


-
香味是稻米重要的食味品质性状和增值指标之一,因此,香味浓郁的香稻品种在市场上深受消费者喜爱,如日本的‘宫香’、美国的‘Jasmine85’和‘Della’等都是世界颇有名气的香稻品种[1]。香稻品种除了根以外,其余各部位均能散发出香味。香稻挥发性代谢物有300多种,包括醛类、酮类、酯类、酸类、醇类、烃类及杂环化合物等,其中杂环类和醛醇类在香米中积累丰富[2-3]。研究表明,2-乙酰基-1-吡咯啉(2-AP)是香稻的主要香味物质[4],然而,对2-AP之外的其他挥发性代谢物研究不足,且相应的合成基因尚未充分解析。
香叶醇是一种高价值单萜化合物,具有玫瑰花香,在植物花器官中含量丰富,如天竺葵(Pelargonium hortorum Bailey)、蜂香薄荷(Mentha canadensis Linnaeus)、玫瑰(Rosa hybrid)等[5]。香叶醇具有广泛的生理活性,如清除过氧化物自由基能力、抗真菌、抗病毒等[6-7];对于多种疾病有治疗作用,如溃疡、血栓、动脉硬化和免疫缺失等[8]。此外,香叶醇还能促进细胞更新和自我修复,提高细胞的抗氧化能力,被广泛用作香化日用产品,如作为玫瑰系精油的主剂、化妆品、护肤品等[9]。然而,香叶醇在植物中含量很低,通过提取天然香叶醇,成本高昂[10]。水稻(Oryza sativa L.)是一种重要的经济作物和模式作物,产量高,易于栽培管理,并拥有完整的基因组信息、成熟的代谢组体系及高效率的转化体系。有研究表明,三叶期的水稻中检测到8种单萜倍半萜类化合物,包括芳樟醇(16.2±7.6) ng∙g−1,香叶醇(797.4±6.9) ng∙g−1和反式橙花叔醇(
1042.0 ±252.4) ng∙g−1等[11],然而香叶醇合成酶功能尚未解析。研究表明,香叶醇合成酶主要包括2个家族,即定位于质体上的TPS家族和NUDIX家族。其中,TPS家族是通过催化由胞质溶胶中的甲羟戊酸(MVA)途径和质体中的2-C-甲基-D-赤藓糖醇-4-磷酸(MEP)途径生成的香叶基二磷酸(GPP)直接合成香叶醇[12],如罗勒(Ocimum basilicum L.)ObGES[13]、柑橘(Citrus reticulata Blanco)CitTPS16[14]及茶树(Camellia sinensis)的CsTPS1[15];而NUDIX家族通过水解GPP的一个磷酸基团生成香叶基磷酸(GP),再由植物中的非特异磷酸酶水解产生香叶醇,如玫瑰RhNUDIX1[16]、天竺葵PgNdx1[17]等。本研究以已报道的香叶醇合成酶基因为靶标,对水稻基因组进行同源序列比对分析,结合组织特异性表达谱、蛋白质理化性质分析、系统发育树分析和亚细胞定位实验,共同筛选水稻香叶醇合成酶基因。研究结果可为香叶醇合成提供新的基因资源,同时为实现在水稻胚乳中高效合成香叶醇提供基因资源。
-
利用公开数据库phytozome(
https://phytozome-next.jgi.doe.gov/ ),分别获得TPS家族ObGES和CsTPS1,以及NUDIX家族RhNUDX1和PgNdx1的氨基酸序列,与水稻和拟南芥基因组进行Blast比对分析,得到其在水稻和拟南芥中的同源序列。使用MEGA11软件中的Clustal W工具将蛋白序列进行比对分析,选择Neighbor-joining Tree方法构建系统进化树,筛选出水稻香叶醇合成酶的候选基因。从国家水稻数据中心(https://www.ricedata.cn/gene/ )获取OsNUDX11的CDS序列和氨基酸序列;利用NCBI(https://www.ncbi.nlm.nih.gov/ )分析蛋白质保守结构域;在Espasy网站(https://web.expasy.org/protparam/ )对蛋白序列进行理化性质分析;使用SWISS-MODEL网站(https://swissmodel.expasy.org/ )对OsNUDX11蛋白质的三维结构进行预测;利用RiceXPro网站(https://ricexpro.dna.affrc.go.jp/index.html )预测OsNUDX11的组织特异性表达谱;在公开的水稻转录组数据库TENOR(https://tenor.dna.affrc.go.jp/ )上预测OsNUDX11的非生物胁迫诱导表达谱;使用Cell-PLoc 2.0在线预测工具(http://www.csbio.sjtu.edu.cn/bioinf/Cell-PLoc-2/ )预测亚细胞定位。 -
选用野生型中花11(ZH11)为研究材料,在自然环境下培养至分蘖期,选取健康的倒一叶片,使用TRIZOL试剂(赛默飞世尔科技公司)提取水稻叶片中的RNA,并用反转录试剂盒Supermix(北京全式金生物技术有限公司)将RNA反转录为cDNA。反应条件为42 ℃ 15 min,80 ℃ 10 s,cDNA保存于−20 ℃冰箱。
-
以水稻叶片的cDNA为模板,利用带有Gateway接头的PCR特异性引物(F:5′-aaaaagcaggcttaATGCTCCTCGTCAGATCCCAC-3′;R:5′-agaaagctgggtaTCACACAGCCAAGTGGTTGTC-3′),使用高保真酶KOD FX neo(大连宝日医生物工程有限公司)扩增。利用琼脂糖凝胶电泳分析基因扩增结果。扩增片段通过Gateway(Thermo Fisher)技术,将扩增片段构建至pDONR207载体,转入大肠杆菌DH5α感受态中,并测序得到含正确片段的载体(擎科生物公司)。将比对正确后的质粒通过Gateway技术,使目的片段构建至烟草瞬时表达载体(pEAQ-HT-DEST2),使用载体正向引物和基因特异性反向引物(F:5′-cttcgcaagacccttcctct-3′;R:5′-TCACACAGCCAAGTGGTTGTC-3′)鉴定。
-
使用Primer 3 Plus设计实时荧光定量PCR引物(F:5′-TGACATACCATTTGATTCTCTAGCA-3′;R:5′-TGTTTATAGTGCAGTAGTGGAATTTGA-3′ ),利用SYBR酶(吐露港生物公司),以水稻Ubiquitin5基因作为内参基因对水稻样品的cDNA的表达量进行检测。反应程序为:95 ℃ 30 s;95 ℃ 10 s,60 ℃ 30 s,40循环;95 ℃ 30 s;65 ℃ 30 s;95 ℃ 30 s。每个样品采取3个生物学重复,采用2−ΔC法计算相对表达量。
-
将野生型水稻ZH11的种子浸种出芽后,点种于水培盒中,置于30 ℃、50%湿度、光照16 h + 黑暗8 h的光照培养箱中培养至分蘖长出,分别取样倒一叶、倒二叶、倒三叶、叶鞘、根、第一节点、第二节点、第三节点8个样品,置于液氮速冻后−80 ℃保存;将水稻种至土中,观察水稻生长至开花期,取倒一叶、倒二叶、倒三叶、叶鞘、花、穗、穗梗、根、第一节点、第二节点、第三节点11个样品,置于液氮速冻后−80 ℃保存;以开花后21 d为成熟期,取倒一叶、倒二叶、倒三叶、叶鞘、穗、穗梗、胚乳、根、第一节点、第二节点、第三节点11个样品,置于液氮速冻后−80 ℃保存。每个样品为3株材料的混合样品。
-
使用两周龄的野生型水稻ZH11幼苗,置于4 ℃环境下48 h,处理期间分别在0、2、4、6、12、24、36及48 h取样幼苗的倒一叶片,每个实验样品为3株材料的混合样品。
-
将携带目的片段的烟草瞬时表达载体转化至GV3101农杆菌感受态细胞中(上海维地生物)。挑取阳性单克隆菌落进行菌落PCR鉴定,接种至含相应抗性的LB培养基中,28 ℃恒温摇床振荡过夜培养。以
5 000 r·min−1转速离心5 min收集全部菌液,加入烟草缓冲液[10 mmol·L−1 MgCl2,10 mmol·L−1 4-吗啉乙磺酸(MES),100 µmol·L−1乙酰丁香酮(Acetosyringone)]重悬菌体并调整菌液OD600至1.0,避光复苏2 h后。采用无针头注射器将菌液注射至烟草叶片背部,经黑暗条件下培养24 h,光照条件下培养24 h后,使用蔡司LSM900共聚焦显微镜观察荧光现象。 -
为了筛选水稻香叶醇合成酶,笔者以罗勒和茶叶中已经报道的萜烯合酶TPS家族香叶醇合成酶基因ObGES和CsTPS1为靶标。对水稻基因组进行BLAST分析(表1)。结果表明,与ObGES相似度最高的是LOC_Os02g02930(Identity=34%),LOC_Os02g02930编码OsLIS,在水稻中发挥芳樟醇合酶的功能;而与CsTPS1相似度最高的是LOC_Os01g23530(Identity=32%),LOC_Os01g23530编码OsTPS1,该基因在水稻中具有红没药醇合成酶的功能。以玫瑰香叶醇合成酶基因RhNUDX1和天竺葵香叶醇合成酶基因PgNdx1为靶标,对水稻基因组进行Blast分析。结果表明,与RhNUDX1和PgNdx1相似度最高的是LOC_Os09g38040(Identity=45%)(表1),且该基因功能并未表征。
表 1 香叶醇合成酶同源序列比对结果
Table 1. Homologous sequence alignment of geraniol synthase
候选基因
Candidate genesBlast 结果
Blast描述
Description长度/aa
Length比特得分
Bit score期望值
E-value相似度/%
IdentityObGES Os02g0121700 OsLIS 596 290 1e-78 34 Os08g0139700 (E)-β-caryophyllene synthase 577 262 5e-70 34 Os08g0139700 Similar to terpene synthase 6 520 261 1e-69 34 CsTPS1 Os01g0337100 OsTPS1 555 276 3e-74 32 Os04t0344100 Similar to OSIGBa0106G08.3 protein. 544 258 7e-69 31 Os08t0168000 Terpene synthase, Resistance to aphids
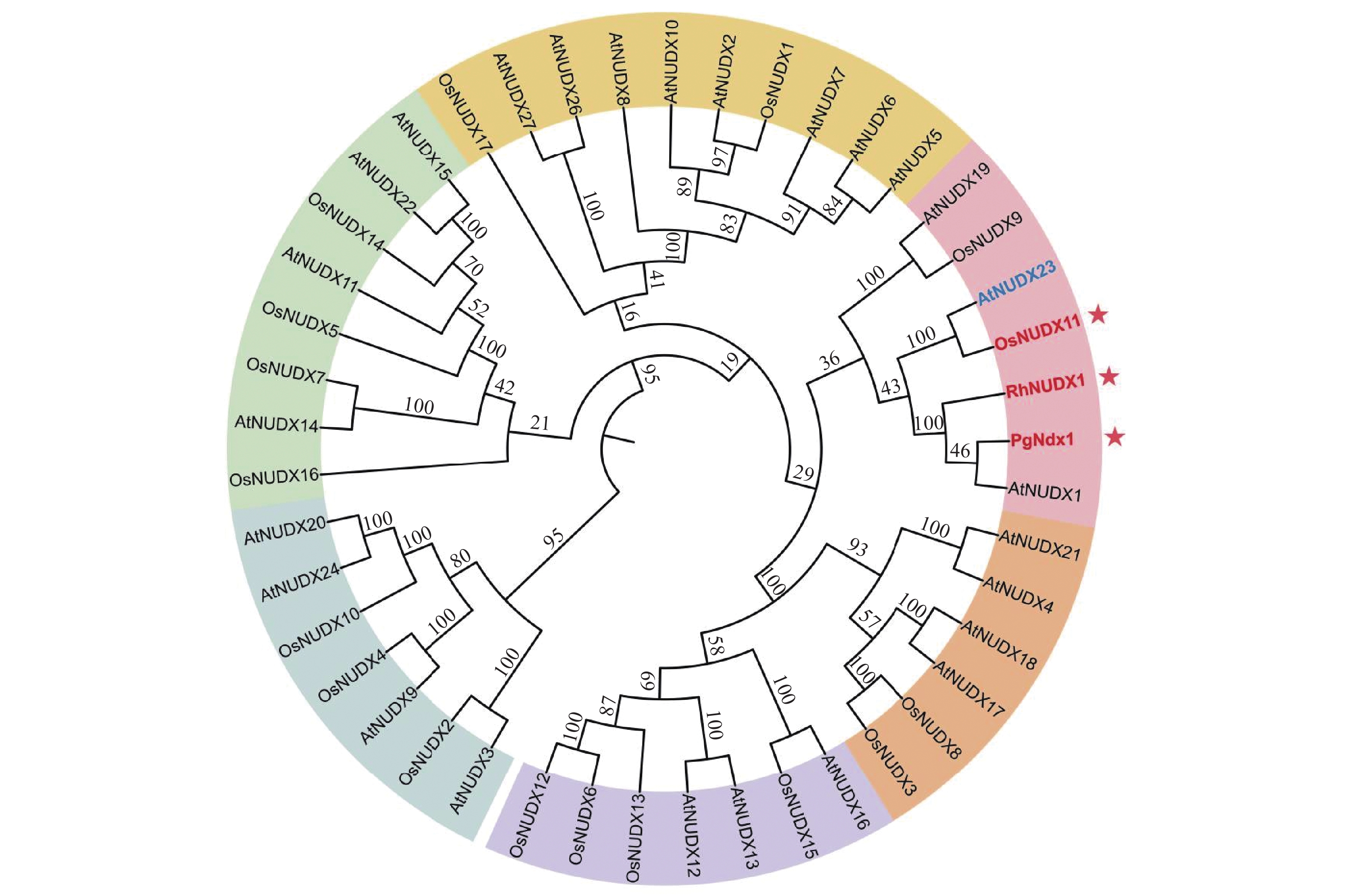
(Rhopalosiphum padi)547 251 8e-67 33 RhNUDX1 Os09g0553300 NUDIX hydrolase domain containing protein 185 40.4 5e-04 45 Os06g0634300 Similar to Nudix hydrolase 2 331 32 1.4e-01 29 Os08g0139700 Similar to terpene synthase 6 520 261 1e-69 31 PgNdx1 Os09g0553300 NUDIX hydrolase domain containing protein 185 37 4e-03 32 Os08g0375900 Similar to Nudix hydrolase 22 251 36.6 7e-03 35 Os06g0129700 Nudix hydrolase 14 326 34.3 2.6e-02 40 LOC_Os09g38040编码OsNUDX11,为进一步确认OsNUDX11是否与香叶醇合成有关,分别检索已命名的水稻和拟南芥NUDIX家族成员,发现27个拟南芥和17个水稻NUDIX水解酶家族成员,将其与玫瑰和天竺葵的香叶醇合成酶蛋白序列构建系统发育树。系统发育树显示,OsNUDX11与AtNUDX23、PgNdx1及RhNUDX1显示出较为紧密的亲缘关系(图1),而AtNUDX23已被鉴定为黄素腺嘌呤二核苷酸(FAD)焦磷酸水解酶[18],一种参与黄素代谢负反馈调控的水解酶,这表明OsNUDX11在水稻中很可能参与香叶醇的生成。
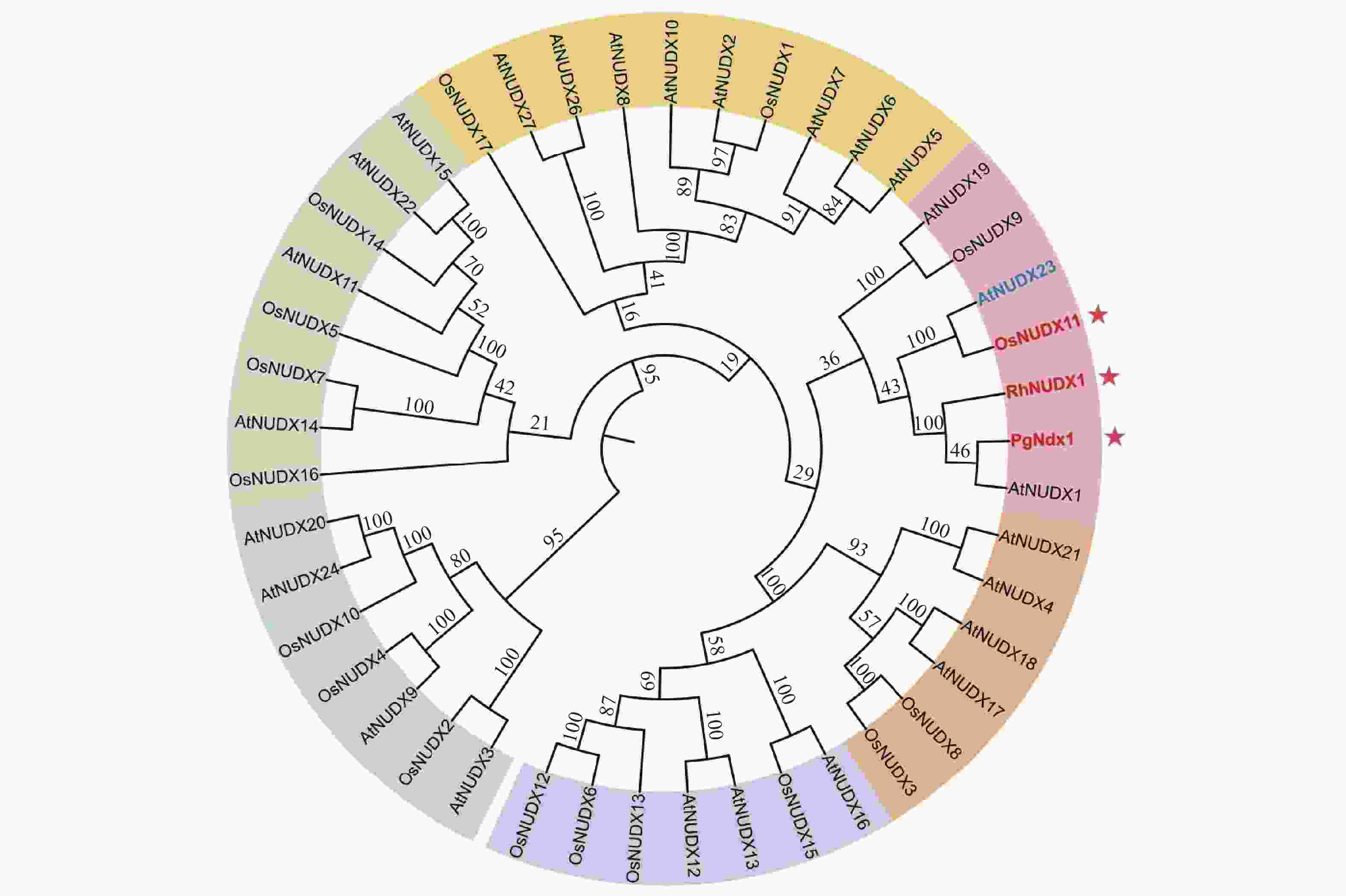
图 1 NUDIX水解酶家族系统发育树
Figure 1. The phylogenetic tree of the NUDIX hydrolase family
基因结构分析结果表明,OsNUDX11基因含有7段外显子和6段内含子,编码区共813个核苷酸,编码271个氨基酸(图2-a)。利用NCBI在线网站的CDD功能对OsNUDX11蛋白质序列的保守结构域进行分析。结果表明,OsNUDX11与RhNUDX1和PgNdx1存在一个相同的保守结构域(图2-b),推测OsNUDX11可能具有香叶醇合成酶的功能。
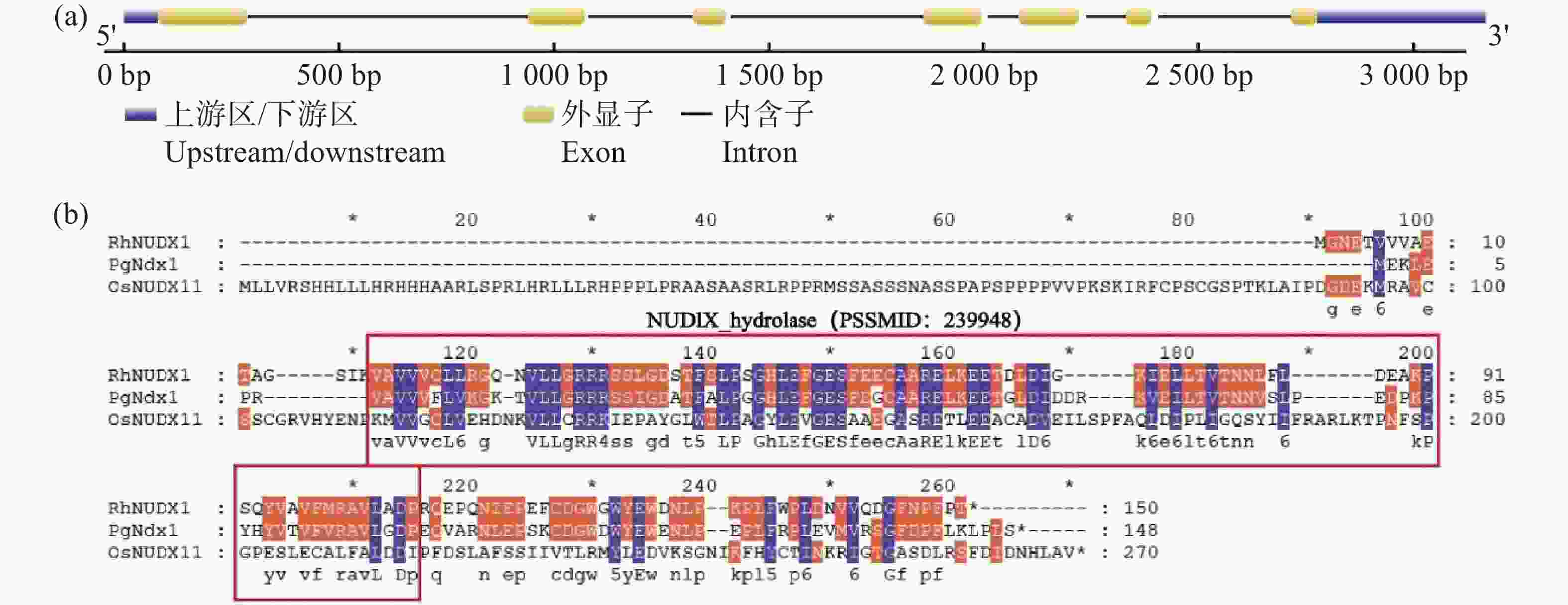
图 2 OsNUDX11基因的结构和蛋白质保守结构域分析
Figure 2. Analysis of the structure and protein conservative domain of OsNUDX11 gene
-
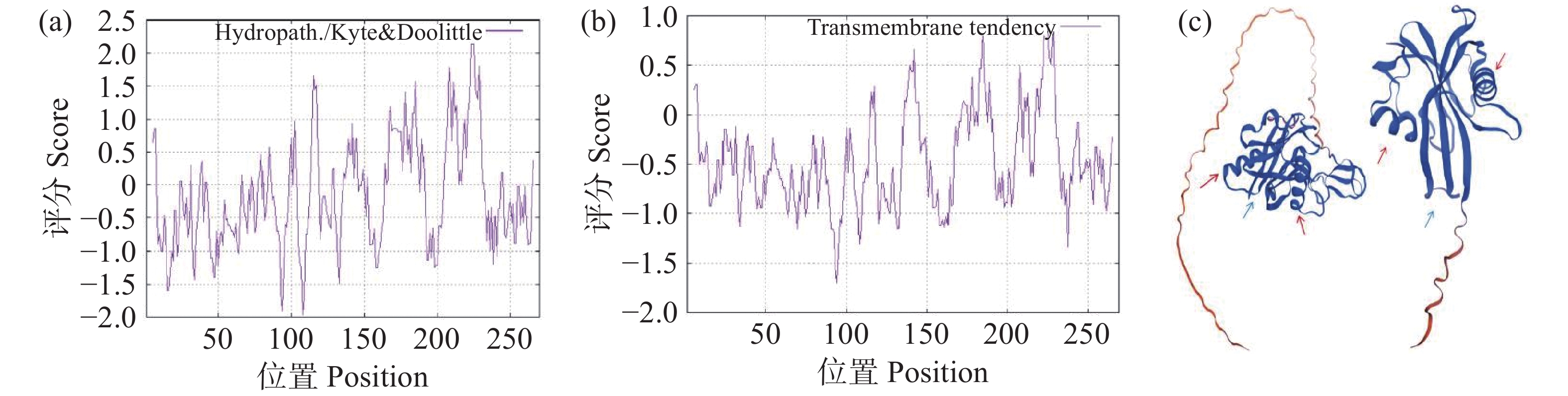
采用Espasy在线网站对OsNUDX11蛋白序列进行理化性质预测,结果表明,OsNUDX11蛋白质相对分子量为29.66 kDa,理论等电点为8.87,总平均亲水性GRAVY为-0.118,疏水性曲线与跨膜趋势分析中氨基酸序列密集均分布在负值以下(图3-a,图3-b),表明该蛋白为亲水性蛋白且不易发生跨膜;不稳定系数为71.95,表明该蛋白在体外环境中易失活;脂肪系数为92.19,表明其热稳定性较好,在温度波动范围内不易因热胁迫失活,但在酶的参与下容易失活。使用SWISS-MODEL在线网站预测OsNUDX11蛋白质的三维结构,结果表明,OsNUDX11与RhNUDX1蛋白的三维结构相似,均具有两个螺旋结构以及排列整齐的网状结构(图3-c)。
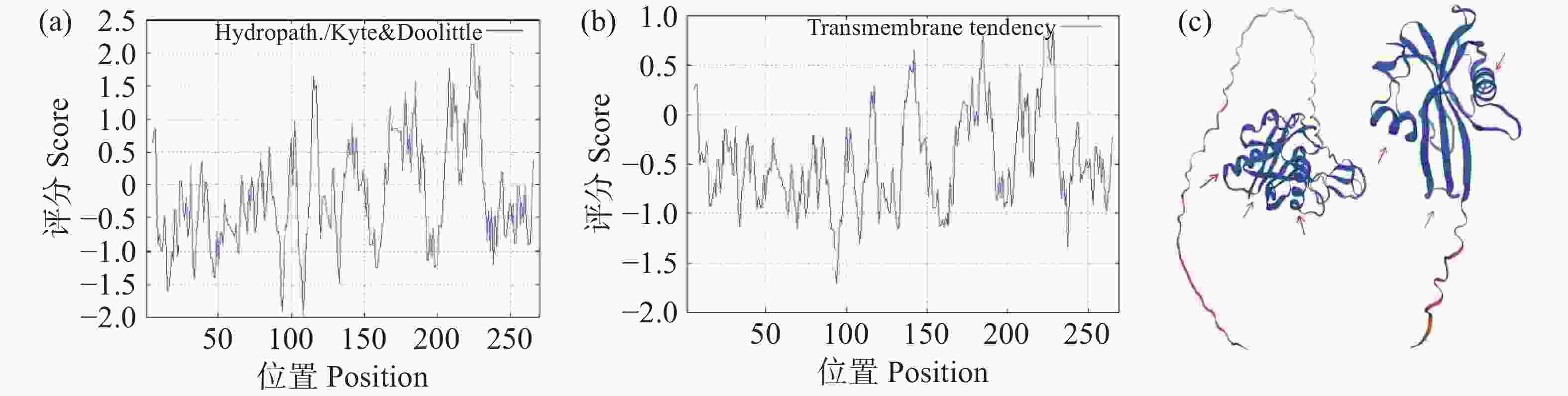
图 3 OsNUDX11蛋白质的理化性质与OsNUDX11和RhNUDX1蛋白质的三维结构
Figure 3. The physicochemical properties of OsNUDX11 protein the tertiary structure of OsNUDX11 and RhNUDX1 proteins
-
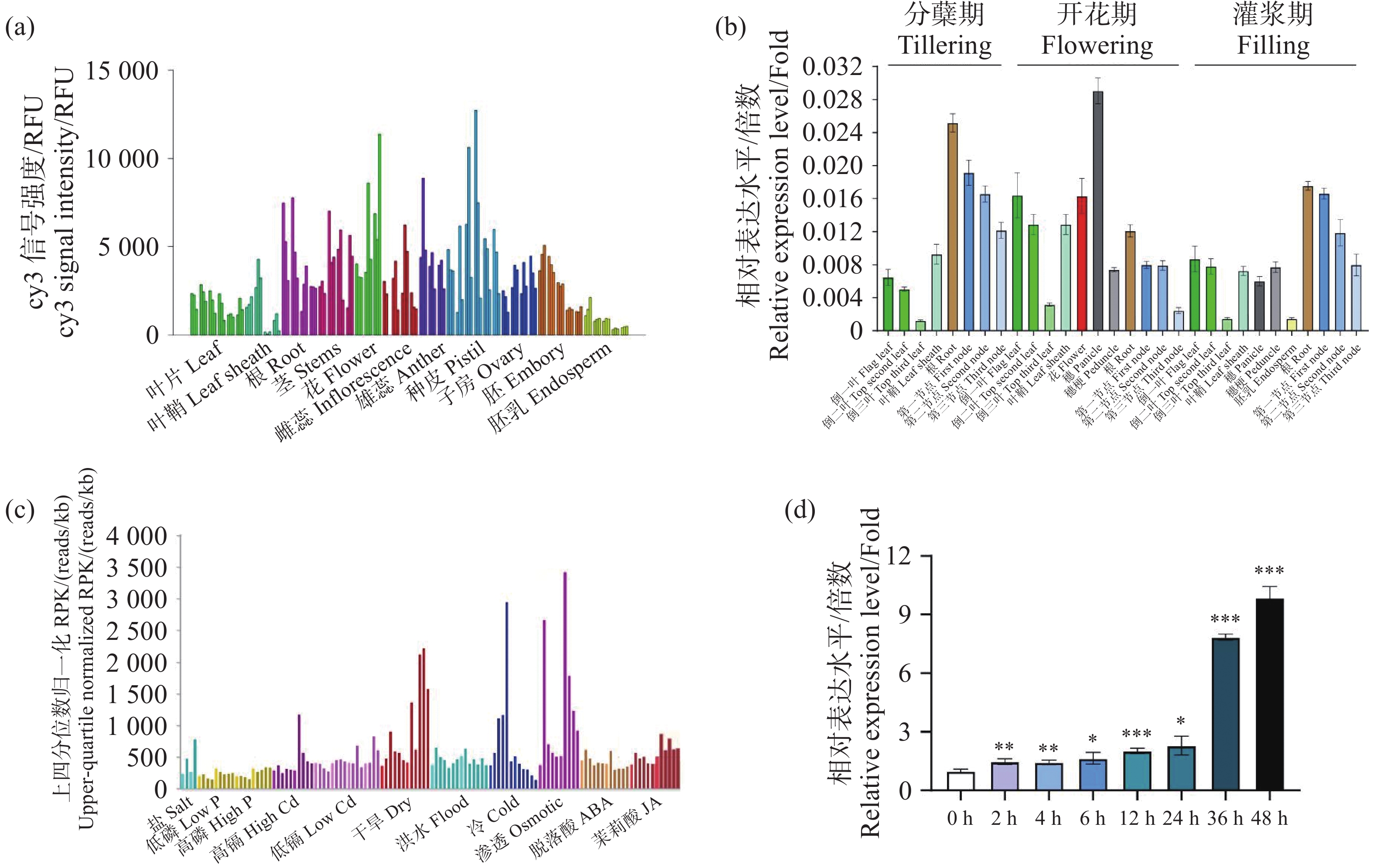
为探究OsNUDX11在水稻中的表达模式,首先使用RiceXPro预测OsNUDX11的组织特异性表达模式,结果表明,OsNUDX11在根和穗中表达量较高(图4-a)。为了进一步探究OsNUDX11的表达模式,分别对分蘖期、开花期以及灌浆期的野生型水稻ZH11的30个不同组织的表达量进行测定。结果表明,OsNUDX11在水稻的根和穗中表达量较高,与网站预测结果一致(图4-b)。为探究OsNUDX11受到非生物胁迫的响应,本研究使用TENOR在线网站对不同胁迫下OsNUDX11表达量进行预测,结果表明,OsNUDX11对渗透胁迫和冷胁迫的响应较为明显(图4-c)。为验证该预测结果,使用野生型水稻ZH11的三叶期幼苗在4 ℃环境下处理48 h,并分别测定0 h、2 h、4 h、6 h、12 h、24 h、36 h以及48 h叶片的基因表达量,结果表明,在冷处理后OsNUDX11的表达量随处理的时间增加,表达量显著提高(图4-d),说明OsNUDX11的表达受冷胁迫调控,与网站预测结果一致。
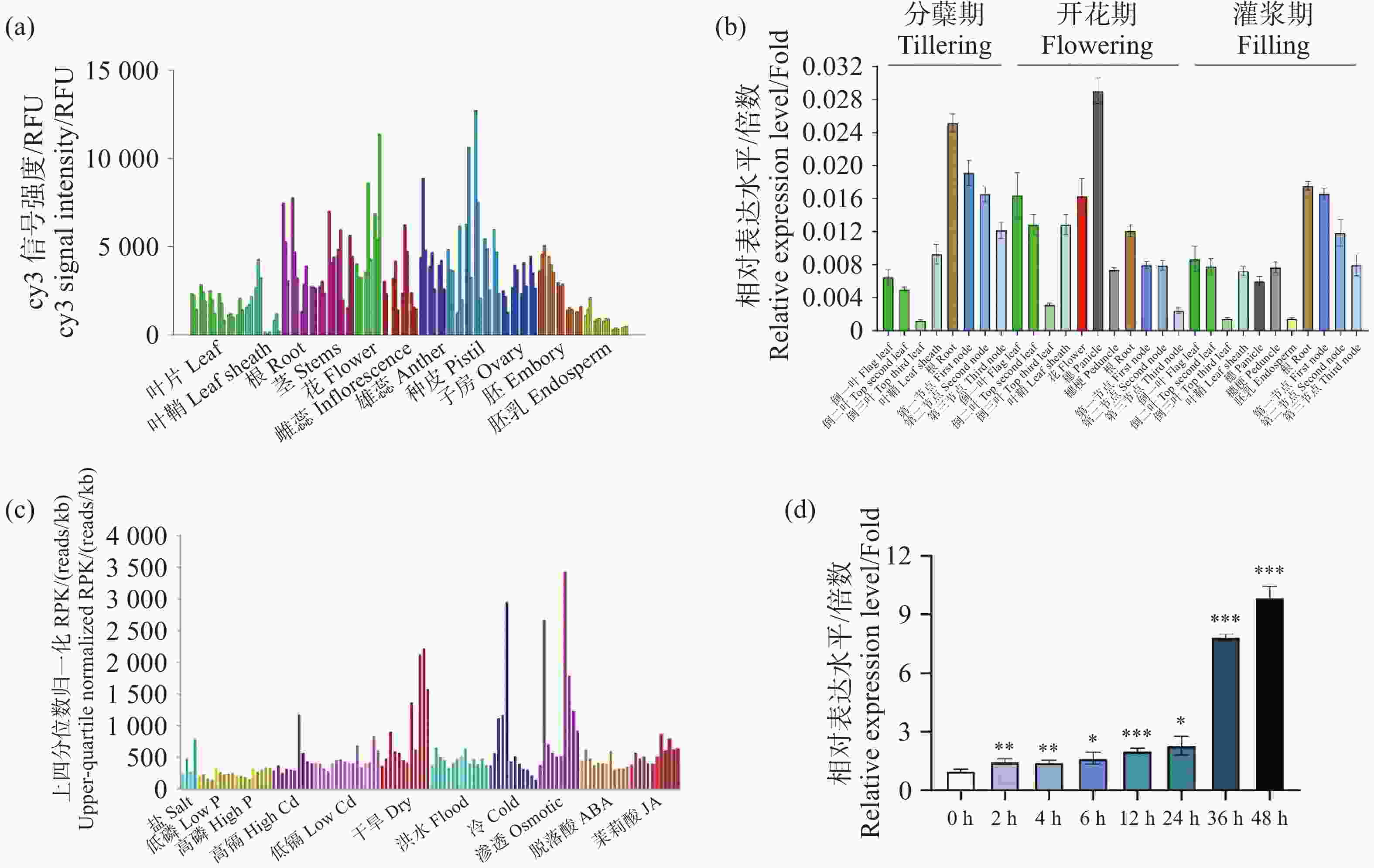
图 4 OsNUDX11的表达谱和诱导表达
Figure 4. OsNUDX11 expression profile and induced expression
-
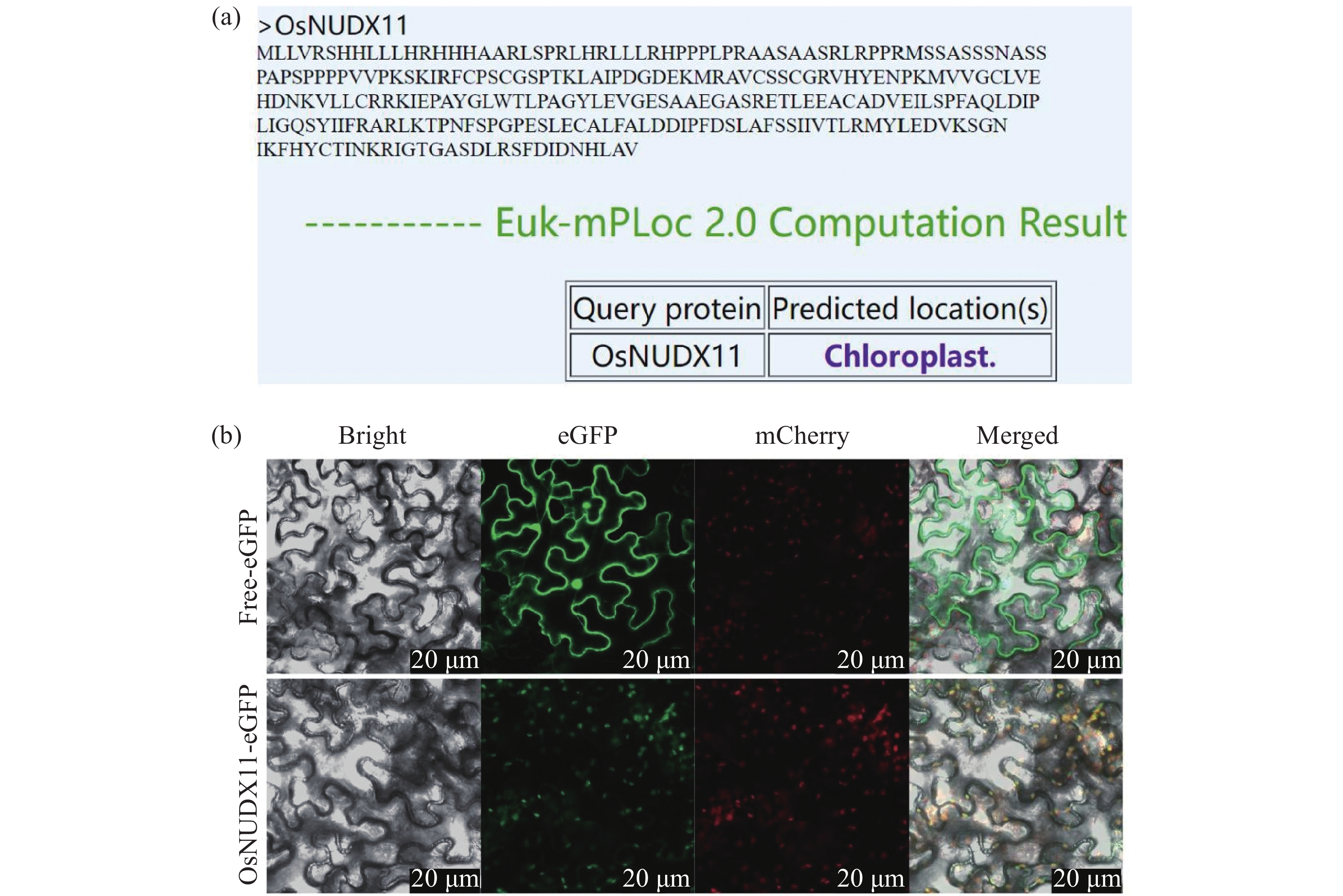
为明确OsNUDX11蛋白的亚细胞定位,利用Cell-PLoc 2.0在线预测工具对OsNUDX11蛋白序列进行分析,预测结果显示其亚细胞定位与大多数萜烯合酶一致,均定位于叶绿体(图5-a)。之后将35S:OsNUDX11-eGFP融合表达载体(实验组)及35S:eGFP空载体(对照组)转入农杆菌感受态细胞GV3101中,分别注射至1月龄烟草叶片,培养48h后使用共聚焦显微镜观察荧光现象。结果表明,与GFP空载相比,35S:OsNUDX11-eGFP的绿色荧光与叶绿体红色自发荧光完全重叠(图5-b),表明OsNUDX11与玫瑰RhNUDX1和天竺葵PgNdx1亚细胞定位一致,三者均在质体中发挥功能,进一步说明OsNUDX11很可能参与香叶醇合成过程。
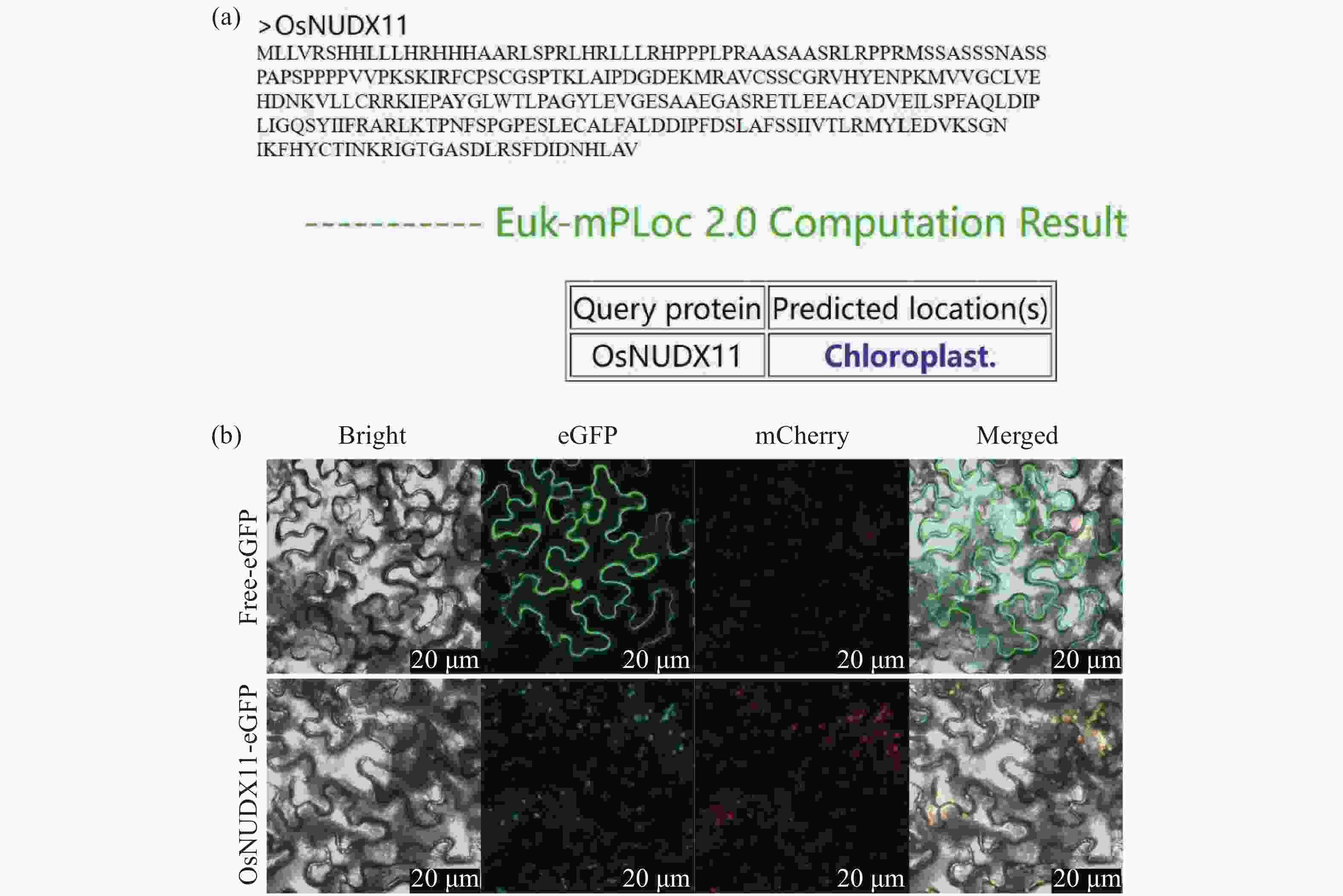
图 5 OsNUDX11亚细胞定位预测结果(a)和OsNUDX11亚细胞定位结果(b)
Figure 5. The subcellular localization prediction results of OsNUDX11 and the subcellular localization results of OsNUDX11
-
NUDIX家族水解酶是一类普遍存在于真核生物中的水解酶,其功能主要是水解核苷二磷酸化合物[19]。在植物体中,NUDIX家族水解酶主要定位在胞质溶胶、线粒体和叶绿体中,控制着多种代谢物的合成,并广泛参与生物和非生物胁迫反应,如干旱、辐射、盐度和病原体攻击[20]。拟南芥AtNUDX9主要负责水解GDP-D-甘露糖生成甘露糖1-磷酸,通过调节根中的蛋白质N-糖基化参与GDP-D-Man水平的调节,从而影响铵敏感性[21];大麦HvNUDX12具有8-氧代-dGTP、四磷酸二腺苷(Ap4A)和鸟苷四磷酸(ppGpp)焦磷酸水解酶活性,该独特的结构可耐受非生物胁迫的反应[22],如受紫外UV-C诱导后,HvNUDX12基因表达量显著上调;水稻OsNUDX2的重组蛋白可在体外水解8-氧代-dGTP,以及二甲基烯丙基二磷酸(DMAPP)和异戊烯基二磷酸(IPP),OsNUDX2通过不同的底物特异性和同一性有效降低紫外线胁迫[23];而OsNUDX14定位在线粒体,参与调控水稻木质素的生物合成,同时调控水稻叶夹角和籽粒垩白度等农艺性状[24]。
本研究利用已报道的香叶醇合成酶进行BLAST比对,鉴定并克隆了一个水稻已被命名的基因OsNUDX11,通过荧光定量实验验证了OsNUDX11在根和穗中表达量较高且受冷处理诱导,亚细胞定位证实了OsNUDX11编码的蛋白质在叶绿体中定位,这一定位与玫瑰和天竺葵中报道的RhNUDX1与PgNdx1一致。但笔者发现OsNUDX11与玫瑰香叶醇合成酶RhNUDX1的蛋白质三维结构仍存在一定差异,因此后续还需要对OsNUDX11催化香叶基二磷酸生成香叶醇的机制进行进一步探究,为解析OsNUDX11催化活性提供理论依据。
氨基酸序列比对分析表明,OsNUDX11与拟南芥AtNUDX23同源关系较近,AtNUDX23在南芥叶片中高度表达,定位在叶绿体中,通过与叶绿体中的八氢番茄红素合酶(PSY,phytoene synthase)和牻牛儿牻牛儿基焦磷酸合酶(geranylgeranyl diphosphate synthase, GGPPS)直接相互作用来调节类胡萝卜素的生物合成[25],同时AtNUDX23在质体中发挥黄素腺嘌呤二核苷酸(FAD)焦磷酸水解酶活性,通过水解黄素单核苷酸(FMN)和5’-腺苷酸(AMP),使二者结合产生FAD,从而调节植物细胞中的黄素稳态[18]。本研究推测OsNUDX11可能通过水解香叶基焦磷酸 GPP 来合成香叶醇。因此,还需继续验证OsNUDX11可能同时调节水稻类胡萝卜素生物合成与调节水稻生长发育过程中黄素的稳态。
Screening and functional characterization of rice geraniol synthase
-
摘要: 香稻因其品质优良、香味浓郁、清醇可口,长期颇受人们喜爱。香叶醇是一种散发玫瑰香味的高价值单萜化合物,具有广泛的医药生理活性和应用。然而,香稻中香叶醇含量较低,且香叶醇合成酶功能未被解析。为筛选和鉴定水稻香叶醇合成酶的候选基因,本研究以已报道的香叶醇合成酶萜烯合酶(TPS)家族和核苷二磷酸酶X(NUDIX)水解酶家族基因为靶标,对水稻进行同源序列比对分析和系统进化树分析,结合组织特异性表达谱,筛选到一个在水稻根和穗中高表达的NUDIX家族成员OsNUDX11,该基因与玫瑰和天竺葵中的香叶醇合成酶亲缘关系较为紧密。通过进一步的蛋白理化性质分析和烟草瞬时表达实验,证实了OsNUDX11与玫瑰和天竺葵中的香叶醇合成酶亚细胞定位一致。故推测OsNUDX11参与了香叶醇合成,为在水稻中大量合成香叶醇提供重要依据。Abstract: Fragrant rice has long been cherished for its superior quality, potent aroma, delicate flavor, and pleasant texture. Geraniol, a valuable monoterpene with a rose-like odor, exhibits a broad spectrum of medicinal and physiological activities. Despite its popularity, fragrant rice varieties typically contain low levels of geraniol, and the biochemical function of geraniol synthase in these plants remains undefined. In this context the previously reported geraniol synthase, terpene synthase (TPS), family and the nucleoside diphosphatase X (NUDIX) hydrolase family were employed to select candidate genes for geraniol synthase in rice by using homologous sequence alignment analysis and phylogenetic tree analysis of rice, combined with tissue-specific expression profiles, and OsNUDX11, a member of the NUDIX family that is highly expressed in rice roots and panicles was identified. This gene is closely related to geraniol synthases from Rosa rugosa (rose) and Pelargonium graveolens (geranium). Further analysis of protein physicochemical properties and tobacco transient expression assays confirmed that OsNUDX11 shares consistent subcellular localization with geraniol synthases from rose and geranium. It is, thus, inferred that OsNUDX11 be involved in geraniol biosynthesis, which provides a crucial basis for the large-scale synthesis of geraniol in rice.
-
Key words:
- rice /
- geraniol /
- NUDIX hydrolases /
- subcellular localization
-
图 2 OsNUDX11基因的结构和蛋白质保守结构域分析
(a)OsNUDX11的基因结构图;(b)OsNUDX11、RhNUDX1和PgNdx1蛋白质序列比对结果。红框部分为其三者共有的NUDIX水解酶保守结构域,PSSMID号为239948。
Fig. 2 Analysis of the structure and protein conservative domain of OsNUDX11 gene
(a) Gene structure of OsNUDX11; (b) OsNUDX11 RhNUDX1 and PgNdx1 protein sequence alignment results. The red box represents the conserved domains of NUDIX hydrolase shared by the three protein sequences.The PSSMID number is 239948.
图 3 OsNUDX11蛋白质的理化性质与OsNUDX11和RhNUDX1蛋白质的三维结构
(a)OsNUDX11蛋白质疏水性分析,纵坐标为疏水性评分,横坐标为氨基酸长度,评分越高说明蛋白质疏水性越好;(b)OsNUDX11蛋白质跨膜区预测,纵坐标为跨膜评分,横坐标为氨基酸长度,评分越高说明该蛋白在发挥功能时可能发生跨膜;(c)OsNUDX11与RhNUDX1蛋白质三级结构,红色箭头标注为螺旋结构,蓝色箭头标注为网状结构。
Fig. 3 The physicochemical properties of OsNUDX11 protein the tertiary structure of OsNUDX11 and RhNUDX1 proteins
(a) Hydrophobicity analysis of OsNUDX11 protein: Hydrophobicity score is on the vertical axis, and amino acid length on the horizontal axis; the higher the score, the better the hydrophobicity of the protein. (b) OsNUDX11 protein transmembrane region prediction: The vertical axis represents transmembrane score and the horizontal axis represents amino acid length; the higher the score, the more likely the protein may undergo transmembrane transition while performing its function. (c) The tertiary structures of OsNUDX11 and RhNUDX1 proteins are indicated by a red arrow indicating a helical structure and a blue arrow indicating a reticular structure.
图 4 OsNUDX11的表达谱和诱导表达
(a)RiceXPro网站由芯片测序测得的OsNUDX11表达谱图;(b)OsNUDX11表达谱图。其中相同颜色代表不同时期的相同组织;(c)TENOR网站预测的OsNUDX11诱导表达谱图;(d)OsNUDX11对冷胁迫的表达谱图。
Fig. 4 OsNUDX11 expression profile and induced expression
(a) OsNUDX11 expression profile obtained from chip sequencing on the RichXPro website; (b) OsNUDX11 expression profile. The same color represents the same tissue from different periods; (c) OsNUDX11 induced expression profile predicted by TENOR website; (d) Expression profile of OsNUDX11 under cold stress.
图 5 OsNUDX11亚细胞定位预测结果(a)和OsNUDX11亚细胞定位结果(b)
注:eGFP-OsNUDX11融合蛋白在1月龄本氏烟草叶片中瞬时表达。eGFP呈绿色荧光,叶绿体在488 nm激发光下呈自发红色荧光,绿色荧光与红色荧光叠加后呈现黄色荧光。标尺为20 μm。
Fig. 5 The subcellular localization prediction results of OsNUDX11 and the subcellular localization results of OsNUDX11
Note: The eGFP-OsNUDX11 fusion protein was transiently expressed in the leaves of 1-month-old Nicotiana benthamiana. eGFP shows green fluorescence, chloroplasts show spontaneous red fluorescence under 488 nm excitation light, and yellow fluorescence appears after the superposition of green fluorescence and red fluorescence. The scale bar is 20 μm.
表 1 香叶醇合成酶同源序列比对结果
Table 1 Homologous sequence alignment of geraniol synthase
候选基因
Candidate genesBlast 结果
Blast描述
Description长度/aa
Length比特得分
Bit score期望值
E-value相似度/%
IdentityObGES Os02g0121700 OsLIS 596 290 1e-78 34 Os08g0139700 (E)-β-caryophyllene synthase 577 262 5e-70 34 Os08g0139700 Similar to terpene synthase 6 520 261 1e-69 34 CsTPS1 Os01g0337100 OsTPS1 555 276 3e-74 32 Os04t0344100 Similar to OSIGBa0106G08.3 protein. 544 258 7e-69 31 Os08t0168000 Terpene synthase, Resistance to aphids
(Rhopalosiphum padi)547 251 8e-67 33 RhNUDX1 Os09g0553300 NUDIX hydrolase domain containing protein 185 40.4 5e-04 45 Os06g0634300 Similar to Nudix hydrolase 2 331 32 1.4e-01 29 Os08g0139700 Similar to terpene synthase 6 520 261 1e-69 31 PgNdx1 Os09g0553300 NUDIX hydrolase domain containing protein 185 37 4e-03 32 Os08g0375900 Similar to Nudix hydrolase 22 251 36.6 7e-03 35 Os06g0129700 Nudix hydrolase 14 326 34.3 2.6e-02 40  下载: 导出CSV
下载: 导出CSV
-
[1] 余亚莹, 邵高能, 圣忠华, 等. 国内外香稻资源遗传多样性研究[J]. 植物分类与资源学报, 2015, 37(6), 871−880 [2] 韩丽欣, 任红波, 孟利. 大米中风味物质的形成与变化[J]. 中国农学通报, 2022, 38(30), 126−134 [3] Hu X Q, Lu L, Guo Z L, et al. Volatile compounds, affecting factors and evaluation methods for rice aroma: a review [J]. Trends in Food Science & Technology, 2020, 97, 136−146. https://doi.org/10.1016/j.jpgs.2020.01.003 doi: 10.1016/j.jpgs.2020.01.003 [4] Buttery R G, Ling L C, Juliano B O, et al. Cooked rice aroma and 2-acetyl-1-pyrroline [J]. Journal of Agricultural and Food Chemistry, 1983, 31 (4), 823−826. https://doi.org/10.1021/jf00118a036 doi: 10.1021/jf00118a036 [5] 周汉琛, 杨霁虹, 徐玉婕, 等. 香叶醇生物合成相关基因NUDX1的进化分析[J]. 茶叶科学, 2022, 42(5), 638−648. https://doi.org/10.3969/j.issn.1000-369X.2022.05.004 doi: 10.3969/j.issn.1000-369X.2022.05.004 [6] Edris A E. Pharmaceutical and therapeutic potentials of essential oils and their individual volatile constituents: a review [J]. Phytotherapy Research, 2007, 21 (4), 308−323. https://doi.org/10.1002/ptr.2072 doi: 10.1002/ptr.2072 [7] Si W, Ni X, Gong J, et al. Antimicrobial activity of essential oils and structurally related synthetic food additives towards Clostridium perfringens [J]. Journal of Applied Microbiology, 2009, 106 (1), 213−220. https://doi.org/10.1111/j.1365-2672.2008.03994.x doi: 10.1111/j.1365-2672.2008.03994.x [8] 周勇, 姚三桃, 翁玉春, 等. 香叶醇抗真菌作用的研究[J]. 真菌学报, 1983, 2(4), 262−264. https://doi.org/10.13346/j.mycosystema.1983.04.013 doi: 10.13346/j.mycosystema.1983.04.013 [9] Bhavaniramya S, Vishnupriya S, Al-Aboody M S, et al. Role of essential oils in food safety: antimicrobial and antioxidant applications [J]. Grain & Oil Science and Technology, 2019, 2 (2), 49−55. https://doi.org/10.1016/j.gaost.2019.03.001 doi: 10.1016/j.gaost.2019.03.001 [10] 罗彪彪. 应用酵母表面展示技术体外重构甲羟戊酸途径合成香叶醇[D]. 武汉: 华中农业大学, 2020. https://doi.org/10.27158/d.cnki.ghznu.2020.001106 [11] 殷焕然, 何玉池, 李洋. GC-MS/MS法分析水稻叶子单萜倍半萜类化合物的组成[J]. 生物过程, 2017, 7(4), 54−59. https://doi.org/10.12677/bp.2017.74008 doi: 10.12677/bp.2017.74008 [12] Degenhardt J, Köllner T G, Gershenzon J. Monoterpene and sesquiterpene synthases and the origin of terpene skeletal diversity in plants [J]. Phytochemistry, 2009, 70 (15/16), 1621−1637. https://doi.org/10.1016/j.phytochem.2009.07.030 doi: 10.1016/j.phytochem.2009.07.030 [13] Iijima Y, Gang D R, Fridman E, et al. Characterization of geraniol synthase from the peltate glands of sweet basil [J]. Plant Physiology, 2004, 134 (1), 370−379. https://doi.org/10.1104/pp.103.032946 doi: 10.1104/pp.103.032946 [14] Li X, Xu Y Y, Shen S L, et al. Transcription factor CitERF71 activates the terpene synthase gene CitTPS16 involved in the synthesis of E-geraniol in sweet orange fruit [J]. Journal of Experimental Botany, 2017, 68 (17), 4929−4938. https://doi.org/10.1093/jxb/erx316 doi: 10.1093/jxb/erx316 [15] Jiang H, Zhang M T, Yu F, et al. A geraniol synthase regulates plant defense via alternative splicing in tea plants [J]. Horticulture Research, 2023, 10 (10), uhad184. https://doi.org/10.1093/hr/uhad184 doi: 10.1093/hr/uhad184 [16] Magnard J L, Roccia A, Caissard J C, et al. Biosynthesis of monoterpene scent compounds in roses [J]. Science, 2015, 349 (6243), 81−83. https://doi.org/10.1126/science.aab0696 doi: 10.1126/science.aab0696 [17] Bergman M E, Bhardwaj M, Phillips M A. Cytosolic geraniol and citronellol biosynthesis require a Nudix hydrolase in rose-scented geranium (Pelargonium graveolens) [J]. The Plant Journal, 2021, 107 (2), 493−510. https://doi.org/10.1111/tpj.15304 doi: 10.1111/tpj.15304 [18] Maruta T, Yoshimoto T, Ito D, et al. An Arabidopsis FAD pyrophosphohydrolase, AtNUDX23, is involved in flavin homeostasis [J]. Plant and Cell Physiology, 2012, 53 (6), 1106−1116. https://doi.org/10.1093/pcp/pcs054 doi: 10.1093/pcp/pcs054 [19] Arczewska K D, Kuśmierek J T. Bacterial DNA repair genes and their eukaryotic homologues: 2. Role of bacterial mutator gene homologues in human disease. Overview of nucleotide pool sanitization and mismatch repair systems [J]. Acta Biochimica Polonica, 2007, 54 (3), 435−457. https://doi.org/10.18388/abp.2007_3220 doi: 10.18388/abp.2007_3220 [20] Ogawa T, Muramoto K, Takada R, et al. Modulation of NADH levels by Arabidopsis Nudix hydrolases, AtNUDX6 and 7, and the respective proteins themselves play distinct roles in the regulation of various cellular responses involved in biotic/abiotic stresses [J]. Plant and Cell Physiology, 2016, 57 (6), 1295−1308. https://doi.org/10.1093/pcp/pcw078 doi: 10.1093/pcp/pcw078 [21] Ogawa T, Yoshimura K, Miyake H, et al. Molecular characterization of organelle-type Nudix hydrolases in Arabidopsis [J]. Plant Physiology, 2008, 148 (3), 1412−1424. https://doi.org/10.1104/pp.108.128413 doi: 10.1104/pp.108.128413 [22] Tanaka S, Kihara M, Sugimoto M. Structure and molecular characterization of barley Nudix hydrolase genes [J]. Bioscience, Biotechnology, and Biochemistry, 2015, 79 (3), 394−401. https://doi.org/10.1080/09168451.2014.978259 doi: 10.1080/09168451.2014.978259 [23] Kondo Y, Rikiishi K, Sugimoto M. Rice Nudix hydrolase OsNUDX2 sanitizes oxidized nucleotides [J]. Antioxidants, 2022, 11 (9), 1805. https://doi.org/10.3390/antiox11091805 doi: 10.3390/antiox11091805 [24] Liu Y R, Zhang W, Wang Y H, et al. Nudix hydrolase 14 influences plant development and grain chalkiness in rice [J]. Frontiers in Plant Science, 2022, 13, 1054917. https://doi.org/10.3389/FPLS.2022.1054917 doi: 10.3389/FPLS.2022.1054917 [25] Rao S, Cao H B, O'hanna F J, et al. Nudix hydrolase 23 post-translationally regulates carotenoid biosynthesis in plants [J]. The Plant Cell, 2024, 36 (5), 1868−1891. https://doi.org/10.1093/plcell/koae030 doi: 10.1093/plcell/koae030 -

 点击查看大图
点击查看大图
计量
- 文章访问数: 639
- HTML全文浏览量: 317
- PDF下载量: 69
- 被引次数: 0
