


 下载:
下载:

-
植物呼吸爆发氧化酶(Respiratory burst oxidase homologue, Rboh)即NADPH氧化酶(NADPH oxidase, NOX)是动物NADPH氧化酶核心亚基gp91phox 的同源物[1]。ROS(Reactive Oxygen Species)包括O2.−,H2O2,.OH等,主要由质膜上的Rboh、过氧化物酶和胺氧化酶催化形成。植物在响应逆境胁迫时,Rboh将电子传递给O2生成超氧阴离子,在超氧化物歧化酶的作用下产生H2O2,ROS迅速增加作为信号分子传递到下游基因。Rboh作为产生ROS的关键活性酶,在植物的生长发育起关键作用,调节细胞中的许多重要代谢途径。在拟南芥(Arabidopsis thaliana)中AtRbohD和AtRbohF是AtRbohs基因家族中表达最高的基因,其他的AtRboh基因则保持在低表达水平[2]。高表达的AtRbohD在细胞程序性死亡、细胞壁木质化、对胁迫的信号传导中发挥着重要作用[3-5]。模式识别受体(PRRs)对病原相关分子模式(PAMPs)的感知导致ROS的爆发主要依赖于AtRbohD。RbohF可能在某些反应中与RbohD协同作用[6]。在拟南芥 atrbohd/ atrbohf双重突变体的ROS含量降低,ABA诱导的气孔关闭,ROS积累及Ca2+通道的活跃程度均削弱[3, 7-8]。植物NADPH氧化酶的活性主要受Ca2+、蛋白磷酸化、Rac蛋白及激素调控。Ca2+的拮抗剂EGTA能有效抑制马铃薯(Solanum tuberosum)StRboh蛋白诱导的ROS[9];Ca2+通过与AtRbohD蛋白N端的EF−手性基序结合,改变其蛋白结构,从而调控AtRbohD活性[10]。磷酸化状态的AtCDPK5能进一步磷酸化AtRbohD蛋白,从而激活活性氧的爆发[11]。AtBik1(botrytis-induced kinase1)能够磷酸化AtRbohD蛋白的第343位和第347位丝氨酸,激活其活性,促进ROS的积累,同时正反馈调节Ca2+信号,促进Ca2+的细胞内流,引起气孔关闭[6]。Rac是一种具有GTP酶活性的蛋白,调控ROS的形成,进而影响细胞的各种生理过程和增强植物抗病能力。烟草中NtRac5基因通过负调控Rboh基因活性参与H2O2的合成[12]。研究发现,水稻(Oryza sativa)OsRac1与OsRbohB蛋白的N端相互作用,同时,细胞质中的Ca2+浓度的升高也促进了RbohB的酶活性的提高[13]。水杨酸(SA)、茉莉酸(JA)和脱落酸(ABA)广泛参与调控植物的抗逆响应过程,它们可促进Rboh的表达及酶活性提高。拟南芥外源施加MeJA可以诱导AtRbohD和AtRbohF表达量升高[14]。外源施加ABA可以直接刺激Rboh氧化酶的活性,产生ROS类物质,参与植株抗逆的生理调节过程[7]。
木薯(Manihot esculenta)作是世界三大薯类之一,是重要的粮食作物和生物质能源作物[15]。木薯作为热区作物对低温,干旱等环境因素非常敏感,病害也是影响木薯产量的一个重要因素[16-18]。植物NADPH氧化酶家族中RbohD基因已被证实在植物与病原微生物的互作中主要负责ROS的形成。目前,木薯中还没有关于MeRbohD相关的报道。推测MeRbohD可能作为信号传导中的重要基因参与木薯的抗逆响应途径。笔者在获取MeRbohD的互作蛋白基础上,利用酵母点对点杂交验证方法验证MeRbohD和MeNOSIP(nitric oxide synthase-interacting protein)的互作关系;从木薯cDNA中扩增得到MeNOSIP的CDs序列,利用生物信息学方法初步分析MeNOSIP基因和蛋白的理化性质;同时,利用实时荧光定量PCR分析MeNOSIP在JA,SA,ABA诱导下的表达情况,旨在为木薯的遗传改良提供参考资料。
-
华南8号木薯品种(SC8),酵母菌株AH109,大肠杆菌E. coli DH5α,酵母双杂交载体质粒pGADT7,pGBKT7,pGADT7-LargeT,pGBKT7-p53,pGBKT7-laminc,pGADT7-MeRbohD均由海南大学生物资源可持续利用生物重点实验室提供。
-
以木薯SC8的cDNA为模板,用Thermo的高保真酶和基于木薯基因组数据库设计的基因编码区引物(primerF:ATGCCTCAGAGACACTCG/primer R: TTAAATCTTCATTGCGGC)进行目的基因CDs序列的扩增,扩增程序为:95 ℃ 3 min;30个循环,95 ℃ 10 s,58 ℃ 10 s,72 ℃ 20 s;72 ℃ 5 min。扩增产物取2 µL进行1%琼脂糖凝胶电泳检测。将扩增获得的目的条带通过TA克隆到中间载体pEASY-Blunt,转化大肠杆菌感受态,筛选阳性克隆送华大基因公司测序。
-
利用蛋白质网站ExPAS(
http://web.expasy.org/protparam/ )鉴定MeNOSIP蛋白的理化性质。利用网站(http://www.csbio.sjtu.edu.cn/bioinf/Cell-PLoc-2/ )对MeNOSIP蛋白进行亚细胞定位预测。利用MapGene2chrom(http://mg2c.iask.in/mg2c_v2.1/ )进行MeNOSIP的染色体定位分析。运用MEGA7.0对木薯(Manihot esculenta)、拟南芥(Arabidopsis thaliana)、水稻(Oryza sativa)、烟草(Nicotiana tomentosiformis)、番茄(Solanum lycopersicum)、马铃薯(Solanum tuberosum)、巴西橡胶树(Hevea brasiliensis)、玉米(Zea mays)、大麦(Hordeum vulgare)等的NOSIP蛋白序列进行多序列比对,构建NJ进化树。 -
为了验证筛选到的MeNOSIP与MeRbohD是否真实互作,笔者将MeNOSIP构建至诱饵载体pGBKT7-BD,与pGAD共转入酵母AH109菌株,涂布在SD/TL平板。鉴定无误后,点在SD/TL, SD/TLHA和SD/TLHA+20 mg·L−1 x-α-gal的平板中,验证MeNOSIP是否有自激活活性。将pGBKT7-MeNOSIP与pGADT7-MeRbohD共同转化酵母菌株AH109。28 ℃暗培养3 d后,挑取单菌落进行菌落PCR,鉴定阳性克隆。将阳性克隆点至SD/TL,SD/TLHA和SD/TLHA+20 mg·L−1 x-α-gal的平板中,28 ℃培养4 d后观察菌落生长情况,验证互作关系。
-
茉莉酸(JA)、水杨酸(SA)、脱落酸(ABA)等激素在植物应对各种胁迫下发挥重要的调节作用。为了探究MeRbohD和MeNOSIP是否会响应这3种激素的诱导,选取生长约30 d长势良好的木薯SC8幼苗,分别用JA(100 μmol·L−1),SA(2 μmol·L−1),ABA(100 μmol·L−1) 溶液喷雾处理。提取处理不同时间点下的木薯RNA进行反转录用于 MeRbohD和MeNOSIP基因的表达模式分析。qRT-PCR程序按SYBR Premix Ex TaqTM(TaKaRa)说明操作:95 ℃预变性30 s;40个循环包括95 ℃变性5 s,58 ℃退火30 s,72 ℃延伸30 s。Ubq10基因作为参照基因,计算2−ΔΔCt值。每个样品、每个时间点设3个生物学重复,重复3次实验。结果利用GraphPad软件绘图。
-
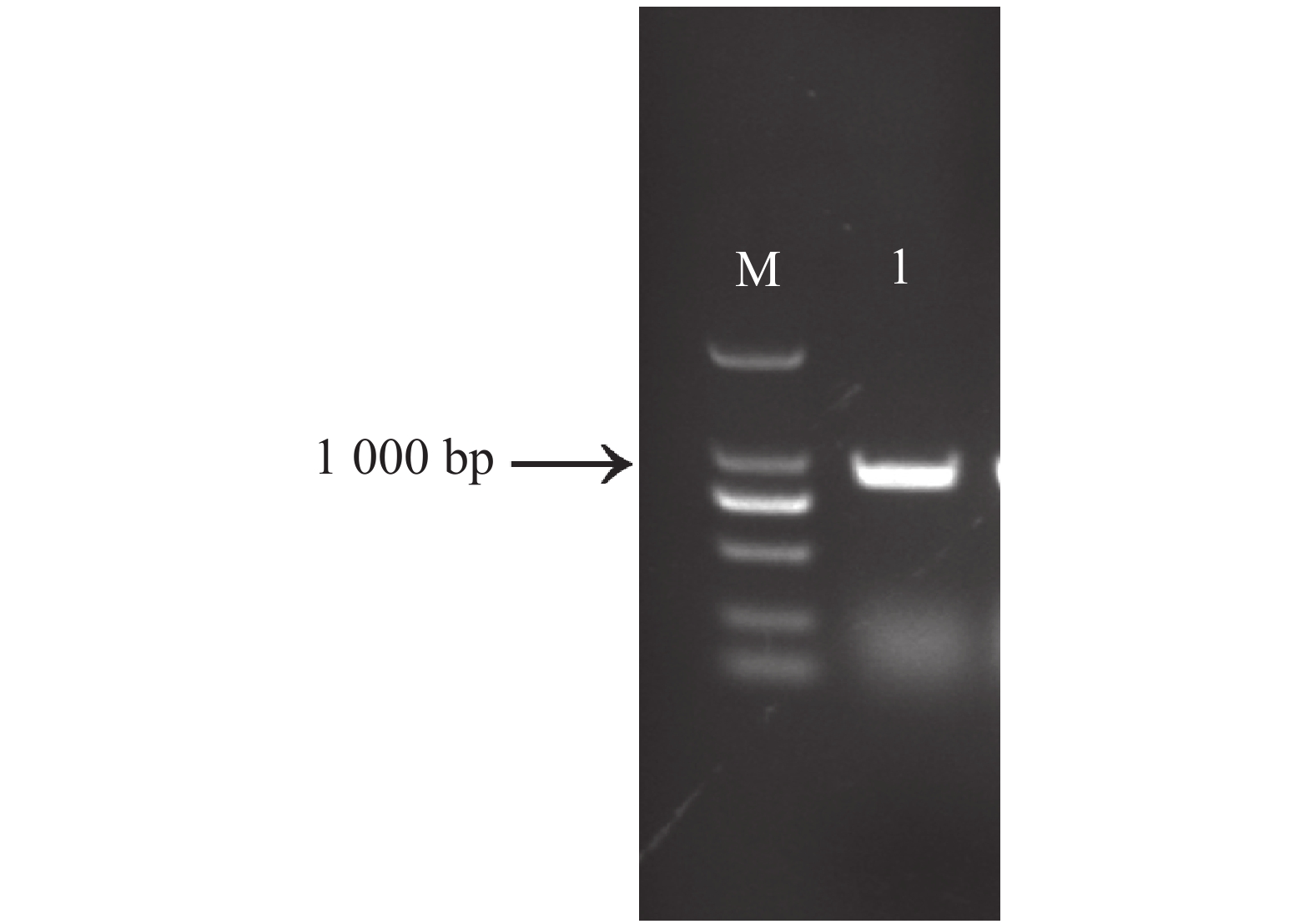
以木薯SC8的cDNA为模板,利用MeNOSIP编码区引物扩增得到1条约900 bp的目的片段(图1),从图1可知,电泳条带单一,无拖尾,特异性强,与预期大小一致。TA克隆到中间载体pEASY-Blunt、转化大肠杆菌感受态后测序。测序结果表明,该序列全长918 bp,编码1条含有305个氨基酸的多肽,命名为MeNOSIP(Phytozome编号:Manes.S039100.1;GeneBank登录号:XM_021746786.1)。
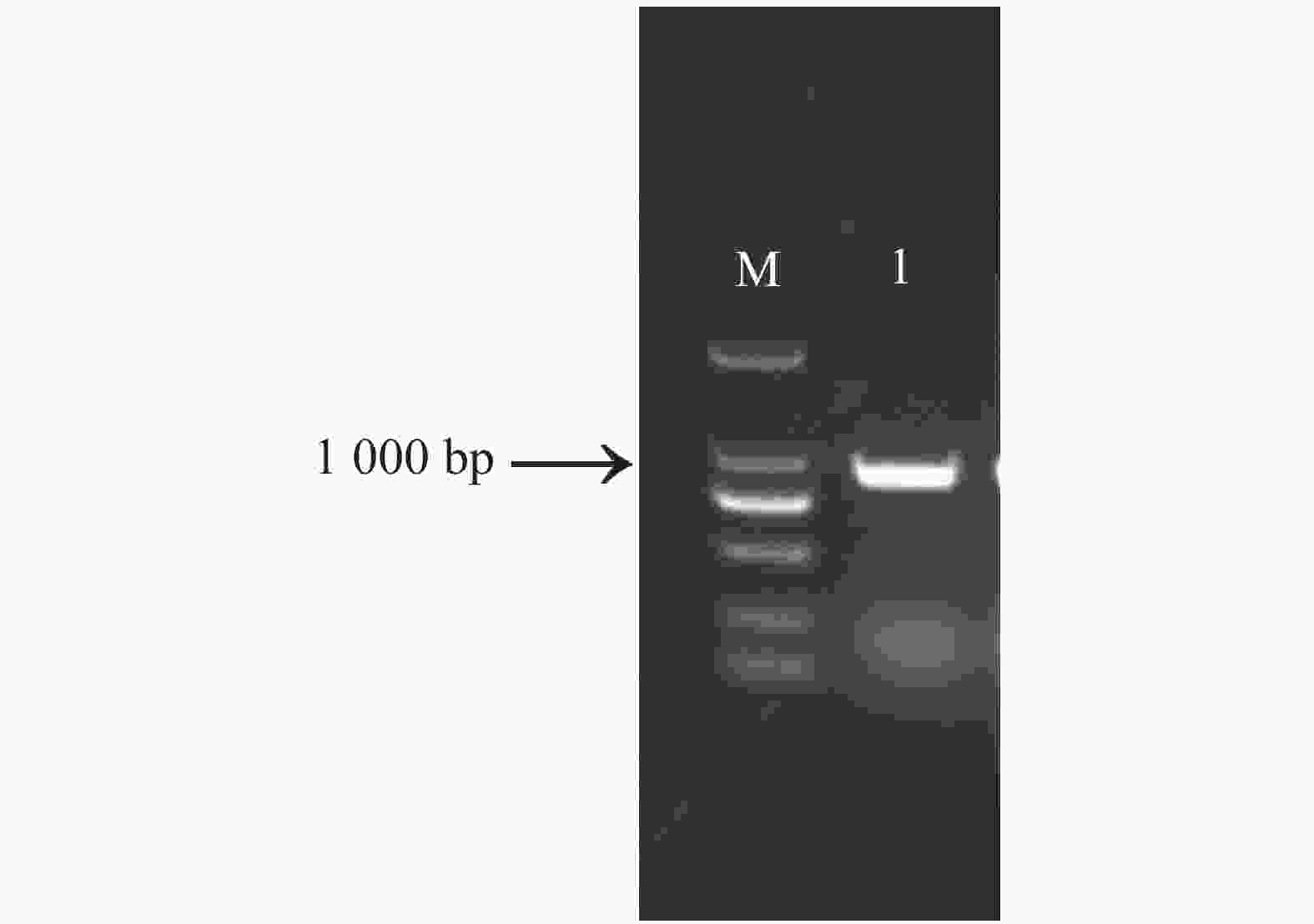
图 1 MeNOSIP PCR扩增
Figure 1. Amplification of the full length cDNA of MeNOSIP
-
对MeNOSIP蛋白进行理化性质分析,MeNOSIP蛋白分子式为C1475H2374N424O453S23,相对分子质量约为34.03×103,等电点约为8.95。此蛋白富含赖氨酸(11.8%),亮氨酸(9.2%),谷氨酸(7.5%),丝氨酸(7.2%)。蛋白不稳定指数为46.82,属于不稳定蛋白。对MeNOSIP做亲水性分析,从图2可知,该蛋白亲水系数为−0.628,属于亲水类蛋白。通过预测发现MeNOSIP定位于细胞膜。
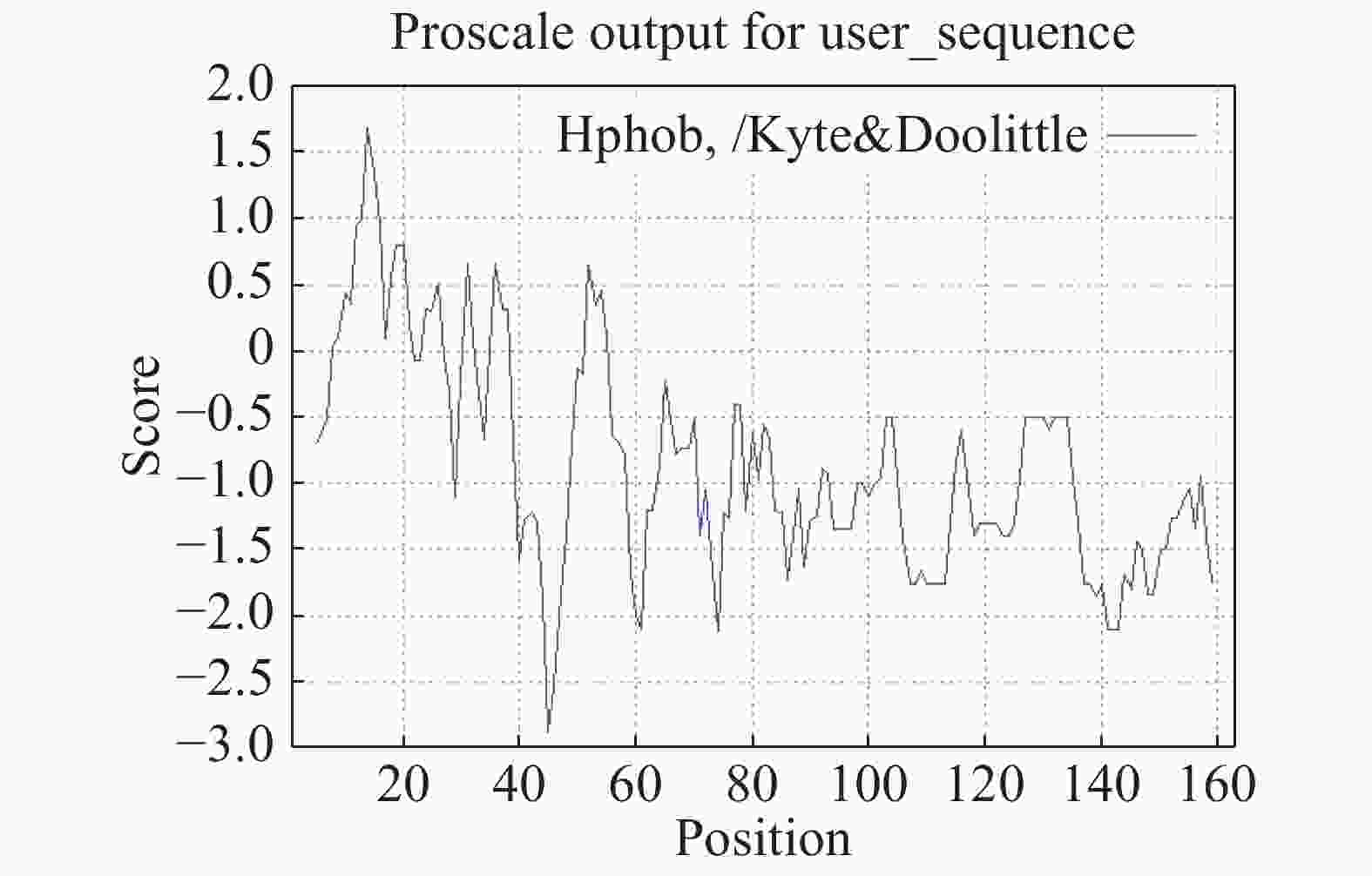
图 2 木薯MeNOSIP蛋白亲水性分析
Figure 2. Hydropilia analysis of MeNOSIP protein
-
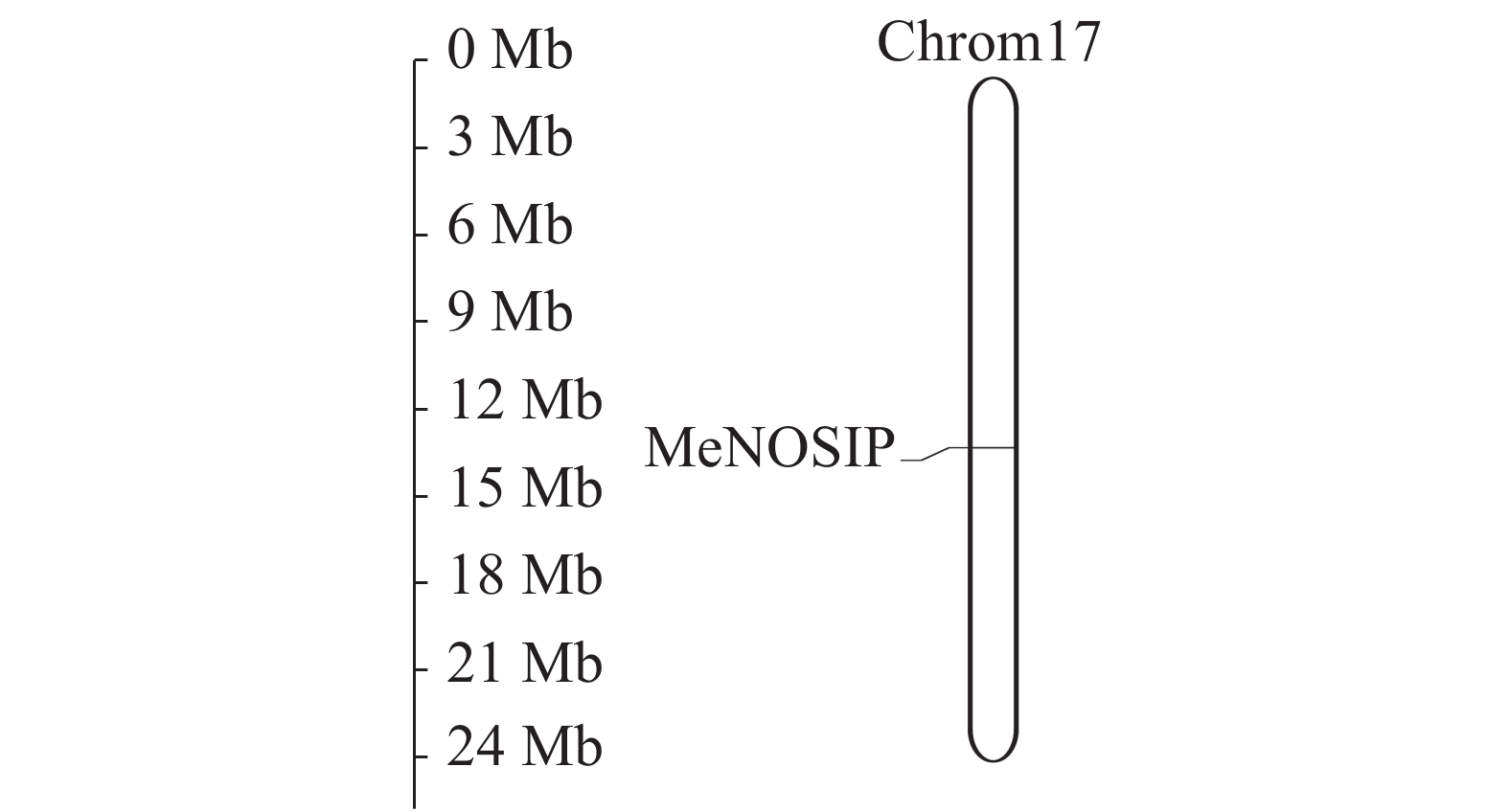
利用Phytozome网站下载木薯全基因组数据,在MapGene2chrom上分析MeNOSIP染色体定位。结果如图3所示:MeNOSIP基因定位在木薯第17号染色体上,基因位置从第14 842 551位碱基到14 844 636位碱基。
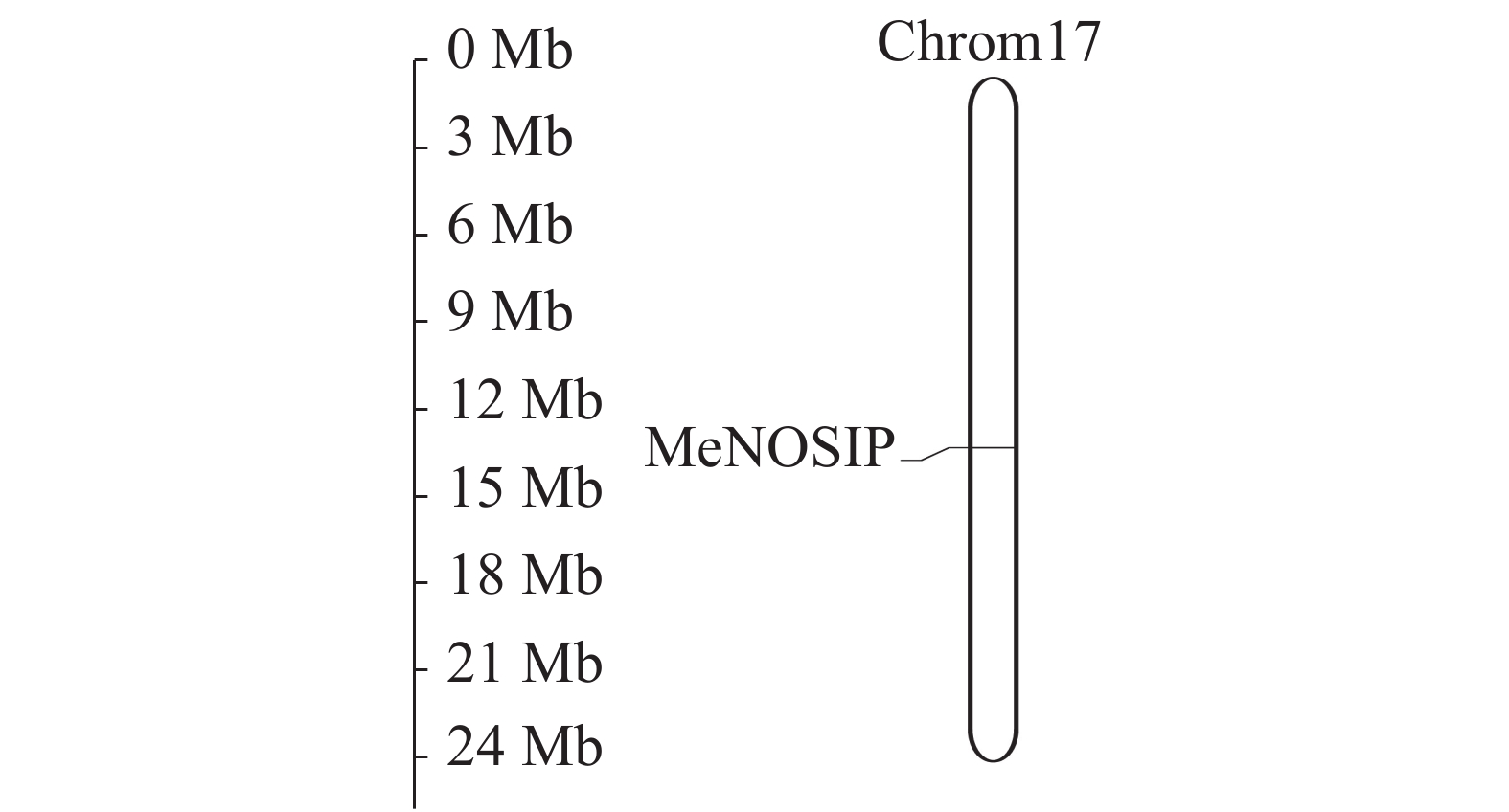
图 3 木薯MeNOSIP染色体定位
Figure 3. Chromosomal localization of the MeNOSIP gene in cassava
-
应用MEGA 7.0对木薯(XM_021746786.1),拟南芥(AT1G61620),水稻(LOC4332437),巴西橡胶树(LOC110661181),烟草(LOC107823767),番茄(LOC101247881),马铃薯(LOC102602935),玉米(LOC1103639300),大麦(AK370636.1)的NOSIP蛋白序列进行多序列比较和构建进化树。从图4可知:木薯NOSIP与巴西橡胶树的NOSIP亲缘关系最近,约为92.81%,与水稻,玉米的亲缘关系较远。
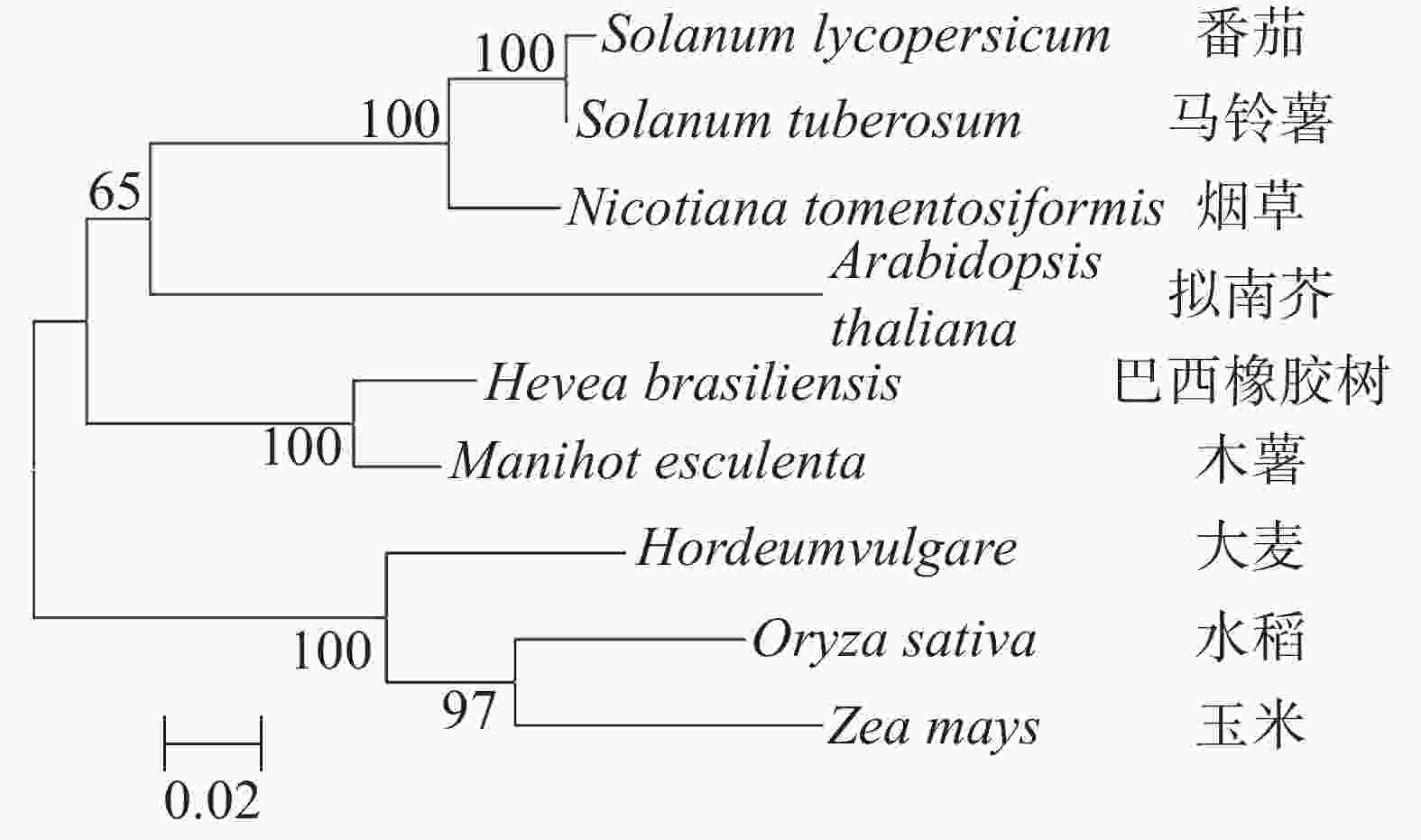
图 4 不同物种的NOSIP聚类分析
Figure 4. NOSIP cluster analysis
-
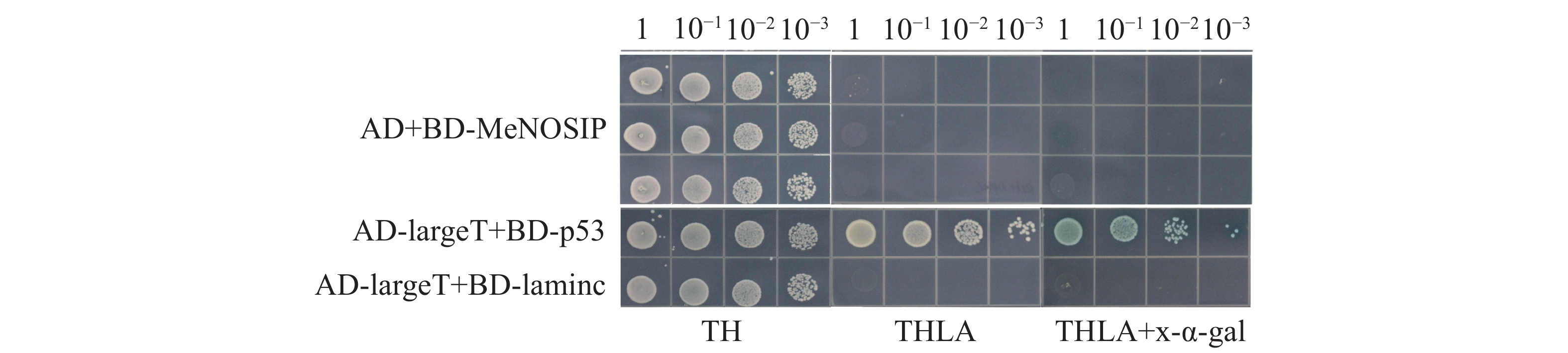
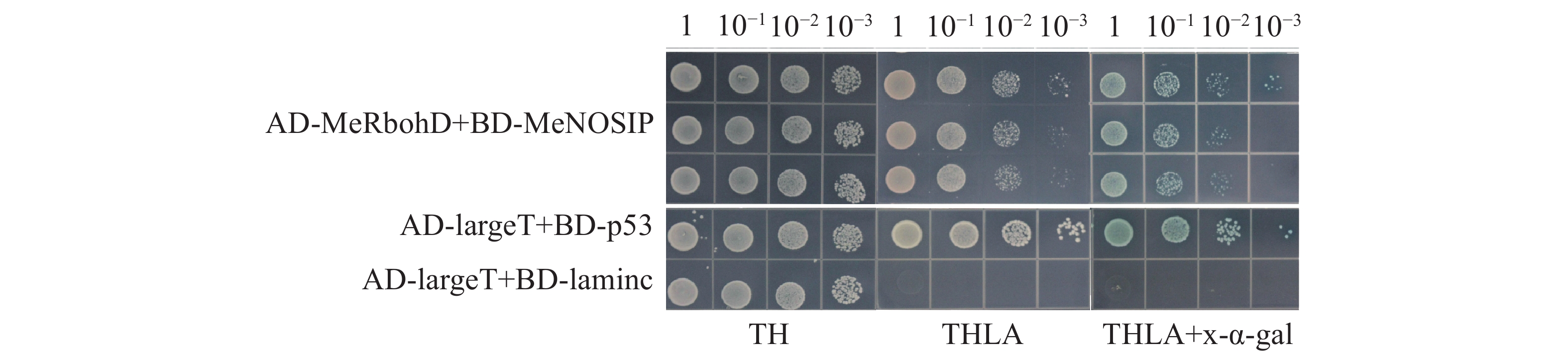
自激活检测中,实验组(BD-MeNOSIP+AD)和阳性对照(AD-largeT+BD-p53)、阴性对照(AD-largeT+BD-laminc)的酵母在SD/TL平板上均能正常生长,但仅有阳性对照在SD/TLHA平板上生长,在SD/TLHA+ x-α-gal平板上酵母菌落变蓝,实验组的生长状态与对照一致(图5),实验表明,MeNOSIP不具有自激活活性。在酵母双杂交验证中,实验组(AD-MeRbohD+BD-MeNOSIP)和对照在SD/TL 平板上均能正常生长,仅有实验组和阳性对照在SD/TLHA平板上生长,且在SD/TLHA+x-α-gal平板上酵母菌落变蓝(图6),结果表明,MeNOSIP蛋白与MeRbohD蛋白存在互相作用。
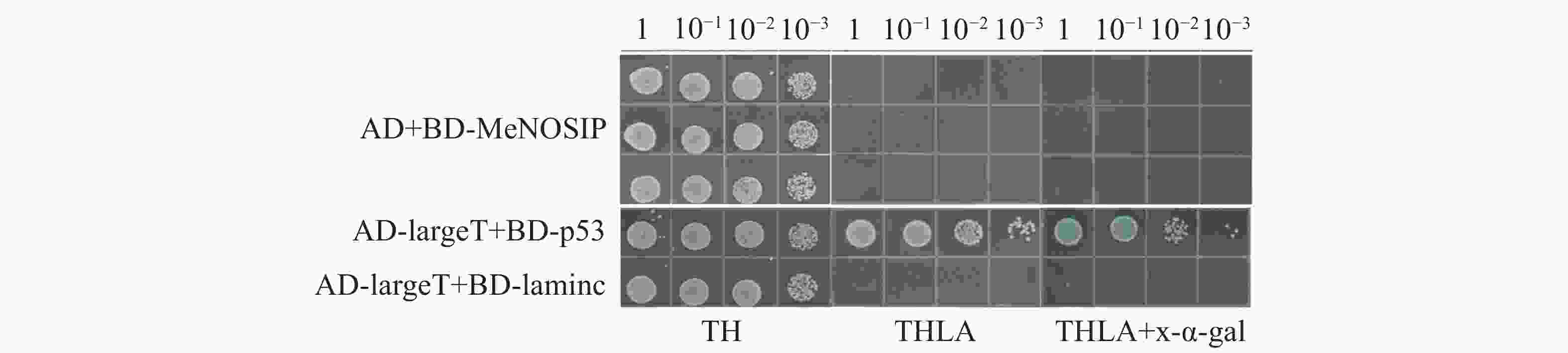
图 5 MeNOSIP蛋白自激活检测
Figure 5. Self-activation assay of MeNOSIP
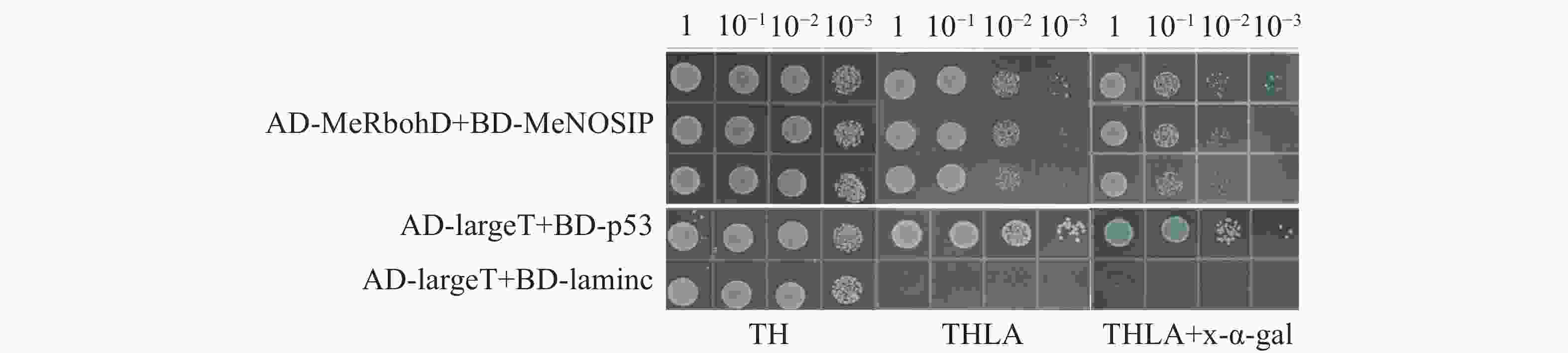
图 6 酵母双杂交验证MeRbohD和MeNOSIP的互作
Figure 6. Identification of the interaction between MeRbohD and MeNOSIP by yeast two-hybrid assays
-
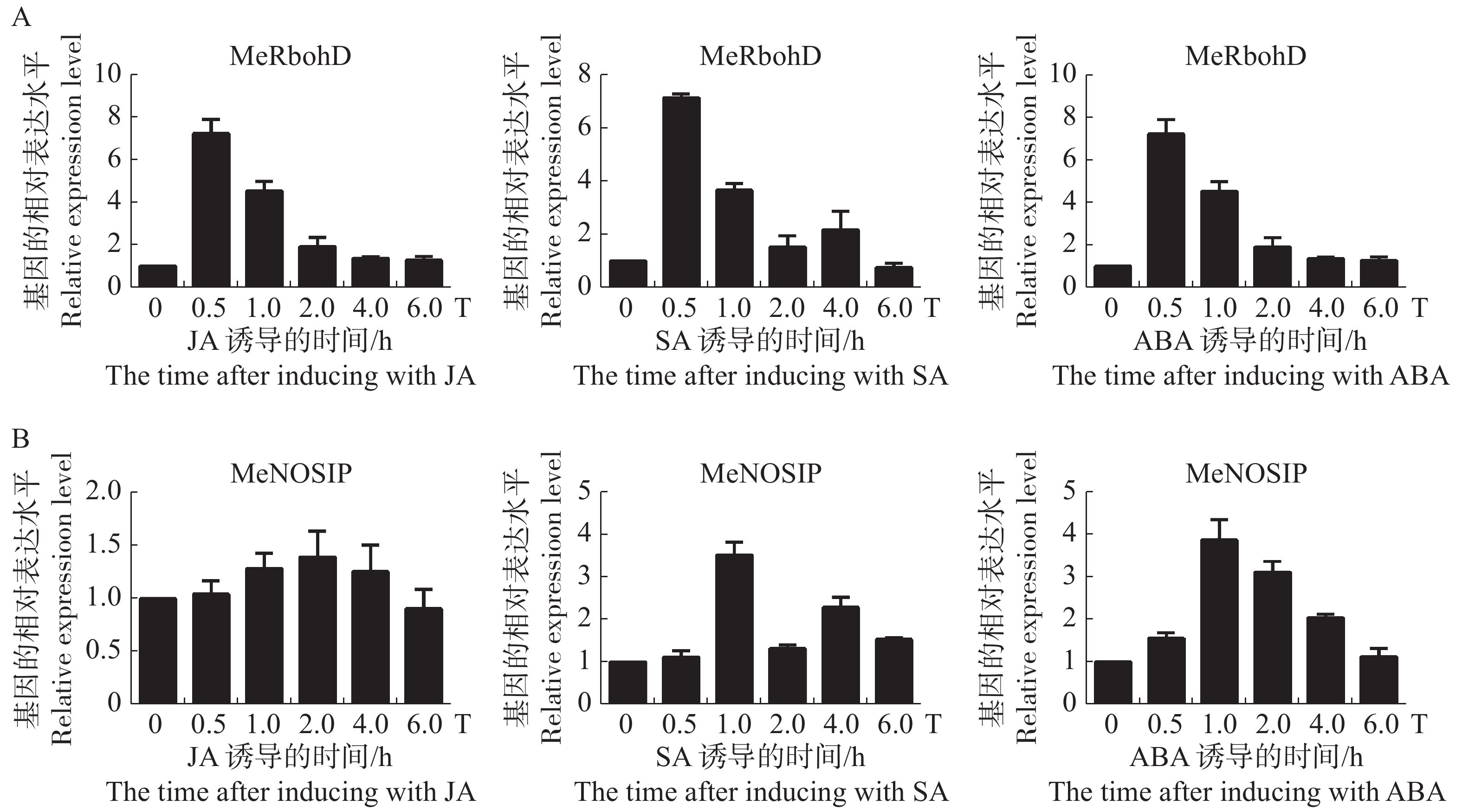
分析木薯MeRbohD和MeNOSIP基因在植物激素JA,SA,ABA诱导下的表达模式,结果分别如图7A,B所示。实验发现,MeRbohD均能在早期响应3种激素的诱导,分别在JA,SA处理30 min后,ABA处理3 h后基因表达上调5~10倍。JA处理下MeNOSIP的表达量没有明显的变化,推测MeNOSIP可能不参与JA引起的信号传导。SA处理1 h后和ABA处理3 h后,MeNOSIP的表达量上调约4倍。在ABA的诱导下,MeRbohD与MeNOSIP基因的表达量变化水平基本一致,因此,推测MeRbohD和MeNOSIP可能共同参与了由ABA介导抗逆通路。
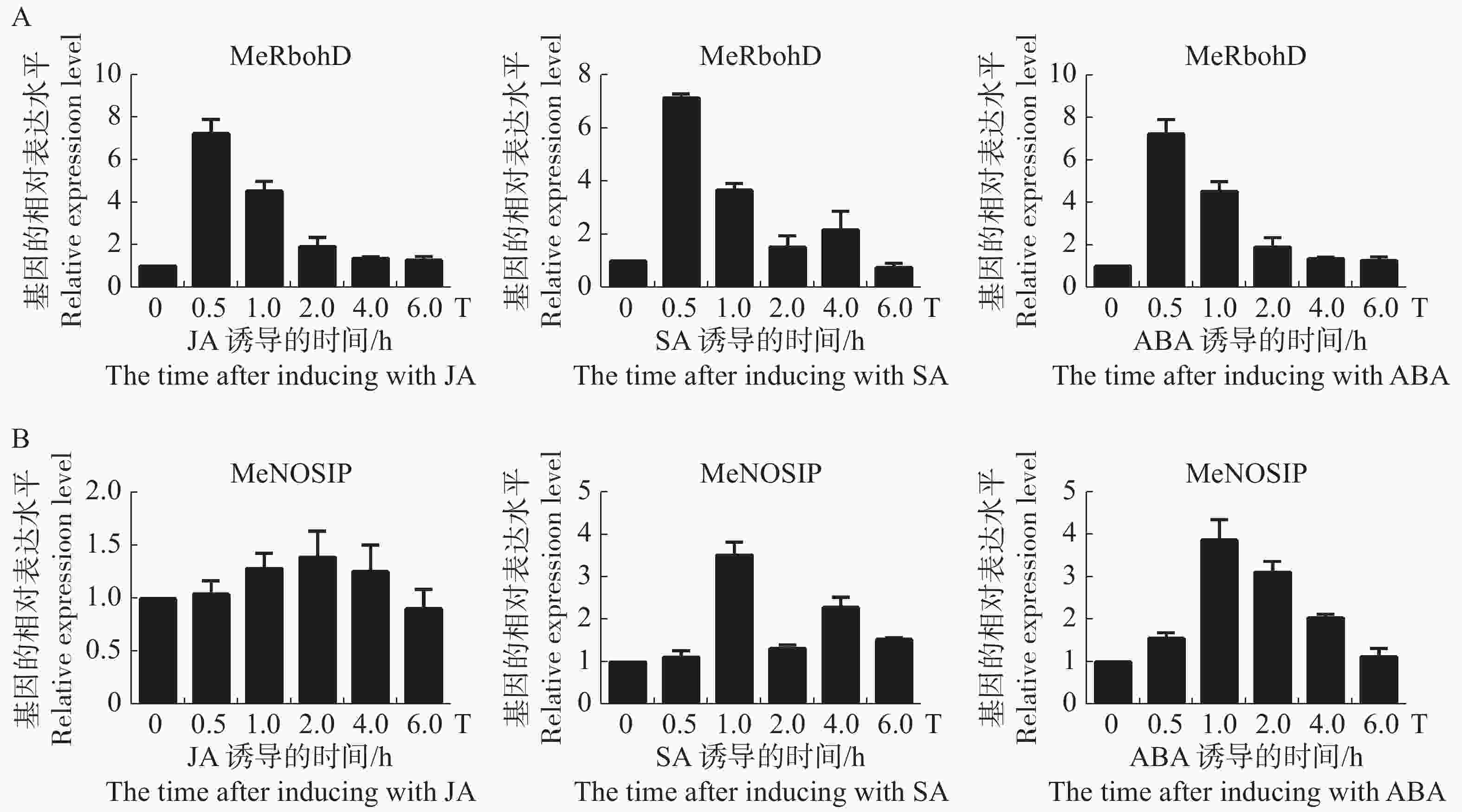
图 7 MeRbohD,MeNOSIP受到激素JA,SA和ABA诱导后的表达分析
Figure 7. Expression analysis of MeRbohD and MeNOSIP induced by JA, SA and ABA, respectively
-
木薯作为热区重要的经济和粮食作物,其产量受到生物胁迫(细菌性枯萎病、褐色条斑病、粉蚧等)和非生物胁迫(低温、干旱)的制约,同时在木薯收获后,块根易发生生理性腐烂,不耐贮藏。ROS是植物中重要的信号分子,在调控生长发育、应对非生物胁迫和激发植物免疫反应中都起着重要的作用[19]。提高木薯块根清除ROS的能力,有利于延缓生理性腐烂的发生。RbohD作为ROS产生过程中的关键基因,在植物响应逆境胁迫过程中发挥着重要作用。植物Rboh的酶活性受到其互作蛋白的调控,如AtBik1,OsRac1。在获得MeRbohD新的候选互作蛋白MeNOSIP的基础上,本研究中笔者克隆了MeNOSIP的编码基因,全长918 bp,编码305个氨基酸,MeNOSIP蛋白分子式为C1475H2374N424O453S23,相对分子质量约为34.03×103,等电点约为8.95。MeNOSIP基因定位于木薯17号染色体,蛋白与巴西橡胶树NOSIP有较高的同源性(92.81%)。亚细胞定位预测MeNOSIP蛋白存在于细胞膜上。
目前,在植物中有还没有关于NOSIP的功能研究报道。NOSIP作为一氧化氮合酶(NOS)的伴侣蛋白,与NOS互作调节生物体内一氧化氮水平[20]。细胞内Ca2+通过与钙调蛋白的相互作用来激活神经元的一氧化氮合酶(NOS)家族催化产生一氧化氮[21]。一氧化氮(NO)作为一种高效的信号分子,与活性氧(ROS)反应生成更强大的氧化剂活性氮(RNS)调节细胞功能[22],信号传导途径和各种病理过程中发挥着重要作用[22]。ROS和RNS)作为细胞内的第2信使参与信号转导,细胞生长,分化。但过量的ROS和RNS的产生会导致严重的细胞损伤。NOSIP则在这一过程中负责调节神经元细胞中的一氧化氮稳态[20]。
本实验成功克隆得到MeRbohD和MeNOSIP的编码基因,采用酵母点对点杂交方法验证了MeNOSIP与MeRbohD互作关系。MeNOSIP蛋白预测定位于细胞膜,推测MeRbohD与MeNOSIP互作位置在细胞膜上。进一步研究发现,JA,SA,ABA均能诱导MeNOSIP与MeRbohD的表达量提高,特别是在ABA的诱导下,MeRbohD与MeNOSIP的表达量变化趋势一致,因此,推测MeRbohD和MeNOSIP可能共同参与了由ABA介导抗逆通路。在神经元细胞中NOSIP与NOS(一氧化氮合酶)互作调控胞内一氧化氮水平[20]。在哺乳动物的吞噬细胞中一氧化氮与活性氧(ROS)反应形成活性氮(RNS)作为第2信使进行信号传导[22]。因此,推测在木薯防卫反应中,RbohD与NOSIP、NOS三者之间相互作用调控胞内RNS的生成,进行信号传导。本研究所用到的酵母cDNA文库是由木薯致病细菌致病细菌Xanthomonas axonopodis pv. manihotis(Xam)处理木薯SC8植株后构建得到的。推测木薯细胞膜表面的模式识别受体接受到病原微生物激发的信号后,激活细胞内一系列信号传导和防卫反应,例如Ca2+增加,MAPK的活化,激活了细胞膜上MeRbohD的活性,导致ROS的爆发,与MeNOSIP互作调控胞内一氧化氮的生成、气孔关闭,胼胝质沉淀等。本实验验证了MeNOSIP与MeRbohD的真实互作,笔者推测在木薯细胞膜上MeNOSIP与MeRbohD互作,并能调控MeRbohD的活性,进而调节植物细胞中ROS的生成。
Verification of the Interaction Between Cassava MeNOSIP and MeRbohD Proteins and Their Response to Exogenous Hormones
-
摘要: 为了解木薯(Manihot esculenta)MeRbohD在抗病途径中的功能,笔者在获得MeRbohD候选互作蛋白MeNOSIP(nitric oxide synthase-interacting protein,一氧化氮合酶互作蛋白)的基础上,通过RT-PCR克隆获得MeNOSIP基因编码区序列,并进行生物信息学分析。结果表明,MeNOSIP的CDs序列全长918 bp,编码含305个氨基酸的多肽,属于不稳定的亲水类蛋白质,亚细胞定位预测MeNOSIP在细胞膜上表达。采用酵母杂交验证发现,MeRbohD与MeNOSI真实互作。MeRbohD和MeNOSIP在JA,SA,ABA诱导下基因均上调表达;ABA处理下,MeRbohD和MeNOSIP基因表达变化趋势一致,推测它们可能共同参与了由ABA介导的抗逆通路。Abstract: MeNOSIP (Nitric oxide synthase interaction protein) gene coding region sequence in cassava was obtained by RT-PCR cloning based on the MeRbohD candidate interaction protein MeNOSIP to study the function of cassava MeRbohD in the disease resistance pathway. Bioinformatics analysis revealed that the CDs sequence of MeNOSIP was 918 bp, encoding a polypeptide containing 305 amino acids, and MeNOSIP belongs to an unstable hydrophilic protein. The subcellular localization predicted the expression of MeNOSIP on the cell membrane. Yeast hybridization was used to verify the true interaction between MeRbohD and MeNOSIP. MeRbohD and MeNOSIP genes were up-regulated under the induction of JA, SA, and ABA treatment, and MeRbohD and MeNOSIP gene expression under the ABA treatment tended to be consistent, which indicated that that they might be involved in the ABA-mediated stress resistance pathway.
-
Key words:
- MeRbohD /
- interaction protein /
- MeNOSIP /
- expression analysis
-
图 2 木薯MeNOSIP蛋白亲水性分析
负值表示亲水性,正值表示疏水性。
Fig. 2 Hydropilia analysis of MeNOSIP protein
Negative values: hydrophilic; positive values: hydrophobicit.
图 6 酵母双杂交验证MeRbohD和MeNOSIP的互作
Fig. 6 Identification of the interaction between MeRbohD and MeNOSIP by yeast two-hybrid assays
-
[1] BABIOR B M, LAMBETH J D, NAUSEEF W. The neutrophil NADPH oxidase [J]. Archives of Biochemistry and Biophysics, 2002, 397(2): 342 − 344. doi: 10.1006/abbi.2001.2642 [2] TORRES M A, DANGL J L, JONES J D G. Arabidopsis gp91phox homologues AtrbohD and AtrbohF are required for accumulation of reactive oxygen intermediates in the plant defense response [J]. Proceedings of the National Academy of Sciences, 2002, 99(1): 517 − 522. doi: 10.1073/pnas.012452499 [3] TORRES M A, DANGL J L. Functions of the respiratory burst oxidase in biotic interactions, abiotic stress and development [J]. Current Opinion in Plant Biology, 2005, 8(4): 397 − 403. doi: 10.1016/j.pbi.2005.05.014 [4] MILLER G, SCHLAUCH K, TAM R, et al. The plant NADPH oxidase RBOHD mediates rapid systemic signaling in response to diverse stimuli [J]. Science Signaling, 2009, 2(84): 45. [5] DENNESS L, MCKENNA J F, SEGONZAC C, et al. Cell wall damage-induced lignin biosynthesis is regulated by a reactive oxygen species-and jasmonic acid-dependent process in Arabidopsis [J]. Plant Physiology, 2011, 156(3): 1364 − 1374. doi: 10.1104/pp.111.175737 [6] KADOTA Y, SHIRASU K, ZIPFEL C. Regulation of the NADPH oxidase RBOHD during plant immunity [J]. Plant and Cell Physiology, 2015, 56(8): 1472 − 1480. doi: 10.1093/pcp/pcv063 [7] MA L, ZHANG H, SUN L, et al. NADPH oxidase AtrbohD and AtrbohF function in ROS-dependent regulation of Na+/K+ homeostasis in Arabidopsis under salt stress [J]. Journal of Experimental Botany, 2012, 63(1): 305 − 317. doi: 10.1093/jxb/err280 [8] TORRES M A, JONES J D G, DANGL J L. Reactive oxygen species signaling in response to pathogens [J]. Plant Physiology, 2006, 141(2): 373 − 378. doi: 10.1104/pp.106.079467 [9] KOBAYASHI M, OHURA I, KAWAKITA K, et al. Calcium-dependent protein kinases regulate the production of reactive oxygen species by potato NADPH oxidase [J]. The plant cell, 2007, 19(3): 1065 − 1080. doi: 10.1105/tpc.106.048884 [10] OGASAWARA Y, KAYA H, HIRAOKA G, et al. Synergistic activation of the Arabidopsis NADPH oxidase AtrbohD by Ca2+ and phosphorylation [J]. Journal of Biological Chemistry, 2008, 283(14): 8885 − 8892. doi: 10.1074/jbc.M708106200 [11] DUBIELLA U, SEYBOLD H, DURIAN G, et al. Calcium-dependent protein kinase/NADPH oxidase activation circuit is required for rapid defense signal propagation [J]. Proceedings of the National Academy of Sciences, 2013, 110(21): 8744 − 8749. doi: 10.1073/pnas.1221294110 [12] HASSANAIN H H, SHARMA Y K, MOLDOVAN L, et al. Plant Rac proteins induce superoxide production in mammalian cells [J]. Biochem Biophys Res Commun, 2000, 272: 783 − 788. doi: 10.1006/bbrc.2000.2791 [13] WONG H L, PINONTOAN R, HAYASHI K, et al. Regulation of rice NADPH oxidase by binding of Rac GTPase to its N-terminal extension [J]. The Plant Cell, 2007, 19(12): 4022 − 4034. doi: 10.1105/tpc.107.055624 [14] MARUTA T, INOUE T, TAMOI M, et al. Arabidopsis NADPH oxidases, AtrbohD and AtrbohF, are essential for jasmonic acid-induced expression of genes regulated by MYC2 transcription factor [J]. Plant Science, 2011, 180(4): 655 − 660. doi: 10.1016/j.plantsci.2011.01.014 [15] HU W, XIA Z Q, YAN Y, et al. Genome-wide gene phylogeny of CIPK family in cassava and expression analysis of partial drought-induced genes [J]. Frontiers in Plant Science, 2015(6): 914. [16] 卢赛清, 盘欢, 马崇熙, 等. 2008年广西木薯低温冻害情况及应对措施[J]. 广西热带农业, 2009(1): 21 − 22. [17] OKOGBNIN E, SETTER T L, FERGUSON M, et al. Phenotypic approaches to drought in cassava [J]. Frontiers in Physiology, 2013(4): 93. [18] 方佳, 濮文辉, 张慧坚. 国内外木薯产业发展近况[J]. 中国农学通报, 2010, 26(16): 353 − 361. [19] VRANOVA E, INZE D, VAN B F. Signal transduction during oxidative stress [J]. Journal of Experimental Botany, 2002, 53(372): 1227 − 1236. doi: 10.1093/jxb/53.372.1227 [20] DREYER J, HIRLINGER D, MULLER-ESTERL W, et al. Spinal upregulation of the nitric oxide synthase-interacting protein NOSIP in a rat model of inflammatory pain [J]. Neuroscience Letters, 2003, 350(1): 13 − 16. doi: 10.1016/S0304-3940(03)00771-7 [21] GARTHWAITE J, CHARLES S L, CHESS-WILLIAMS R. Endothelium-derived relaxing factor release on activation of NMDA receptors suggests role as intercellular messenger in the brain [J]. Nature, 1988, 336(6197): 385 − 388. doi: 10.1038/336385a0 [22] HALLIWELL B. Phagocyte-derived reactive species: salvation or suicide? [J]. Trends in biochemical sciences, 2006, 31(9): 509 − 515. doi: 10.1016/j.tibs.2006.07.005 -

 点击查看大图
点击查看大图
图(7)
计量
- 文章访问数: 850
- HTML全文浏览量: 195
- PDF下载量: 50
- 被引次数: 0
