

-
海洋作为地球生态系统中最大的碳资源库,对CO2的循环与固定起着极其重要的作用[1]。浮游植物是海洋中最主要的初级生产者,不仅在海洋碳循环中发挥着极其重要作用,而且对海洋中的氮、硫循环也具有着重要作用[2-4]。然而,由于各种污水、废水的排放,导致近岸海域生态环境极易发生改变,造成藻类大量繁殖,暴发近岸海水赤潮,最终对海洋生态环境安全构成严重威胁,同时严重威胁着海水养殖、人类健康和生态安全[5]。浮游植物的群落结构和时空变化可间接反映浮游植物所处生态环境状态的变化[6], 因此,通过监测浮游植物群落结构的变化,可及时了解周围生态环境和水质健康的状况。
近年来,随着三亚市旅游业、加工业及港口航运等的迅猛发展,其近岸海洋生态环境受到空前压力[7],环境质量受到了负面影响[8]。三亚湾紧挨三亚市区,是当地居民和游客重要的休闲活动场所,近年来,其海滩不断被侵蚀,近岸植被被破坏,这已影响到三亚湾的娱乐休闲功能[9],并对三亚湾生态环境构成严重威胁。贾磊等[10]对三亚湾表层沉积物重金属分布特征进行研究,发现该区域已受到Cd污染。然而,目前对三亚湾浮游植物群落特征和水质的调查研究报道并不多。本研究拟通过对海南三亚湾近岸海域网采浮游植物和水质的采样调查,探究浮游植物的群落结构、优势种种类、多样性特征和对水质进行测定,分析浮游植物与水环境因子之间的响应关系,评估水体环境的污染状况,旨在了解三亚湾生态环境的变化,便于当地相关部门及早采取风险管控措施,减少环境恶化所带来的危害和为赤潮预警与海洋生态环境保护提供科学依据。
-
通过参考《海洋调查规范》[11]和结合三亚湾实际的地理特征和水文环境,在海南岛三亚湾附近海域(109°23′07.64″~109°29′15.72″E,18°15′03.03″~18°17′11.49″N)设置了4个调查站位,分别于2020年4月和2021年4月进行调查,调查站位地理坐标见表1。
表 1 三亚湾调查站位地理坐标
站位 经度 纬度 SYW1-1 109°29′15.70″ 18°15′03.03″ SYW2-1 109°27′46.06″ 18°16′25.97″ SYW3-1 109°25′18.58″ 18°17′09.23″ SYW4-1 109°23′09.19″ 18°17′07.35″ -
根据《海洋调查规范》[12]中的调查规定对三亚湾浮游植物样本进行采样与处理。利用浅水Ⅲ型浮游生物网采样,采集的样品被甲醛溶液固定,最终浓度为4%。样品静置、浓缩后,取0.1 mL均匀样品置于计数框中,在生物显微镜(LEICADM6000B)下进行计数和鉴定。
-
用1.5 L采水器采集混合水样,装入1 L样品瓶中,水质物理指标(pH、电导率、溶解氧、盐度、水温)采用KEDIDDA便携式相关指标水质检测仪现场测定,TN、TP、NH4+-N等在水样采集后带回实验室,利用哈希DR3900多参数水质检测仪来分析,具体操作按照哈希水质仪配套的相关检测试剂使用说明进行。
-
采用Shannon-Wiener多样性指数(H ′)、Pielou 均匀度指数(J)、优势度(Y)和丰富度指数(d)对浮游植物群落结构特征进行分析评价[13-14]。
$$ P_{i}=n_i/N \;, $$ (1) 式中, ni为第i种的个体数量;N为全部物种的个体数。
$$ {H}^{{'}}=-{\sum }_{i=1}^{s}{P}_{i}\mathrm{l}\mathrm{o}\mathrm{g}_2{P}_{i} \;, $$ (2) $$ J=H ′/{\rm{log}}_{2}S\;, $$ (3) 式中,S表示样品中浮游植物的总种类数;H′为多样性指数。
$$d=(S-1)/{\rm{log}}_{2}N \;, $$ (4) 式中,N为全部物种的个体数;S表示样品中浮游植物的总种类数。
$$ Y= P_{i}×f_{i} \;, $$ (5) 式中,Pi=ni/N;fi为第i种的出现频率。
-
相关数据经Excel初步处理,统计学差异性分析使用DPS 18.10软件,Kruskal-Wallis检验采用SPSS 25.0软件,采用origin.2022和ArcMap 10.8作图,分别将浮游植物的2020年和2021年优势度排名前10的物种数据和环境数据经过 lg(X+1)转换,通过R 4.2.1去趋势对应分析(DCA),结果显示2020年和2021年4个排序轴中的最大值分别为0.404 29和0.980 64,均小于3,故在软件Canoco 5.0中选择线性模型(RDA)对三亚湾浮游植物与环境因子进行排序分析。
-
在2020年和2021年前后2个航次采集到的样品中,分别鉴定出浮游植物4门98种和3门65种(图1),其中,均以硅藻门种类占多数,分别为78种和62种,占浮游植物总种类数的79.60%和96.92%。甲藻门和蓝藻门种类2021年的比2020年的减少,2020年和2021年的甲藻门分别为14种和1种,2020年和2021年的蓝藻门分别为5种和2种。2020年还鉴定出一种隐藻,占总种类数的1%(图1)。
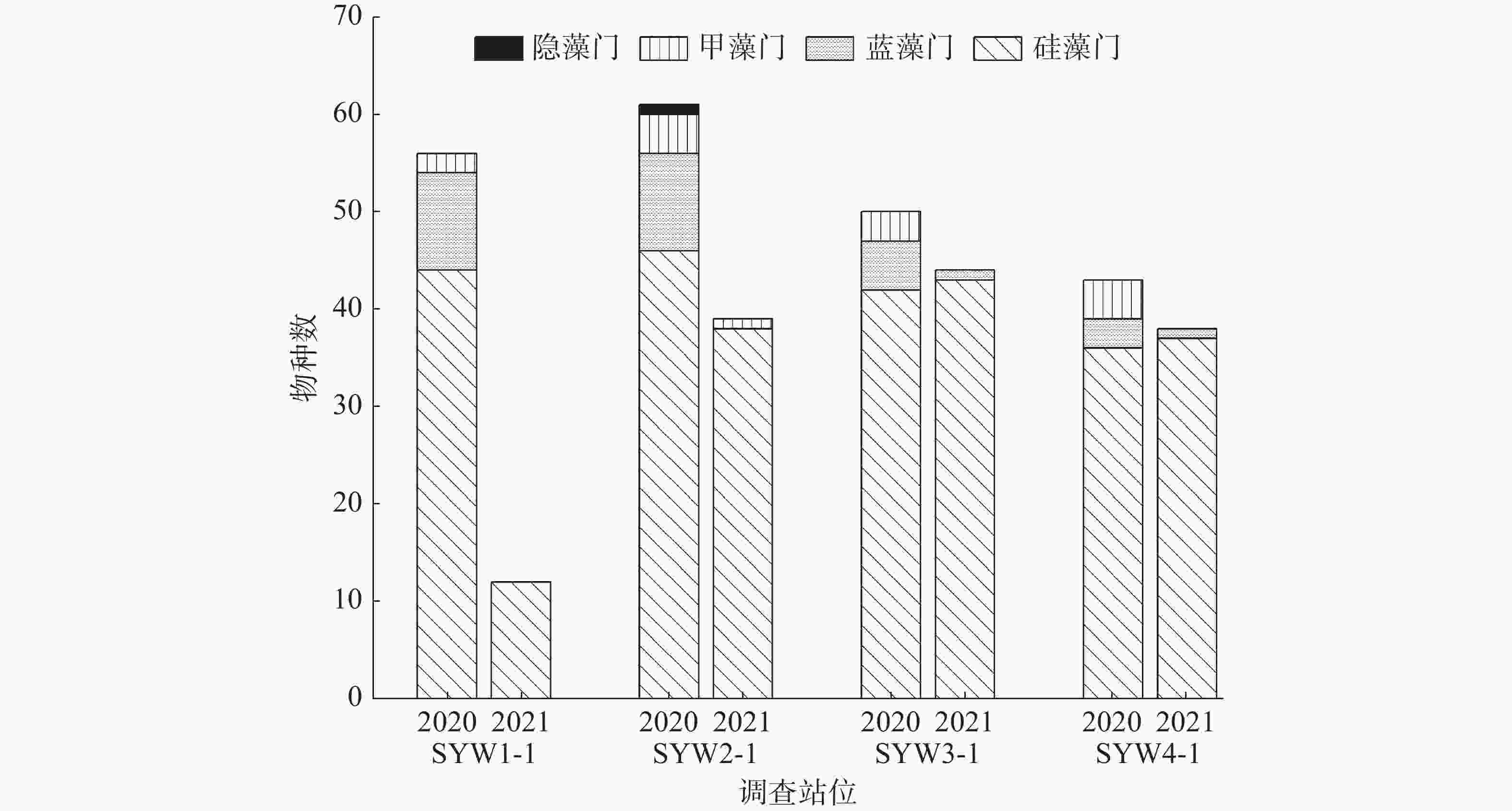
图 1 三亚湾各调查站位浮游植物物种数
-
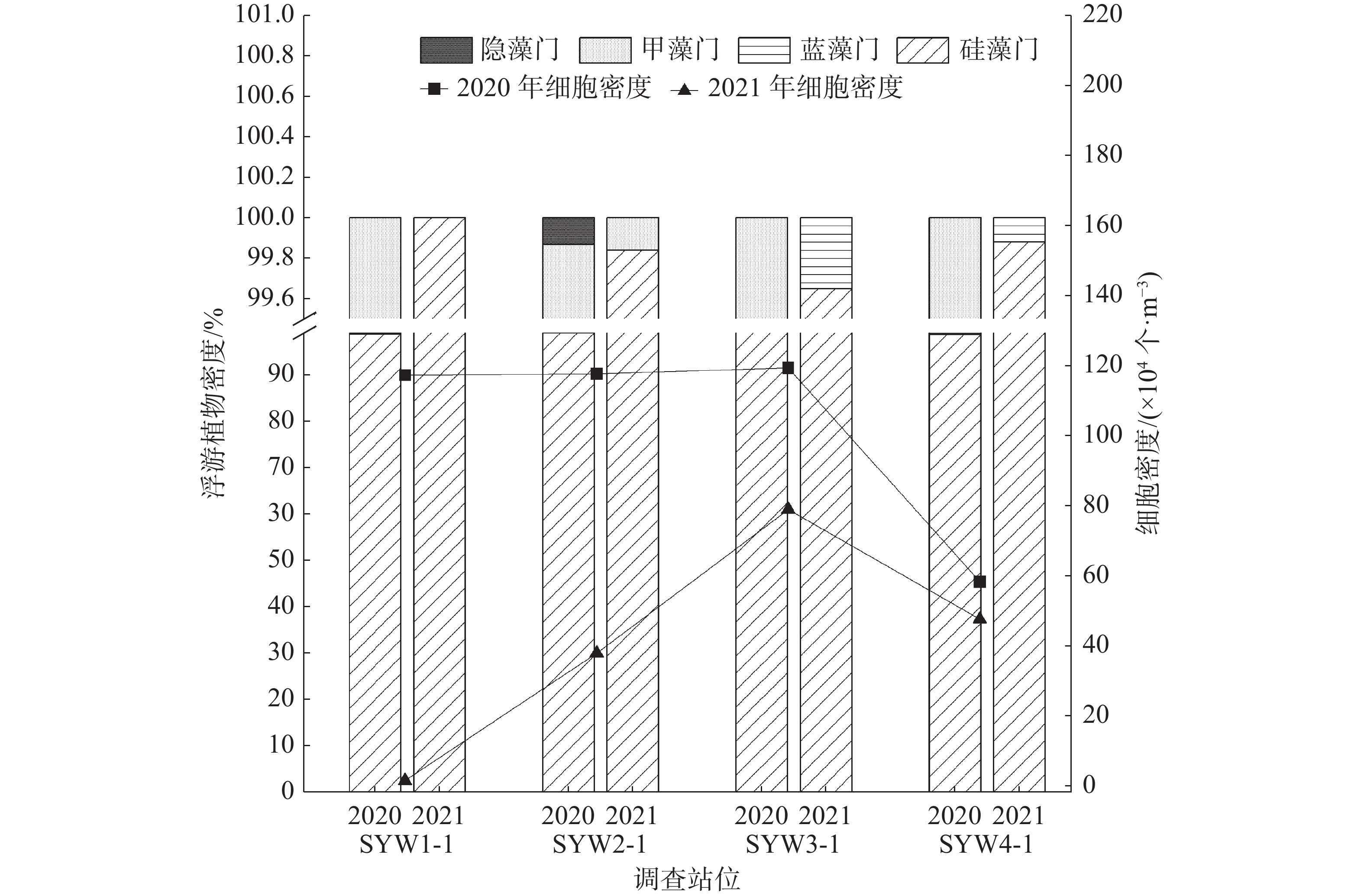
调查区域内,2020年浮游植物细胞密度明显高于2021年,2020年浮游植物细胞密度介于58.21×104~119.32×104个·m−3,平均值为103.1×104个·m−3,2021年浮游植物的细胞密度介于1.47×104~78.78×104个·m−3,平均值为41.42×104个·m−3,比2020年降低了59.83%。物种组成方面,2020年和2021年浮游植物均以硅藻类占绝大多数,分布最为广泛,2020年硅藻门浮游植物细胞密度平均占比为98.85%,2021年平均占比为99.84%,为2个航次的调查中的优势类群。除此以外,2021年的硅藻门、甲藻门、蓝藻门的种类数均低于2020年(图2)。

图 2 各门浮游植物空间分布
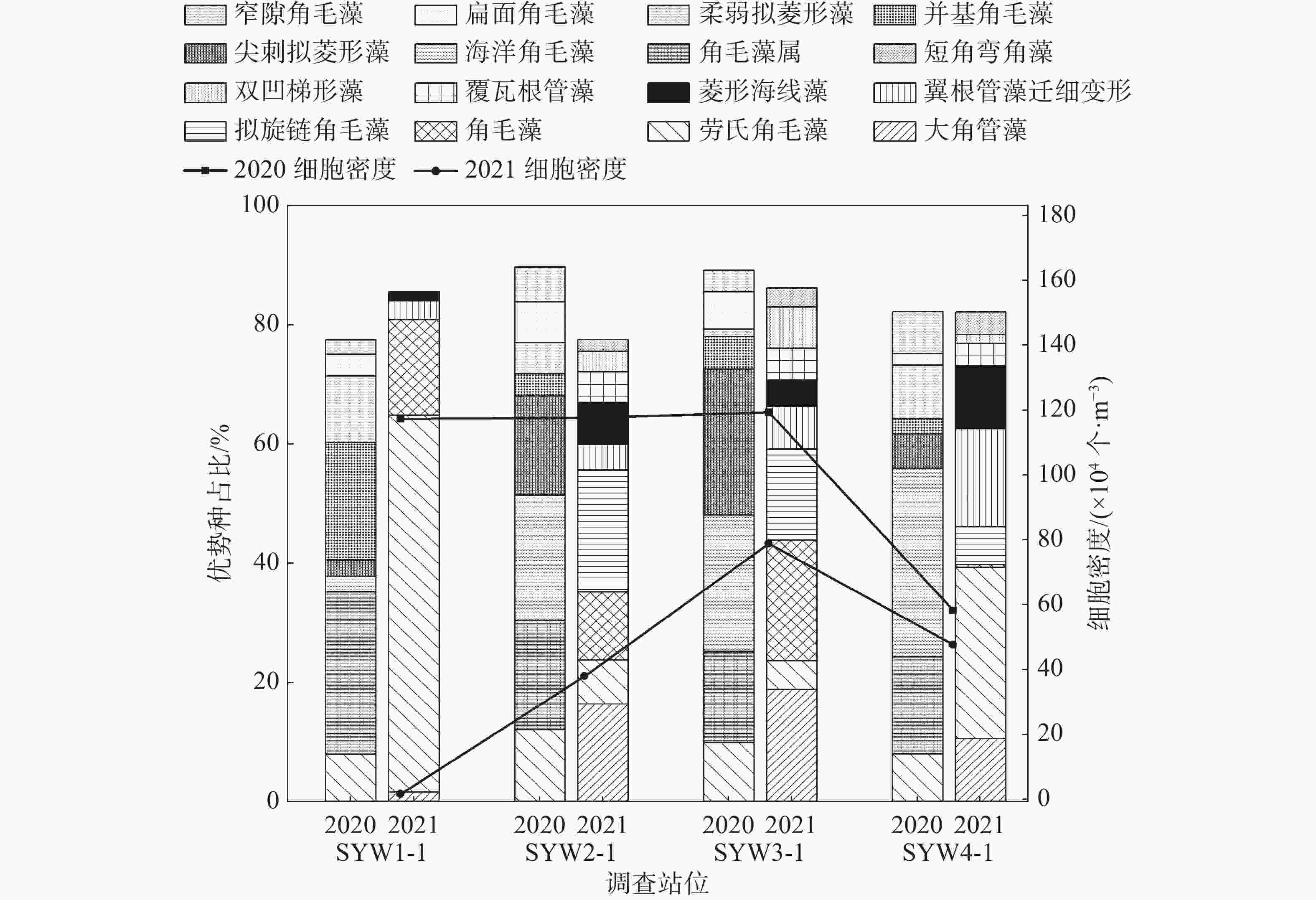
图 3 优势种空间分布
-
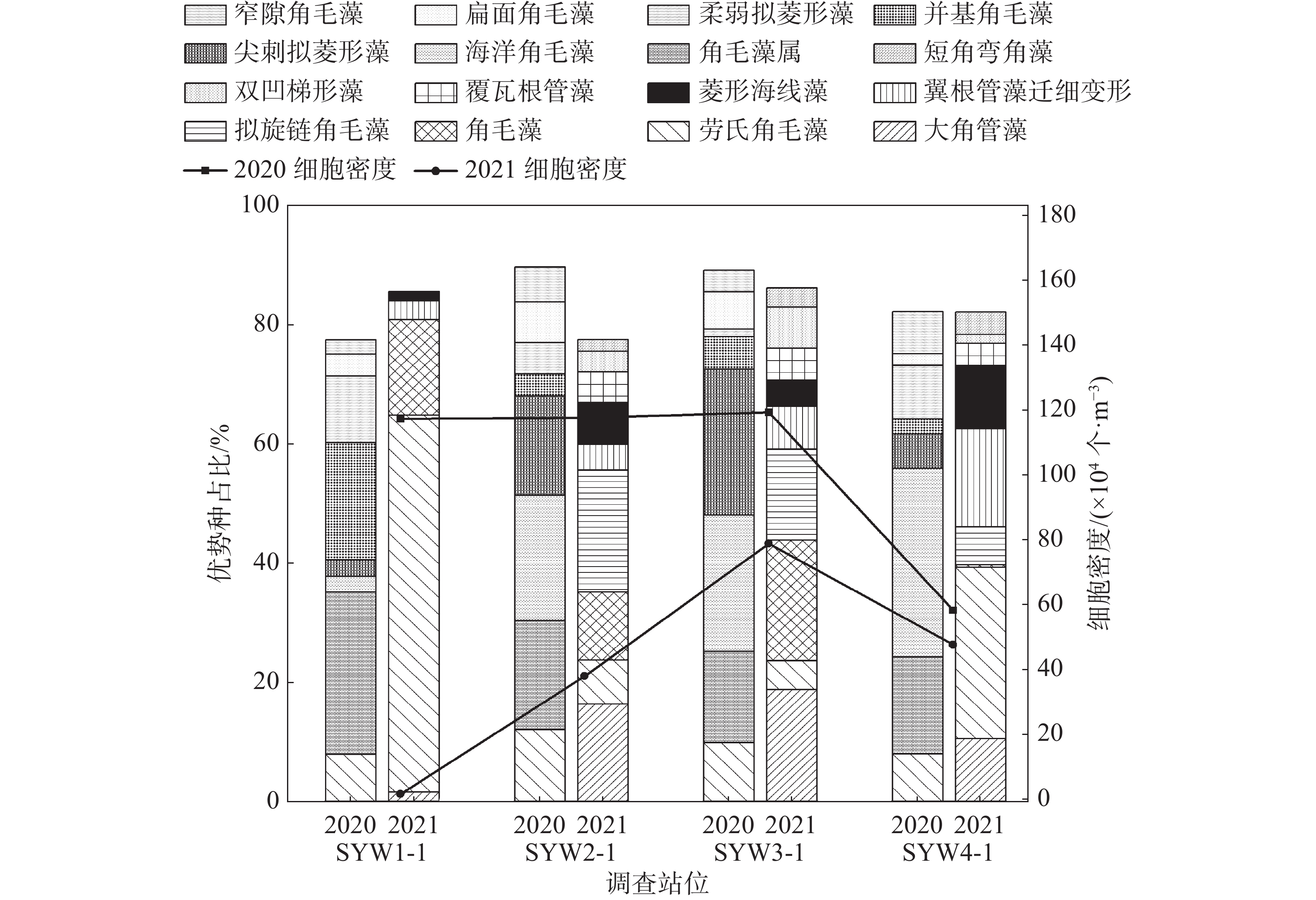
将浮游植物优势度≥0.02的种类作为该海域的优势种类。2020年和2021年浮游植物优势种分别为8种和9种,按优势度大小依次排列,2020年:角毛藻属>海洋角毛藻>劳氏角毛藻>尖刺拟菱形藻>并基角毛藻>柔弱拟菱形藻>扁面角毛藻>窄隙角毛藻,2021年:大角管藻>劳氏角毛藻>角毛藻>拟旋链角毛藻>翼根管藻纤细变型>菱形海线藻>覆瓦根管藻>双凹梯形藻>短角弯角藻。角毛藻属为第一优势种,优势度为0.221,平均密度为15.74×104个·m−3,在SYW1-1号站位密度最大。大角管藻为2021年第一优势种,优势度为0.157,平均细胞密度为6.51×104个·m−3,在SYW3-1号站位密度最大。具体详见表2和图3。
表 2 浮游植物优势种
时间 优势种 平均密度/
(104个·m−3)占总密
度比例/%出现
频率/%优势
度2020年4月 角毛藻属 15.74 22.06 100 0.221 海洋角毛藻 14.65 20.53 91.67 0.188 劳氏角毛藻 9.21 12.91 100.00 0.129 尖刺拟菱形藻 8.16 11.44 100.00 0.114 并基角毛藻 4.02 5.64 100.00 0.056 柔弱拟菱形藻 3.40 4.76 100.00 0.048 扁面角毛藻 3.31 4.64 100.00 0.046 窄隙角毛藻 2.58 3.62 91.67 0.033 2021年5月 大角管藻 6.51 15.72 100.00 0.157 劳氏角毛藻 5.31 12.82 100.00 0.128 角毛藻 5.15 12.44 100.00 0.124 拟旋链角毛藻 5.71 13.77 75.00 0.103 翼根管藻
纤细变型3.80 9.17 100.00 0.092 菱形海线藻 2.79 6.72 100.00 0.067 覆瓦根管藻 1.98 4.78 75.00 0.036 双凹梯形藻 1.86 4.49 75.00 0.034 短角弯角藻 1.27 3.08 75.00 0.023 -
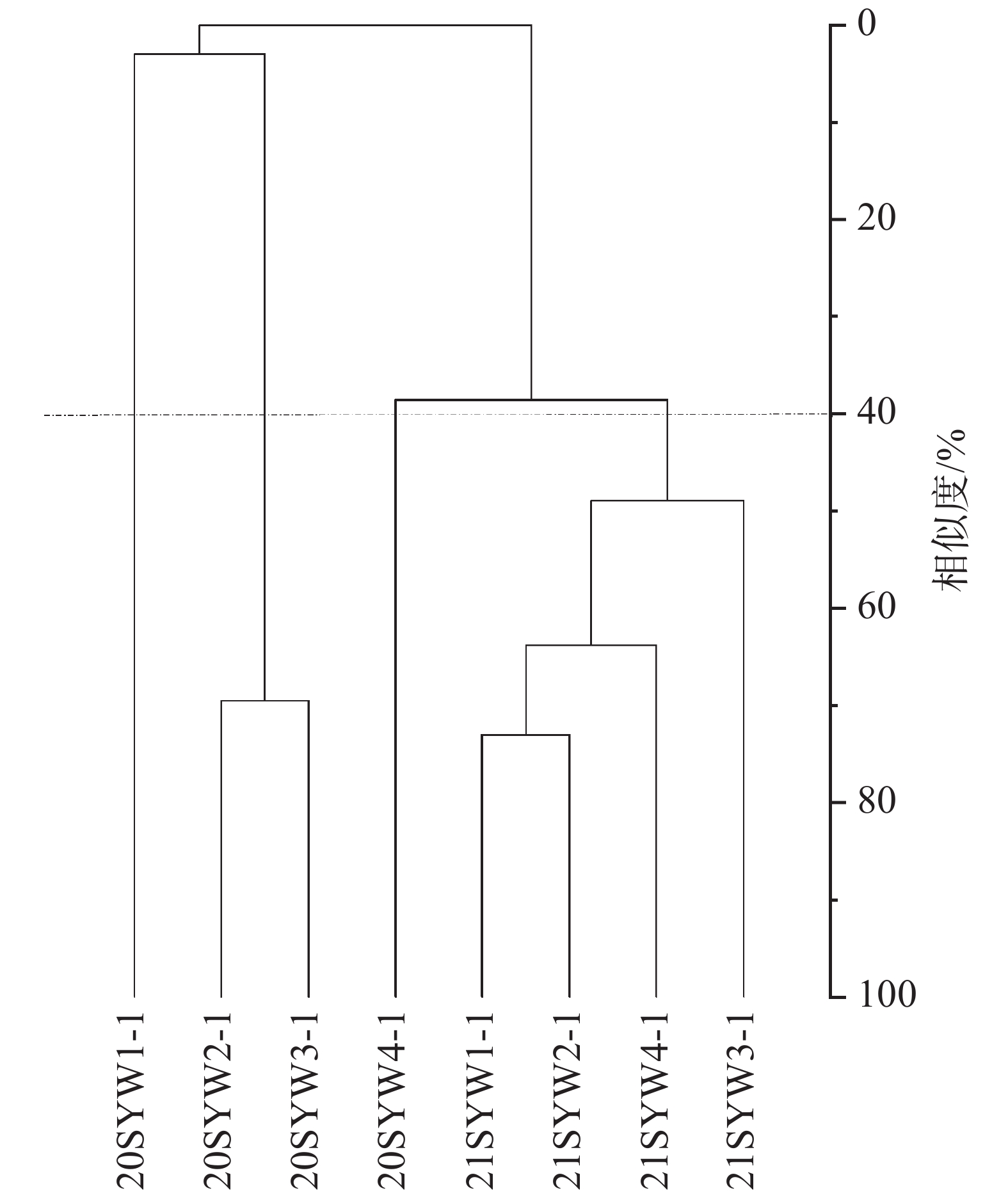
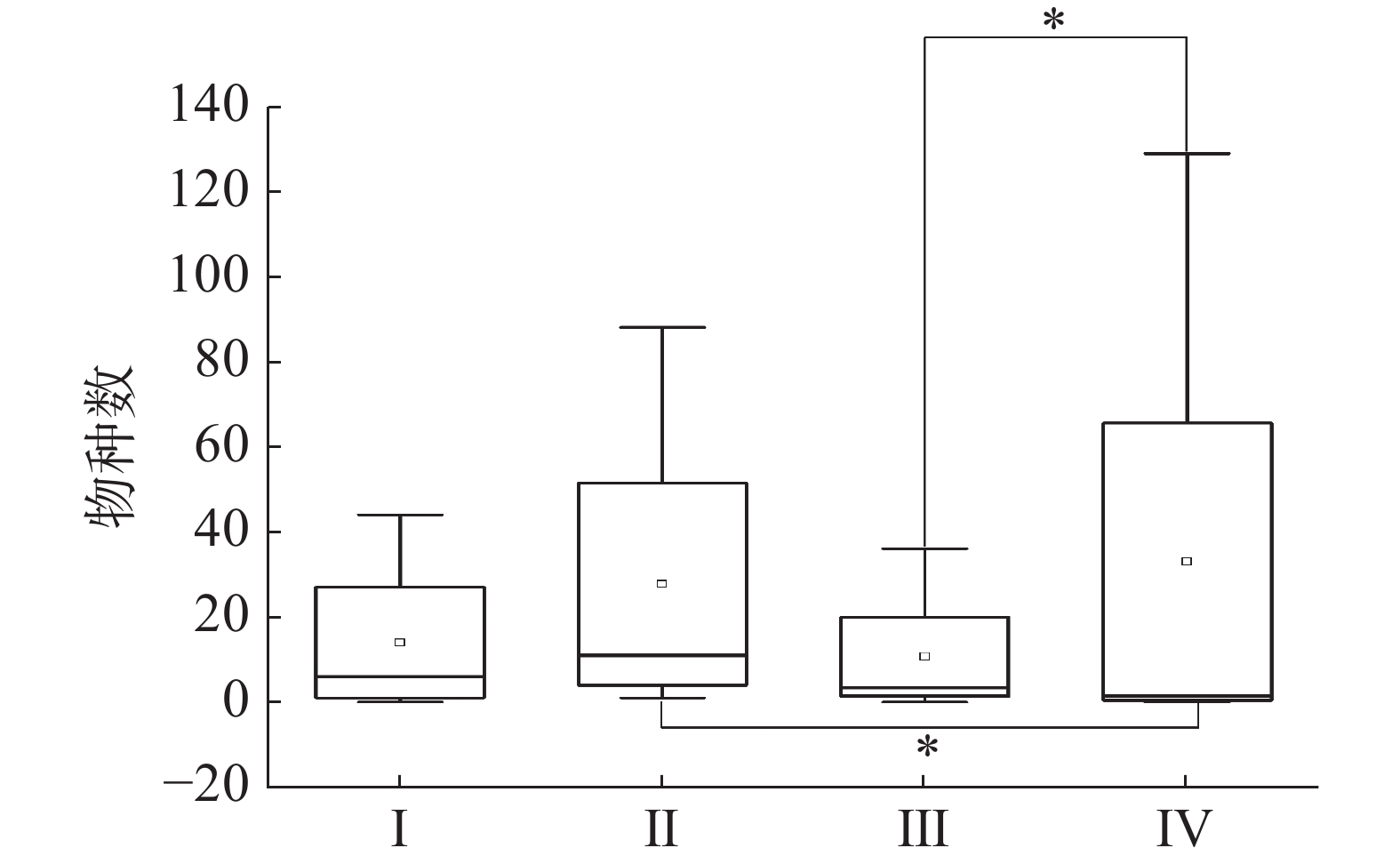
当相似尺度为40%时,2020年和2021年的调查站位可分为四类群落,2020年SYW1-1为Ⅰ类,SYW2-1和SYW3-1为Ⅱ类,SYW3-1为Ⅲ类,2021年的4个调查站位为Ⅳ类(图4)。Kruskal-Wallis检验表明Ⅳ类浮游植物群落与Ⅱ和Ⅲ类浮游植物群落存在显著性差异(P<0.05),与Ⅰ类则不存在显著性差异(P>0.05),综合说明2021年浮游植物群落结构与2020年存在明显差异(图5)。
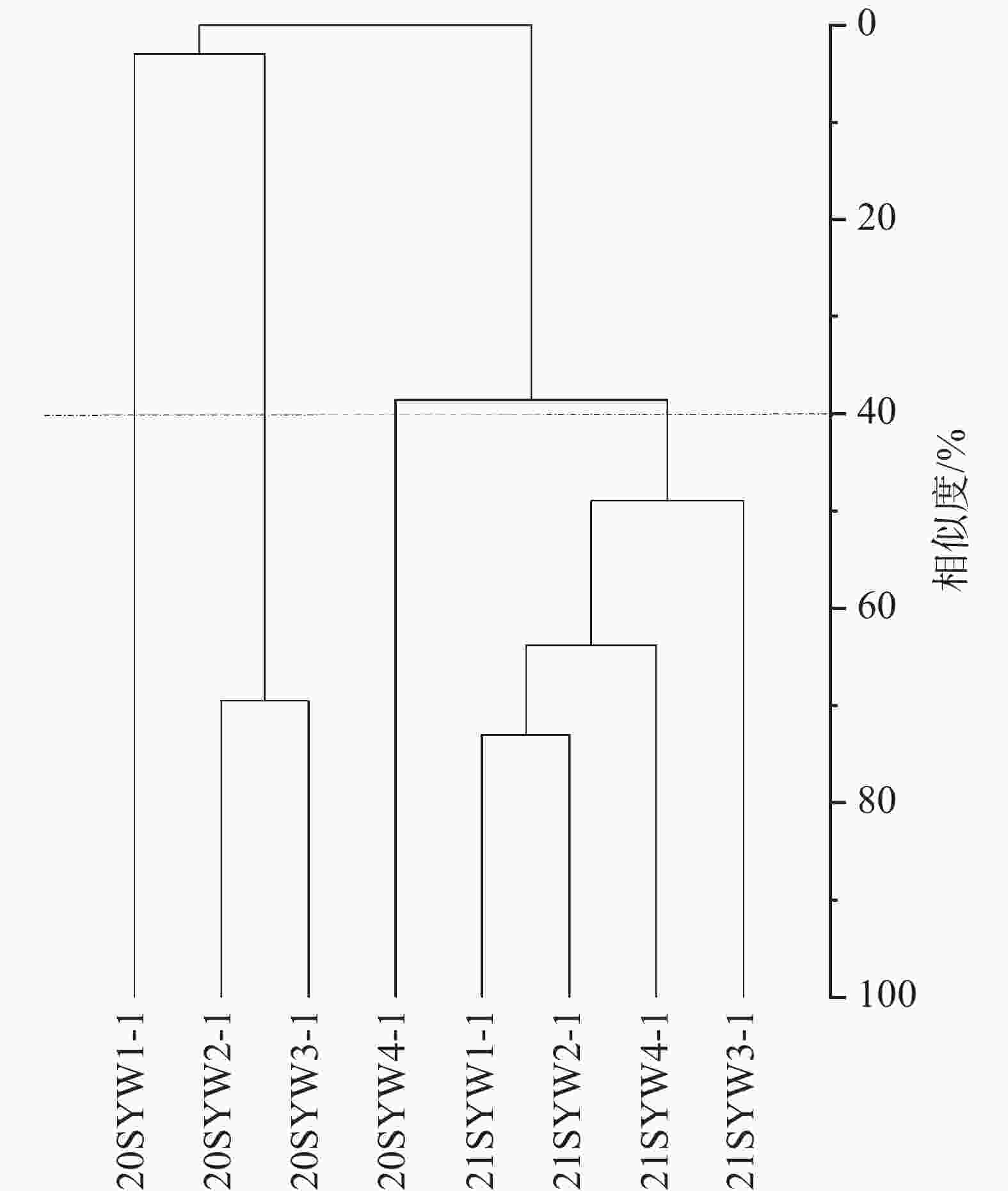
图 4 三亚湾2020年和2021年浮游植物聚类分析
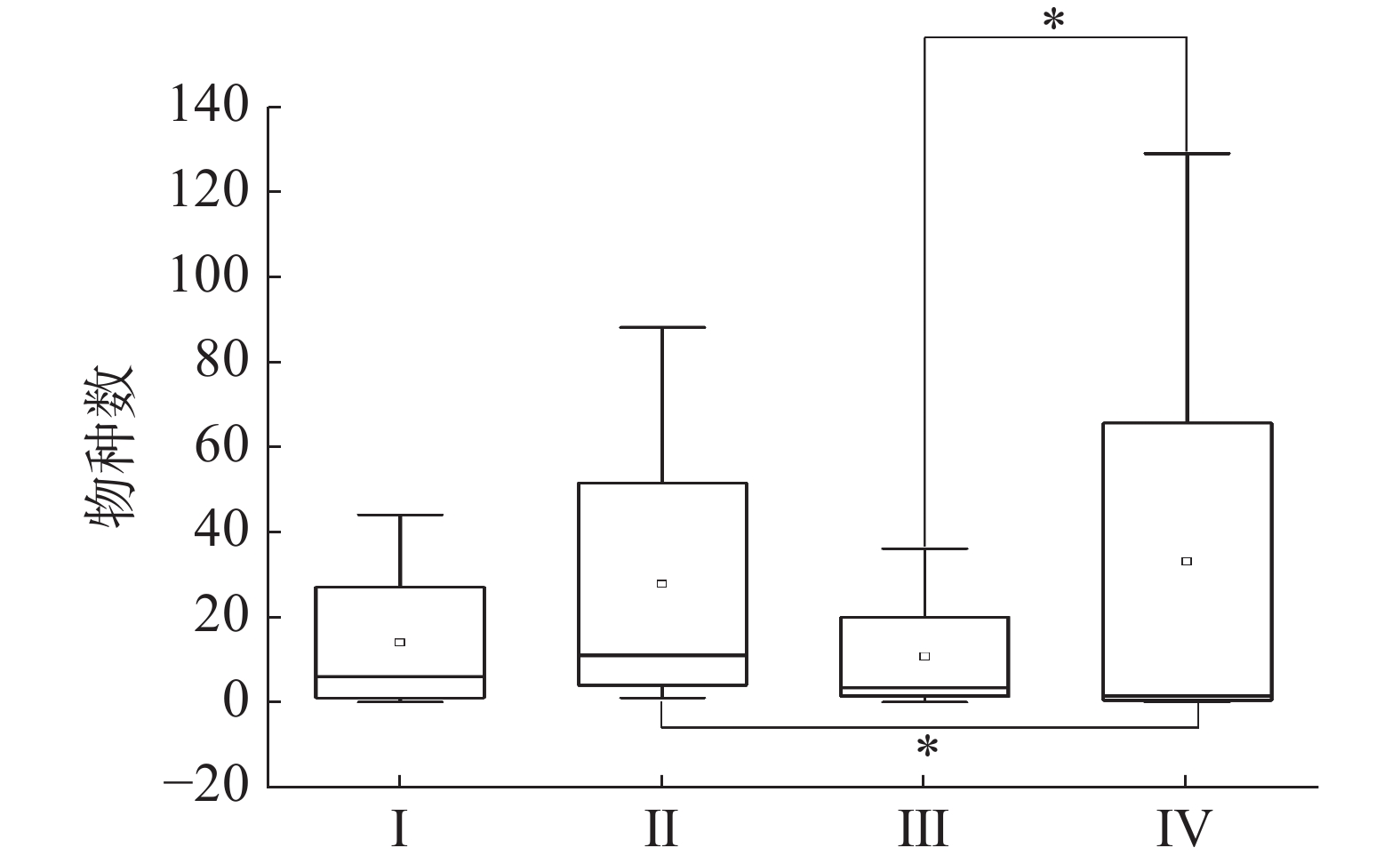
图 5 Kruskal-Wallis检验
-
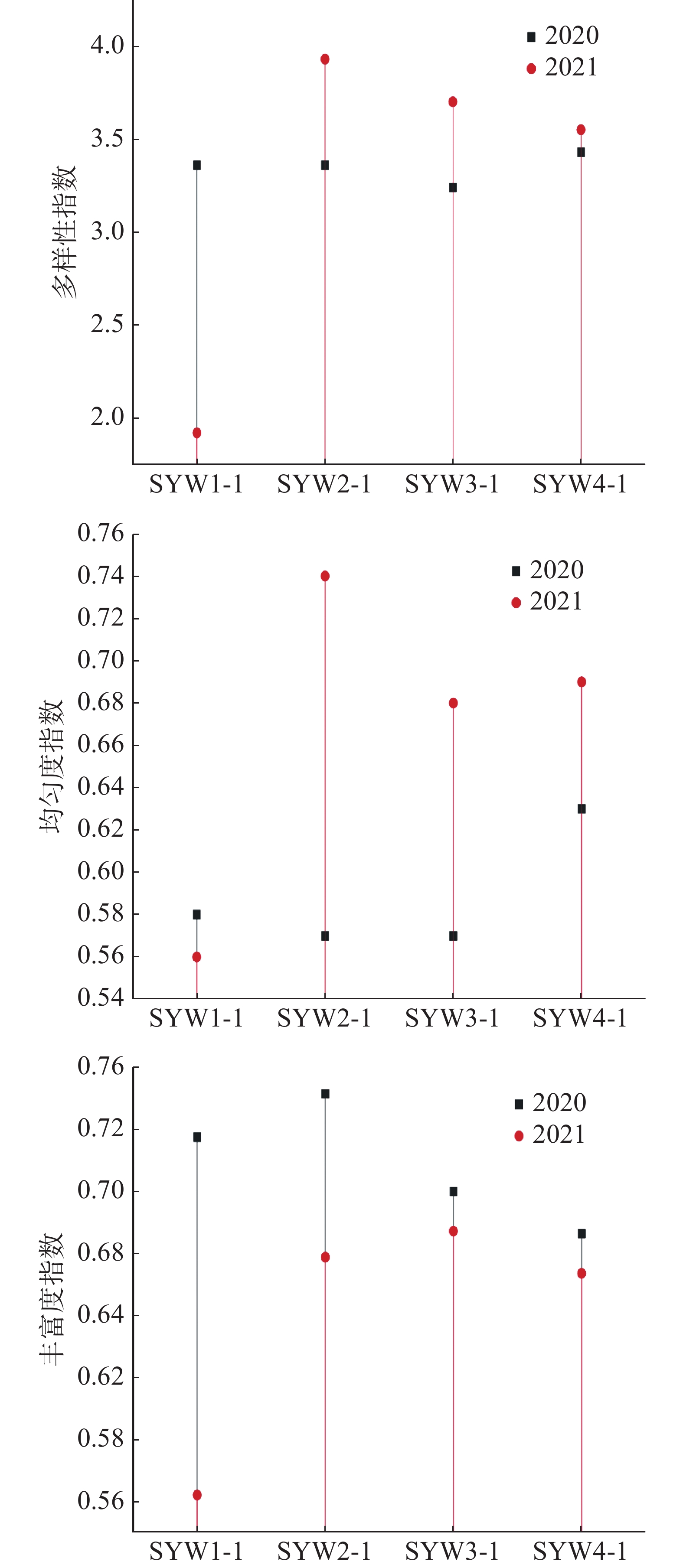
浮游植物多样性指数是反映其种类的多寡和各个种类数量差异的函数关系,均匀度则反映其种类数量的分布情况,可以作为生态监测的参数。一般多样性指数大于等于3.0时, 表明生态环境优良;丰富度指数大于等于3时,表明水质轻度污染或无污染,大于等于1小于3时,表明水质中度污染,小于1时,表明水质重度污染。三亚湾2020年和2021年浮游植物4个站位的多样性指数和均匀度指数的平均值差异不大,2020年多样性指数均值为3.35,高于2021年的3.27;2020年均匀度指数均值为0.59,低于2021年的0.66;2020年丰富度指数均值为3.72,高于2021年的2.5。2021年SYW1-1站位的多样性指数比2020年明显降低了43.15%,丰富度指数明显降低了73.60%,而2021年SYW2-1站位的均匀度指数比2020年明显升高了29.82%(图6)。
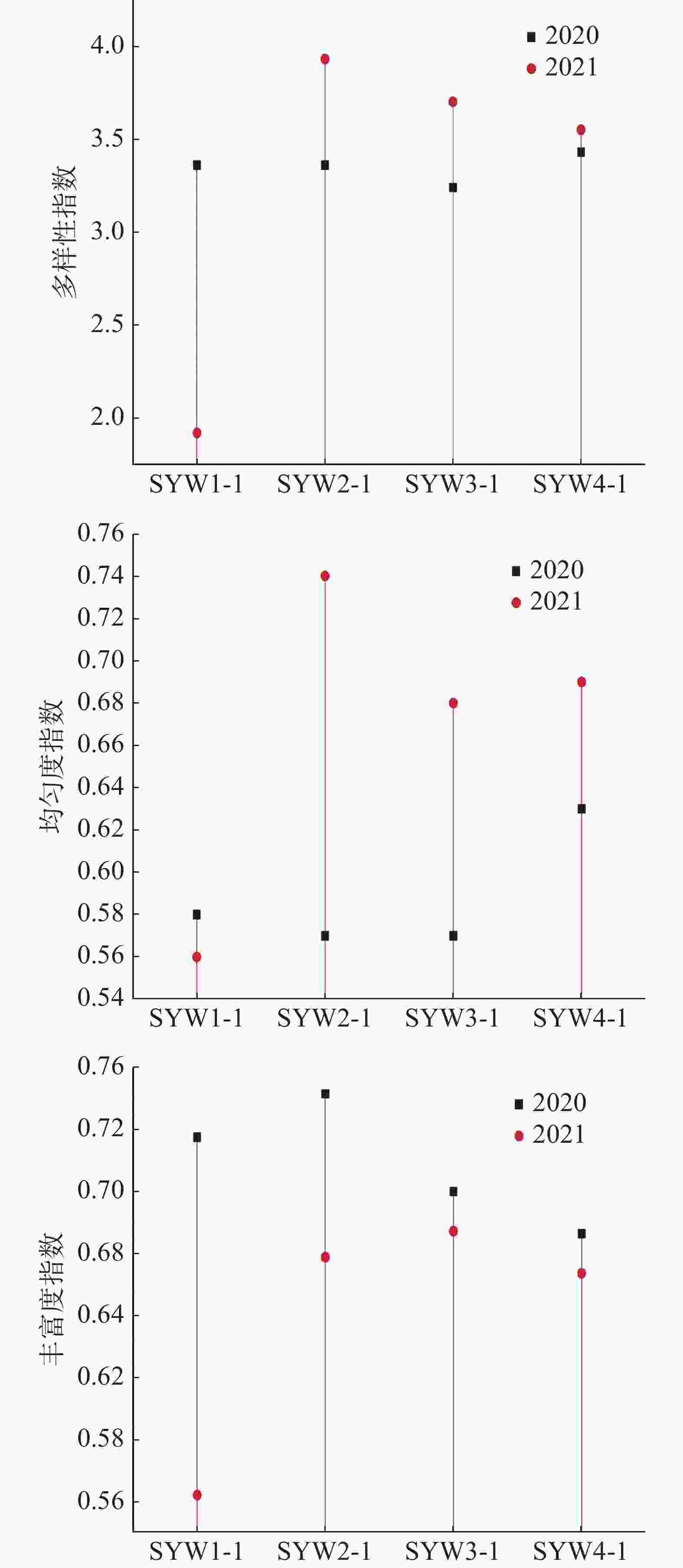
图 6 三亚湾各站位浮游植物多样性指数、均匀度和丰富度指数
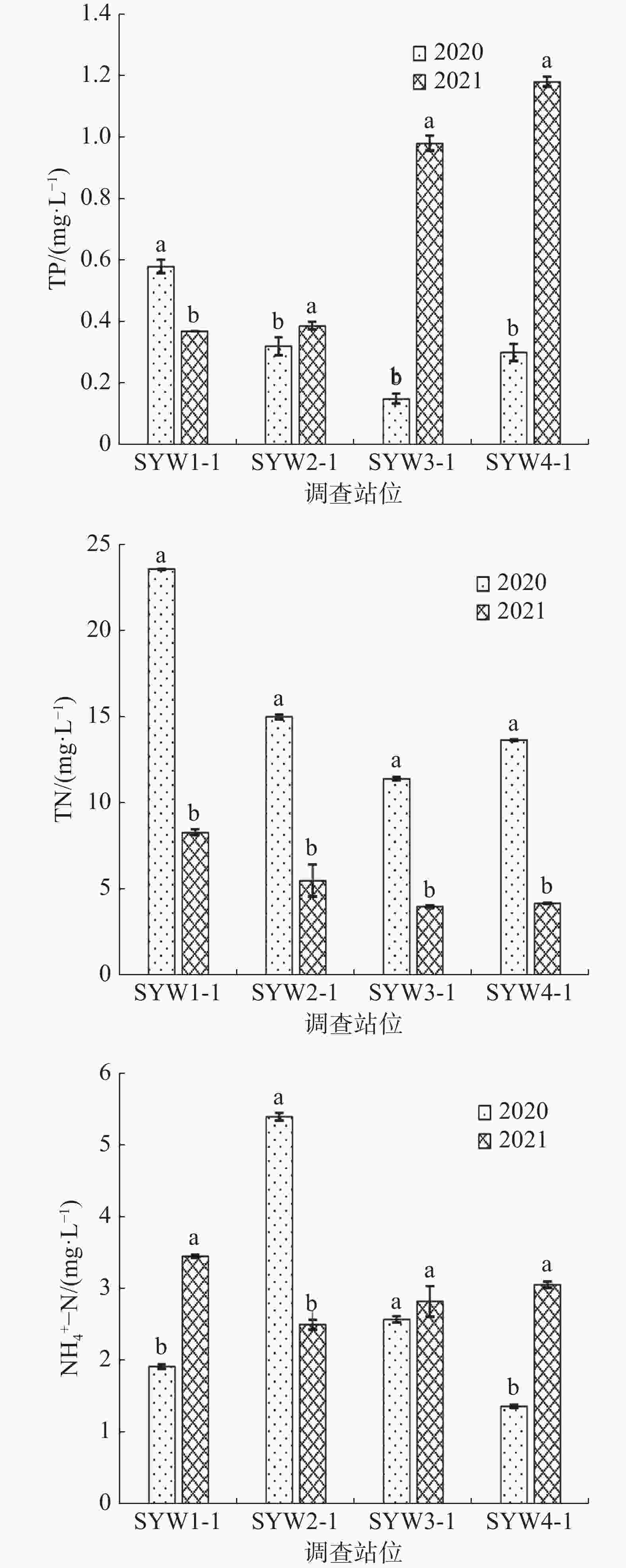
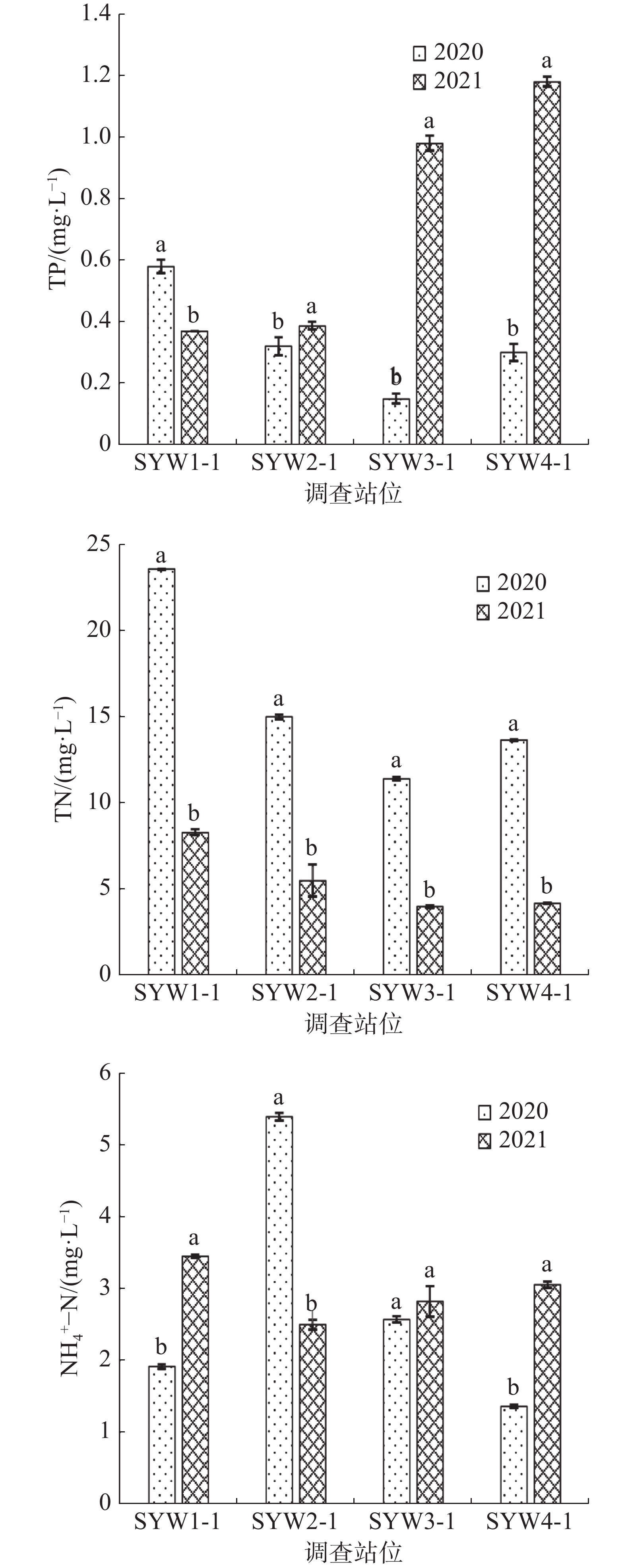
图 7 三亚湾水质化学指标
-
如表3所示,2020年和2021年三亚湾调查期间平均气温差别不大,而2021年的电导率比2020年明显升高了70.5%(表3)。各站位总氮(TN)、总磷(TP)、氨氮(NH4+-N)含量如图7所示,2021年水质中总氮(TN)含量平均值为5.47 mg·L−1且4个调查站位的TN含量均显著低于2020年(P<0.05);而2021年水质中的总磷(TP)和氨氮(NH4+-N)的平均值分别为0.73 和2.95 mg·L−1,均高于2020年的0.34和2.81 mg·L−1。
表 3 三亚湾各调查站位环境参数表
站位 气温/℃ 水温/℃ 电导率/
(µs·cm−1)盐度/% 溶解氧/
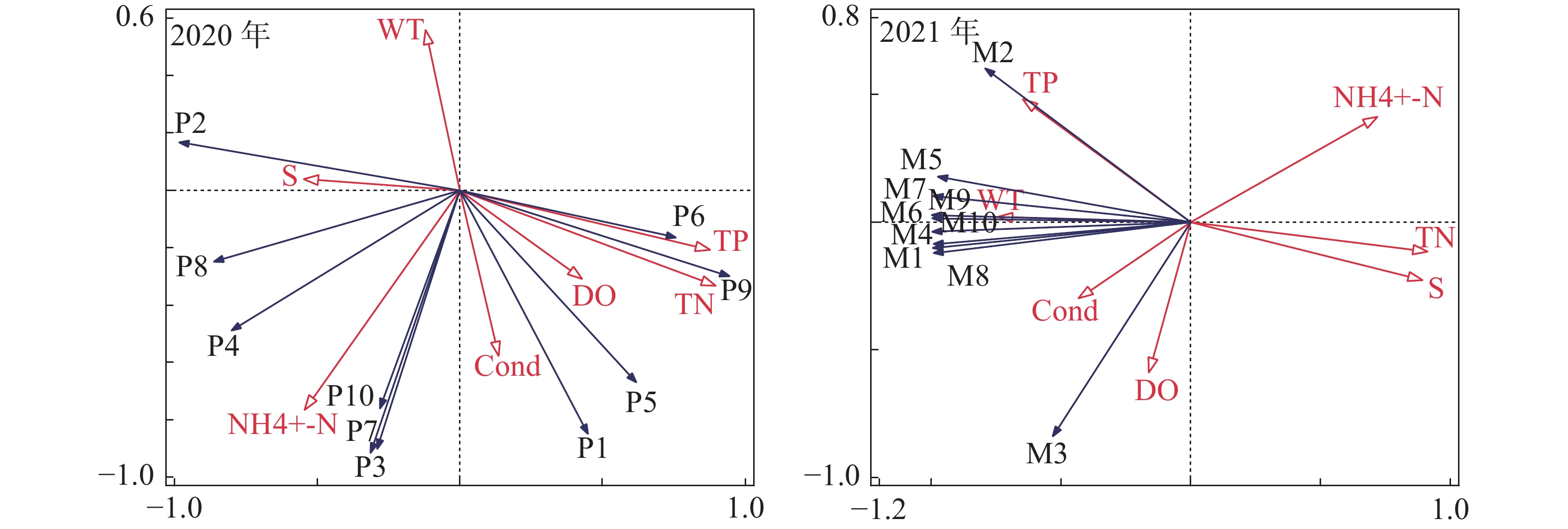
(mg·L−1)2020 2021 2020 2021 2020 2021 2020 2021 2020 2021 SYW1-1 32.5 32.4 30.1 27.0 22.14 37.6 2.9 3.4 5.7 8.0 SYW2-1 32.2 33.1 30.0 27.1 22.14 38.1 3.0 3.4 6.6 8.1 SYW3-1 32.0 33.2 29.9 27.6 22.27 37.8 3.1 3.3 4.0 8.0 SYW4-1 31.9 32.6 32.0 27.4 22.13 37.8 3.0 3.3 4.6 7.9 平均值 32.2 32.8 30.5 27.3 22.17 37.8 3.0 3.3 5.2 8.0 将三亚湾2020年和2021年的浮游植物与环境因子进行RDA排序(图8)的分析结果表明:0.4665和0.3848分别为2020年前两个排序轴的特征值,累计解释了85.13%的物种变化信息;0.8791和0.1144分别为2021年前两个排序轴的特征值,累计解释了99.35%的物种变化信息。
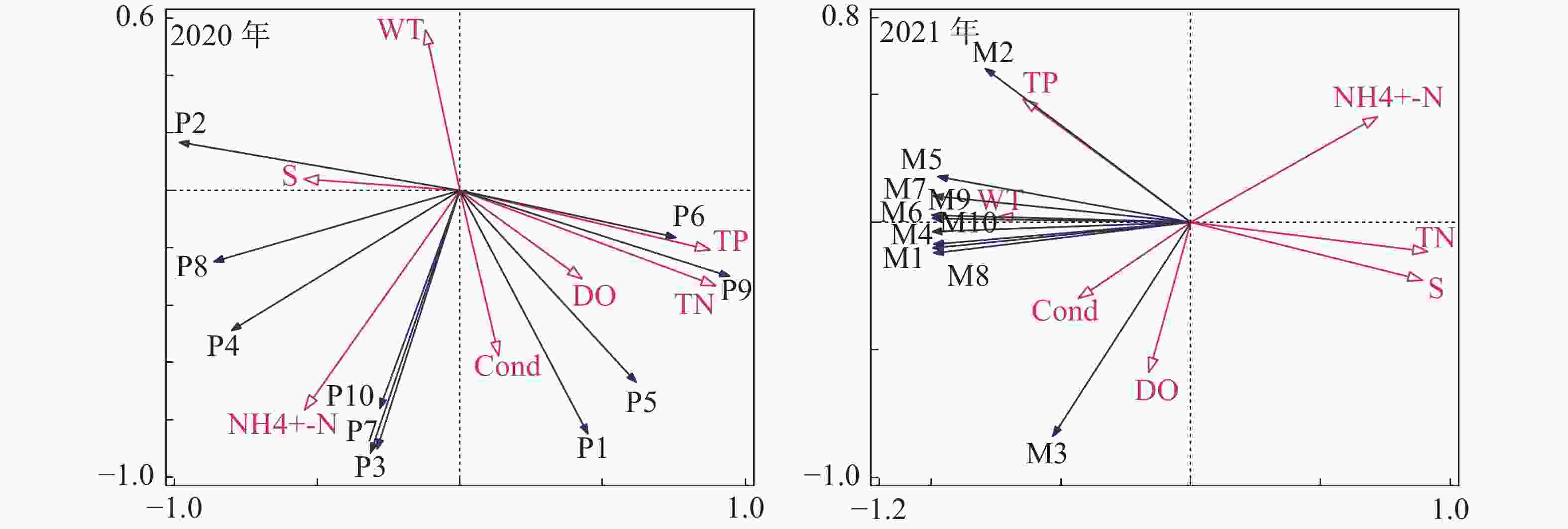
图 8 三亚湾2020年和2021年浮游植物与环境因子的RDA排序图
利用向前引入法逐步筛选环境因子,在7个环境因子中,总氮(TN)(F=7.4,P=0.002),氨氮(NH4+-N)(F=15,P=0.002)和总磷(TP)(F=7.2,P=0.03)及电导率(Cond)(F=5.3, P=0.034)的检验结果在2020年的调查结果中达到显著水平;氨氮(NH4+-N)(F=12.5,P=0.018)、电导率(Cond)(F=17.3, P=0.006)、总磷(TP)(F=11.4,P=0.002)和盐度(S)(F=60.3,P=0.002)的检验结果在2021年的调查结果中达到显著水平。其中氨氮、电导率、总磷在2020年和2021年的检验结果中均达到显著水平,说明这3个环境因子在影响三亚湾浮游植物群落结中占据着更重要的地位。
RDA排序结果表明,2020年的海洋角毛藻与WT、S、NH4+-N呈正相关关系,与Cond、DO、TN、TP呈负相关关系;尖刺拟菱形藻、窄隙角毛躁与S、NH4+-N、Cond呈正相关关系,与WT、DO、TN、TP呈负相关系;劳氏角毛藻、扁面角毛藻、异角毛躁与WT呈负相关关系,与其他环境因子呈正相关关系;角毛藻属和并基角毛藻与WT、S呈负相关关系,与其他环境因子呈正相关关系;柔弱拟菱形藻和短孢角毛藻与WT、S呈负相关关系,与其他环境因子呈正相关关系;2021年浮游植物与环境因子的关系发生明显变化,劳氏角毛躁与TP、WT、Cond呈正相关关系,与其他4种环境因子呈负相关关系;角毛藻除与溶解氧呈正相关关系和与TP呈负相关关系外,与其他环境因子的相关关系与劳氏角毛藻相同;其余8种浮游植物与TP、WT、Cond、DO呈正相关关系,与其他3种环境因子呈负相关关系。
-
2020年浮游植物平均细胞密度为1.031×106 个·m−3,2021年为4.142×105个·m−3,根据浮游植物细胞密度评价水质的标准[15],2020年水体为贫营养状态,2021年为极贫营养状态,营养减弱可能导致了2021年浮游植物的门类数和种类数少于2020年。但在本研究的水质测定结果中,2021年的营养盐元素氨氮和总磷含量略高于2020年,这可能是利用哈希DR3900多参数水质检测仪检测时出现误差造成的。总的来看,2020年和2021年的水体营养都处于较低水平,未有富营养化趋势。在营养水平较低的水体中浮游植物常以硅藻为主[16],这与本研究硅藻门种类数和细胞丰度在2020年和2021年均占最大比例的调查结果相互印证,杨志浩等[17]和Yanying等[18]对三亚湾浮游植物的调查结果也与此一致。隐藻门植物更偏向于生活在温度较低的水体中[19],而4月份三亚湾的水体温度较高,因此2021年的调查结果中未发现任何隐藻门植物,且2020年也仅鉴别到一种隐藻门植物,也可能是因为其体积较小,游泳速率快[20],网采取样难以捕捉到。在2020年的蓝藻门中鉴定到三种束毛藻属(其中包括红海束毛藻)和一种鱼腥藻属(多变鱼腥藻),束毛藻是蓝藻中赤潮的主要种类,尤其是红海束毛藻已在中国近海引发多次赤潮[21],而三亚湾在调查时并未出现赤潮,可能是因为水体营养水平较低,未能达到形成赤潮时所需的细胞丰度,随着2021年水体营养水平的进一步降低,该藻种消失,只鉴定到1种束毛藻属,这与前面的研究结果一致;鱼腥藻属可产生对肝有毒性的微囊藻毒素,鱼腥藻种类还可以产生生物碱毒素[22],这可能会通过食物链的积累传递对动物和人类构成潜在的健康风险,但在2021年未发现该藻种,降低了该风险发生的可能性。
优势种在群落结构的演替方向中起着支配作用[23],2020年优势种中多为角毛藻属,角毛藻类最佳生长繁殖的温度为30 ℃[24],而2020年的水温正好在30 ℃左右。2021年优势种种类发生变化,其中角毛藻属数减少,浮游植物的分布与水体环境密切相关,优势种的变化说明2020—2021年三亚湾水体环境发生了较大改变。2020年三亚湾浮游植物的多样性指数和丰富度指数大于3,均匀度指数大于0.3,表明此时三亚湾生态环境优良,浮游植物种类丰富,分布均匀且群落结构较为稳定[25],水质无污染;2021年多样性指数大于3,均匀度指数大于0.3,而丰富度指数大于1小于3,表明三亚湾生态环境虽仍保持在优良内,但水质已受到中度污染,三亚湾临近市区,其污染程度主要受人为活动和城市废水排放的影响,说明可能是2021年三亚湾的人为活动强度增大和城市废水排放量增加导致的。
RDA结果表明,氨氮、电导率、总磷是影响三亚湾2020年和2021年浮游植物群落结构的主要环境因子。2020年总磷和总氮与浮游植物优势度排名前10中的9种浮游植物密度呈显著正相关,N、P元素是浮游植物生长所必需的营养元素,适量的N、P浓度能够促进浮游植物的生长[26],这与Bi等[27]的研究结果一致。电导率可间接反映水中无机盐浓度,密切关联着水体营养状态[28],较高的营养盐含量有利于浮游植物生长,这使得2020年和2021年优势度排名前10的浮游植物密度均与电导率呈正相关。除此以外,电导率的大小对浮游植物群落的构建有一定程度的影响[29],电导率的改变也会导致浮游植物α多样性发生显著变化[30],本研究中2021年电导率显著高于2020年,这可能是导致聚类分析和Kruskal-Wallis检验结果中2020年的浮游植物群落结构与2021年的存在明显差异的原因之一,而群落结构差异主要是由上述的水体环境及营养状态发生改变造成的。
综上所述,三亚湾2020年和2021年浮游植物群落结构主要以硅藻门为主,总体较为稳定,由于水体处于贫营养状态,尽管出现赤潮藻种,但也达不到形成赤潮时所需的细胞丰度,所以没有形成赤潮和出现水体富营养化的可能,且2020年和2021年三亚湾生态环境总体较为优良。但值得注意的是,2021年三亚湾的水质已受到污染,当地相关部门应提高警惕,及早采取应对措施,减少废水、污水的排放,降低人为活动的影响,防止三亚湾水体进一步恶化,保护海洋生态环境。
Characteristics of phytoplankton community in the offshore area of Sanya Bay
-
摘要: 为研究海南三亚湾浮游植物的群落特征和水质状况,于2020年和2021年的4月对三亚湾近岸海域进行了2次网采取样调查。采用Shannon-Wiener多样性指数、Pielou 均匀度指数、优势度和丰富度指数对浮游植物群落特征和水质环境状况进行分析评价。镜检结果显示,2020年鉴定出浮游植物4门98种;2021年鉴定出浮游植物为3门65种;硅藻类在2次调查结果中均为优势类群。浮游植物优势种2020年为7种,2021年为9种,聚类分析与秩和检验(Kruskal-Wallis)结果显示,浮游植物群落结构2020年和2021年存在显著差异。2020年的多样性指数和丰富度指数分别为3.35和3.72,均高于2021年的多样性指数(3.27)和丰富度指数(2.5),而2020年的均匀度指数为0.59,低于2021年(0.66)的,且2021年比2020年的浮游植物平均细胞密度降低了41.95%。冗余分析(RDA)结果表明,氨氮、电导率、总磷是影响三亚湾2020年和2021年浮游植物群落结构的主要环境因子。以上结果表明,三亚湾2020年和2021年生态环境总体较为优良,浮游植物群落结构主要以硅藻门为主,水体处于贫营养状态,没有形成赤潮和出现水体富营养化的可能。Abstract: In order to study the characteristics of phytoplankton community and water quality in Sanya Bay, Hainan Province, net sampling surveys were conducted in April 2020 and 2021, respectively. Samples of phytoplankton and seawater were collected in 4 stations, and their community characteristics and water quality and environmental status were analyzed and evaluated by using Shannon-Wiener diversity index, Pielou evenness index, dominance index and richness index. Microscopic examination results showed that 98 species in 4 phyla of phytoplankton were identified in 2020 and 65 species in 3 phyla in 2021. Diatoms were the dominant group in both surveys. The number of dominant phytoplankton species was 8 in 2020 and 9 in 2021. Cluster analysis and Kruskal-Wallis test showed that the phytoplankton community structure in 2020 was significantly different from that in 2021. The diversity index and richness index in 2020 are 3.35 and 3.72, respectively, both of which are higher than those (3.27 and 2.5, respectively) in 2021, while the evenness index is 0.59, which is lower than that (0.66) in 2021. The average phytoplankton cell density in 2021 was 41.95% lower than that in 2020. The redundancy analysis (RDA) showed that ammonia nitrogen, conductivity and total phosphorus were the main environmental factors affecting the phytoplankton community structure in Sanya Bay in 2020 and 2021. All the results showed that the sea water in Sanya Bay is generally excellent ecologically in 2020 and 2021. The phytoplankton community structure is mainly composed of diatoms. The sea water has poor nutrients, indicating no possibility of occurrence of red tides or eutrophication.
-
Key words:
- Sanya Bay /
- phytoplankton /
- water quality /
- community characteristics
-
图 8 三亚湾2020年和2021年浮游植物与环境因子的RDA排序图
TN:总氮;TP:总磷;NH4+-N:氨氮; WT:水温;S:盐度;DO:溶解氧;Cond:电导率;P1:角毛藻属;P2:海洋角毛藻;P3:劳氏角毛藻;P4:尖刺拟菱形藻;P5:并基角毛藻;P6:柔弱拟菱形藻;P7:扁面角毛藻;P8:窄隙角毛藻;P9:短孢角毛藻;P10:异角毛藻;M1:大角管藻;M2:劳氏角毛藻;M3:角毛藻;M4:拟旋链角毛藻;M5:翼根管藻纤细变型;M6:覆瓦根管藻;M7:菱形海线藻;M8:双凹梯形藻;M9:短角弯角藻;M10:笔尖形根管藻。
表 1 三亚湾调查站位地理坐标
站位 经度 纬度 SYW1-1 109°29′15.70″ 18°15′03.03″ SYW2-1 109°27′46.06″ 18°16′25.97″ SYW3-1 109°25′18.58″ 18°17′09.23″ SYW4-1 109°23′09.19″ 18°17′07.35″  下载: 导出CSV
下载: 导出CSV
表 2 浮游植物优势种
时间 优势种 平均密度/
(104个·m−3)占总密
度比例/%出现
频率/%优势
度2020年4月 角毛藻属 15.74 22.06 100 0.221 海洋角毛藻 14.65 20.53 91.67 0.188 劳氏角毛藻 9.21 12.91 100.00 0.129 尖刺拟菱形藻 8.16 11.44 100.00 0.114 并基角毛藻 4.02 5.64 100.00 0.056 柔弱拟菱形藻 3.40 4.76 100.00 0.048 扁面角毛藻 3.31 4.64 100.00 0.046 窄隙角毛藻 2.58 3.62 91.67 0.033 2021年5月 大角管藻 6.51 15.72 100.00 0.157 劳氏角毛藻 5.31 12.82 100.00 0.128 角毛藻 5.15 12.44 100.00 0.124 拟旋链角毛藻 5.71 13.77 75.00 0.103 翼根管藻
纤细变型3.80 9.17 100.00 0.092 菱形海线藻 2.79 6.72 100.00 0.067 覆瓦根管藻 1.98 4.78 75.00 0.036 双凹梯形藻 1.86 4.49 75.00 0.034 短角弯角藻 1.27 3.08 75.00 0.023
下载: 导出CSV
表 3 三亚湾各调查站位环境参数表
站位 气温/℃ 水温/℃ 电导率/
(µs·cm−1)盐度/% 溶解氧/
(mg·L−1)2020 2021 2020 2021 2020 2021 2020 2021 2020 2021 SYW1-1 32.5 32.4 30.1 27.0 22.14 37.6 2.9 3.4 5.7 8.0 SYW2-1 32.2 33.1 30.0 27.1 22.14 38.1 3.0 3.4 6.6 8.1 SYW3-1 32.0 33.2 29.9 27.6 22.27 37.8 3.1 3.3 4.0 8.0 SYW4-1 31.9 32.6 32.0 27.4 22.13 37.8 3.0 3.3 4.6 7.9 平均值 32.2 32.8 30.5 27.3 22.17 37.8 3.0 3.3 5.2 8.0
下载: 导出CSV
-
[1] CARBON B. A Rapid Response Assessment[M]. Norway: Birkeland Trykkeri AS, Norway, 2009. [2] 沈国英, 施并章. 海洋生态学[M]. 2版. 北京: 科学出版社, 2002. [3] HOOD R R, LAWS E A, ARMSTRONG R A, et al. Pelagic functional group modeling: progress, challenges and prospects[J]. Deep Sea Research Part II:Topical Studies in Oceanography, 2006, 53(5/6/7): 459 − 512. [4] CHEN C C, GONG G C, SHIAH F K. Hypoxia in the East China Sea: one of the largest coastal low-oxygen areas in the world[J]. Marine Environmental Research, 2007, 64(4): 399 − 408. doi: 10.1016/j.marenvres.2007.01.007 [5] 李阳东, 谢洋洋, 李仁虎, 等. 浙江近岸海域赤潮事件时空分布特征研究[J]. 海洋预报, 2021, 38(1): 54 − 60. doi: 10.11737/j.issn.1003-0239.2021.01.007 [6] CROSSETTI L O, DE M BICUDO C E, Bicudo C E. Phytoplankton as a monitoring tool in a tropical urban shallow reservoir (Garças Pond): the assemblage index application[J]. Hydrobiologia, 2008, 610(1): 161 − 173. doi: 10.1007/s10750-008-9431-z [7] 亚力士(PHANMAHA ALEK). 三亚湾近岸海域及其珊瑚生长区重金属污染现状与评价[D]. 南宁: 广西大学, 2018. [8] 李亚男, 庞富广, 臧瑜, 等. 三亚湾沙滩小型底栖动物时空异质性及影响因素[J]. 海洋湖沼通报, 2022, 44(3): 139 − 148. [9] 王艳红, 曾成杰, 陆培东, 等. 三亚湾东段海岸侵蚀与海滩养护[J]. 自然资源学报, 2022, 37(4): 1049 − 1060. [10] 贾磊, 刘文涛, 唐得昊, 等. 三亚湾及周边海域表层沉积物重金属分布特征及生态风险评价[J]. 海洋地质前沿, 2020, 36(12): 22 − 31. doi: 10.16028/j.1009-2722.2020.029 [11] 中华人民共和国国家质量监督检疫总局, 中国国家标准化管理委员会. 海洋调查规范 第9部分: 海洋生态调查指南:GB/T 12763.9——2007 [S]. 北京: 中国标准出版社, 2008.2007. [12] 国家质量监督检验检疫总局, 中国国家标准化管理委员会. 海洋监测规范 第7部分: 近海污染生态调查和生物监测:GB 17378.7——2007[S]. 北京: 中国标准出版社. 2008. [13] 李亚军, 王先明, 程贤松, 等. 海南岛海尾湾浮游植物群落结构及水质的调查[J]. 热带生物学报, 2020, 11(3): 257 − 265. doi: 10.15886/j.cnki.rdswxb.2020.03.001 [14] 徐明, 许静波, 唐春燕, 等. 大纵湖浮游植物群落特征及其与环境因子的关系[J]. 水生态学杂志, 2021, 42(6): 64 − 69. doi: 10.15928/j.1674-3075.201908030196 [15] 况琪军, 马沛明, 胡征宇,等. 湖泊富营养化的藻类生物学评价与治理研究进展[J]. 安全与环境学报, 2005, 5(2): 87 − 91. doi: 10.3969/j.issn.1009-6094.2005.02.024 [16] 张春梅, 朱宇轩, 宋高飞, 等. 南水北调中线干渠浮游植物群落时空格局及其决定因子[J]. 湖泊科学, 2021, 33(3): 675 − 686. doi: 10.18307/2021.0305 [17] 杨志浩, 董俊德, 吴梅林, 等. 三亚湾网采浮游植物群落结构特征分析[J]. 热带海洋学报, 2007, 26(6): 62 − 66. doi: 10.3969/j.issn.1009-5470.2007.06.011 [18] ZHANG Y, DONG J, LING J, et al. Phytoplankton distribution and their relationship to environmental variables in Sanya Bay, South China Sea[J]. Scientia Marina, 2010, 74(4): 783 − 792. doi: 10.3989/scimar.2010.74n4783 [19] BARLOW S B, KUGRENS P. Cryptomonads from the Salton Sea, California[M]. Hydrobiologia, 2002, 473: 129 − 137. [20] 夏爽, 刘国祥, 胡征宇. 武汉东湖隐藻门植物的分类学研究[J]. 植物科学学报, 2011, 29(2): 250 − 255. [21] KRISHNAN A A, KRISHNAKUMAR P K, RAJAGOPALAN M. Trichodesmium erythraeum (Ehrenberg) bloom along the southwest coast of India (Arabian Sea) and its impact on trace metal concentrations in seawater[J]. Estuarine, Coastal and Shelf Science, 2007, 71(3/4): 641 − 646. doi: 10.1016/j.ecss.2006.09.012 [22] 赵文倩, 刘振中, 郭文莉, 等. 浅水湖泊浮水植物芡实收割对浮游植物群落的影响[J]. 生态学报, 2023,43(13): 5558 − 5570. [23] 陈思, 陈海刚, 田斐, 等. 深圳湾浮游植物群落结构特征及其与环境因子的关系[J]. 生态科学, 2021, 40(1): 9 − 16. doi: 10.14108/j.cnki.1008-8873.2021.01.002 [24] 栾会妮, 魏佳丽, 杨磊, 等. 盐度、温度、光照强度对耐高温角毛藻藻株生长的影响[J]. 科学养鱼, 2019(2): 69 − 70. doi: 10.14184/j.cnki.issn1004-843x.2019.02.037 [25] 马建新, 郑振虎, 李云平,等. 莱州湾浮游植物分布特征[J]. 海洋湖沼通报, 2002(4): 63 − 67. doi: 10.3969/j.issn.1003-6482.2002.04.011 [26] 王雅雯, 李迎鹤, 张博, 等. 嘉兴南湖不同湖区浮游动植物群落结构特征与环境因子关系[J]. 环境科学, 2022, 43(6): 3106 − 3117. [27] BI R, ARNDT C, SOMMER U. Stoichiometric responses of phytoplankton species to the interactive effect of nutrient supply ratios and growth rates[J]. Journal of Phycology, 2012, 48(3): 539 − 549. doi: 10.1111/j.1529-8817.2012.01163.x [28] 李磊, 张莹, 陈宁,等. 大庆龙凤湿地浮游植物功能类群季节变化及其驱动因子[J]. 生态学杂志, 2021, 40(9): 2850 − 2859. doi: 10.13292/j.1000-4890.202109.027 [29] 张静, 胡愈炘, 胡圣, 等. 长江流域浮游植物群落的环境驱动及生态评价[J]. 环境科学, 2023, 44(4): 2072 − 2082. [30] NATASSA S, MATINA K, DIMITRIS T, et al. Patterns in alpha and beta phytoplankton diversity along a conductivity gradient in coastal Mediterranean lagoons [J]. Diversity, 2020, 12(1): 38-(起止页码?). STEFANIDOU N , KATSIAPI M , TSIANIS D , et al. Patterns in alpha and beta phytoplankton diversity along a conductivity gradient in coastal Mediterranean lagoons[J]. Diversity, 2020, 12(1): 38. -

 点击查看大图
点击查看大图

图(8) / 表 (3)
计量
- 文章访问数: 1583
- HTML全文浏览量: 731
- PDF下载量: 35
- 被引次数: 0





