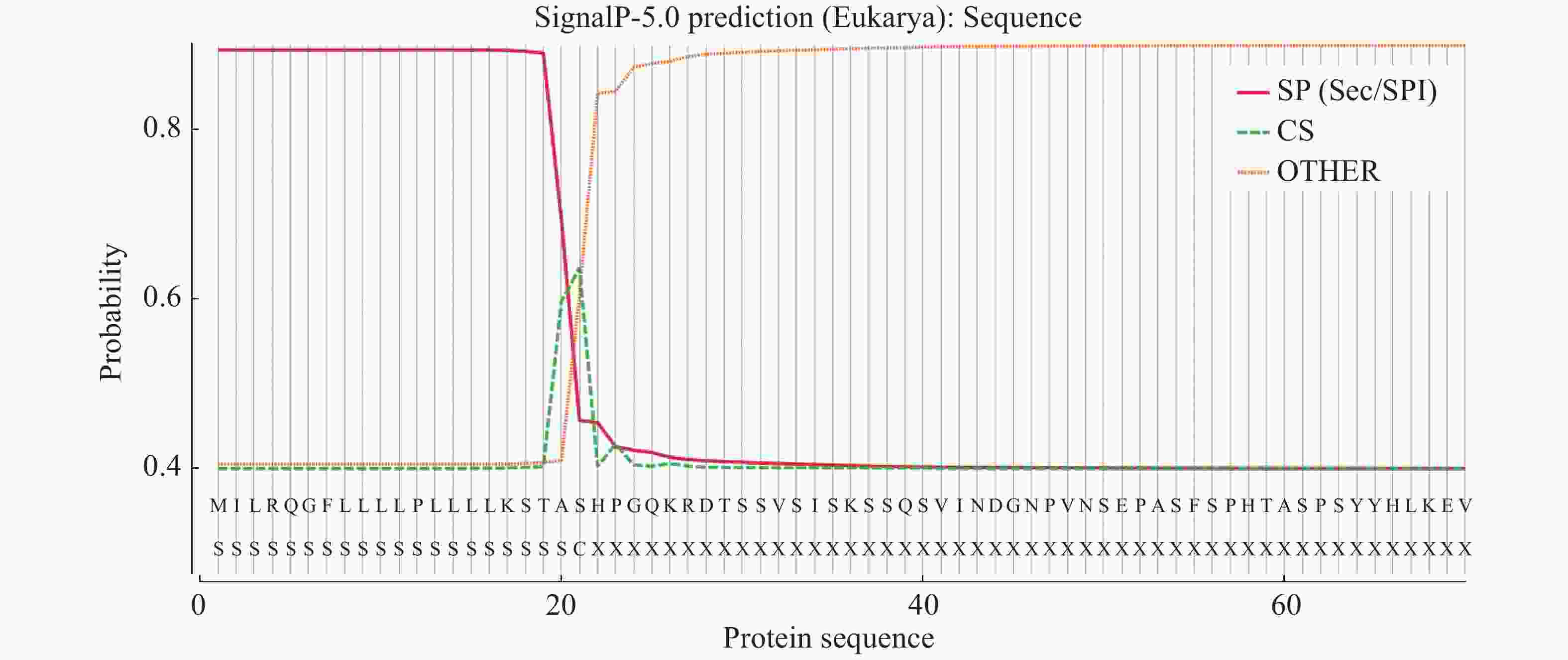
图 1 效应蛋白OhEF 2信号肽序列分析
Figure 1. Signal peptide analysis of effector protein OhEF 2

图 2 橡胶树白粉菌效应蛋白OhEF 2拟南芥转基因植株的构建
Figure 2. The construction of Oidium heveae effector OhEF 2 transgenic plants in Arabidopsis Col-0 background
-
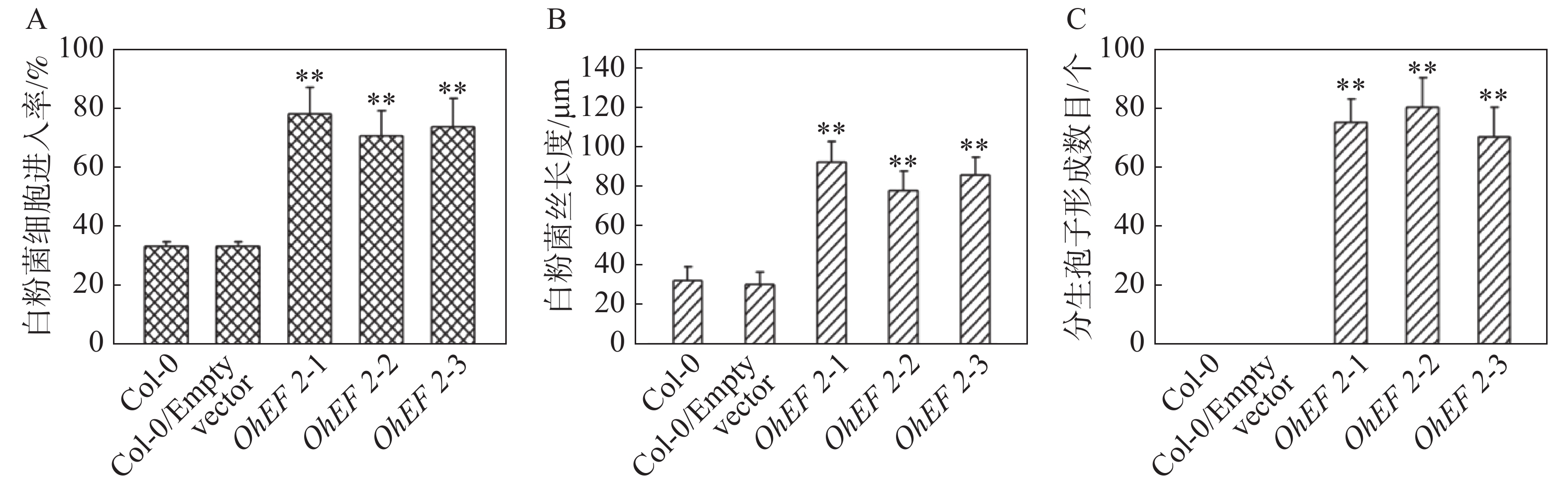
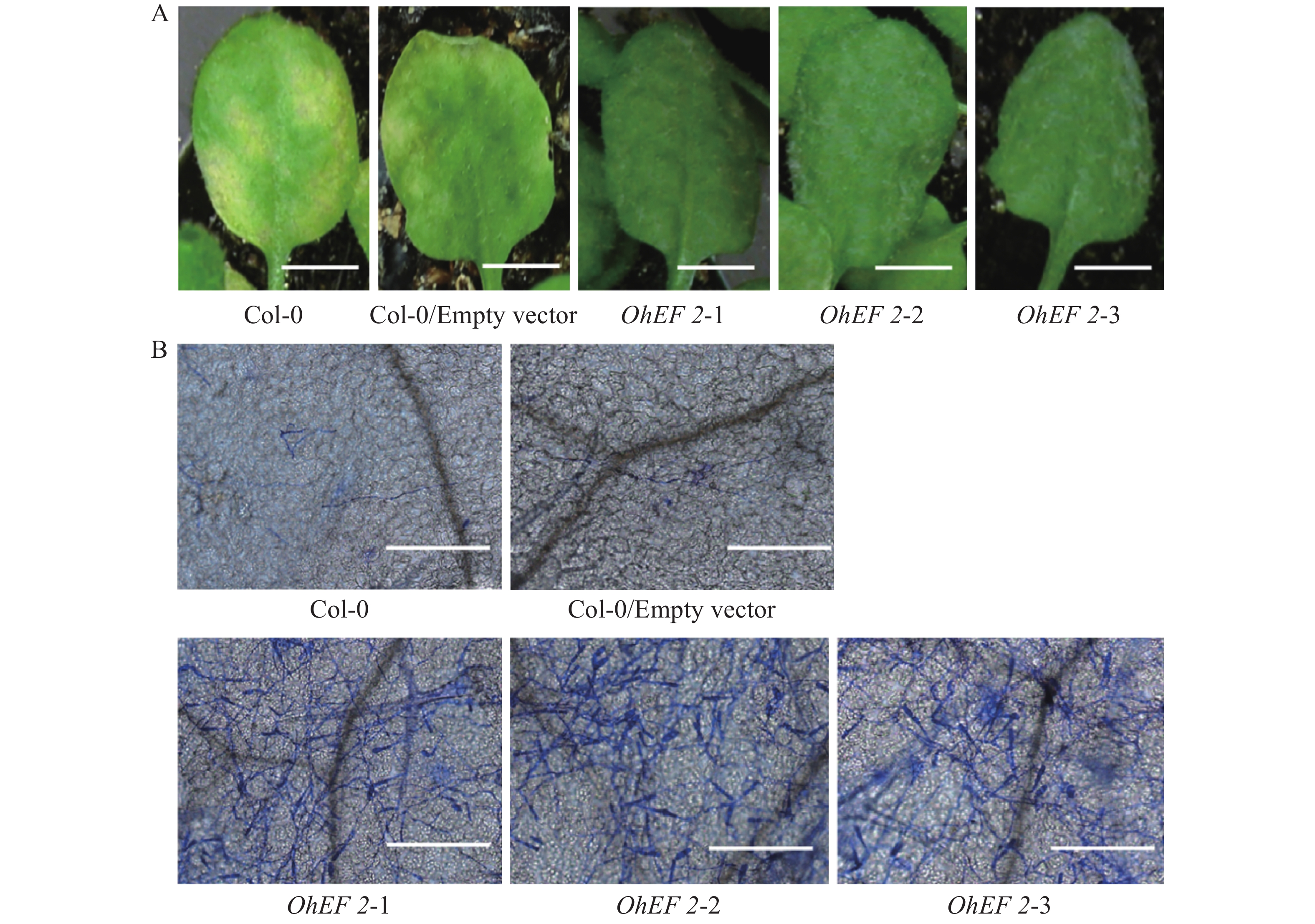
为了进一步研究效应蛋白OhEF 2在植物体内的作用机理,在野生型Col-0,空载体转基因植株和OhEF 2转基因植株OhEF 2-1,OhEF 2-2和OhEF 2-3上分别接种橡胶树白粉菌O.Heveae HN1106。接种1 d后,白粉菌细胞进入率统计分析结果表明,3个转基因植株显著高于野生型Col-0和空载体转基因植株(图3A),Col-0和空载体转基因植株白粉菌细胞进入率约为35%,无明显差异,3个OhEF 2转基因植株白粉菌细胞进入率约为75%,与野生型植物相比,差异极显著(P<0.01)。接种2 d后的菌丝生长长度计算结果表明,3个OhEF 2转基因植株同样显著高于野生型Col-0和空载体转基因植株(图3B),Col-0和空载体转基因植株菌丝长度约为30 μm,无明显差异,3个OhEF 2转基因植株菌丝长度约为90 μm,与野生型植物相比,差异极显著(P<0.01)。接种10 d后,3个OhEF 2转基因植株出现典型的白粉病病斑(图4A),野生型Col-0和空载体转基因植株并未出现白粉病病斑(图4A),而是表现出叶片发黄的症状(图4A)。分生孢子计数分析结果表明,橡胶树白粉菌不能在野生型Col-0和空载体转基因植株上产生分生孢子,而在3个OhEF 2转基因植株上形成了大量的分生孢子(图3C,4B)。综上所述,效应蛋白OhEF 2可以明显增强拟南芥对橡胶树白粉菌感病的表型。
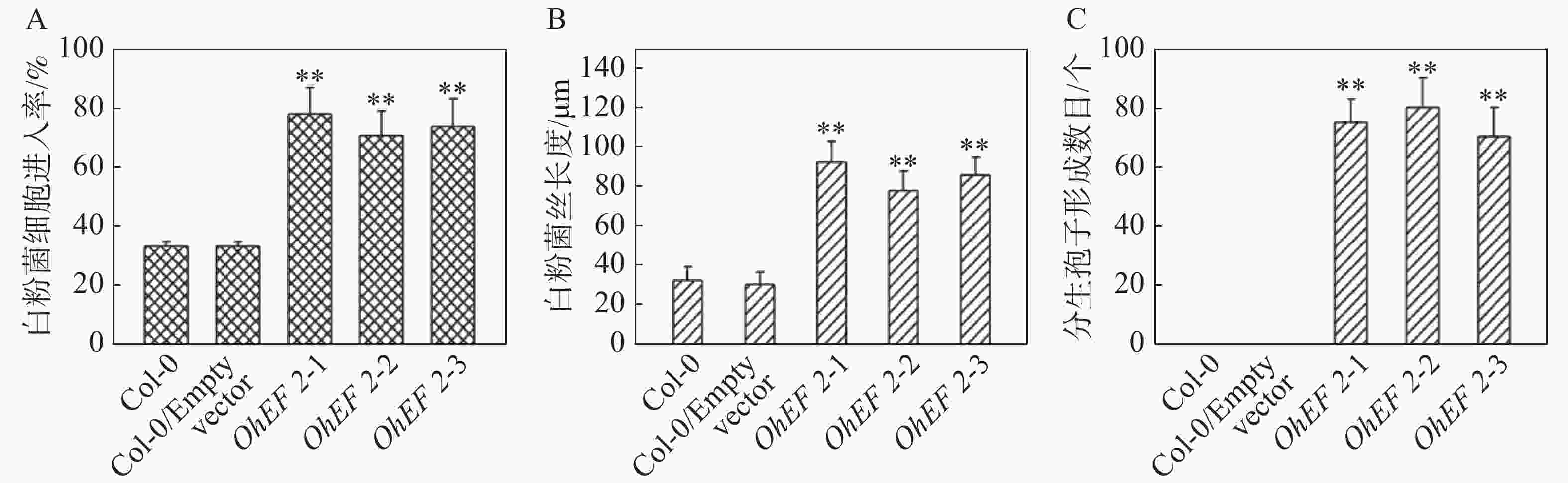
图 3 效应蛋白OhEF 2正调控拟南芥对橡胶树白粉菌的感病性
Figure 3. Effector protein OhEF 2 positively regulates the susceptibility of Arabidopsis to Oidium heveae
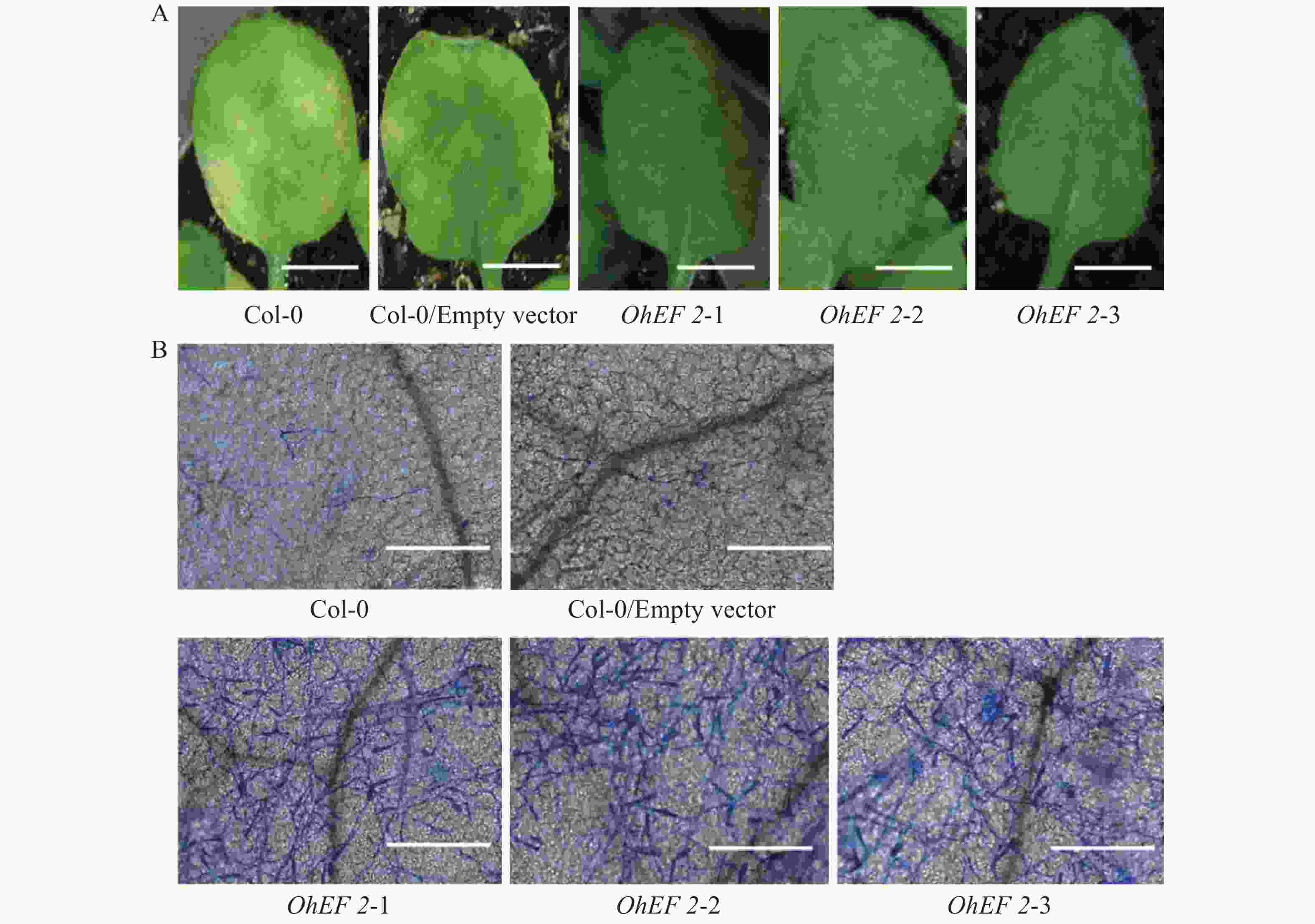
图 4 OhEF 2转基因植株叶片发病症状及显微观察结果
Figure 4. Symptoms and light microscopy of leaves of OhEF2 transgenic plants inoculated with Oidium heveae
-
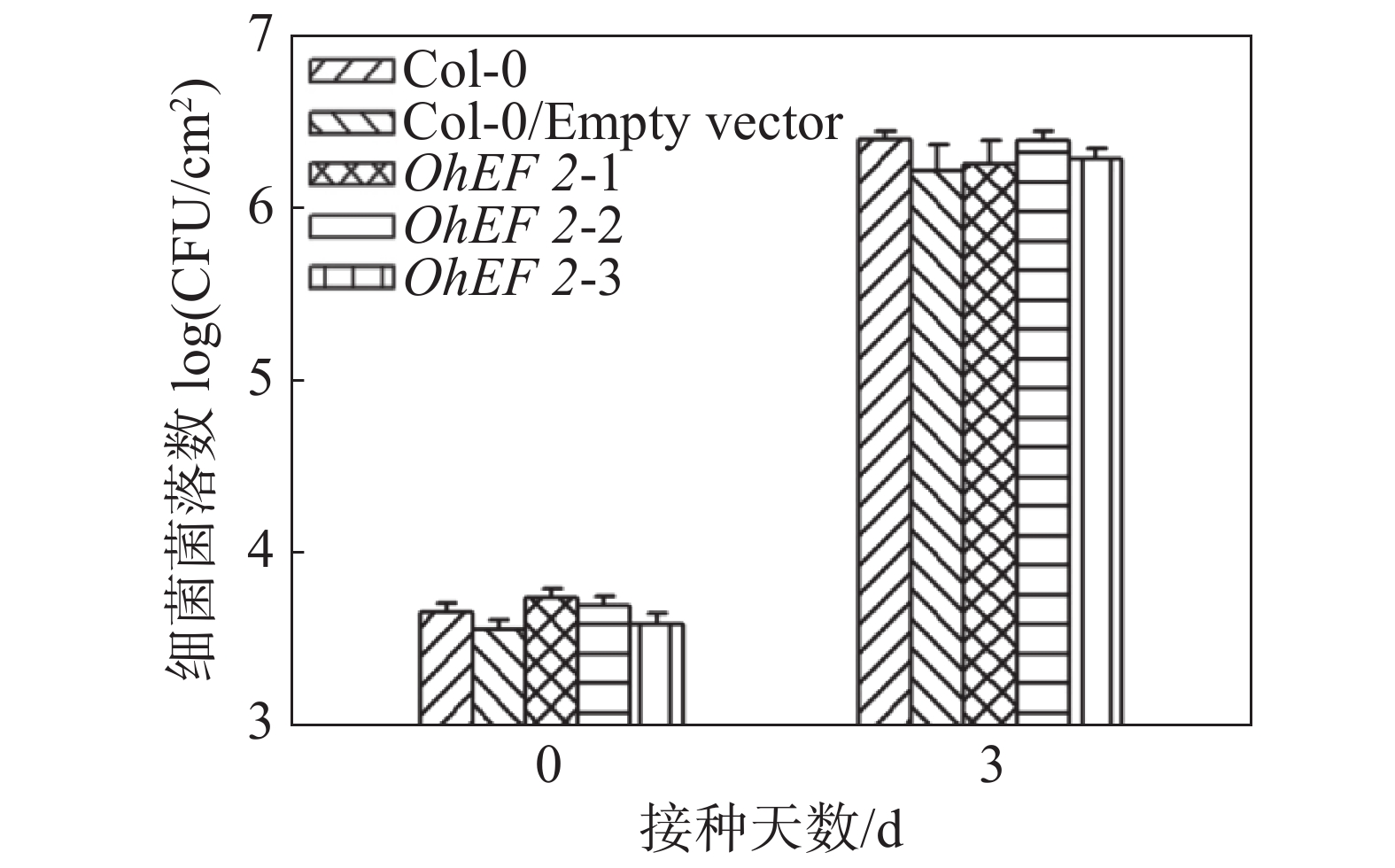
效应蛋白OhEF 2可以明显提高橡胶树白粉菌在拟南芥上的毒性,为了验证OhEF 2是否也可以促进其他病原菌的毒性,笔者在野生型拟南芥Col-0、空载体转基因植株和OhEF 2转基因植株OhEF 2-1,OhEF 2-2,OhEF 2-3上接种了假单胞菌DC 3000。假单胞菌作为活体寄生病原微生物,其与寄主植物的互作机制已有比较深入的了解。分别在接种后0 d和3 d取样、研磨、涂板,进行细菌计数。结果表明,假单胞菌DC 3000在野生型植物和空载体转基因植株以及OhEF 2转基因植株上并未表现出明显的生长差异(图5),结果表明,效应蛋白OhEF 2不能促进假单胞菌DC 3000的毒性。
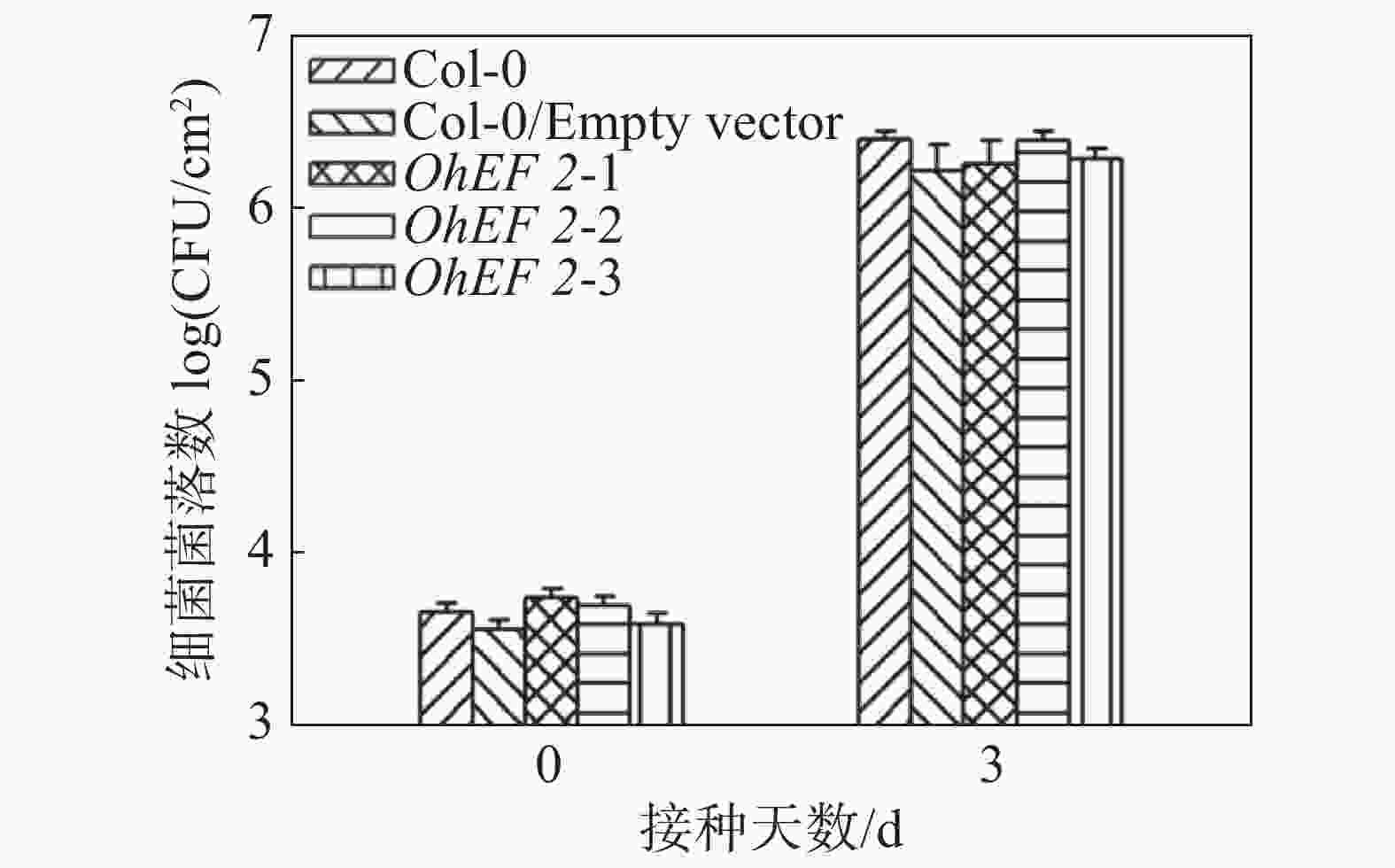
图 5 假单胞菌DC 3000细菌生长结果
Figure 5. Bacterial growth of Pseudomonas syringae DC 3000
-
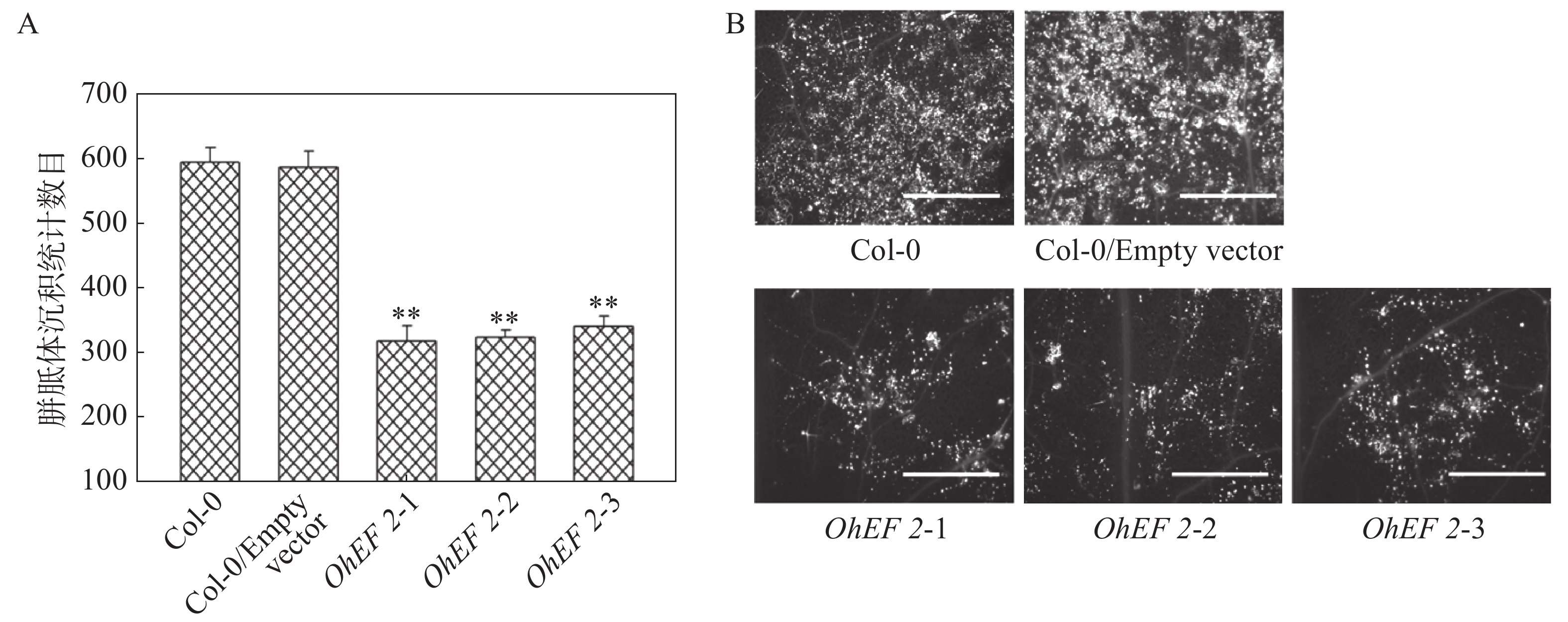
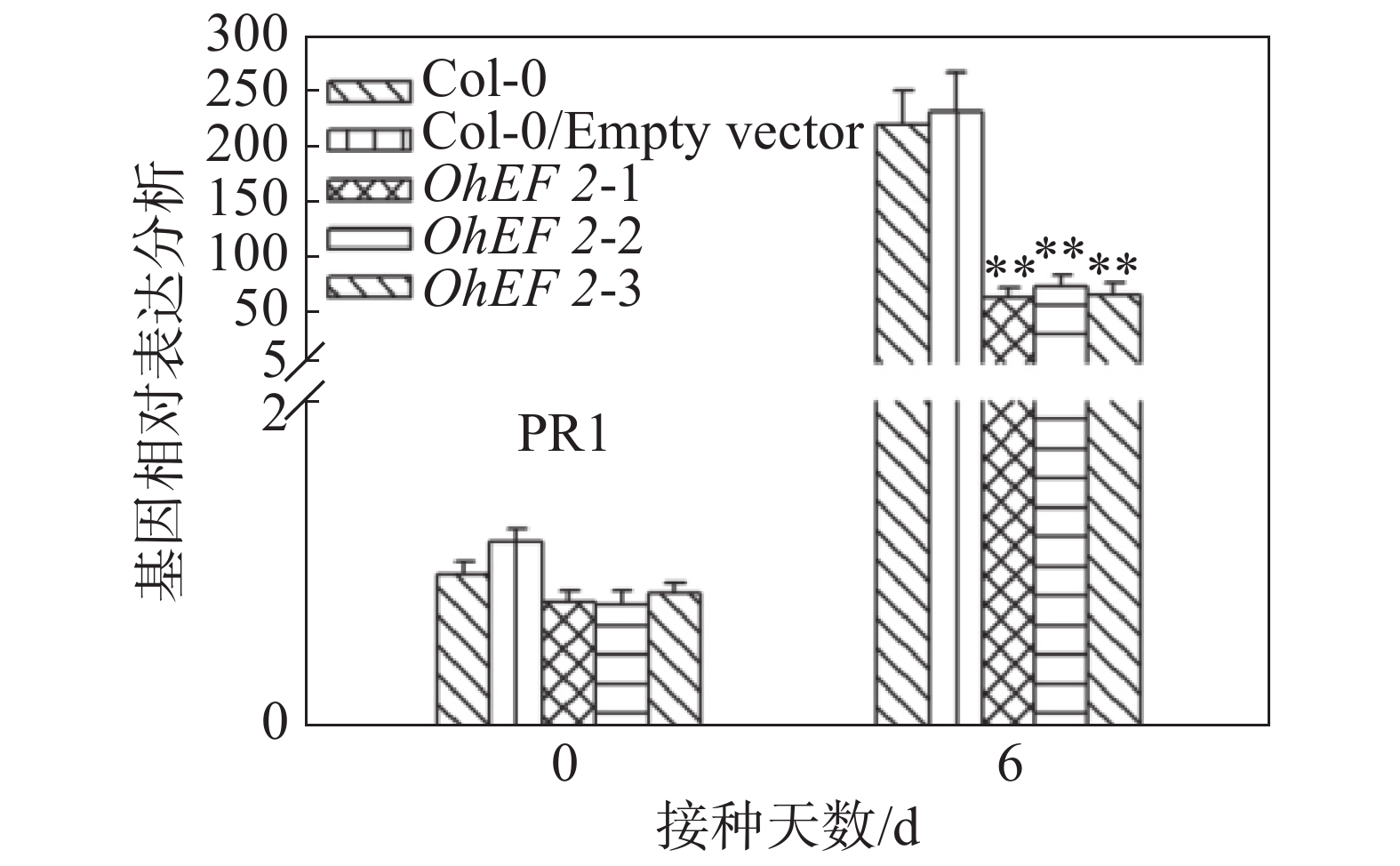
效应蛋白OhEF 2显著增强了拟南芥对橡胶树白粉菌的感病性,推测OhEF 2可能抑制了橡胶树白粉菌在拟南芥上激活的抗病反应。为了确定这一可能性,笔者在野生型Col-0、空载体转基因植株和OhEF 2转基因植株OhEF 2-1、OhEF 2-2、OhEF 2-3上分别接种橡胶树白粉菌O.Heveae HN1106。接种6 d后,剪取接种叶片,进行了胼胝体沉积分析和PR1基因表达分析。结果表明,与野生型相比,橡胶树白粉菌在3个转基因植株上诱导的胼胝体沉积(图6A,B)和PR1基因表达(图7)都显著降低,表明效应蛋白OhEF 2有效地抑制了橡胶树白粉菌在拟南芥上激发的抗病反应。
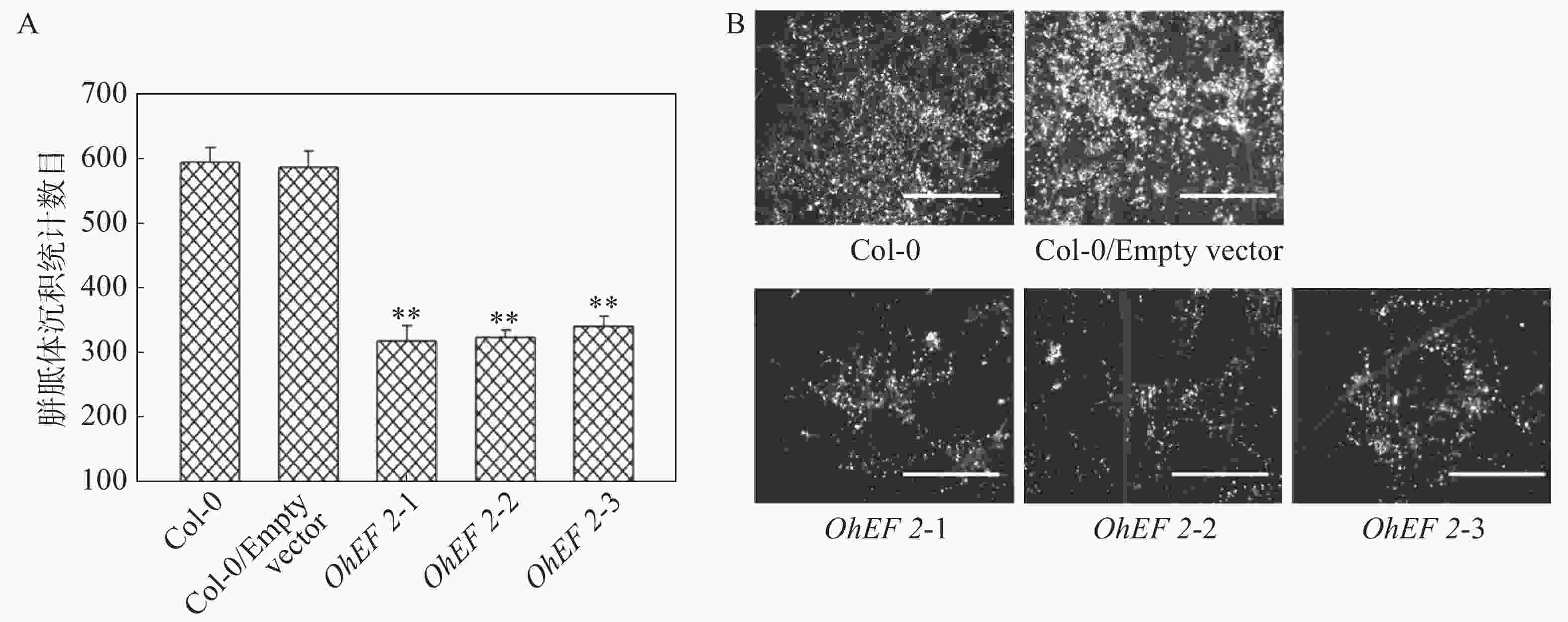
图 6 效应蛋白OhEF 2抑制橡胶树白粉菌在拟南芥上激发的胼胝体沉积
Figure 6. Effector protein OhEF 2 inhibits the callose deposition triggered by Oidium heveae in Arabidopsis
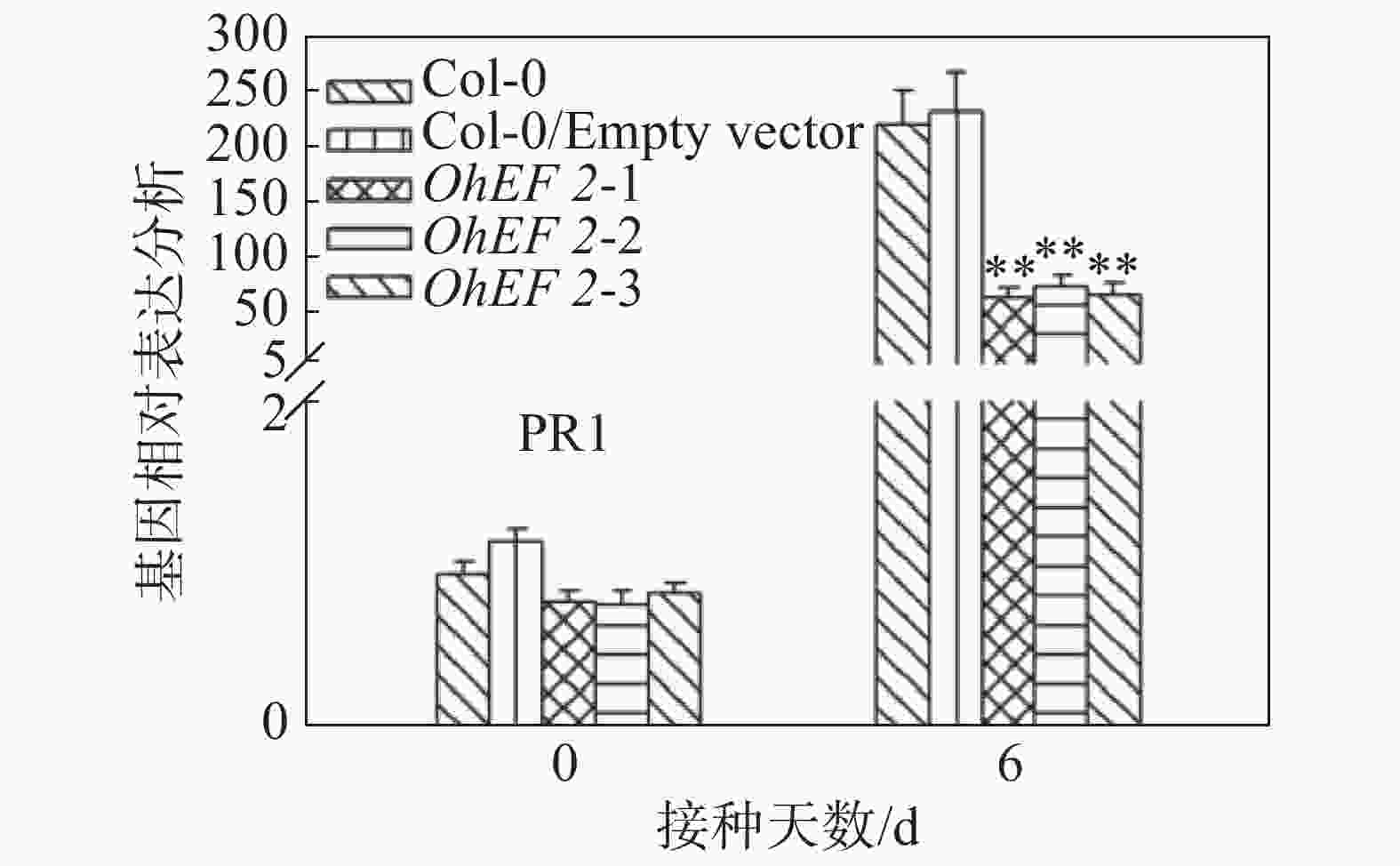
图 7 效应蛋白OhEF 2抑制橡胶树白粉菌在拟南芥上激发的PR1基因表达
Figure 7. Effector protein OhEF 2 inhibits the PR1 gene expression triggered by Oidium heveae in Arabidopsis
-
与拟南芥的相互作用过程中,橡胶树白粉菌对拟南芥野生型Col-0激发抗病反应,并且这种抗病反应依赖于EDS1和PAD4,但并不依赖于NDR1,推测拟南芥TIR-NB-LRR类抗性蛋白可以识别橡胶树白粉菌的效应蛋白,从而激活了下游的抗病信号通路[14]。笔者在前人研究的基础上,克隆了橡胶树白粉菌的1个潜在效应蛋白基因OhEF 2,并构建了该基因在野生型Col-0背景下雌激素诱导的过表达转基因植株。通过雌激素诱导后,与野生型Col-0和空载体转基因植株相比,OhEF 2过表达转基因植物并未出现明显的植物叶片发黄表型,暗示着效应蛋白OhEF 2在植物体内并不是发挥无毒蛋白的功能。
白粉菌作为专性活体寄生真菌,通过分泌大量的效应蛋白到寄主细胞内,发挥毒性功能,帮助白粉菌在寄主体内的侵染和繁殖[16-17]。大麦白粉菌效应蛋白CSEP0064可以与植物细胞内PR10蛋白发生相互作用,干扰宿主细胞内核糖体RNA的降解,抑制植物的免疫反应[18]。通过寄主诱导的基因沉默技术,发现白粉菌Podosphaera xanthii效应蛋白PEC可以抑制植物细胞内活性氧的产生[10],从而促进白粉菌的繁殖。笔者通过在野生型Col-0过表达效应蛋白OhEF 2,发现效应蛋白OhEF 2显著增强了橡胶树白粉菌的毒性功能,并有效降低了橡胶树白粉菌在拟南芥上激发的胼胝体沉积和PR1基因表达。胼胝体沉积在白粉菌的侵染过程中发挥非常重要的作用,在白粉菌侵染早期形成的刺入钉位置,植物细胞产生大量的胼胝体,从而抑制白粉菌的侵染。胼胝体合成酶基因PMR4突变后,植物对白粉菌的感病性大大增强[19],表明效应蛋白OhEF 2可能参与了白粉菌侵染的早期阶段,与胼胝体形成的相关基因发生相互作用,从而抑制了胼胝体沉积。另外,PR1基因作为效应蛋白激发免疫信号通路ETI(Effector triggered immunity)的标志基因[20],暗示着效应蛋白OhEF 2抑制了植物抗性蛋白识别橡胶树白粉菌后激发的免疫反应。然而,OhEF 2并不能促进活体寄生假单胞菌DC 3000在拟南芥上的毒性,表明橡胶树白粉菌与假单胞菌在植物体内的毒性作用机理可能是不相同的。OhEF 2能否促进其他白粉菌的毒性功能,尚需进一步验证。
Effector protein OhEF 2 enhances the susceptibility of Arabidopsis to Oidium heveae
-
摘要: 白粉菌通过在植物细胞内形成吸器,产生大量的效应蛋白,从而实现在寄主细胞内的侵染。前期有研究人员对Oidium heveae进行基因组和转录组测序分析,预测出133个潜在的效应蛋白。笔者克隆了其中的一个基因OhEP2 (Oidium heveae Effector Protein 2),并构建了OhEF 2基因在拟南芥Col-0背景下的过表达转基因植株。通过接种发现,过量表达OhEF 2可以明显促进拟南芥对橡胶树白粉菌的感病性,但不能提高假单胞菌DC 3000的毒性功能,表明OhEF 2可能只在白粉菌侵染的过程中发挥作用。进一步研究发现,OhEF 2显著降低了橡胶树白粉菌在拟南芥上激发的胼胝体沉积和PR1 (Pathogen-Related Gene1)基因表达,这为进一步研究效应蛋白OhEF 2在植物体内的毒性作用机理奠定了基础。Abstract: Powdery mildew fungi complete its infection process on living host plants through the establishment of haustoria which secretes a plethora of effectors into plant cells. A previous study showed that 133 potential effectors of Oidium heveae have been predicted by genome and transcriptome sequencing. However, the biological function of these effectors are still unknown. In this study, an effector gene, named OhEF 2, was cloned and transformed into Arabidopsis, and the transgenic plants of OhEF 2 gene in Arabidopsis Col-0 background were constructed and inoculated with O. heveae. Inoculation assay showed that OhEF 2 obviously enhanced the susceptibility of Arabidopsis to O. heveae. However, OhEF 2 did not increase the virulence of Pseudomonas syringae DC 3000, suggesting that OhEF 2 probably plays a role only in the process of powdery mildew infection. Further study showed that OhEF 2 decreased the callose deposition and PR1 gene expression triggered by O. heveae in Arabidopsis, which lays a foundation for the future study of OhEF 2 virulence mechanisms in host plants.
-
Key words:
- Oidium heveae /
- effector protein /
- OhEF 2 /
- virulence mechanism
-
图 2 橡胶树白粉菌效应蛋白OhEF 2拟南芥转基因植株的构建
A.效应蛋白OhEF 2转基因植株表型观察;B.效应蛋白OhEF 2转基因植株蛋白表达分析。Bar=1 cm。
Fig. 2 The construction of Oidium heveae effector OhEF 2 transgenic plants in Arabidopsis Col-0 background
A. The phenotype observation of effector OhEF-2 transgenic plants; B. Protein detection of effector OhEF-2 transgenic plants. Bar=1 cm.
图 3 效应蛋白OhEF 2正调控拟南芥对橡胶树白粉菌的感病性
A.橡胶树白粉菌细胞进入率测定结果;B.橡胶树白粉菌菌丝生长测定结果;C.分生孢子计数结果。**:P<0.01。
Fig. 3 Effector protein OhEF 2 positively regulates the susceptibility of Arabidopsis to Oidium heveae
A. Quantitative assessment of host cell entry rates; B. Quantitative analysis of hyphal growth of O. heveae; C. The numbers of conidiospores per colony were counted at 10 dpi. **: P<0.01.
图 4 OhEF 2转基因植株叶片发病症状及显微观察结果
A. 叶片症状观察结果,Bar=5 mm;B. 显微观察结果,Bar=200 μm。
Fig. 4 Symptoms and light microscopy of leaves of OhEF2 transgenic plants inoculated with Oidium heveae
A. Symptoms of WT Col-0/ Empty vector and OhEF 2 transgenic plants at 10 dpi, Bar=5 mm; B. Light microscopy images, Bar=200 μm.
图 6 效应蛋白OhEF 2抑制橡胶树白粉菌在拟南芥上激发的胼胝体沉积
A. 胼胝体计数结果;B. 叶片显微观察结果。**:P<0.01。
Fig. 6 Effector protein OhEF 2 inhibits the callose deposition triggered by Oidium heveae in Arabidopsis
A. Average number of callose deposits per microscope field of 0.1 mm2 on Col-0 and OhEF 2 transgenic plants; B. Light microscopy images. Bar=200 μm. **: P<0.01.
-
[1] WILSON I W, SCHIFF C L, HUGHES D E, et al. Quantitative trait loci analysis of powdery mildew disease resistance in the Arabidopsis thaliana accession kashmir-1 [J]. Genetics, 2001, 158(3): 1301 − 1309. [2] XIAO S, CALIS O, PATRICK E, et al. The atypical resistance gene, RPW8, recruits components of basal defence for powdery mildew resistance in Arabidopsis [J]. The Plant Journal: for cell and molecular biology, 2005, 42(1): 95 − 110. doi: 10.1111/j.1365-313X.2005.02356.x [3] GOLLNER K, SCHWEIZER P, BAI Y, et al. Natural genetic resources of Arabidopsis thaliana reveal a high prevalence and unexpected phenotypic plasticity of RPW8-mediated powdery mildew resistance [J]. The New Phytologist, 2008, 177: 725 − 742. doi: 10.1111/j.1469-8137.2007.02339.x [4] WANG Y, NISHIMURA M T, ZHAO T, et al. ATG2, an autophagy-related protein, negatively affects powdery mildew resistance and mildew-induced cell death in Arabidopsis [J]. The Plant Journal: for cell and molecular biology, 2011, 68: 74 − 87. doi: 10.1111/j.1365-313X.2011.04669.x [5] SHARMA G, AMINEDI R, SAXENA D, et al. Effector mining from the Erysiphe pisi haustorial transcriptome identifies novel candidates involved in pea powdery mildew pathogenesis [J]. Molecular Plant Pathology, 2019, 20(11): 1506 − 1522. doi: 10.1111/mpp.12862 [6] SPANU P D, ABBOTT J C, AMSELEM J, et al. Genome expansion and gene loss in powdery mildew fungi reveal tradeoffs in extreme parasitism [J]. Science, 2010, 330: 1543 − 1546. doi: 10.1126/science.1194573 [7] GODFREY D, BOHLENIUS H, PEDERSEN C, et al. Powdery mildew fungal effector candidates share N-terminal Y/F/WxC-motif [J]. BMC genomics, 2010, 11: 317. doi: 10.1186/1471-2164-11-317 [8] LIANG P, LIU S, XU F, et al. Powdery mildews are characterized by contracted carbohydrate metabolism and diverse effectors to adapt to obligate biotrophic lifestyle [J]. Front Microbiol., 2018, 9: 3160. doi: 10.3389/fmicb.2018.03160 [9] WU Y, MA X, PAN Z, et al. Comparative genome analyses reveal sequence features reflecting distinct modes of host-adaptation between dicot and monocot powdery mildew [J]. BMC Genomics., 2018, 19(1): 705. doi: 10.1186/s12864-018-5069-z [10] MARTÍNEZ-CRUZ J, ROMERO D, TORRE F N, et al. The functional characterization of Podosphaera xanthii candidate effector genes reveals novel target functions for fungal pathogenicity [J]. Mol. Plant-Microbe Interact., 2018, 32: 914 − 931. [11] BEELEY, F. Oidium heveae: report on the 1933 outbreak of Hevea leaf mildew [J]. J. Rubber Res. Inst. Malaysia, 1933, 5: 5 − 13. [12] MITRA M, MEHTA P R. Some leaf diseases of Hevea brasiliensis new to India [J]. Indian J. Agric. Sci., 1938, 8: 185 − 188. [13] SARANYA L, SAWANEE K, EDSON L, et al. Molecular phylogenetic and morphological analyses of Oidium heveae, a powdery mildew of rubber tree [J]. J. Mol. Evol., 2005, 46: 220 − 226. [14] MEI S, HOU G, CUI T, et al. Characterization of the interaction between Oidium heveae and Arabidopsis thaliana [J]. Mol Plant Pathol, 2016, 17(9): 1331 − 1343. doi: 10.1111/mpp.12363 [15] ZHANG J, SHAO F, LI Y, et al. A Pseudomonas syringae effector inactivates MAPKs to suppress PAMP-induced immunity in plants [J]. Cell Host Microbe, 2007, 1: 175 − 185. doi: 10.1016/j.chom.2007.03.006 [16] SATO K, KADOTA Y, SHIRASU, K. Plant immune responses to parasitic nematodes [J]. Front Plant Sci., 2019, 10: 1 − 14. doi: 10.3389/fpls.2019.00001 [17] THORDAL-CHRISTENSEN H, BIRCH P R J, SPANU P D, et al. Why did filamentous plant pathogens evolve the potential to secrete hundreds of effectors to enable disease? [J]. Mol Plant Pathol, 2018, 19: 781 − 785. doi: 10.1111/mpp.12649 [18] PENNINGTON H G, JONES R, KWON S, et al. The fungal ribonuclease-like effector protein CSEP0064/BEC1054 represses plant immunity and interferes with degradation of host ribosomal RNA [J]. PLoS Pathog, 2019, 15: e1007620. doi: 10.1371/journal.ppat.1007620 [19] NISHIMURA M T, STEIN M, HOU B H, et al. Loss of a callose synthase results in salicylic acid-dependent disease resistance [J]. Science, 2003, 301: 969 − 972. doi: 10.1126/science.1086716 [20] WU Y J, GAO Y, ZHAN Y Y, et al. Loss of the common immune coreceptor BAK1 leads to NLR-dependent cell death [J]. Proceedings of the National Academy of Sciences of the United States of America, 2020, 117(43): 27044 − 27053. doi: 10.1073/pnas.1915339117 -

 点击查看大图
点击查看大图
计量
- 文章访问数: 860
- HTML全文浏览量: 268
- PDF下载量: 36
- 被引次数: 0
