


-
水稻(Oryza sativa L.),原产于中国和印度,可分为籼稻(Oryza sativa L. subsp. indica Kato)和粳稻(Oryza sativa L. subsp. japonica Kato)。作为单子叶模式植物,水稻具有易培养、生长快、基因组较小且具有精细的基因组序列等优点[1-2]。植物激素是调控水稻植株最终表型的因素之一[3],同时水稻体内的代谢物变化又是植物表型的直接体现者[4],因此研究植物激素对水稻代谢的影响可以为水稻表型形成提供有价值的理论数据和生长发育模型。独脚金内酯作为一种新型植物激素,可以抑制水稻分蘖,并在水稻种子发芽过程中负调控细胞分裂,抑制侧根的形成和初生根的伸长[5-6]。水稻中,独脚金内酯的生物合成首先是由全反式−β−胡萝卜素在异构酶D27的作用下形成9−反式−β−胡萝卜素[7-8]。9−反式−β−胡萝卜素在类胡萝卜素裂解双加氧酶CCD7(D17)的作用下形成9−顺式−β−胡萝卜素[9],其在由类胡萝卜素裂解双加氧酶CCD8(D10)的催化下形成R−构型−己内酯[10],R−构型−己内酯在细胞色素P450(Os900)的作用下形成5−脱氧独脚金醇[11],最终在细胞色素P450(Os1400)的作用下,形成不同形式的独脚金内酯[12]。
水稻中存在大量的初生及次生代谢产物,据报道,目前从水稻中已鉴定出276种次级代谢产物,主要包括酚酸、类黄酮、萜类、类固醇、生物碱及其衍生物等[13],其广泛参与了水稻的生长发育及抗氧化、抗病、抗虫等生理过程[14-15]。有研究表明,独脚金内酯可以影响水稻中可溶性糖、碳水化合物和氨基酸等代谢途径的重要酶的含量[16-17]。在低磷酸盐的条件下,独脚金内酯可以调节水稻中叶绿素的含量[18];在拟南芥中,独脚金内酯能够调节花青素的积累、抑制褪黑素的合成、诱导磷酸盐代谢相关基因的表达和酸性磷酸酶的产生[19-21];此外,独脚金内酯能够影响类黄酮生物合成基因的表达,外源施加独脚金内酯类似物GR24会导致黄酮生物合成相关的酶的丰度增加,从而影响黄酮醇的积累[22]。虽然有关独脚金内酯在水稻中的生物学功能研究已有不少,然而与独脚金内酯调节水稻次生代谢的相关研究还比较缺乏,因此,笔者以水稻独脚金内酯合成突变体d17及其野生型中花11(Oryza Sativa L. ssp. japonica cv. zhonghua 11, ZH11)为材料,进行转录组和代谢组分析,旨在发现独脚金内酯对水稻初生和次生代谢过程的影响,为进一步研究独脚金内酯调控水稻体内初生和次生代谢及生长发育提供参考。
-
本实验以水稻独脚金内酯合成缺失突变体d17及其野生型栽培品种ZH11为研究对象。为了获取生长一致的实验材料,首先通过无菌培养的方式将上述材料培育成幼苗,随后进行水培。简要过程如下:将d17和ZH11种子脱壳,放入锥形瓶中,加入50 mL ω=0.15%的升汞,37 ℃摇床中摇晃灭菌15 min。倒掉升汞,先用无菌蒸馏水清洗2次,回收至5 mL的无菌试管中,并用无菌蒸馏水清洗5次。将灭菌的水稻种子转移到1/2 MS固体培养基中培养14 d。将生长14 d的水稻幼苗转移至水培槽中,用Hoagland’s营养液进行培养,每3天更换1次营养液。取生长30 d水稻幼苗的幼叶和根,立即放入液氮中速冻,随后储存于−80 ℃冰箱备用。
-
用Trizol试剂盒(Trans Gen Biotech)提取样品的总RNA,并送到武汉未来组生物科技公司进行质检,以保证所有样品最终都获得高质量的总RNA。利用Illumina HiSeq X Ten测序平台(Illumina Inc, CA, USA)进行同批次的转录组建库测序及分析。流程如下:(1)用带有Oligo(dT)的磁珠富集mRNA,去除rRNA;(2)将RNA随机打断,用随机引物反转录,末端修复3′,5′端加A尾,连接测序接头;(3)扩增产生DNA的聚集片段;(4)利用Illumina测序平台,进行双末端测序;(5)去低质量序列、去接头污染,得到高质量的序列;(6)将测序结果用CASAVA软件进行碱基识别,转化为原始测序序列;(7)用fastp软件去除接头污染的Reads,去除低质量的Reads,去除含N比例大于5%的Reads;(8)用Tophat软件进行比对,比对时调用软件Bwtie 2,再用Integrative Genomics Viewer将比对文件可视化;(9)用HTseq-count软件计算每个基因的reads数量即count值,用count值表示基因表达量。
-
运用Deseq 2进行不同样品间差异表达基因定量分析。采用Benjamini-Hochberg方法对得到的P值进行校正,校正后的P值,即FDR(false discovery rate),作为差异表达基因筛选的关键指标。将得到的FDR<0.01,且差异倍数FC(foldchange)≥2作为差异表达基因的筛选标准。
-
将筛选得到的差异基因用PANTHER在线工具(
http://www.pantherdb.org/ )对差异基因进行Gene Ontology (GO)富集分析,用特定的GO terms给差异表达基因的表达模式注释,包括分子功能(MF)、生物过程(BP)、细胞组分(CC)。用Kobas和Mapman软件对差异基因进行KEGG富集分析,注释文件下载于https://Mapman.gabipd.org/Mapmanstore 。 -
分别将0.1 g的d17和ZH11水稻种子冷冻干燥,将干燥后的样品研磨成粉末,置于2 mL离心管中。向样品中加入1 mL ψ=70%的甲醇,4 ℃条件下进行低温萃取。每隔0.5 h涡旋1次,涡旋3次后于4 ℃条件下过夜。将萃取的样品于12 000 g,4 ℃条件下离心10 min,用注射器过滤(SCAA-104, 0.22 μm, 上海安普)进代谢样品检测瓶。将准备好的样品上样到LC-MS (ABI 4000Q-TRAP)进行检测,具体参数参考文献[23]。获得代谢的原始数据后,首先将LC-MS中的MIM数据和MRM数据进行去重整合,得到低分辨质谱的数据,将低分辨质谱的数据与LC-MS的数据进行去重整合,建立代谢数据库。其次,将代谢数据库中物质的保留时间和二级质谱碎片与标准品进行比对,确定物质种类与结构。通过与Metlin、Massbank、HMDB等公开质谱数据库进行比对,确定数据库中未鉴定物质的种类和结构。最后依据其保留时间和特征碎片,将仍未鉴定的物质进行人工解谱,推测物质结构。
-
为了分析水稻独脚金内酯合成缺失突变体d17及其野生型栽培品种ZH11中基因表达差异,对生长30 d的d17突变体及野生型ZH11水稻的根和叶进行了转录组测序,每个样品设2次生物学重复。转录组原始数据质控结果(表1)表明,所有样本的Q30均大于94%,表明测序数据质量可靠。在过滤掉低质量reads后,平均每个样本获得47.87 M reads 。将各样本的clean reads与日本晴(MSU 7.0)参考基因组进行比对后显示,95%以上的reads均可匹配到参考基因组上。转录组分析结果显示,从d17和ZH11的根和叶中共检测到44 432个表达基因,其中在根中表达的基因有41 694 (93.84%)个,在叶中表达的基因有38 692 (87.08%)个。
表 1 8个样品的转录组测序原始和过滤数据
Table 1. Raw and clean reads of 8 samples used for transcriptome sequencing
Name Q20/% Q30/% Raw reads/M Clean reads/M Mapped reads/M Reads mapped/% Expressed gene/NO. d17 leaf rep1 98.31 94.94 52.41 52.14 48.70 97.64 34 446 d17 leaf rep2 98.15 94.58 50.52 50.21 46.66 97.55 34 244 ZH11 leaf rep1 97.98 94.36 45.00 44.75 41.58 97.43 33 784 ZH11 leaf rep2 97.98 94.15 47.45 47.11 43.83 97.52 34 190 Gene expressed in leaf 38 692 d17 root rep1 98.16 94.6 43.87 43.58 40.05 96.03 36 983 d17 root rep2 98.03 94.28 51.47 51.12 46.34 94.98 37 463 ZH11 root rep1 98.08 94.43 47.40 47.10 43.26 96.08 37 475 ZH11 root rep2 98.14 94.58 47.21 46.92 42.56 94.98 37 096 Gene expressed in root 41 694 -
为了获得d17和ZH11的根和叶中的差异表达基因,以|log2(fold change)| ≥ 1,FDR < 0.01为筛选条件[24],从d17水稻的根和叶中共获得248和441个差异表达基因(图1-A),去重后共获得610个差异基因。与野生型ZH11相比,d17的叶中有102个基因上调表达,146个基因下调表达;根中分别有127个基因上调表达,314个基因下调表达(图1-A)。从差异表达基因的数量上来看,根中下调表达的差异基因数量是叶中的2.15倍,而上调表达的基因数量与叶中接近。此外,这些差异表达基因在根和叶的分布相对独立,根和叶共有的差异表达基因只有38个(图1-B),说明独脚金内酯调控水稻根和叶发育的基因不同。
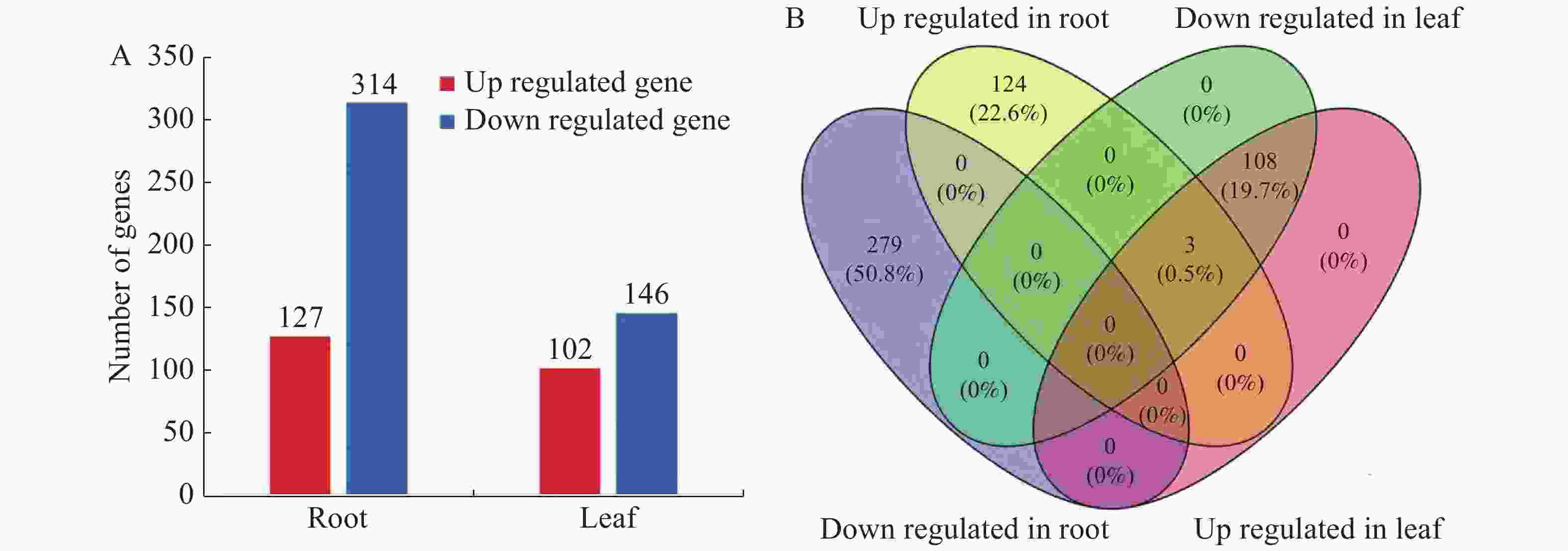
图 1 独脚金内酯d17突变体中的差异表达基因数量与分布
Figure 1. Number and distribution of differential expressed genes in d17 mutant
-
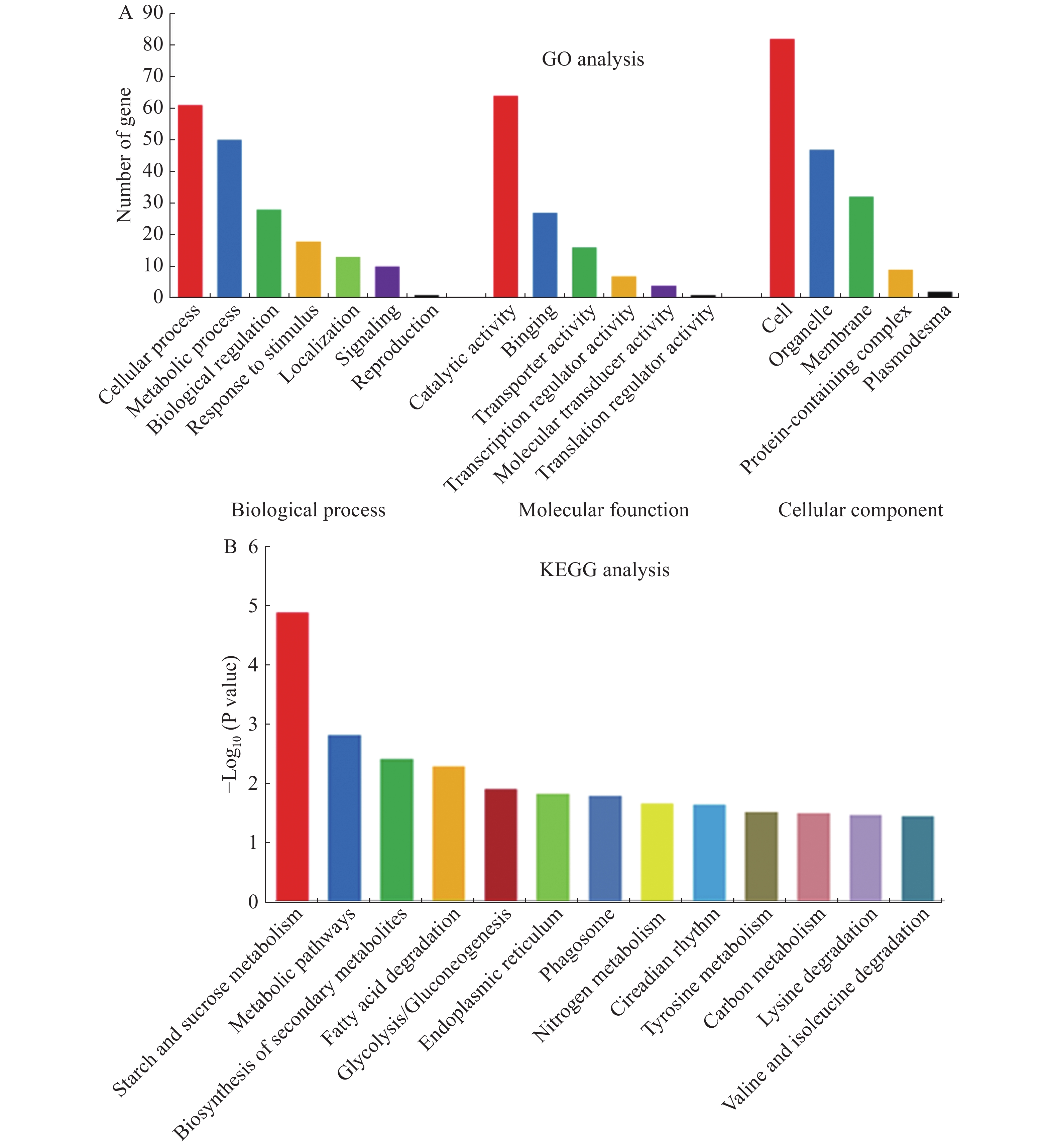
为了了解独脚金内酯d17突变体根和叶中差异基因的分布特征,对所有的610个差异基因进行GO注释和富集分析。结果显示,共有303个差异基因被注释,并主要富集到18个GO项中(图2)。在生物学过程(BP)这一项中,差异基因主要在细胞过程(cellular process)和代谢过程(metabolic process)中富集。在分子功能(MF)中,差异基因则大多富集在催化活性(catalytic activity)中。在细胞组分(CC)中,差异基因则主要在细胞(cell)和细胞器(organelle)中富集(图2-A)。由于大多数基因在根和叶中都是特异表达的,因此本研究对根和叶中的差异基因单独进行了GO分析。比较分析结果表明,根和叶中差异基因富集结果类似,主要富集于细胞、催化和代谢过程中。GO分析的结果说明,虽然水稻根和叶中的差异基因不同,但其功能相似。
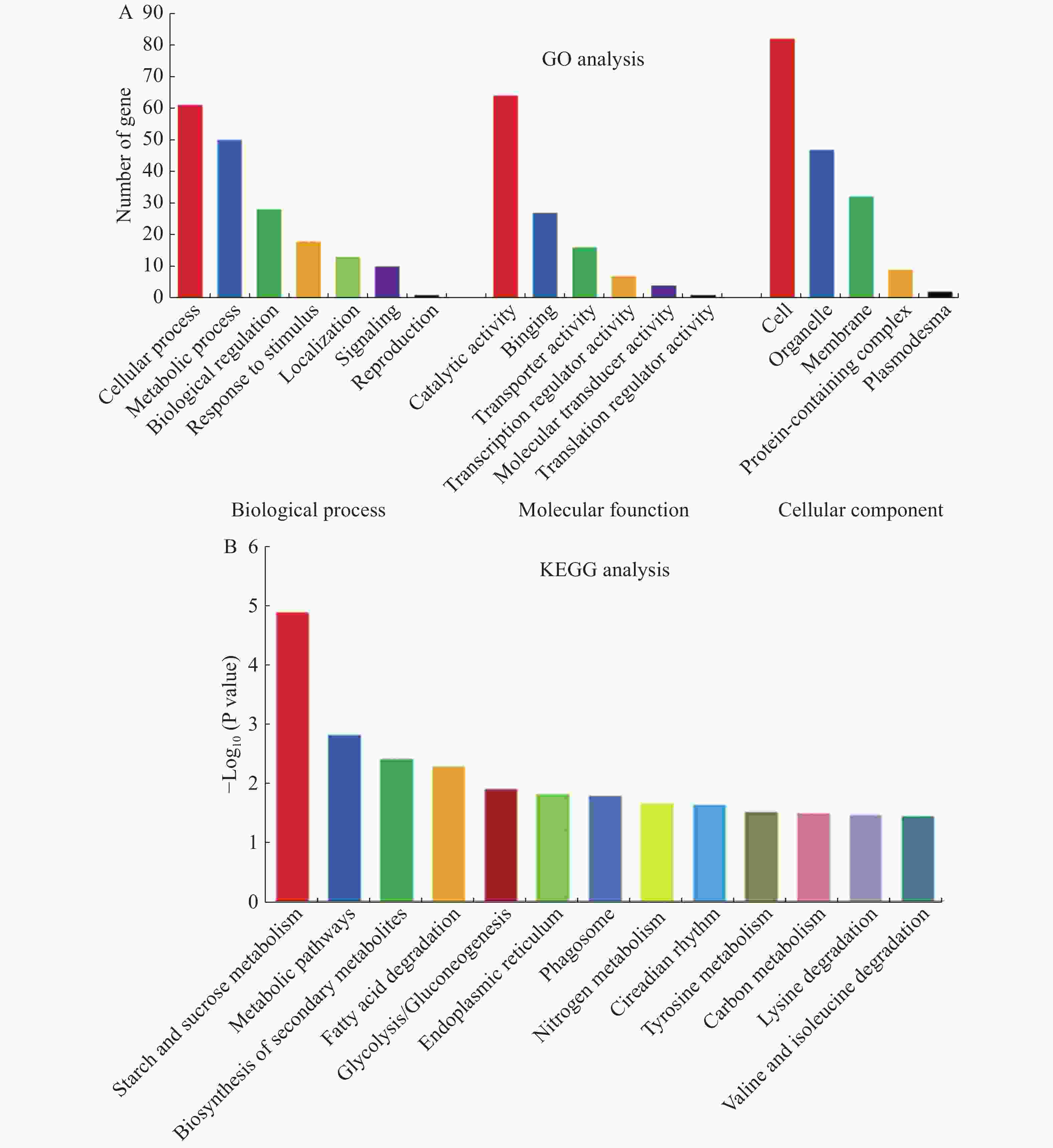
图 2 独脚金内酯d17突变体根和叶中差异表达基因的GO和KEGG富集分析
Figure 2. GO and KEGG enrichment analysis with differential expressed genes in roots and leaves of d17
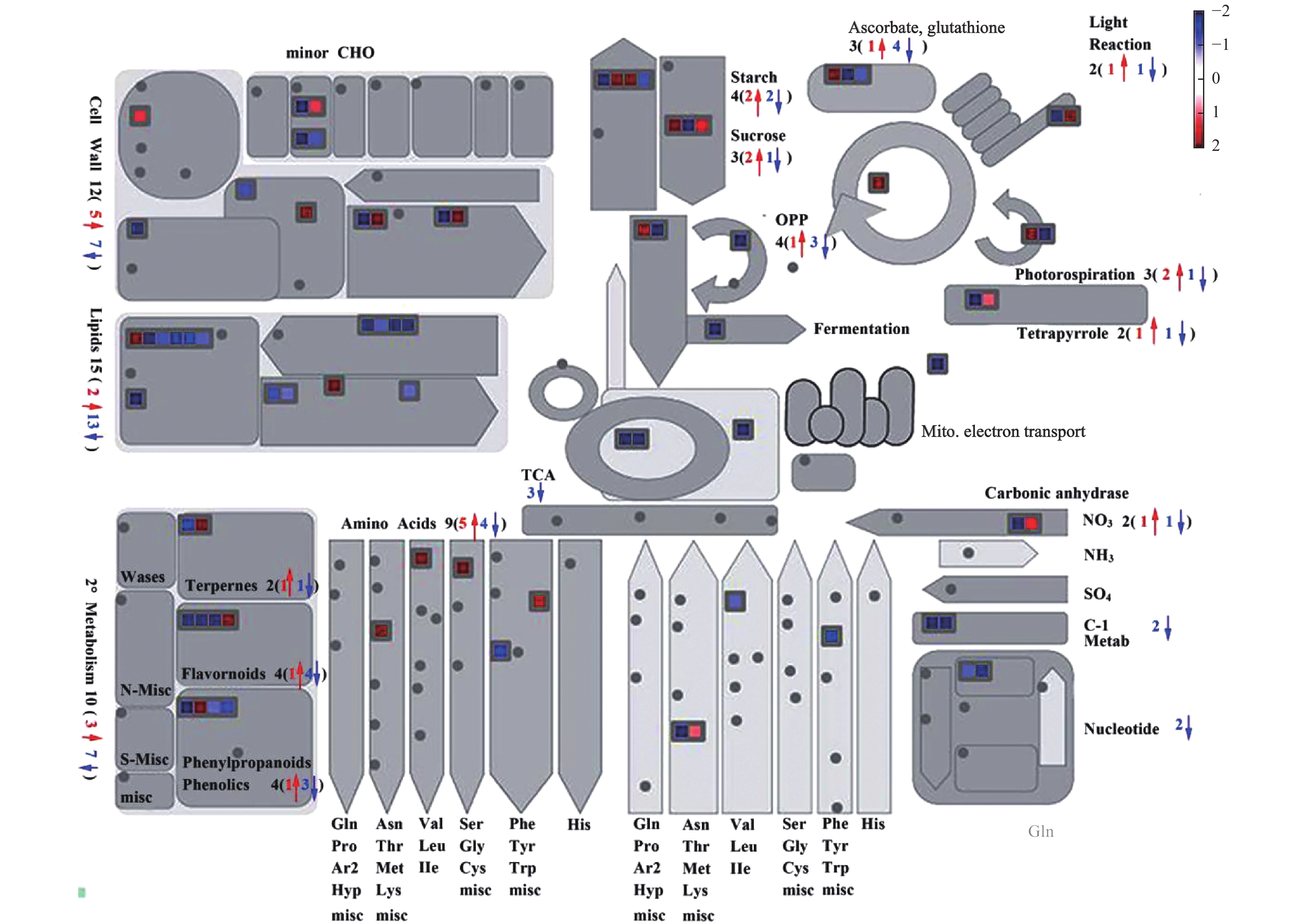
为了进一步了解独脚金内酯对水稻代谢过程的影响,对获得的610个差异基因进行了KEGG pathway的注释和富集分析。结果显示610个差异基因中有337个得到注释,并富集到89个代谢途径中。其中,显著富集的途径有13个(P < 0.05),主要包括淀粉和蔗糖代谢、脂质代谢和次生代谢(图2-B)。将水稻根和叶中注释到的203和153个差异基因分别进行KEGG pathway富集分析,结果分别富集到8和18条代谢路径。其中,糖类、脂质和次生代谢在根和叶中均显著富集,根中的植物激素信号转导、α−亚麻酸代谢等也发生了富集,而在叶中则氨基酸代谢发生了富集等。综上所述,独脚金内酯的缺失不仅影响水稻的糖类、脂质等初级代谢物的代谢过程,对水稻中的次生代谢过程也有影响。
-
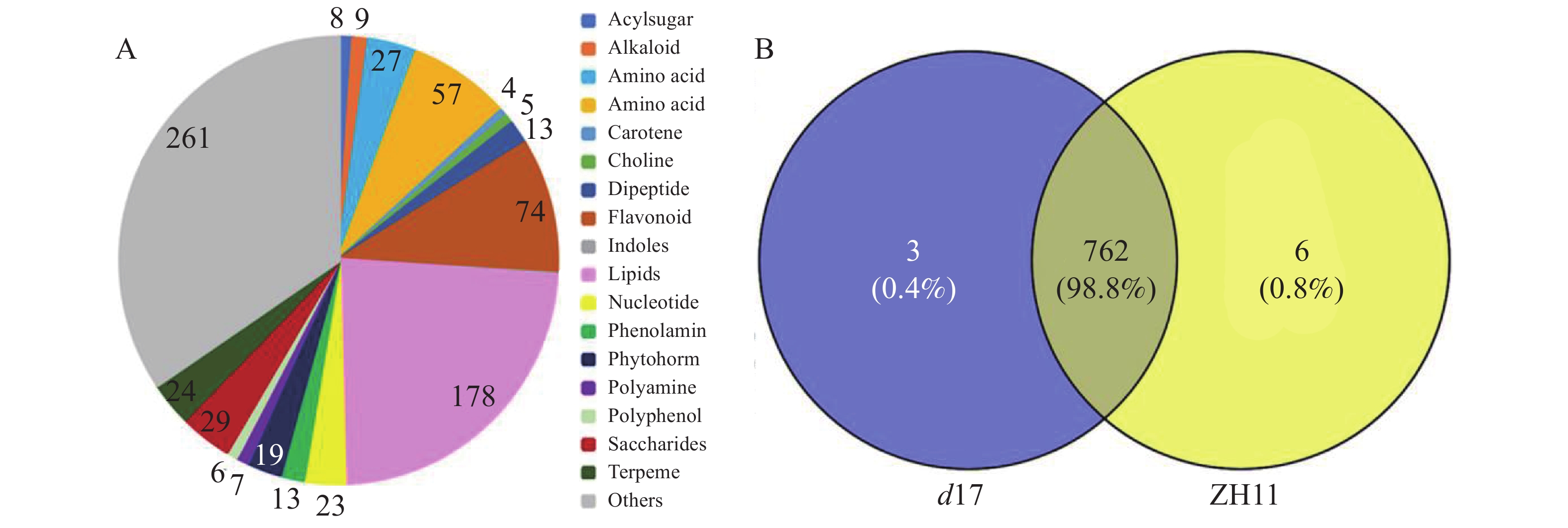
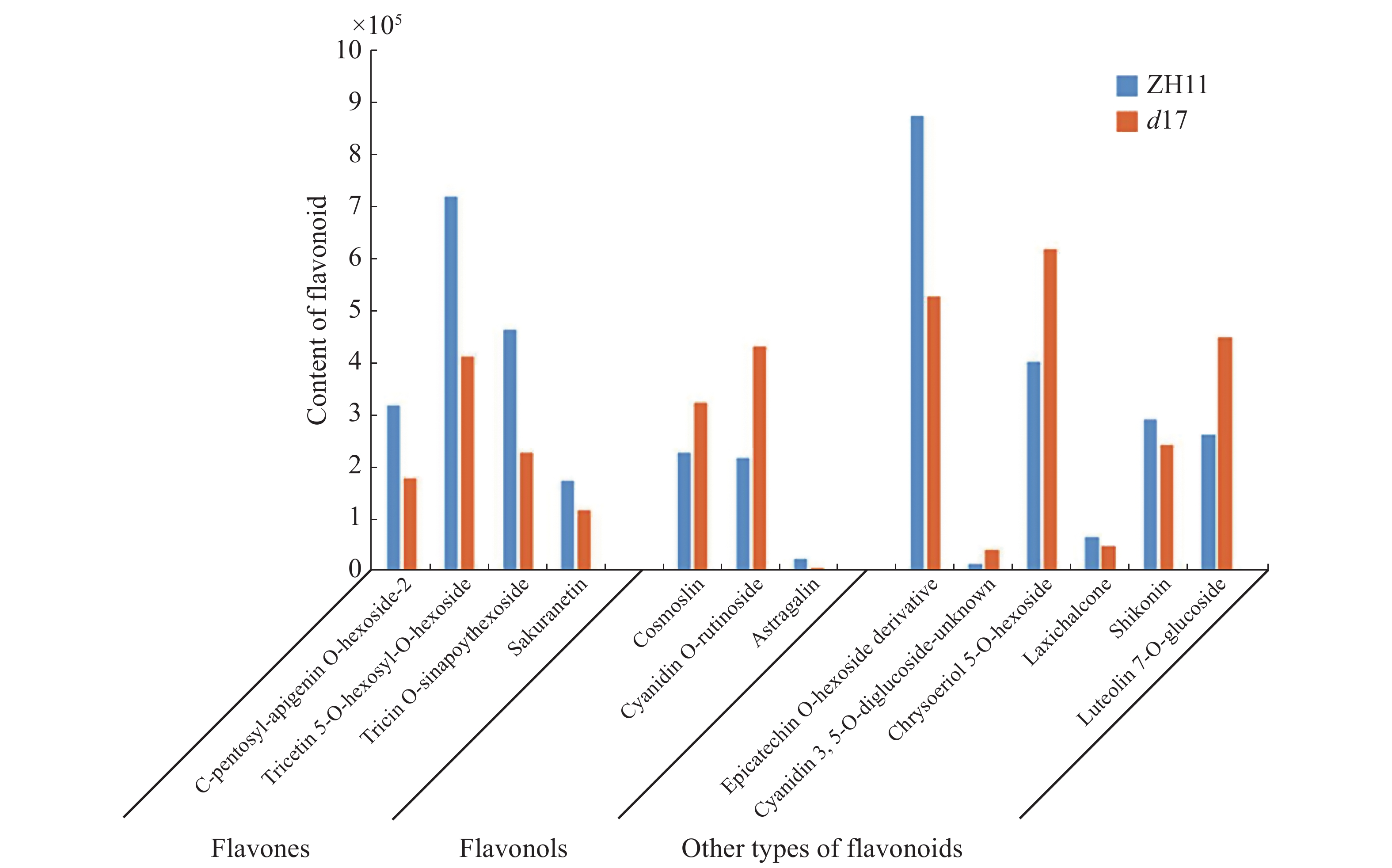
为了解独脚金内酯对水稻代谢的影响,对d17和ZH11的水稻种子进行代谢组检测分析。将不同样本的代谢物去重后,共检测到781个代谢物。将781个代谢物划分为20个类别,其中数量最多的几类物质是脂质(178个,22.79%)、黄酮类(74个,9.48%)以及氨基酸和氨基酸衍生物(85个,10.88%)(图3-A)。其中,在d17和ZH11中分别检测到765和768个代谢物,两者共有的代谢物有762个(图3-B)。为了分析独脚金内酯对水稻代谢过程的影响,以P < 0.05,|log2(fold change)| > 0.75为筛选条件[25],对所有检测到的代谢物进行定量筛选后,结果在d17突变体中共得到52个差异代谢物,包括脂质(20个, 38%)、黄酮(5个, 10%)、核酸(3个, 6%)等。差异代谢物含量的热图分析结果显示,在d17突变体中,脂质的含量大多下降,包括脂肪酸(FA)、甘油磷脂酰乙醇胺(GPE)、溶血磷脂酰胆碱(LPC)、溶血磷脂酰乙醇胺(LysoPE)和磷酸酯(Phosphate)等(图4)。此外,黄芪素(Astragalin)、C−戊糖芹菜素−O−己糖苷2(C-pentosyl-apigenin-O-hexoside 2)、三羟甲基甘氨酸−O−芥子基己糖苷(tricin O-sinapoylhexoside)和丙烯腈5−O−己糖苷(tricetin 5-O-hexosyl-O-hexoside)等黄酮类物质的含量下降也极为明显(图4)。
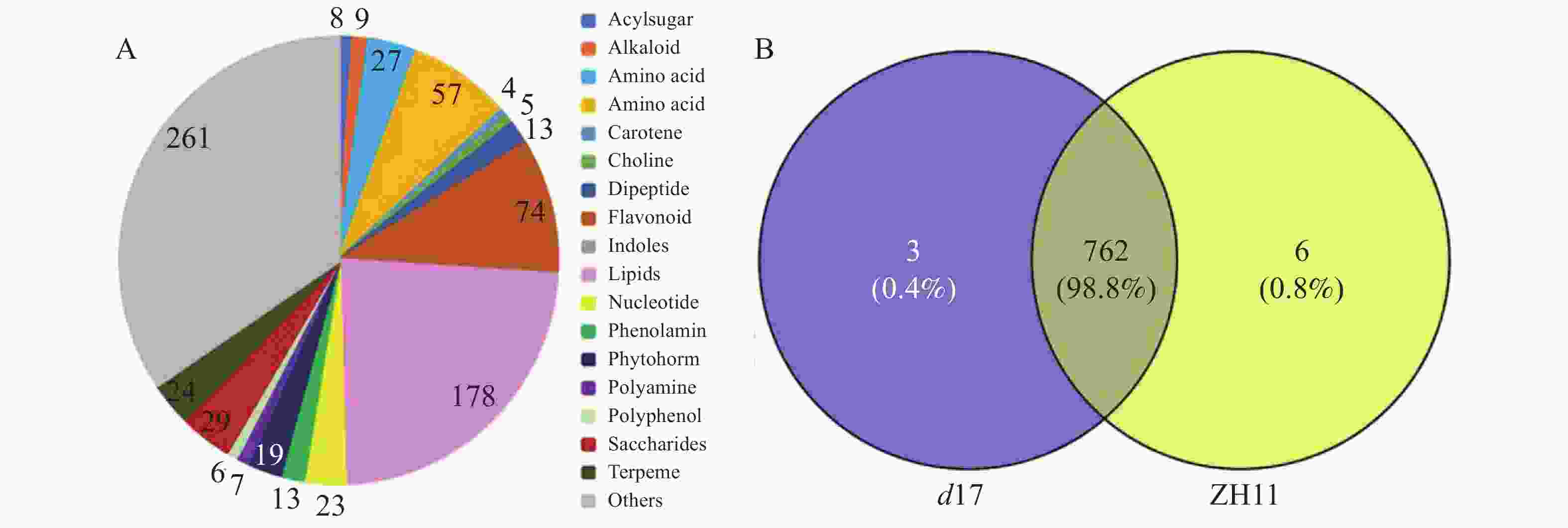
图 3 独脚金内酯d17突变体中的代谢物
Figure 3. The metabolites in the strigolactone d17 mutant
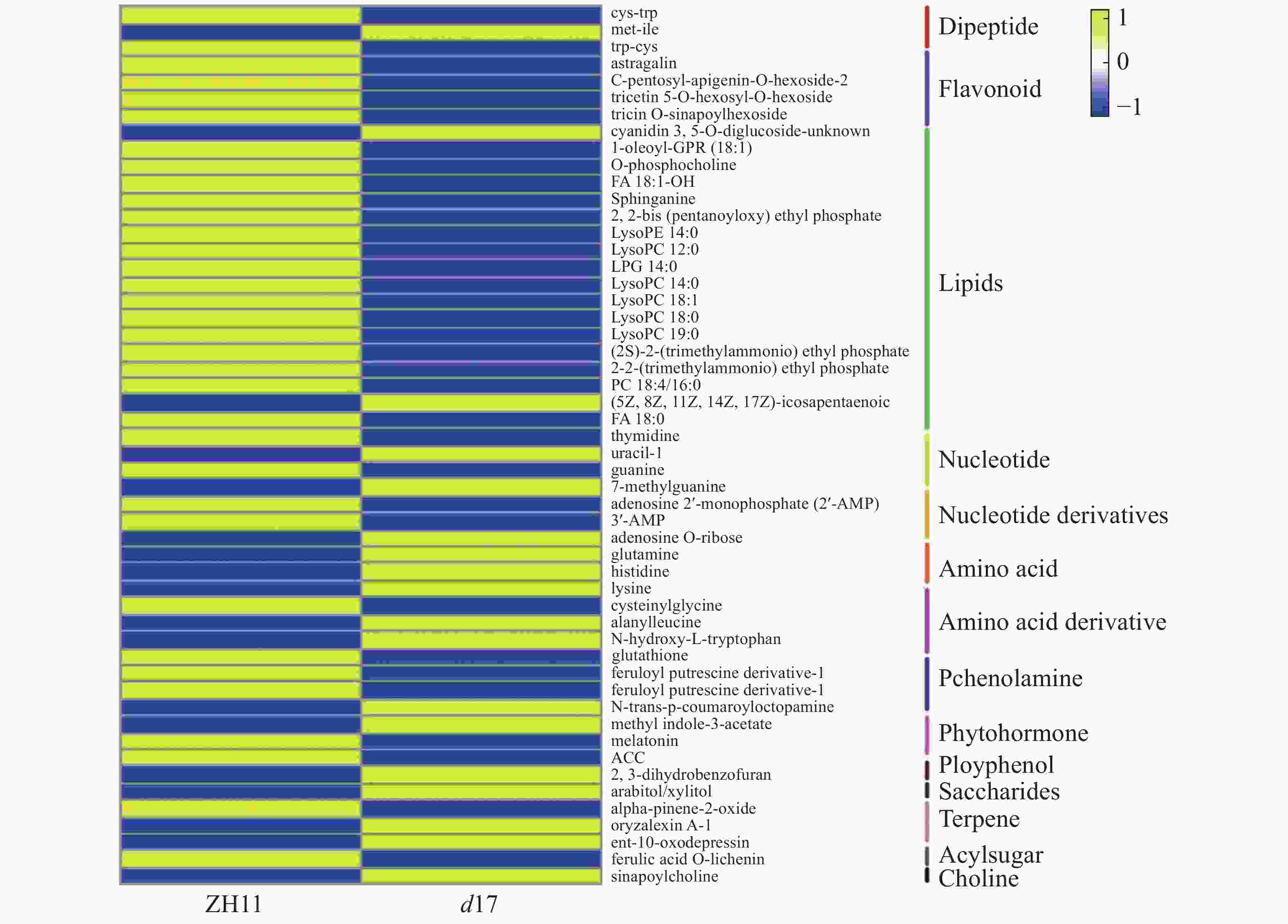
图 4 52个独脚金内酯d17突变体中含量显著变化的代谢物热图
Figure 4. Heatmap showed the contents of 52 metabolites were significantly different in strigolactone d17 mutants
-
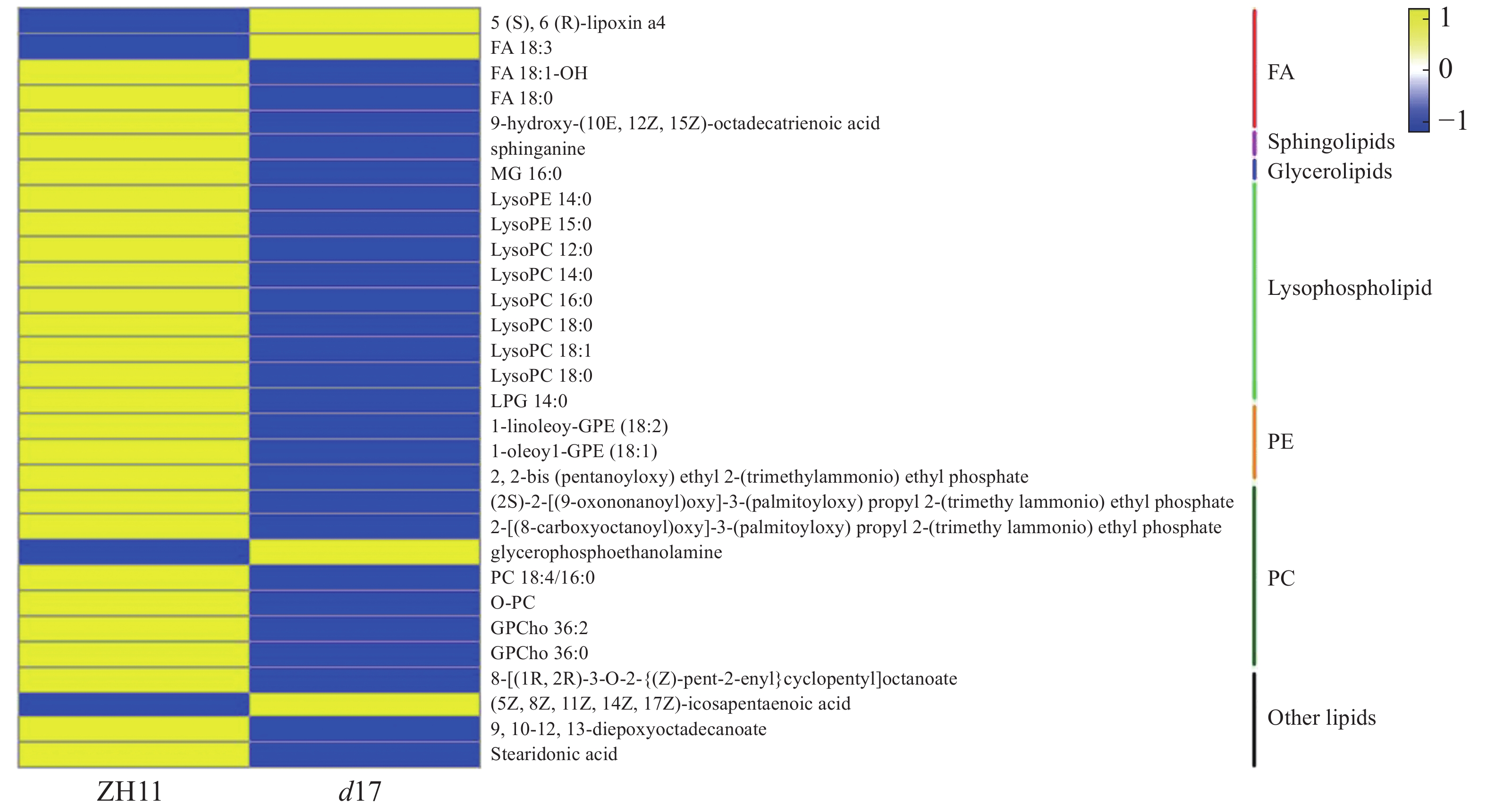
为了进一步了解独脚金内酯对水稻脂质含量的影响,将独脚金内酯突变体d17中显著变化(P < 0.05)的脂质单独提出,最终获得30个脂质物质(图5)。热图分析结果表明,溶血磷脂(Lysophospholipid, 9个)、磷脂酰胆碱(PC, 4个)、甘油磷脂酰乙醇胺(GPE, 2个)、脂肪酸(FA, 2个)等脂质的含量下降;而甘油磷酸乙醇胺(GPE, 1个)等4个脂质的含量上升。从结果可以看出,独脚金内酯的缺失会导致水稻中脂质的含量下降。对d17中610个差异基因进行通路分析显示,d17突变体中共有15个差异基因在脂质代谢中富集,且绝大多数(13/15)下调表达(图6)。其中,与磷脂合成相关的绝大多数基因(5/6)的表达下调,包括3个二酰基甘油激酶(LOC_Os12g12260.1,LOC_Os02g54650.1和LOC_Os01g57420.1)、1个胆碱激酶(LOC_Os01g51920.1)和1个磷脂酰丝氨酸脱羧酶(LOC_Os01g72940.1)。因此笔者推测独脚金内酯的缺失会导致水稻中脂质尤其是磷脂含量的下降。
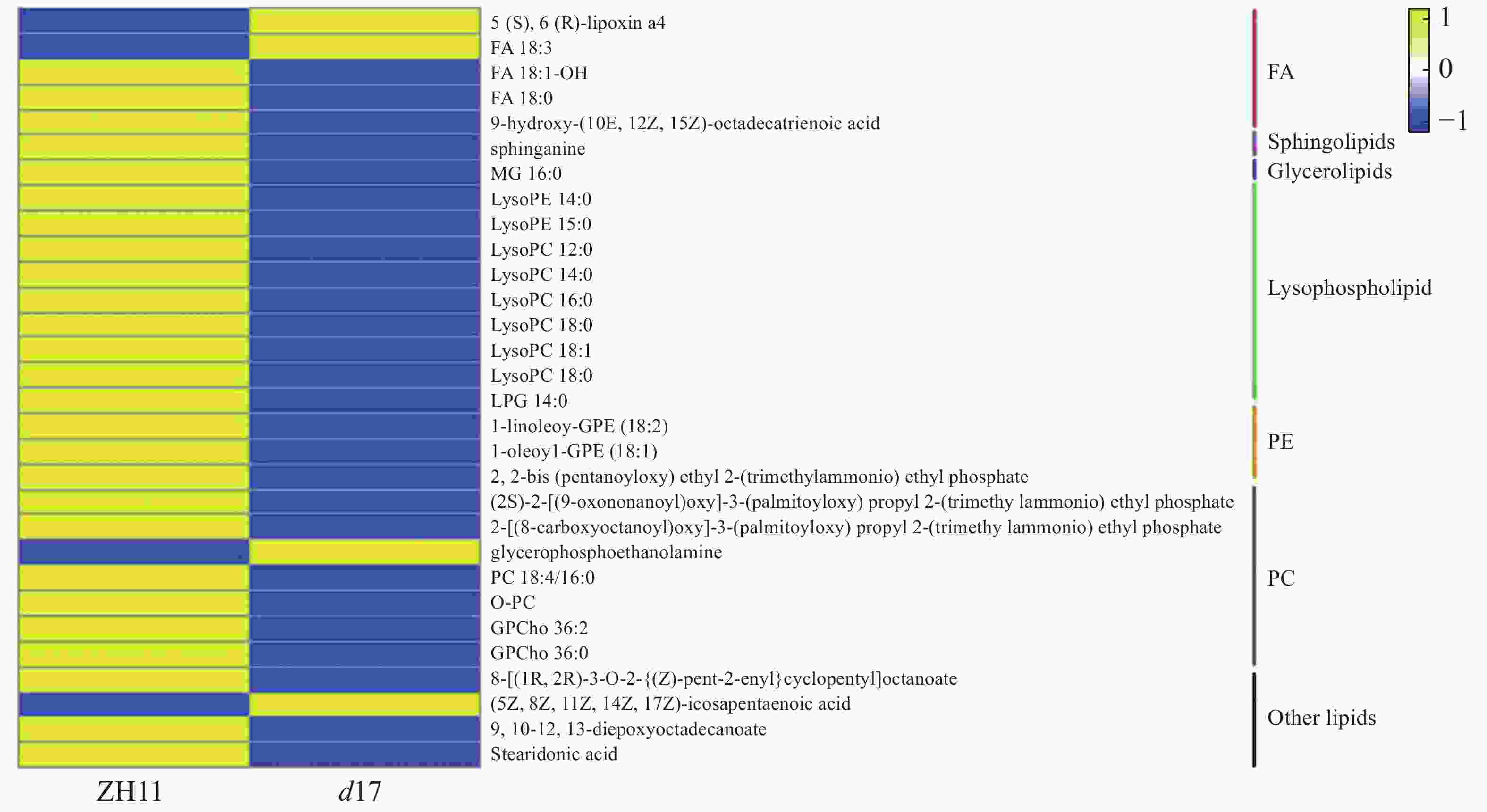
图 5 独脚金内酯d17突变体中含量差异显著的脂质的热图分析
Figure 5. Heat map analysis of the contents of lipids significantly changed in strigolactone d17 mutants
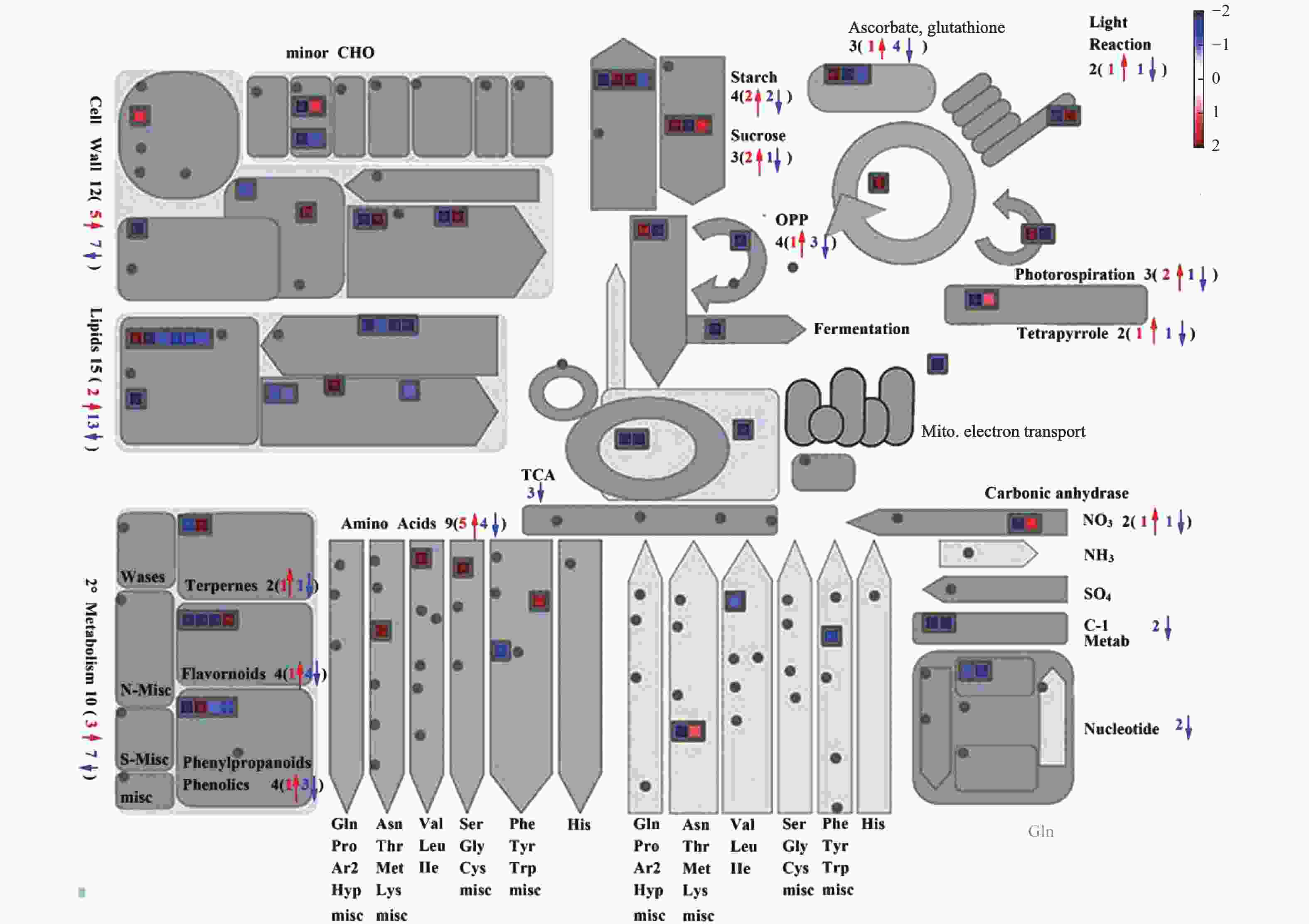
图 6 独脚金内酯d17突变体中差异表达基因富集的代谢通路
Figure 6. Enrichment of metabolic pathways of differentially expressed genes in strigolactone d17 mutants
-
将独脚金内酯突变体d17中显著变化(P < 0.05)的黄酮单独提出,最终获得13个黄酮类(Flavonoid)物质(图7)。其中,4种黄酮(Flavones)的含量下降,包括C−戊糖芹菜素−O−己糖−2、5−O−己糖基−O−己糖苷五羟黄酮、O−芥子基己糖苷麦黄酮和樱红素(图7)。大波斯菊苷和矢车菊素−O−芸香糖苷和黄芪苷,3种黄酮醇(Flavonols)的含量升高(图7)。另外,矢车菊素−3, 5−O−双葡萄糖苷等其他6种黄酮的含量变化无明显规律(图7)。通路分析结果表明,d17中黄酮(Flavones)代谢相关的4个差异基因中有3个差异基因的表达下降(图6)。将水稻根和叶中黄酮代谢相关的基因列出,并以|log2(fold change)| > 4为条件进行筛选,共发现13个差异基因(表2)。与黄酮醇代谢相关的基因有5个,其中与黄酮醇合成相关的4个基因上调表达,1个下调表达。结果表明,基因的表达变化与黄酮和黄酮醇的含量变化相吻合,因此,笔者推断独脚金内酯的缺失影响水稻中黄酮和黄酮醇的代谢。
表 2 独脚金内酯d17突变体中黄酮代谢相关基因的变化
Table 2. The changes of flavonoid metabolism-related genes in d17
MSU_Locus Gene name or annotation log2 (fold change) LOC_Os03g18030.2 flavonol synthase/flavanone −8.61 LOC_Os04g57160.1 FLS_SOLTU flavonol synthase −5.25 LOC_Os06g07923.2 flavonol synthase 4.3 LOC_Os01g25010.1 flavonol synthase 5.06 LOC_Os01g27490.1 flavonol synthase 4.79 LOC_Os02g52840.1 flavonol synthase 4.62 LOC_Os04g23940.1 naringenin-chalcone synthase 4.78 LOC_Os06g08060.1 leucocyanidin oxygenase −4.22 LOC_Os12g16410.3 isoflavone reductase 26.43 LOC_Os04g56700.1 flavonone-3-hydroxylase −4.41 LOC_Os02g28220.1 forward transferase protein 6.87 LOC_Os06g05284.1 reverse transferase protein −4.22 LOC_Os11g03510.1 1-aminocyclopropane-1-carboxylate oxidase 1 −4.22 LOC_Os02g28410.1 5-aromatic acyltransferase 5.37 
图 7 独脚金内酯d17突变体及ZH11中黄酮类代谢物的含量
Figure 7. Contents of flavonoid metabolites in the strigolactone d17 mutant
-
独脚金内酯作为一种新型植物激素,具有调控水稻分枝、分蘖、株高、根的伸长等多种作用[23,24]。目前,独脚金内酯在水稻中的研究主要聚焦在其对水稻分蘖、产量等性状的影响上[7,25],对水稻体内初生和次生代谢影响的相关研究较少。本研究对独脚金内酯合成突变体d17及其野生型ZH11进行转录组和代谢组学的分析,发现水稻中脂质和黄酮类的代谢过程受到独脚金内酯缺失的影响。
独脚金内酯合成突变体d17中差异表达基因的GO富集分析结果表明,独脚金内酯的缺失会影响水稻的细胞和代谢过程(图2-B),且对水稻中绝大部分脂质代谢相关基因的表达有抑制作用(图6)。代谢组的分析发现,独脚金内酯的缺失使水稻中脂质尤其是磷脂的含量下降(图5)。磷脂为细胞膜的重要组成成分[26],有研究结果表明,在缺磷和富磷的情况下独脚金内酯使拟南芥中磷脂的含量发生改变[27]。本研究发现,含量下降的磷脂中绝大多数是溶血磷脂(图5),而溶血磷脂常以淀粉和脂质复合物的形式存在[28],且独脚金内酯可以影响柑橘中淀粉的含量[29]。此外,独脚金内酯还影响水稻根和叶中淀粉和蔗糖相关基因的表达(图2)。有研究结果表明,独脚金内酯的缺失会抑制水稻中可溶性糖的积累[16],因此,笔者推测独脚金内酯可能协同调控了淀粉和脂质的代谢。
独脚金内酯除了影响水稻的脂质代谢外,其缺失还影响水稻体内黄酮和黄酮醇的代谢(图7)。独脚金内酯的缺失导致水稻中黄酮类物质的含量下降,特别是黄酮的含量下降(图7),同时大多数黄酮类物质代谢相关基因的表达量下降(图6)。前人研究结果表明,独脚金内酯可以使拟南芥中黄酮醇合成酶的含量增加,进而诱导黄酮醇的积累[19]。此外,独脚金内酯能够提高低磷低氮条件下和正常条件下拟南芥中花青素的含量[30-31]。在葡萄中,独脚金内酯能够抑制ABA对葡萄中花青素积累的促进作用[20]。因此,笔者推测独脚金内酯可以促进水稻体内黄酮类的积累,而其缺失会抑制水稻中黄酮类的代谢。独脚金内酯的缺失使得水稻体内多数的黄酮醇含量升高(图7),且使多数黄酮醇合成相关基因的表达上升(表2),因此,笔者推测独脚金内酯的缺失对黄酮醇的合成有促进作用。
在本研究中,对独脚金内酯d17合成突变体及其野生型ZH11的代谢组和转录组的分析结果表明,独脚金内酯可以抑制水稻的脂质代谢和黄酮代谢,为独脚金内酯调控水稻的初生和次生代谢提供了实验证据和基础。
Transcriptome and Metabolome Reveal that Loss of Strigolactone Affects Lipid and Flavonoid Metabolism in Rice Roots and Leaves
-
摘要: 对水稻的独脚金内酯合成突变体d17及野生型水稻ZH11进行转录组研究,结果表明,在d17的根和叶中的248和441个差异表达基因在脂质代谢路径上出现富集。代谢组检测结果表明,脂质和黄酮类物质的含量在d17突变体和ZH11中表现出显著差异。进一步研究发现,d17中的溶血磷脂和黄酮的含量显著降低,相关基因的表达水平下降,表明独脚金内酯参与调控水稻体内的黄酮和脂质代谢。研究结果可为研究独脚金内酯调控水稻体内次生代谢及生长发育提供参考。Abstract: Transcriptome-based analysis of the strigolactone-deficient mutant d17 of rice (Oryza sativa L ssp. japonica) and its wild-type ZH11 (O. sativa L. ssp. japonica cv. Zhonghua11) were performed. A total of 248 and 441 differentially expressed genes were found in the roots and leaves of d17, which were enriched on the lipid metabolism pathway. Metabolomics analysis revealed that lipid and flavonoid contents were significantly different between the d17 mutant and ZH11. Further research found that the contents of lysophospholipid and flavonoids in the d17 was decreased significantly, and that the expression levels of related genes were decreased. Our results revealed that strigolactone was involved in regulating of flavonoids and lipid metabolism in rice, which provides references for further research on strigolactone in regulating of secondary metabolism and growth in rice.
-
Key words:
- strigolactone /
- rice /
- transcriptome /
- metabolome /
- flavonoids /
- lipids
-
图 2 独脚金内酯d17突变体根和叶中差异表达基因的GO和KEGG富集分析
A:d17突变体中差异表达基因的GO富集分析;B:d17突变体中差异表达基因的KEGG富集分析(P < 0.05)。
Fig. 2 GO and KEGG enrichment analysis with differential expressed genes in roots and leaves of d17
A: GO enrichment analysis with differential expressed genes in d17;B: KEGG pathway enrichment analysis of differentially expressed genes in d17 (P < 0.05).
图 3 独脚金内酯d17突变体中的代谢物
A:独脚金内酯d17突变体和野生型ZH11中代谢物种类分布;781个代谢物被分为21类,主要包括脂质、黄酮以及氨基酸和氨基酸衍生物。B:独脚金内酯d17突变体和野生型ZH11中代谢物维恩图。
Fig. 3 The metabolites in the strigolactone d17 mutant
A: Distribution of metabolite species in strigolactone d17 mutant and wild-type ZH11;The 781 metabolites were divided into 21 categories, mainly including lipids, flavones, and amino acids and amino acid derivatives. B: Venn diagram of strigolactone d17 mutant and wild-type ZH11.
图 4 52个独脚金内酯d17突变体中含量显著变化的代谢物热图
52个代谢物被分为14类,主要包括脂质、黄酮和核酸。蓝色代表与ZH11相比,代谢物的含量在d17突变体中降低,黄色表示升高。
Fig. 4 Heatmap showed the contents of 52 metabolites were significantly different in strigolactone d17 mutants
All of 52 metabolites were divided into 14 categories, mainly including lipids, flavonoids and nucleotide. Blue indicates the decreased metabolites in d17 mutant compared with ZH11, and yellow represents increased metabolites.
图 5 独脚金内酯d17突变体中含量差异显著的脂质的热图分析
蓝色代表代谢物的含量在d17突变体中降低,黄色表示升高。
Fig. 5 Heat map analysis of the contents of lipids significantly changed in strigolactone d17 mutants
Blue indicates the decreased metabolites in the d17 mutant, and yellow represents increased metabolites.
图 6 独脚金内酯d17突变体中差异表达基因富集的代谢通路
蓝色表示基因下调,红色表示基因上调。
Fig. 6 Enrichment of metabolic pathways of differentially expressed genes in strigolactone d17 mutants
Blue means genes down-regulated and red means up-regulated.
图 7 独脚金内酯d17突变体及ZH11中黄酮类代谢物的含量
所有代谢物的含量均为两次生物学重复的平均值。
Fig. 7 Contents of flavonoid metabolites in the strigolactone d17 mutant
The average of two biological replicates was merged together as the content of metabolites.
表 1 8个样品的转录组测序原始和过滤数据
Table 1 Raw and clean reads of 8 samples used for transcriptome sequencing
Name Q20/% Q30/% Raw reads/M Clean reads/M Mapped reads/M Reads mapped/% Expressed gene/NO. d17 leaf rep1 98.31 94.94 52.41 52.14 48.70 97.64 34 446 d17 leaf rep2 98.15 94.58 50.52 50.21 46.66 97.55 34 244 ZH11 leaf rep1 97.98 94.36 45.00 44.75 41.58 97.43 33 784 ZH11 leaf rep2 97.98 94.15 47.45 47.11 43.83 97.52 34 190 Gene expressed in leaf 38 692 d17 root rep1 98.16 94.6 43.87 43.58 40.05 96.03 36 983 d17 root rep2 98.03 94.28 51.47 51.12 46.34 94.98 37 463 ZH11 root rep1 98.08 94.43 47.40 47.10 43.26 96.08 37 475 ZH11 root rep2 98.14 94.58 47.21 46.92 42.56 94.98 37 096 Gene expressed in root 41 694  下载: 导出CSV
下载: 导出CSV
表 2 独脚金内酯d17突变体中黄酮代谢相关基因的变化
Table 2 The changes of flavonoid metabolism-related genes in d17
MSU_Locus Gene name or annotation log2 (fold change) LOC_Os03g18030.2 flavonol synthase/flavanone −8.61 LOC_Os04g57160.1 FLS_SOLTU flavonol synthase −5.25 LOC_Os06g07923.2 flavonol synthase 4.3 LOC_Os01g25010.1 flavonol synthase 5.06 LOC_Os01g27490.1 flavonol synthase 4.79 LOC_Os02g52840.1 flavonol synthase 4.62 LOC_Os04g23940.1 naringenin-chalcone synthase 4.78 LOC_Os06g08060.1 leucocyanidin oxygenase −4.22 LOC_Os12g16410.3 isoflavone reductase 26.43 LOC_Os04g56700.1 flavonone-3-hydroxylase −4.41 LOC_Os02g28220.1 forward transferase protein 6.87 LOC_Os06g05284.1 reverse transferase protein −4.22 LOC_Os11g03510.1 1-aminocyclopropane-1-carboxylate oxidase 1 −4.22 LOC_Os02g28410.1 5-aromatic acyltransferase 5.37
下载: 导出CSV
-
[1] TAKESHI I, KO S. Becoming a model plant: The importance of rice to plant science [J]. Trends Plant Sci, 2009, 1(3): 95 − 99. [2] 肖景华, 吴昌银, 袁猛, 等. 中国水稻功能基因组研究进展与展望[J]. 科学通报, 2015, 60(18): 1711 − 1712. [3] WANG B, SMITH M, LI J. Genetic regulation of shoot architecture [J]. Annu Rev Plant Biol, 2018, 69(25): 437 − 468. [4] FIEHN O. Metabolomics: the link between genotypes and phenotypes [J]. Plant Mol Biol, 2002, 48(1/2): 155 − 171. doi: 10.1023/A:1013713905833 [5] LIANG W H, SHANG F, LIN Q T, et al. Tillering and panicle branching genes in rice [J]. Gene, 2014, 537(1): 1 − 5. doi: 10.1016/j.gene.2013.11.058 [6] HU Z, YAN H, YANG J, et al. Strigolactones negatively regulate mesocotyl elongation in rice during germination and growth in darkness [J]. Plant Cell Physiol, 2010, 51(7): 1136 − 1142. doi: 10.1093/pcp/pcq075 [7] LIN H, WANG R, QIAN, Q, et al. DWARF27, an iron-containing protein required for the biosynthesis of strigolactones, regulates rice tiller bud outgrowth [J]. Plant Cell, 2009, 21(5): 1512 − 1525. doi: 10.1105/tpc.109.065987 [8] 李丛丛, 马小定, 马建. 一个新的水稻D17/HTD1基因等位突变体的分子鉴定[J]. 植物遗传资源学报, 2019, 20(5): 1255 − 1261. [9] ZOU J, ZHANG S, ZHANG W, et al. The rice HIGH‐TILLERING DWARF1 encoding an ortholog of Arabidopsis MAX3 is required for negative regulation of the outgrowth of axillary buds [J]. Plant J, 2006, 48(5): 687 − 698. doi: 10.1111/j.1365-313X.2006.02916.x [10] ARITE T, IWATA H, OHSHIMA K, et al. DWARF10, an RMS1/MAX4/DAD1 ortholog, controls lateral bud outgrowth in rice [J]. Plant J, 2007, 51(6): 1019 − 1029. doi: 10.1111/j.1365-313X.2007.03210.x [11] ZHANG Y, VAN DIJK AD, SCAFFIDI A, et al. Rice cytochrome P450 MAX1 homologs catalyze distinct steps in strigolactone biosynthesis [J]. Nat Chem Biol, 2014, 10(12): 1028 − 1033. doi: 10.1038/nchembio.1660 [12] ISHIKAWA S, MAEKAWA M, ARITE T, et al. Suppression of tiller bud activity in tillering dwarf mutants of rice [J]. Plant Cell Physiol, 2005, 46(1): 79 − 86. doi: 10.1093/pcp/pci022 [13] WANG W, LI Y, DANG P, et al. Rice secondary metabolites: structures, roles, biosynthesis, and metabolic regulation [J]. Molecules, 2018, 23(12): 3098. doi: 10.3390/molecules23123098 [14] ALI G, NEDA G. Flavonoids and phenolic acids: role and biochemical activity in plants and human [J]. J Med Plants Res, 2011, 5(31): 6697 − 6703. [15] DAYAN F E, CANTRELL C L, DUKE S O. Natural products in crop protection [J]. Bioorg Med Chem, 2009, 17(12): 4022 − 4034. doi: 10.1016/j.bmc.2009.01.046 [16] NASIR F, TIAN L, SHI S, et al. Strigolactones positively regulate defense against Magnaporthe oryzae in rice (Oryza sativa) [J]. Plant Physiol Bioch, 2019, 142(4): 106 − 116. [17] CHEN F, JIANG L, ZHENG J, et al. Identification of differentially expressed proteins and phosphorylated proteins in rice seedlings in response to strigolactone treatment [J]. Plos One, 2014, 9(4): e93947. doi: 10.1371/journal.pone.0093947 [18] YAMADA Y, FURUSAWA S, NAGASAKA S, et al. Strigolactone signaling regulates rice leaf senescence in response to a phosphate deficiency [J]. Planta, 2014, 240(2): 399 − 408. doi: 10.1007/s00425-014-2096-0 [19] KUMAR M, PANDYA-KUMAR N, KAPULNIK Y, et al. Strigolactone signaling in root development and phosphate starvation [J]. Plant Signal Behav, 2015, 10(7): e1045174. doi: 10.1080/15592324.2015.1045174 [20] ITO S, NOZOYE T, SASAKI E, et al. Strigolactone regulates anthocyanin accumulation, acid phosphatases production and plant growth under low phosphate condition in Arabidopsis [J]. Plos One, 2015, 10(3): e0119724. doi: 10.1371/journal.pone.0119724 [21] ZHANG Z, HU Q, LIU Y, CHENG P, et al. Strigolactone represses the synthesis of melatonin, thereby inducing floral transition in Arabidopsis thaliana in an FLC-dependent manner [J]. J Pineal Res, 2019, 67(2): e12582. [22] WALTON A, STES E, GOEMINNE G, et al. The response of the root proteome to the synthetic strigolactone GR24 in Arabidopsis [J]. Mol Cell Proteomics, 2016, 15(8): 2744 − 2755. doi: 10.1074/mcp.M115.050062 [23] LIU H, LI X, XIAO J, et al. A convenient method for simultaneous quantification of multiple phytohormones and metabolites: application in study of rice-bacterium interaction [J]. Plant Methods, 2012, 8(2): 2. [24] YOAV B DANIEL Y. The control of the false discovery rate in multiple testing under dependency [J]. Ann Stat, 2001, 29(4): 1165 − 1188. doi: 10.1214/aos/1013699998 [25] GAUVREAU K, PAGANO M. Why 5%[J] Nutrition, 1994, 10(1): 93 − 94. [26] 许佳妮, 邓丽莉, 曾凯芳. 磷脂酶D在果蔬采后逆境胁迫及衰老过程中的作用[J]. 食品工业科技, 2014, 36(5): 393 − 399. [27] VISMANS G, MEER T, LANGEVOORT O, et al. Low phosphate induction of plastidal stromules is dependent on strigolactones but not on the canonical strigolactone signaling component MAX2 [J]. Plant Physiol, 2016, 172(4): 2235 − 2244. doi: 10.1104/pp.16.01146 [28] TONG C, LIE L, DANIEL L E W, et al. Association mapping and marker development of genes for starch lysophospholipid synthesis in rice [J]. Rice Sci, 2016, 23(6): 287 − 296. doi: 10.1016/j.rsci.2016.09.002 [29] YONG Q, ZHENG N K, PEDRO G. Strigolactones restore vegetative and reproductive developments in Huanglongbing (HLB) affected, greenhouse-grown citrus trees by modulating carbohydrate distribution [J]. Sci Hortic-amstetdam, 2018, 237(1): 89 − 95. [30] THUSSAGUNPANIT J, NAGAI Y, NAGAE M, et al. Involvement of STH7 in light-adapted development in Arabidopsis thaliana promoted by both strigolactone and karrikin [J]. Biosci Biotechnol Biochem, 2017, 81(2): 292 − 301. doi: 10.1080/09168451.2016.1254536 [31] FERRERO M, PAGLIARANI C, NOVAK O, et al. Exogenous strigolactone interacts with abscisic acid-mediated accumulation of anthocyanins in grapevine berries [J]. J Exp Bot, 2018, 69(9): 2391 − 2401. doi: 10.1093/jxb/ery033 -

 点击查看大图
点击查看大图

计量
- 文章访问数: 820
- HTML全文浏览量: 518
- PDF下载量: 60
- 被引次数: 0
