


-
溶藻弧菌(Vibrio alginolyticus)是一种海洋中常见的细菌,属于弧菌科(Vibrionaceae)弧菌属(Vibrio)一类不产荚膜与芽孢的革兰氏阴性细菌,外形呈杆棒状,具有端生鞭毛与周生鞭毛,耐盐性极强[1-4]。溶藻弧菌对水产养殖中的鱼类、贝类和对虾都具有极强的侵染性和致病性,其导致的弧菌病给养殖业带来了严重的经济损失[5-7]。溶藻弧菌的致病性主要由宿主、致病环境与细菌本身3个要素构成。对宿主进行黏附与侵袭、在宿主体内进行大量增殖、产生毒素等是细菌致病的基本过程,而其中生物被膜(bacterial biofilm,BF)是由细菌本身分泌的多糖、蛋白、eDNA等物质与细胞聚集体结合而成的胞外共生聚合物,是病原菌在宿主中持续反复感染的关键,可以增强细菌的毒力[8-9]。生物被膜成熟后细菌的新陈代谢速率会降低,从而形成休眠体,此时在被膜中的耐药基因被激活,产生胞外酶,进而使细菌产生耐药性,同时也增强对于不良环境的抵抗力[10-11]。生物被膜功能大多数与细菌产生的胞外聚合物质有关,eDNA分子在生物被膜的骨架结构中发挥着至关重要的作用。对eDNA具有降解作用的DNase,同样可以有效抑制与清除生物被膜。研究表明,肺炎球菌与金黄色葡萄球菌可以分泌一些脱氧核糖核酸酶以促进毒力的表达,这些核酸酶可以水解宿主中性粒细胞为抑制病原菌所产生的DNA骨架结构,从而增强其毒力[12];但也有着发挥解聚酶作用的DNase,如细菌的TatD蛋白。TatD蛋白是一种在原核生物与真核生物中都存在的蛋白质,大多数只具有1个与TIM-桶状折叠相似的DNase结构域[13-15]。然而,真核生物恶性疟原虫TatD蛋白具有与常见TatD蛋白有所区别的特殊结构,其TatD蛋白具有1个长为24~26个氨基酸分子的信号肽,氨基酸序列表明该蛋白能够被分泌到细胞外部[16-17]。在TatD蛋白的探究初期,人们对模式生物大肠杆菌的TatD蛋白进行了深入分析,解析其晶体结构并模拟了该蛋白与DNA分子之间结合作用的机制[18]。研究者通过解析大肠杆菌中的TatD蛋白,辅助多种方式确定了其晶体构造,TatD蛋白TIM桶状结构具有DNA酶的性质,与DNA分子的结合模式依赖于金属离子。而与金属离子结合的区域为TIM桶状上方的Loop结构,该结构类似于酸性布袋,经过X射线衍射,表明其由α螺旋与8个β折叠共同形成,与核酸内切酶Endo Ⅳ结构相似[19-20]。本研究构建了tatD基因缺失突变菌株,通过探究tatD基因在溶藻弧菌生长与生物被膜形成上的功能,可为溶藻弧菌病害防控提供参考。
-
实验所用溶藻弧菌由海南大学南海微生物资源开发与利用课题组提供,菌株编号为HN08155(WT),分离自1只患病石斑鱼。实验所用具有氯霉素与苄青霉素抗性的自杀质粒pDM4由海南大学微生物资源开发与利用课题组保藏,所用DH5α、Top10感受态细胞购自上海唯地生物技术有限公司。
-
1)LB培养基:胰蛋白胨10 g·L−1,酵母膏5 g·L−1,氯化钠20 g·L−1;2)LBS培养基:胰蛋白胨10 g·L−1,酵母膏5 g·L−1,氯化钠30 g·L−1;3)2216E培养基:胰蛋白胨1.5 g·L−1,酵母膏0.3 g·L−1,氯化钠6 g·L−1,磷酸铁0.01 g·L−1;4)M9培养基:M9药品11.4 g·L−1,葡萄糖4 g·L−1,氯化钠19.5 g·L−1。
-
DNA提取试剂盒购自天根生化科技北京有限公司,DNA凝胶回收试剂盒、质粒提取试剂盒购自OMEGA生物技术公司,限制性内切酶、DNA 聚合酶和 T4 连接酶与PCR相关试剂均由 TaKaRa 公司提供,引物合成与测序分别由上海生工生物工程股份有限公司与华大基因科技股份有限公司完成。实验所用其他试剂均为常规试剂。
-
实验前期已对溶藻弧菌HN08155进行全基因组测序分析。从溶藻弧菌全基因组DNA(美吉生信云:
https://www.majorbio.com )中挑选全部tatD基因进行基因信息比对,将tatD基因序列翻译为氨基酸序列,使用Uniprot蛋白数据库,进行蛋白信息查询比对,以验证tatD基因翻译后的氨基酸序列。此外,利用NCBI进行TatD蛋白的结构域分析,同时通过蛋白质数据库Pfam进行结构域的预测验证。应用ProtParam tool预测TatD蛋白的理化性质,包括分子质量、等电点等重要参数。溶藻弧菌DNA利用天根生化科技北京有限公司 DNA提取试剂盒进行提取,以提取DNA为模板,利用tatD基因相关引物(表1)扩增tatD基因ORF框,PCR产物经过切胶、纯化后,由华大基因科技股份有限公司进行测序。表 1 PCR扩增所使用的引物
Table 1. Primers used in PCR
引物
Primer序列
SequencetatD1-T-F ATGAAGCTCTTTGATACCCA tatD1-T-R TTATTCACATATCGAGAAAG tatD1-UF GAAGATCTTCAGAACCAAGCAAACTTAGCA tatD1-UR CCATAACTGAGAGGCAATCGAAGTCACCTTGGAATACATC tatD1-DF GATGTATTCCAAGGTGACTTCGATTGCCTCTCAGTTATGG tatD1-DR CCCTCGAGGGACCGTAGTTGATAACGTTAG tatD1-TF GCATTAACGTTGCAACTGCG tatD1-TR GAATGCGAAGATGAAACCGC tatD2-T-F ATGTTCGTAGATTCTCATTG tatD2-T-R TCATCGCAAAAAAAGATTTC tatD2-UF GAAGATCTTCAACTGAATATGGGCTCTCCG tatD2-UR GCACGATTTGAGTTGCGCAATCCACATTGATGTGGAGATC tatD2-DF GATCTCCACATCAATGTGGATTGCGCAACTCAAATCGTGC tatD2-DR CCCTCGAGGGGTAGCCTAAAGGTTAGCACC tatD2-TF TTACCCACAATCAGGAGCGT tatD2-TR CTGATACTGGAAGCATCAGC tatD3-T-F ATGATCGATACCCACGCGCA tatD3-T-R TTAAATATTAAACAGCTTTT tatD3-UF GAAGATCTTCCGATTGCTGGAACGCCTAGA tatD3-UR AGTTGCGACATCGACATCACACTACTTCATCGCGGTCGTT tatD3-DF AACGACCGCGATGAAGTAGTGTGATGTCGATGTCGCAACT tatD3-DR CCCTCGAGGGGACGTAACCACACTTTGTGC tatD3-TF TCTTCATCGATGATGCCGTC tatD3-TR GCCATTCCGATGTGCTTACT pDM4-TF CACAGGAACACTTAACGGCT pDM4-TR TCCTGTTCAGCTACTGACGG -
使用DNA提取试剂盒TIANGEN提取溶藻弧菌HN08155基因组DNA。利用软件SnapGene viewer以测序获得的HN08155全基因组序列中的tatD基因为模板设计上游/下游同源臂PCR引物(tatD-U、tatD-D,表1)。利用引物tatD-UF+UR、tatD-DF+DR分别扩增tatD基因上下游片段,将扩增上下游片段纯化回收后作为模板,利用tatD-UF+DR引物扩增tatD基因框内缺失片段,PCR产物经过纯化回收与自杀质粒PDM4进行双酶切后连接,构建重组载体PDM4-tatD载体,将重组载体转化至溶藻弧菌(WT)。自杀质粒在宿主内无法正常进行复制,要么会被识别为外源DNA而被清除,要么整合到宿主染色体上,与其一同复制。自杀质粒PDM4含有蔗糖致死基因,当同源性DNA片段发生重组时,在蔗糖培养条件下,无法进行正常的复制,从而筛选出tatD基因缺失株。利用 tatD-TF+TR(表1)引物分别进行验证,在构建成功的单基因缺失菌株上重复上述步骤,从而构建双基因缺失株与三基因缺失株。
-
将保藏的菌株进行活化,挑选单菌落到液体培养基中30 ℃、180 r·min−1培养12 h,将菌液按百分之一的接种量转接到无菌液体培养基中,相同条件下培养至对数生长期。按上述接种方式与接种量将菌液接种至新鲜液体培养基中,混匀后按每孔200 μL培养液转入96孔板,重复6次,酶标仪测定菌液OD600数值,连续监测48 h,通过OD600数值反映菌株的生长情况。LB培养基为富营养条件,M9为低营养条件。
-
将菌株活化后选择单菌落转接至新鲜液体培养基中培养过夜,培养后的菌液按上述菌种量转接至液体培养基中培养至菌株生长对数期,将培养液在4 ℃、5 000 r·min−1条件下离心10 min,弃沉淀取上清液。将从北京索莱宝科技有限公司购来的小牛胸腺DNA溶于无菌ddH2O中,移液枪吸取10×反应缓冲液(MgCl2)1 μL与小牛胸腺DNA 1 μL配制降解反应体系,加入各菌株上清液4 μL,添加无酶ddH2O至10 μL反应体系;EDTA具有螯合金属离子的作用,添加DNase I酶与EDTA作为对照,将反应体系在30 ℃下反应30 min,使用琼脂糖凝胶电泳测定菌液上清液对DNA的降解能力。
-
将菌株在LB液体培养基中活化培养,30 ℃振荡培养至对数生长后期,在4 ℃、4 000 r·min−1条件下离心8 min,收集菌体,用 PBS缓冲液(pH7.4)洗涤2次,重复3次重悬在Triton X-100( 体积百分数为0.1% )缓冲液中,测定其OD600值, 随后在 30 ℃、180 r·min−1下振荡培养。每隔1 h测定菌株OD600值。细菌的自溶率可以通过以下公式得出:
自溶率(%)= 即时吸光度/初始吸光度×100,
其中,即时吸光度表示细菌每间隔1 h后的OD600值,初始吸光度表示初始重悬后菌液的OD600值。
-
将菌种活化后挑选单菌落在5 mL LBS液体培养基培养14 h,按1:100 (L:μL)接种量将菌液转入新鲜LB培养基中振荡培养至菌液OD600值为0.6,将所有菌株菌液OD600值调整至相等,按上述接种方式与接种量将菌液接种至新鲜LB液培养基中,混匀后按每孔200 μL培养液转入96孔板,重复5次。孔板置于30 ℃培养箱中培养,在4、8、16、20、24 h后分别取出孔板,弃上清,用1×PBS(pH7.4)洗脱放置55 ℃条件下烘干15 min,加入0.1%结晶紫溶液(CV)220 μL,30 min后,弃CV溶液,用1×PBS(pH7.4)进行3次洗脱,将孔板放置55 ℃条件下烘干15 min,每孔加200 μL 95%乙醇进行30 min再溶,酶标仪测定孔板OD570值,以时间为横坐标,以OD570值为纵坐标绘制生物被膜形成量曲线。
-
通过全基因组系列分析发现溶藻弧菌HN08155含有3个tatD基因,分别对其命名为tatD1(tatD2409)、tatD2(tatD2024)、tatD3(tatD0097),通过PCR扩增与生物信息学分析发现其碱基对tatD1全长为774bp, tatD2为768 bp,tatD3为762 bp(美吉:MJ20211021082,图1)。Pafm分析工具分析发现3个同源蛋白都带有TIM-桶状的TatD结构域,该结构域具有Mg2+依赖金属水解酶与3’-5’ ssDNA/RNA核酸外切酶特性。
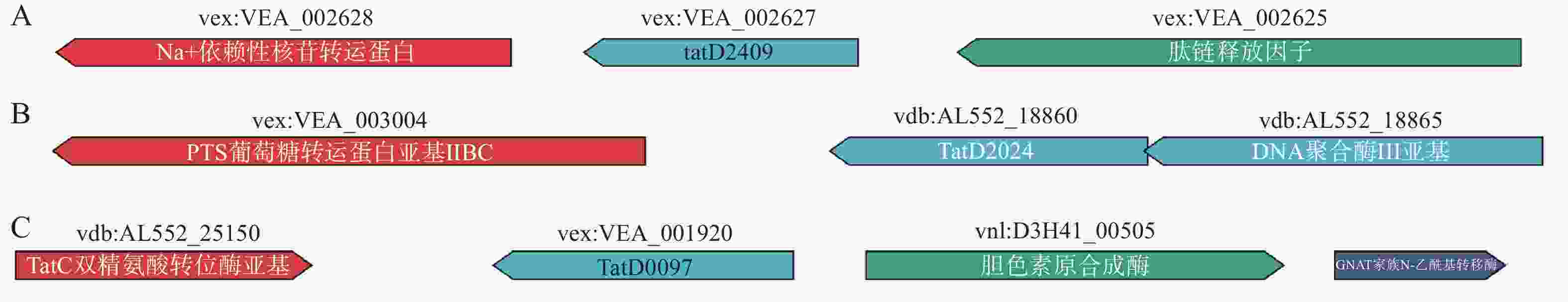
图 1 溶藻弧菌tatD基因示意图
Figure 1. Illustration of Vibrio alginolyticus tatD Gene
-
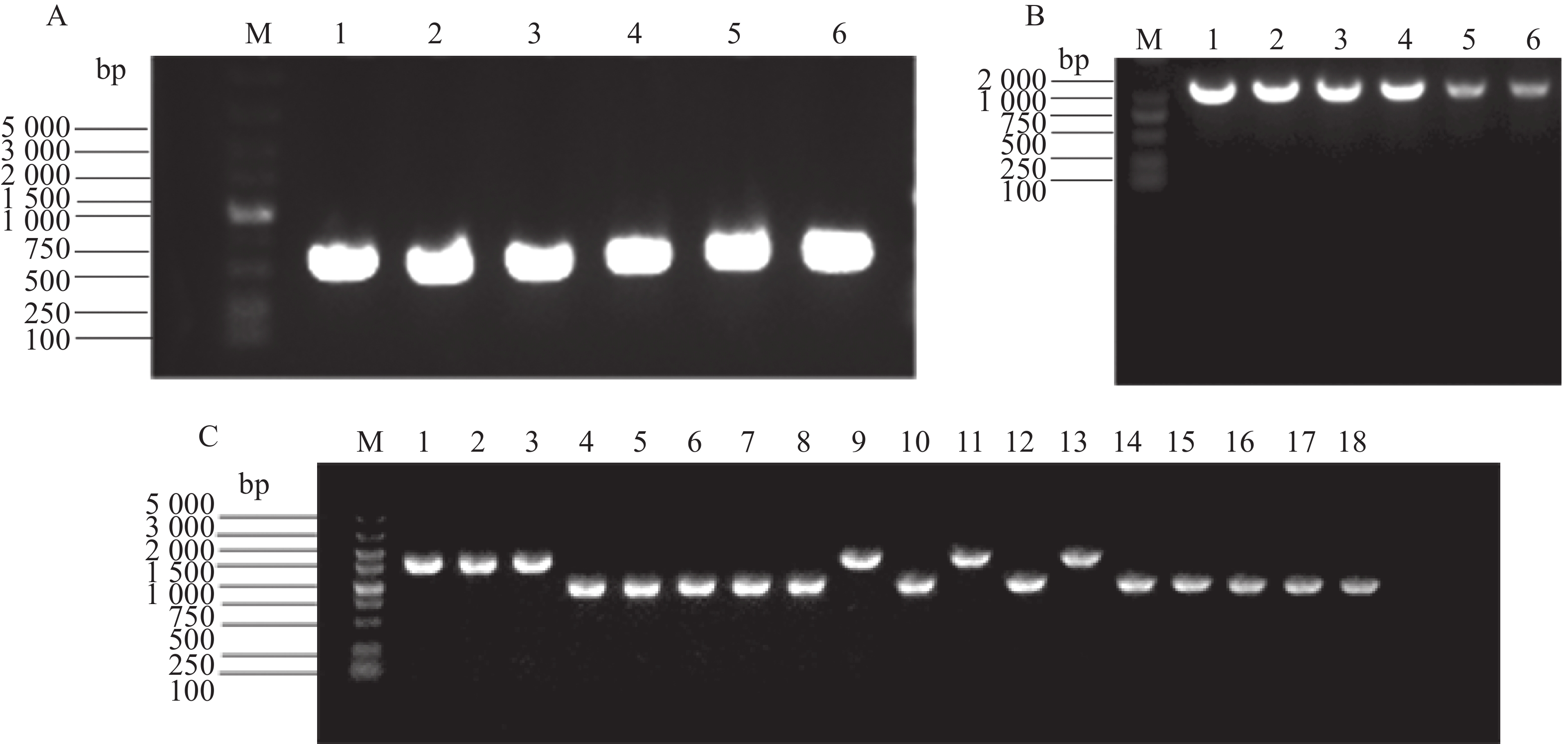
PCR扩增tatD基因上下游片段条带清晰(图2-A),目的基因与自杀质粒经双切后连接构建重组载体,利用tatD-T-F+T-R引物验证条带清晰(图2-B)。将重组载体PDM4-tatD转化至溶藻弧菌HN08155,通过自杀质粒的特点构建tatD基因缺失型菌株,利用tatD1-TF+TR、tatD2-TF+TR、tatD3-TF+TR引物筛选并验证目的基因缺失菌株。结果表明,HN08155菌株中的完整条带在2 000 bp左右,tatD基因缺失菌株条带在1 100 bp左右(图2-C),将PCR产物纯化回收送测,测序结果证明正确。构建成功的基因缺失菌株分别命名为△tatD1、△tatD2、△tatD3、△tatD12、△tatD13、△tatD23、△tatD123。
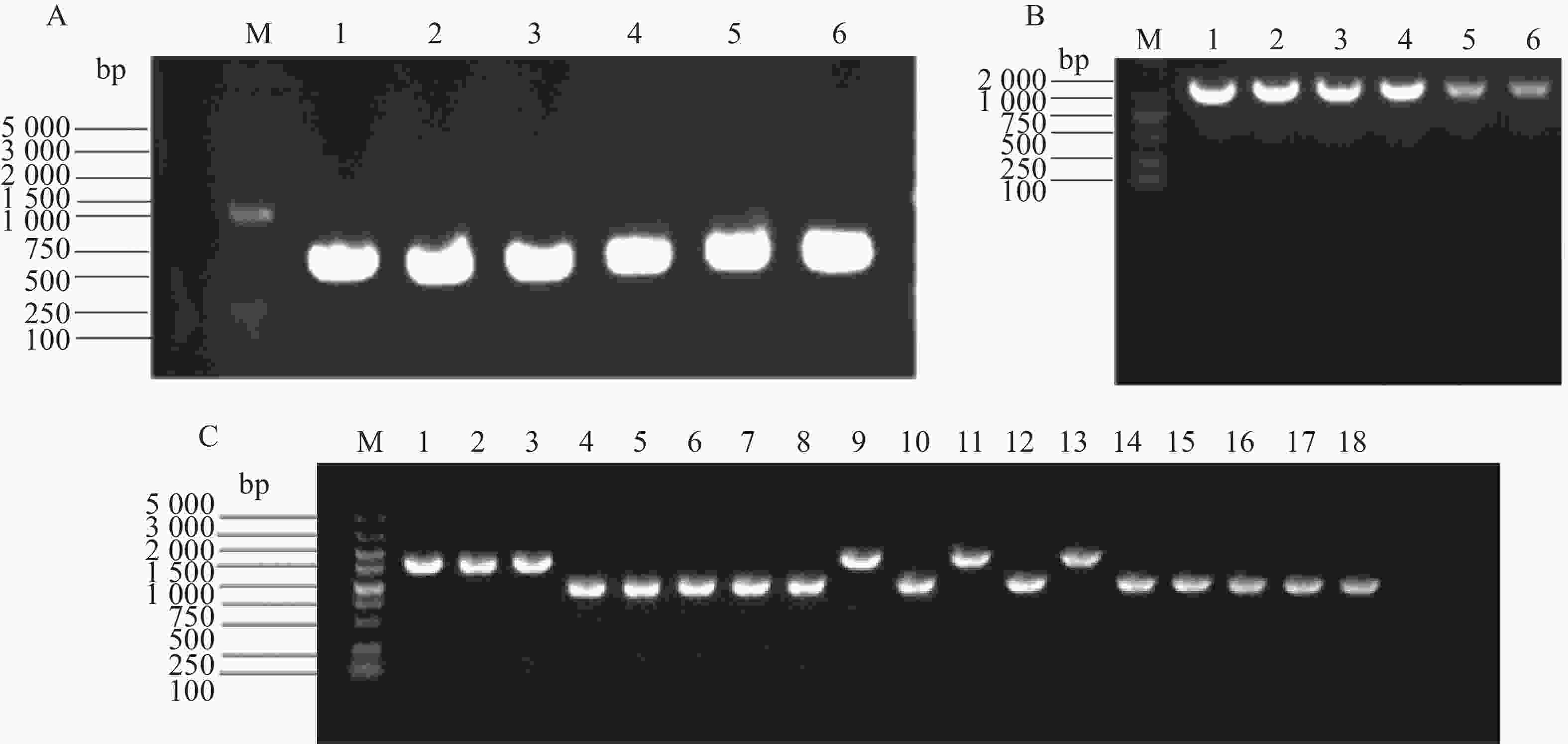
图 2 溶藻弧菌tatD基因克隆与tatD基因缺失菌株筛选
Figure 2. Cloning of tatD gene from Vibrio alginolyticus and screening of tatD gene deletion strains.
-
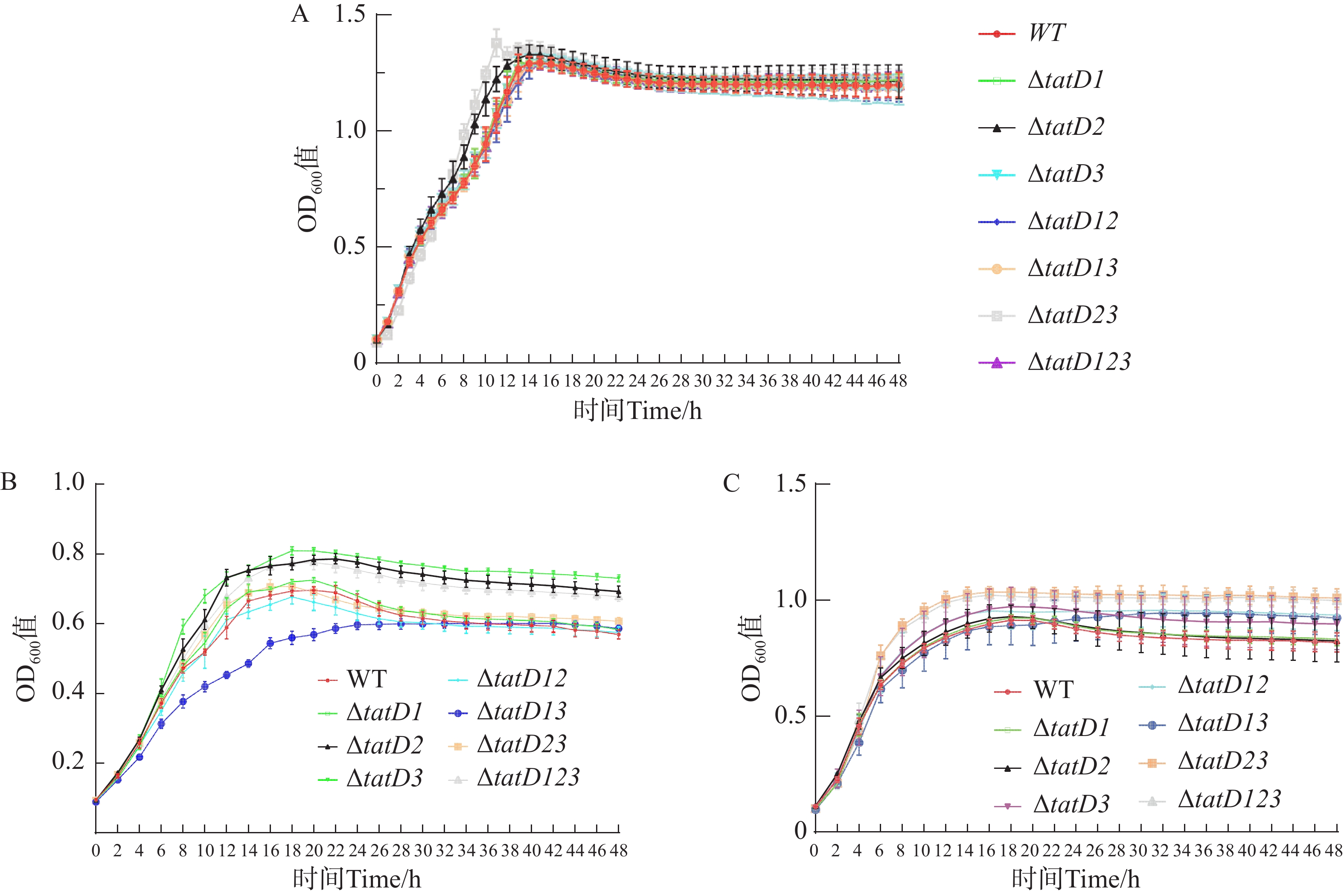
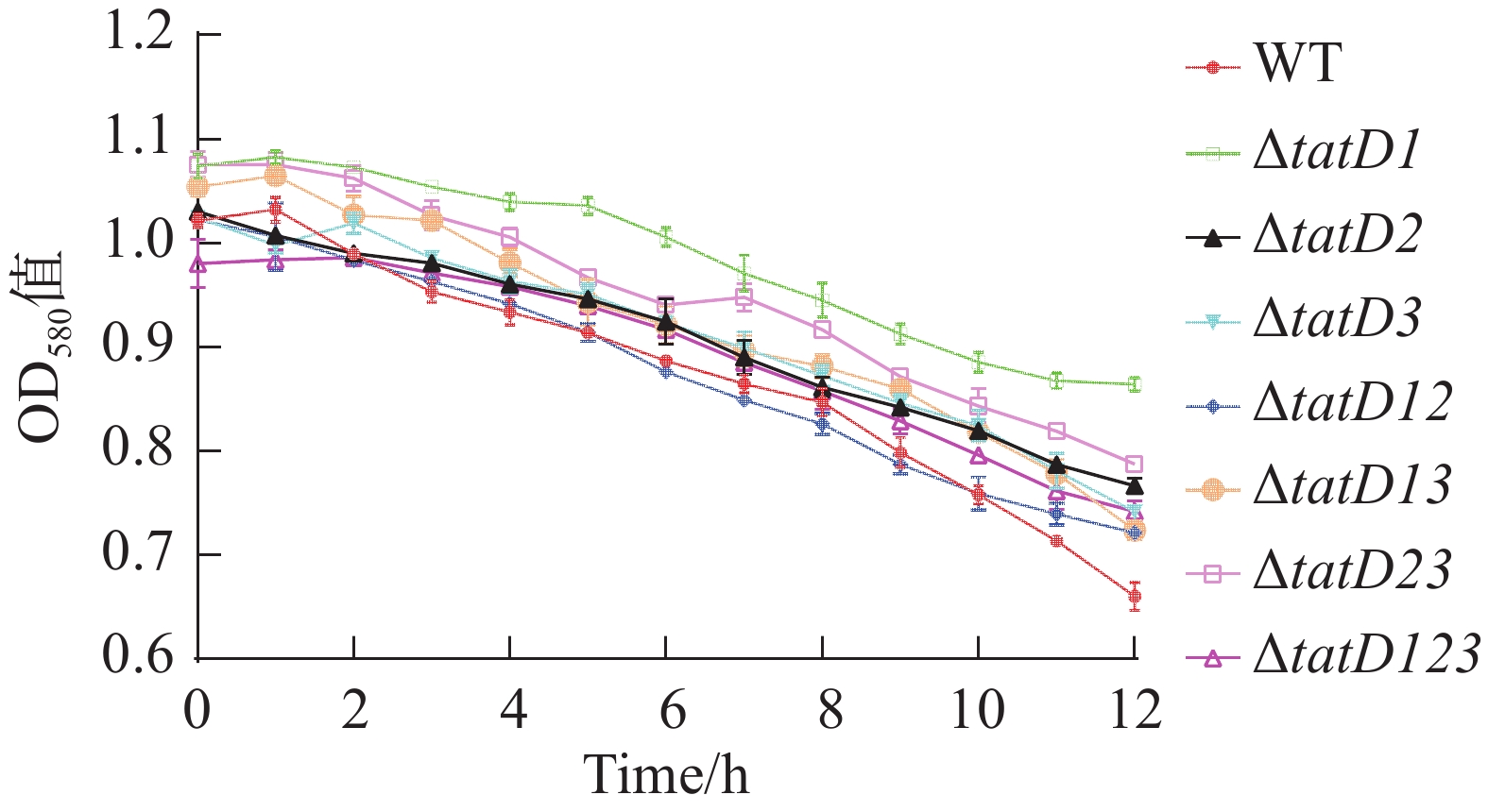
溶藻弧菌HN08155(WT)与tatD基因缺失株在正常营养条件下,48 h内的生长曲线如图3-A所示, 溶藻弧菌野生株与tatD基因的缺失株生长趋势相似。最初各种菌株都处于生长延迟期,在这个时期内,各菌株生长速度较为缓慢;在对数生长期时,细菌生长迅速,并且细菌数量呈指数级增加,在各菌株进入了生长平台期后,细菌增长稳定,生长曲线基本上趋于一致。结果表明,单独缺失tatD任何一个基因,或者缺失两个基因,以及三者同时缺失,均未对溶藻弧菌HN08155的生长特性产生明显影响。在营养条件较低的M9培养基中,各tatD基因缺失菌株的生长能力明显高于野生株。其中△tatD123、△tatD2与△tatD3菌株生长趋势显著上升(P<0.000 1)(图3-B),在培养48 h后,tatD基因缺失株菌体浓度均高于野生株。同时,在不添加Fe2+时,野生菌株与缺失菌株生长差异明显,而在添加Fe2+后,各菌株生长差异不显著,菌株生长趋势与菌体浓度差异缩小(图3-C)。这表明溶藻弧菌tatD基因在不同营养条件下表现出了不一样的功能。
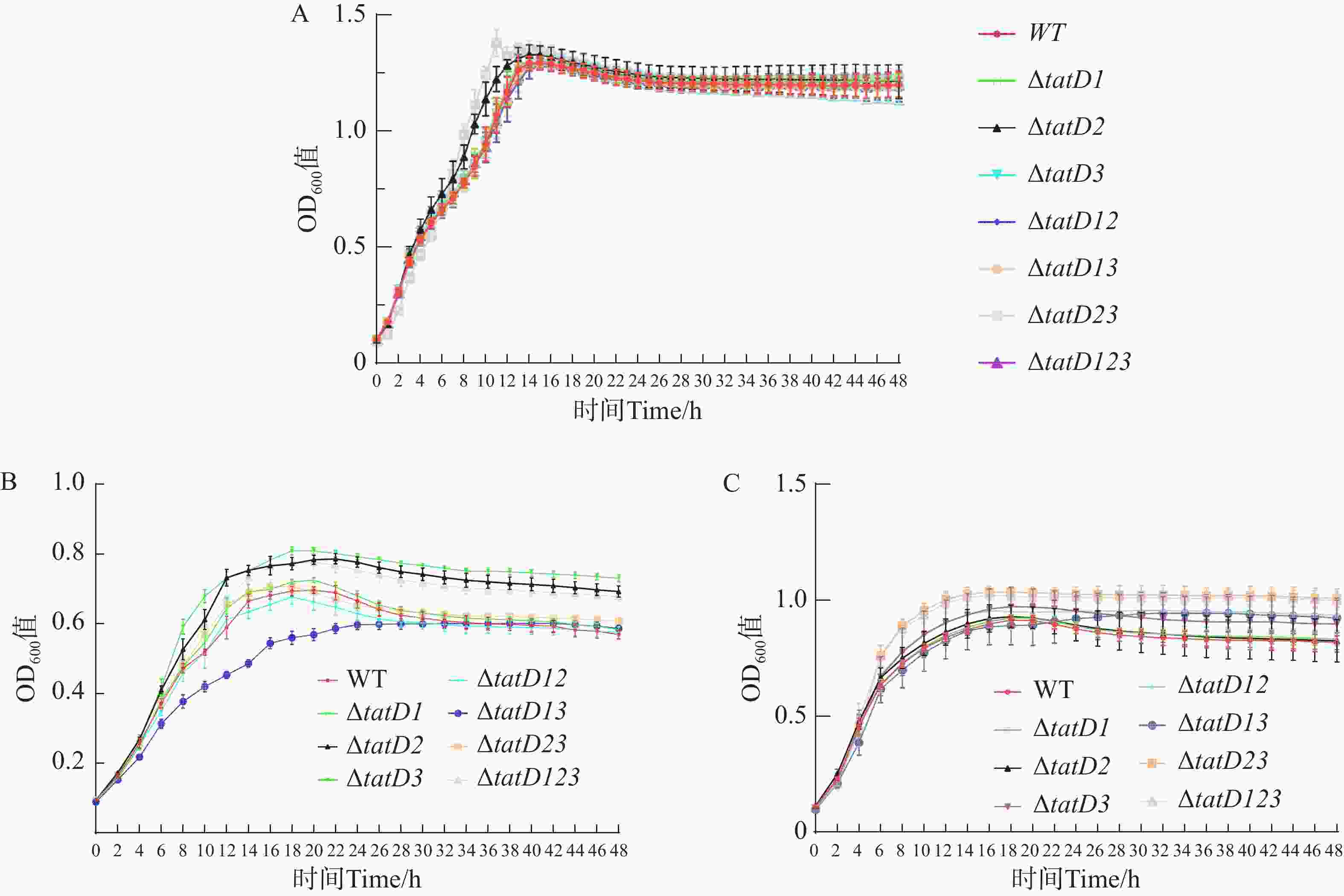
图 3 HN08155野生菌株和tatD基因缺失菌株生长能力测定
Figure 3. Determination of the growth capabilities of wild-type strain HN08155 and tatD gene deletion strains
-
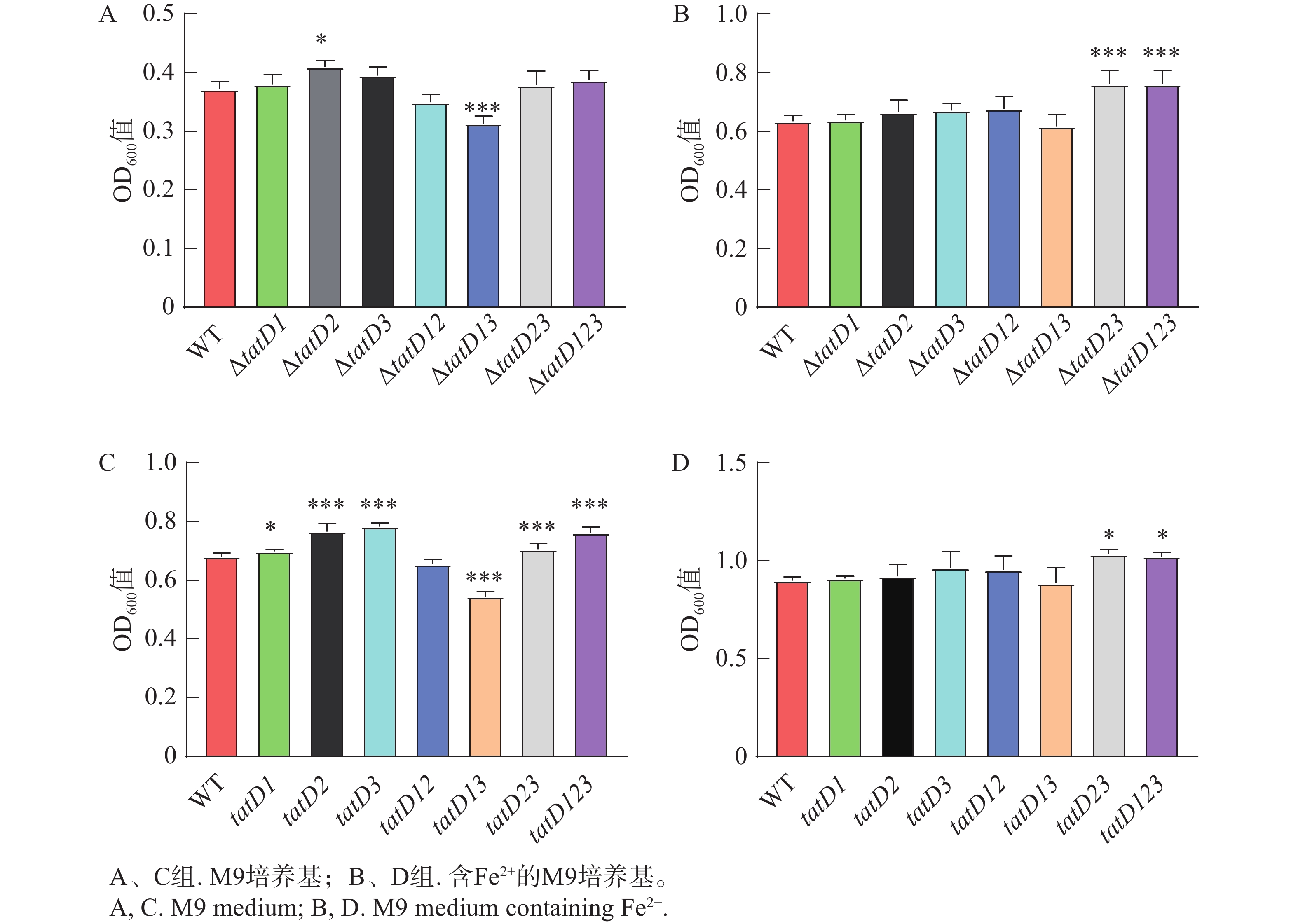
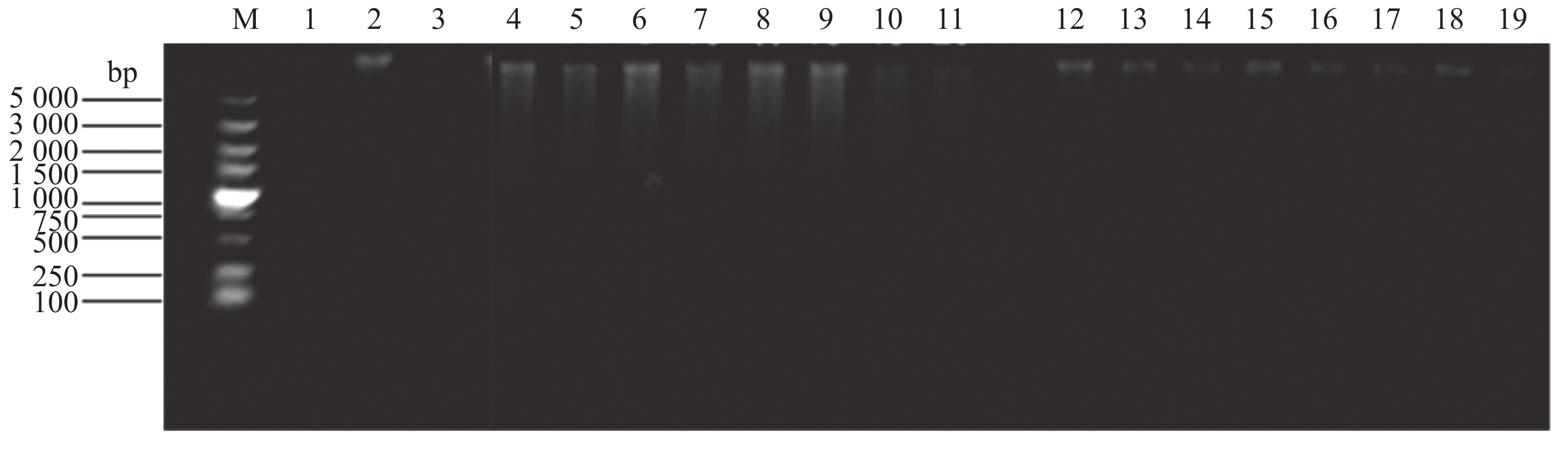
溶藻弧菌野生株与tatD基因缺失株上清液对DNA的降解能力结果见图4、图5,野生株上清液的降解DNA效果明显高于缺失株,而三基因缺失株的降解效果显著下降。△tatD12菌株上清液的降解能力比其他双基因缺失株要弱。在单一基因缺失菌株中,△tatD3菌株的上清液降解能力明显高于其他单基因缺失株。此外,在添加EDTA后,无论是野生株还是基因缺失菌株,均失去降解DNA的能力。结果表明,溶藻弧菌tatD基因的表达提升了其核酸酶活性,表现出对金属离子的需求,与DNase I酶发挥作用时表现相似。其中,tatD2基因降解DNA分子的能力最高。
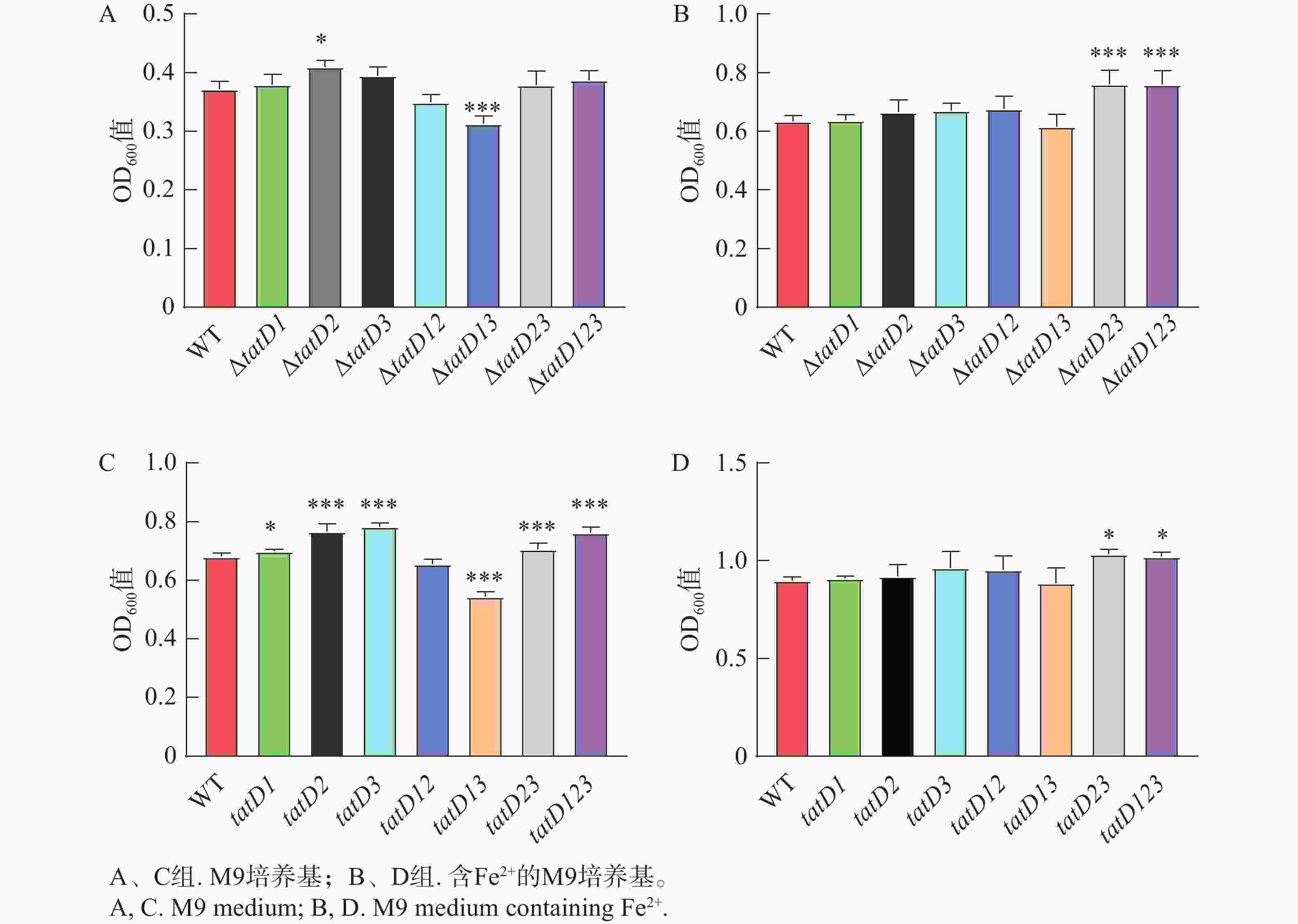
图 4 HN08155野生菌株和tatD基因缺失菌株在M9培养基中培养6 h和8 h的菌体浓度
Figure 4. Bacterial cell density of the wild-type strain HN08155 and the tatD gene knockout strain in M9 medium containing Fe2+ after 6 hours and 8 hours

图 5 培养物上清液降解DNA能力琼脂糖凝胶电泳分析
Figure 5. Analysis of DNA degradation using agarose gel electrophoresis
-
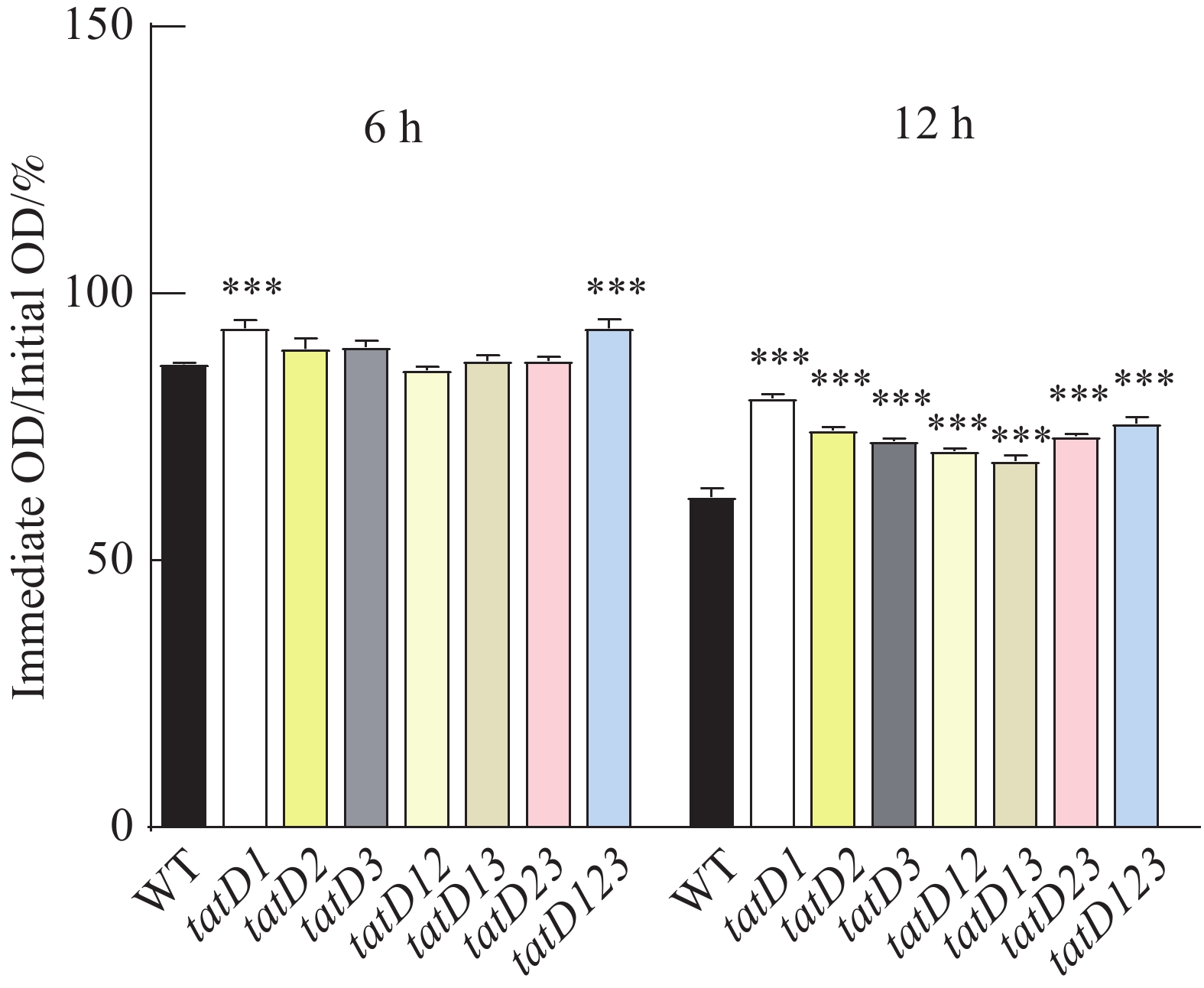
在Triton X-100(体积浓度为0.1%)的诱导下。对溶藻弧菌HN08155(WT)及其tatD缺失菌株进行的自溶活性分析结果如图6所示。图7显示了在6和12 h,测定的野生菌株与tatD缺失菌株的自溶率。在0~6 h内,野生株和tatD基因缺失株的自溶活性曲线呈现缓慢下降趋势,在6 h的时候,两者自溶率之间尚无显著性差异,而在6~12 h,野生株和tatD基因缺失株的自溶活性曲线下降速度加快,12 h的时候,tatD基因缺失株自溶率显著高于野生株。结果表明,与野生型菌株相比,tatD基因缺失株的自溶活性显著降低。
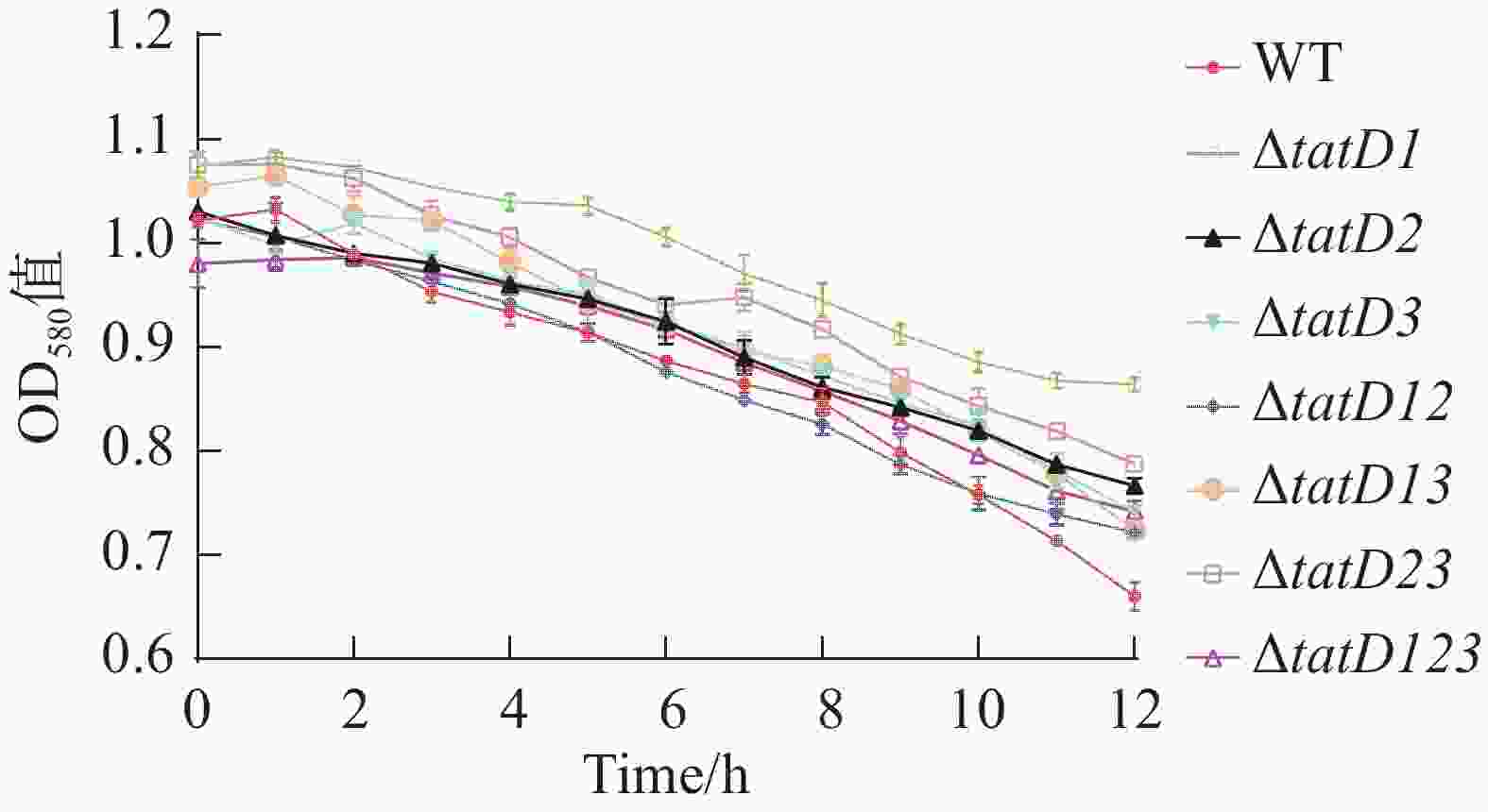
图 6 HN08155和tatD基因敲除菌株的自溶能力曲线图
Figure 6. Self-lysis curve of HN08155 and tatD gene knockout strains
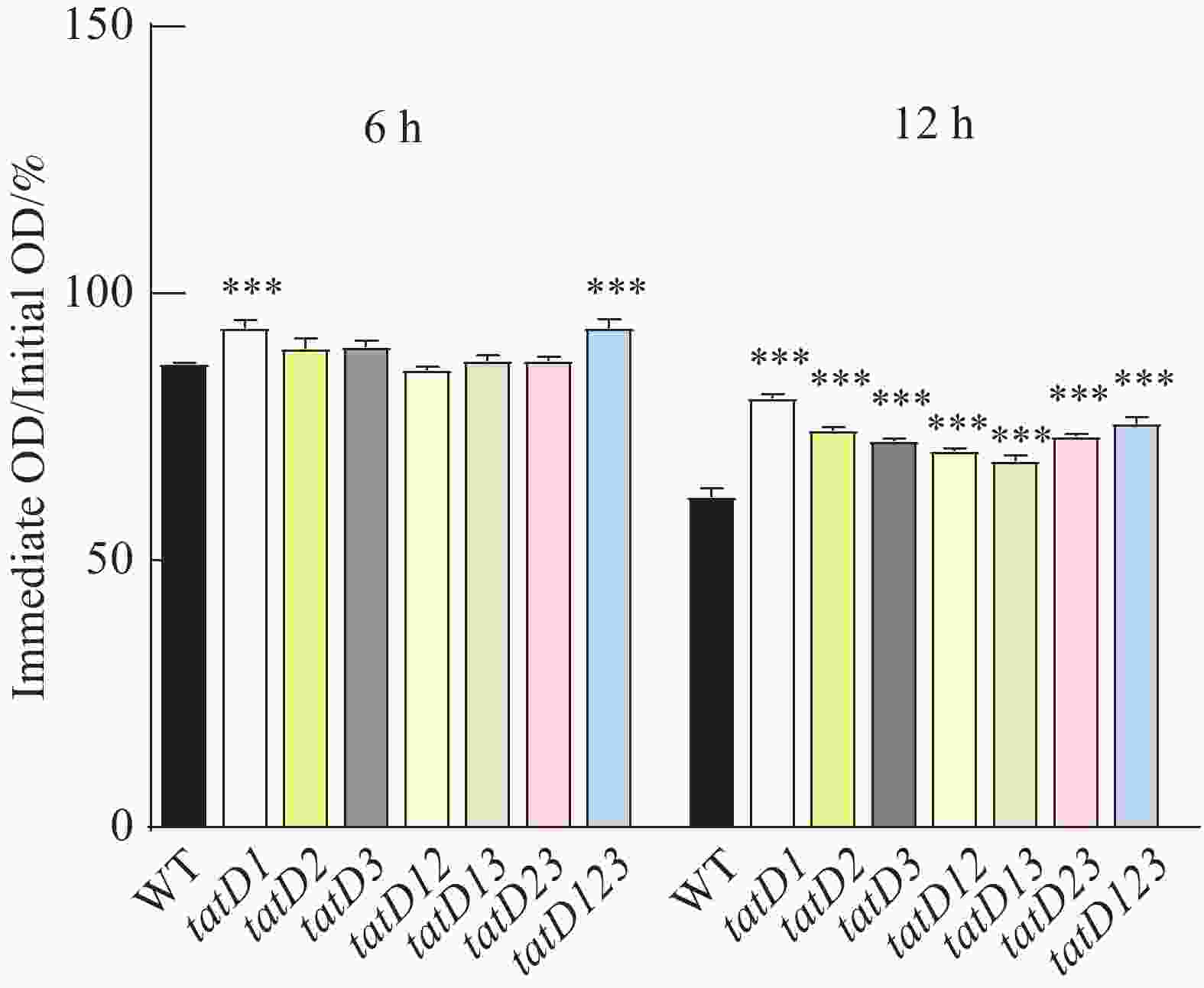
图 7 HN08155和tatD基因敲除菌株的自溶能力对比
Figure 7. Comparison of autolysis ability between HN08155 and tatD gene knockout strains
-
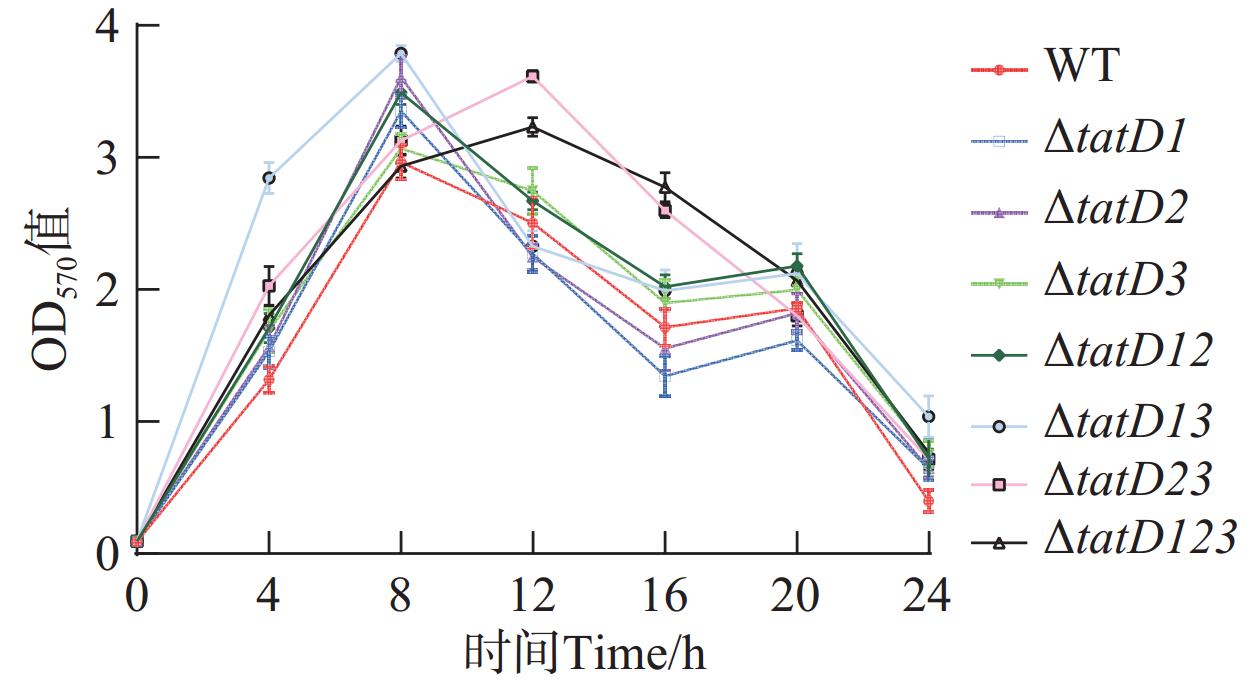
本实验测定了野生株和tatD基因缺失株在24 h内生物被膜形成量(图8)。图9显示了在8 h时野生型和tatD基因缺失株的生物被膜形成量。实验显示tatD基因缺失株生物被膜形成量与生物被膜形成趋势均高于野生株。在这一阶段,各菌株的生物被膜量快速增加,在经过8 h后,除了△tatD13基因双缺失株和△tatD123三基因缺失株外,其他所有菌株都已达到了其生物被膜形成量的峰值。其中溶藻弧菌△tatD13菌株的生物被膜形成量最多;在8~16 h时,除了△tatD13基因双缺失株△tatD123三基因缺失株之外,其他菌株的生物被膜逐渐出现溶解下降的趋势。而在16~20 h的时间段内,所有菌株的生物被膜形成量都呈现了先上升后下降的波动。这种波动趋势的出现可能是后期菌体聚集所造成的,进而导致生物膜形成量曲线出现异常上升现象。在8 h时,△tatD2、△tatD12、△tatD123菌株形成的生物被膜量与厚度、密度均高于野生株。研究结果表明,溶藻弧菌在缺失tatD基因后,其生物被膜形成量增加,而降解生物被膜的能力减弱。这证实了tatD基因具有调控生物被膜形成和抑制生物被膜形成的能力。
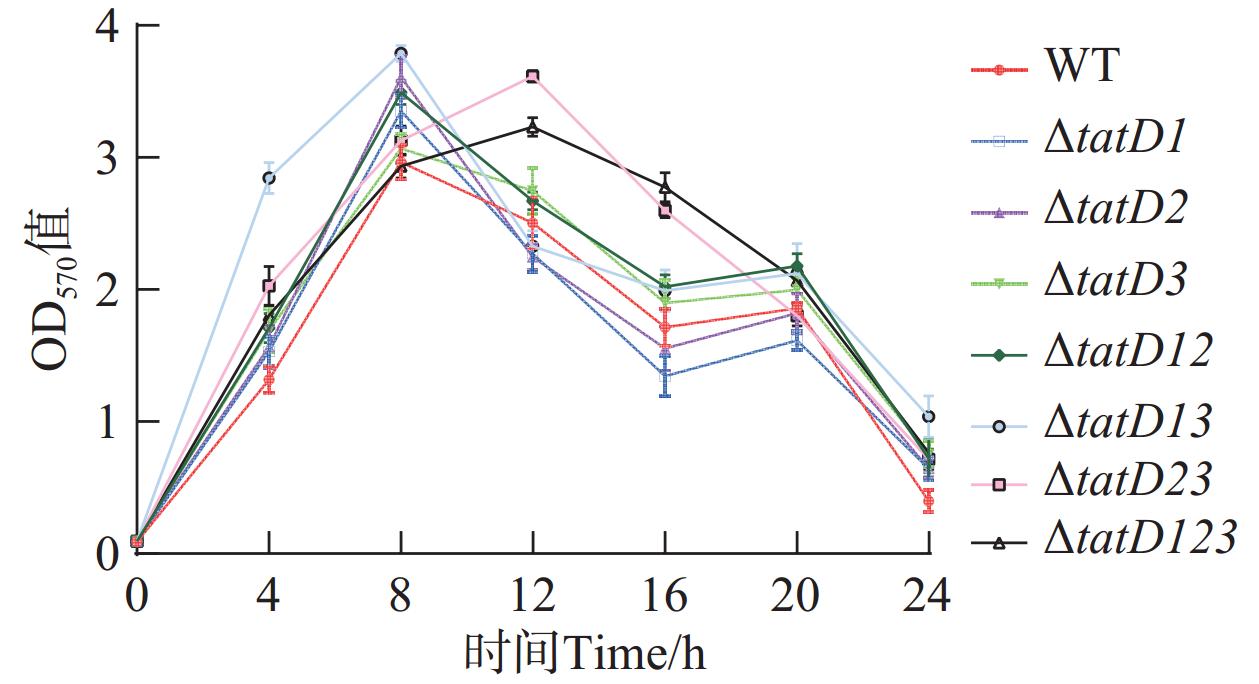
图 8 野生株与tatD基因缺失株生物被膜形成量
Figure 8. Biofilm formation of wild-type (WT) strain and tatD gene deletion mutant
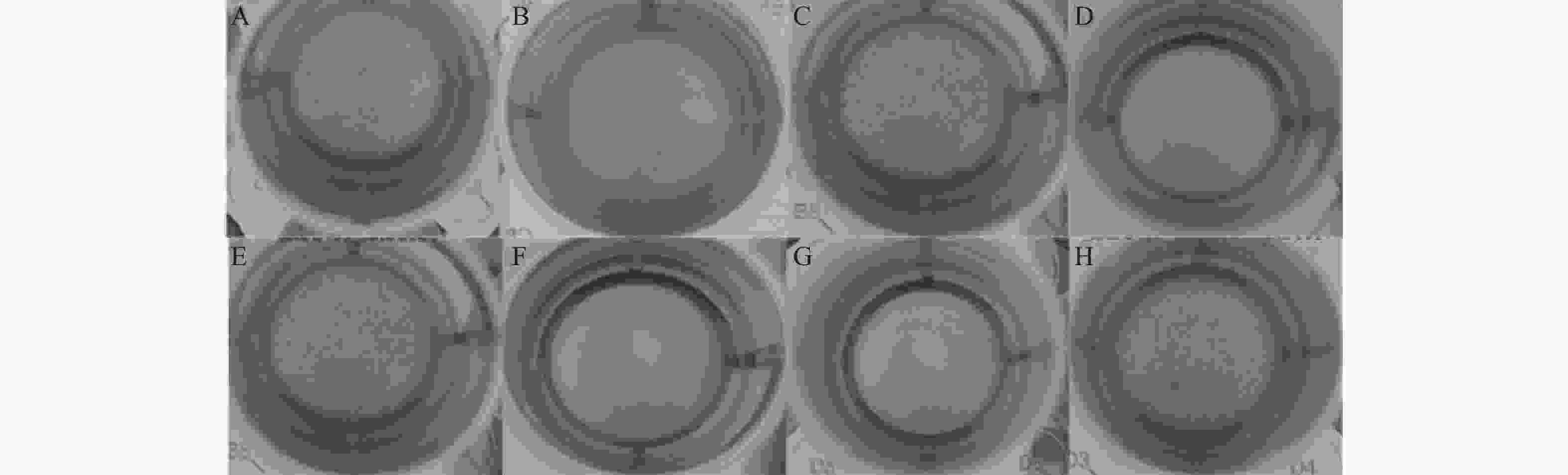
图 9 8 h时野生株与tatD基因缺失株生物被膜形成量
Figure 9. Biofilm formation of the wild-type (WT) strain and a tatD gene deletion mutant after 8 hours
-
为了深入研究溶藻弧菌HN08155菌株中tatD基因的功能和作用机制,本研究构建了tatD基因缺失突变株,并对其进行生物学特性分析。实验结果显示,在正常营养条件下,tatD基因的缺失对溶藻弧菌的生长能力没有影响。一些类似研究结果也表明,tatD基因不参与细菌生长调控,如单增李氏特菌、化脓性隐球菌、肺炎链球菌和迟缓型艾德华氏菌在失去tatD基因后,其生长能力并未受到影响[21]。不过,进一步的实验表明,在营养较为匮乏的环境下,tatD基因敲除菌株相较于野生型菌株表现出了更好的生长能力,各缺失株与野生株相比具有明显的生长趋势上的差异,在添加Fe2+后,各菌株生长差异会显著降低。这种差异可能是由于tatD基因缺失菌株能够形成较野生株更多的生物膜,膜中细菌生长缓慢及被膜的保护能力增强了tatD基因缺失株的生长能力;也有研究结果表明,tatD是铁硫蛋白质控制中心组成成分之一,对于蛋白的输出至关重要[22],在M9培养条件下可能影响了离子相关蛋白的输出,进而影响了菌株的生长;以上实验结果证明了tatD基因并非溶藻弧菌生长核心基因,在正常营养条件下缺失了tatD基因并不会影响溶藻细菌的生长。而在不利的环境条件下,溶藻弧菌可能通过tatD基因调控形成生物被膜,或影响了离子相关蛋白的输出,进而影响了菌株的生长。
在应对不利的环境条件及维持群体稳态中,生物被膜发挥着至关重要的作用。研究发现tatD基因缺失后缺失株生物被膜生成量均高于野生株,其中,△tatD13缺失株形成的生物被膜最多,以上实验结果说明tatD基因参与了生物被膜形成的调控。生物被膜保护性取决于其胞外聚合物(EPS)的组成,由于不同细菌能够产生多种多样的EPS。在这个复杂的过程中,胞外多糖和胞外蛋白差异显得尤为关键[23]。与胞外多糖、蛋白不同,eDNA是生物被膜网状分子结构的组成成分。本研究团队在上清液降解小牛胸腺DNA的实验中,分析野生株与缺失株对于DNA分子的降解能力,结果发现,tatD基因的存在增强溶藻弧菌对于DNA分子的降解能力,同时对于金属离子的需求,与DNase I酶发挥作用时表现出了相似性。
eDNA从细胞内部释放到胞外的方式之一就是自溶。在深入研究tatD基因时,发现tatD DNases 参与多种微生物的程序性凋亡[24-26],推测tatD可能参与了细菌自溶,从而释放eDNA。对此,本研究开展了溶藻弧菌自溶性分析实验,结果表明,溶藻弧菌在缺失tatD基因后自身自溶活性受到了影响,自溶能力下降。tatD基因参与了细菌自溶,但仍没有直接证据表明溶藻弧菌通过菌体自溶释放eDNA进而促进细菌生物被膜的形成。
-
本研究通过构建tatD基因缺失菌株,初步探究了tatD基因在溶藻弧菌生长与生物被膜形成等方面的功能,结果表明,在正常营养条件下缺失tatD基因不会影响细菌的生长,在营养贫瘠的条件下,tatD基因可能参与了细菌的生长调节。同时,tatD基因也参与溶藻弧菌生物被膜形成与细菌自溶性的调节,缺失tatD基因,生物被膜形成量增加;自溶性下降。这些结果为研究tatD基因在弧菌致病机制中的功能奠定了基础。
-
摘要: 溶藻弧菌是一种海洋常见革兰氏阴性细菌,广泛存在于水产养殖环境中,它对养殖鱼类、贝类和对虾都具有极强的侵染性和致病性,其导致的弧菌病给养殖业造成了巨大的经济损失。tatD基因编码具有核酸酶活性的蛋白,其功能与细菌生物被膜形成及毒力调控相关。本研究利用基因敲除技术研究了溶藻弧菌HN08155菌株的tatD基因在菌体生长与生物被膜形成过程中的功能。研究结果表明,溶藻弧菌HN08155菌株含有3个tatD基因,在正常营养条件下tatD基因不影响细菌生长,在低营养条件下对细菌的生长具有调节作用;分泌到胞外的TatD蛋白具有核酸酶活性,野生株培养物上清液降解DNA的能力高于tatD基因缺失菌株;在0.1% Triton X-100 的诱导下,tatD基因缺失株自溶率显著高于野生株;tatD基因缺失后,溶藻弧菌生物被膜量与厚度显著增加。Abstract: Vibrio alginolyticus is a common marine Gram-negative bacterium widely found in aquaculture environments. It exhibits strong infectivity and pathogenicity toward cultured fish, shellfish, and shrimp, causing significant economic losses in the aquaculture industry due to vibriosis. The tatD gene encodes a protein with nuclease activity, which is related to the formation of bacterial biofilms and the regulation of virulence. An attempt was made to investigate the role of tatD gene in the growth and biofilm formation of V. alginolyticus strain HN08155 by utilizing gene knockout technology. The results indicate that the strain HN08155 contains three tatD genes. Under normal nutrient conditions, the tatD gene does not affect bacterial growth, whereas it plays a regulatory role in bacterial growth under low-nutrient conditions. The extracellular tatD protein demonstrates nuclease activity, with the wild-type strain showing a greater capacity for DNA degradation in the culture supernatant compared to the tatD knockout strain. Induction with 0.1% Triton X-100 significantly increased the autolysis rate of the tatD knockout strain compared to the wild-type strain. Additionally, the absence of the tatD gene resulted in a significant increase in both the quantity and thickness of the biofilm formed by V. alginolyticus.
-
Key words:
- Vibrio alginolyticus /
- tatD gene /
- gene knockout /
- bacterial biofilm /
- autolysis
-
图 2 溶藻弧菌tatD基因克隆与tatD基因缺失菌株筛选
A.tatD基因的上游和下游片段(M,5000 bp;泳道1~2,tatD1;泳道3~4,tatD2;泳道5~6,tatD3);B.重组载体的构建(M,2 000 bp Marker;泳道1~2,tatD1;泳道3~4,tatD2;泳道5~6,tatD3);C.tatD基因缺失菌株(M.5 000 bp Marker;泳道1~3,野生型(WT);泳道4~6,△tatD1、△tatD2、△tatD3;泳道7~9,△tatD12;泳道10~12,△tatD13;泳道13~15,△tatD23;泳道16~18,△tatD123;泳道1、4、7、10、13、16,使用tatD1引物扩增;泳道2、5、8、11、14、17,使用tatD2引物扩增;泳道3、6、9、12、15、18,使用tatD3引物扩增)。
Fig. 2 Cloning of tatD gene from Vibrio alginolyticus and screening of tatD gene deletion strains.
A. Upstream and downstream fragments of the tatD gene (M, 5 000 bp marker; Lanes 1−2, tatD1; Lanes 3−4, tatD2; Lanes 5−6, tatD3); B. Construction of the recombinant vector (M, 2000 bp marker; Lanes 1−2, tatD1; Lanes 3−4, tatD2; Lanes 5−6, tatD3); C. tatD gene deletion strains (M, 5 000 bp marker; Lanes 1−3, WT; Lanes 4−6, △tatD1, △tatD2, △tatD3; Lanes 7−9, △tatD12; Lanes 10−12, △tatD13; Lanes 13−15,△tatD23; Lanes 16−18, △tatD123; Lanes 1, 4, 7, 10, 13, 16, tatD1 primers; Lanes 2, 5, 8, 11, 14, 17, tatD2 primers; Lanes 3, 6, 9, 12, 15, 18, tatD3 primers).
图 3 HN08155野生菌株和tatD基因缺失菌株生长能力测定
A. LB培养基;B. M9培养基;C.含Fe2+的M9培养基。
Fig. 3 Determination of the growth capabilities of wild-type strain HN08155 and tatD gene deletion strains
A. LB medium; B. M9 medium; C. M9 medium supplemented with Fe2+.
图 4 HN08155野生菌株和tatD基因缺失菌株在M9培养基中培养6 h和8 h的菌体浓度
注: "*"表示P< 0.05,指野生型菌株与敲除菌株自溶能力存在显著差异;"***"表示P<0.0001,指野生型菌株与敲除菌株自溶能力存在极显著差异。
Fig. 4 Bacterial cell density of the wild-type strain HN08155 and the tatD gene knockout strain in M9 medium containing Fe2+ after 6 hours and 8 hours
Note: “*” indicates P < 0.05, denoting a significant difference in autolytic capacity between the wild-type strain and the knockout strain. “***” indicates P < 0.001, denoting a highly significant difference in autolytic capacity between the wild-type strain and the knockout strain.
图 5 培养物上清液降解DNA能力琼脂糖凝胶电泳分析
泳道1.DNA分子量标准(5000 bp);泳道2.未添加DNase I酶;泳道3.添加DNase I酶;泳道4~11、12~19.△tatD123、△tatD13、△tatD12、△tatD23、△tatD1、△tatD2、△tatD3及野生型(WT)菌株上清液;泳道12~19中均添加EDTA。
Fig. 5 Analysis of DNA degradation using agarose gel electrophoresis
Lane 1. DNA Marker 5000; Lane 2. Without DNase I enzyme; Lane 3. With DNase I enzyme; Lanes 4−11, 12−19. Supernatants from △tatD123, △tatD13, △tatD12, △tatD23, △tatD1, △tatD2, △tatD3, and WT; EDTA was added to Lanes 12−19.
图 6 HN08155和tatD基因敲除菌株的自溶能力曲线图
Fig. 6 Self-lysis curve of HN08155 and tatD gene knockout strains
图 7 HN08155和tatD基因敲除菌株的自溶能力对比
注:“***”表示P<0.001,野生株与缺失株自溶能力对比差异极显著。
Fig. 7 Comparison of autolysis ability between HN08155 and tatD gene knockout strains
Note: "***" indicates P<0.001, denoting a highly significant difference in autolytic capacity between the wild-type strain and the knockout strain.
图 8 野生株与tatD基因缺失株生物被膜形成量
Fig. 8 Biofilm formation of wild-type (WT) strain and tatD gene deletion mutant
图 9 8 h时野生株与tatD基因缺失株生物被膜形成量
A.WT菌株;B.△tatD1菌株;C.△tatD2菌株;D.△tatD3菌株;E.△tatD12菌株;F.△tatD13菌株;G.△tatD23菌株;H.△tatD123菌株。
Fig. 9 Biofilm formation of the wild-type (WT) strain and a tatD gene deletion mutant after 8 hours
A. WT strain;B. △tatD1 strain;C. △tatD2 strain;D. △tatD3 strain;E. △tatD12 strain;F. △tatD13 strain;G. △tatD23 strain;H. △tatD123 strain.
表 1 PCR扩增所使用的引物
Table 1 Primers used in PCR
引物
Primer序列
SequencetatD1-T-F ATGAAGCTCTTTGATACCCA tatD1-T-R TTATTCACATATCGAGAAAG tatD1-UF GAAGATCTTCAGAACCAAGCAAACTTAGCA tatD1-UR CCATAACTGAGAGGCAATCGAAGTCACCTTGGAATACATC tatD1-DF GATGTATTCCAAGGTGACTTCGATTGCCTCTCAGTTATGG tatD1-DR CCCTCGAGGGACCGTAGTTGATAACGTTAG tatD1-TF GCATTAACGTTGCAACTGCG tatD1-TR GAATGCGAAGATGAAACCGC tatD2-T-F ATGTTCGTAGATTCTCATTG tatD2-T-R TCATCGCAAAAAAAGATTTC tatD2-UF GAAGATCTTCAACTGAATATGGGCTCTCCG tatD2-UR GCACGATTTGAGTTGCGCAATCCACATTGATGTGGAGATC tatD2-DF GATCTCCACATCAATGTGGATTGCGCAACTCAAATCGTGC tatD2-DR CCCTCGAGGGGTAGCCTAAAGGTTAGCACC tatD2-TF TTACCCACAATCAGGAGCGT tatD2-TR CTGATACTGGAAGCATCAGC tatD3-T-F ATGATCGATACCCACGCGCA tatD3-T-R TTAAATATTAAACAGCTTTT tatD3-UF GAAGATCTTCCGATTGCTGGAACGCCTAGA tatD3-UR AGTTGCGACATCGACATCACACTACTTCATCGCGGTCGTT tatD3-DF AACGACCGCGATGAAGTAGTGTGATGTCGATGTCGCAACT tatD3-DR CCCTCGAGGGGACGTAACCACACTTTGTGC tatD3-TF TCTTCATCGATGATGCCGTC tatD3-TR GCCATTCCGATGTGCTTACT pDM4-TF CACAGGAACACTTAACGGCT pDM4-TR TCCTGTTCAGCTACTGACGG  下载: 导出CSV
下载: 导出CSV
-
[1] 赵伟, 王雷, 刘梅, 等. 副溶血弧菌对凡纳滨对虾肝胰腺抗氧化酶活性和基因表达的影响[J]. 中国水产科学, 2017, 24(6): 1261−1270. https://doi.org/10.3724/SP.J.1118.2017.16221 doi: 10.3724/SP.J.1118.2017.16221 [2] Park M Y, Kim H J, Choi S T, et al. Pathogenic factors of Vibrio spp. isolated from seawater of gwangan beach in Busan [J]. Fisheries and Aquatic Sciences, 2002, 5(3): 178−182. https://doi.org/10.5657/fas.2002.5.3.178 doi: 10.5657/fas.2002.5.3.178 [3] Sakuma M, Nishikawa S, Inaba S, et al. Structure of the periplasmic domain of SflA involved in spatial regulation of the flagellar biogenesis of Vibrio reveals a TPR/SLR-like fold [J]. The Journal of Biochemistry, 2019, 166(2): 197−204. https://doi.org/10.1093/jb/mvz027 doi: 10.1093/jb/mvz027 [4] Ono H, Takashima A, Hirata H, et al. The MinD homolog FlhG regulates the synthesis of the single polar flagellum of Vibrio alginolyticus [J]. Molecular Microbiology, 2015, 98(1): 130−141. https://doi.org/10.1111/mmi.13109 doi: 10.1111/mmi.13109 [5] Hormansdorfer S, Wentges H, Neugebaur-Buchler K, et al. Isolation of Vibrio alginolyticus from seawater aquaria [J]. International Journal of Hygiene and Environmental Health, 2000, 203(2): 169−175. https://doi.org/10.1078/S1438-4639(04)70024-3 doi: 10.1078/S1438-4639(04)70024-3 [6] Xie J S, Bu L F, Jin S, et al. Outbreak of vibriosis caused by Vibrio harveyi and Vibrio alginolyticus in farmed seahorse Hippocampus kuda in China [J]. Aquaculture, 2020, 523: 735168. https://doi.org/10.1016/j.aquaculture.2020.735168 doi: 10.1016/j.aquaculture.2020.735168 [7] Liang H Y, Xia L Q, Wu Z H, et al. Expression, purification and antibody preparation of flagellin FlaA from Vibrio alginolyticus strain HY9901 [J]. Letters in Applied Microbiology, 2010, 50(2): 181−186. https://doi.org/10.1111/j.1472-765X.2009.02768.x doi: 10.1111/j.1472-765X.2009.02768.x [8] 权太淑, 李薇, 杨喜玲. 自腹泻患者中分离溶藻性弧菌及其病原性的探讨[J]. 中国公共卫生, 1985, 4(5): 15−18. [9] Balebona M C, Andreu M J, Bordas M A, et al. Pathogenicity of Vibrio alginolyticus for cultured gilt-head sea bream (Sparus aurata L.)[J]. Applied and Environmental Microbiology, 1998, 64(11): 4269−4275. [10] Høiby N, Bjarnsholt T, Givskov M, et al. Antibiotic resistance of bacterial biofilms [J]. International Journal of Antimicrobial Agents, 2010, 35(4): 322−332. https://doi.org/10.1016/j.ijantimicag.2009.12.011 doi: 10.1016/j.ijantimicag.2009.12.011 [11] Battin T J, Kaplan L A, Denis Newbold J, et al. Contributions of microbial biofilms to ecosystem processes in stream mesocosms [J]. Nature, 2003, 426(6965): 439−442. https://doi.org/10.1038/nature02152 doi: 10.1038/nature02152 [12] Jhelum H, Sori H, Sehgal D. A novel extracellular vesicle-associated endodeoxyribonuclease helps Streptococcus pneumoniae evade neutrophil extracellular traps and is required for full virulence [J]. Scientific Reports, 2018, 8(1): 7985. https://doi.org/10.1038/s41598-018-25865-z doi: 10.1038/s41598-018-25865-z [13] Zhao Y X, Xiang X, Liu X P, et al. The biochemical characterization of a TatD nuclease from Thermus thermophilus [J]. Protein Expression and Purification, 2024, 223: 106557. https://doi.org/10.1016/j.pep.2024.106557 doi: 10.1016/j.pep.2024.106557 [14] Yu T, Sun Z, Cao X Y, et al. Identification and characterization of TatD DNase in planarian Dugesia japonica and its antibiofilm effect [J]. Environmental Research, 2024, 251: 118534. https://doi.org/10.1016/j.envres.2024.118534 doi: 10.1016/j.envres.2024.118534 [15] Wexler M, Sargent F, Jack R L, et al. TatD is a cytoplasmic protein with DNase activity: no requirement for TatD family proteins in sec-independent protein export [J]. Journal of Biological Chemistry, 2000, 225(22): 16717−16722. https://doi.org/10.1074/jbc.M000800200 doi: 10.1074/jbc.M000800200 [16] Chang Z G, Jiang N, Zhang Y Y, et al. The TatD-like DNase of Plasmodium is a virulence factor and a potential malaria vaccine candidate [J]. Nature Communications, 2016, 7: 11537. https://doi.org/10.1038/ncomms11537 doi: 10.1038/ncomms11537 [17] Nag S, Banerjee C, Goyal M, et al. Plasmodium falciparum Alba6 exhibits DNase activity and participates in stress response [J]. iScience, 2024, 27(4): 109467. https://doi.org/10.1016/j.isci.2024.109467 doi: 10.1016/j.isci.2024.109467 [18] Chen L L, Shen D Y, Sun N N, et al. Phytophthora sojae TatD nuclease positively regulates sporulation and negatively regulates pathogenesis [J]. Molecular Plant-Microbe Interactions, 2014, 27(10): 1070−1080. https://doi.org/10.1094/MPMI-05-14-0153-R doi: 10.1094/MPMI-05-14-0153-R [19] Lovett S T. The DNA exonucleases of Escherichia coli [J]. EcoSal Plus, 2011, 4(2): 19−25. https://doi.org/10.1128/ecosalplus.4.4.7 doi: 10.1128/ecosalplus.4.4.7 [20] Lee K Y, Cheon S H, Kim D G, et al. A structural study of TatD from Staphylococcus aureus elucidates a putative DNA-binding mode of a Mg2+-dependent nuclease [J]. IUCrJ, 2020, 7(3): 509−521. https://doi.org/10.1107/S2052252520003917 doi: 10.1107/S2052252520003917 [21] Matos C F R O, Di Cola A, Robinson C. TatD is a central component of a tat translocon-initiated quality control system for exported FeS proteins in Escherichia coli [J]. Embo Reports, 2009, 10(5): 474−479. https://doi.org/10.1038/embor.2009.34 doi: 10.1038/embor.2009.34 [22] Lyu P C, Li H Q, Xue J Y, et al. Synthesis and biological evaluation of novel luteolin derivatives as antibacterial agents [J]. European Journal of Medicinal Chemistry, 2009, 44(2): 908−914. https://doi.org/10.1016/j.ejmech.2008.01.013 doi: 10.1016/j.ejmech.2008.01.013 [23] Li J P, Chen Y X, Qi J, et al. Characterization of EPS subfractions from a mixed culture predominated by partial-denitrification functional bacteria [J]. Water Research X, 2024, 24: 100250. https://doi.org/10.1016/j.wroa.2024.100250 doi: 10.1016/j.wroa.2024.100250 [24] Wang Q, Liu Q, Ma Y, et al. LuxO controls extracellular protease, haemolytic activities and siderophore production in fish pathogen Vibrio alginolyticus [J]. Journal of Applied Microbiology, 2007, 103(5): 1525−1534. https://doi.org/10.1111/j.1365-2672.2007.03380.x doi: 10.1111/j.1365-2672.2007.03380.x [25] Bosedasgupta S, Das B B, Sengupta S, et al. The caspase-independent algorithm of programmed cell death in Leishmania induced by baicalein: the role of LdEndoG, LdFEN-1 and LdTatD as a DNA 'degradesome'[J]. Cell Death & Differentiation, 2008, 15(10): 1629−1640. https://doi.org/10.1038/cdd.2008.85 doi: 10.1038/cdd.2008.85 [26] Gannavaram S, Debrabant A. Involvement of TatD nuclease during programmed cell death in the protozoan parasite Trypanosoma brucei [J]. Molecular Microbiology, 2012, 83(5): 926−935. https://doi.org/10.1111/j.1365-2958.2012.07978.x doi: 10.1111/j.1365-2958.2012.07978.x -

 点击查看大图
点击查看大图
计量
- 文章访问数: 516
- HTML全文浏览量: 285
- PDF下载量: 53
- 被引次数: 0
