


-
角蛋白是自然界中丰度最高的纤维状蛋白[1],也是已知最坚硬的生物材料之一,是羽毛、龟壳和禽爪等常见的表皮附属物的主要构成成分[2]。生产生活中每年都会产生百万吨以上富含角蛋白的生物副产品[3]。这些副产品中蛋白质含量很高,其中禽类羽毛中角蛋白质含量约占90 % [4],具有回收再利用的价值。若处理不当,则会污染大陆与海洋[5]。例如,海鸭(Anas platyrhynchos)养殖中产生的大量鸭毛若不进行妥善处理,不仅造成蛋白资源的浪费,还会污染环境,甚至传播疾病[6]。角蛋白富含半胱氨酸和甘氨酸。两分子半胱氨酸易形成分子间二硫键转化为胱氨酸。甘氨酸具有疏水性、硬度较高且不易降解的特点。这使得角蛋白难以被自然降解,且在水中的溶解度比大多数蛋白质都低[7]。角蛋白也难以被动物来源的胃蛋白酶、胰蛋白酶直接降解,需进一步加工后才能利用,这也导致羽毛经常被填埋或焚烧处理[8]。而角蛋白资源难以被有效开发利用,在增加环境压力的同时,也造成巨大的蛋白质资源浪费。利用酶法处理废弃的角蛋白资源绿色环保,且降解产物适宜应用于有高附加值的科技领域。目前已筛选出多种角蛋白酶,例如Huang等[9]发现一种S8家族角蛋白酶。Kang等[10] 发现S16、M16和M38等多个家族的角蛋白酶。已报道的多数角蛋白酶对于温度和离子浓度的耐受性较差。为适应工业化生产,需要筛选出耐高温、耐高盐的角蛋白酶。笔者从角蛋白的结构特征、降解机理以及降解产物的利用等方面综述了角蛋白降解的进展,并着重介绍了酶法降解角蛋白这一最具潜力的降解方式。
-
根据蛋白质二级结构可将角蛋白分为α−角蛋白和β−角蛋白。α−角蛋白主要存在于哺乳动物中,而β−角蛋白则主要存在于鸟类和爬行动物中[11]。
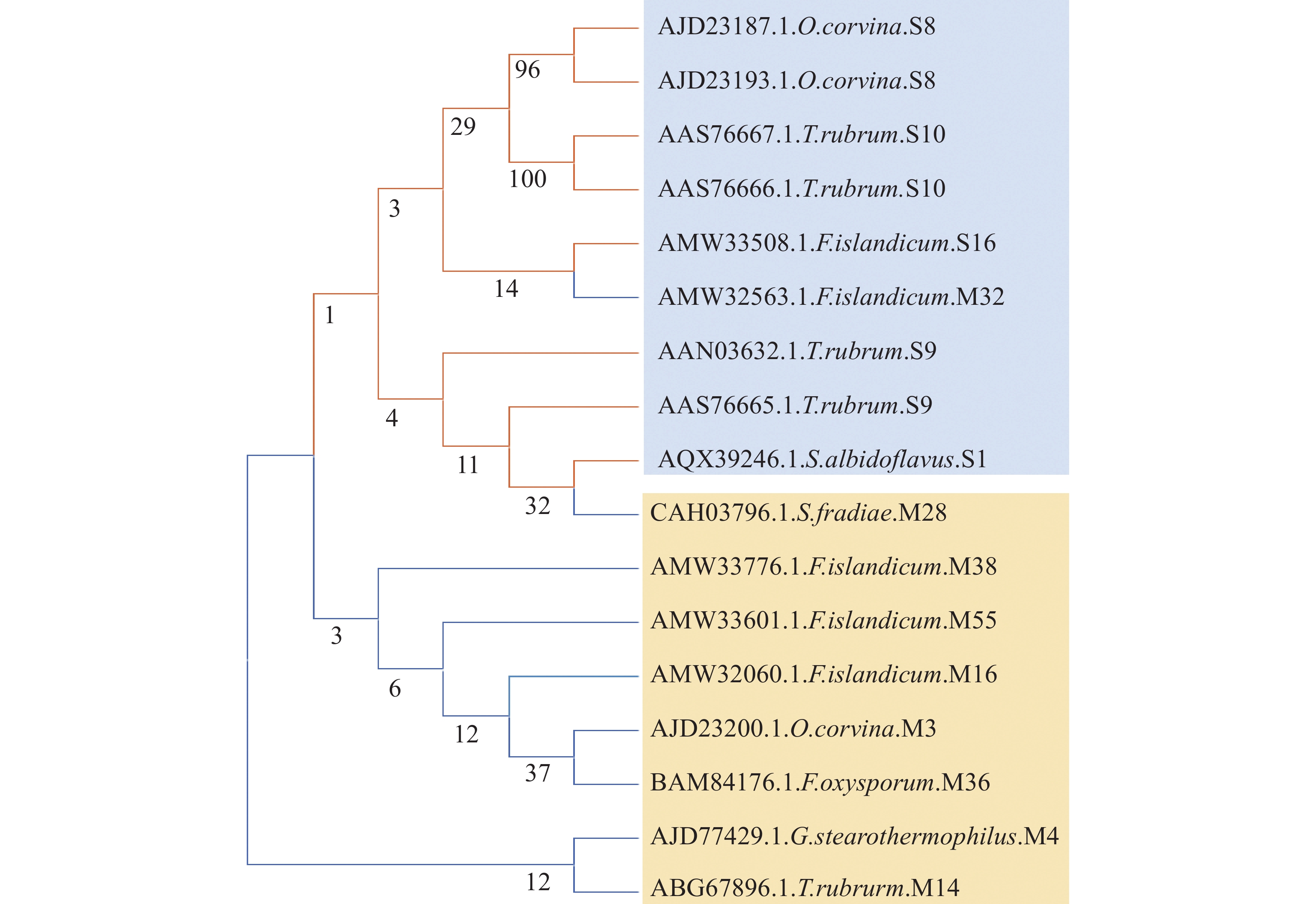
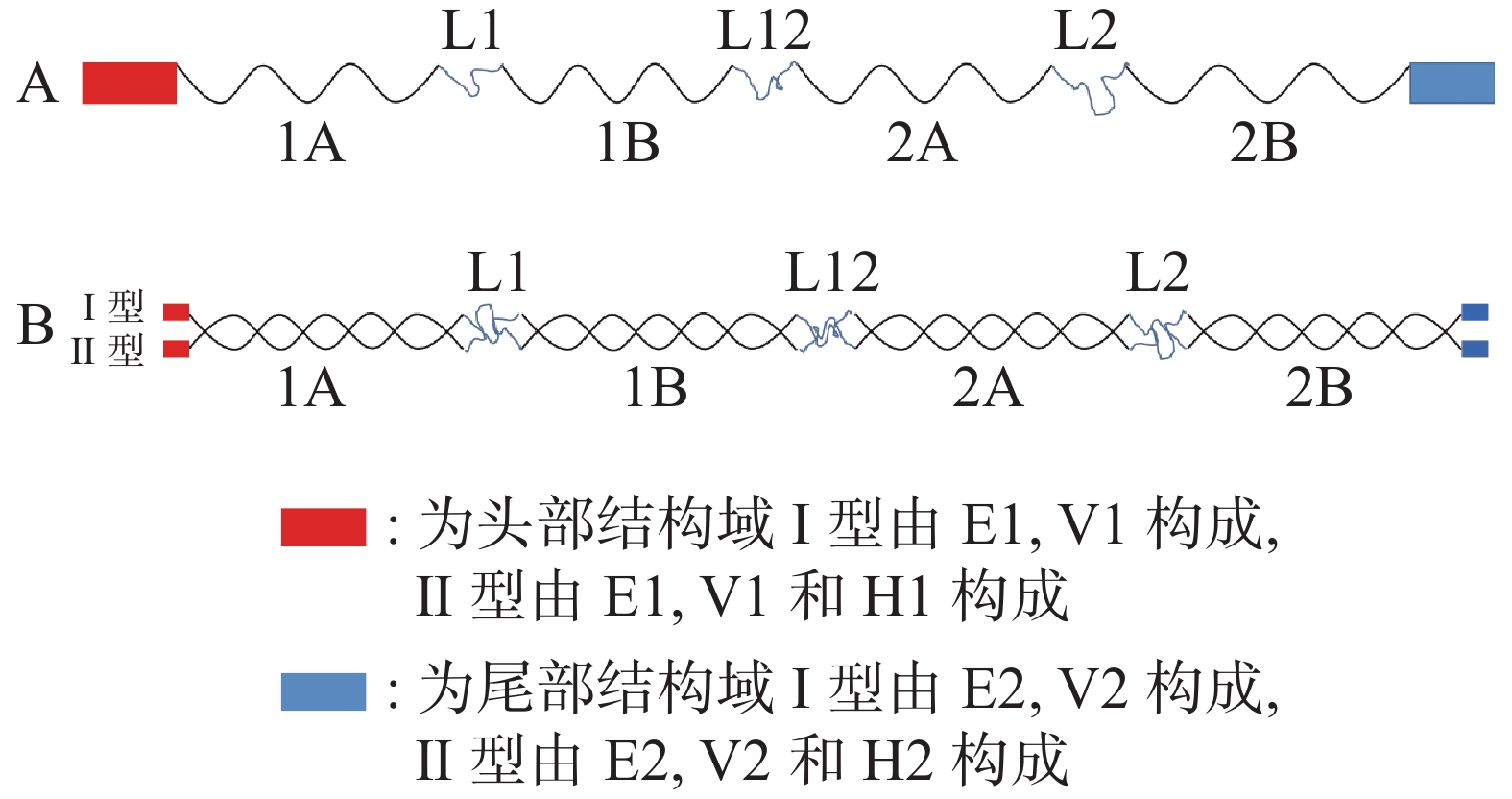
α−角蛋白的分子单体由3个结构域构成,包括头部结构域、中央棒状结构域和末端结构域。头部结构域由3部分组成,即末端子域E1、中间可变域V1和同源子域H1。棒状结构域由4个右旋的α螺旋和3个β转角构成,4个α螺旋分别命名为1A、1B、2A和2B,3个β转角分别命名为L1、L12和L2(图1) [12]。α螺旋和β转角依次交替排列,共同组成由7个部分构成的棒状结构域。尾端结构域与头部结构域类似,由E2、V2和H2构成[13](图1)。角蛋白分子单体可以分为Ⅰ型与Ⅱ型,Ⅰ型角蛋白分子不包含H1和H2,所以其分子量较Ⅱ型小6~9 kDa [14]。一分子Ⅰ型角蛋白可以和一分子Ⅱ型角蛋白形成左旋螺旋,聚合为异二聚体(图1)。两分子异二聚体可以聚合形成四聚体,两分子四聚体可以结合形成原纤维,而四分子原纤维聚合会形成α−角蛋白中间丝,若干个α−角蛋白中间丝会与富含硫化物的角蛋白相关蛋白通过二硫键互相结合并交叉连接形成粗纤维[15]。角蛋白相关蛋白对α−角蛋白中间丝的组装具有重要影响,直接决定了角蛋白组织的属性,如强度、惰性和刚性[16]。由于角蛋白分子内含有大量二硫键、氢键和离子键,极大地增加了角蛋白的结构稳定性,使其难以被降解。
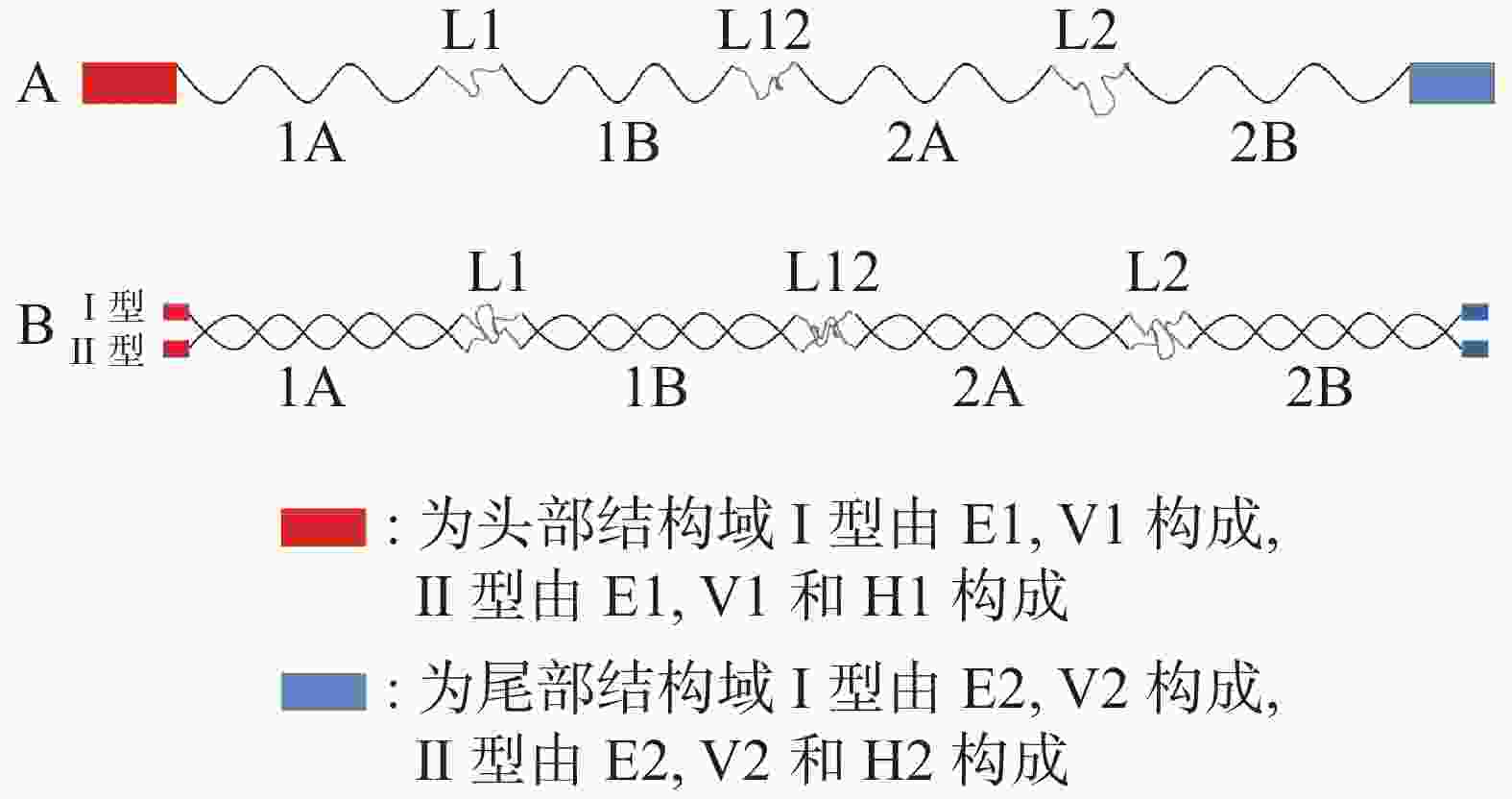
图 1 α−角蛋白单体与二聚体分子结构示意图
β−角蛋白以羽毛和爬行动物的鳞片为代表,二者具有很高的同源性。以羽毛为例,羽毛由羽根、羽轴和羽枝3部分组成,在这3部分中都包含32个蛋白残基片段,构成了β−角蛋白的纤维骨架。每个β−角蛋白分子单体由4条β链构成[17]。4条链由疏水作用力平行或反平行结合在一起,非极性氨基酸(如缬氨酸和异亮氨酸)常位于片状褶皱内表面,而带电残基和半胱氨酸残基位于侧面或者转弯处。2分子β−角蛋白单体可以通过疏水相互作用形成二聚体,在2个分子间特定氨基酸之间存在盐桥、氢键和二硫键,共同维持二聚体的稳定性。4个二聚体可以通过氢键和二硫键组装形成螺旋状重复单元,堆积的二聚体渐进旋转上升,每次旋转约为45°,构成β−角蛋白丝(图2)[18]。
图 2 β−角蛋白单体与二聚体分子结构示意图
-
物理法降解角蛋白包括高温高压水解法和膨化法。高温高压水解法是指在高温(100~200 ℃)和高压(0.294~0.980 MPa)的条件下彻底破坏角蛋白稳定的空间结构,降解产生可溶性多肽混合物。膨化法是指在高温(230 ℃)、高压(1.0~1.5 MPa)和高剪切力的条件下,快速(2.5 min)减压膨化,使角蛋白纤维降解为小的蛋白亚单位和肽链群[19]。但是,物理法降解过程中,高温可能会破坏热敏性氨基酸,降低某些氨基酸的消化率和利用率;且由于设备参数不易控制,水解效果并不理想,也难以保持产品质量的稳定性。同时,生产过程中所需耐高温高压的设备较多,生产成本和维护成本都相对较高[20]。
-
化学法降解角蛋白包括酸碱水解法、氧化还原降解法和电化学还原法。酸碱水解法是指利用强酸或强碱破坏角蛋白分子空间结构,这就需要精确控制反应的pH值、温度和反应时间,其中反应时间对产物分子量的大小和产率具有重要影响[21]。氧化还原降解法是利用氧化剂(如过氧化氢、过甲酸、过乙酸等)将角蛋白中的二硫键氧化为磺酸基,或利用还原剂(如巯基乙酸、巯基乙酸钠等)将角蛋白分子中的二硫键还原成巯基,获得可溶性角蛋白。其中,氧化法易断裂肽键,不易形成高分子量产品;而还原法对肽键的影响较少,易获得长链肽;但巯基的化学性质较为活泼,需要加入保护剂(如鼠李糖脂)避免巯基被氧化,以获得稳定的角蛋白产品[22]。电化学还原法是指将角蛋白置于电解液中,利用电流使角蛋白发生还原反应,断裂二硫键形成巯基。同时巯基会与电解液中其他离子结合避免再次被氧化。在反复地渗透、透析后可以获得可溶性角蛋白。但电化学还原法降解工艺操作较为复杂,且产物成分多样。
化学法降解角蛋白存在的主要问题是会破坏角蛋白中部分氨基酸,且工艺较为复杂,成本偏高。同时,在降解角蛋白的过程中会产生大量的工业废水,严重污染环境[23],这些问题都阻碍了化学法降解角蛋白的应用与推广。
-
微生物降解法是指在一种或多种微生物的共同作用下降解角蛋白,具有反应条件温和,产物利用率较高的特点。目前,已发现的可降解角蛋白的微生物包括真菌、放线菌和细菌。1899年Ward[24]首次发现可以降解羽毛的真菌——马爪甲团囊菌Onygena equina。后来,发现多种真菌都具有降解角蛋白的能力[25-26],例如Microsporum canis[27],Trichophyton mentagrophytes[28],Scopulariopsis brevicaulis[29]等。然而,降解角蛋白的真菌大多数为皮肤致病菌,存在环境外溢污染的风险,在工业中难以被直接利用。当前,大多数可以降解角蛋白的放线菌属于链霉菌属,已筛选出多株可以降解头发、羽毛和羊毛的链霉菌菌株,比真菌安全性高,如Streptomyces pactim [30], Streptomyces fradiae [31]等。部分细菌也可以降解角蛋白[26],已发现了部分可耐受高温或耐受碱性环境的角蛋白降解细菌,如Fervidobacterium islandicum[10],Thermoanaerobacter sp.[32]等,这类细菌在工业应用中具有显著的优势。
-
生物法降解角蛋白相较于物理法或化学法更具优势。然而,部分微生物由于是致病菌或条件致病菌,使用过程中外溢扩散导致的环境风险很高,极大地限制了其在工业生产中的大范围应用,且菌体生长过程中也会利用降解的角蛋白作为碳氮源,直接影响了最终的降解得率。因此,使用重组表达、纯化获得的角蛋白酶进行角蛋白降解的优势凸显。
目前已被鉴定的角蛋白酶都属于丝氨酸蛋白酶家族和金属蛋白酶家族,根据MEROPS肽酶数据库(
https://www.ebi.ac.uk/merops/ )对角蛋白酶的分类,可细分为14个蛋白家族,分别为丝氨酸蛋白酶S1、S8、S9、S10、S16和金属蛋白酶M3、M4、M14、M16、M28、M32、M36、M38、M55 [7]。大多数角蛋白酶不能单独降解角蛋白,需要与能够打开二硫键的酶联用共同降解角蛋白 [33]。根据角蛋白酶攻击角蛋白的位点,又可以分为内切角蛋白酶、外切角蛋白酶和寡肽酶(表1)。表 1 角蛋白酶分类及其降解机制
酶 蛋白酶家族 降解方式 核心催化单元 S1
S8
S9
S10
S16
M3
M4
M14
M16
M28
M32
M36
M38丝氨酸蛋白酶家族
丝氨酸蛋白酶家族
丝氨酸蛋白酶家族
丝氨酸蛋白酶家族
丝氨酸蛋白酶家族
金属蛋白酶家族
金属蛋白酶家族
金属蛋白酶家族
金属蛋白酶家族
金属蛋白酶家族
金属蛋白酶家族
金属蛋白酶家族
金属蛋白酶家族蛋白内切酶
蛋白内切酶
蛋白外切酶(N端)
蛋白外切酶(C端)
蛋白内切酶
寡肽酶
蛋白内切酶
蛋白外切酶(C端)
蛋白内切酶
蛋白外切酶(C端)
寡肽酶
蛋白内切酶
蛋白外切酶(N端)Ser-Asp-His催化三联体
Ser-Asp-His催化三联体
Ser-Asp-His催化三联体
Ser,-Asp-His催化三联体
Ser-Lys二元催化机制
未确定
His-Glu-His催化三联体
未确定
未确定
未确定
未确定
未确定
未确定M55 金属蛋白酶家族 蛋白外切酶(N端) 未确定 内切角蛋白酶催化角蛋白内部肽键的断裂,S1、S8、S16、M4、M16、M36蛋白家族的角蛋白酶属于内切酶(表1)[7]。S1与S8蛋白酶家族的角蛋白酶通过经典的催化三联体(Ser-Asp-His)行使功能。马怡茗等[30] 将Streptomyces albidoflavus基因组中发现的基因gm2886构建到穿梭质粒pSET152上,在Streptomyces pactum中异源表达,获得一种S1蛋白家族角蛋白酶, 在还原剂DTT的存在时可以水解天青角蛋白和羽毛。Huang等 [9] 挖掘非致病菌Onygena corvina基因组,将获取的角蛋白酶基因于毕赤酵母中表达,获得一种S8蛋白家族的角蛋白酶6877,这似乎是一种真正的角蛋白酶,因为它能够单独催化猪鬃降解,而不需要添加还原剂来破坏角蛋白中的二硫键。Kang等[10] 从可降解羽毛的极端嗜热菌Fervidobacterium islandicum中分离出S16和M16蛋白家族的角蛋白酶。与S1与S8蛋白家族的角蛋白酶不同,S16蛋白家族的角蛋白酶催化活性位点为Ser-Lys二元催化机制。将S16或M16角蛋白酶添加到F. islandicum细胞提取物中,可以提高羽毛在体外的降解效率[10]。Gegeckas等[34] 从嗜热细菌Geobacillus stearothermophilus发现M4蛋白家族的角蛋白酶GEOker。该酶属于锌金属蛋白酶,His372-Glu373-His376催化三联体对GEOker酶的催化能力起至关重要的作用,可以与DTT共同作用降解羊毛。Chaya等[35] 从Fusarium oxysporum中鉴定出2种蛋白酶KrtA和KrtC。对编码这些蛋白的基因krtA和krtC进行了分离和鉴定。在Aspergillus oryzae中成功表达并分泌rKrtA和rKrtC。同时添加rKrtA和rKrtC可以完全去除羊毛纤维的角质层,经鉴定KrtA属于S8蛋白家族,KrtC为M36蛋白家族的角蛋白酶。
外切角蛋白酶攻击角蛋白多肽链的末端,S9、M38、M55蛋白家族的角蛋白酶由N端开始降解肽链,S10、M14、M28蛋白家族的角蛋白酶则由C端开始降解角蛋白肽链[7]。Monod等 [36] 从Trichophyton rubrum中分离出来2种来自于S9蛋白家族的经典角蛋白酶,也是通过Ser-Asp-His催化三联体行使功能。Kang等[10] 从F. islandicum中基因组中挖掘到M38和M55蛋白家族的角蛋白酶基因,与仅使用F. islandicum的细胞提取物相比,加入纯化后的M38和M55蛋白家族角蛋白酶可以提高游离氨基酸的释放率,M38蛋白家族角蛋白酶可能负责调节角蛋白水解,而M55蛋白家族角蛋白酶可能在饥饿条件下被激活,控制细胞内被降解的小肽片段的流动。Zaugg等[37] 从人类致病性皮肤癣菌T. rubrum中分离出可以降解致密角质的角蛋白酶TruScpA和TruScpB,由于这2个酶与细胞膜上的糖基磷脂酰肌醇锚定,不能分泌到胞外,它们都属于S10蛋白家族,其核心催化组件依然是Ser-Asp-His催化三联体。在感染过程中糖基磷脂酰肌醇锚定的TruScpA和TruScpB与内切角蛋白酶和寡肽酶联用共同降解致密的角化组织,从而增强T. rubrum的侵染能力。T. rubrum在以角蛋白和大豆为唯一碳源和氮源的培养基中可以分泌表达角蛋白酶McpA,它属于M14蛋白家族,氨基酸残基Arg237、Arg255、Tyr311、Tyr362和Glu385对M14家族角蛋白酶的底物结合和催化非常重要,而His179、Glu182和His309则作为锌离子的配体[37- 38]。Wu等[39] 从角蛋白降解菌S. fradiae变种K11中发现了属于M28蛋白家族的角蛋白酶,并在大肠杆菌表达系统中成功异源表达。EDTA可抑制该酶的酶活,而Ca2+与Co2+则对该酶有激活作用。
目前发现的寡肽酶仅存在于2个金属蛋白酶家族M3与M32,皆为锌依赖性金属蛋白酶[7]。Huang等[9] 通过对Onygena corvina基因组的分析,推测有18种酶可能有降解角蛋白的功能,并最终在毕赤酵母中成功异源表达了分属3个蛋白家族(S8,M28和M3)的5个角蛋白酶,其中M3蛋白家族角蛋白酶为寡肽酶,由这5个蛋白酶中的3种(每个蛋白家族各1种)组成的混合酶在体外表现出高度的角蛋白降解活性。Lee等[40] 通过比较分析具有角蛋白降解活性的F. islandicum AW-1和无角蛋白降解活性的Fervidobacterium nodosum的基因组信息,发现FIAW1_1600基因在角蛋白降解过程中发挥着重要作用,并在大肠杆菌中成功异源表达出分子量为107 kDa的蛋白,为同型二聚体,且在Co2+存在时,80℃、pH 7.0条件下酶活性最高。该酶主要裂解具有碱性氨基酸序列的多肽链C端,加入细胞提取物后,显著促进了鸡毛的降解。
-
为探究角蛋白酶在自然界中的发育进化关系,笔者使用软件MEGA11中最大简约法(Maximum parsimony, MP)[41] 构建部分角蛋白酶系统发育进化树。笔者从NCBI下载了17个角蛋白酶氨基酸序列。聚类分析发现,(1)由同种微生物产生的同家族角蛋白酶的亲缘关系十分接近,如由O. corvina分泌的2种S8家族角蛋白酶在进化树中的同一簇上(图3)。(2)同种微生物产生的不同蛋白家族的角蛋白酶亲缘关系则不一定接近,如F. islandicum可以分泌分属5种不同家族(S16,M32,M38,M55和M16蛋白家族)角蛋白酶,其中S16与M32蛋白家族的角蛋白酶亲缘关系较近,而M38,M55和M16蛋白家族的角蛋白酶亲缘关系则较为接近。T. rubrum可以分泌分属于3个不同蛋白家族(S10,S9和M14)的5个角蛋白酶——S9和S10蛋白家族各2个,M14蛋白家族1个,其中S9和S10蛋白家族内2种角蛋白酶亲缘关系较近,M14蛋白家族与S9和S10蛋白家族的角蛋白酶亲缘关系较远(图3)。总体来看,丝氨酸蛋白酶家族或金属蛋白酶家族内部的角蛋白酶亲缘关系相对较近,不同蛋白家族的角蛋白酶亲缘关系较远。不过也存在例外,如F. islandicum分泌的属于S16丝氨酸蛋白家族角蛋白酶与M32金属蛋白酶家族角蛋白酶的亲缘关系较近,与金属蛋白家族M38,M55和M16角蛋白酶亲缘关系较远。
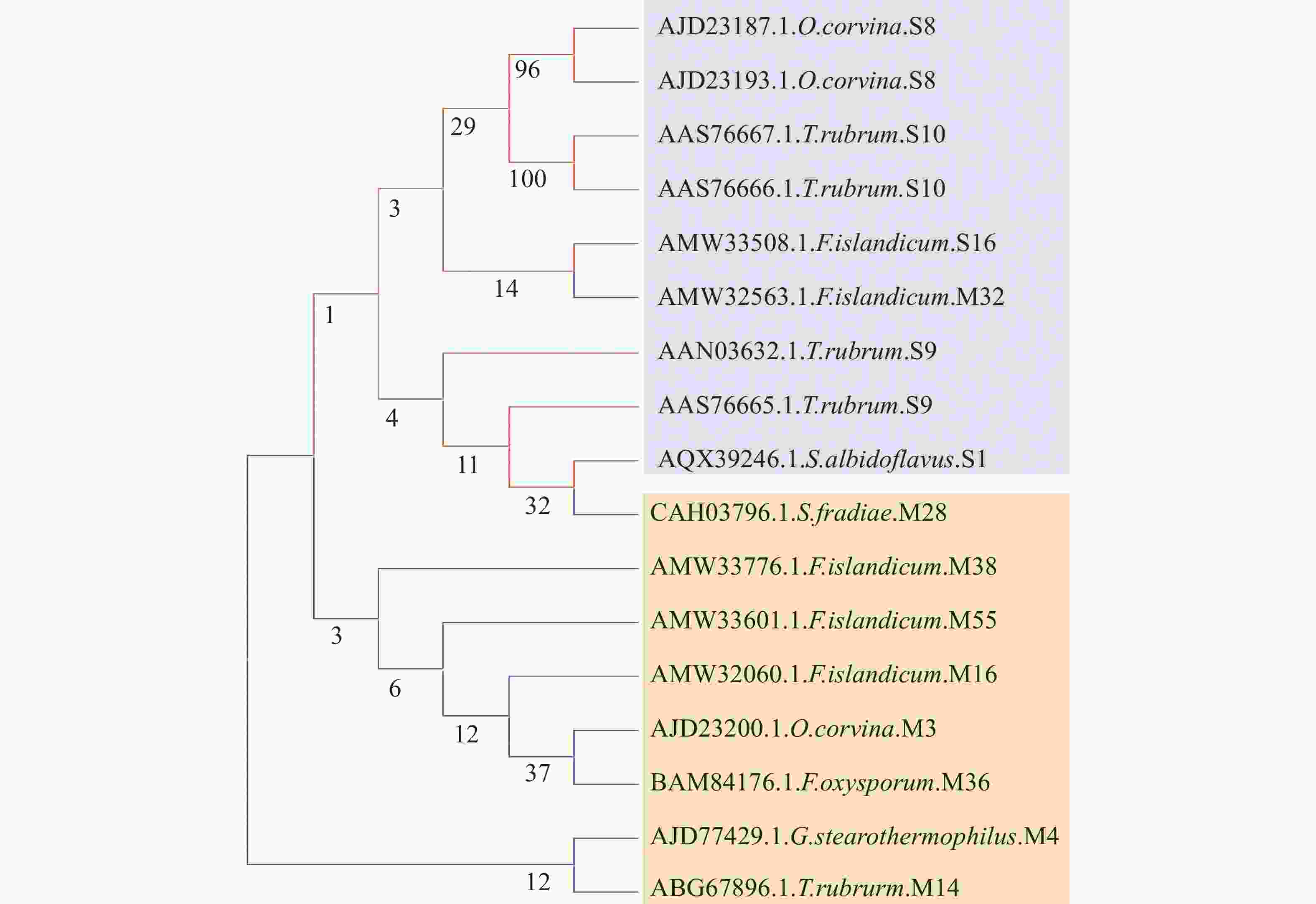
图 3 角蛋白酶氨基酸序列聚类系统发育树
-
降解角蛋白的第一步反应是还原角蛋白中的二硫键,能够催化二硫键断裂的酶包括二氢硫辛酰胺脱氢酶(EC 1.8.1.4)、谷胱甘肽还原酶(EC 1.8.1.7)、硫氧还蛋白还原酶(EC 1.8.1.9)、磷酸腺苷硫酸盐还原酶(EC 1.8.4.8)、肽甲硫氨酸亚砜还原酶(EC 1.8.4.11)、烷基过氧化氢还原酶(EC 1.11.1.15)、半胱氨酸双加氧酶(EC 1.13.11.20)、核糖核苷二磷酸还原酶(EC 1.17.4.1),这些酶都可能与角蛋白的降解相关[42]。其中,二硫键还原酶、半胱氨酸双加氧酶和谷胱甘肽还原酶在角质分解中发挥重要作用,如皮肤癣菌在角质组织生长时,半胱氨酸双加氧酶Cdo1会催化半胱氨酸形成亚硫酸盐,随后由亚硫酸盐外排泵Ssu1排出,而亚硫酸盐作为一种还原剂,可以直接破坏角蛋白中的二硫键,从而促进角蛋白降解[43]。
尽管角蛋白酶降解角蛋白的详细分子机制尚未完全清楚,但已有部分假说尝试解释角蛋白的酶解过程及分子机制。角蛋白含有大量的二硫键和氢键,在角蛋白酶开始降解前,需要对二硫键进行初步的降解以松动角蛋白的结构,使角蛋白酶便于攻击肽链[42]。因此,对于角蛋白的降解主要分为2个过程,即二硫键的还原和角蛋白水解酶降解蛋白多肽链。如果二硫键不被破坏,大多数角蛋白酶无法有效降解天然角蛋白,如在没有还原剂存在的情况下,Bacillus sp.中分离出的角蛋白酶WF146无法降解羽毛[44];角蛋白酶Cibenza DP100无法在没有还原剂的条件下降解毛发,产生可溶性多肽[45]。也有部分微生物无法分泌二硫键还原酶或亚硫酸盐,但是可以依赖细胞代谢还原二硫键,这可能是膜电位发挥了关键作用[46]。角蛋白致密的结构松动后,角蛋白酶即可发挥降解作用。这些微生物利用了代谢中产生的化学物质或膜电位与角蛋白酶共同作用降解角蛋白。例如,在皮肤癣菌侵染角质组织时,一旦角蛋白中二硫键被破坏,外切角蛋白酶家族(S9、S10、M14和M28)从两端切割肽链,内切角蛋白酶家族(S1、8、M4和M36)成员将长链角蛋白多肽切割为短肽,而寡肽酶家族(M3和M32)则作用于寡肽以释放单个氨基酸或短肽[47]。羊毛纤维表面含有大量脂质,脂质层包含18−甲基二十碳烯酸和其他脂肪酸,它们主要通过硫酯键与富含半胱氨酸的底层蛋白质结合[48],推测脂肪酸降解相关的酶有助于破坏角蛋白的外部脂质层,从而使蛋白水解酶更容易接近内部角蛋白结构。此外,角质酶(EC 3.1.1.74)也可能有助于角蛋白的消化。与角蛋白酶相似,角质酶可以与疏水性极强的底物结合,不少天然角蛋白材料中含有角质成分,例如羊毛等。推测在降解天然角蛋白材料的过程中,角蛋白酶与角质酶之间可能存在协同作用。
综上所述,单靠一种角蛋白酶一般难以将角蛋白高效降解为游离氨基酸,通常需要多种酶互相配合才能完成。例如,Aspergillus spp.和Lactobacillus spp.,通过Laps(M28)和Dpp Ⅳ(S9)协同消化由角蛋白内切酶产生的肽链[49]。对O. corvina的角蛋白酶的研究表明,由S8蛋白家族、M28蛋白家族和M3蛋白家族构成的角蛋白酶混合物协同分解猪鬃角蛋白[9]。近期报道的F. islandicum AW-1降解羽毛的酶学性质也指出,不同类型角蛋白酶之间的协同作用有助于天然角蛋白的降解,羽毛亚硫酸化后,膜内金属蛋白酶(M48)和至少3种其他的膜蛋白酶(分别属于S41、S54和S8蛋白家族)参与催化羽毛降解。随后,释放的氨基酸和多肽被转运到F. islandicum细胞内进行进一步降解,所涉及的角蛋白酶包括属于M16,M38,M55和S16蛋白家族的蛋白酶[10]。
-
角蛋白的降解液中含有多种氨基酸和小肽,包括必需氨基酸和非必需氨基酸,具有一定的医药、工业生产原料和饲料添加的应用价值。角蛋白的降解液有较高的抗氧化活性,具有维持还原环境的应用前景[50]。不同角蛋白降解后获得的多肽成分也有明显差异。
-
Fontoura等[51] 发现Chryseobacterium sp. kr6降解羽毛的发酵产物有抗氧化和抗高血压的活性,可以有效抑制血管紧张素转换酶和二胎基肽酶-4的活性,这为防治高血压和糖尿病提供了新的可能性。角蛋白具有良好的细胞亲和性和生物相容性,这使得角蛋白具有成为医疗材料的潜力,例如羽毛的降解产物可以与其他高分子材料聚合应用于人工支架或人工关节材料[52]。
-
角蛋白的降解液也可以直接作为其他发酵过程的培养基添加剂。Hassan等[53] 利用Bacillus thuringiensis MT1分泌的角蛋白酶降解驴毛,在酿酒酵母Saccharomyces cerevisiae ATCC 64712发酵生产维生素B的过程中额外添加驴毛降解产物,添加量为1.5%时,显著提高了维生素B1、维生素B2和维生素B12的产量。
-
我国的角蛋白资源十分丰富,禽肉产量在2019年已经达到2 239万t,按照鸡毛质量占总质量的7 %计算,羽毛的年产量约为156.73万t [54]。羽毛中含有蛋白质、微量元素、维生素以及未知生长因子,是一种优质的蛋白质饲料,合理地利用角蛋白资源可以缓解我国蛋白质饲料的短缺。
-
角蛋白在自然界中丰度高,但结构稳定难以被降解。如无法妥善处理,则会造成资源浪费,污染环境甚至传播疾病。目前,通过物理或化学方法处理角蛋白的成本较高,对环境不友好并且无法获得高价值的降解产物。利用生物法,尤其是使用角蛋白酶直接分解角蛋白可以克服物理、化学法的劣势,在不破坏环境的前提下获得可以在医药,工业生产及养殖业中有效利用的高价值的降解产物。
角蛋白作为一种难溶于水的蛋白质,探明角蛋白酶如何在疏水界面识别、结合角蛋白具有重要的生物学意义和应用潜力。这对于明确角蛋白酶如何在两性界面正常发挥作用也具有重要的科学研究价值。同时这些研究成果势必在极大程度上促进角蛋白在医药、健康、工业和养殖业等领域的应用。当前,对于角蛋白酶的研究还不够完善,大多数筛选出的角蛋白酶无法单独降解角蛋白,需要在还原环境中破坏角蛋白二硫键后才能发挥作用,或者同时使用多种不同的酶才能完全降解羽毛。目前发现有单独降解能力的角蛋白酶数量很少,如Huang等[9] 发现的蛋白酶6877 (S8),这些角蛋白酶资源无法满足大规模的工业应用需求。筛选和纯化耐高温、耐酸碱和更加高效的角蛋白酶、探明角蛋白酶降解角蛋白的分子机制以及充分利用角蛋白酶对角蛋白进行有目的的剪切,获得高纯度、高附加值的目标降解产物仍是今后研究的重点。
Research progress of keratin degradation
-
摘要: 介绍了角蛋白的结构特征,物理、化学和生物3种降解方法及降解产物的利用,并对3种降解方法进行了比较,相较于物理和化学降解方法,生物降解法尤其是酶解法具有对环境友好、成本低、可以获得在医药、工业生产及养殖业中有效利用的高价值的降解产物等优点;重点从酶的分类、进化关系和降解方式等角度介绍了角蛋白酶的研究和应用现状,目前对于角蛋白酶的研究还不够完善,大多数筛选出的角蛋白酶无法单独降解角蛋白,需要在还原环境中破坏角蛋白二硫键后才能发挥作用,或者同时使用多种不同的酶才能完全降解,无法满足大规模的工业应用需求。筛选和纯化耐高温、耐酸碱和更加高效的角蛋白酶、探明角蛋白酶降解角蛋白的分子机制以及充分利用角蛋白酶对角蛋白进行有目的的剪切,获得高纯度、高附加值的目标降解产物仍是今后研究的重点。Abstract: The structural features of keratins, the three degradation methods for keratins including physical, chemical and biological degradation, and the utilization of degradation products were reviewed, and the three degradation methods were compared. Compared with the physical and chemical degradation methods, the biological degradation approach, especially the enzymatic hydrolysis method, has the advantages of environmental friendliness, low-cost, and high-value degradation products which can be effectively utilized in the sectors of medicine, industry and aquaculture. The recent research and application of keratinase from the perspectives of classification, developmental evolution relationship and degradation methods and modes were overviewed. At present, the existing research on keratinases needs to improve. For example, most of the selected keratinases are unable to degrade keratin alone without disruption of the keratin disulfide bond in a reducing environment. Or multiple different keratinases are used simultaneously to complete the degradation process. These shortcomings and deficiencies prevent the large-scale industrial application of keratinase. It is still the focus of future research to screen and purify more efficient keratinases tolerant of high temperature, acid and alkali to explore the molecular mechanism of keratin degradation by keratinases, and to make full use of keratinases for targeted cleavage of keratin to obtain high-purity and high-value-added degradation products.
-
Key words:
- keratin /
- degradation /
- keratinase /
- degradation product
-
表 1 角蛋白酶分类及其降解机制
酶 蛋白酶家族 降解方式 核心催化单元 S1
S8
S9
S10
S16
M3
M4
M14
M16
M28
M32
M36
M38丝氨酸蛋白酶家族
丝氨酸蛋白酶家族
丝氨酸蛋白酶家族
丝氨酸蛋白酶家族
丝氨酸蛋白酶家族
金属蛋白酶家族
金属蛋白酶家族
金属蛋白酶家族
金属蛋白酶家族
金属蛋白酶家族
金属蛋白酶家族
金属蛋白酶家族
金属蛋白酶家族蛋白内切酶
蛋白内切酶
蛋白外切酶(N端)
蛋白外切酶(C端)
蛋白内切酶
寡肽酶
蛋白内切酶
蛋白外切酶(C端)
蛋白内切酶
蛋白外切酶(C端)
寡肽酶
蛋白内切酶
蛋白外切酶(N端)Ser-Asp-His催化三联体
Ser-Asp-His催化三联体
Ser-Asp-His催化三联体
Ser,-Asp-His催化三联体
Ser-Lys二元催化机制
未确定
His-Glu-His催化三联体
未确定
未确定
未确定
未确定
未确定
未确定M55 金属蛋白酶家族 蛋白外切酶(N端) 未确定  下载: 导出CSV
下载: 导出CSV
-
[1] NNOLIM N E, UDENIGWE C C, OKOH A I, et al. Microbial keratinase: Next generation green catalyst and prospective applications [J]. Frontiers in Microbiology, 2020, 11: 580164. doi: 10.3389/fmicb.2020.580164 [2] WANG B, YANG W, MCKITTRICK J, et al. Keratin: Structure, mechanical properties, occurrence in biological organisms, and efforts at bioinspiration [J]. Progress in Materials Science, 2016, 76: 229 − 318. doi: 10.1016/j.pmatsci.2015.06.001 [3] 彭俏丽, 张辉华. 羽毛角蛋白降解的研究进展[J]. 广东饲料, 2020, 29(10): 43 − 46. [4] BEN HAMAD BOUHAMED S, KECHAOU N. Kinetic study of sulphuric acid hydrolysis of protein feathers [J]. Bioprocess and Biosystems Engineering, 2017, 40(5): 715 − 721. doi: 10.1007/s00449-017-1737-7 [5] HASSAN M A, ABOL-FOTOUH D, OMER A M, et al. Comprehensive insights into microbial keratinases and their implication in various biotechnological and industrial sectors: A review [J]. International Journal of Biological Macromolecules, 2020, 154: 567 − 583. doi: 10.1016/j.ijbiomac.2020.03.116 [6] 张红岩, 何瑾, 杨梦莹, 等. 海洋来源羽毛降解菌株的筛选、鉴定及酶学性质研究[J]. 应用海洋学学报, 2022, 41(2): 338 − 346. [7] QIU J, WILKENS C, BARRETT K, et al. Microbial enzymes catalyzing keratin degradation: Classification, structure, function [J]. Biotechnology Advances, 2020, 44: 107607. doi: 10.1016/j.biotechadv.2020.107607 [8] 柯野, 朱艳媚, 余国兴, 等. 羽毛降解菌株Streptomyces sp. DJ产生的蛋白酶酶学性质[J]. 中山大学学报, 2017, 56(6): 147 − 152. [9] HUANG Y, BUSK P K, HERBST F A, et al. Genome and secretome analyses provide insights into keratin decomposition by novel proteases from the non-pathogenic fungus Onygena corvina [J]. Applied Microbiology and Biotechnology, 2015, 99(22): 9635 − 9649. doi: 10.1007/s00253-015-6805-9 [10] KANG E, JIN H S, LA J W, et al. Identification of keratinases from Fervidobacterium islandicum AW-1 using dynamic gene expression profiling [J]. Microbial Biotechnology, 2020, 13(2): 442 − 457. doi: 10.1111/1751-7915.13493 [11] VIDMAR B, VODOVNIK M. Microbial keratinases: Enzymes with promising biotechnological applications [J]. Food Technology and Biotechnology, 2018, 56(3): 312 − 328. [12] SMITH T A, STEINERT P M, PARRY D A. Modeling effects of mutations in coiled-coil structures: Case study using epidermolysis bullosa simplex mutations in segment 1a of K5/K14 intermediate filaments [J]. Proteins, 2004, 55(4): 1043 − 1052. doi: 10.1002/prot.20089 [13] BRAGULLA H H, HOMBERGER D G. Structure and functions of keratin proteins in simple, stratified, keratinized and cornified epithelia [J]. Journal of Anatomy, 2009, 214(4): 516 − 559. doi: 10.1111/j.1469-7580.2009.01066.x [14] STEINERT P M, STEVEN A C, ROOP D R. The molecular biology of intermediate filaments [J]. Cell, 1985, 42(2): 411 − 420. doi: 10.1016/0092-8674(85)90098-4 [15] MARSHALL R C, ORWIN D F, GILLESPIE J M. Structure and biochemistry of mammalian hard keratin [J]. Electron Microscopy Reviews, 1991, 4(1): 47 − 83. doi: 10.1016/0892-0354(91)90016-6 [16] GONG H, ZHOU H, MCKENZIE G W, et al. An updated nomenclature for keratin-associated proteins (KAPs) [J]. International Journal of Biological Sciences, 2012, 8(2): 258 − 264. doi: 10.7150/ijbs.3278 [17] FRASER R D, PARRY D A. Molecular packing in the feather keratin filament [J]. Journal of Structural Biology, 2008, 162(1): 1 − 13. doi: 10.1016/j.jsb.2008.01.011 [18] CALVARESI M, ECKHART L, ALIBARDI L. The molecular organization of the beta-sheet region in Corneous beta-proteins (beta-keratins) of sauropsids explains its stability and polymerization into filaments [J]. Journal of Structural Biology, 2016, 194(3): 282 − 291. doi: 10.1016/j.jsb.2016.03.004 [19] 陈吉英. 羽毛粉的五种加工方法[J]. 农家顾问, 2015, 1(19): 56. [20] 王晶. 一株新的羽毛角蛋白降解菌的鉴定、研究与应用 [D]; 东华大学, 2007. [21] 贾如琰, 何玉凤, 王荣民, 等. 角蛋白的分子构成、提取及应用[J]. 化学通报, 2008(4): 265 − 271. [22] OZDEMIR G, SEZGIN O E. Keratin-rhamnolipids and keratin-sodium dodecyl sulfate interactions at the air/water interface [J]. Colloids and Surfaces B, Biointerfaces, 2006, 52(1): 1 − 7. doi: 10.1016/j.colsurfb.2006.05.008 [23] 张荣, 凌晓宁, 李昆太. 微生物降解角蛋白机制及角蛋白酶应用研究进展[J]. 生物灾害科学, 2020, 43(4): 331 − 337. [24] WARD H M. Onygena equina, willd., a horn-destroying fungus [J]. Philosophical Transactions of the Royal Society of London Series B:Containing Papers of a Biological Character, 1899, 191: 269 − 291. doi: 10.1098/rstb.1899.0008 [25] 李欣, 韩淑梅, 张芝元, 等. 嗜角蛋白真菌的界定、研究方法及其应用价值[J]. 微生物学通报, 2022, 49(1): 292 − 305. [26] 韩淑梅, 李欣, 张芝元, 等. 微生物角蛋白酶的特性及其应用研究进展[J]. 微生物学通报, 2021, 48(11): 4315 − 4326. [27] BROUTA F, DESCAMPS F, FETT T, et al. Purification and characterization of a 43.5 kDa keratinolytic metalloprotease from Microsporum canis [J]. Medical Mycology, 2001, 39(3): 269 − 275. doi: 10.1080/mmy.39.3.269.275 [28] GNAT S, ŁAGOWSKI D, NOWAKIEWICZ A, et al. Population differentiation, antifungal susceptibility, and host range of Trichophyton mentagrophytes isolates causing recalcitrant infections in humans and animals [J]. European Journal of Clinical Microbiology & Infectious Diseases:Official Publication of the European Society of Clinical Microbiology, 2020, 39(11): 2099 − 2113. [29] ANBU P, GOPINATH S C, HILDA A, et al. Optimization of extracellular keratinase production by poultry farm isolate Scopulariopsis brevicaulis [J]. Bioresour Technol, 2007, 98(6): 1298 − 1303. doi: 10.1016/j.biortech.2006.05.047 [30] 马怡茗, 柯欣, 李晓霞, 等. 角蛋白酶基因gm2886在密旋链霉菌ACT12中的表达及鉴定[J]. 生物工程学报, 2017, 33(12): 1968 − 1978. [31] NOVAL J J, NICKERSON W J. Decomposition of native keratin by Streptomyces fradiae [J]. Journal of Bacteriology, 1959, 77(3): 251 − 263. doi: 10.1128/jb.77.3.251-263.1959 [32] KUBLANOV I V, TSIRUL'NIKOV K B, KALIBERDA E N, et al. Keratinase of an anaerobic thermophilic bacterium Thermoanaerobacter sp. strain 1004-09 isolated from a hot spring in the Baikal Rift zone [J]. Mikrobiologiia, 2009, 78(1): 79 − 88. [33] LI Q. Structure, application, and biochemistry of microbial keratinases [J]. Frontiers in Microbiology, 2021, 12: 674345. doi: 10.3389/fmicb.2021.674345 [34] GEGECKAS A, GUDIUKAITĖ R, DEBSKI J, et al. Keratinous waste decomposition and peptide production by keratinase from Geobacillus stearothermophilus AD-11 [J]. Int J Biol Macromol, 2015, 75: 158 − 165. doi: 10.1016/j.ijbiomac.2015.01.031 [35] CHAYA E, SUZUKI T, KARITA S, et al. Sequence analysis and heterologous expression of the wool cuticle-degrading enzyme encoding genes in Fusarium oxysporum 26-1 [J]. Journal of Bioscience and Bioengineering, 2014, 117(6): 711 − 714. doi: 10.1016/j.jbiosc.2013.11.012 [36] MONOD M, LÉCHENNE B, JOUSSON O, et al. Aminopeptidases and dipeptidyl-peptidases secreted by the dermatophyte Trichophyton rubrum [J]. Microbiology (Reading, England), 2005, 151(1): 145 − 155. doi: 10.1099/mic.0.27484-0 [37] ZAUGG C, JOUSSON O, LÉCHENNE B, et al. Trichophyton rubrum secreted and membrane-associated carboxypeptidases [J]. International Journal of Medical Microbiology:IJMM, 2008, 298(7-8): 669 − 682. doi: 10.1016/j.ijmm.2007.11.005 [38] ZAUGG C, MONOD M, WEBER J, et al. Gene expression profiling in the human pathogenic dermatophyte Trichophyton rubrum during growth on proteins [J]. Eukaryotic Cell, 2009, 8(2): 241 − 250. doi: 10.1128/EC.00208-08 [39] WU B, SHI P, LI J, et al. A new aminopeptidase from the keratin-degrading strain Streptomyces fradiae var. k11 [J]. Applied Biochemistry and Biotechnology, 2010, 160(3): 730 − 739. doi: 10.1007/s12010-009-8537-8 [40] LEE Y J, DHANASINGH I, AHN J S, et al. Biochemical and structural characterization of a keratin-degrading M32 carboxypeptidase from Fervidobacterium islandicum AW-1 [J]. Biochemical and Biophysical Research Communications, 2015, 468(4): 927 − 933. doi: 10.1016/j.bbrc.2015.11.058 [41] TAMURA K, PETERSON D, PETERSON N, et al. MEGA5: Molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods [J]. Molecular Biology and Evolution, 2011, 28(10): 2731 − 2739. doi: 10.1093/molbev/msr121 [42] PENG Z, ZHANG J, DU G, et al. Keratin waste recycling based on microbial degradation: Mechanisms and prospects [J]. ACS Sustainable Chemistry & Engineering, 2019, 7(11): 9727 − 9736. [43] ŁABA W, CHOIŃSKA A, RODZIEWICZ A. The release of sulfur compounds during degradation of feather keratin by two Bacillus strains [J]. Wrocław University of Environmental and Life Sciences, 2013, 12(2): 29 − 40. [44] LIANG X, BIAN Y, TANG X F, et al. Enhancement of keratinolytic activity of a thermophilic subtilase by improving its autolysis resistance and thermostability under reducing conditions [J]. Applied Microbiology and Biotechnology, 2010, 87(3): 999 − 1006. doi: 10.1007/s00253-010-2534-2 [45] NAVONE L, SPEIGHT R. Understanding the dynamics of keratin weakening and hydrolysis by proteases [J]. PloS One, 2018, 13(8): 1 − 21. [46] BOCKLE B, MULLER R. Reduction of disulfide bonds by Streptomyces pactum during growth on chicken feathers [J]. Applied and Environmental Microbiology, 1997, 63(2): 790 − 792. doi: 10.1128/aem.63.2.790-792.1997 [47] MERCER D K, STEWART C S. Keratin hydrolysis by dermatophytes [J]. Medical Mycology, 2019, 57(1): 13 − 22. doi: 10.1093/mmy/myx160 [48] GHOSH A, CLERENS S, DEB-CHOUDHURY S, et al. Thermal effects of ionic liquid dissolution on the structures and properties of regenerated wool keratin [J]. Polymer Degradation and Stability, 2014, 108: 108 − 115. doi: 10.1016/j.polymdegradstab.2014.06.007 [49] BYUN T, KOFOD L, BLINKOVSKY A. Synergistic action of an X-prolyl dipeptidyl aminopeptidase and a non-specific aminopeptidase in protein hydrolysis [J]. Journal of Agricultural and Food Chemistry, 2001, 49(4): 2061 − 2063. doi: 10.1021/jf001091m [50] SHARMA I, KANGO N. Production and characterization of keratinase by Ochrobactrum intermedium for feather keratin utilization [J]. Int J Biol Macromol, 2021, 166: 1046 − 1056. doi: 10.1016/j.ijbiomac.2020.10.260 [51] FONTOURA R, DAROIT D J, CORREA A P, et al. Production of feather hydrolysates with antioxidant, angiotensin-I converting enzyme- and dipeptidyl peptidase-IV-inhibitory activities [J]. New Biotechnology, 2014, 31(5): 506 − 513. doi: 10.1016/j.nbt.2014.07.002 [52] MELKE J, MIDHA S, GHOSH S, et al. Silk fibroin as biomaterial for bone tissue engineering [J]. Acta Biomaterialia, 2016, 31: 1 − 16. doi: 10.1016/j.actbio.2015.09.005 [53] HASSAN M A, TAHA T H, HAMAD G M, et al. Biochemical characterisation and application of keratinase from Bacillus thuringiensis MT1 to enable valorisation of hair wastes through biosynthesis of vitamin B-complex [J]. Int J Biol Macromol, 2020, 153: 561 − 572. doi: 10.1016/j.ijbiomac.2020.03.032 [54] 腰文颖. 我国家禽生产情况及趋势分析[J]. 兽医导刊, 2021(3): 5 − 7. -

 点击查看大图
点击查看大图
图(3) / 表 (1)
计量
- 文章访问数: 698
- HTML全文浏览量: 221
- PDF下载量: 39
- 被引次数: 0
