



-
土壤盐碱化是制约全球农业生产的主要非生物胁迫因子之一。其对作物的危害主要由于过量钠离子(Na+)和氯离子(Cl-)的积累,这些离子的过量积累会引发植物的渗透胁迫、离子毒害与养分失衡等,进一步破坏抗氧化系统、干扰蛋白质的合成,显著抑制植物的生长发育[1]。全球约20%的灌溉农田受到盐渍化影响,且其面积仍在持续扩大。由于大多数作物对盐分高度敏感,土壤盐渍化已造成严重的农作物减产、土地生产力下降、荒漠化加剧,极大地威胁到人类粮食和生态安全[2]。因此,提高植物的耐盐性已成为当前农业可持续发展的重要研究方向。
海雀稗(Paspalum vaginatum)为禾本科(Poaceae)雀稗属(Paspalum)多年生草本植物,具匍匐茎且根系较为发达,秆高20~60 cm,叶片线形,叶长5~15 cm、叶宽2~5 mm,叶鞘松弛包茎,叶舌短膜质。耐盐性强,耐水淹与贫瘠土壤,适生于滨海滩涂、盐渍草地兼具生态与经济价值,可用于滨海湿地植被恢复、盐碱地改良,亦是热带亚热带地区优质牧草与高尔夫球场果岭草坪草种,在盐渍环境生态修复中应用前景显著[3-5]。John等通过高粱(Sorghum bicolor)与海雀稗的对比研究表明,在盐胁迫下,海雀稗能够将Na+储存在叶乳突内,从而有效减少Na+向光合组织和代谢活跃部位的运输,缓解胁迫,维护细胞的正常生理功能[6]。该机制显著增强了海雀稗的耐盐性,为其在盐渍环境中的生存和生长提供了重要保障。叶片的形态、大小和表皮结构直接影响海雀稗的生长,叶片表皮的多种特化细胞和结构能够抵抗生物和非生物胁迫。同时,表皮结构对光反射及水分和气体交换的调控也会影响植物的光合性能[7]。
乳突(papillae)是由表皮细胞壁或角质层外凸形成的特化结构,广泛存在于多种植物组织中[8],其在植物生理生态过程中具有重要功能,尤其在苔藓植物叶片中表现显著[9]。这些由表皮细胞特化形成的微小物理结构,通常分布于气孔周围或叶片沟槽区域,能够有效增加叶片表面疏水性,促进气膜形成,从而减少盐分通过蒸腾作用在叶片表面的沉积与侵入[10]。此外,有研究表明,该结构可能作为物理屏障延缓水分蒸发、调节离子(如Na+、Cl-)的吸收与运输,进而缓解离子毒害与渗透胁迫[11-12]。高等植物叶片在长期进化过程中分化出多种特化的细胞类型与表皮结构,例如乳突、气孔以及毛状体等。这些结构不仅可有效反射过量辐射,还可以抵御病原体侵入[13]。部分植物的叶乳突可增加叶片粗糙度,使昆虫难以立足、爬行,干扰其取食与产卵,像一些表面布满密集叶乳突的植物,可减少害虫啃食[14]。在抵御病原菌方面,叶乳突可作为物理屏障,阻碍真菌孢子附着、萌发及菌丝侵入,降低植物染病几率[15]。在调节植物与环境的物质交换上也有明显效果,以荷叶为例,其叶乳突呈独特的结构,和表皮蜡质协同,使叶面形成超疏水表面。雨水落在荷叶上,因叶乳突存在,只能与叶表面形成点接触,在自身表面张力作用下聚成水珠滚落,带走灰尘等污染物,实现自清洁,这就是著名的“荷叶效应”[16-17]。当前植物叶表乳突研究多集中于形态特征及紫外损伤防护、病原菌侵染抵御等生态功能,但海雀稗叶乳突与其耐盐性的关联性尚未明确[18]。因此,本研究以40份海雀稗种质资源为研究材料,通过指甲油拓印法将叶乳突拓印至载玻片上进行观察,在先前研究的基础上进一步了解海雀稗叶乳突盐处理前后的变化特征,旨在揭示植物叶乳突特征是否与耐盐性相关,为后续研究海雀稗耐盐机制奠定了基础。
-
供试的40份海雀稗材料分别来自于美国、阿根廷、南非、中国广东和海南等地(表1),现存在海南大学热带草坪草育种实验地资源圃。实验于2025年5—6月在海南大学海甸校区农学基地10号大棚内进行。先将实验群体进行培养,选择生长一致、无病害的匍匐茎作为试验材料,种植在育苗杯内,每个育苗杯内放置5根匍匐茎且每个匍匐茎上的节点数相同,每个种质5个重复。实验采用水培培养,NaCl处理浓度为200 mmol·L−1,将材料放在水培盆中,容量为5 L,每盆内放置4个育苗杯,等到实验开始时将其剪至同一高度进行试验。实验进行期间每4 d加1次清水,7天添加1次霍格兰营养液A、B、C液各15 mL,在实验的第28天时,取完全展开且完整的倒四叶,每个重复取1片叶子,将叶子平均的分为前、中、后3部分,取叶子的中部进行保存。将采集的叶片储存在装有70%(V/V) FAA固定液的小棕瓶内,避光保存,后续使用。
表 1 40份海雀稗种质信息来源
Table 1. Source information on 40 accessions of the germplasm of Paspalum vaginatum
种质来源
Germplasm sources材料名称
Name of germplasm阿根廷
Argentina17USA-34,PI509018-2,17USA-06 南非
South Africa17USA-11 马尔代夫
Maldives马尔代夫 中国广东
Guangdong, China18HN-90 中国海南
Hainan, China18HN-101,17HN-02,琼2,17HN-45,17HN-54,17HN-25 美国
America17USA-03,KaiLuna,17USA-17,TFP7-4,17USA-45,17USA-30,HYB7,17-35,17USA-07,HYB5,17USA-31,18HN-100,17USA-41,18HN-98,Taliaferra,17USA-42,TCR3,TGkona,VeroBeach,Polo,Saiam,17USA-23,TOCGC, 17USA-18,Kim1 ,17USA-15,Sealsle2000,Wailual kauai -
乳突是植物表皮细胞壁或角质层向外凸起形成的产物,在植物受到盐胁迫时可以将植物体内的Na+封存在叶乳突内。叶乳突临时装片的制作采用指甲油拓印法[18](图1),将叶片从FAA固定液里取出来后风干,取叶片的中部,之后涂薄薄的一层透明甲油在叶片的上表皮,再使用手持小风扇将其吹干并用高透的透明胶带覆盖在上表皮上,随后轻轻的按压使胶带和甲油胶更加的贴合慢慢的把胶带撕下粘贴在载玻片上写好相应的编号。将制作好的临时装片放在光学显微镜(Olym-pus BX41,Olympus,Tokyo,Japan)
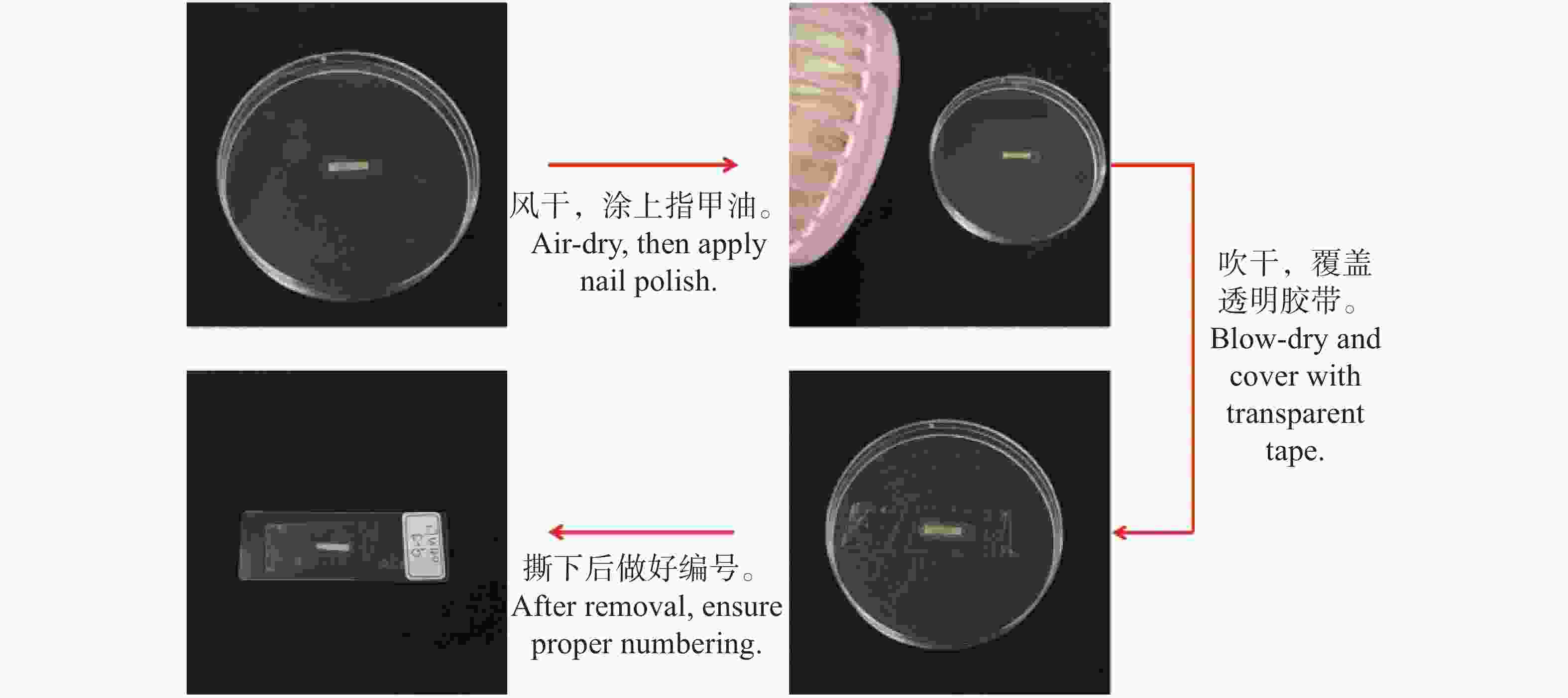
图 1 指甲油拓印法
Figure 1. Nail polish stamping technique
下进行图像的采集。使用Image J软件对叶乳突的形态特征进行测定(图2),包括单个叶乳突的长(μm,Lpl)、单个叶乳突的宽(μm,Lpw)、叶乳突簇宽(μm,Lpv)、单个叶乳突的长宽比(μm,Lpr)、叶乳突总簇宽(μm,Lpg)、叶乳突簇间隙距离(μm,Lg)、叶乳突面积(μm2,Ap)(每次测量6个乳突为1个整体)。

图 2 海雀稗叶乳突测量方案
Figure 2. Measurement scheme of Paspalum vaginatum leaf papillae
-
先使用Microsoft Excel 2022软件对原始数据进行计算与整理,将数据标准化。在R(1.4.1)软件中使用corrplot包进行相关性分析和聚类分析,通过聚类分析划分出功能类群相似的种质类群,以明确不同种质间的功能关联性,图表中所呈现的数据均以 “平均值±标准误差” 形式表示。在IMB SPSS Statistics 26中进行盐处理前后描述性统计分析和主成分分析。计算公式如下:
$$ U({\mathrm{X}}_{f})=\frac{{X}_{f}-{X}_{fmin}}{{X}_{fmax}-{X}_{fmin}} \text{,} $$ (1) $$ {W}_{f}=\frac{{P}_{f}}{\displaystyle\sum \limits_{f=1}^{n}{\mathrm{P}}_{f}} \text{,} $$ (2) $$ D=\sum \limits_{f=1}^{n}\left[U({X}_{f})\times {W}_{f}\right] \text{,} $$ (3) 式中:Xf 代表第f个指标,Xfmin、Xfmax分别为第f个指标的最小值与最大值;Wf为权重,用于体现第f个指标在评价体系中的重要程度;Pf是第f个综合指标对应的贡献率;D值则表示叶乳突排布密集程度的综合评价值。
-
由表2可知,海雀稗群体在盐处理下,叶乳突长宽比、簇宽、簇间隙距离和总簇宽胁迫前较胁迫后种质差异不大,没有显著差异。叶乳突单个长、单个宽、面积胁迫前较胁迫后种质明显变小,有显著差异(P<0.05)。胁迫前叶乳突簇间隙距离和面积变异系数最大,分别为28%、27%,两者最大值为最小值的倍数分别是4.79和3.47,该指标群体数值分布较分散,最大值、最小值相差较多。胁迫前后叶乳突单个长的变异系数增加最多,增加了3%,叶乳突长宽比的变异系数不变,叶乳突簇宽的变异系数减少1%。
表 2 盐胁迫前后叶乳突特征描述统计
Table 2. Statistical description of leaf papilla characteristics before and after salt stress
指标
Parameter胁迫前
Non-stress处理
Salt stress最大值
Max最小值
Min均值±标准差
Meam±SD变异系数/%
CV/%最大值
Max最小值
Min均值±标准差
Meam±SD变异系数/%
CV/%Lpl/μm 95 28.74 57.91±11.03a 19 83.72 20.93 52.13±10.78b 21 Lpw/μm 31.35 9.36 20.36±4.02a 19 30.67 6.37 18.41±4.08b 22 Lpr/μm 5.58 1.82 2.89±0.50a 17 5.70 1.79 2.89±0.51a 17 Lpv/μm 122.01 31.92 77.86±13.81a 17 106.57 39.06 77.97±12.62a 16 Ap/(ind.μm−2) 107502.41 22249.62 58414.07 ±15913.37 a27 84782.22 10099.44 48332.26 ±13994.94 b29 Lg/μm 47.93 6.56 21.50±6.01a 28 44.08 8.58 21.07±5.83a 28 Lpg/μm 363.03 162.9 275.77±30.94a 11 338.53 182.96 276.08±31.33a 11 注: Lpl.叶乳突单个长,Lpw.单个叶乳突宽,Lpr.叶乳突长宽比,Lpv.叶乳突簇宽,Ap.6个叶乳突面积,Lg.叶乳突簇间隙距离,Lpg.叶乳突总簇宽。均值±标准差后不同小写字母表示同一处理时间,不同指标在0.05水平下差异显著;下同。 Note: Lpl. Single length of papillae; Lpw.Width of single papillae; Lpr. Length and width ratio of papillae; Lpv. Width of papillae clusters; Ap.6leaf mastoid areas; Lg. Wide gap of papillae; Lpg. Width of total clusters of papillae. Different lowercase letters following the mean ± standard deviation denote significant differences at the 0.05 level for the same treatment time across different indicators, similarly hereinafter. -
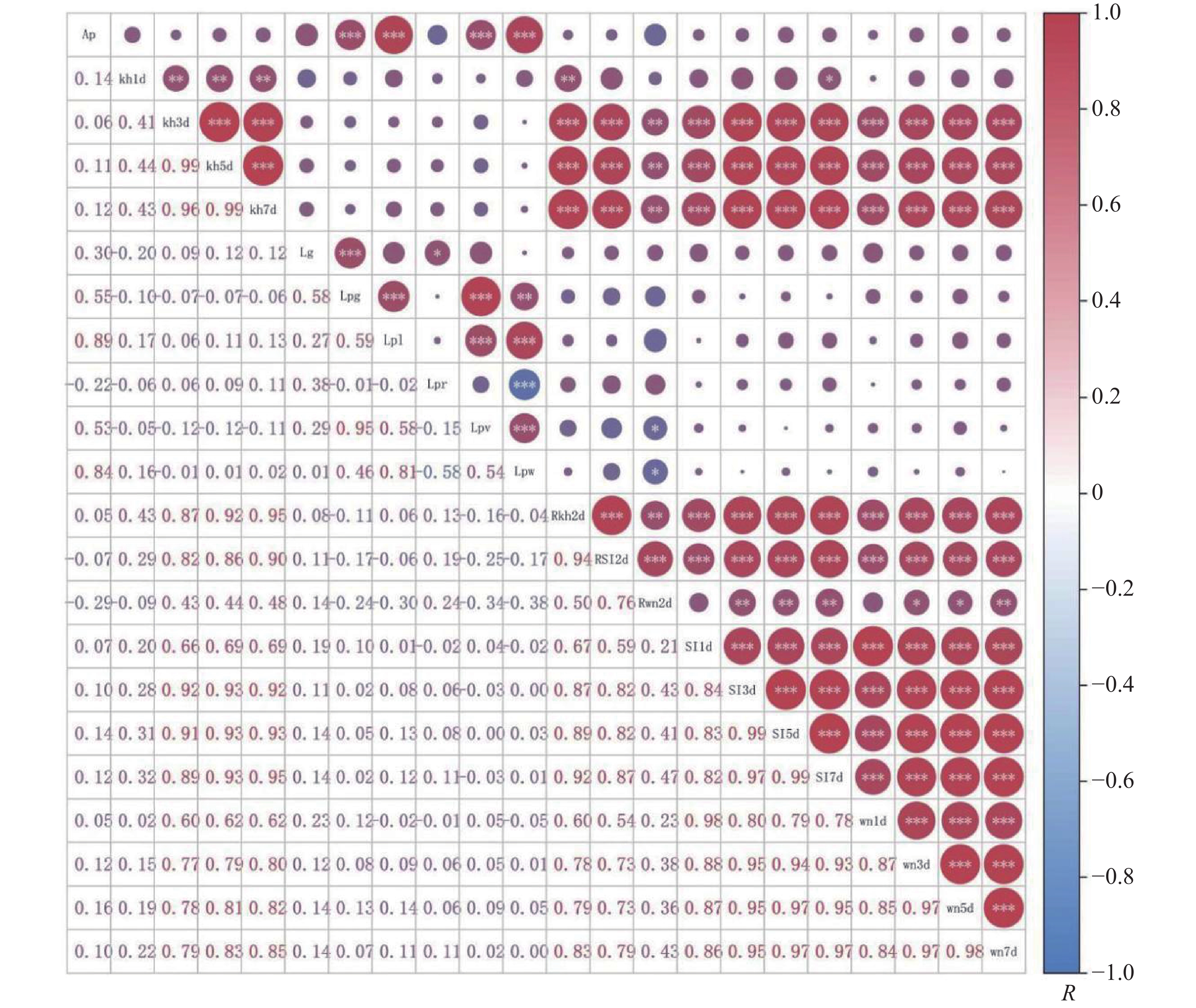
利用Pearson模型对盐处理前后叶乳突特征相关性进行分析,胁迫前结果表明(图1),叶乳突面积与叶乳突总簇宽、簇宽呈显著正相关(P<0.01),相关系数分别为0.46、0.45,与叶乳突单个长、单个宽呈极显著正相关(P<0.001),相关系数分别为0.94、0.84。叶乳突簇间隙距离与叶乳突总簇宽呈极显著正相关(P<0.001)。叶乳突总簇宽与叶乳突单个长、叶乳突单个宽呈显著正相关(P<0.01),相关系数分别为0.44、0.45,与叶乳突簇宽呈极显著正相关(P<0.001)。叶乳突单个长与叶乳突簇宽呈显著正相关(P<0.01),与叶乳突单个宽呈极显著正相关(P<0.001)。叶乳突长宽比与叶乳突单个宽呈显著负相关(P<0.01)。叶乳突簇宽与叶乳突单个宽呈极显著正相关(P<0.001)。叶乳突簇间隙距离与总簇宽呈极显著相关,与其他指标无相关性,叶乳突长宽比与簇宽、单个宽有相关性,与其他指标无相关性。
胁迫前后结果表明(图2),叶乳突面积与叶乳突簇宽、总簇宽、单个长、单个宽呈极显著正相关(P<0.005),相关系数分别为0.53、0.55、0.89、0.84。叶乳突簇间隙距离与叶乳突叶乳突总簇宽呈极显著正相关(P<0.005),与叶乳突长宽比呈显著正相关(P<0.05)。叶乳突总簇宽与叶乳突单个长、簇宽呈极显著正相关(P<0.005),相关系数分别为0.59、0.95,与叶乳突单个宽呈显著正相关(P<0.01)。叶乳突单个长与叶乳突簇宽、单个宽呈极显著正相关(P<0.005),相关系数分别为0.58、0.81。叶乳突长宽比与叶乳突单个宽呈极显著负相关(P<0.005)。叶乳突簇宽与叶乳突单个宽呈极显著正相关(P<0.005),与胁迫复水后第2天(Rwn2d)呈显著负相关(P<0.05),相关系数为−0.34。叶乳突单个宽与盐处理复水第2天萎蔫度(Rwn2d)呈显著负相关(P<0.05),相关系数为−0.38。叶乳突各指标除叶乳突单个宽和簇宽与Rwn2d呈显著负相关外,其他指标与盐处理指标无相关性。胁迫前叶乳突长宽比与叶乳突簇间隙距离无明显相关性,胁迫后呈显著相关(P<0.05)。盐处理指标中,Rwn2d与胁迫第1天综合指标(SI1d)和胁迫第1天萎蔫度(wn1d)无相关性,与其他指标均有相关性。胁迫第1天枯黄率(kn1d)与胁迫第3、5、7天枯黄率 、胁迫复水后第二天枯黄率(Rkn2d)呈显著正相关(P<0.01),与胁迫第7天综合指标呈显著相关(P<0.05),与其他指标均无相关性,其他盐胁迫指标之间均有相关性。

图 4 胁迫后不同种质叶乳突特征相关性
Figure 4. Correlation of leaf papilla characteristics among different accessions of the germplasm following stress exposure
-
对叶乳突各指标进行主成分分析,主成分1和主成分2综合指标的贡献率分别为55.54%、21.21%,累计贡献率达到79.75%,表明这2个主成分贡献率符合主成分分析要求可以代表叶乳突测量指标的大部分信息(表3)。在主成分1中,载荷值较大的因子是叶乳突单个长和叶乳突面积,其特征向量都为0.89,最小的是叶乳突的长宽比为-0.24,这些性状主要与叶乳突的大小相关。在主成分2中,载荷值较大的因子是叶乳突长宽比和叶乳突簇间隙距离,其特征向量分别为0.83、0.76,最小的是叶乳突单个宽为-0.49,这些性状主要与叶乳突排布有关。叶乳突单个长、面积和长宽比是2个主成分的主导因子,对海雀稗叶乳突特征划分起主导作用。
表 3 海雀稗种质盐胁迫后叶乳突主成分分析
Table 3. Principal component analysis of leaf papillae in Paspalum vaginatum following salt stress
性状
Traits主成分1
Comp.1主成分2
Comp.2Lpl/μm 0.89 −0.02 Lpw/μm 0.84 −0.49 Lpr/μm −0.24 0.83 Lpv/μm 0.83 0.15 Ap/μm 0.89 −0.13 Lg/μm 0.40 0.76 Lpg/(ind·μm−2) 0.84 0.38 特征值
Characteristic value3.89 1.69 贡献率/%
Contribution rate/%55.54 24.21 累计贡献率/%
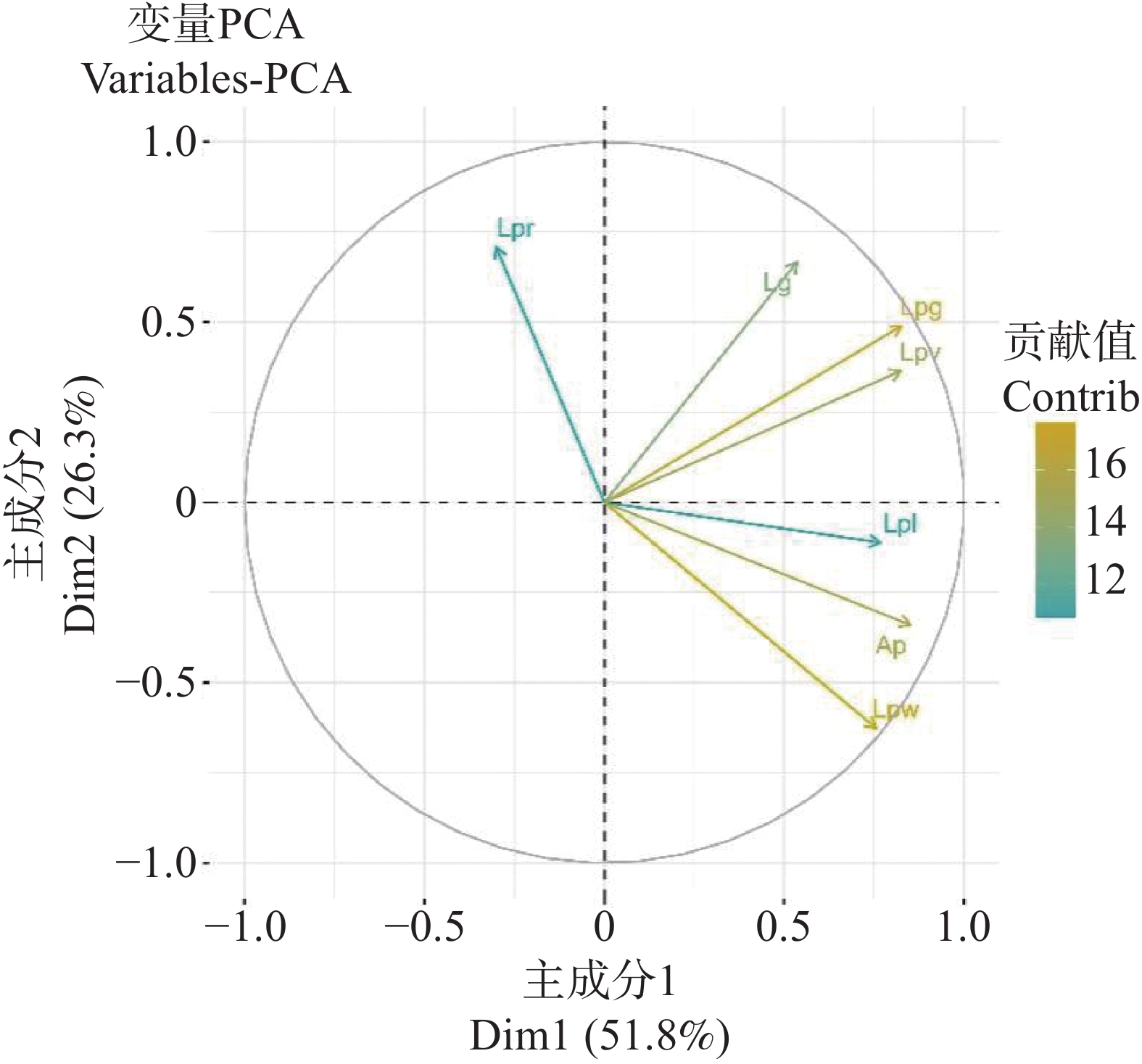
Cumulative contribution rate/%55.54 79.75 为清晰呈现2个主成分之间各指标的特征,依据各指标在主成分1(PC1)与主成分2(PC2)的载荷系数,构建GGE双标图(图3)并纳入所有参试种质与指标。在该双标图中,指标间相关性可通过向量夹角直观判断:当2个指标的向量夹角小于90°时,二者呈正相关关系;若向量夹角大于90°,则表明2个指标存在负相关关系。
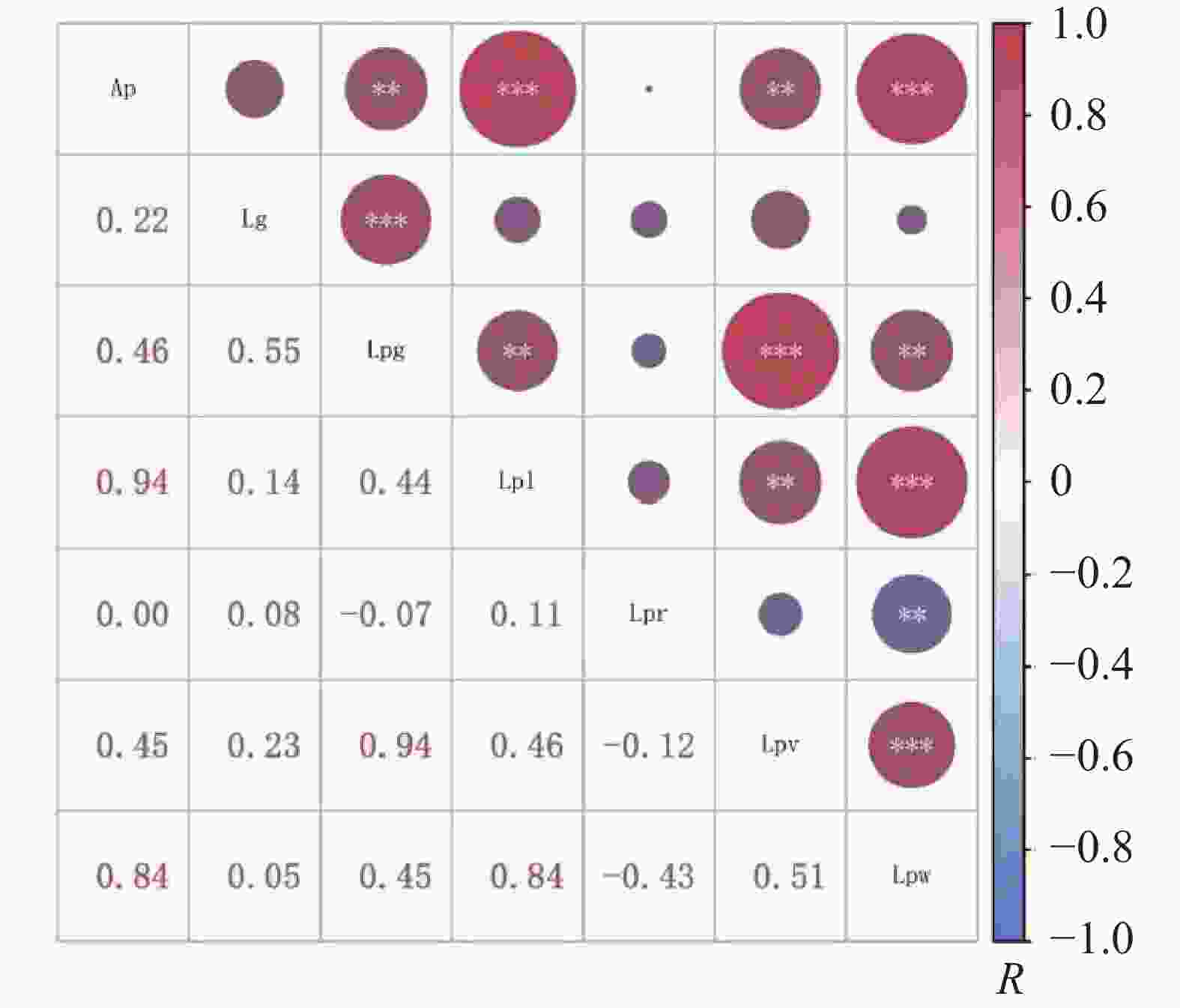
图 3 胁迫前不同种质叶乳突特征相关性
Figure 3. Correlation of different genetic backgrounds in the characteristics of leaf papillae before stress treatment

图 5 海雀稗叶乳突各指标的GGE双标图
Figure 5. GGE dual-scale diagram of various indicators for the papillae of the leaves of Paspalum vaginatum
-
为了进一步了解各种质叶乳突性状的优异,根据1.3中公式(1)、(2)、(3)计算出各种质的D值进行排名,筛选出叶乳突覆盖程度较高的种质(表4)。结果如下,排名前5的分别是17USA-03、17USA-07、18HN-90、Kai Luna、17USA-35,排名后5的分别是17HN-25、Sealsle2000、TCR3、17USA-15、Wailua Kauai。前5的种质叶乳突排布更密集,覆盖率更高,后5个种质的叶乳突排布较稀疏,覆盖率较低。D值的算式为D=0.613
$ \times $ Lpl+0.436$ \times $ Lpw+0.084$ \times $ Lpr+0.623$ \times $ Lpv+0.580$ \times $ Ap+0.509$ \times $ Lg+0.7$ \times $ Lpg。表 4 40份海雀稗种质叶乳突性状排名
Table 4. Ranking of leaf papillae traits in 40 accessions of Paspalum vaginatum germplasm
种质
SamplesD值
D values排名
Ranking种质
SamplesD值
D values排名
Ranking17USA-03 3.60 1 Kim 1 0.11 21 17USA-07 2.14 2 Salam 0.10 22 18HN-90 1.75 3 TOCGC 0.07 23 Kai Luna 1.43 4 17USA-34 0.07 24 17USA-35 1.35 5 TFP 7-4 −0.12 25 17USA-30 1.27 6 琼2 −0.15 26 17HN-54 1.20 7 17USA-17 −0.33 27 马尔代夫 1.10 8 17USA-45 −0.57 28 17USA-06 1.01 9 Taliaferra −0.68 29 17USA-31 0.88 10 17USA-18 −0.74 30 17USA-42 0.82 11 18HN-101 −0.79 31 17HN-02 0.77 12 Polo −0.80 32 18HN-98 0.70 13 17USA-23 −1.00 33 HYB 5 0.58 14 TG Kona −1.17 34 Vero Beach 0.58 15 PI 509018-2 −1.53 35 18HN-100 0.53 16 17HN-25 −1.72 36 17USA-11 0.53 17 SeaIsle2000 −1.85 37 17HN-45 0.32 18 TCR 3 −2.23 38 17USA-41 0.18 19 17USA-15 −3.26 39 HYB 7 0.15 20 Wailua Kauai −4.31 40 -
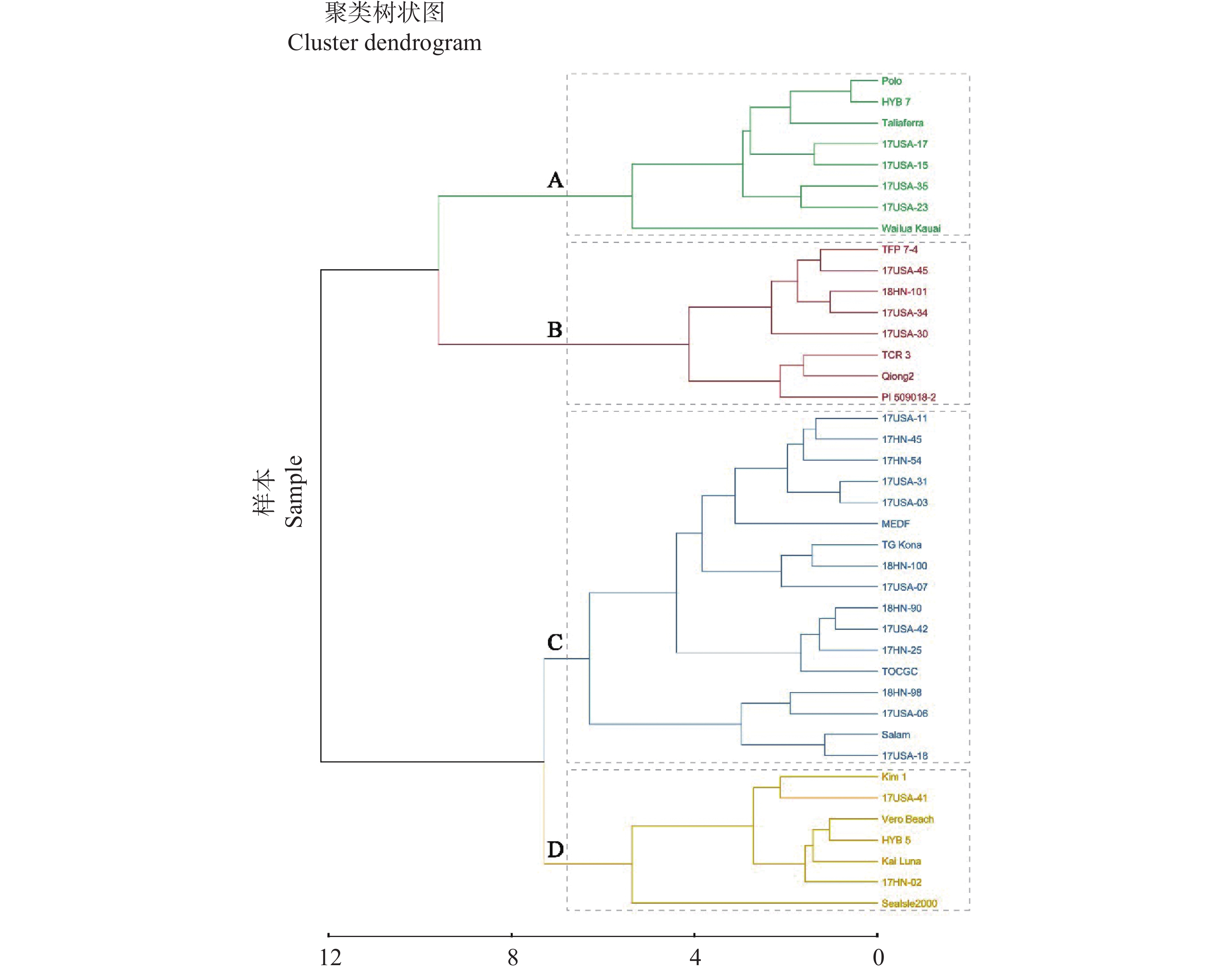
以40份供试材料为基础,按照叶乳突的各个指标进行聚类划分(图2),将其划分为4大类(A、B、C、D)。其中,A类包括Wailuakauai 、Taliaferra、17USA-17、17USA-15、17USA-23、17USA-35、HYB7、Polo等8个种质;B类包括TFP7-4、17USA-45、18HN-101、17USA-34、17USA-30、TCR3、琼2(qiong2)、PI509018-2等8个种质;C类包括17HN-45、17HN-54、17HN-25、17USA-11、17USA-31、17USA-03、17USA-07、17USA-06、17USA-42、17USA-18、18HN-100、18HN-90、18HN-98、马尔代夫(MEDF)、TGKona、TOCGC、Salam等17个种质;D类包括Kim1、17USA-41、VeroBeach、HYB5、17HN-02、Sealsle2000、KaiLuna等7个种质。
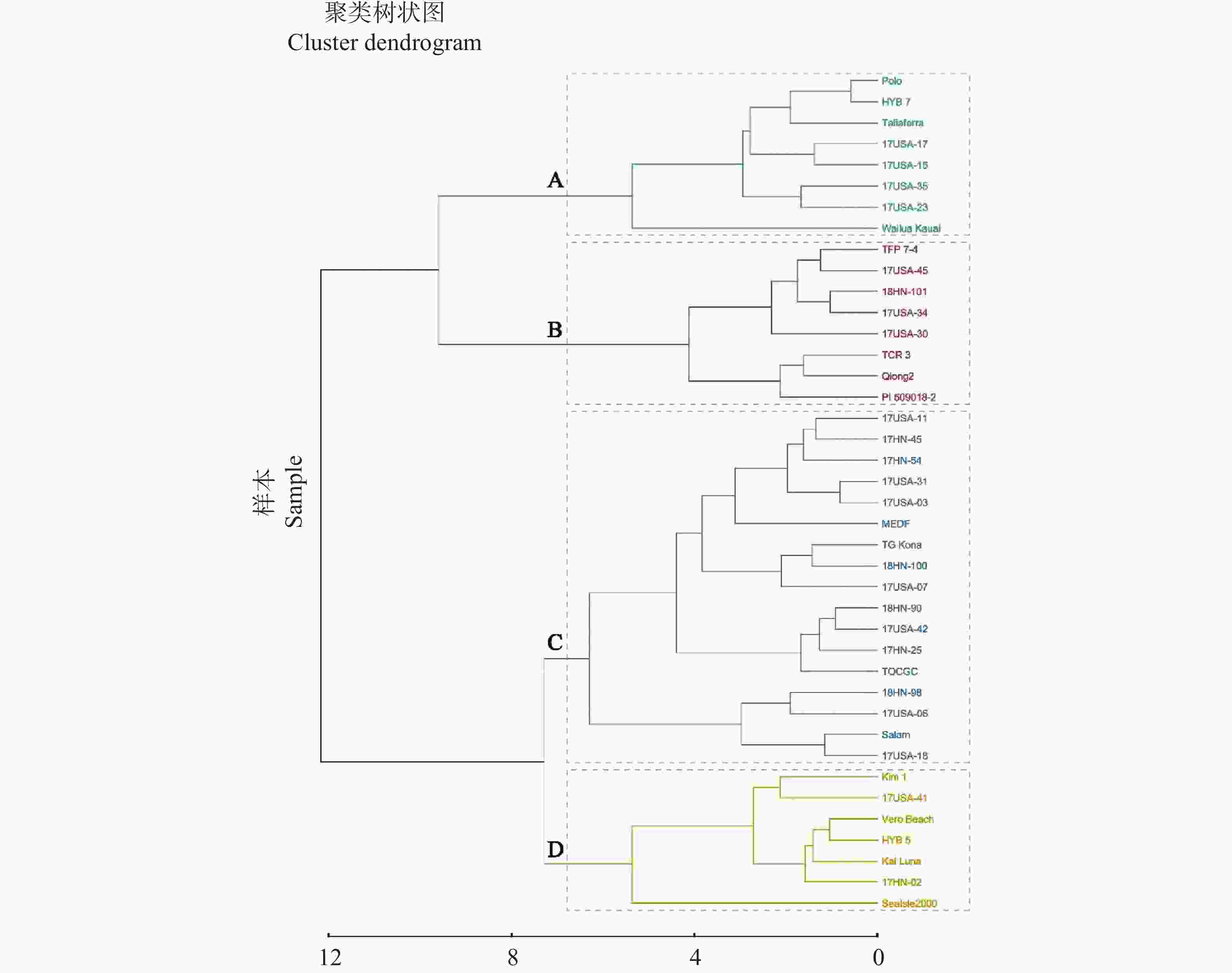
图 6 40份海雀稗胁迫后主成分综合得分聚类分析
Figure 6. Cluster analysis of the composite principal component scores for 40 accessions of Paspalum vaginatum following stress exposure
将聚类结果分为4类群进行差异分析,从表5可知,A类群叶乳突单个宽、簇间隙距离大于B、C、D类群,其均值分别为21.14 、22.43 μm;B类群叶乳突单个长、长宽比、簇宽、总簇宽、面积明显大于A、C、D类群,其均值分别为61.73 μm、3.14 μm、81.81 μm、289.78 μm、61 634.79 μm2;D类群叶乳突单个长、单个宽、簇宽、总簇宽、面积、簇间隙距离明显小于A、B、C类群,其均值分别为55.27 μm、18.39 μm、68.25 μm、241.89 μm、51 038.51 μm2、18.56 μm。胁迫前D类群叶乳突平均值较小,B类群较大。
表 5 40份群海雀稗各性状不同类群差异分析
Table 5. Differential analysis of traits among different groups of 40 accessions of Paspalum vaginatum
A类群
Group AB类群
Group BC类群
Group CD类群
Group DLpl/μm 57.34±13.96a 61.73±6.03a 57.48±6.56b 55.27±7.48b Lpw/μm 21.12±4.88a 20.25±3.73a 20.86±2.34b 18.39±2.14b Lpr/μm 2.72±0.12a 3.14±0.42a 2.79±0.23a 3.04±0.23a Lpv/μm 78.33±8.15a 81.81±8.11a 79.09±6.90b 68.25±9.44b Ap/μm 57237.71 ±20909.92 a61634.79 ±10928.60 a60489.02 ±11242.87 b51038.51 ±12962.65 bLg/μm 22.43±3.63a 22.18±2.01a 21.96±7.23a 18.56±5.20a Lpg/(ind.μm-2) 279.86±19.61a 289.78±24.72a 281.19±26.97b 241.89±37.12b 注:不同小写字母表示同一处理时间,不同指标在0.05水平下差异显著;下同。 Note: Different lowercase letters denote significant differences between parameters at the 0.05 level at the same treatment time, similarly hereinafter. 从表6中可知,胁迫后叶乳突长宽比在4类群间差异均不显著。A类种质叶乳突长宽比大于B、C、D类群,但叶乳突面积、单个长、单个宽明显低于B、C、D类群;B类群叶乳突单个宽、面积、簇间隙距离均大于A、C、D类群,但叶乳突长宽比小于A、C、D类群;C类群叶乳突簇宽和总簇宽大于A、B、D类群;D类群叶乳突单个长大于A、B、C类群,但叶乳突簇间隙距离低于其他类群。A类群叶乳突长宽比变大,其他指标均变小;B类群叶乳突簇间隙距离变大,其余指标均变小;C类群叶乳突长宽比、簇间隙距离、总簇宽均变大,其余指标均变小;D类群叶乳突簇宽变小,叶乳突单个长、单个宽、长宽比、面积、簇间隙距离、总簇宽均变大。4个类群枯黄率排名为A>B>C>D,A类群的枯黄率较高,耐盐性较差,D类群的枯黄率较低,耐盐性较好,B、C类群的耐盐性中等。从表7中可知,盐处理后D类群除叶乳突簇宽以外其余指标均增大,叶乳突总簇宽、长宽比分别增加了14.77%、13.65%;A类群叶乳突各指标均减小,叶乳突单个宽、面积,分别减小了28.31%、38.09%。
表 6 40份群海雀稗各性状胁迫后不同类群差异分析
Table 6. Differential analysis of traits among different groups of 40 accessions of Paspalum vaginatum following stress exposure
A类群
Group AB类群
Group BC类群
Group CD类群
Group DLpl/μm 43.72±10.87a 53.85±7.21b 53.61±7.17b 56.18±7.58b Lpw/μm 15.14±4.47a 20.04±3.80b 18.72±2.29b 19.52±2.42b Lpr/μm 2.96±0.29a 2.78±0.42a 2.90±0.34a 2.90±0.20a Lpv/μm 73.33±13.77a 73.86±7.40a 81.93±6.20b 78.33±5.58ab Ap/μm 35436.55 ±14216.59 a52480.548 ±10798.80 b51011.52 ±10917.29 b51822.52 ±11698.97 bLg/μm 20.88±4.41a 24.36±9.26a 23.21±6.00a 19.96±3.25a Lpg/(ind.μm−2) 260.75±40.00a 257.97±29.54a 292.30±24.59b 274.90±18.75ab 表 7 叶乳突盐处理前后缩小比例
Table 7. Reduction in proportion of leaf papillae following salt treatment
指标
ParameterA类群
Group AB类群
Group BC类群
Group CD类群
Group DLpl/μm 23.75% 12.77% 6.73% −1.65% Lpw/μm 28.31% 1.04% 10.26% −6.14% Lpv/μm −8.82% 11.46% −3.94% 4.61% Lpg/(ind.μm−2) 6.38% 9.72% −3.59% −14.77% Ap/μm 38.09% 14.85% 15.67% −1.54% Lg/μm 6.91% −9.83% 13.80% −1.99% Lpr/μm 6.83% 10.98% −3.95% −13.65% 注:正数代表减少的百分比,负数代表增加的百分比。 Note: Positive numbers represent a percentage decrease, while negative numbers represent a percentage increase. -
已有研究表明,海雀稗可通过将Na+储存于叶乳突结构内,降低离子毒害对植株的损伤,进而增强盐胁迫适应性[6]。然而,之前研究仅以少量种质资源为试验材料,结论代表性受限,且盐胁迫下海雀稗叶乳突是否存在普遍性变化特征与耐盐性的内在关联仍不明确。基于此,本研究以40份海雀稗种质为试验材料,系统探究盐胁迫对叶乳突形态的影响及其与耐盐性的相关性。结果表明,盐胁迫处理后,耐盐种质叶乳突体积增大,敏盐种质则呈明显缩小趋势,叶乳突单个长(Lpl)、单个宽(Lpw)和面积(Ap)相比于胁迫前显著减小,说明盐分胁迫使乳突发育受到抑制或发生形态收缩。这一现象可能与植物为减少蒸腾表面积、限制离子侵入而作出的适应性调整有关[19]。叶乳突作为由表皮细胞外凸形成的特化结构,其形态可塑性是植物应对环境胁迫的重要策略之一[20]。乳突结构并非海雀稗所特有,在部分禾本科植物中也发现存在类似的表皮特化结构。例如,一些耐盐或抗旱的禾本科作物及野生近缘种(如小麦族Triticeae、稻属Oryza等)的叶片表面同样分布有乳突,其在形态和功能上与海雀稗有一定的相似性[21-23],是了解叶乳突阐明海雀稗耐盐机制的关键一步。
主成分分析结果显示,主成分1和主成分2累计贡献率达79.75%,表明所选形态指标能够充分代表叶乳突的结构变异。其中,第1主成分主要代表乳突的大小特征(如Lpl、Lpw和Ap),第2主成分则更多反映乳突的排列特征(如Lg和Lpg)。这一结果说明,海雀稗叶乳突的形态变异主要体现在尺寸上,而非构型或排列方式上。聚类分析将40份种质划分为4个类群,各类群在乳突形态上存在明显区别。B类群在Lpw、Lg及Ap方面均显著大于A、C、D类群,表明该类群种质具有更为发达的乳突结构且叶乳突较大,具有更好的离子储存能力。然而,值得注意的是,4个类群间叶乳突长宽比(Lpr)无显著差异,再次印证了乳突形态在构型上的保守性[24]。A类群所有测量指标胁迫后均显著变小,Lpl、Lpw和Ap分别变小了23.75%、28.31%和38.09%,且A类群耐盐性较差。D类群胁迫后除Lpr外其余指标均增大,最大增加了14.77%,且D类群耐盐性较好。虽然叶乳突形态在盐胁迫下发生变化,且可能通过物理阻隔、调节蒸腾和影响叶片表面离子再分配等方式参与耐盐响应,但并非决定海雀稗耐盐性的唯一或主导因素。植物的耐盐性是一个由形态、生理、生化及分子多层次机制共同构成的复杂性状[25]。研究表明[26-27],叶乳突可将植物体内危害植物生长的Na+封存在叶乳突内,但是否与植物的耐盐性有关仍需进一步研究。在盐处理后,4个类群Lpl、Lpw和Ap普遍显著减小,表明盐胁迫对海雀稗叶乳突形态存在抑制作用,可能是植物在盐环境下通过缩减乳突结构以降低能量消耗的适应性响应[28]。部分耐盐种质可通过盐胁迫下叶乳突特征增大的方式响应胁迫,其余耐盐种质则可能通过其他生理或形态调控途径规避盐胁迫伤害[29]。相关性分析显示,胁迫前后Lpl、Lpw和Ap均呈极显著(P<0.001)正相关,说明这些形态指标在结构上具有高度一致性,共同构成了叶乳突的物理基础。但胁迫后Lpr与Lpw呈极显著(P<0.001)负相关,说明在胁迫环境下,叶乳突可能倾向于变得更短而宽,这种形态变化可能有助于增强其结构的稳定性和功能效率。胁迫后叶乳突簇宽(Lpv)、Lpw与Rwn2d呈显著(P<0.05)负相关,说明Lpv、Lpw越小,植物在复水后的恢复情况反而越好,为种质筛选提供了新思路,并不是叶乳突越大越好。
本研究首次构建了海雀稗叶乳突的观察和测量方法,Lpl、Lpw、Ap和Lpv可作为研究叶乳突耐盐性的主要表型指标。Lpl、Lpw和Ap呈极显著正相关,利用主成分分析将40份种质进行聚类得到4个不同类群种质,4个类群按照耐盐性大小强弱顺序为D>C>B>A类群。由叶乳突盐处理前后的缩小比例可知,海雀稗耐盐性高,叶乳突有增大的趋势;耐盐性差,叶乳突有减小的趋势,反映出叶乳突形态与耐盐能力之间存在一定关联。相关性分析表明,海雀稗叶乳突特征与植株耐盐性存在一定关联,尤其与Rwn2d呈现显著相关性,推测该关联可能与植株渗透压调节能力及水分储存功能密切相关。本研究结果为解析海雀稗耐盐机制提供了新视角,同时为后续大规模开展海雀稗叶乳突相关研究奠定了理论基础。
Effects of salt treatment on leaf papillae characteristics of Paspalum vaginatum
-
摘要: 为了探讨不同种质海雀稗叶乳突与耐盐性之间的潜在关系,本研究以40份海雀稗种质(Paspalum vaginatum)为材料,在200 mmol·L-1的NaCl处理条件下进行研究,通过测量叶乳突单个长、单个宽、簇宽等指标并通过相关性分析、主成分分析及聚类分析开展系统研究。结果表明,不同种质海雀稗的叶乳突各性状指标间均存在显著差异(P<0.05),且性状间呈显著(P<0.05)正相关或负相关;胁迫前后叶乳突单个长、单个宽和面积均呈极显著正相关(P<0.001),胁迫后叶乳突簇宽和单个宽与胁迫复水后第二天呈显著负相关(P<0.05)。主成分分析表明,主成分1和主成分2累计贡献率79.75%,叶乳突形态变异主要源于叶乳突单个长、单个宽、面积。聚类分析将种质划分为4个类群, A类群叶乳突胁迫后明显变小,叶乳突面积减小了38.09%,且A类群耐盐性较差;D类群胁迫后叶乳突变大,叶乳突总簇宽增加了14.77%,耐盐性较好;B、C类群叶乳突除个别指标外均减小。Abstract: To explore the potential relationship between the leaf papillae of different accessions of Paspalum vaginatum and salt tolerance, 40 accessions of Paspalum vaginatum were selected and treated with 200mmol/L NaCl salt in the experiment, and their leaf papillae traits such as the length and width of individual leaf papilla, and cluster width were measured and analyzed by using correlation analysis, principal component analysis, and cluster analysis. The results showed that there were significant differences (P<0.05) in leaf papillae traits among different accessions of P. vaginatum, and that the traits were significantly (P<0.05) positively or negatively correlated. Before and after salt stress, the length, width and area of individual leaf papilla were highly significantly positively correlated (P<0.001). After stress, the cluster width of leaf papillae and the individual width showed a significant negative correlation with the second day after rehydration (P<0.05). Principal component analysis indicates that principal components 1 and 2 collectively account for 79.75% of the variance. Variation in leaf papilla morphology primarily stems from the individual length, individual width, and area of the leaf papillae. Cluster analysis grouped the accessions of the germplasm into four. Group A exhibited significantly reduced leaf papillae following stress, with papillae area decreasing by 38.09%, and demonstrated poor salt tolerance. Group D showed enlarged leaf papillae after stress, with total papilla cluster width increasing by 14.77%, and exhibited good salt tolerance. Group B and C displayed that most of the parameters in leaf papillae were reduced except for few parameters.
-
Key words:
- Paspalum vaginatum /
- leaf papillae /
- salt tolerance /
- change characteristics
-
图 2 海雀稗叶乳突测量方案
注:1表示叶乳突簇宽;2表示叶乳突总簇宽;3表示叶乳突簇间隙距离;4表示叶乳突面积;5表示叶乳突单个长;6表示叶乳突单个宽。
Fig. 2 Measurement scheme of Paspalum vaginatum leaf papillae
Note: 1 means the width of the leaf papilla cluster; 2 means the total width of the leaf papilla cluster; 3 means the width of the gaps between leaf papillae; 4 means the area of the leaf papilla; 5 means the length of a single leaf papilla; 6 means the width of a single leaf papilla.
图 4 胁迫后不同种质叶乳突特征相关性
注:Kh.胁迫枯黄率;Rkh.胁迫复水后枯黄率;SI.胁迫综合指标;RSI.胁迫复水后盐害指数;wn.胁迫萎蔫率;1~7 d.处理天数,下同。
Fig. 4 Correlation of leaf papilla characteristics among different accessions of the germplasm following stress exposure
Note: Kh: Wilting rate under stress; Rkh: Wilting rate after rehydration under stress; SI: Comprehensive stress index; RSI: Salt damage index after rehydration under stress; wn: Wilting rate under stress; 1~7 d:.Number of treatment days; similarly hereinafter.
图 3 胁迫前不同种质叶乳突特征相关性
注:*表示P<0.05;**表示P<0.01;***表示P<0.001;下同。
Fig. 3 Correlation of different genetic backgrounds in the characteristics of leaf papillae before stress treatment
Note: *mean P<0.05; ** mean P<0.01; *** mean P<0.001; similarly hereinafter.
图 5 海雀稗叶乳突各指标的GGE双标图
Fig. 5 GGE dual-scale diagram of various indicators for the papillae of the leaves of Paspalum vaginatum
图 6 40份海雀稗胁迫后主成分综合得分聚类分析
Fig. 6 Cluster analysis of the composite principal component scores for 40 accessions of Paspalum vaginatum following stress exposure
表 1 40份海雀稗种质信息来源
Table 1 Source information on 40 accessions of the germplasm of Paspalum vaginatum
种质来源
Germplasm sources材料名称
Name of germplasm阿根廷
Argentina17USA-34,PI509018-2,17USA-06 南非
South Africa17USA-11 马尔代夫
Maldives马尔代夫 中国广东
Guangdong, China18HN-90 中国海南
Hainan, China18HN-101,17HN-02,琼2,17HN-45,17HN-54,17HN-25 美国
America17USA-03,KaiLuna,17USA-17,TFP7-4,17USA-45,17USA-30,HYB7,17-35,17USA-07,HYB5,17USA-31,18HN-100,17USA-41,18HN-98,Taliaferra,17USA-42,TCR3,TGkona,VeroBeach,Polo,Saiam,17USA-23,TOCGC, 17USA-18,Kim1 ,17USA-15,Sealsle2000,Wailual kauai  下载: 导出CSV
下载: 导出CSV
表 2 盐胁迫前后叶乳突特征描述统计
Table 2 Statistical description of leaf papilla characteristics before and after salt stress
指标
Parameter胁迫前
Non-stress处理
Salt stress最大值
Max最小值
Min均值±标准差
Meam±SD变异系数/%
CV/%最大值
Max最小值
Min均值±标准差
Meam±SD变异系数/%
CV/%Lpl/μm 95 28.74 57.91±11.03a 19 83.72 20.93 52.13±10.78b 21 Lpw/μm 31.35 9.36 20.36±4.02a 19 30.67 6.37 18.41±4.08b 22 Lpr/μm 5.58 1.82 2.89±0.50a 17 5.70 1.79 2.89±0.51a 17 Lpv/μm 122.01 31.92 77.86±13.81a 17 106.57 39.06 77.97±12.62a 16 Ap/(ind.μm−2) 107502.41 22249.62 58414.07 ±15913.37 a27 84782.22 10099.44 48332.26 ±13994.94 b29 Lg/μm 47.93 6.56 21.50±6.01a 28 44.08 8.58 21.07±5.83a 28 Lpg/μm 363.03 162.9 275.77±30.94a 11 338.53 182.96 276.08±31.33a 11 注: Lpl.叶乳突单个长,Lpw.单个叶乳突宽,Lpr.叶乳突长宽比,Lpv.叶乳突簇宽,Ap.6个叶乳突面积,Lg.叶乳突簇间隙距离,Lpg.叶乳突总簇宽。均值±标准差后不同小写字母表示同一处理时间,不同指标在0.05水平下差异显著;下同。 Note: Lpl. Single length of papillae; Lpw.Width of single papillae; Lpr. Length and width ratio of papillae; Lpv. Width of papillae clusters; Ap.6leaf mastoid areas; Lg. Wide gap of papillae; Lpg. Width of total clusters of papillae. Different lowercase letters following the mean ± standard deviation denote significant differences at the 0.05 level for the same treatment time across different indicators, similarly hereinafter.
下载: 导出CSV
表 3 海雀稗种质盐胁迫后叶乳突主成分分析
Table 3 Principal component analysis of leaf papillae in Paspalum vaginatum following salt stress
性状
Traits主成分1
Comp.1主成分2
Comp.2Lpl/μm 0.89 −0.02 Lpw/μm 0.84 −0.49 Lpr/μm −0.24 0.83 Lpv/μm 0.83 0.15 Ap/μm 0.89 −0.13 Lg/μm 0.40 0.76 Lpg/(ind·μm−2) 0.84 0.38 特征值
Characteristic value3.89 1.69 贡献率/%
Contribution rate/%55.54 24.21 累计贡献率/%
Cumulative contribution rate/%55.54 79.75
下载: 导出CSV
表 4 40份海雀稗种质叶乳突性状排名
Table 4 Ranking of leaf papillae traits in 40 accessions of Paspalum vaginatum germplasm
种质
SamplesD值
D values排名
Ranking种质
SamplesD值
D values排名
Ranking17USA-03 3.60 1 Kim 1 0.11 21 17USA-07 2.14 2 Salam 0.10 22 18HN-90 1.75 3 TOCGC 0.07 23 Kai Luna 1.43 4 17USA-34 0.07 24 17USA-35 1.35 5 TFP 7-4 −0.12 25 17USA-30 1.27 6 琼2 −0.15 26 17HN-54 1.20 7 17USA-17 −0.33 27 马尔代夫 1.10 8 17USA-45 −0.57 28 17USA-06 1.01 9 Taliaferra −0.68 29 17USA-31 0.88 10 17USA-18 −0.74 30 17USA-42 0.82 11 18HN-101 −0.79 31 17HN-02 0.77 12 Polo −0.80 32 18HN-98 0.70 13 17USA-23 −1.00 33 HYB 5 0.58 14 TG Kona −1.17 34 Vero Beach 0.58 15 PI 509018-2 −1.53 35 18HN-100 0.53 16 17HN-25 −1.72 36 17USA-11 0.53 17 SeaIsle2000 −1.85 37 17HN-45 0.32 18 TCR 3 −2.23 38 17USA-41 0.18 19 17USA-15 −3.26 39 HYB 7 0.15 20 Wailua Kauai −4.31 40
下载: 导出CSV
表 5 40份群海雀稗各性状不同类群差异分析
Table 5 Differential analysis of traits among different groups of 40 accessions of Paspalum vaginatum
A类群
Group AB类群
Group BC类群
Group CD类群
Group DLpl/μm 57.34±13.96a 61.73±6.03a 57.48±6.56b 55.27±7.48b Lpw/μm 21.12±4.88a 20.25±3.73a 20.86±2.34b 18.39±2.14b Lpr/μm 2.72±0.12a 3.14±0.42a 2.79±0.23a 3.04±0.23a Lpv/μm 78.33±8.15a 81.81±8.11a 79.09±6.90b 68.25±9.44b Ap/μm 57237.71 ±20909.92 a61634.79 ±10928.60 a60489.02 ±11242.87 b51038.51 ±12962.65 bLg/μm 22.43±3.63a 22.18±2.01a 21.96±7.23a 18.56±5.20a Lpg/(ind.μm-2) 279.86±19.61a 289.78±24.72a 281.19±26.97b 241.89±37.12b 注:不同小写字母表示同一处理时间,不同指标在0.05水平下差异显著;下同。 Note: Different lowercase letters denote significant differences between parameters at the 0.05 level at the same treatment time, similarly hereinafter.
下载: 导出CSV
表 6 40份群海雀稗各性状胁迫后不同类群差异分析
Table 6 Differential analysis of traits among different groups of 40 accessions of Paspalum vaginatum following stress exposure
A类群
Group AB类群
Group BC类群
Group CD类群
Group DLpl/μm 43.72±10.87a 53.85±7.21b 53.61±7.17b 56.18±7.58b Lpw/μm 15.14±4.47a 20.04±3.80b 18.72±2.29b 19.52±2.42b Lpr/μm 2.96±0.29a 2.78±0.42a 2.90±0.34a 2.90±0.20a Lpv/μm 73.33±13.77a 73.86±7.40a 81.93±6.20b 78.33±5.58ab Ap/μm 35436.55 ±14216.59 a52480.548 ±10798.80 b51011.52 ±10917.29 b51822.52 ±11698.97 bLg/μm 20.88±4.41a 24.36±9.26a 23.21±6.00a 19.96±3.25a Lpg/(ind.μm−2) 260.75±40.00a 257.97±29.54a 292.30±24.59b 274.90±18.75ab
下载: 导出CSV
表 7 叶乳突盐处理前后缩小比例
Table 7 Reduction in proportion of leaf papillae following salt treatment
指标
ParameterA类群
Group AB类群
Group BC类群
Group CD类群
Group DLpl/μm 23.75% 12.77% 6.73% −1.65% Lpw/μm 28.31% 1.04% 10.26% −6.14% Lpv/μm −8.82% 11.46% −3.94% 4.61% Lpg/(ind.μm−2) 6.38% 9.72% −3.59% −14.77% Ap/μm 38.09% 14.85% 15.67% −1.54% Lg/μm 6.91% −9.83% 13.80% −1.99% Lpr/μm 6.83% 10.98% −3.95% −13.65% 注:正数代表减少的百分比,负数代表增加的百分比。 Note: Positive numbers represent a percentage decrease, while negative numbers represent a percentage increase.
下载: 导出CSV
-
[1] Reddy M P K, Talekar N, Kumar A, et al. A review on understanding the effects and mechanisms of salinity tolerance in rice (Oryza sativa L. ) [J]. International Journal of Plant & Soil Science, 2023, 35(17): 209−219. https://doi.org/10.9734/IJPSS/2023/v35i173201 doi: 10.9734/IJPSS/2023/v35i173201 [2] 杨文涛, 王琦, 郭二丹, 等. 土壤盐渍化对植物影响的研究进展[J]. 农业与技术, 2024, 44(18): 95−99. https://doi.org/10.19754/j.nyyjs.20240930022 doi: 10.19754/j.nyyjs.20240930022 [3] 申晴, 韦海燕, 卞华, 等. 海雀稗种质资源的耐盐性评价[J]. 热带生物学报, 2020, 11(1): 11−19. https://doi.org/10.15886/j.cnki.rdswxb.2020.01.003 doi: 10.15886/j.cnki.rdswxb.2020.01.003 [4] 洪佳琦. 海雀稗种质资源遗传多样性研究[D]. 乌鲁木齐: 新疆农业大学, 2019. https://doi.org/10.27431/d.cnki.gxnyu.2019.000360 [5] Trenholm L E, Carrow R N, Duncan R R. Mechanisms of wear tolerance in seashore paspalum and bermudagrass [J]. Crop Science, 2000, 40(5): 1350−1357. https://doi.org/10.2135/cropsci2000.4051350x doi: 10.2135/cropsci2000.4051350x [6] Spiekerman J J, Devos K M. The halophyte seashore paspalum uses adaxial leaf papillae for sodium sequestration [J]. Plant Physiology, 2020, 184(4): 2107−2119. https://doi.org/10.1104/pp.20.00796 doi: 10.1104/pp.20.00796 [7] Lv Y P, Gu L H, Man R Z, et al. Response of stomatal conductance, transpiration, and photosynthesis to light and CO2 for Rice leaves with different appearance days [J]. Frontiers in Plant Science, 2024, 15: 1397948. https://doi.org/10.3389/fpls.2024.1397948 doi: 10.3389/fpls.2024.1397948 [8] Chaves M, Davies B. Drought effects and water use efficiency: improving crop production in dry environments [J]. Functional Plant Biology, 2010, 37(2): iii−vi. https://doi.org/10.1071/FPv37n2_FO doi: 10.1071/FPv37n2_FO [9] 马秋雨, 袁芳. 植物盐腺泌盐及发育研究进展[J]. 生物技术通报, 2023, 39(11): 74−85. https://doi.org/10.13560/j.cnki.biotech.bull.1985.2023-0581 doi: 10.13560/j.cnki.biotech.bull.1985.2023-0581 [10] 孙劝劝, 祝青, 胡旭, 等. 盐生植物海雀稗的气孔特征与光合特性[J]. 热带生物学报, 2025, 16(4): 528−536. https://doi.org/10.15886/j.cnki.rdswxb.20240085 doi: 10.15886/j.cnki.rdswxb.20240085 [11] Wu Y H, Cheng J Q, Feng H Y, et al. Advances of research on desiccation-tolerant moss [J]. Journal of Desert Research, 2004, 24(1): 23−29. (查阅网上资料,本条文献为中文文献,请确认) https://doi.org/10.3321/j.issn:1000-694X.2004.01.004 [12] Aist J R, Israel H W. Autofluorescent and ultraviolet-absorbing components in cell walls and papillae of barley coleoptiles and their relationship to disease resistance [J]. Canadian Journal of Botany, 1986, 64(2): 266−272. https://doi.org/10.1139/b86-039 doi: 10.1139/b86-039 [13] 毛学文. 植物的毛状体[J]. 生物学教学, 2002, 27(11): 38−39. https://doi.org/10.3969/j.issn.1004-7549.2002.11.030 doi: 10.3969/j.issn.1004-7549.2002.11.030 [14] Arteaga N, Savic M, Méndez-Vigo B, et al. MYB transcription factors drive evolutionary innovations in Arabidopsis fruit trichome patterning [J]. The Plant Cell, 2021, 33(3): 548−565. https://doi.org/10.1093/plcell/koaa041 doi: 10.1093/plcell/koaa041 [15] Zhu L, Zeng F L, Liang Y P, et al. Small papillae regulated by SPD25 are critical for balancing photosynthetic CO2 assimilation and water loss in rice [J]. Rice, 2023, 16(1): 58. https://doi.org/10.1186/s12284-023-00676-7 doi: 10.1186/s12284-023-00676-7 [16] 刘双平, 俞熹. 荷叶效应的研究[J]. 大学物理, 2011, 30(9): 50−54. https://doi.org/10.3969/j.issn.1000-0712.2011.09.015 doi: 10.3969/j.issn.1000-0712.2011.09.015 [17] 王力, 胡琴, 舒立春, 等. 荷叶超疏水表面防覆冰试验及机理研究[J]. 电工技术学报, 2025, 40(3): 855−863. https://doi.org/10.19595/j.cnki.1000-6753.tces.232155 doi: 10.19595/j.cnki.1000-6753.tces.232155 [18] Câmara P E A S, Kellogg E A. Morphology and development of leaf papillae in Sematophyllaceae [J]. The Bryologist, 2010, 113(1): 22−33. https://doi.org/10.1639/0007-2745-113.1.22 doi: 10.1639/0007-2745-113.1.22 [19] Ghannoum O. C4 photosynthesis and water stress [J]. Annals of Botany, 2009, 103(4): 635−644. https://doi.org/10.1093/aob/mcn093 doi: 10.1093/aob/mcn093 [20] Yoo J H, Park J H, Cho S H, et al. The rice bright green leaf (bgl) locus encodes OsRopGEF10, which activates the development of small cuticular papillae on leaf surfaces [J]. Plant Molecular Biology, 2011, 77(6): 631−641. https://doi.org/10.1007/s11103-011-9839-0 doi: 10.1007/s11103-011-9839-0 [21] 朱琳. 水稻叶表皮小乳突发育基因SPD25和粒型基因SRG3的克隆和功能分析[D]. 沈阳: 沈阳农业大学, 2024. https://doi.org/10.27327/d.cnki.gshnu.2024.000017 [22] 郭昱. 谷子核心种质叶片气孔、乳突密度全基因组关联分析[D]. 太原: 山西农业大学, 2020. https://doi.org/10.27285/d.cnki.gsxnu.2020.000577 [23] Jacobs A K, Lipka V, Burton R A, et al. An Arabidopsis callose synthase, GSL5, is required for wound and papillary callose formation [J]. The Plant Cell, 2003, 15(11): 2503−2513. https://doi.org/10.1105/tpc.016097 doi: 10.1105/tpc.016097 [24] 葛星辰. 谷子叶片乳突发育特征及关键调控基因SiICS的功能研究[D]. 太原: 山西农业大学, 2023. doi: 10.27285/d.cnki.gsxnu.2023.000342 [25] Munns R. Genes and salt tolerance: bringing them together [J]. New Phytologist, 2005, 167(3): 645−663. https://doi.org/10.1111/j.1469-8137.2005.01487.x doi: 10.1111/j.1469-8137.2005.01487.x [26] Meng X Q, Zhou J, Sui N. Mechanisms of salt tolerance in halophytes: current understanding and recent advances [J]. Open Life Sciences, 2018, 13(1): 149−154. https://doi.org/10.1515/biol-2018-0020 doi: 10.1515/biol-2018-0020 [27] Guo H L, Wang Y, Li D D, et al. Growth response and ion regulation of seashore paspalum accessions to increasing salinity [J]. Environmental and Experimental Botany, 2016, 131: 137−145. https://doi.org/10.1016/j.envexpbot.2016.07.003 doi: 10.1016/j.envexpbot.2016.07.003 [28] 吴欣欣, 白天惠, 张乐, 等. 泌盐盐生植物的泌盐机理研究进展[J]. 植物生理学报, 2020, 56(12): 2526−2532. https://doi.org/10.13592/j.cnki.ppj.2020.0370 doi: 10.13592/j.cnki.ppj.2020.0370 [29] Duarte-Silva A G, Carvalho-Silva M, Câmara P E A S. Morphology and development of leaf papillae in the Pilotrichaceae [J]. Acta Botanica Brasilica, 2013, 27(4): 737−742. https://doi.org/10.1590/S0102-33062013000400013 doi: 10.1590/S0102-33062013000400013 -

 点击查看大图
点击查看大图
计量
- 文章访问数: 300
- HTML全文浏览量: 226
- 被引次数: 0










