


-
石蒜绵粉蚧(Phenacoccus solani Ferris)隶属半翅目(Hemiptera)粉蚧科(Pseudococcidae)绵粉蚧属(Phenacoccus),体小,体长约2~3 mm、体宽约1~2 mm,体表覆盖蜡粉层,以口针刺入植物内部组织吸食汁液,食性杂,寄主范围广,喜群集生活,庞大种群的为害可在短时间内引起植物枯萎、死亡[1]。石蒜绵粉蚧以若虫、雌性成虫等虫态营固定取食,个体转移能力相对较弱,但随着全球物流业的日趋发达及人为长距离跨境跨域活动的日益频繁,加之石蒜绵粉蚧寄主范围广泛,使得该类昆虫随着各种产品的运输,尤其是多肉类观赏植物,在全球范围内扩散传播。近年来,各地不断出现石蒜绵粉蚧的危害报道[2-5]。同时,在我国进境植物及植物产品检疫中,石蒜绵粉蚧不断被截获[6],对我国农林生产具有极大的威胁[7-8]。目前,国内外针对石蒜绵粉蚧开展了部分基本生物学特征[9-11]、除害处理[12]等方面的研究,但其致害性评价或相关机制方面的内容鲜见报道。
刺探电位图谱技术(electrical penetration graph technique, EPG),又称刺吸电位技术,是目前用于研究刺吸式口器昆虫取食行为的最佳技术[13]。EPG技术可实现实时监控刺吸式昆虫取食时其口针所在的植物组织部位,并对相关的行为,如唾液分泌、口针穿刺叶肉细胞、吸食韧皮部汁液等进行有效的区分。该技术多用于分析研究刺吸式口器昆虫的取食习性[13-15]、致害性评价[16-18]、病毒传播[19]以及植物抗性评价[5]等。笔者采用该技术研究石蒜绵粉蚧的取食行为,定义其刺探电位图谱,旨在为其致害力评价及其他入侵生物学机制的进一步研究提供参考及基础数据。
-
石蒜绵粉蚧(Phenacoccus solani Ferris)截获自入境杭州邮检的日本进口植物,鉴定于浙江省检验检疫科学技术研究院植检实验室。在植物检疫实验室昆虫人工气候箱内进行室内种群维持,以景天科风车草属多肉植物胧月(Graptopetalum paraguayense)作为寄主植物,环境温度(25±1)℃,相对湿度(70±5)%,光周期14L︰10D。挑取3龄末幼虫进行隔离饲养,将羽化后2 d的成虫转移至铺有干燥滤纸的培养皿内进行饥饿处理,12 h后用于EPG测试。
已有研究报道了其他粉蚧在寄主上的取食波形,如扶桑绵粉蚧[15]、木薯绵粉蚧[20]等。由于EPG波形图谱与植物内部结构存在相关性,考虑到波形的可比性,将番茄(Lycopersicon esculentum)作为初步研究石蒜绵粉蚧EPG信号图谱的寄主植物,受试植物为盆栽的5叶苗,在上述环境下培养。为进一步研究石蒜绵粉蚧的食性,再以多肉植物胧月(株高2~3 cm)作为适应性寄主植物用于EPG测试,同等环境下培养。
-
供试仪器及软件为Giga-8刺探电位仪,购自荷兰瓦赫尼根大学;正极通过铜丝、银胶、金线(直径为18 μm)与粉蚧体背相连,负极铜针插入盆栽土壤内。每日早上9:00开始监测,连续监测时间为8 h。依据前期研究关于同属粉蚧取食行为EPG图谱定义的原则[15],利用Stylet+程序(
http://www.epgsystems.eu )记录及分析取食行为的EPG信号。 -
EPG数据参数分析参考SARRIA等[21]的方法,利用SPSS软件进行数据统计分析。
-
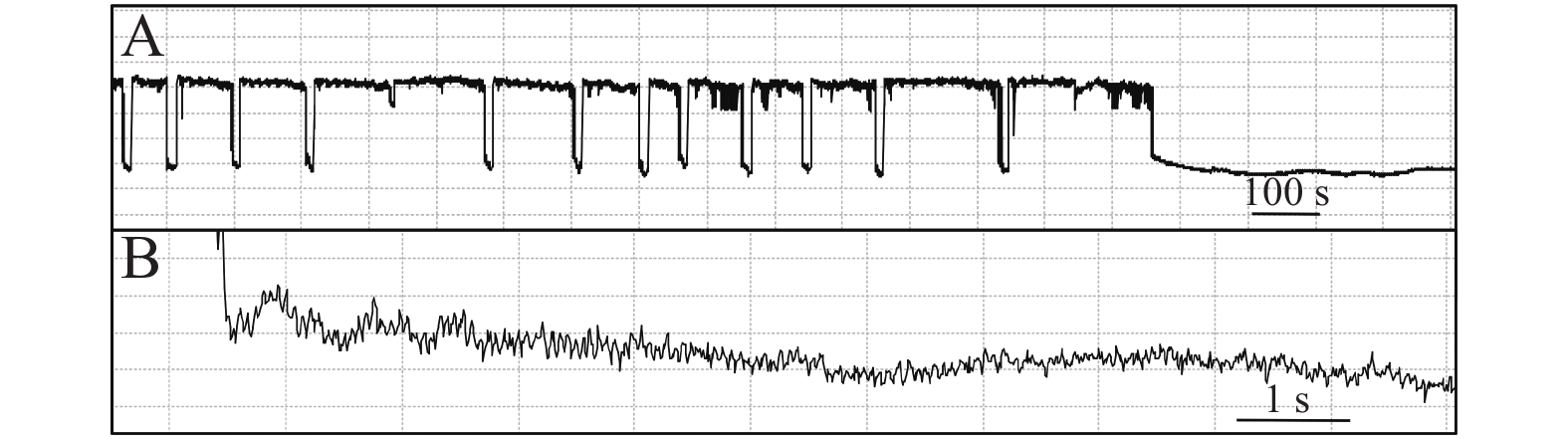
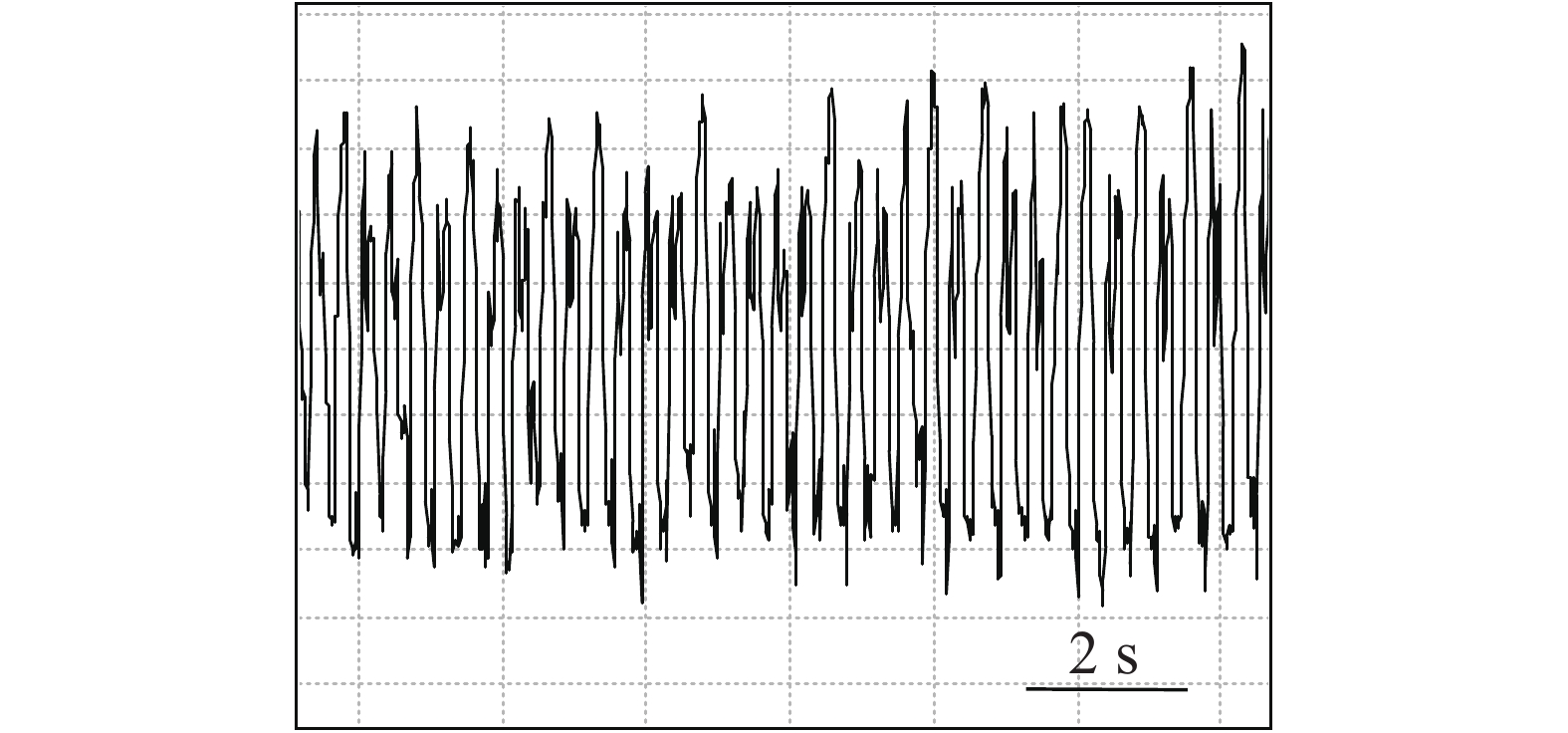
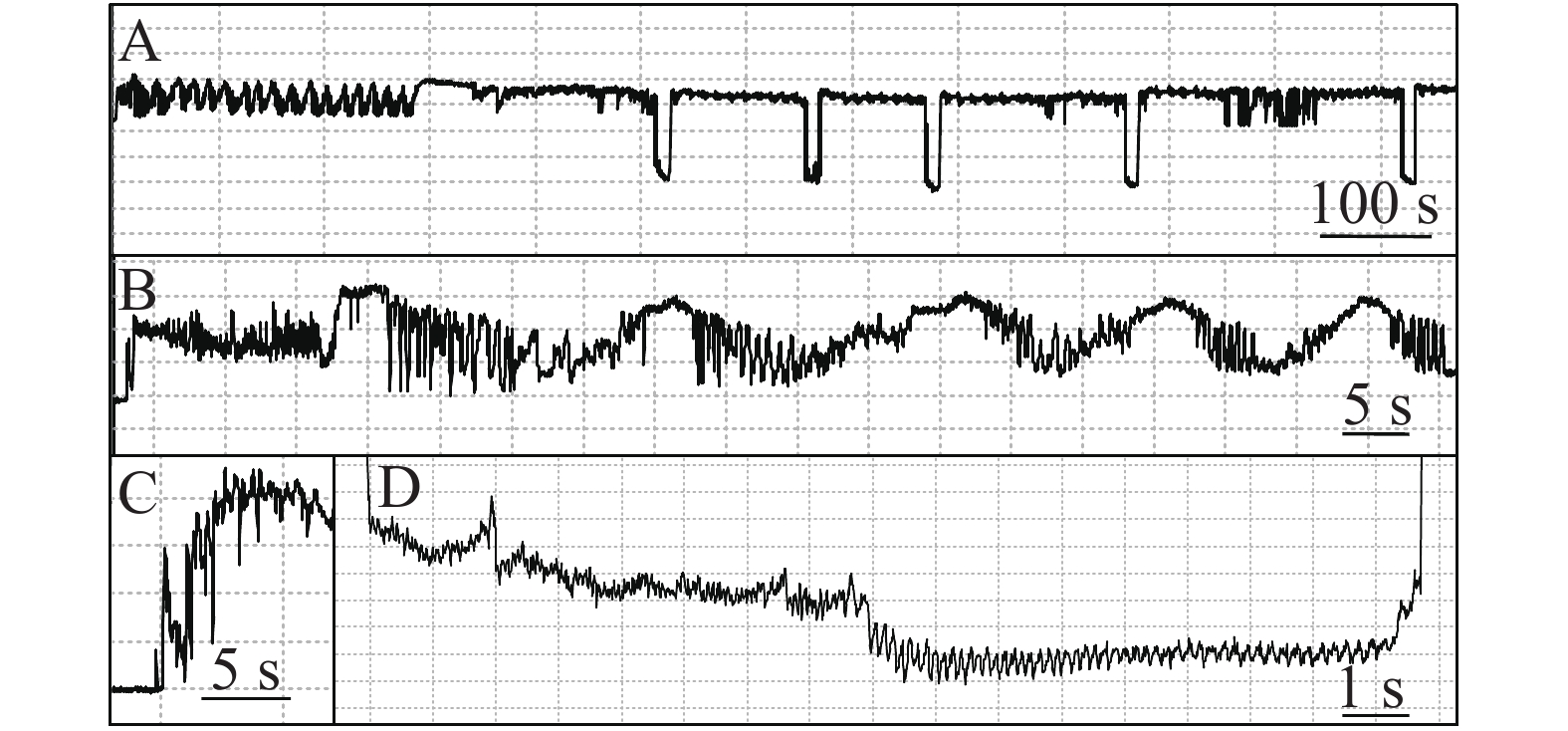
石蒜绵粉蚧的单次取食行为开始于以口针刺入终结于口针脱离叶片,记为1次刺探,记为Probe(Pr,图1-A)。期间出现的主要波型:A波:整个Pr过程中正压振幅最大的波(图1-B、图1-C),频率不规则,持续时长视不同种类的寄主而异,在番茄和胧月上分别为(9.6 ± 2.1)s(n = 30)和(4.4 ± 1.5)s(n = 30),后者显著短于前者(P < 0.01)。B波:首个B波一般跟随于A波发生,初始正压振幅与A波相同,频率为(0.083 ± 0.033) Hz;随着时间发展,振幅逐渐变小直至降至系统水平线(零压强)(图1-B)。在口针处于叶肉细胞层时,B波也可间歇性发生,持续时间由116 s至265 s不等。C波:跟随于A波发生,B波发生时,整合于C波发生(图1-A、图1-B)。C波为零压强波,振幅为A波的20%~30%。pd(potential drop)波:负压波,不规律地发生在C波阶段(图1-A、图1-D)。单个pd波持续时间因寄主种类而异,取食同一寄主时单个pd波持续时间变异很小。在番茄和胧月上,石蒜绵粉蚧单个pd波的平均时长分别为(16.2 ± 1.1) s(n = 30)和(30.7 ± 3.1) s(n = 30),后者显著长于前者(P < 0.01)。每个pd波内,根据波形发生的频率,可分为pd1波和pd2波,两者所占单个pd波的时长比例接近1,且该比例不随寄主种类变化而变化。E波:负压波,跟随于C波发生,振幅接近于pd波,但持续时间显著长于pd波(图2-A)。E波不发生其他粉蚧或蚜虫E波内典型的被动取食波形(E2波)(图2-B)。E1e(extracellular E1)波:发生于C波之后,电动势水平与C波相同(图3A虚线框内),持续时间数分钟至几小时不等。除电动势水平外,波形发生的其他特性与其他粉蚧或蚜虫的韧皮部相关波形E1波相似(图3-B)。G波:频率为8~10 Hz(图4)。
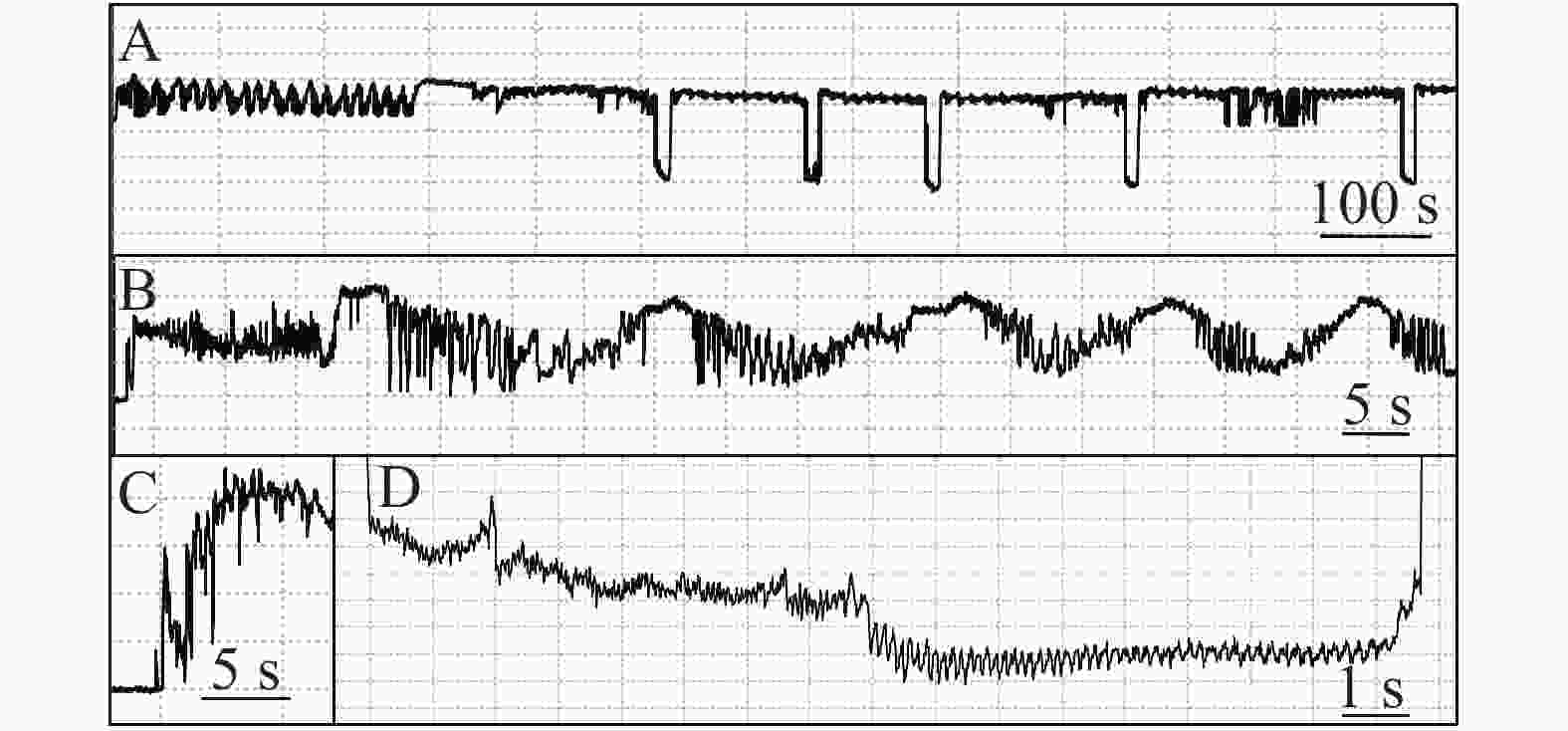
图 1 石蒜绵粉蚧取食行为EPG图谱
Figure 1. Electropenetrographs (EPG) of Phenacoccus solani.
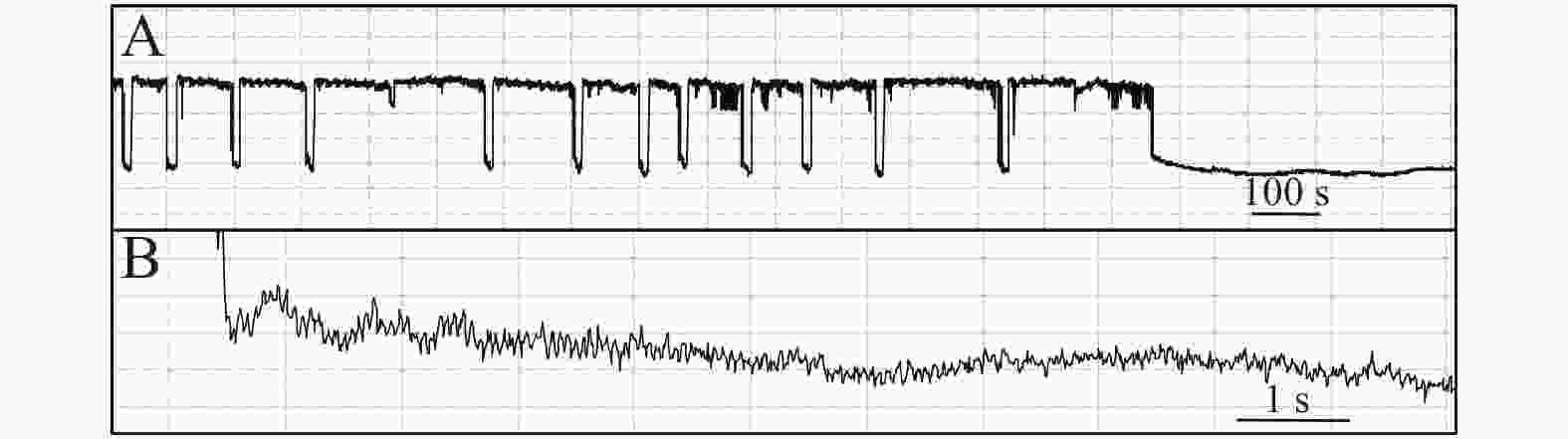
图 2 石蒜绵粉蚧取食行为EPG图谱(E波)
Figure 2. Electropenetrographs (EPG) of Phenacoccus solani (waveform E).
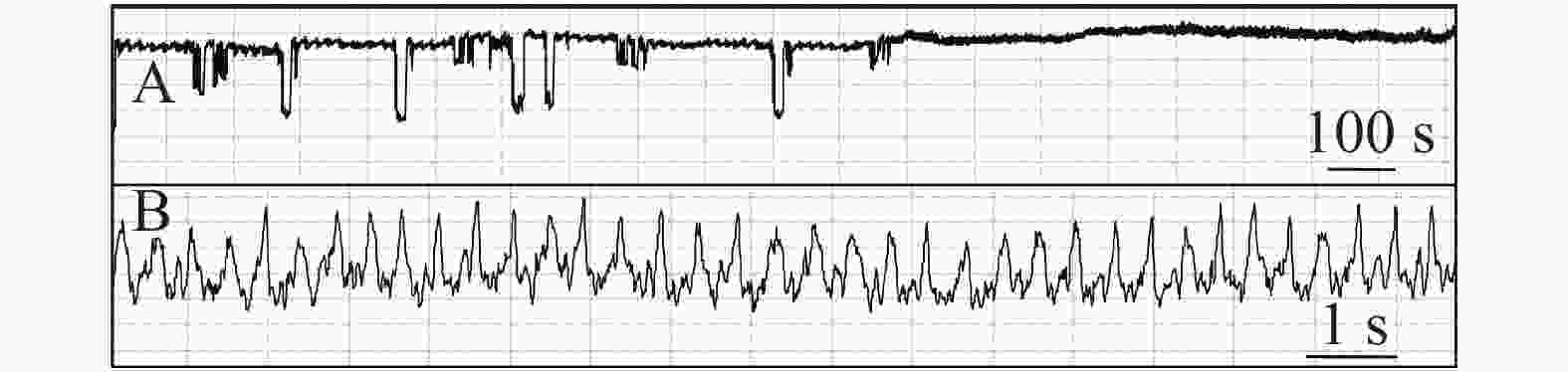
图 3 石蒜绵粉蚧取食行为EPG图谱(E1e波)
Figure 3. Electropenetrographs (EPG) of Phenacoccus solani (waveform E1e).
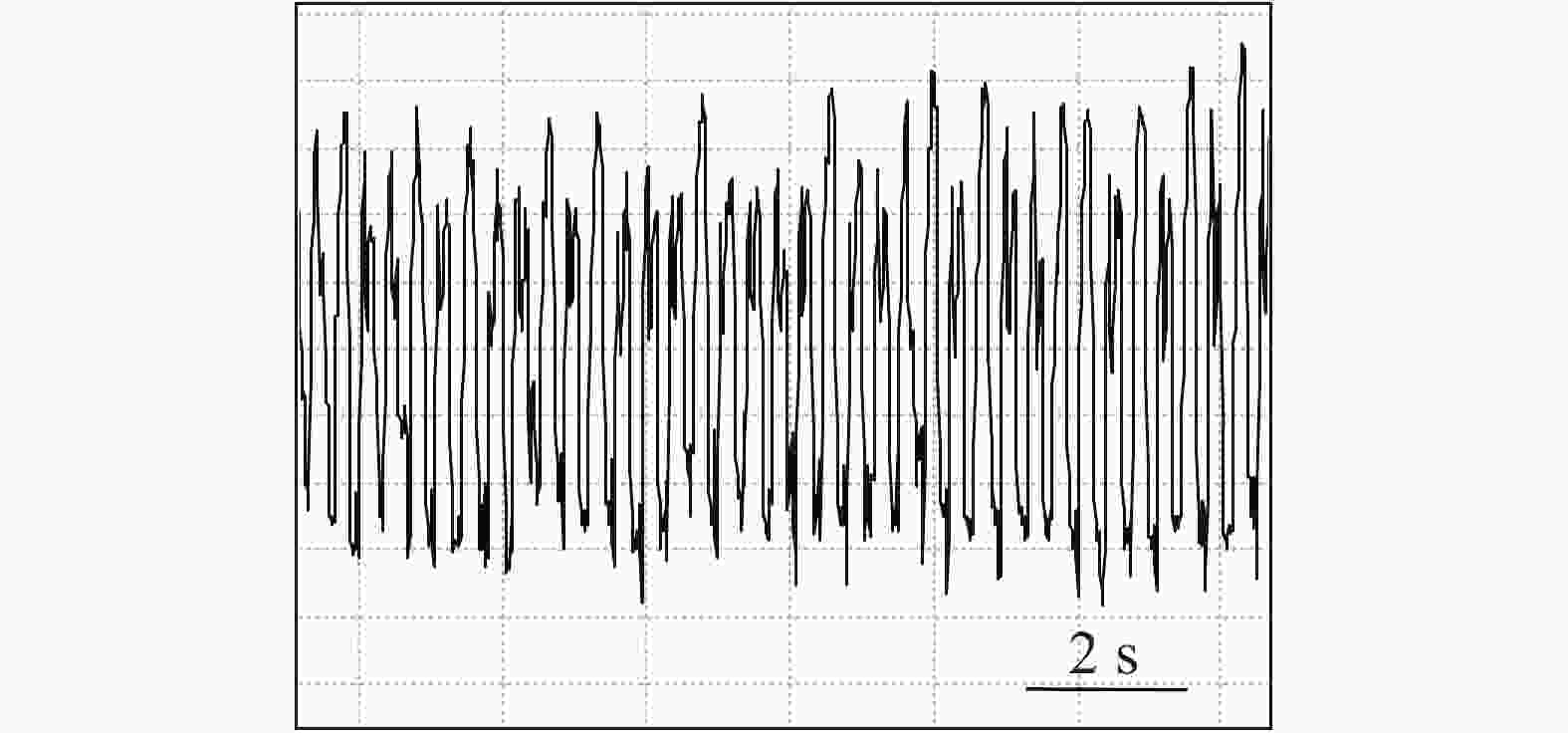
图 4 石蒜绵粉蚧取食行为EPG图谱(G波)
Figure 4. Electropenetrographs (EPG) of Phenacoccus solani (waveform G)
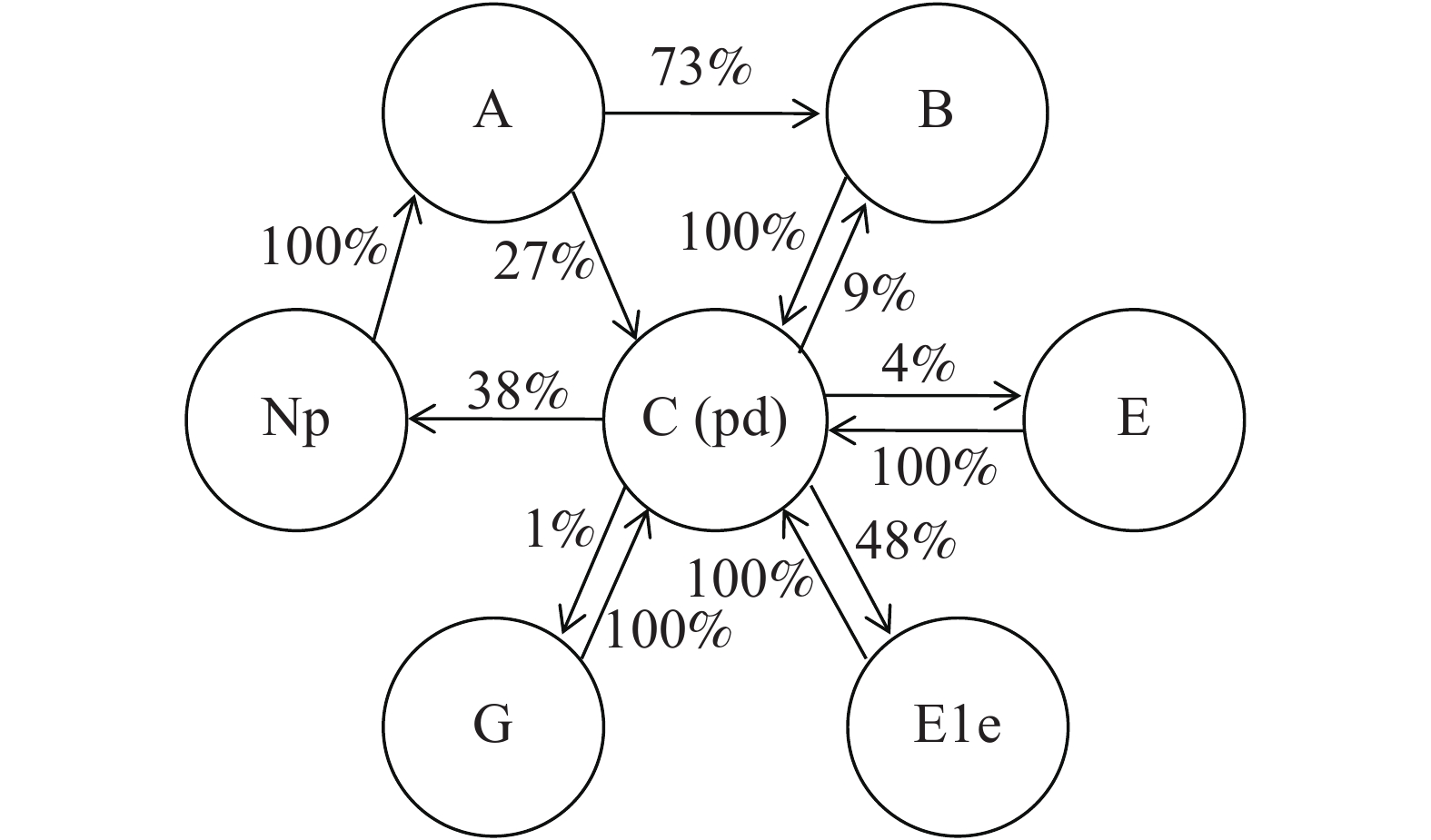
将EPG监测的所有结果进行统计,各波形发生的关系及概率如图5所示。73%的probe发生过程中会有B波的发生,除了紧随A波发生的情况外,还在路径波(C波)发生的过程中有9%的出现概率。除A波外,所有的波形发生后均回复到C波后再进行其他波形的转换。跟随C波发生的主要波形是E1e波形和np波形,发生概率分别为48%和38%,此外还有较小的概率发生E波和G波。
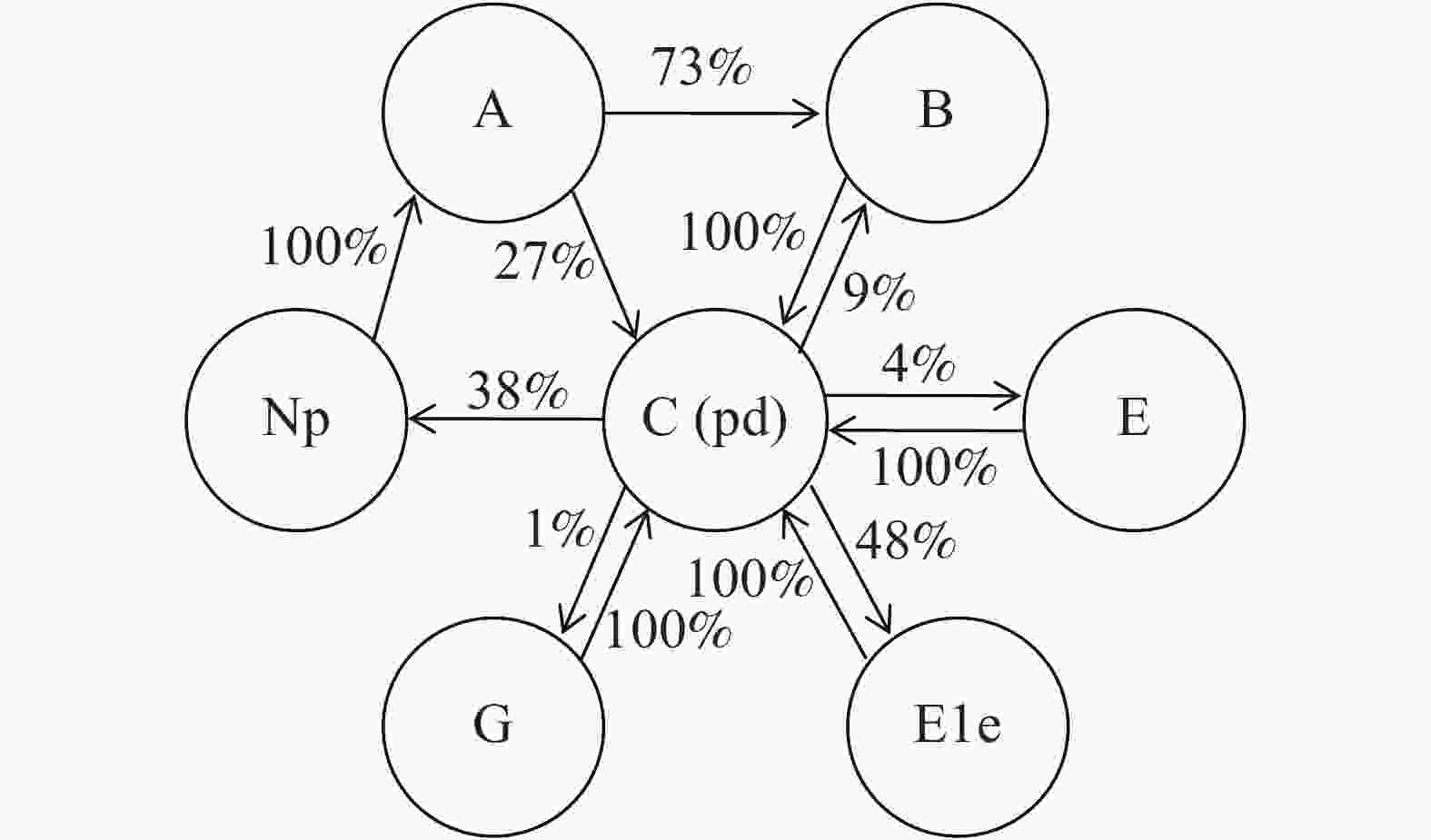
图 5 石蒜绵粉蚧取食行为EPG各波形发生关系。
Figure 5. Correlations of electropenetrograph (EPG) waveforms of Phenacoccus solani
-
石蒜绵粉蚧在番茄和胧月上的取食波形在类别和发生情况上存在一定程度的差异。
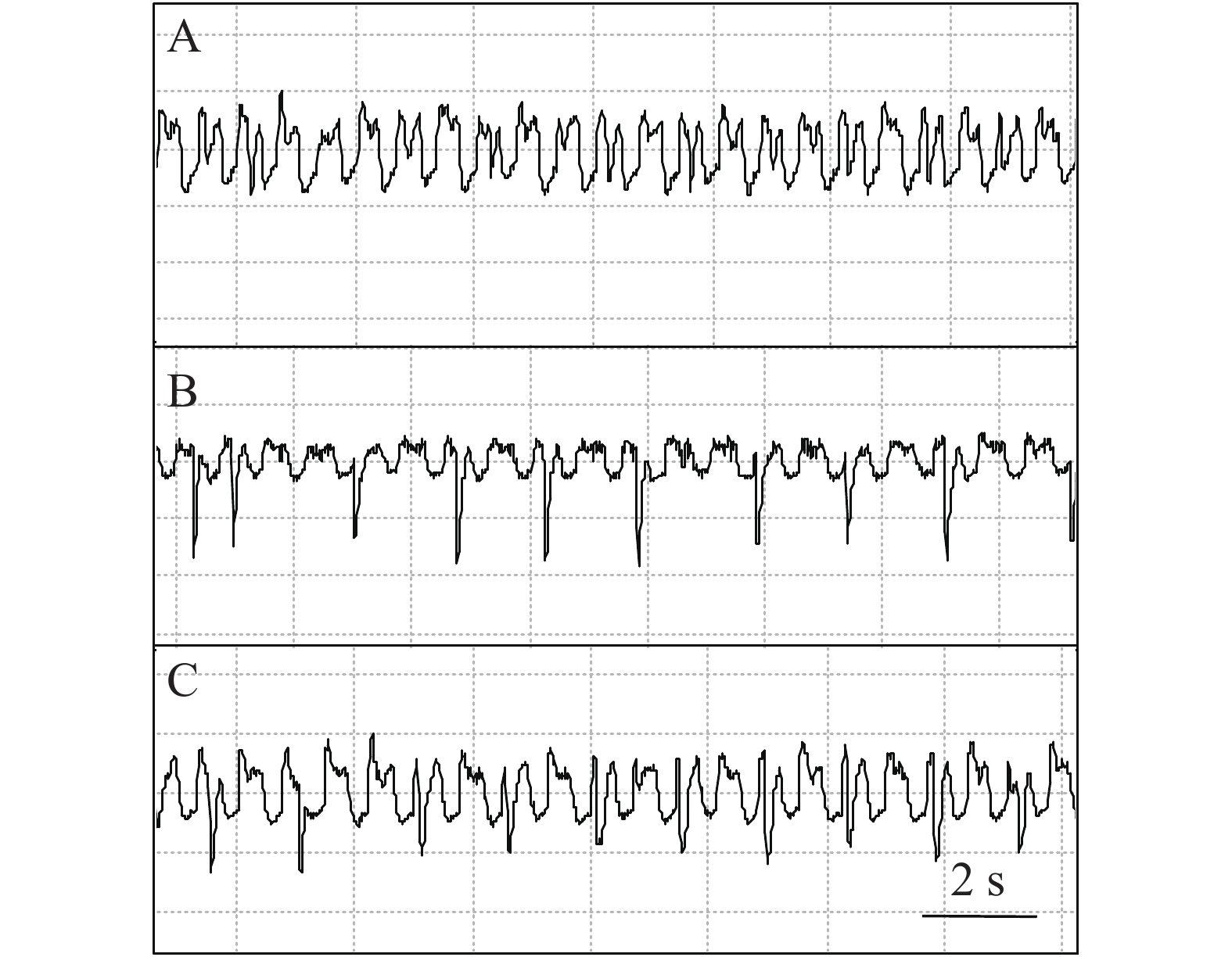
在波形种类上,石蒜绵粉蚧取食番茄时可发生E波,但是在已观察的所有取食波形(> 100 probe)中,发生E波的次数很少,仅存在数次。石蒜绵粉蚧取食胧月时从未发生E波,但是出现大量E1e波形,且在发生过程中出现水溶性唾液分泌波“E1”波相似的亚波型(图6-A)和主动取食波形“E2”相似的亚波型(图6-B)以及两种波形的混合波形(图6-C)。在已定义的粉蚧和蚜虫的EPG图谱中,韧皮部相关的水溶性唾液分泌波和主动取食波形的发生情况均与消化效率相关;同时,考虑到EPG图谱定义的统一性,因此,对于胞外水平的E1和E2暂不加以区分而都归并于E1e波。
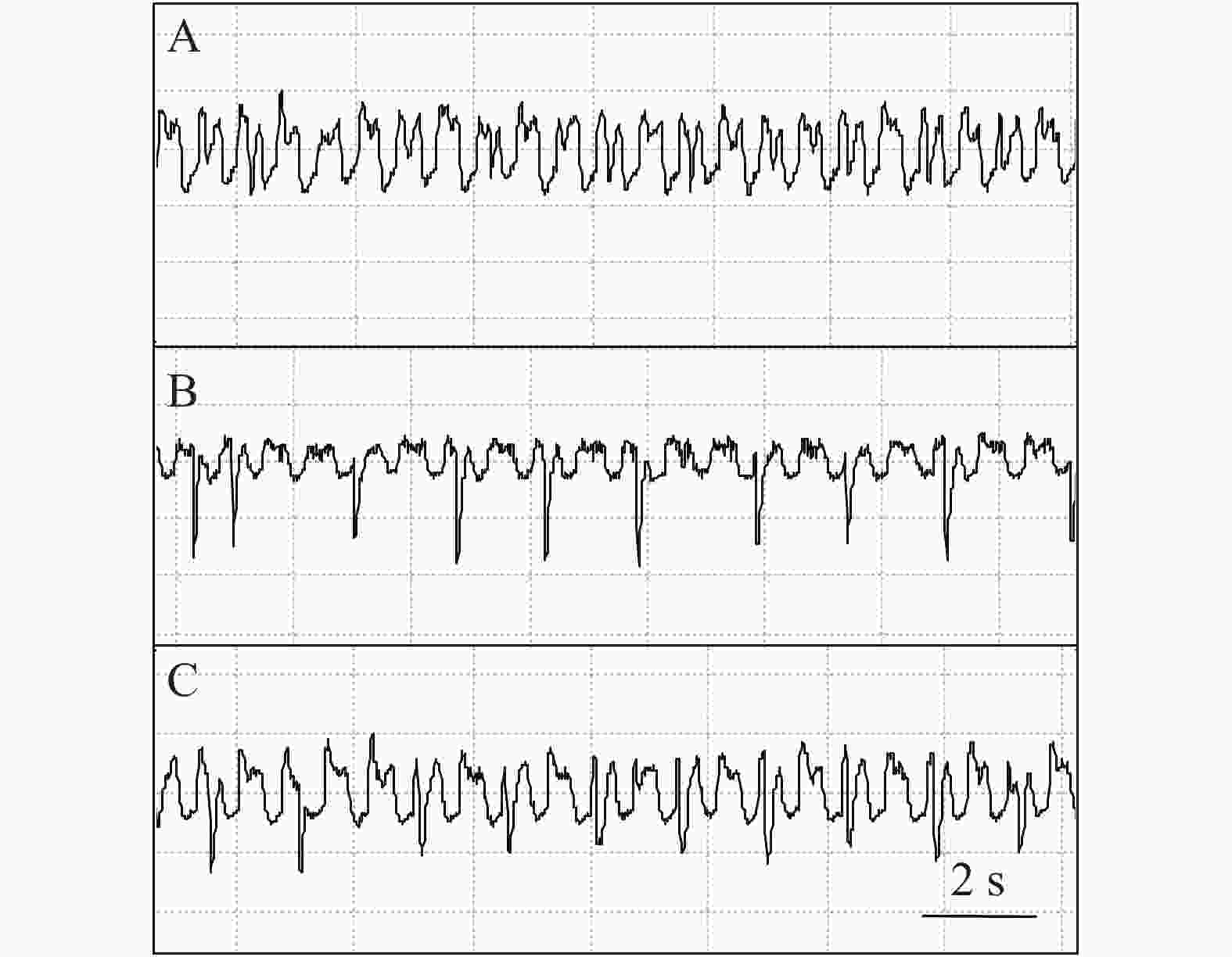
图 6 石蒜绵粉蚧在胧月上的取食行为EPG图谱(E1e波)
Figure 6. Electropenetrographs (EPG) of Phenacoccus solani on Graptopetalum paraguayense (waveform E1e)
在波形发生的相关参数上,石蒜绵粉蚧在胧月的取食行为对比于其在番茄上的取食行为,具体如表1所示。在监测时间范围内的刺探总体情况而言,前者的刺探数显著少于后者,表明石蒜绵粉蚧在胧月上取食位点更加固定,相对而言,其在番茄上时会较为频繁地变换取食位点;同时,前者的刺探总时长显著长于后者的刺探总时长。虽然两者的刺探总时长存在显著差异,但是路径波总时长却非常接近,主要的差异表现在持续性取食波的总时长,从数据上看,前者几乎是后者的4倍。前者首个刺探发生的时间要显著快于后者,且首个刺探波的时长是后者的3倍有余。单个刺探内口针穿透叶表皮的时间,前者显著短于后者。首个pd波发生的时间,两者间并不存在显著差异。然而,前者pd波的次数显著多于后者。石蒜绵粉蚧在两种植物上的持续性取食波都以E1e波为主,首个E1e波发生之前所需的刺探数,前者显著少于后者。
表 1 石蒜绵粉蚧在不同寄主植物上的取食行为
Table 1. Feeding behaviors of Phenacoccus solani on different plants
参数 Variables/time 生物学意义 Biological implications 植物 Plants 胧月 G. paraguayense 番茄 L. esculentum n_Pr 刺探的个数 1.05 ± 0.15 3.33 ± 0.97* s_Pr/hr 刺探时间总和 7.98 ± 0.41 6.03 ± 0.95* s_C/hr 口针在细胞间的时间总和 4.98 ± 0.92 5.02 ± 0.56 s_E1e/hr 持续取食时间 3.03 ± 0.24 0.71 ± 0.19* t_1Pr/hr 昆虫从接触植物到第一次口针插入叶片表面的时间 15.65 ± 5.97 68.11±36.25* ec_1Pr/hr 第一个Prb的总时长 3.33 ± 0.97 1.05 ± 0.15* s_A+B/sec 单个刺探内口针穿透叶表皮的时间 16.42 ± 5.32 29.67 ± 5.12* ec_1pd/sec 昆虫从接触植物到第一次口针插入叶肉细胞的时间 163.54 ± 15.06 147.04±18.40 n_pd 叶肉细胞内的尝试性取食次数 115.78 ± 10.56 57.57±12.33* n_Pr1E1e 开始第一次取食之前的刺探的个数 0.69 ± 0.06 1.09 ± 0.12* 注:*表示同行不同列数据间存在显著差异(P < 0.05)。
Note: * means there is a significant difference (P < 0.05) between data in the same row. -
参考扶桑绵粉蚧(Phenaoccus solenopsis) [15]、木薯绵粉蚧(Phenacoccus manihoti) [20]、橘臀纹粉蚧(Planococcus citri)[22]、暗色粉蚧(Pseudococcus viburni) [23]等取食行为的EPG图谱,石蒜绵粉蚧取食行为的刺探电位图谱也包含7种基本波形,即A波、B波、C波、pd波、E1e波、E波以及G波,以上波形分别对应口针穿透叶表面时产生的波形、唾液鞘形成时产生的波形、口针游走于叶肉细胞间是产生的波形、在叶肉细胞内尝试性取食时产生的波形、胞外取食波形、持续性取食波形和木质部取食波形。
扶桑绵粉蚧EPG相关研究[15]表明,不同取食位点叶表皮层的消化难易度不同,导致B波变化较大,有时在某些刺探内几乎没有显著的体现。在粉蚧和蚜虫单次刺探取食行为的EPG信号中,B波紧随于A波,仅出现一段,且发生时间通常少于20 s。与前期研究不同的是,石蒜绵粉蚧取食时,B波可在单个刺探内多次发生。因B波为唾液鞘形成时产生的波形,推测可能与其取食习惯相关,但仍有待证实。口针游走于叶肉细胞间时,会发生对周围细胞进行尝试性的穿刺并伴随虫体主动吸食的过程,因此pd波是相对于C波的负压波。有研究认为pd波是刺吸式口器传毒媒介昆虫获毒传毒的主要行为表现[24-25]。虽然目前尚无研究发现石蒜绵粉蚧可传带某种病毒,但该虫的取食特性使其具备获毒传毒的生理学基础,因此,在制定针对石蒜绵粉蚧的综合防治措施时还应考虑其寄主植物上可能携带病毒的发生情况。根据主要摄食部位的不同,刺吸式口器昆虫通常分为木质部取食型昆虫,如叶蝉等,韧皮部取食型昆虫,如蚜虫、烟粉虱等,以及叶肉汁液取食型昆虫,如棉盲蝽等。目前关于粉蚧取食行为的EPG研究中,所有的结果都显示粉蚧为典型的韧皮部取食性昆虫,如扶桑绵粉蚧[15]、桔臀纹粉蚧[22]、木薯绵粉蚧[26]等,且持续取食时间通常可长达数小时。本文研究结果显示石蒜绵粉蚧虽也可发生韧皮部取食,但受试个体中出现的韧皮部取食的次数非常少,虽然持续时间亦可长达数小时以上,但波形上并未呈现典型的被动取食波形(E2波)。在长尾粉蚧Pseudococcus longispinus[27]的相关取食波研究中也发现了这种类似的情况,即没有典型E波亚波型。虽然报道中也对相关波形进行了E1和E2的标记,但其图例结果中并没有呈现类似于其他粉蚧E1及E2波形的典型特征。同时,研究者在讨论中提及,在长尾粉蚧的EPG记录中同样出现了E1e波形,该波形被整合入C波进行统计分析。对比于前期针对扶桑绵粉蚧的研究,其原因可能是E1e出现的频率较低且时长较短。然而迄今为止,E1e波形的生物学意义仍未确定[15],在刺吸式口器昆虫EPG波型相关的所有研究中也未见针对E1e波形进行详细的统计与分析。对比于其他粉蚧与韧皮部取食昆虫产生的E波,如扶桑绵粉蚧[15]、木薯绵粉蚧[26]、桔臀纹粉蚧和暗色粉蚧[23]等,石蒜绵粉蚧在本研究供试植物上的持续取食波更多的是以胞外形式出现,即E1e波的形式为主,在EPG信号上表现为零压强波,而典型的E波则是强烈的负压强波。除电动势外,不管发生的频率、持续的时长以及包含的波形,石蒜绵粉蚧的E1e波与扶桑绵粉蚧E波(包括E1和E2)的EPG性质非常相似。推测可能与石蒜绵粉蚧喜好取食多肉植物有关[11],而石蒜绵粉蚧在胧月和番茄上取食行为的不同则说明了石蒜绵粉蚧在多肉植物上具有更强的适应性和为害风险。本实验结果证明,相较于番茄,石蒜绵粉蚧在胧月上取食时,取食位点转换频率低、穿透叶表速度快、获取叶肉营养多、持续性取食时间长,即石蒜绵粉蚧更加适应寄主植物胧月。有研究者[28]认为,多肉植物的叶片结构具有其特殊性,主要体现在叶片厚度、角质层的厚薄有无、气孔的分布密度以及叶肉细胞的分化种类和输导组织的发达程度等方面。据此推测,石蒜绵粉蚧取食多肉植物时发生的E1e波,包括期间出现的类似E1和E2的亚波型,是其主要的取食波形;而电动势方面的差异,推测主要和植物结构本身有关,但仍需做进一步深入研究。
本实验结果说明,石蒜绵粉蚧取食行为的EPG波形与韧皮部取食型昆虫类似,但其主要取食波形为E1e波,证实了石蒜绵粉蚧在多肉植物胧月上具有更强的适应性和为害风险。本研究结果也为进一步研究石蒜绵粉蚧的致害性和寄主偏好性的行为学机制提供了基础数据和技术参考。
Characterization of Feeding Behaviors of Phenacoccus solani on Two Plants Based on EPGs
-
摘要: 利用刺探电位图谱(Electronic penetration graph, EPG)技术研究了石蒜绵粉蚧(Phenacoccus solani)在胧月(Graptopetalum paraguayense)和番茄(Lycopersicon esculentum)上的取食行为。结果表明,石蒜绵粉蚧在这2种植物上取食时,可产生A波、B波、C波、pd波、E波、E1e波以及G波等基本波型,其中,E1e波为主要的持续性取食波形;取食番茄时,E波偶发且该波内未出现典型的被动取食波(E2波)。石蒜绵粉蚧在2种植物上的取食行为差异显著,相较于后者,石蒜绵粉蚧在胧月上取食时,位点转换频率更低、穿透叶表速度更快、获取叶肉营养更多、持续性取食时间更长,且期间未出现E波而长时间发生E1e波,E1e波内出现典型的E1波和E2波(但电动势水平为零)的亚波型。本研究结果说明了石蒜绵粉蚧在2种植物上取食时产生的EPG波形与韧皮部取食型昆虫类似,但其主要取食波形为E1e波,表明石蒜绵粉蚧在多肉植物胧月上具有更强的适应性和为害风险。Abstract: Feeding behaviors of Phenacoccus solani on Graptopetalum paraguayense and Lycopersicon esculentum were characterized by using electropenetrationgraphy (EPG), respectively. The results showed that the basic waveforms including waveforms A, B, C, pd, E, E1e and F were generated when P. solani was feeding on the two plants. Waveform E1e was the main persistent feeding waveform, while waveform E occurred occasionally without typical passive feeding waveform (waveform E2) when P. solani was feeding on L. esculentum. Feeding behaviors of P. solani on these two plants were significantly different. Compared to the performance on L. esculentum, P. solani feeding on G. paraguayense showed lower site transition frequency, faster penetration of the leaf surface, more access to foliar nutrients, and longer duration of persistent feeding, and generated no waveform E but waveform E1e with prolonged occurrence. There were typical subwaveforms of the waveforms E1 and E2 (but with the zero level of electric potential) within the waveform Ele. These results indicated that the EPG waveforms of feeding behaviors of P. solani on these two plants were similar to those of the phloem-feeding insects, but the waveform Ele was the main feeding waveform, which confirms that P. solani have greater adaptability and infestation risk on the succulent plant G. paraguayense.
-
图 2 石蒜绵粉蚧取食行为EPG图谱(E波)
Fig. 2 Electropenetrographs (EPG) of Phenacoccus solani (waveform E).
图 3 石蒜绵粉蚧取食行为EPG图谱(E1e波)
Fig. 3 Electropenetrographs (EPG) of Phenacoccus solani (waveform E1e).
图 4 石蒜绵粉蚧取食行为EPG图谱(G波)
Fig. 4 Electropenetrographs (EPG) of Phenacoccus solani (waveform G)
图 5 石蒜绵粉蚧取食行为EPG各波形发生关系。
Fig. 5 Correlations of electropenetrograph (EPG) waveforms of Phenacoccus solani
图 6 石蒜绵粉蚧在胧月上的取食行为EPG图谱(E1e波)
A:水溶性唾液分泌波;B:主动取食波;C:以上两种波形的叠加。
Fig. 6 Electropenetrographs (EPG) of Phenacoccus solani on Graptopetalum paraguayense (waveform E1e)
A: Waveform associated with salivation; B: Waveform associated with ingestion; C: Superposition of the two waveforms above.
表 1 石蒜绵粉蚧在不同寄主植物上的取食行为
Table 1 Feeding behaviors of Phenacoccus solani on different plants
参数 Variables/time 生物学意义 Biological implications 植物 Plants 胧月 G. paraguayense 番茄 L. esculentum n_Pr 刺探的个数 1.05 ± 0.15 3.33 ± 0.97* s_Pr/hr 刺探时间总和 7.98 ± 0.41 6.03 ± 0.95* s_C/hr 口针在细胞间的时间总和 4.98 ± 0.92 5.02 ± 0.56 s_E1e/hr 持续取食时间 3.03 ± 0.24 0.71 ± 0.19* t_1Pr/hr 昆虫从接触植物到第一次口针插入叶片表面的时间 15.65 ± 5.97 68.11±36.25* ec_1Pr/hr 第一个Prb的总时长 3.33 ± 0.97 1.05 ± 0.15* s_A+B/sec 单个刺探内口针穿透叶表皮的时间 16.42 ± 5.32 29.67 ± 5.12* ec_1pd/sec 昆虫从接触植物到第一次口针插入叶肉细胞的时间 163.54 ± 15.06 147.04±18.40 n_pd 叶肉细胞内的尝试性取食次数 115.78 ± 10.56 57.57±12.33* n_Pr1E1e 开始第一次取食之前的刺探的个数 0.69 ± 0.06 1.09 ± 0.12* 注:*表示同行不同列数据间存在显著差异(P < 0.05)。
Note: * means there is a significant difference (P < 0.05) between data in the same row. 下载: 导出CSV
下载: 导出CSV
-
[1] 王珊珊, 武三安. 中国大陆新纪录种: 石蒜绵粉蚧(Phenacoccus solani Ferris)[J]. 植物检疫, 2009, 23(4): 35 − 37. doi: 10.3969/j.issn.1005-2755.2009.04.012 [2] CHEN S P, CHEN C N, WONG C Y. New record of a pest - Phenacoccus solani Ferris (Homoptera: Pseudococcidae) in Taiwan [J]. Journal of Agricultural Research of China, 2002, 51(2): 79 − 82. [3] MOGHADDAM M, HATAMI B, ZIBAII K, et al. Report of Phenacoccus solani (Hom. : Coccoidea: Pseudococcidae) from Iran [J]. Journal of Entomological Society of Iran, 2004(1): 135 − 136. [4] BEN-DOV Y. Note: The solanum mealybug, Phenacoccus solani ferris (Hemiptera: Coccoidea: Pseudococcidae), extends its distribution range in the Mediteranean basin [J]. Phytoparasitica, 2005, 33(1): 15 − 16. doi: 10.1007/BF02980920 [5] ALAM S, KHAM, MURUGAN, et al. Electrical penetration graph recording of Russian wheat aphid (Hemiptera: Aphididae) feeding on aphid-resistant wheat and barley [J]. Journal of Economic Entomology, 2015, 108(5): 2465 − 2470. doi: 10.1093/jee/tov183 [6] 万志伟. 杭州邮路口岸首次截获石蒜绵粉蚧[J]. 中国检验检疫, 2019(4): 52 − 52. [7] 郑斯竹, 高渊, 樊新华. 石蒜绵粉蚧传入我国风险分析[J]. 中国植保导刊, 2015, 35(4): 75 − 77. doi: 10.3969/j.issn.1672-6820.2015.04.018 [8] 黄芳, 张文俊, 张建成, 等. 基于BIOCLIM模型的石蒜绵粉蚧在中国的适生区分析[J]. 浙江农业学报, 2019, 31(8): 1331 − 1336. doi: 10.3969/j.issn.1004-1524.2019.08.14 [9] 智伏英, 黄芳, 黄俊, 等. 石蒜绵粉蚧生物学特性[J]. 昆虫学报, 2018, 61(7): 871 − 876. [10] 李思怡. 石蒜绵粉蚧的生物学和生态学特性研究[D]. 杭州: 浙江农林大学, 2019. [11] 潘颖佳, 崔旭红, 邵崟, 等. 石蒜绵粉蚧在七种多肉植物上的生物学特性[J]. 昆虫学报, 2020, 63(10): 1223 − 1233. [12] 黄鹏, 陈汉鑫, 姚锦爱, 等. 金龟子绿僵菌对石蒜绵粉蚧的室内毒力与防治效果[J]. 中国生物防治学报, 2019, 35(6): 884 − 890. [13] TJALLINGII W F. Electrical recording of stylet penetration activities. In: Minks AK, Harrewijn, ed. Aphids, their biology, natural enemies and control[M]. The Netherlands: Elsevier, Amsterdam, 1988: 95 − 108. [14] CIVOLANI S, LEIS M, GRANDI G, et al. Stylet penetration of Cacopsylla pyri; an electrical penetration graph (EPG) study [J]. Journal of Insect Physiology, 2011, 57(10): 1407 − 1419. doi: 10.1016/j.jinsphys.2011.07.008 [15] HUANG F, TJALLINGII W F, ZHANG P J, et al. EPG waveform characteristics of solenopsis mealybug stylet penetration on cotton [J]. Entomologia Experimentalis et Applicata, 2012, 143(1): 47 − 54. doi: 10.1111/j.1570-7458.2012.01233.x [16] 黄芳, 吕要斌, 吴志毅, 等. 扶桑绵粉蚧对马铃薯幼苗的致害性研究[J]. 环境昆虫学报, 2017, 39(4): 888 − 892. [17] HUANG F, WANG F F, ZHANG Z J, et al. EPG analysis of feeding behavior of the solenopsis mealybug, Phenacoccus solenopsis (Hemiptera: Pseudococcidae) before and after host shift [J]. Acta Entomologica Sinica, 2014, 57(4): 503 − 508. [18] HUANG F, ZHANG Z J, LI W D, et al. Host plant probing analysis reveals quick settlement of the solenopsis mealybug during host shift [J]. Journal of Economic Entomology, 2014, 107(4): 1419 − 1425. doi: 10.1603/EC13510 [19] FARMER A, MCGUFFIN P. Analysis of circulative transmission by electrical penetration graphs [J]. Virus-Insect-Plant Interactions, 2001, 178(3): 69 − 85. [20] CALATAYUD P A, RAHBE Y, TJALLINGII W F, et al. Electrically recorded feeding behaviour of cassava mealybug on host and non-host plants [J]. Entomologia Experimentalis Et Applicata, 1994, 72: 219 − 232. doi: 10.1111/j.1570-7458.1994.tb01821.x [21] SARRIA E, CID M, GARRO E, et al. Excel Workbook for automatic parameter calculation of EPG data [J]. Computers & Electronics in Agriculture, 2009, 67(1/2): 35 − 42. [22] CID M, FERERES A. Characterization of the probing and feeding behavior of Planococcus citri (Hemiptera: Pseudococcidae) on grapevine [J]. Annals of the Entomological Society of America, 2010, 103: 404 − 417. doi: 10.1603/AN09079 [23] OBOK E, WETTEN A, ALLAINGUILLAUME J. Electropenetrography application and molecular-based virus detection in mealybug (Hemiptera: Pseudococcidae) vectors of cacao swollen shoot virus on Theobroma cacao L[J]. Annals of Agricultural Sciences, 2018: S0570178318300095. [24] LETT J M, GRANIER M, GRONDIN M, et al. Electrical penetration graphs from Cicadulina mbila on maize, the fine structure of its stylet pathways and consequences for virus transmission efficiency [J]. Entomologia Experimentalis Et Applicata, 2010, 101(2): 93 − 109. [25] STAFFORD C A, WALKER G P, CREAMER R. Stylet penetration behavior resulting in inoculation of beet severe curly top virus by beet leafhopper, Circulifer tenellus [J]. Entomologia Experimentalis Et Applicata, 2010, 130(2): 130 − 137. [26] RENARD S, CALATAYUD P A, PIERRE J S, et al. Recognition behavior of the cassava mealybug Phenacoccus manihoti Matile-Ferrero (Homoptera: Pseudococcidae) at the leaf surface of different host plants [J]. Journal of Insect Behavior, 1998, 11(3): 429 − 450. doi: 10.1023/A:1020911016042 [27] SANDANAYAKA W R M, BLOUIN A G, PRADO E, et al. Stylet penetration behaviour of Pseudococcus longispinus in relation to acquisition of grapevine leafroll virus 3 [J]. Arthropod-Plant Interactions, 2013(7): 137 − 146. [28] 张力凡, 孙凉, 李茜, 等. 多肉植物叶片结构与生境条件研究[J]. 南方农机, 2017, 48(22): 11 − 12. doi: 10.3969/j.issn.1672-3872.2017.22.006 -

 点击查看大图
点击查看大图
计量
- 文章访问数: 598
- HTML全文浏览量: 223
- PDF下载量: 84
- 被引次数: 0
