

-
植物在生长过程中常遭受生物和非生物胁迫,这2种胁迫都会影响植物的健康、生长和产量[1]。在极端温度、干旱和盐胁迫等非生物胁迫中,盐胁迫尤其具有挑战性。全球有100多个国家存在盐渍化土壤(电导率EC>2 dS·m−1)[2],据估计全球盐碱化土壤面积已达8.33亿hm2[3]。当土壤中的盐质量浓度明显高于植物生长的适宜质量浓度时,植物质膜的选择渗透性被破坏,导致细胞内溶质快速外渗和大量盐离子进入细胞,从而使植物受到盐毒害。在严重的盐胁迫下,植物脱水、气孔闭合、叶绿素含量下降及光合作用中相关酶的活性降低,从而抑制了植物地上部分的生长[4-6]。同时,也抑制了地下根系的生长。
研究表明,植物主要通过离子区域化、选择性摄取和分配等方式启动相关调节机制来适应高盐环境,以抵抗不利影响并维持离子稳态[7]。多项研究表明,MYB转录因子在植物胁迫响应中发挥关键作用,特别是在受非生物胁迫中[8]。在玉米(Zea mays)中鉴定出植物中第一个MYB基因C1,ZmMYBC1可以调节花青素的生物合成[9]。随后,越来越多的植物MYB超家族成员被鉴定出来[10],许多植物物种编码超过 100 种 R2R3 MYB 蛋白[11]。到目前为止,在拟南芥(Arabidopsis thaliana)、水稻(Oryza sativa)中分别报道了126[12]、155个MYB基因[13]。在拟南芥中,定位在细胞核上的AtMYB44能够增强转基因拟南芥气孔闭合的功能,从而提高植物的耐旱性和耐盐性[14]。同样定位于细胞核中的OsMYB2通过抑制或激活相关基因及通路在盐胁迫响应过程中发挥作用,ABA通路中的 OsMYB2在水稻盐毒害时,通过转录调控相关基因来调节水稻的生长和耐盐性[15]。因此,MYB家族成员在响应盐胁迫、干旱胁迫、生物合成、气孔调节等方面发挥了作用。
海雀稗广泛分布于全球各地,特别是在热带和亚热带的沿海地区。这种植物以其鲜艳的颜色、优良的草皮质地、旺盛的生长能力、出色的适应性和抗逆性而闻名。海雀稗适应海水灌溉,是盐碱地区和热带、亚热带沿海地区绿化工程的理想选择[16]。通过对不同品种的海雀稗耐盐能力进行评价分析,发现维持较高的 K+含量以及较高的地下 K+/Na+,可能是海雀稗耐盐的离子调节机制[17]。目前,未有MYB基因通过离子调节响应海雀稗盐胁迫机制的报道。因此,本研究分析了极端耐盐种质‘USA17-18’对盐胁迫的生长响应,克隆并分析了PvMYB1基因,为解析海雀稗响应盐胁迫提供理论依据。
-
供试材料为海雀稗‘USA17-18’种质资源,现保存于海南大学儋州校区农科基地。
-
参照陈静波[18]等的试验方法,剪下‘USA17-18’海雀稗顶端三节匍匐茎,挑选生长一致的匍匐茎用棉花固定在剪去尾部的离心管中于1/2 Hoagland营养液(pH5.8)中预培养17 d,参考申晴等[19]方法,分别进行以下处理:1)分别在0、15、30 g·L−1 NaCl处理液中(pH5.8)持续水培14 d,采集叶片与根系组织样本,用于后续RNA提取及相关基因表达研究;2)在30 g·L−1 NaCl处理液中(pH5.8),设置不同时间处理:0(CK)、5、10 d,收取叶和根系样品用于RNA提取和基因表达分析。
-
参考黄杰等[20]方法,使用 EPSON 12000XL 扫描仪(日本)扫描海雀稗根系图像。借助WinRhizo Pro软件(Regent Instruments,加拿大)测定盐胁迫下植株的总根长、根表面积及根体积等参数,完成定量评估。
-
参照邹晓燕等[21]的方法用TRNzol试剂(TIANGEN,中国)提取海雀稗RNA。海雀稗根叶组织的RNA提取步骤如下:称取0.1 g样本,加入1 mL TRIzol提取液,经破碎仪充分破碎。加入200 µL三氯甲烷,剧烈震荡混匀,室温静置3 min。4 ℃条件下11 999 r·min−1离心15 min,收集上清液转移至新的1.5 mL离心管。加入等体积的异丙醇,震荡混匀后置于−20 ℃反应2 h。4 ℃ 11 999 r·min−1离心10 min,去除上清液。加入500 µL无水乙醇清洗沉淀,4 ℃ 10 000 r·min−1离心5 min后去除上清液。最后加入900 µL 体积分数为75%的乙醇,4 ℃ 10 000 r·min−1离心5 min。弃上清,用枪头吸出残留液体,置于超净工作台中使沉淀风干,加入30 µL无RNase ddH2O溶解RNA。
cDNA合成参考HiScript III RT SuperMix for qPCR(+gDNA wiper)试剂盒方法。在PCR管中加入4 μL RNA、4 μL 4 × gDNA wiper Mix和8 μL RNase ddH2O,轻轻混匀,42 ℃反应2 min,置于冰上,加入4 μL 5 × HiScript III qRT SuperMix,轻轻混匀,37 ℃ 15 min,85 ℃ 5 s反应,获得cDNA,并置于−20 ℃保存备用。
-
根据本实验室前期海雀稗全长转录组结果,筛选得到PvMYB1基因全长序列。PvMYB1基因全长片段由生工生物工程(上海)股份有限公司连接到pUC-SP载体后,对其进行测序分析,确认PvMYB1序列。
-
利用Expasy(
https://web.expasy.org/ )对PvMYB1进行蛋白理化性质分析,在NCBI(https://www.ncbi.nlm.nih.gov/cdd/ )中对蛋白保守结构域进行分析;借助 SignalP v5.0在线分析平台(https://novopro.cn/tools/signalp ),完成PvMYB1蛋白信号肽预测,通过WoLF PSORT(https://wolfpsort.hgc.jp/ )预测亚细胞定位;采用MEGA v11软件构建蛋白进化树。 -
参考刘莉婷等[22]的方法,采用 QuantStudioTM Real-Time PCR仪器(Thermo Fisher,美国)进行实时荧光定量分析。20 μL 反应体系含:10 μL ChamQ Universal SYBR qPCR Master Mix(Vazyme,中国)、6 μL ddH2O、上下游引物各1 μL及被30倍稀释的cDNA模板。95 °C预变性 10 min;随后进行40个循环的扩增,每个循环包括 95 °C变性15 s、58 °C退火30 s、72 °C延伸30 s;循环结束后,于 95 °C 保持15 s 采集终末信号,随后进行熔解曲线分析。基于PvMYB1基因表达量与看家基因PvEF1α表达量的比值计算相对表达水平[23]。qPCR引物信息详见表1。
表 1 PvMYB1克隆 和 qPCR 引物分析
Table 1. Primers for PvMYB1 cloning and qPCR analysis
引物名称
Primer引物序列(5′—3′)
SequencePvMYB1-ORF-F ATGGGGCGGATCCCGTGCTGC PvMYB1-ORF-R TTACACCATGTGATTTGTCAGGTCA PvMYB1-RT-F GGCGCCTCATCCCCAAGAAT PvMYB1-RT-R GCCGCGATCACCGACCAC PvMYB1a-RT-F CACTGTCGAGTGGGTTCGCTAC PvMYB1a-RT-R GCCGATGAATTTTACTTGTTGC -
通过Microsoft Excel 2013与 SPSS v20.0软件完成数据处理与统计分析工作。
-
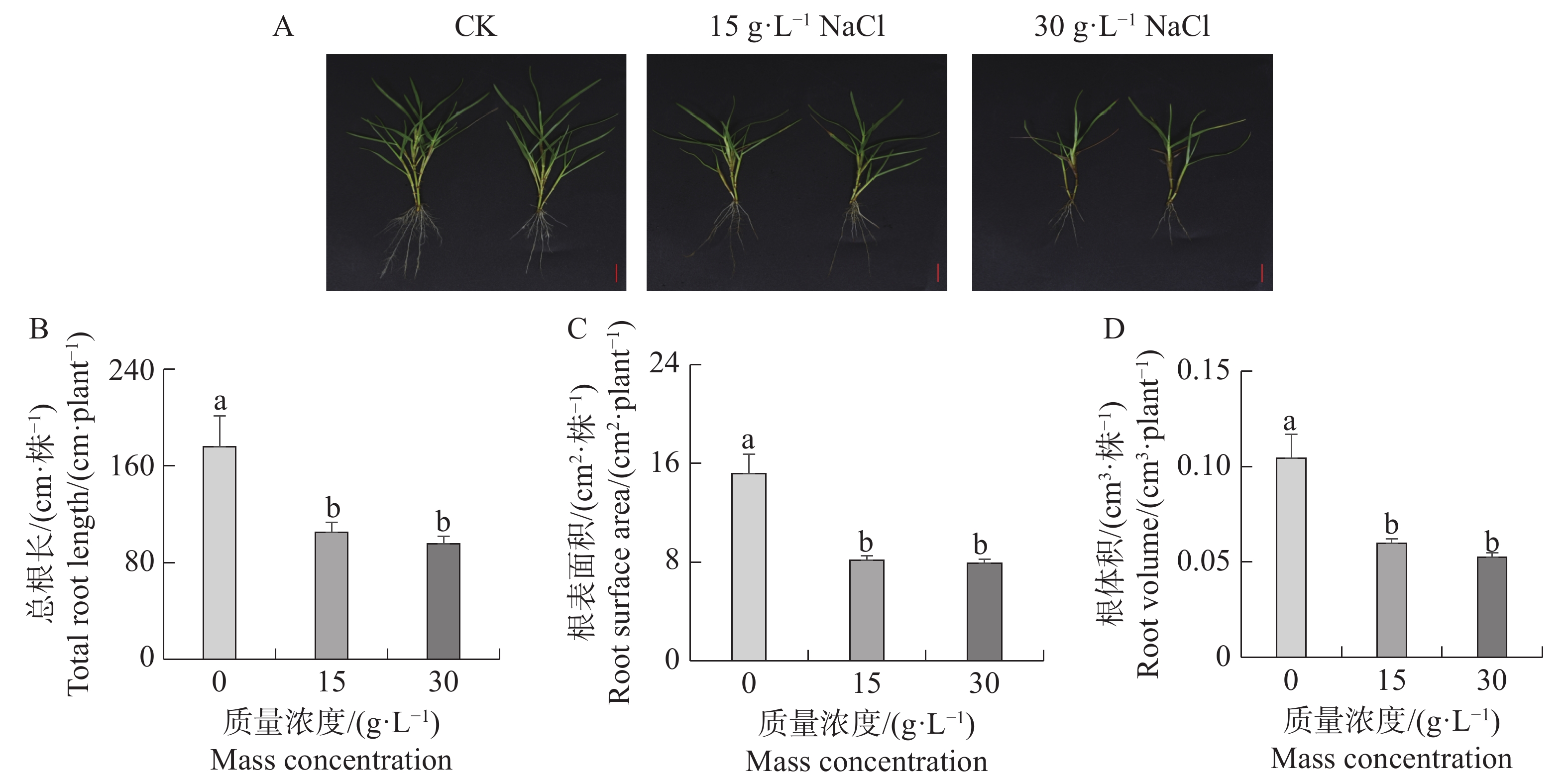
本研究分析了海雀稗根系受不同NaCl质量浓度胁迫的影响。从图1-A可以看出,盐胁迫抑制了海雀稗根系生长,NaCl处理浓度越高,根系生长受抑制程度越大。相对对照(0 g·L−1)处理,15、30 g·L−1 NaCl处理下海雀稗总根长被显著抑制。(图1-B—D)。
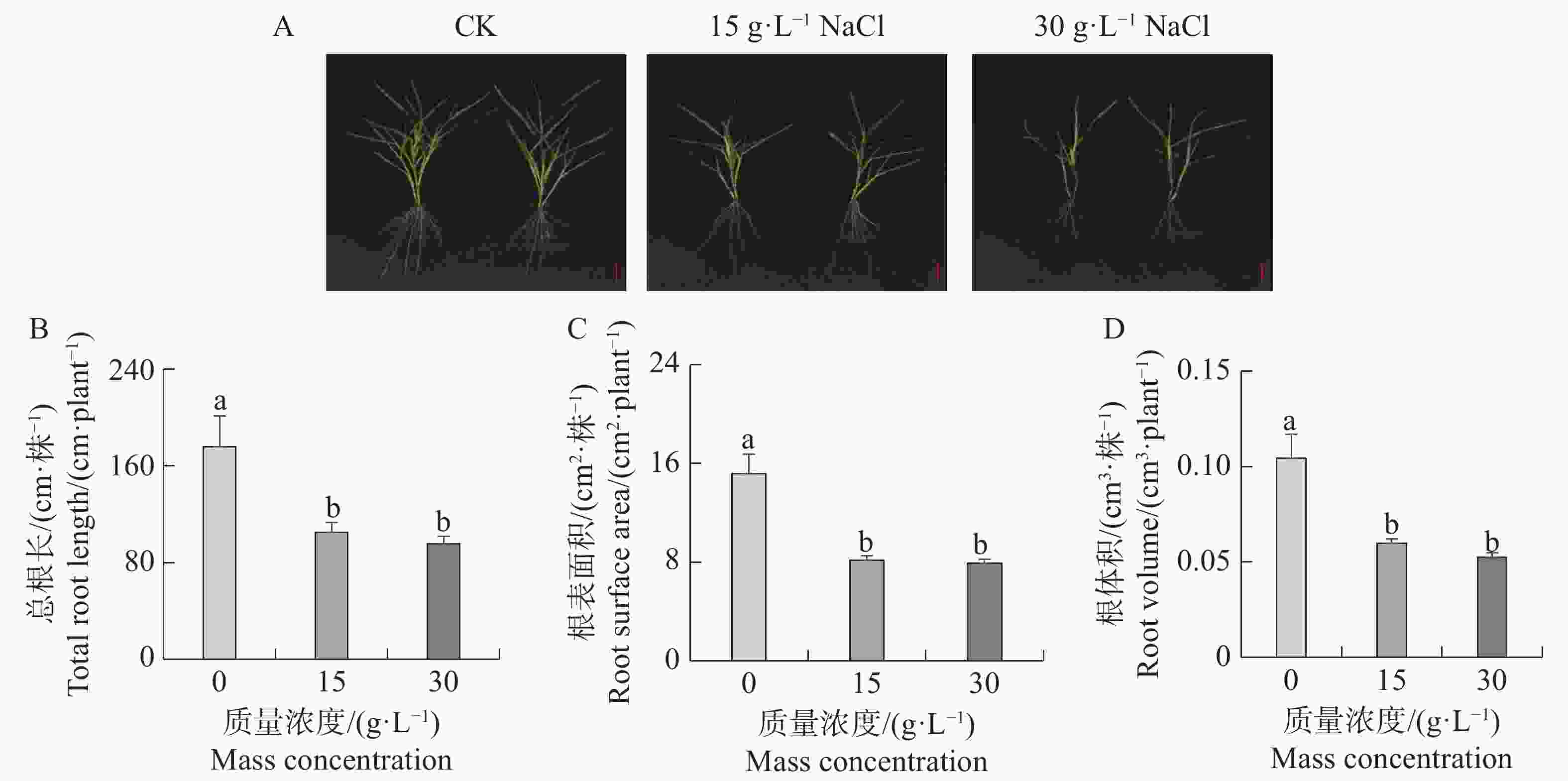
图 1 不同盐处理对海雀稗根系生长的影响
Figure 1. Effects of different NaCl treatments on root growth of Paspalum vaginatum
-
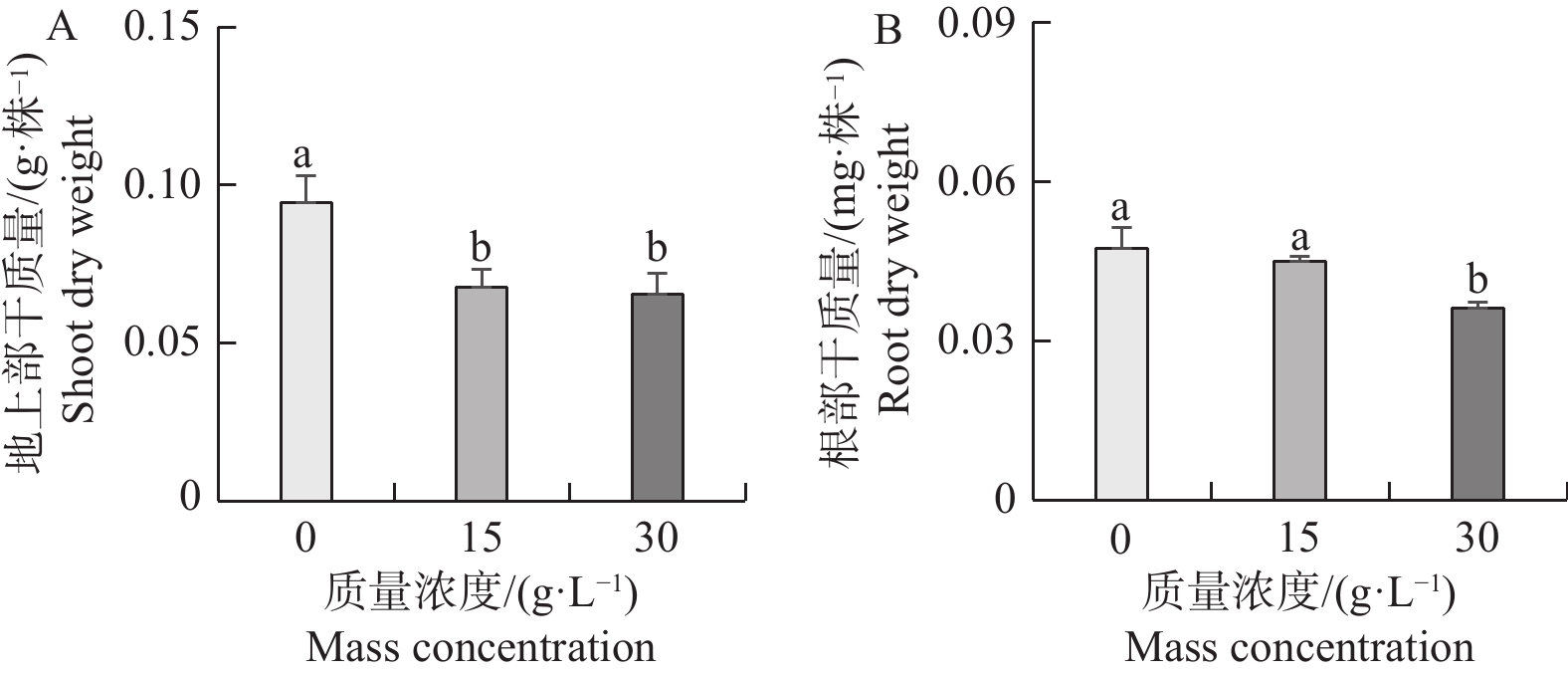
从图2-A,B可以看出,不同浓度NaCl处理不仅造成了地上部差异,同时也抑制了海雀稗根部干质量。相对对照(0 g·L−1)处理,15、30 g·L−1NaCl处理下海雀稗根部干质量被抑制了5.0%、23.7%,海雀稗地上部干质量被抑制了23.1%、35.9%。结果表明,随着NaCl浓度的升高,海雀稗的生长被抑制的程度有越来越严重的趋势。
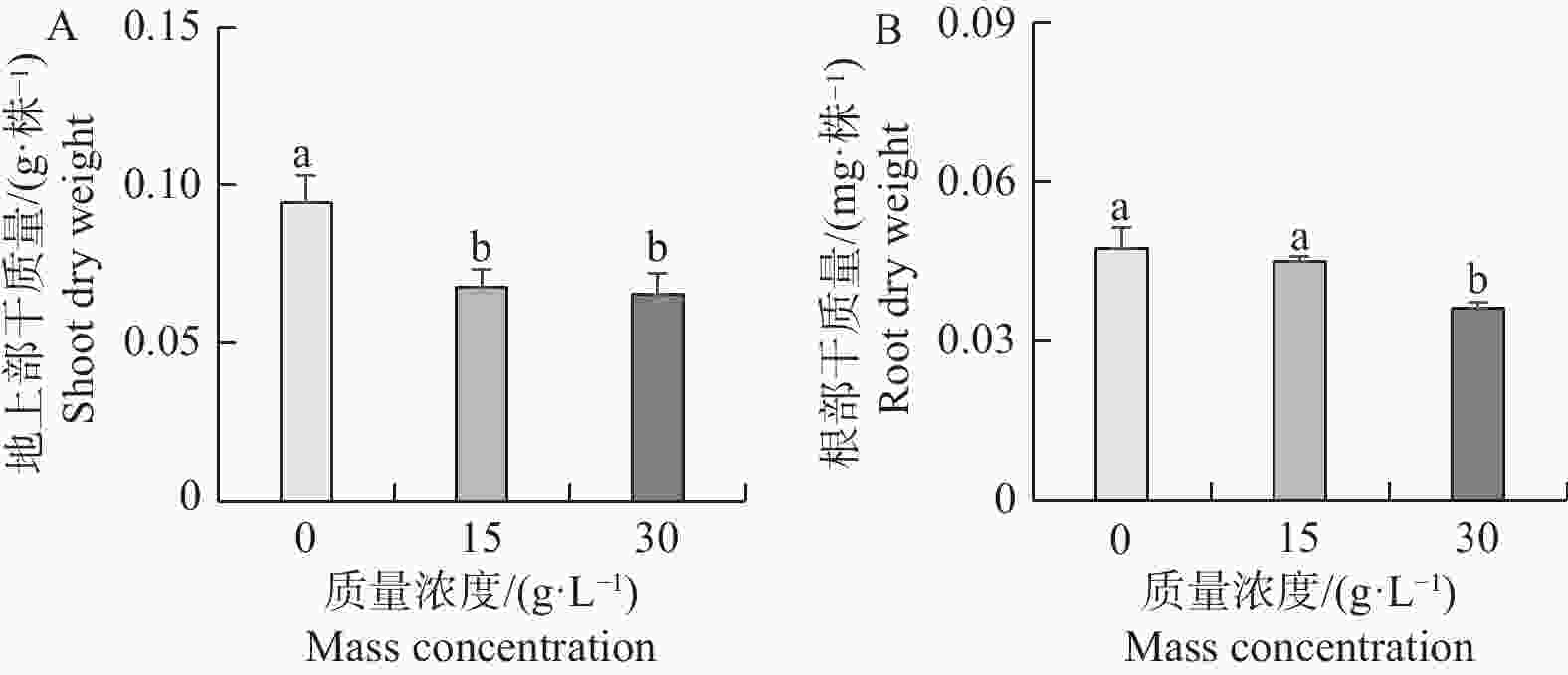
图 2 不同盐处理对海雀稗干质量的影响
Figure 2. Dry weight of Paspalum vaginatum under different NaCl treatments
-
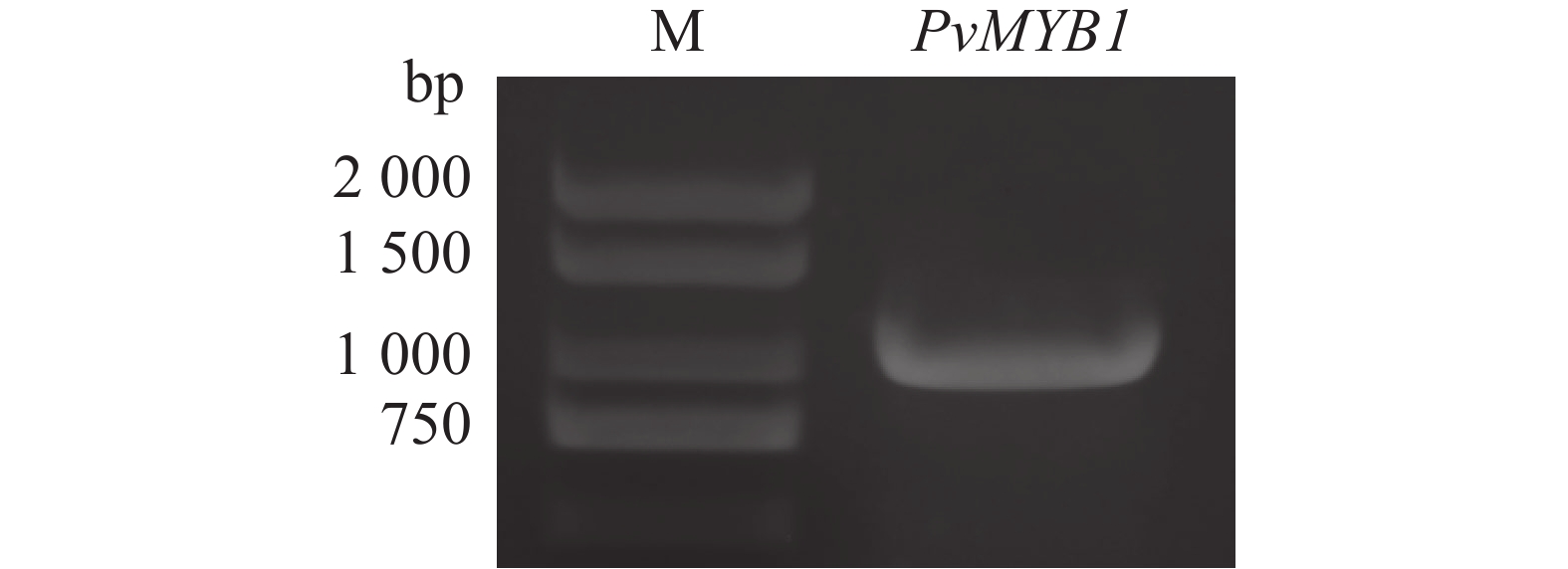
本研究通过PCR方法,利用PvMYB1-pYES2质粒扩增出PvMYB1基因(图3)。经PCR产物测序分析,PvMYB1基因包含
1 098 bp核苷酸序列,编码365个氨基酸残基。该蛋白分子质量大小为39.3 kDa,理论等电点(pI)5.7,属于亲水性蛋白。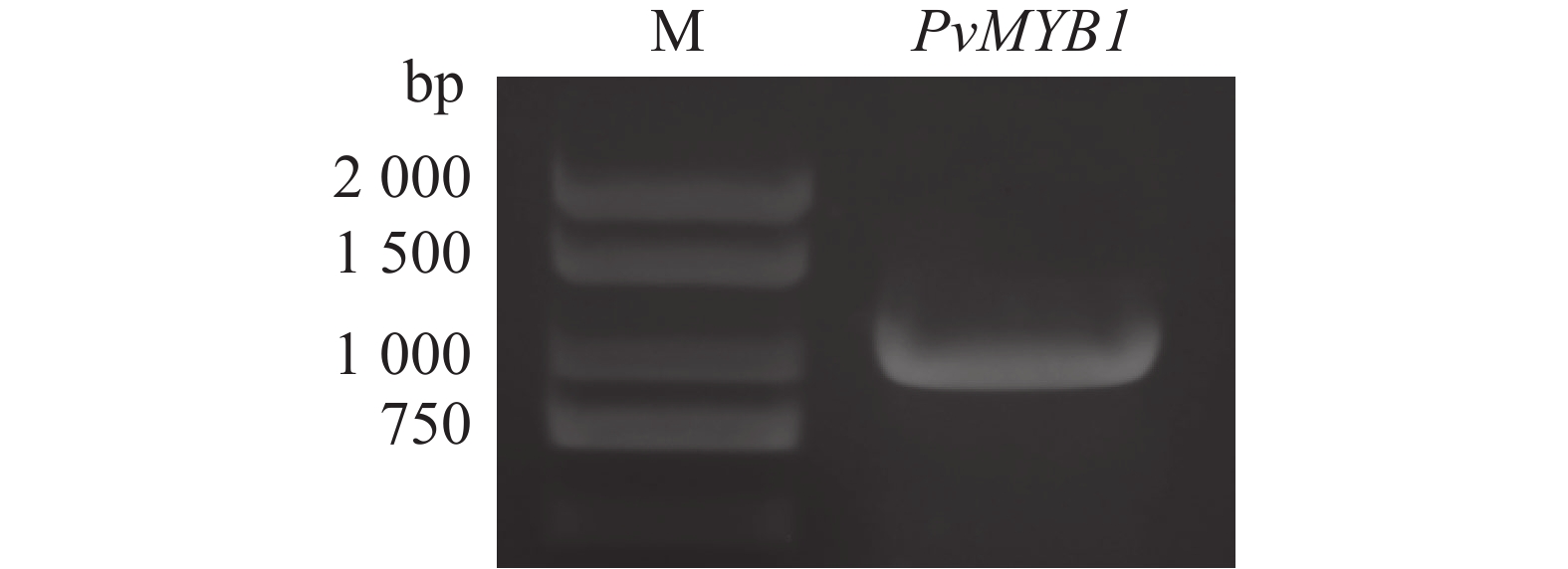
图 3 PvMYB1基因的克隆
Figure 3. Cloning of PvMYB1 gene
-
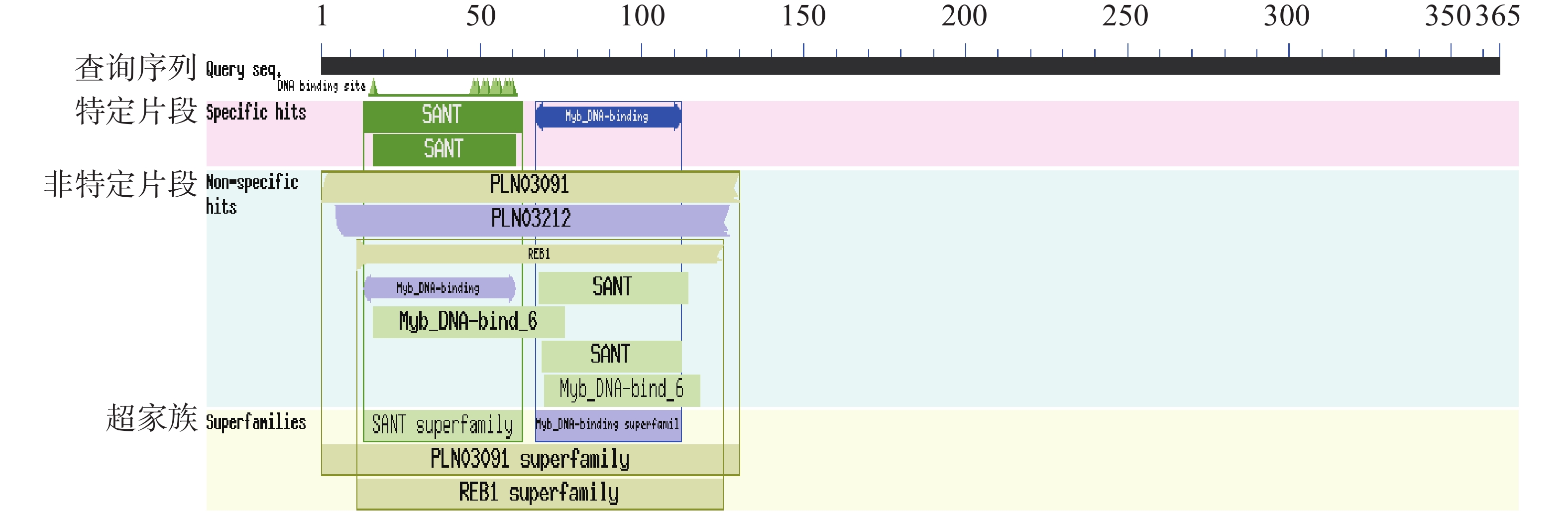
通过NCBI Conserved Domains数据库分析,蛋白质结构域(图4)预测显示PvMYB1蛋白N端具备两个保守SANT结构域,属于MYB转录因子R2R3亚家族。其中,第一个SANT域(14~63位)含50个氨基酸残基,第二个SANT域(17~61位)由45个氨基酸残基组成。经WoLF PSORT预测,该蛋白定位于细胞核。
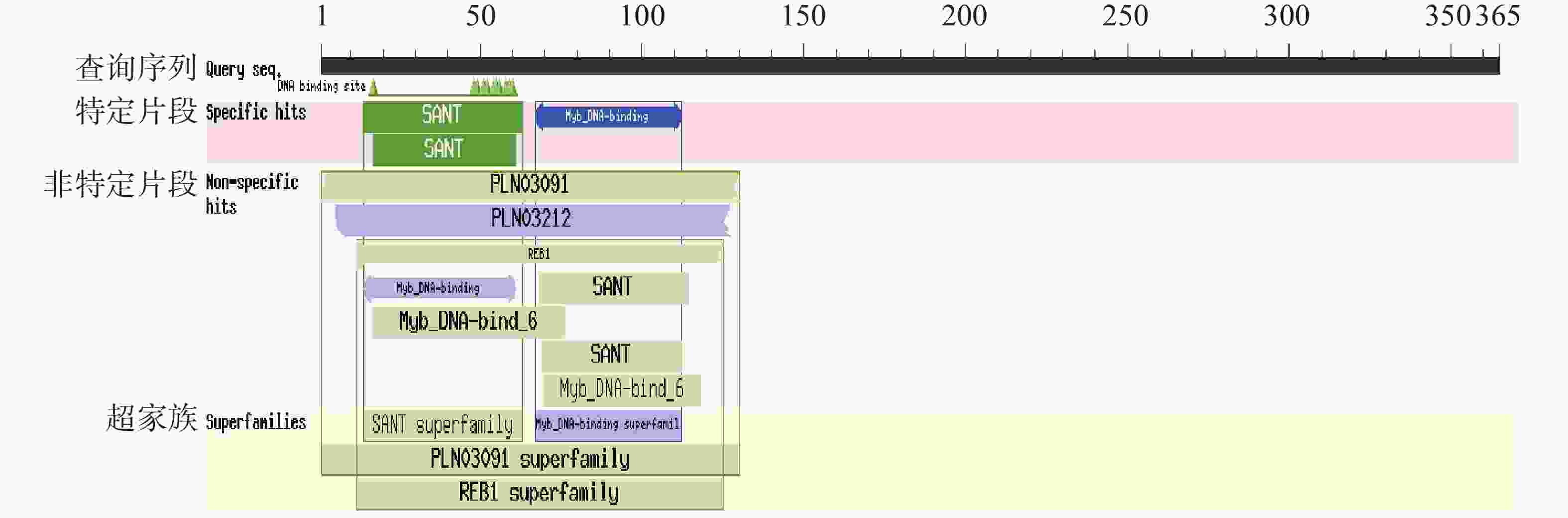
图 4 PvMYB1 保守结构域分析
Figure 4. Analysis of conserved domains of PvMYB1
-
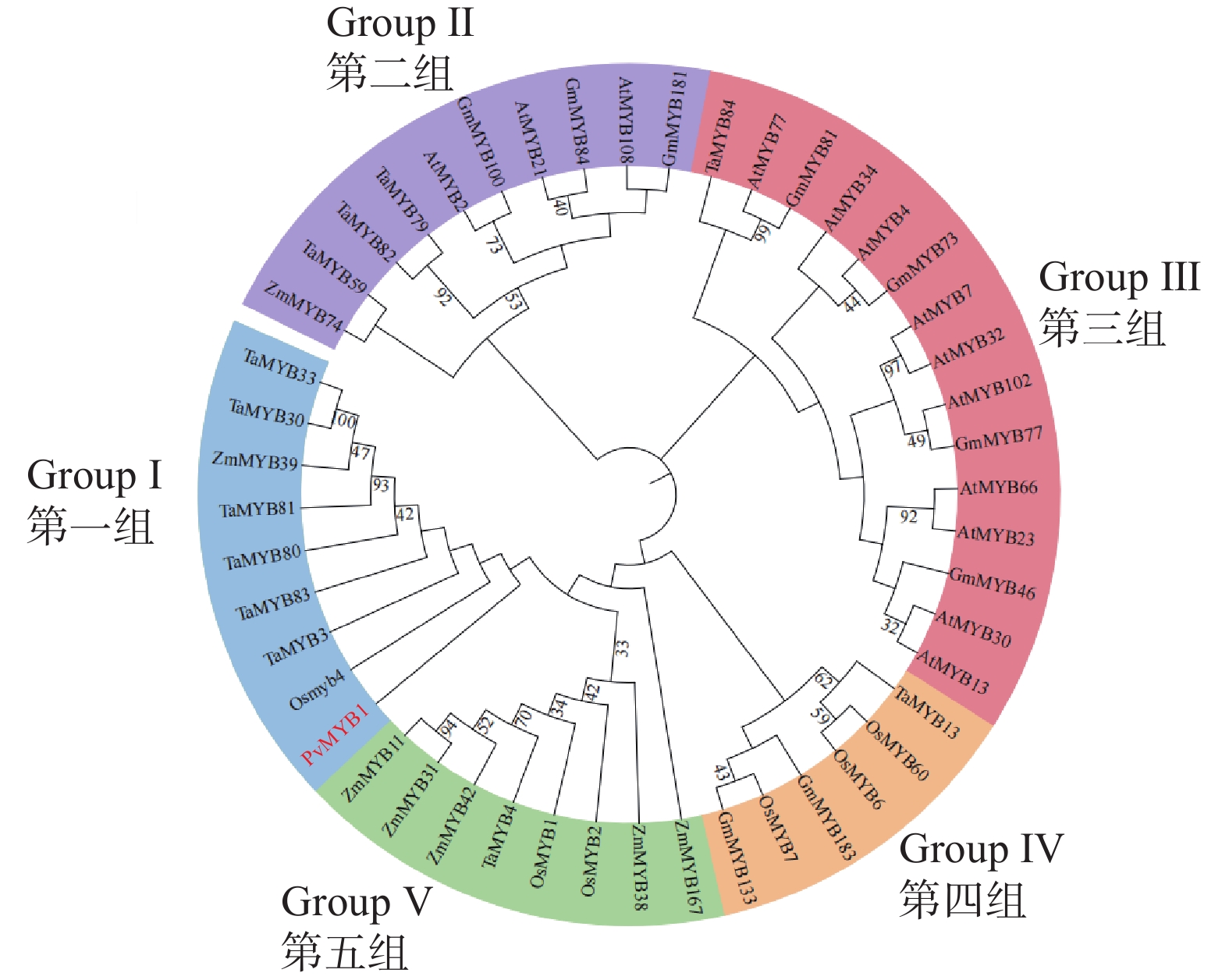
为分析PvMYB1蛋白与不同物种MYB蛋白的关系,本研究将PvMYB1与水稻、玉米、大豆(Glycine max)、小麦(Triticum aestivum)、拟南芥等MYB蛋白构建进化树。基于图5系统发育分析,多种植物MYB蛋白聚类为5组。PvMYB1与水稻OsMYB4同源性较高,被分在第一组中。这表明PvMYB1可能具有与OsMYB4相似的增加植物对干旱胁迫的生理生化适应性的功能。
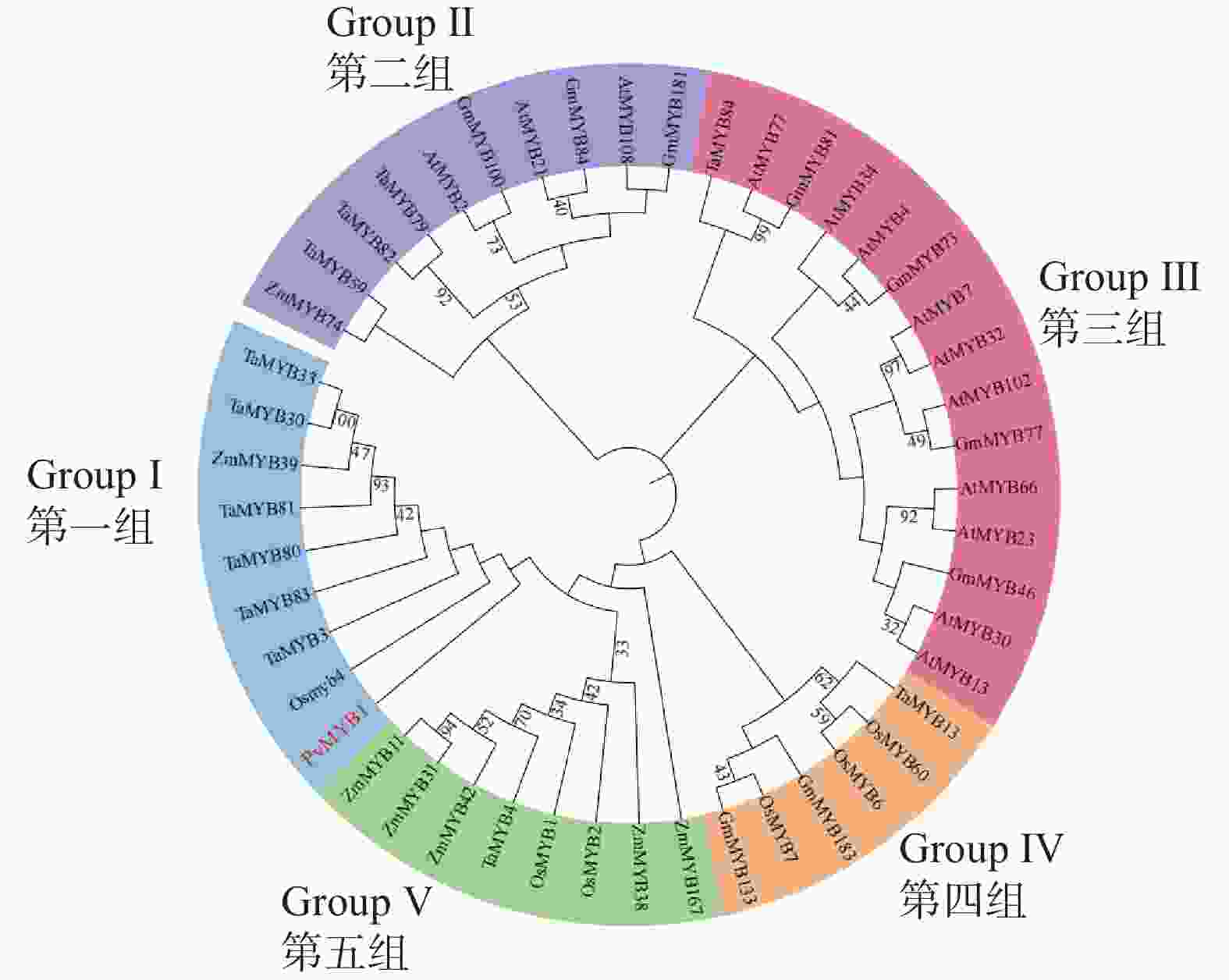
图 5 MYB 蛋白进化树分析
Figure 5. Phylogenetic analysis of MYB proteins
-
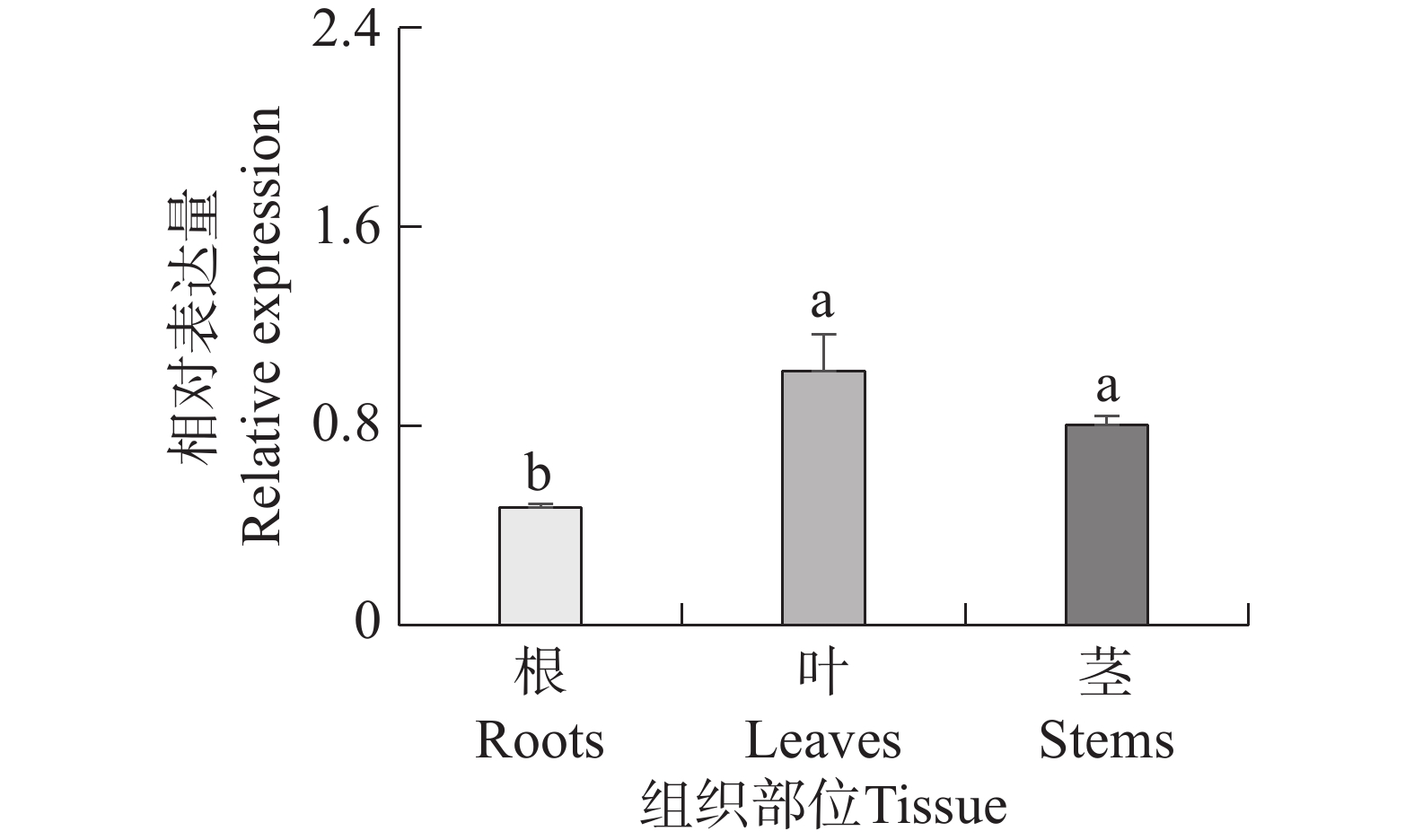
本研究通过定量PCR的方法分析了PvMYB1基因在海雀稗不同组织中的表达情况。如图6所示,PvMYB1在海雀稗根、茎和叶中均有所表达,且PvMYB1在叶和茎中的表达量显著高于在根中的表达量。
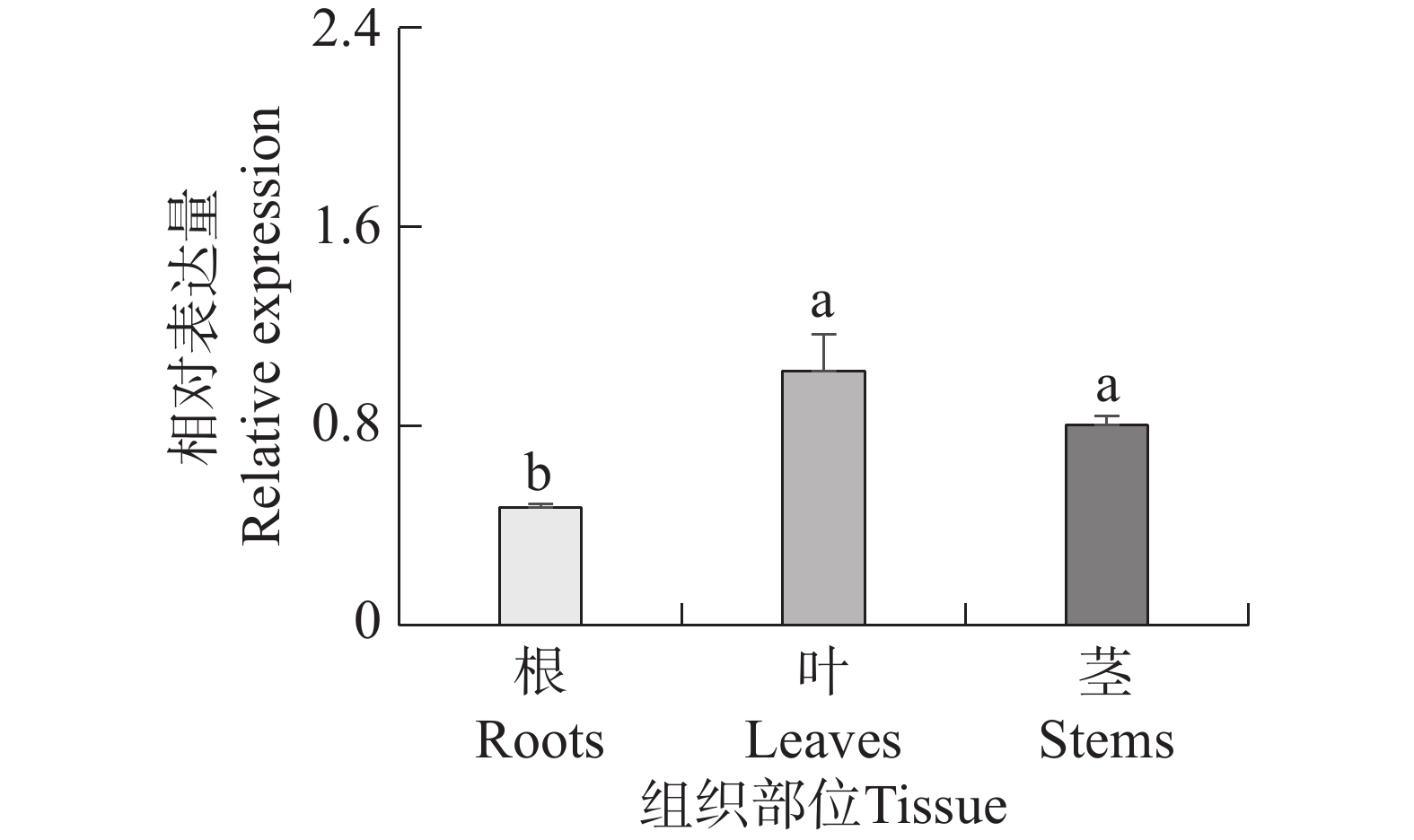
图 6 海雀稗 PvMYB1 基因组织表达分析
Figure 6. Expression analysis of PvMYB1 in different tissues
-
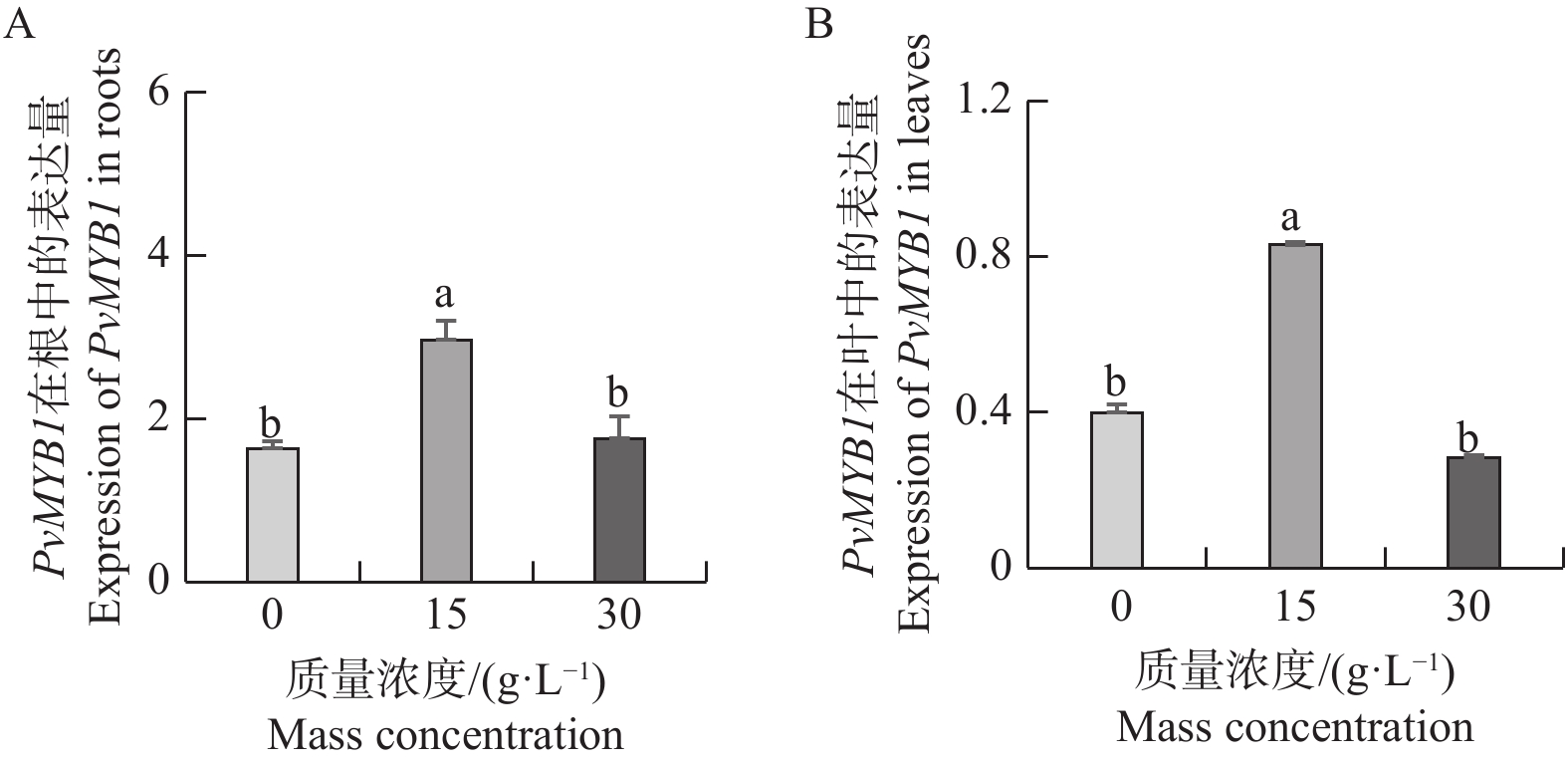
图7显示,PvMYB1基因在NaCl质量浓度升至15 g·L−1时,在根和叶中的表达量显著上调。然而,随着NaCl质量浓度进一步增至30 g·L−1,表达量又显著下降。15 g·L−1盐处理条件下PvMYB1基因在根和叶中的表达量与对照和30 g·L−1盐处理组之间差异达显著水平。PvMYB1对盐胁迫呈先升后降的表达模式,其可能在15 g·L−1 NaCl范围内发挥早期应答作用。
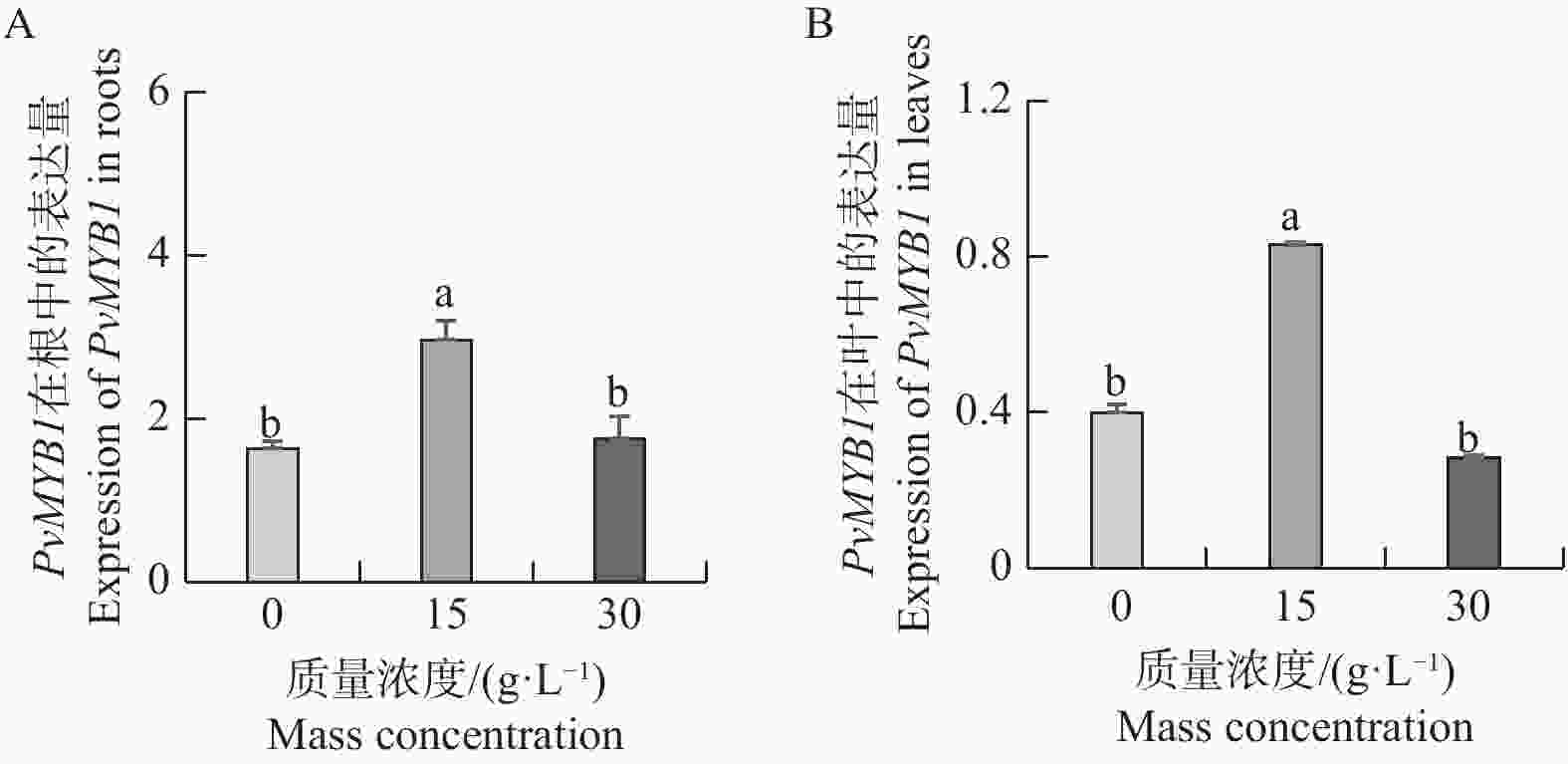
图 7 不同盐质量浓度处理对 PvMYB1 基因表达的影响
Figure 7. Expression analysis of PvMYB1 at different salt mass concentration treatments
-
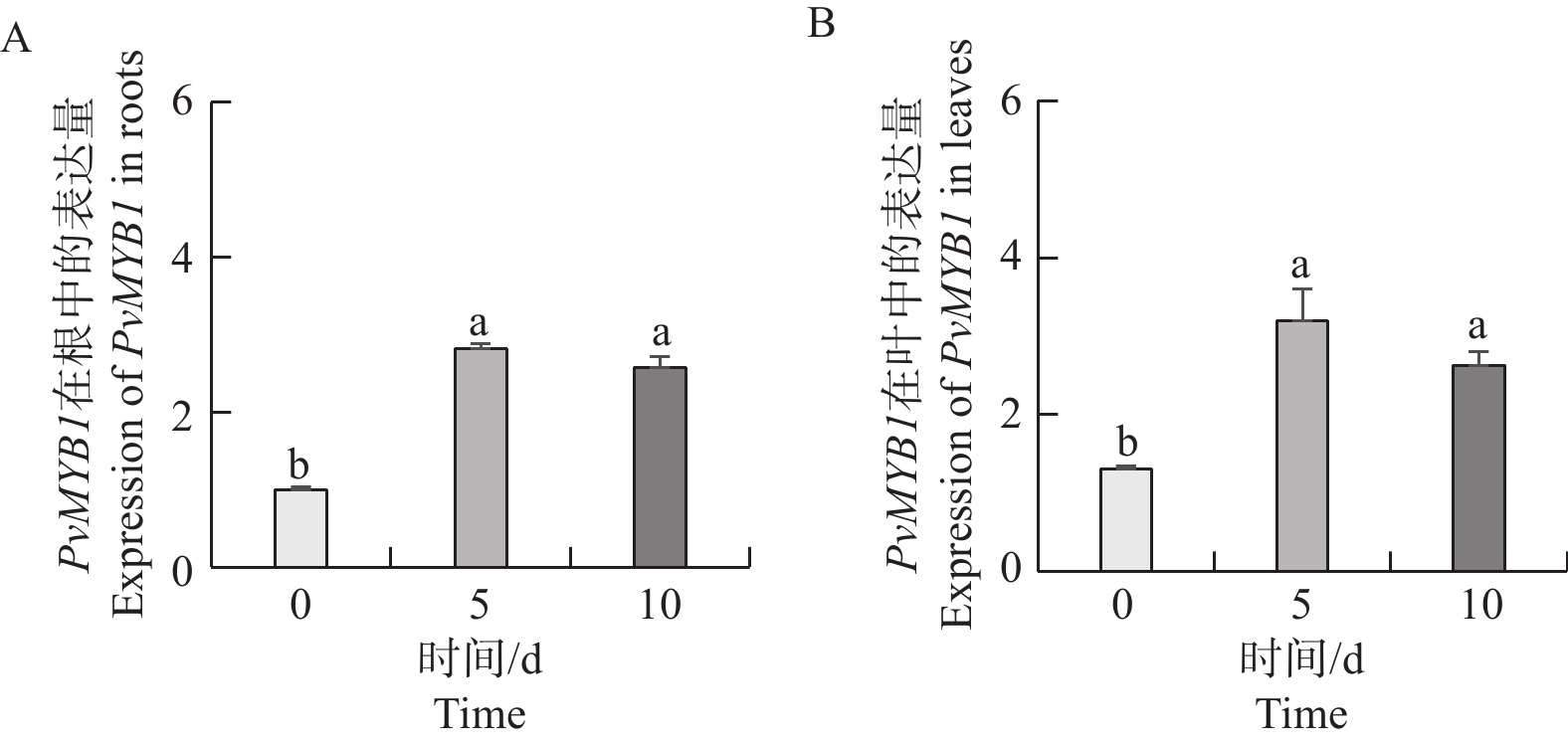
如图8所示,与0 d相比,NaCl处理5 d和10 d的PvMYB1基因表达量在海雀稗的根和叶中都显著上升。但是NaCl处理5 d的海雀稗的根和叶中的PvMYB1基因表达量与NaCl处理10 d的PvMYB1基因表达量无显著差异。
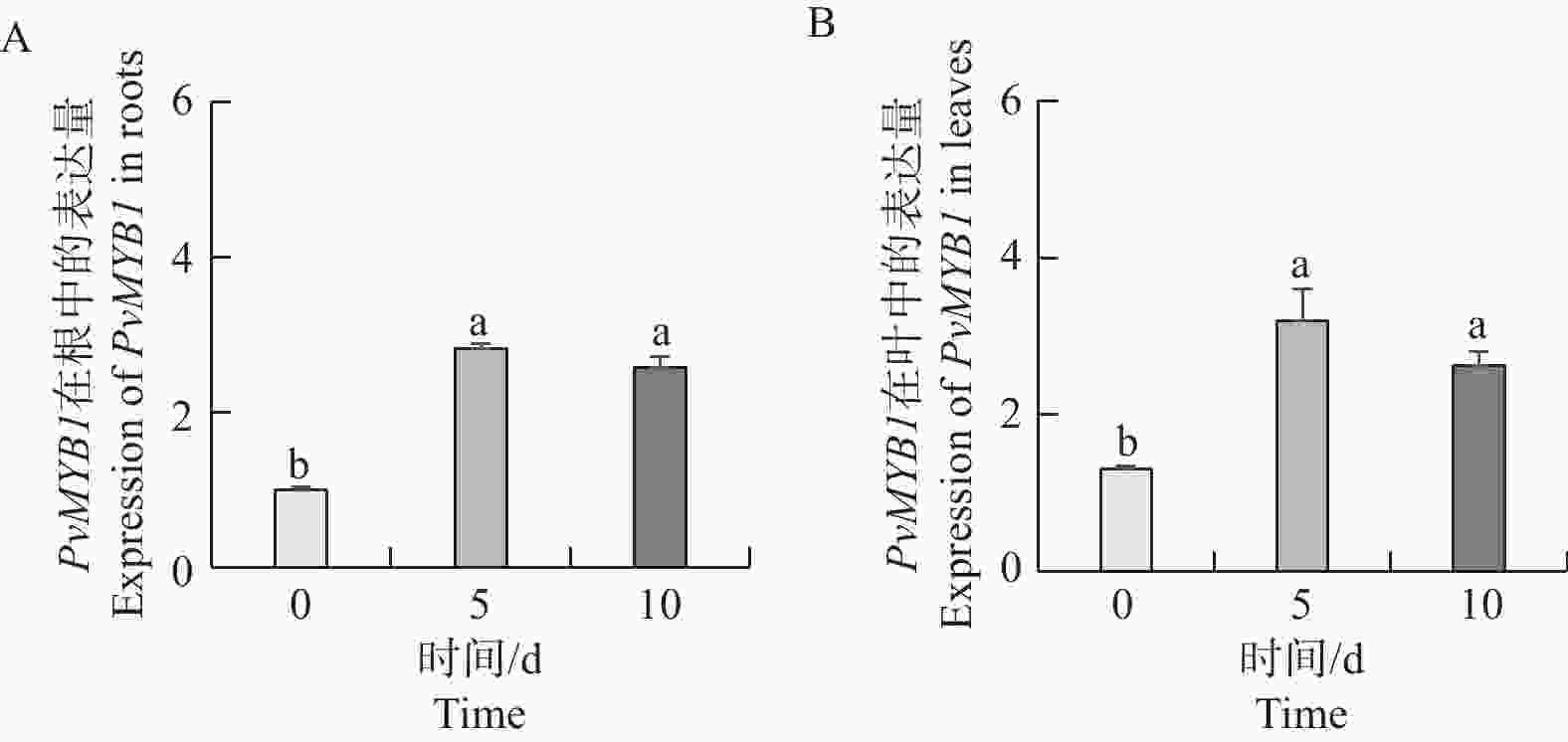
图 8 30 g·L−1 NaCl 不同时间梯度处理对 PvMYB1 基因表达的影响
Figure 8. Expression analysis of PvMYB1 at different time treatment in 30 g·L−1 NaCl
-
自然环境恶化、不良的灌溉方式和气候变化加剧了土壤盐碱化问题。高盐环境胁迫下,土壤中过量可溶性盐分对多数植物是有不利影响的[24]。种植耐盐植物可以提高盐碱地的利用率,但是由于耐盐性差,包括紫花苜蓿(Medicago sativa)在内的传统牧草植物的产量和质量受到盐胁迫的影响[25]。海雀稗作为优良的草坪草、牧草和盐碱地改良草种,在对海雀稗、杂交狗牙根(Cynodon dactylon × C. transvaalensis)、结缕草(Zoysia japonica)、假俭草(Eremochloa ophiuroides)等草坪草的耐盐性分析中发现,海雀稗在盐胁迫下表现出更好的生长性能,表现为盐处理下生长被抑制的程度较小以及较高的草坪质量[26]。同一草种中不同品种或品系的耐盐性有明显差异,因此对海雀稗种质资源进行耐盐性评价,从中筛选出了极端耐盐种‘USA17-18’[17]。本研究中以‘USA17-18’海雀稗为实验材料,为了更好地探究海雀稗耐盐机制,本研究克隆了海雀稗 PvMYB1基因,并对其进行生物信息学及基因表达分析。
MYB转录因子广泛分布于植物体内,在调控植物生长发育的各个阶段中发挥着作用[11]。MYB蛋白可以通过它们在N末端高度保守的DNA 结合结构域(DBD)来代表,该结构域也称为 MYB结构域[27]。它在功能上与 DNA 结合和蛋白质-蛋白质相互作用相关,由1到4个不完全重复序列(R)组成。根据不完全重复的氨基酸序列(R)的数量,MYB蛋白分为4R-MYB、R1R2R3-MYB(3R)、R2R3-MYB(2R)和1R-MYB4类亚家族[7]。其中,R2R3-MYB属于在植物中普遍存在的转录因子类型之一[28]。多数研究表明,R2R3-MYB亚家族的基因具有MYB-DNA-binding和2个SANT结构域[29],通过在线工具对PvMYB1蛋白保守结构域进行分析,结果表明,PvMYB1蛋白具有2个SANT结构域、MYB-DNA结合域和稳定的PLN03091结构域等,符合MYB基因家族保守结构域特点,是典型的R2R3型MYB转录因子。
系统进化分析显示,PvMYB1蛋白与水稻OsMYB4、小麦TaMYB33等已知参与非生物胁迫响应的MYB蛋白亲缘关系较近,同属一个进化支,提示PvMYB1可能具有类似的功能。在该组中,定位于细胞核内的TaMYB33促进了渗透压平衡和活性氧清除的能力,从而增强了转基因拟南芥的耐盐性和耐旱性[30]。与之类似,TaMYB80同样定位于细胞核上,在转基因拟南芥中过表达TaMYB80增加了植物对高温和干旱的耐受性[31]。MYB转录因子对靶基因的转录调控通常发生在细胞核内,定位于细胞核内的ZmMYB39通过与胁迫相关基因的表达的相互作用和促进其表达增强了玉米的耐旱性[32]。TaMYB30可以通过识别相关顺式元件直接与小麦蜡的生物合成相关的基因的启动子区域结合,并激活其表达,从而正向调控小麦蜡的生物合成[33]。与之类似,本研究中PvMYB1定位于细胞核中,PvMYB1可能参与了海雀稗响应非生物胁迫的不同生理过程。
基因表达分析表明,盐处理增强了PvMYB1基因在根和叶中在30 g·L−1盐处理5 d、10 d以及15 g·L−1盐处理14 d条件下的表达量。研究表明,盐处理能增强拟南芥[34],水稻[35],小麦[30],大豆[27]、苹果(Malus Domestica)[36]等植物MYB基因的表达。例如,过表达OsMYB6的植株表现出更高的耐旱性和耐盐性,且OsMYB6转基因植株中非生物胁迫响应基因的表达量与对照相比显著增加[35]。类似的,过表达TaMYB32的转基因拟南芥的耐盐性得到提高[37]。AtMYB30激活AtAOX1a表达以保持细胞氧化还原稳态,从而赋予植物耐盐性[38]。AtMYB49可以通过调节叶片角质层的形成和维持叶片水分来提高植物的耐盐性,此外还可以增加叶片中的钙离子水平,并在盐胁迫下维持生物膜的稳定性[39]。MYB转录因子可以介导多种途径来增强植物的耐盐性,因此推测PvMYB1可能也通过调控离子稳态、渗透平衡、氧化还原平衡参与海雀稗响应盐胁迫的过程。
-
本研究成功克隆了海雀稗PvMYB1基因,其编码一个R2R3-MYB型转录因子。表达分析表明,PvMYB1受盐胁迫显著诱导,尤其在中等盐质量浓度(15 g·L−1)和胁迫早期表达上调,提示该基因在海雀稗响应盐胁迫过程中可能发挥重要的早期调控作用。
Cloning and expression analysis of PvMYB1 in Paspalum vaginatum
-
摘要: 为探究MYB转录因子基因是否参与了盐胁迫下对海雀稗(Paspalum vaginatum Sw.)生长发育的调控,本研究分析了盐胁迫对海雀稗生长的影响,克隆了PvMYB1基因,通过生物信息学及基因表达2种分析方法展开研究。结果表明,盐胁迫抑制了海雀稗地上部和根系生长,具体表现为总根长、根表面积及根体积的降低。海雀稗PvMYB1基因全长为1 098 bp,编码365个氨基酸残基,蛋白分子质量为 39.26 kDa,属于R2R3-MYB家族,亚细胞定位预测 PvMYB1蛋白定位于细胞核中。实时荧光定量PCR结果显示,PvMYB1基因在叶中的表达量高于在根中的表达量;不同时段NaCl处理结果显示,质量浓度为30 g·L−1 的NaCl处理显著提升了PvMYB1基因的转录水平。此外,在不同质量浓度的NaCl处理的条件下,当NaCl质量浓度升至15 g·L−1时,根和叶中的PvMYB1基因的表达量显著上调。本研究结果揭示了PvMYB1可能参与了海雀稗对NaCl胁迫的应答。Abstract: Seashore paspalum (Paspalum vaginatum Sw.) were treated with different concentrations of salt to observe the effect of salt treatment on the growth of seashore paspalum, and the PvMYB1 gene was cloned from seashore paspalum under different treatments and analyzed by using bioinformatics and gene expression analysis methods in a view to investigating whether MYB transcription factor genes are involved in the regulation of growth and development of seashore paspalum under salt stress. The results showed that salt stress inhibited the growth of both shoots and roots in seashore paspalum, manifested by reduction in total root length, root surface area, and root volume. The full length of the seashore paspalum PvMYB1 gene was
1098 bp, encoding 365 amino acid residues, with a protein molecular weight of 39.26 kDa, and.this gene belongs to the R2R3-MYB family. Subcellular localization prediction indicated that the PvMYB1 protein was located in the nucleus. RT-qPCR analysis demonstrated that the expression of PvMYB1 was higher in leaves than in roots. The results of salt treatment at different time periods showed that 30 g·L-1 salt treatment significantly increased the transcription level of the PvMYB1 gene. Moreover, under different salt concentration treatments, the expression levels of the PvMYB1 gene in the roots and leaves were significantly upregulated when the salt concentration was 15 g·L-1. All these results revealed that PvMYB1 is implicated in the response of seashore paspalum to salt stress.-
Key words:
- Paspalum vaginatum /
- salt stress /
- gene expression /
- MYB
-
图 2 不同盐处理对海雀稗干质量的影响
Fig. 2 Dry weight of Paspalum vaginatum under different NaCl treatments
图 7 不同盐质量浓度处理对 PvMYB1 基因表达的影响
Fig. 7 Expression analysis of PvMYB1 at different salt mass concentration treatments
图 8 30 g·L−1 NaCl 不同时间梯度处理对 PvMYB1 基因表达的影响
Fig. 8 Expression analysis of PvMYB1 at different time treatment in 30 g·L−1 NaCl
表 1 PvMYB1克隆 和 qPCR 引物分析
Table 1 Primers for PvMYB1 cloning and qPCR analysis
引物名称
Primer引物序列(5′—3′)
SequencePvMYB1-ORF-F ATGGGGCGGATCCCGTGCTGC PvMYB1-ORF-R TTACACCATGTGATTTGTCAGGTCA PvMYB1-RT-F GGCGCCTCATCCCCAAGAAT PvMYB1-RT-R GCCGCGATCACCGACCAC PvMYB1a-RT-F CACTGTCGAGTGGGTTCGCTAC PvMYB1a-RT-R GCCGATGAATTTTACTTGTTGC  下载: 导出CSV
下载: 导出CSV
-
[1] Balfagón D, Zandalinas S I, Mittler R, et al. High temperatures modify plant responses to abiotic stress conditions [J]. Physiologia Plantarum, 2020, 170(3): 335−344. https://doi.org/10.1111/ppl.13151 doi: 10.1111/ppl.13151 [2] Zaman M, Shahid S A, Heng L. Guideline for salinity assessment, mitigation and adaptation using nuclear and related techniques [M]. Cham: Springer, 2018. https://doi.org/10.1007/978-3-319-96190-3 [3] Fao. Global map of salt-affected soils [R]. Rome: FAO, 2021: 1-20. [4] Liu G Y, Yu X, Zeng Y L, et al. SOS2-FREE1 regulates SOS1 tonoplast sorting to promote Na+ compartmentalization in vacuole duringsalt stress response [J]. Journal of Integrative Plant Biology, 2025, 67(10): 2545−2560. https://doi.org/10.1111/jipb.13970 doi: 10.1111/jipb.13970 [5] Zhou H P, Shi H F, Yang Y Q, et al. Insights into plant salt stress signaling and tolerance [J]. Journal of Genetics and Genomics, 2024, 51(1): 16−34. https://doi.org/10.1016/j.jgg.2023.08.007 doi: 10.1016/j.jgg.2023.08.007 [6] Acosta-Motos J, Ortuño M, Bernal-Vicente A, et al. Plant responses to salt stress: adaptive mechanisms [J]. Agronomy, 2017, 7(1): 18. https://doi.org/10.3390/agronomy7010018 doi: 10.3390/agronomy7010018 [7] Zhang D W, Zhou H P, Zhang Y, et al. Diverse roles of MYB transcription factors in plants [J]. Journal of Integrative Plant Biology, 2025, 67(3): 539−562. https://doi.org/10.1111/jipb.13869 doi: 10.1111/jipb.13869 [8] Li J L, Han G L, Sun C F, et al. Research advances of MYB transcription factors in plant stress resistance and breeding [J]. Plant Signaling & Behavior, 2019, 14(8): 1613131. https://doi.org/10.1080/15592324.2019.1613131 doi: 10.1080/15592324.2019.1613131 [9] Paz-Ares J, Ghosal D, Wienand U, et al. The regulatory c1 locus of Zea mays encodes a protein with homology to myb proto-oncogene products and with structural similarities to transcriptional activators [J]. The EMBO Journal, 1987, 6(12): 3553−3558. https://doi.org/10.1002/j.1460-2075.1987.tb02684 doi: 10.1002/j.1460-2075.1987.tb02684 [10] 李志强, 罗正乾, 徐琳黎, 等. 基于T2T基因组鉴定大豆R2R3-MYB基因家族及干旱和盐胁迫下的表达分析[J]. 生物技术通报, 2025, 41(5): 141−152. https://doi.org/10.13560/j.cnki.biotech.bull.1985.2024-0745 doi: 10.13560/j.cnki.biotech.bull.1985.2024-0745 [11] Dubos C, Stracke R, Grotewold E, et al. MYB transcription factors in Arabidopsis [J]. Trends in Plant Science, 2010, 15(10): 573−581. https://doi.org/10.1016/j.tplants.2010.06.005 doi: 10.1016/j.tplants.2010.06.005 [12] Li J Y, Liu S Q, Chen P F, et al. Systematic analysis of the R2R3-MYB family in Camellia sinensis: evidence for galloylated catechins biosynthesis regulation [J]. Frontiers in Plant Science, 2022, 12: 782220. https://doi.org/10.3389/fpls.2021.782220 doi: 10.3389/fpls.2021.782220 [13] Katiyar A, Smita S, Lenka S K, et al. Genome-wide classification and expression analysis of MYB transcription factor families in rice and Arabidopsis [J]. BMC Genomics, 2012, 13(1): 544. https://doi.org/10.1186/1471-2164-13-544 doi: 10.1186/1471-2164-13-544 [14] YOO J H, PARK C Y, KIM J C, et al. Direct interaction of a divergent CaM isoform and the transcription factor, MYB2, enhances salt tolerance in Arabidopsis [J]. Journal of Biological Chemistry, 2005, 280(5): 3697−3706. https://doi.org/10.1074/jbc.m408237200 doi: 10.1074/jbc.m408237200 [15] Yang A, Dai XY, Zhang W H. A R2R3-type MYB gene, OsMYB2, is involved in salt, cold, and dehydration tolerance in rice [J]. Journal of Experimental Botany, 2012, 63(7): 2541−2556. https://doi.org/10.1093/jxb/err431 doi: 10.1093/jxb/err431 [16] Karimi I Y M, Kurup S S, Ali Salem M A M, et al. Evaluation of Bermuda and Paspalum grass types for urban landscapes under saline water irrigation [J]. Journal of Plant Nutrition, 2018, 41(7): 888−902. https://doi.org/10.1080/01904167.2018.1431669 doi: 10.1080/01904167.2018.1431669 [17] 申晴. 海雀稗种质资源遗传多样性与优异耐盐种质筛选[D]. 海口: 海南大学, 2019. https://doi.org/10.27073/d.cnki.ghadu.2019.001140 [18] 陈静波, 阎君, 姜燕琴, 等. 暖季型草坪草优良选系和品种抗盐性的初步评价[J]. 草业学报, 2009, 18(5): 107−114. https://doi.org/10.3321/j.issn:1004-5759.2009.05.016 doi: 10.3321/j.issn:1004-5759.2009.05.016 [19] 申晴, 韦海燕, 卞华, 等. 海雀稗种质资源的耐盐性评价[J]. 热带生物学报, 2020, 11(1): 11−19. https://doi.org/10.15886/j.cnki.rdswxb.2020.01.003 doi: 10.15886/j.cnki.rdswxb.2020.01.003 [20] 黄杰, 宋剑灵, 安娜, 等. 柱花草磷饥饿响应基因SgPHR1和SgPHR2的克隆与表达分析[J]. 热带作物学报, 2021, 42(8): 2158−2166. https://doi.org/10.3969/j.issn.1000-2561.2021.08.006 doi: 10.3969/j.issn.1000-2561.2021.08.006 [21] 邹晓燕, 王林杰, 黄杰, 等. 柱花草SgNramp1和SgNramp2基因克隆与表达分析[J]. 草地学报, 2022, 30(6): 1388−1395. https://doi.org/10.11733/j.issn.1007-0435.2022.06.009 doi: 10.11733/j.issn.1007-0435.2022.06.009 [22] 刘莉婷, 高佳淼, 周鸿明, 等. 地毯草扩张蛋白基因AcEXPA1的克隆与表达分析[J]. 草地学报, 2025, 33(2): 351−359. https://doi.org/10.11733/j.issn.1007-0435.2025.02.003 doi: 10.11733/j.issn.1007-0435.2025.02.003 [23] 张郎织, 李季肤, 黄杰, 等. 地毯草AcMATE1基因的克隆与表达分析[J]. 热带作物学报, 2021, 42(7): 1860−1867. https://doi.org/10.3969/j.issn.1000-2561.2021.07.007 doi: 10.3969/j.issn.1000-2561.2021.07.007 [24] Park H J, Kim W Y, Yun D J. A new insight of salt stress signaling in plant [J]. Molecules and Cells, 2016, 39(6): 447−459. https://doi.org/10.14348/molcells.2016.0083 doi: 10.14348/molcells.2016.0083 [25] Campanelli A, Ruta C, Morone-Fortunato I, et al. Alfalfa (Medicago sativa L.) clones tolerant to salt stress: in vitro selection [J]. Open Life Sciences, 2013, 8(8): 765−776. https://doi.org/10.2478/s11535-013-0194-1 doi: 10.2478/s11535-013-0194-1 [26] Marcum K B, Murdoch C L. Salinity tolerance mechanisms of Six C4 turfgrasses [J]. Journal of the American Society for Horticultural Science, 1994, 119(4): 779−784. https://doi.org/10.21273/jashs.119.4.779 doi: 10.21273/jashs.119.4.779 [27] Liao Y, Zou H F, Wang H W, et al. Soybean GmMYB76, GmMYB92, and GmMYB177 genes confer stress tolerance in transgenic Arabidopsis plants [J]. Cell Research, 2008, 18(10): 1047−1060. https://doi.org/10.1038/cr.2008.280 doi: 10.1038/cr.2008.280 [28] 陈娜, 迟晓元, 潘丽娟, 等. MYB转录因子在植物盐胁迫调控中的研究进展[J]. 植物生理学报, 2015, 51(9): 1395−1399. https://doi.org/10.13592/j.cnki.ppj.2015.0239 doi: 10.13592/j.cnki.ppj.2015.0239 [29] 王文林, 陈海生, 郑树芳, 等. 澳洲坚果MiMYB2基因克隆及结构与功能分析[J]. 植物研究, 2020, 40(6): 913−922. https://doi.org/10.7525/j.issn.1673-5102.2020.06.014 doi: 10.7525/j.issn.1673-5102.2020.06.014 [30] Sairam R, Tyagi A. Physiology and molecular biology of salinity stress tolerance in plants [J]. Current Science, 2004, 86(3): 407−421. [31] Zhao Y, Tian X J, Wang F, et al. Characterization of wheat MYB genes responsive to high temperatures [J]. BMC Plant Biology, 2017, 17(1): 208. https://doi.org/10.1186/s12870-017-1158-4 doi: 10.1186/s12870-017-1158-4 [32] Ren Z Z, Zhang P Y, Su H H, et al. Regulatory mechanisms used by ZmMYB39 to enhance drought tolerance in maize (Zea mays) seedlings [J]. Plant Physiology and Biochemistry, 2024, 211: 108696. https://doi.org/10.1016/j.plaphy.2024.108696 doi: 10.1016/j.plaphy.2024.108696 [33] Liu L, Li H Y, Wang X Y, et al. Transcription factor TaMYB30 activates wheat wax biosynthesis [J]. International Journal of Molecular Sciences, 2023, 24(12): 10235. https://doi.org/10.3390/ijms241210235 doi: 10.3390/ijms241210235 [34] Li W H, Zhong J L, Zhang L H, et al. Overexpression of a Fragaria vesca MYB transcription factor gene (FvMYB82) increases salt and cold tolerance in Arabidopsis thaliana [J]. International Journal of Molecular Sciences, 2022, 23(18): 10538. https://doi.org/10.3390/ijms231810538 doi: 10.3390/ijms231810538 [35] Tang Y H, Bao X X, Zhi Y L, et al. Overexpression of a MYB family gene, OsMYB6, increases drought and salinity stress tolerance in transgenic rice [J]. Frontiers in Plant Science, 2019, 10: 168. https://doi.org/10.3389/fpls.2019.00168 doi: 10.3389/fpls.2019.00168 [36] Du B Y, Liu H, Dong K T, et al. Over-expression of an R2R3 MYB gene, MdMYB108L, enhances tolerance to salt stress in transgenic plants [J]. International Journal of Molecular Sciences, 2022, 23(16): 9428. https://doi.org/10.18488/journal.105 doi: 10.18488/journal.105 [37] Zhang L C, Zhao G Y, Jia J Z, et al. Molecular characterization of 60 isolated wheat MYB genes and analysis of their expression during abiotic stress [J]. Journal of Experimental Botany, 2012, 63(1): 203−214. https://doi.org/10.1093/jxb/err264 doi: 10.1093/jxb/err264 [38] Gong Q Y, Li S, Zheng Y, et al. SUMOylation of MYB30 enhances salt tolerance by elevating alternative respiration via transcriptionally upregulating AOX1a in Arabidopsis [J]. The Plant Journal, 2020, 102(6): 1157−1171. https://doi.org/10.1111/tpj.14689 doi: 10.1111/tpj.14689 [39] Zhang L Y, Jiang X C, Liu Q Y, et al. The HY5 and MYB15 transcription factors positively regulate cold tolerance in tomato via the CBF pathway [J]. Plant, Cell & Environment, 2020, 43(11): 2712−2726. https://doi.org/10.1111/pce.13868 doi: 10.1111/pce.13868 -

 点击查看大图
点击查看大图
计量
- 文章访问数: 668
- HTML全文浏览量: 178
- PDF下载量: 68
- 被引次数: 0
