

 下载:
下载:
-
油棕(Elaeis guineensis Jacq.)又名油椰子,为原产于热带非洲的棕榈科多年生单子叶木本植物[1],在中国主要分布于海南省南部、云南省西双版纳等地区,寿命长、经济价值高,年均单位面积产油量(CPO)高达 3.7 t·hm−2[2],是世界油料的重要来源之一。油棕果实的中果皮是重要的油脂积累组织,主要产物为棕榈油,棕榈油的用途广泛,在食品加工、轻工业、机械和生物清洁能源等领域均有应用[3-6]。棕榈油中饱和脂肪酸与不饱和脂肪酸的比例约为 1∶1 。新鲜油棕果肉含有约 38%~45% 的棕榈酸(C16:0),38%~44% 的油酸(C18:1),10%~12% 的亚油酸(C18:2)和 3%~5% 硬脂酸(C18:0),总饱和度约为 50%。提高食用油中油酸(C18:1)等不饱和脂肪酸含量,降低饱和脂肪酸在总油脂中的比例,一直是油棕遗传改良的重要目标之一。
乙酰基酰基载体蛋白硫酯酶(fatty acyl-ACP thioesterase, FAT)是一种由核基因编码的可溶性酶,主要作用于酰基和ACP间的硫酯键,水解硫酯键、释放ACP和游离的脂肪酸,从而停止碳链的延长[7-8]。目前研究的高等植物中酰基-ACP硫酯酶根据其分解底物的特异性可分为两类[9]:一类是FATA(酰基载体蛋白硫酯酶A),在油棕中主要作用于十八碳链脂肪酸链(C18:X-ACP),但在不同的物种中具体对应关系有较大差异;另一类则是FATB(酰基载体蛋白硫酯酶B),主要作用饱和脂酰基-ACP[10],对C16:0-ACP的催化活性最高[11]。
病毒诱导的基因沉默 (virus induced gene silencing, VIGS)利用插入了植物靶基因cDNA片段的病毒载体侵染植物,进入植株的病毒在复制和转录过程中能够特异性识别与插入片段序列同源的mRNA,使其降解从而抑制靶基因的表达[12]。常规遗传转化验证基因功能时,常需要植物生长至F2代甚至F3代时才能展开研究,而VIGS体系的应用,可在较短时间内通过观察植株的表型或生理指标的变化,应用VIGS技术对基因的功能进行验证。VIGS体系的应用特别适合于多年生木本植物中基因的功能鉴定[13]。在本研究中,油棕胚状体实验材料的选择可以克服油棕遗传转化周期长的困难,加快基因功能的鉴定与研究,同时保持了油棕的物种特性。
油棕是热带重要油料作物,产油量较高且用途广泛。然而,作为食用油来说,棕榈油脂肪酸比例的改善一直是油棕的重要育种目标之一[14-15]。其中,油棕乙酰基-ACP硫酯酶决定了脂肪酸链的长度和饱和度,被认为对脂肪酸组分的改善有重要的价值。目前,油棕中油棕乙酰基-ACP硫酯酶的具体功能尚不明确,沉默EgFATA基因的表达并测定基因沉默前后脂肪酸种类的变化,能够解析EgFATA的具体功能及其在脂肪酸调控中的作用。
-
试验使用的油棕胚状体由笔者所在的实验室提供、诱导并保存。保存方式:温度 28 ℃,相对湿度 65%,光照强度 400 μmol·m−2·s−1, 28 d继代培养1次,光照 16 h,黑暗 8 h交替培养。pTRV1、pTRV2载体由山东农业大学李媛媛老师惠赠;pEASY-T1-Simple-Cloning Kit试剂盒购自北京全式金公司;大肠杆菌DH5α感受态细胞、农杆菌EHA105感受态细胞购自上海唯地生物公司。
-
参照NCBI公布的基因序列(
https://www.ncbi.nlm.nih.gov/nuccore/XM_029265988.1?report=fasta ),使用Primer Premier 5软件设计带接头的FATA引物(F:atgaattcATGCTGAAGGGGTACCACGTG;R:atgagctcTCATCGAACCAACTTCCTCCA),酶切位点为Ecor1和Spe1,以实验室保存的油棕果cDNA为克隆模板,使用诺唯赞的Phanta Max Super-Fidelity DNA Polymerase试剂盒进行扩增,按照说明书进行操作,其中,退火温度为57 ℃,延伸时间为2 min。使用Cycle-Pure Kit试剂盒纯化PCR产物,将目的片段37 ℃、30 min构建到pEASY-T1 Simple Cloning Kit中间载体上,用热激法转化至大肠杆菌感受态DH5α,挑取单菌落进行PCR验证,重组载体酶切鉴定、测序均验证无误后,保藏菌种。对EgFATA (XP_010927699.1)的蛋白序列使用ProtParam软件分析理化性质, 使用ProtScale对EgFATA蛋白序列的疏水性进行预测;对FATA使用TMHMM蛋白跨膜结构区分析;使用MEGA 7.0对NCBI数据库对检索出的EgFATA的同源蛋白聚类分析,构建系统发育进化树。
-
以测序正确的EgFATA-pEASY-Blunt质粒为模板,于NCBI在线选取EgFATA 200~500 bp的特异性片段,使用Primer Premier 5软件设计EgFATA特异性片段的引物(F:atgaattcTTGGATGGGTCCTCGAAAGC;R:atgagctcGTGCGGCCCCGATTTATT),使用Cycle-Pure Kit试剂盒纯化PCR产物后,通过Takara T4 DNA Ligase试剂盒22 ℃、1 h连接目的片段与线性化pTRV2载体,测序无误后使用冻融法转化至EHA105农杆菌感受态中,甘油法保藏菌种。
-
于含有Kana+、Lif +的液体培养基(LB)中,28 ℃、220 r·min−1培养pTRV1-EHA105、pTRV2-EHA105、pTRV2-EgFATA-EHA105农杆菌菌种,用分光光度计调节菌液 OD600=0.5。混合pTRV1-EHA105和pTRV2-EHA105菌液作为病毒空白对照,混合pTRV1-EHA105与pTRV2-EgFATA-EHA105作为实验组,离心收集农杆菌菌体后,加入MSO液体培养基悬浮,保持菌液 OD600=0.5。使用针扎、浸泡的方式侵染油棕胚状体,浸泡 5 min 后,将胚状体放置在灭菌后的滤纸上,吸取多余菌液,之后转移至共培养固体植物培养基(WPM)中(含 20 mg·L−1的AS(乙酰丁香酮)和 100 mg·L−1 的半胱氨酸),19 ℃暗培养2 d。无菌水清洗胚状体后将其转移到继代培养基中,在 28 ℃ 温度下培养。期间淘汰长势不好以及死亡的胚状体,培养12 d后获得成功存活下来的转基因胚状体,用于后续实验(qRT-PCR和FAs)分析。
-
使用诺唯赞公司的多糖多酚植物RNA提取试剂盒,提取油棕胚状体RNA,液氮研磨样品后,不可再次冻融,严格按照说明书操作。分别计算野生型、对照组(病毒组)和实验组胚状体RNA浓度,保证RNA加入量相同,反转录获得cDNA。
以油棕保守基因Eg-β-actin为内参,根据EgFATA和Eg-β-actin基因分别设计荧光定量引物(qRT-FATA-F:TATCGCCAATCTCCTCCAG;qRT-FATA-R:GTTCATCATCACCCACTTGC;qRT-actin-F:TGGAAGCTGCTGGAATCCAT;qRT-actin-R:TCCTCCACTGAGCACAACGTT)使用TaKaRa TD600荧光定量PCR仪和2xQ3 SYBR qPCR Master Mix(22204)试剂盒进行实时荧光定量,检测EgFATA基因在实验组和病毒对照组胚状体中的表达量,反应体系(20 µL):上下游引物各0.5 µL,cDNA 1 µL,2xQ3 SYBR qPCR Master Mix(Universal)10 µL,ddH2O 8 µL;PCR反应程序为预变性 95 ℃;循环反应95 ℃ 10 s、60 ℃ 30 s,40个循环;溶解曲线采集95 ℃ 15 s、60 ℃ 60 s、95 ℃ 15 s,实验中每个样品设置 3~4 个平行,严格按照说明书操作。
-
实验用的枪头、Ep管进行121 ℃、20 min高压灭菌处理,无水乙醇灼烧研钵与研磨棒,冷却后于研钵中研磨等量胚状体,油脂瓶收集样品后,使用V氯仿∶V甲醇=2∶1的提取液萃取2~3次,收集上清液经无菌水与0.9% NaCl溶液漂洗后,氮吹仪吹干获得油脂,加入2.5%浓硫酸的甲醇溶液(V/V),并加入10 µg的C17烷酸标准品,80 ℃水浴2 h进行甲酯化,氮吹仪吹干后,加入适量正己烷溶解,漩涡混匀后加入2 mL 0.9% NaCl溶液,静置、离心后取上层正己烷有机相用于高效气相色谱分析。
-
以油棕果cDNA为模板,PCR方法扩增获得EgFATA基因的CDS区 1 155 bp,1.2%琼脂糖凝胶回收后,构建EgFATA-pEASY-Blunt载体,并对单菌落PCR检测,条带大小与预期一致,测序结果与NCBI公布的参考序列相同。使用ProtParam对EgFATA进行理化性质分析,结果表明:EgFATA的编码区由 1 155 个氨基酸的序列组成,编码 385 个氨基酸,相对分子量为91.89 kDa 。通过NCBI在线网站的BLAST功能,比对EgFATA(XP_010927699.1)蛋白的相似序列,结果表明:EgFATA与海藻PdFATA(XP_008794588.1)、椰子CnFATA(AZZ09172.1)、凤梨AcFATA(XP_020106065.1)、香蕉MaFATA( XP_009411855.1)、野蕉MbFATA(THU68832.1)有极高的同源性,分别为94.28%、91.13%、92.76%、89.76%、89.42%(图1-A);为进一步确定油棕EgFATA与其他物种间的亲缘关系,于NCBI在线网站BLAST中继续筛选其他物种的FATA基因,使用 MEGA 7.0构建系统进化树,结果表明,EgFATA与海藻PdFATA蛋白相似度最高,亲缘关系最近(图1-B)。
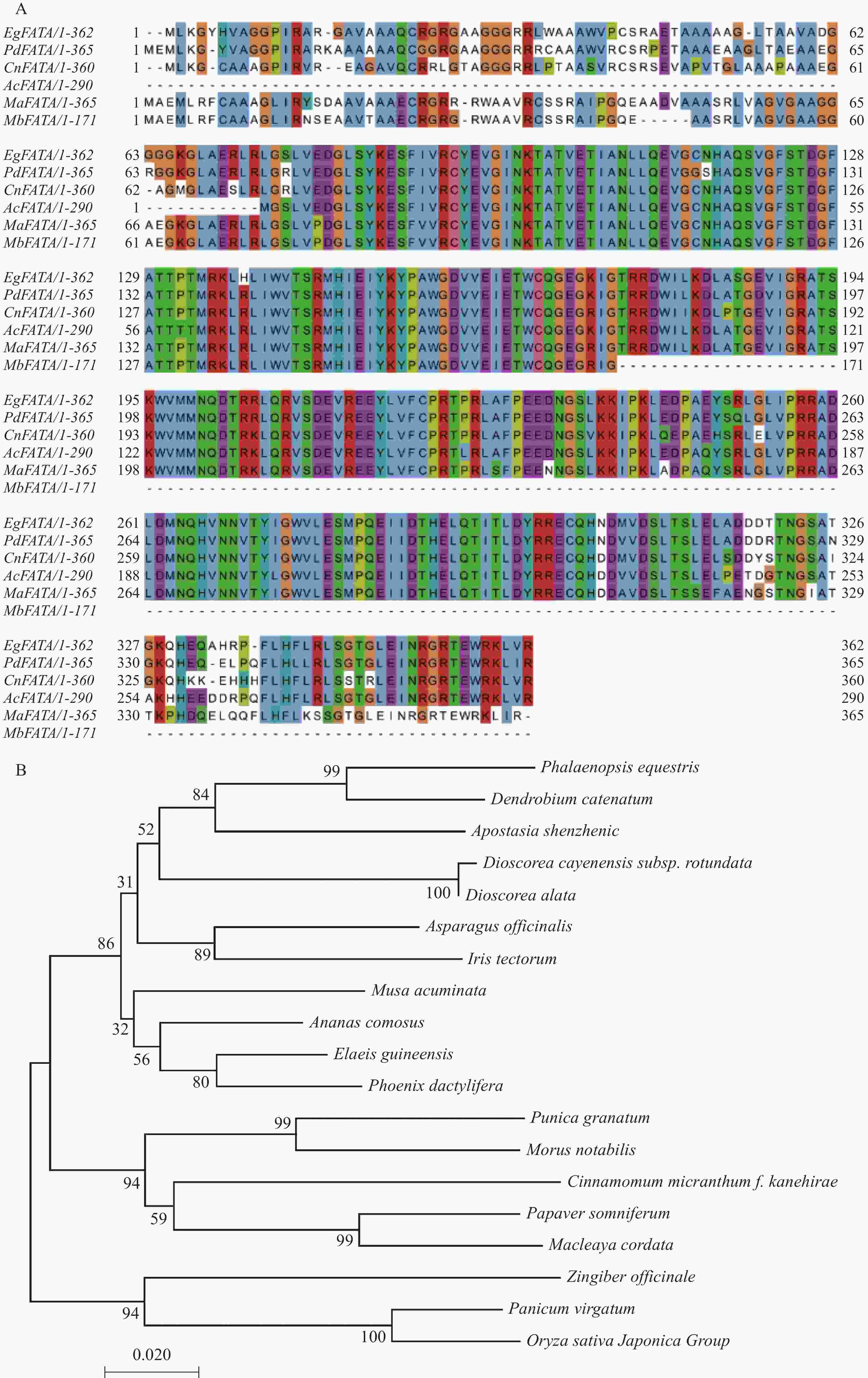
图 1 EgFATA蛋白的生物信息学分析
-
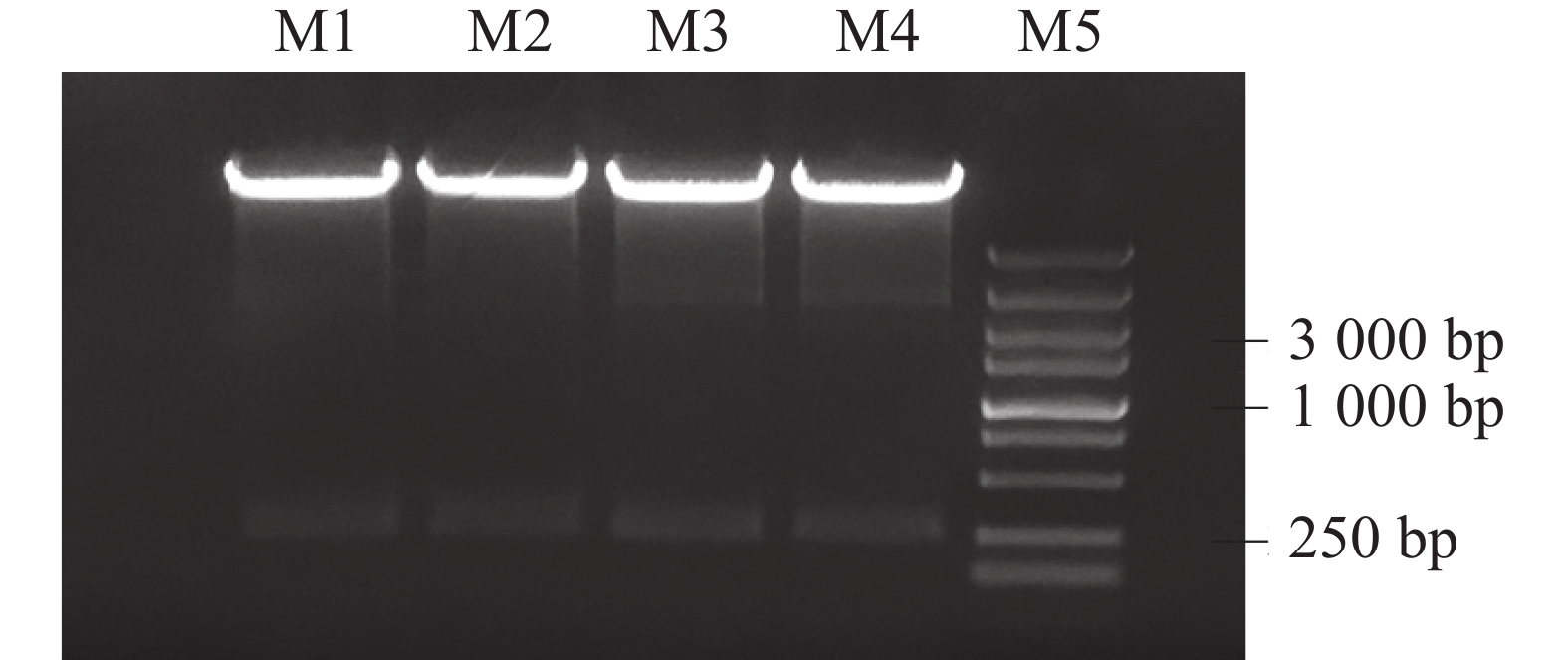
设计带接头的引物,以EgFATA-pEASY-Blunt质粒为模板,PCR扩增方法扩增获得EgFATA基因的特异性片段247 bp,1.2%琼脂糖凝胶回收后,与经Ecor1和Sac1双酶切的pTRV2线性化载体连接并进行酶切鉴定(图2),测序结果表明, EgFATA- pTRV2基因沉默载体构建成功,使用冻融法将其转入EHA105农杆菌感受态中。
图 2 EgFATA- pTRV2重组质粒的酶切鉴定
-
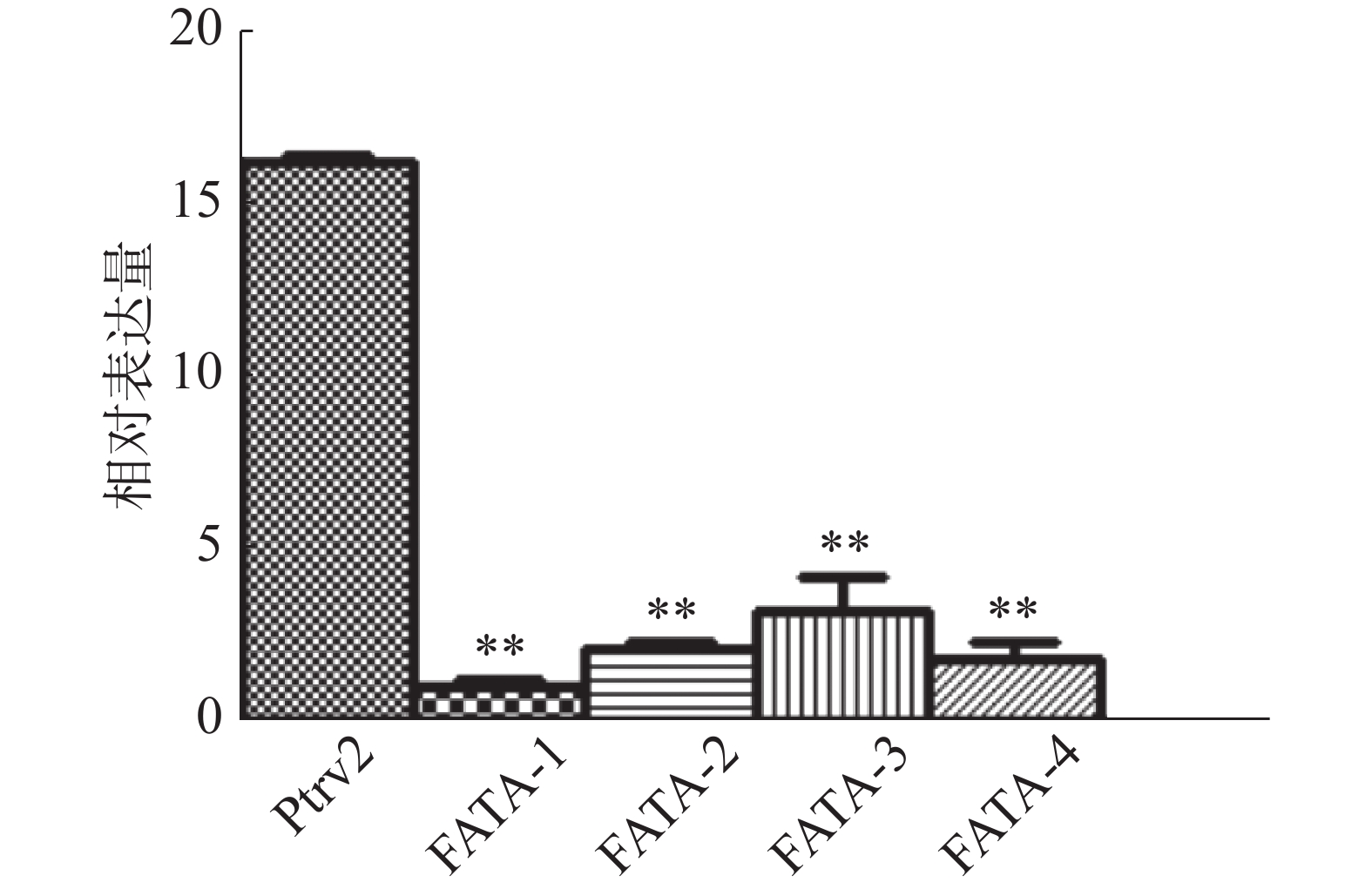
分别提取基因沉默实验组、空白病毒对照组的胚状体RNA,反转录为cDNA,以Eg-β-actin为内参基因,进行实时定量PCR检测,结果显示,与病毒对照组相比,基因沉默实验组不同株系的EgFATA表达量均出现极显著性下调,说明VIGS体系成功将不同株系中的EgFATA基因的表达量下调了85%~90%(图3)。
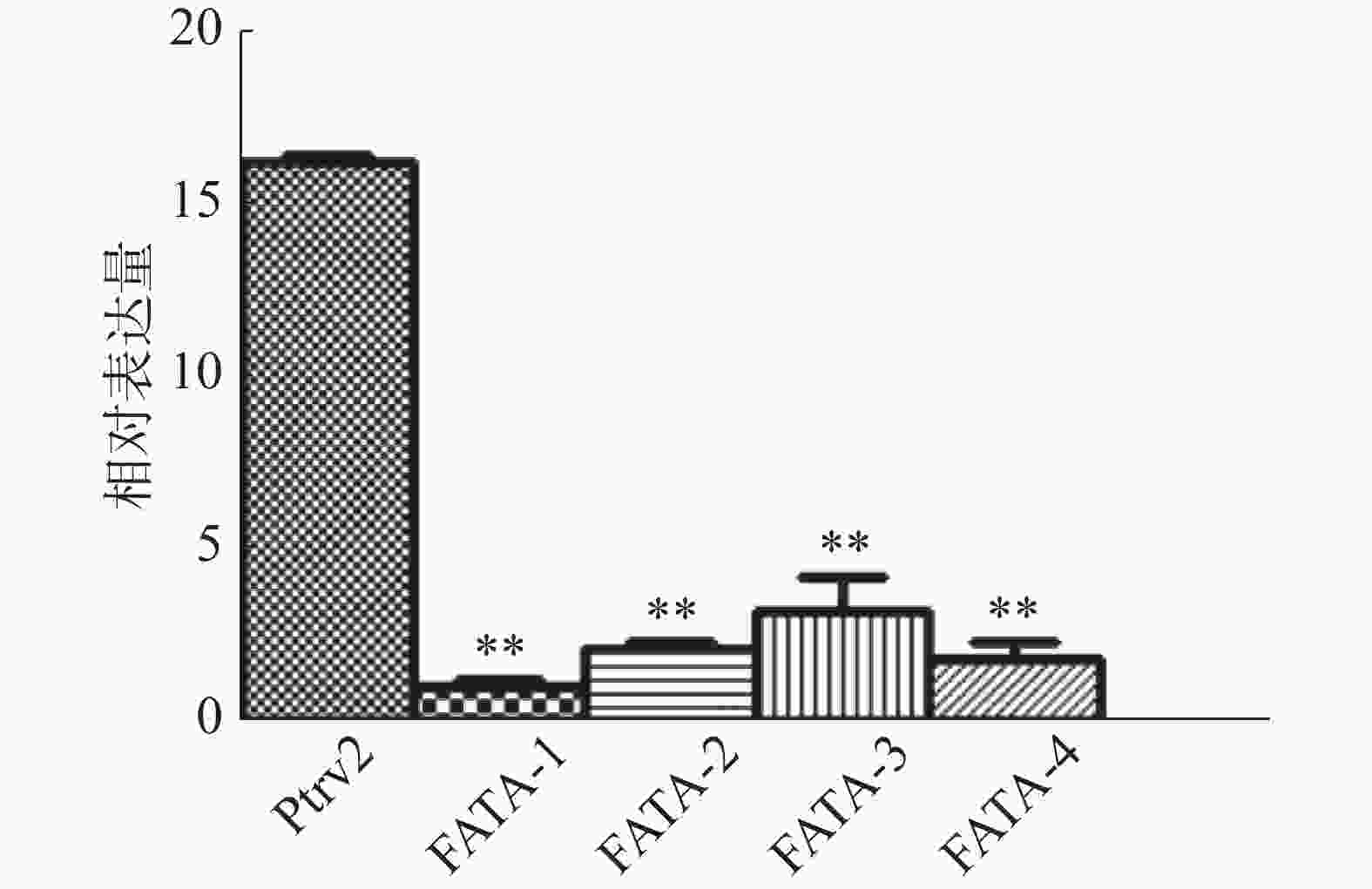
图 3 油棕胚状体中实验组及病毒对照组EgFATA基因表达量的检测
-
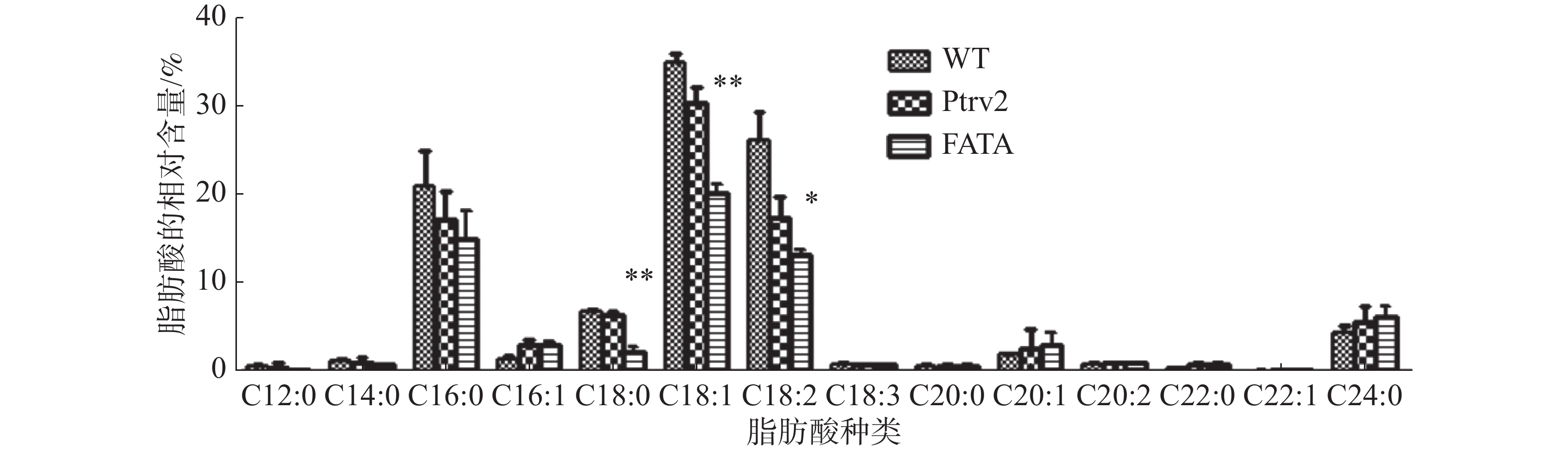
分别提取实验组、空白病毒对照组和野生型的胚状体总油脂,经甲酯化处理后、进行高效气相色谱分析。结果表明,与野生型和空载病毒对照相比,沉默EgFATA基因的油棕胚状体脂肪酸含量发生变化,C18:0和C18:1极显著性下降,与C18:0相比,C18:1下调的程度极显著;除此之外,C18:2显著性下降,其他种类的脂肪酸未呈现显著性变化(图4)。该结果说明,下调EgFATA基因的表达,抑制了油棕胚状体中十八烷基脂肪酸的合成代谢,从而使C18:0、C18:1、C18:2的含量下降,即油棕胚状体中EgFATA的催化底物为C18:0-ACP、C18:1-ACP、C18:2-ACP,其中对C18:1-ACP的偏好性更强。
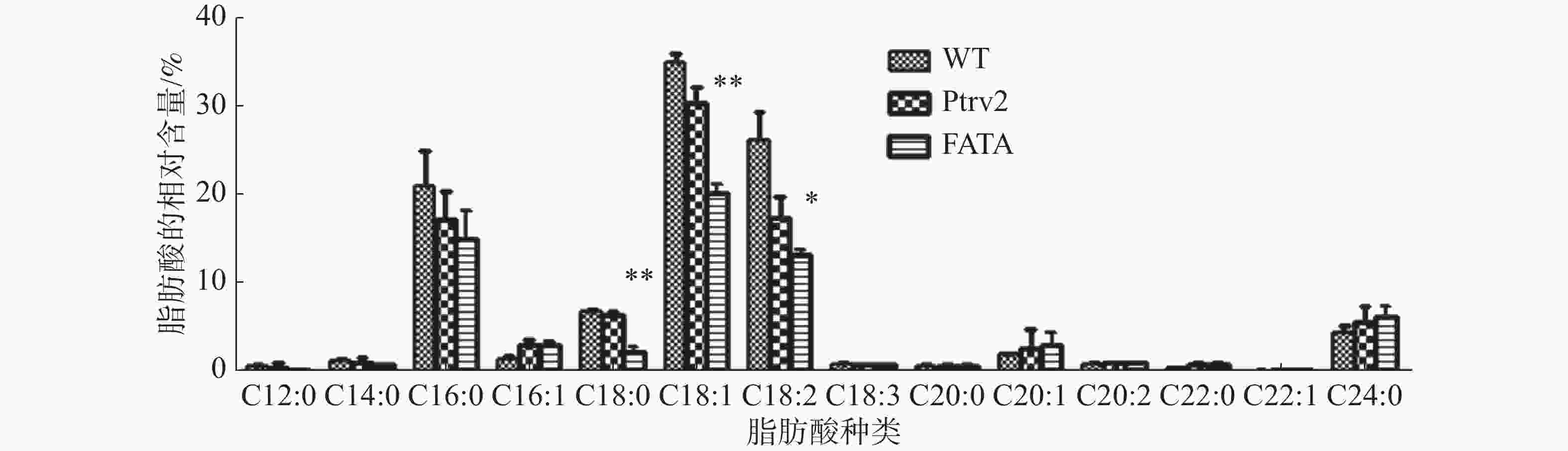
图 4 EgFATA基因沉默胚状体中脂肪酸相对含量的变化
-
研究结果表明,EgFATA氨基酸序列与海藻、椰子、凤梨、香蕉、野蕉同源性极高,在89%以上,说明FATA蛋白在不同物种中具有高度的保守性。系统进化树分析发现,EgFATA与PdFATA亲缘关系最近,二者在功能上可能存在相似性。对沉默EgFATA油棕胚状体进行RT-qPCR实验、油脂的提取与脂肪酸分析,与野生型和空载病毒对照组胚状体比较发现,沉默油棕胚状体中脂肪酸的种类未发生改变,但含量发生了显著性变化,如C18:0、C18:1和C18:2均显著性降低,说明EgFATA表达量的下调影响了十八碳链脂肪酸(C18:X)的积累。
EgFATA是影响油脂积累的重要基因,Kaczmarzyk[16]和Tan等[17]认为,多数植物FATA基因功能上具有相似性,即对C18:X-ACP具有催化活性,且对C18:1-ACP的偏好性更高,这与笔者的研究对象EgFATA功能相似,下调油棕胚状体中EgFATA基因的表达量后,C18:0和C18:1分别下降75%、90%,C18:1下调程度更大,与先前报道相符。然而,文献报道显示有些植物中的FATA基因对C18:0-ACP催化活性更强,如油菜中过表达山竹GmFATA基因会使转基因油菜中C18:0的含量显著提升约20%[18]。除此之外,其他物种中FATA基因还存在不同的对应关系,Moreno-Pérez等发现沉默拟南芥AtFATA基因后,其种子的亚麻酸(C18:3)和芥酸(C22:1)含量显著性增加[19]。
常见的油料作物如油菜、花生、大豆、芝麻等,基因的功能研究均存在实验周期长、转化效率低、操作难度大、研究成本高的问题,因此以往的研究多在拟南芥、大肠杆菌、酵母菌[20]等模式物种中开展,本研究选择油棕胚状体作为实验材料,缩短了实验周期,并且保留物种特性,功能研究更为便捷。与以往研究报道不同的是,本次发现EgFATA表达量的下调对C18:2的积累也产生了影响。在分析沉默EgFATA的转基因胚状体中脂肪酸的含量发现,C18:2约被下调 51%,脂肪酸去饱和酶(△12 Fatty acid desaturase, FAD2)是合成C18:2的重要基因,催化C18:1向C18:2的转变[21]。本实验中C18:2的下调一方面可能是由于合成底物C18:1的减少,影响了C18:2的积累;另一方面EgFATA可能对C18:2-ACP也具有催化活性,因此C18:2的积累可能受到EgFATA与EgFAD2的共同调控。
油棕EgFATA功能的研究探索了EgFATA基因的生物学功能,解析了EgFATA基因在油棕中与脂肪酸积累的对应关系,并为进一步研究 EgFATA调控油脂代谢的分子机制奠定了基础。
Cloning and functional analysis of the fatty acyl-ACP thioesterase EgFATA in oil palm
-
摘要: 为了探索油棕(Elaeis guineensis Jacq.)EgFATA基因的生物学功能,解析EgFATA基因在油棕中与脂肪酸积累的对应关系,为进一步研究EgFATA作用机制奠定基础,从油棕果cDNA中克隆得到了EgFATA基因的全长序列,经生物信息学分析发现,EgFATA编码区由1 155个碱基组成,可编码385 个氨基酸,相对分子量为91.89 kDa,与海藻PdFATA亲缘关系最近。在此基础上,构建EgFATA的病毒沉默载体并侵染油棕胚状体,RT-qPCR结果表明,EgFATA的表达量被下调85%~90%。同时,对基因沉默胚状体进行脂肪酸气相色谱法分析,硬脂酸(C18:0)、油酸(C18:1)、亚油酸(C18:2)的含量均显著性降低。以上结果表明在油棕胚状体中,EgFATA对C18:0-ACP、C18:1-ACP、C18:2-ACP的水解具有催化作用,且对C18:1-ACP的催化活性最强。
-
关键词:
- 油棕 /
- 乙酰基酰基载体蛋白硫酯酶 /
- 油脂代谢调控
Abstract: Oil palm (Elaeis guineensis Jacq.) is an important tropical palm with high oil production. Fatty acyl-ACP thioesterase (FAT) is the key enzyme for hydrolysis of acyl and ACP and termination of carbon chain extension. The oil palm FATA family mainly acts on eighteen-carbon chain fatty acids (18: X-ACP), but the specific hydrolysis substrates and preferences are unknown. In this study, the full-length sequence of the EgFATA gene was first cloned from oil palm fruit. Bioinformatics analysis indicated that the EgFATA coding region consists of 1155 bases and can encode 385 amino acids with a molecular weight of 91.89 kDa, and is most closely related to the alga PdFATA. A viral silencing vector for EgFATA was constructed and then infected the embryoids of oil palm, and RT-qPCR results showed that the expression of EgFATA was down regulated by 85%-90%. At the same time, fatty acid GC analysis of the silenced embryoids showed that the contents of stearic acid (C18:0), oleic acid (C18:1) and linoleic acid (C18:2) were significantly reduced. The aforesaid results indicated that EgFATA had catalytic effect on the hydrolysis of C18:0-ACP, C18:1-ACP and C18:2-ACP in the oil palm embryoids, with the highest catalytic activity observed in the hydrolysis of C18:1-ACP. This study initially explored the biological functions of EgFATA genes in oil palm, providing a theoretical basis for further exploration of the molecular mechanisms of EgFATA in regulation of lipid metabolism.-
Key words:
- oil palm /
- fatty acyl-ACP thioesterase /
- regulation of lipid metabolism
-
[1] 冯美利, 曾鹏, 刘立云. 海南发展油棕概况与前景[J]. 广西热带农业, 2006(4): 37 − 38. [2] NGANDOEBONGUE G F, AJAMBANG W N, KOONA P, et al. Oil palm[M]// Technological Innovations in Major World Oil Crops, Volume 1. New York: Springer, 2012, 79(2): 165 − 200. [3] SYLVIE, WECKX, DIRK, et al. Tissue Culture of Oil Palm: Finding the Balance Between Mass Propagation and Somaclonal Variation. [J]. Frontiers in plant science, 2019, 10: 722. doi: 10.3389/fpls.2019.00722 [4] 高尚士. 高产油料树──油棕[J]. 林业科技通讯, 1994(11): 41. [5] ITHNIN M, XU Y, MARJUNI M, et al. Multiple locus genome-wide association studies for important economic traits of oil palm [J]. Tree Genetics & Genomes, 2017, 13(5): 103. [6] 陈张彬. 甘蓝型油菜中5个脂酰-ACP硫酯酶基因的克隆及表达分析[D]. 长沙: 湖南农业大学, 2018. [7] 元冬娟, 吴湃, 江黎明. 高等植物的酰基-ACP硫酯酶研究进展[J]. 中国油料作物学报, 2009, 31(1): 103-108. [8] 王凯. 木姜子FATB基因的克隆和功能分析[D]. 杨凌: 西北农林科技大学, 2019. [9] SALAS J J, OHLROGGE J B. Characterization of substrate specificity of plant FatA and FatB acyl-ACP thioesterases [J]. Archives of Biochemistry and Biophysics, 2002, 403(1): 25 − 34. [10] DONG S, LIU Y, XIONG B, et al. Transcriptomic analysis of a potential bioenergy tree, Pistacia chinensis Bunge, and identification of candidate genes involved in the biosynthesis of oil [J]. BioEnergy Research, 2016, 9(3): 740 − 749. doi: 10.1007/s12155-016-9716-4 [11] 李昊远, 郝翠翠, 潘丽娟, 等. 花生酰基载体蛋白硫酯酶(FATB2)基因的克隆与表达分析[J]. 花生学报, 2017, 46(4): 7 − 14. doi: 10.14001/j.issn.1002-4093.2017.04.002 [12] CHEN X, DUAN X, WANG S, et al. Virus-induced gene silencing (VIGS) for functional analysis of MYB80 gene involved in Solanum lycopersicum cold tolerance [J]. Protoplasma, 2019, 256(2): 409 − 418. doi: 10.1007/s00709-018-1302-5 [13] 金龙飞, 尹欣幸, 曹红星. 油棕体细胞胚胎发生的研究进展[J]. 江苏农业科学, 2021, 49(13): 29-35. doi: 10.15889/j.issn.1002-1302.2021.13.006 [14] WOITTIEZ L S, VAN WIJK M T, SLINGERLAND M, et al. Yield gaps in oil palm: a quantitative review of contributing factors [J]. European Journal of Agronomy, 2017, 83: 57 − 77. doi: 10.1016/j.eja.2016.11.002 [15] 范世航, 刘念, 华玮. 油料作物油脂合成调控研究进展[J]. 中国油料作物学报, 2021, 43(3): 361-375. doi: 10.19802/j.issn.1007-9084.2021097 [16] KACZMARZYK D, HUDSON E P, FULDA M. Arabidopsis acyl-acyl carrier protein synthetase AAE15 with medium chain fatty acid specificity is functional in cyanobacteria [J]. AMB Express, 2016, 6(1): 7. doi: 10.1186/s13568-015-0169-5 [17] TAN K W M, LEE Y K. Expression of the heterologous Dunaliella tertiolecta fatty acyl-ACP thioesterase leads to increased lipid production in Chlamydomonas reinhardtii [J]. Journal of Biotechnology, 2017, 247: 60 − 67. doi: 10.1016/j.jbiotec.2017.03.004 [18] HAWKINS D J, KRIDL J C. Characterization of acyl-ACP thioesterases of mangosteen (Garcinia mangostana) seed and high levels of stearate production in transgenic canola. [J]. The Plant Journal: for Cell and Molecular Biology, 1998, 13(6): 743 − 752. [19] MORENO-PÉREZ A J, VENEGAS-CALERÓN M, VAISTIJ F E, et al. Reduced expression of FatA thioesterases in Arabidopsis affects the oil content and fatty acid composition of the seeds [J]. Planta, 2012, 235(3): 629 − 639. doi: 10.1007/s00425-011-1534-5 [20] 郝昭程, 王腾飞, 李忠奎, 等. 拟南芥硫酯酶基因在毕赤酵母中的表达[J]. 生物工程学报, 2015, 31(1): 115-122. [21] JARVIS B A, ROMSDAHL T B, MCGINN M G , et al. CRISPR/Cas9-induced fad2 and rod1 mutations stacked with fae1 confer high oleic acid seed oil in pennycress (Thlaspi arvense L.) [J]. Frontiers in Plant Science, 2021(12):652319. -

 点击查看大图
点击查看大图

图(4)
计量
- 文章访问数: 485
- HTML全文浏览量: 123
- PDF下载量: 10
- 被引次数: 0
