



-
水稻细菌性条斑病是水稻粮食生产中的重要病害,由稻黄单胞菌稻生致病变种(Xanthomonas oryzae pv. oryzicola,Xoc)引起,主要发病区域为热带和亚热带[1]。Xoc通过气孔进入叶片,先进入气孔下腔,后在薄壁组织的细胞间隙中定殖,使叶片表面产生条斑病症,造成水稻减产[2]。在定植过程中,黄单胞菌产生的胞外酶、胞外多糖以及其他效应因子都起到重要作用[3]。如通过分泌蛋白酶、淀粉酶和纤维素酶来降解植物相应的细胞壁成分,使细菌成功入侵植物组织[4]。
乙酰化修饰是细胞内蛋白的主要修饰方式之一,乙酰基团通常以乙酰辅酶A(AcCoA)或乙酰磷酸酯(AcP)的形式存在[5]。细菌蛋白乙酰化具有多种功能,如调节DNA复制、转录和修复等DNA过程和通过修饰组蛋白和转录因子调节基因表达等,对于细菌致病力、抗逆性和生长都具有重要作用[6]。
细菌蛋白乙酰化修饰主要为赖氨酸乙酰化,而GNAT(GCN5 related N-acetyltransferases)是最大的赖氨酸乙酰转移酶家族。GNAT结构高度保守,根据其结构特征可以分为3大类(ClassⅠ-Ⅲ)5种小类(Type Ⅰ-Ⅴ)[7]。Xoc编码1个Type Ⅰ和5个Type Ⅳ 类GNAT乙酰转移酶。Type Ⅰ类是包含C-端 GNAT结构域的多结构域乙酰转移酶,以YfiQ及其同源物为代表。YfiQ在细菌酸、高温和活性氧胁迫中发挥作用,但其作用机制还不清楚[8]。Type Ⅳ 类为单GNAT结构域乙酰转移酶,通常长度为150~200个氨基酸。Type Ⅳ 类乙酰转移酶在蛋白质乙酰化作用上弱于Type Ⅰ类。但许多Ⅳ型GNAT具有赖氨酸乙酰化以外的活性。例如,大肠杆菌中的RimI除乙酰化赖氨酸外,也是N-α-乙酰转移酶[9–10]。黄单胞菌中乙酰转移酶的相关研究很少。白叶枯病菌乙酰转移酶Pxo_00987参与细菌鞭毛糖基化的修饰,突变导致病原菌致病性下降[11]。Xcv(X. campestris pv.vesicatoria)和Xcc(X. campestrispv. campestris)中的Ⅲ型效应因子AvrBsT(YpoJ同源蛋白)可通过 O-乙酰化方式修饰宿主蛋白,调控病原菌与植物互作[12,13]。而在Xoc中,乙酰转移酶的作用还未见报道。为分析GNAT类乙酰转移酶在Xoc中的作用,本研究首先通过生物信息学方法预测Xoc中的GNAT家族蛋白,然后通过同源重组双交换的方法创建这些基因的单突和多突突变菌株,进而比较这些突变体与野生型Xoc的生长速度和致病力,以及致病相关因子(胞外酶和运动能力)的差异。该研究将为进一步解析GNAT类乙酰转移酶的功能及调控通路奠定了基础,对水稻细菌性条斑病的防治有一定的促进作用。
-
Xoc模式菌株RS105和载体pK18mobsacB[14]为笔者所在实验室保存,大肠杆菌感受态购自基睿生物科技有限公司。
-
蔗糖蛋白胨培养基(Peptone Sucrose Agar,PSA):1%胰蛋白胨,1%蔗糖,0.1% L-谷氨酸钠;蔗糖蛋白胨筛选培养基:1%胰蛋白胨,15%蔗糖,0.1%L-谷氨酸钠;葡萄糖蛋白胨培养基(Peptone Glucose Agar,PGA):1%胰蛋白胨,1%葡萄糖,0.1%L-谷氨酸钠;卢里亚-贝尔塔尼培养基(Luria-Bertani,LB):1%胰蛋白胨,1%NaCl,0.5%酵母提取物;改良M4培养基(modified M4 medium,M4M):0.048% Na2HPO4,0.03% KH2PO4,0.05%柠檬酸钠,0.1%(NH4)2SO4,0.02% MgSO4·7H2O,0.05%酶水解酪素,0.05%葡萄糖;相应固体培养基另加1.5%琼脂。通用型DNA回收试剂盒和质粒小提试剂盒购自天根生化科技有限公司,高保真DNA聚合酶购自诺唯赞生物科技有限公司,限制性核酸内切酶及T4连接酶购自NEB公司,其他试剂购自Sigma-Aldrich公司或索莱宝生物科技有限公司。
-
根据被敲除基因两端序列,使用SnapGene软件进行序列分析和引物设计,由生工生物公司(广州)分公司合成引物(表1)。
表 1 引物序列
名称 序列 xocmindFF GACGAAGCTTCAGCCAGAAGTAGCTGGCAC xocmindFR ACACGGATCCATCGAGCTCGTGCTCGCGCAC xoc1292ddRF ACACGGATCCGTGTCGGTCGCGCGTTCTATG xoc1292ddRR GACGTCTAGACACGTCCGGCACCTGCGACAG xoc2699ddFF ACTGAAGCTTGATCAATACCTCGGTCGGTC xoc2699ddFR CTGCTCTAGACATGTGGTCAGTCGATGTAG xoc2699ddRF CTGCTCTAGACCTCCTTCTGATGAAAACCC xoc2699ddRR CTACGAATTCCATCTTGCCGAAGAATCCCA xoc0450ddFF ACTGAAGCTTCGGTGGTCCCGTATCCGAAG xoc0450ddFR CTGCTCTAGAGTGGTACGTACTCATCGGAC xoc0450ddRF CTGCTCTAGAGGACGATGCGACCATGATGG xoc0450ddRR CTACGAATTCGACGGATAGTTGTCGGTCAG xoc3825ddFF ACTGAAGCTTCAGAGCTGCCATGAGCGTGT xoc3825ddFR CTGCTCTAGAGGCACGTTCAATGCGCTCAC xoc3825ddRF CTGCTCTAGACGTTGGTGATGGCGATCGAG xoc3825ddRR CTACGAATTCCGTTAGTCGTTGCAACCCGT xoc1598ddFF ACTGAAGCTTCTCAAGAAGCATCTCGGCGA xoc1598ddFR CTGCTCTAGATGGCCATGACCGGTGTTCCT xoc1598ddRF CTGCTCTAGATGGAGCTGGATGCCTGAGGT xoc1598ddRR CTACGAATTCGAAGGTAGGCGACGGCATCA xoc4123ddFF ACTGAAGCTTGGTAGGCGTCACGAACAGAT xoc4123ddFR CTGCTCTAGAGTGCATTGGGTCCGAGAAAC xoc4123ddRF CTGCTCTAGAGACTAGGTGCGGTGACCAAG xoc4123ddRR CTACGAATTCCCTTGACCATGACCTTGGTC 注:下划线表示酶切位点。 -
通过KEGG获取乙酰转移酶的基因序列与氨基酸序列,利用MEGA进行聚类分析[15]。通过NCBI CDD进行蛋白结构域预测,并用TBtools作图[16]。
-
以Xoc模式菌株RS105的基因组为模板,PCR扩增目的基因上下游约500 bp的片段,酶切回收后连接在pK18mobsacB载体上,转化大肠杆菌感受态,经PCR鉴定及测序确定后获得敲除载体pK18-xoc_1292、pK18-xoc_2699、pK18-xoc_0450、pK18-xoc_3825、pK18-xoc_1598、pK18-xoc_4123,通过三亲接合法将构好的pK18mobsacB自杀质粒转入RS105菌株中,经过同源交换后获得突变体菌株。
-
将RS105和突变体菌株于28 ℃培养24 h,离心收集菌体,ddH2O清洗2遍,调OD600=1.0,稀释至10−6,共6个梯度,取1 μL菌悬液分别在PSA和M4M固体培养基上点板,28 ℃倒置培养,2~3 d观察拍照。
-
采用打孔法接种‘TP309’水稻[17]。待接种菌株培养至对数生长期,调OD600=1.0,用直径3 mm打孔器蘸取菌液,于叶片距叶尖约5 cm叶脉处打孔接种,分别接种野生型与突变体。21 d后量取病斑长度,记录并拍照,分析实验结果。
-
将待测菌株于28 ℃培养24 h,离心收集菌体,ddH2O清洗2次,调OD600=1.0,取2 μL菌液分别点在加入脱脂奶粉,可溶性淀粉和羧甲基纤维素的PSA固体培养基上,培养48 h。含有可溶性淀粉和羧甲基纤维素的平板分别使用 1∶100的I2-KI混合液(0.08 mol·L−1 I2,3.2 mol·L−1 KI )和0.1%刚果红染液染色。胞外酶活性可以通过比较D/d(水解产物直径D和菌落直径d)的大小来鉴定,比值越大,菌株产酶能力越强[18]。
-
(1)游动性:将待测菌株于28 ℃培养24 h,离心收集菌体,ddH2O
清洗2次,调OD600=1.0。用牙签蘸取菌液,垂直接种于游动性检测培养基NY2底部,28 ℃正置培养4 d。WT作为阳性对照。拍照记录,并测量菌落游动直径。(2)蠕动性:将待测菌株于28 ℃培养24 h,离心收集菌体,ddH2O清洗2次,调OD600=1.0。取2 μL菌液接种于蠕动性检测培养基NY1表面,28 ℃正置培养,WT作为阳性对照。拍照记录,并测量菌落蠕动直径[19]。
-
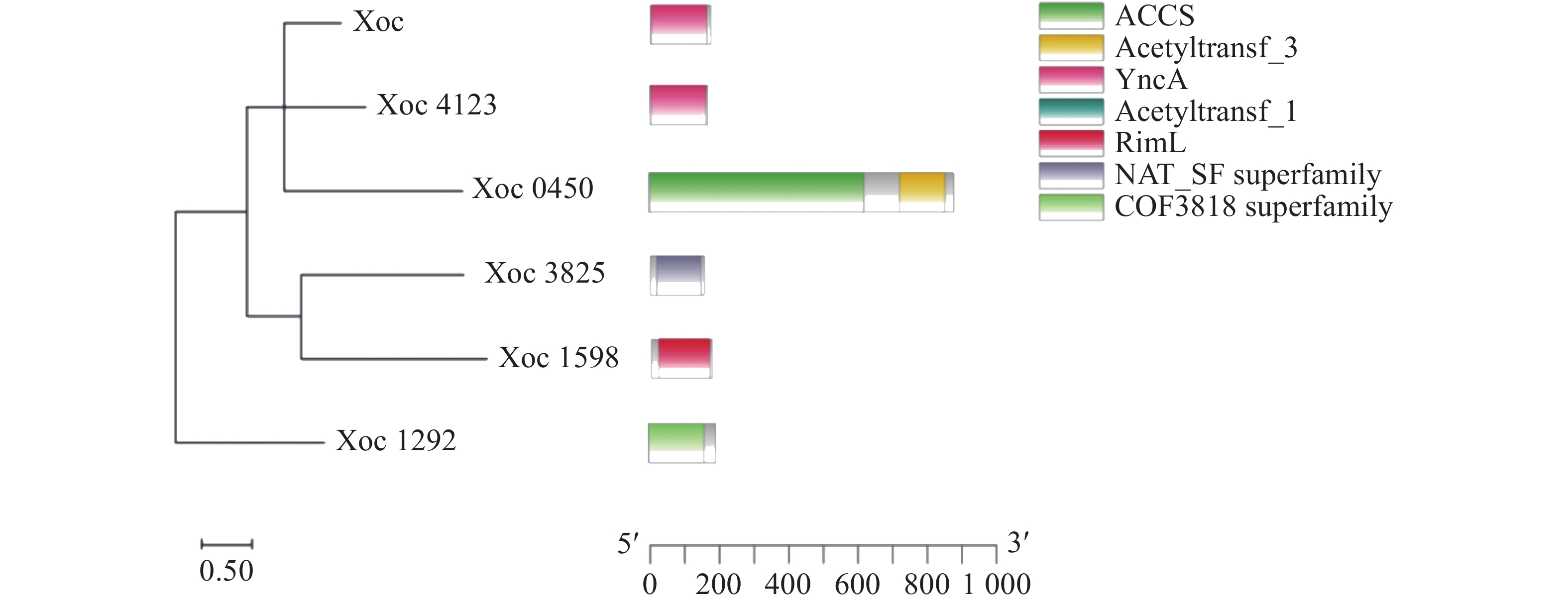
利用大肠杆菌乙酰转移酶SylD对Xoc RS105蛋白数据库进行同源搜索,获得了6个可能的GNAT乙酰转移酶(Xoc_1292、Xoc_2699、Xoc_0450、Xoc_3825、Xoc_1598和Xoc_4123)。并对这6个乙酰转移酶进行了结构域预测和聚类分析。首先通过KEGG网站获取Xoc乙酰转移酶的氨基酸序列,利用MEGA软件对其进行了聚类分析。结果发现,Xoc_2699和Xoc_4123具有较近的亲缘关系,Xoc_1292与其他5个乙酰转移酶的关系较远。利用NCBI CDD(
https://www.ncbi.nlm.nih.gov/cdd )和SMART(https://smart.embl.de )等网站进行结构域预测,并使用TBtools作图(图1)。Xoc_1292结构域为COG3818,是一种 GNAT 超家族结构域。Xoc_2699和Xoc_4123结构域为YncA结构域,一种GNAT家族L-氨基酸N-酰基转移酶,可以帮助沙门氏菌抵御甲硫氨酸磺酸肟和蛋氨酸砜的毒害作用[20]。Xoc_0450含有2个结构域,一种为酰基辅酶A合成酶结构域,另一种则为GNAT家族N-乙酰转移酶。Xoc_3825 所含结构域也是GNAT家族N-乙酰转移酶。Xoc_1598为RimJ/RimL家族GNAT乙酰转移酶。RimJ和RimL在大肠杆菌中可以乙酰化核糖体蛋白S18、S5和L12[21]。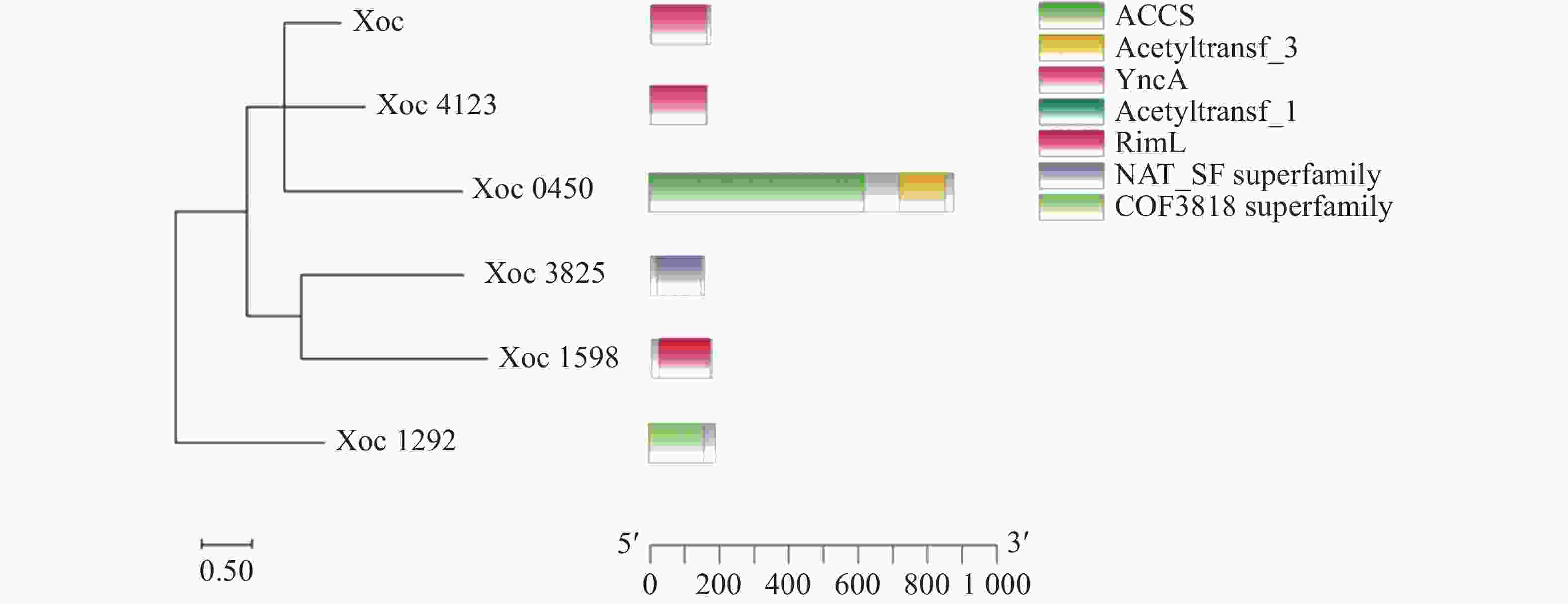
图 1 乙酰转移酶结构域与聚类分析
-
为了探究乙酰转移酶在Xoc中的功能,以RS105为野生型菌株,使用同源重组法将Xoc 6个乙酰转移酶基因敲除,得到了相应的单突突变体、然后以xoc_1292单突突变体(Δxoc_1292)为基础菌株,分别敲除其他乙酰转移酶得到5个双突突变体、2个三突突变体以及四、五和六突突变体各1个,合计16个乙酰转移酶突变体。用目的基因两侧引物进行PCR鉴定,以RS105基因组为阳性对照,ddH2O为阴性对照,xoc_1292基因长度为600 bp、xoc_2699为540 bp、xoc_0450为2 709 bp、xoc_3825为483 bp、xoc_1598为543 bp、xoc_4123为522 bp。如图2所示,对应突变体中野生型扩增片段与突变体扩增片段大小之差与被敲除基因大小相符合,证明突变体构建成功。为了叙述方便,后续将Δxoc_1292/2699/0450/3825简写为Δ4,Δxoc_1292/2699/0450/3825/4123简写为Δ5,Δxoc_1292/2699/0450/3825/4123/1598简写为Δ6。

图 2 Xoc乙酰转移酶突变体的PCR验证
-
为探究乙酰转移酶是否调控Xoc的生长,在营养丰富型培养基PSA和营养缺乏型培养基M4M上进行梯度稀释点板培养细菌。结果显示,PSA平板上突变体与野生型生长速率差异不明显(图3-A),而在M4M培养基上,Δxoc_1292、Δxoc_2699、Δxoc_0450、Δxoc_3825、Δxoc_4123、Δxoc_1292/2699、Δxoc_1292/0450、Δxoc_1292/3825、Δxoc_1292/1598、Δxoc_1292/4123、Δxoc_1292/2699/0450、Δ4、Δ5这些突变体生长速率均明显弱于野生型(图3-B)。这说明除了xoc_1598外,其余乙酰转移酶的缺失会影响Xoc的生长速率,但突变全部乙酰转移酶的六突变体生长速率却接近野生型,这可能是细菌的非酶乙酰化反应发挥了一定的作用,但具体原因还有待进一步研究。
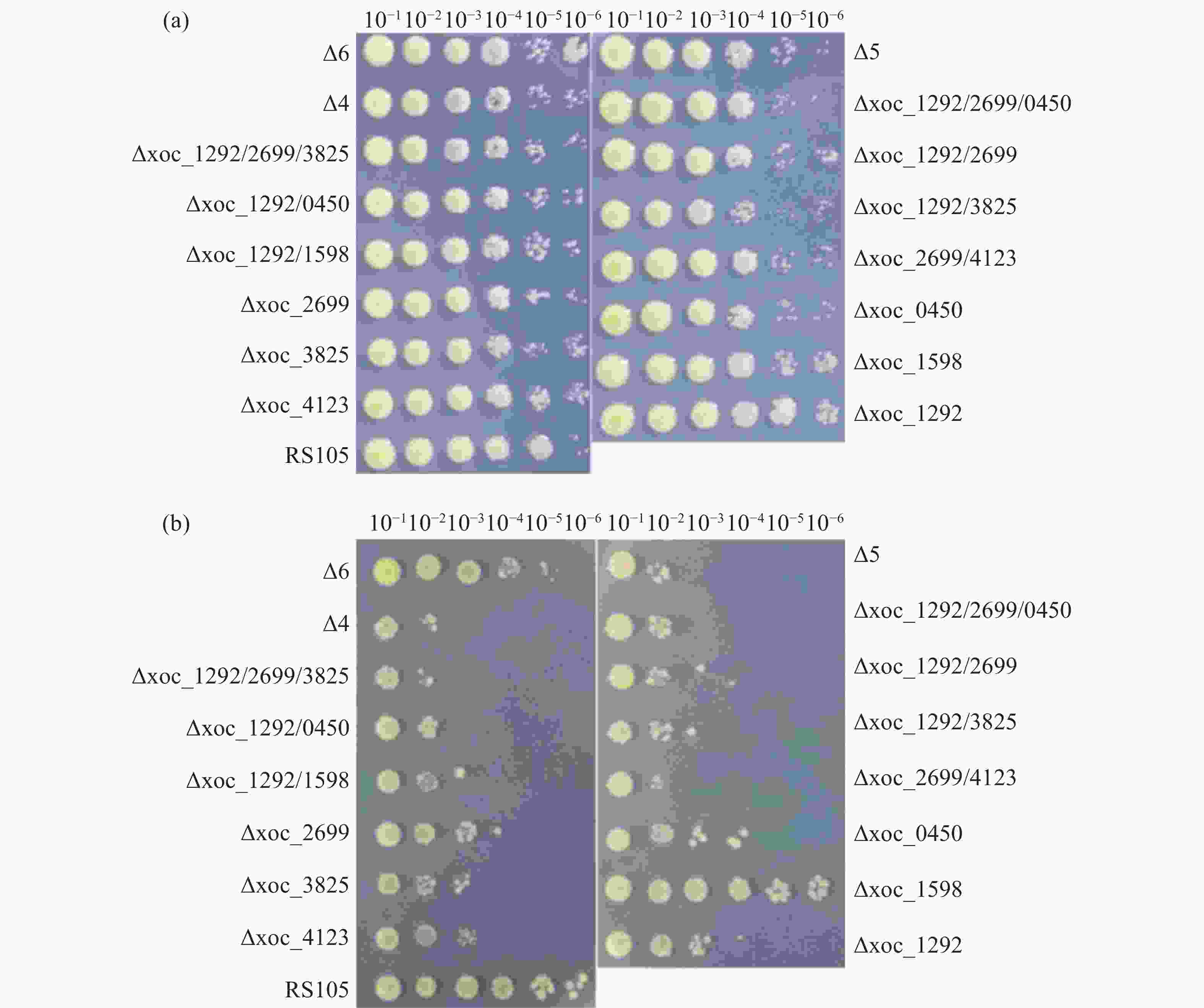
图 3 乙酰转移酶缺失对细菌生长的影响
-
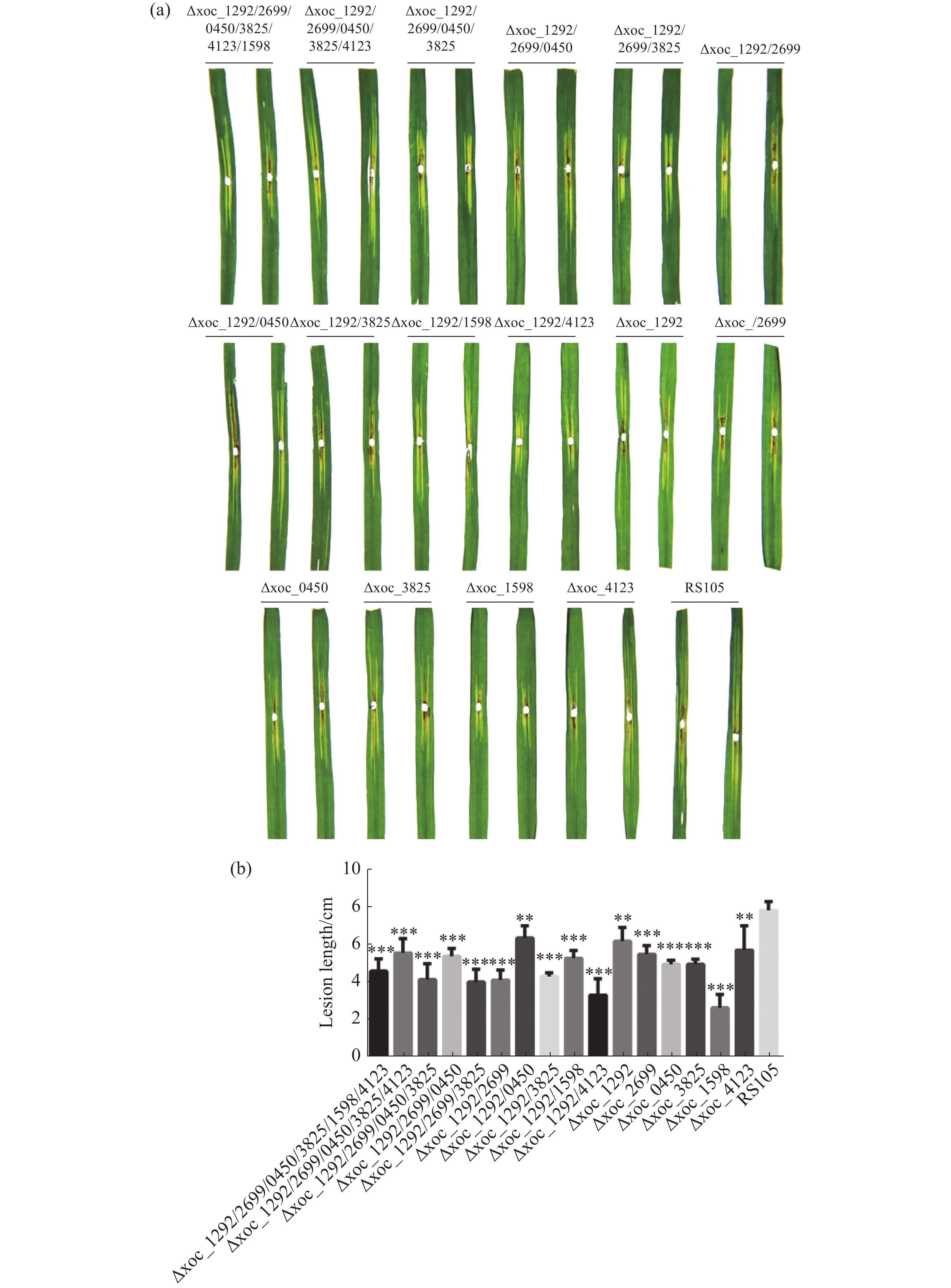
以上实验说明Xoc乙酰转移酶参与调控细菌生长,但其也可能影响Xoc的致病力。因此,将乙酰转移酶突变体和野生型RS105打孔接种TP309水稻,21天后量病斑长度。结果显示,单突突变体和多突突变体菌株致病力较野生型均有所下降(图4)。其中Δxoc_1292/0450、Δxoc_1292、Δxoc_4123病斑长度与野生型相比有显著性差异,其余突变体接种结果与野生型相比差异达到极显著水平,特别是Δxoc_1598的病斑长度与野生型相差达5 cm。这说明乙酰转移酶是调控Xoc致病力的重要因子。

图 4 Xoc乙酰转移酶突变体的致病力
-
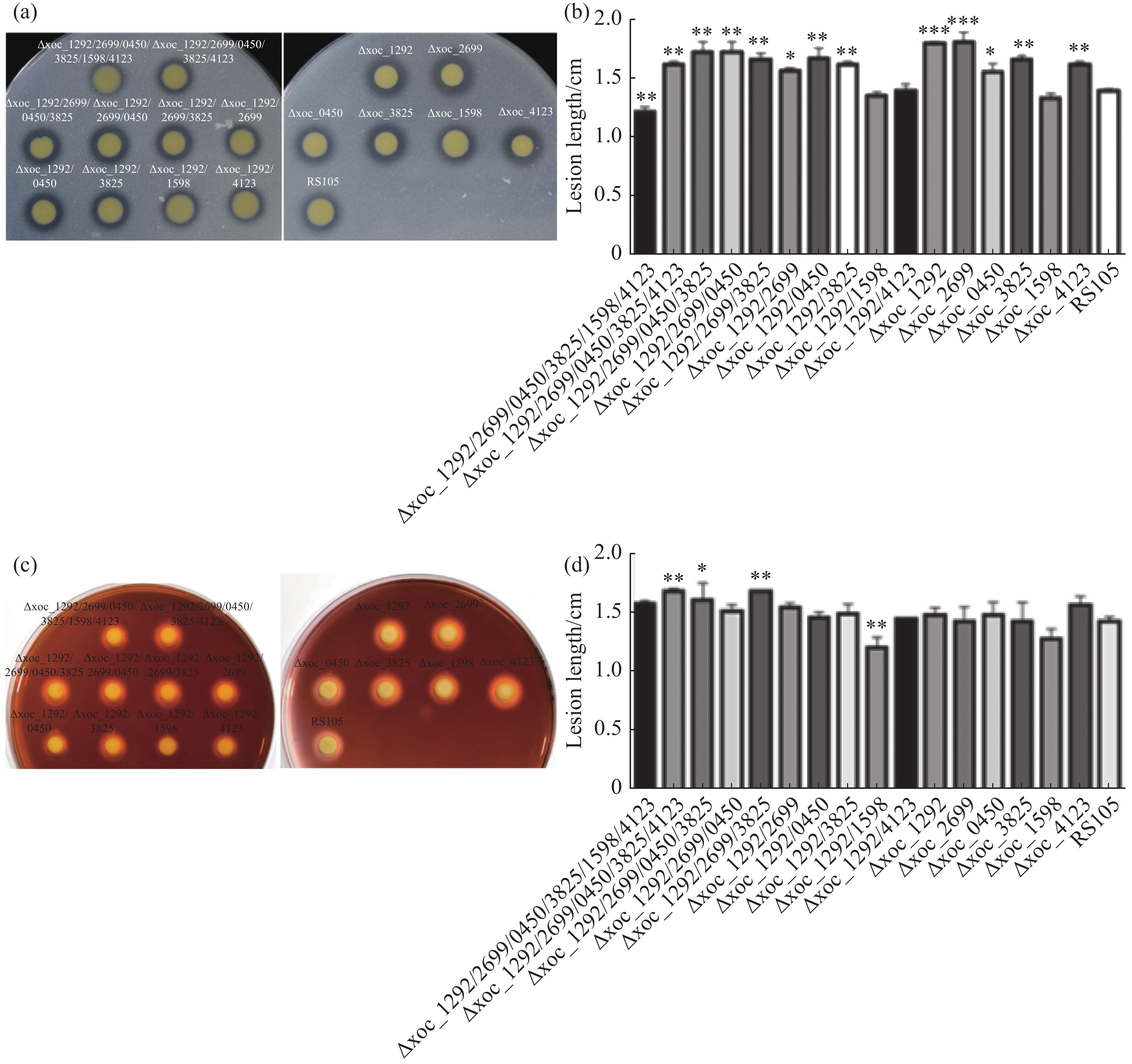
为了探究乙酰转移酶突变是否影响Xoc胞外酶产量,本实验通过平板检测法测定了Xoc胞外淀粉酶和蛋白酶活性。蛋白酶以蛋白质肽链为底物,若细菌分泌胞外蛋白酶,则可以在加入脱脂奶粉的PSA平板上形成透明水解圈。同胞外蛋白酶类似,细菌分泌胞外淀粉酶也可在含淀粉的PSA平板上形成水解圈。结果发现,六突突变体菌株胞外蛋白酶活性几乎完全消失,双突突变体Δxoc_1292/1598、Δxoc_1292/4123和单突突变体Δxoc_1598胞外蛋白酶活性接近野生型,其余突变体蛋白酶活性均高于野生型(图5-a、5-b)。而淀粉酶活性分析显示,双突突变体Δxoc_1292/1598胞外淀粉酶活性几乎完全丧失,而Δ5、Δ4和Δxoc_1292/2699/3825胞外淀粉酶活性大于野生型(图5-c、5-d)。这说明,Xoc乙酰转移酶参与调控胞外蛋白酶活性和胞外淀粉酶活性。
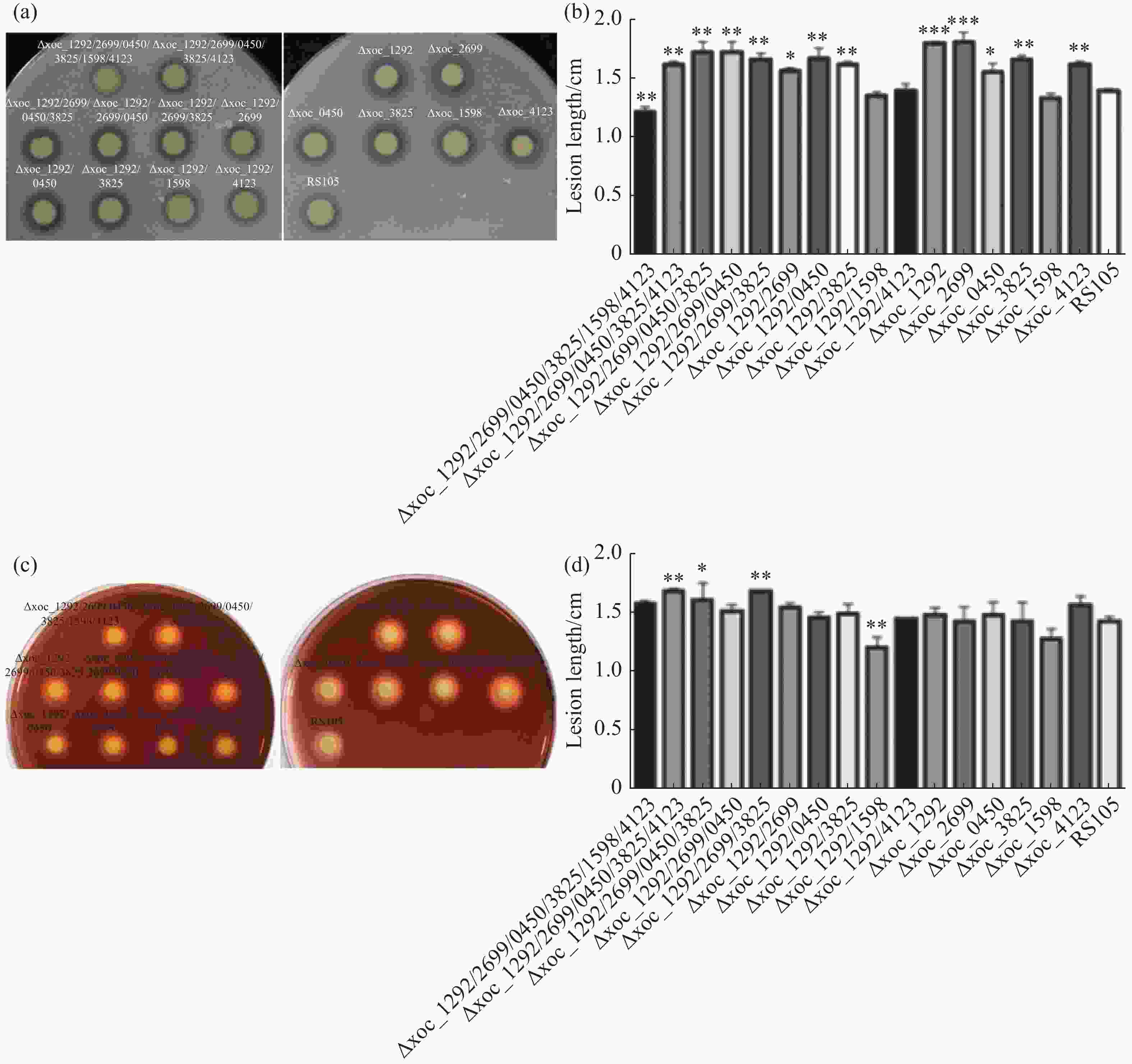
图 5 Xoc乙酰转移酶突变体的胞外淀粉酶和蛋白酶活性
-
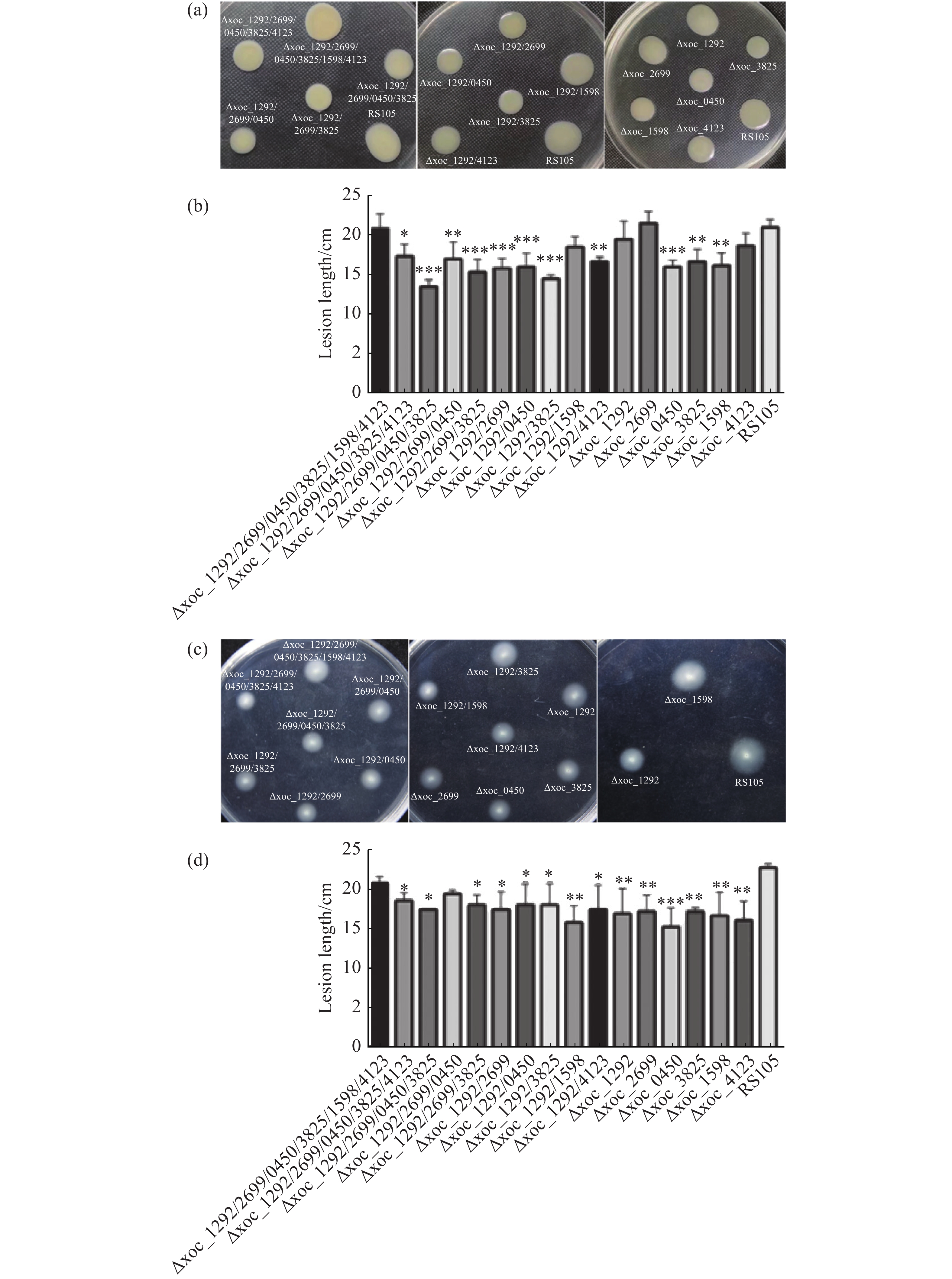
为分析乙酰转移酶是否调控Xoc的运动能力,,本实验分析了由鞭毛控制的游动能力(Swimming)和由菌毛控制的蠕动能力(Swarming)。将细菌悬液OD600调至1.0,分别接种到含有0.6%琼脂的NY1和0.25%琼脂的NY2培养基上,观察野生型和突变体的运动能力差异。在蠕动性方面,Δ4、Δxoc_1292/2699/3825、Δxoc_1292/2699、Δxoc_1292/0450、Δxoc_1292/3825和Δxoc_0450菌落蠕动直径与野生型菌株相比差异达到极显著水平。此外Δ5、Δxoc_1292/2699/0450、Δxoc_1292/4123、Δxoc_3825和Δxoc_1598菌落直径与野生型相比也达到显著水平。而Δ6、Δxoc_1292/1598、Δxoc_1292、Δxoc_2699和Δxoc_4123菌落直径与野生型菌株相比没有显著差异(图6-a、6-b)。
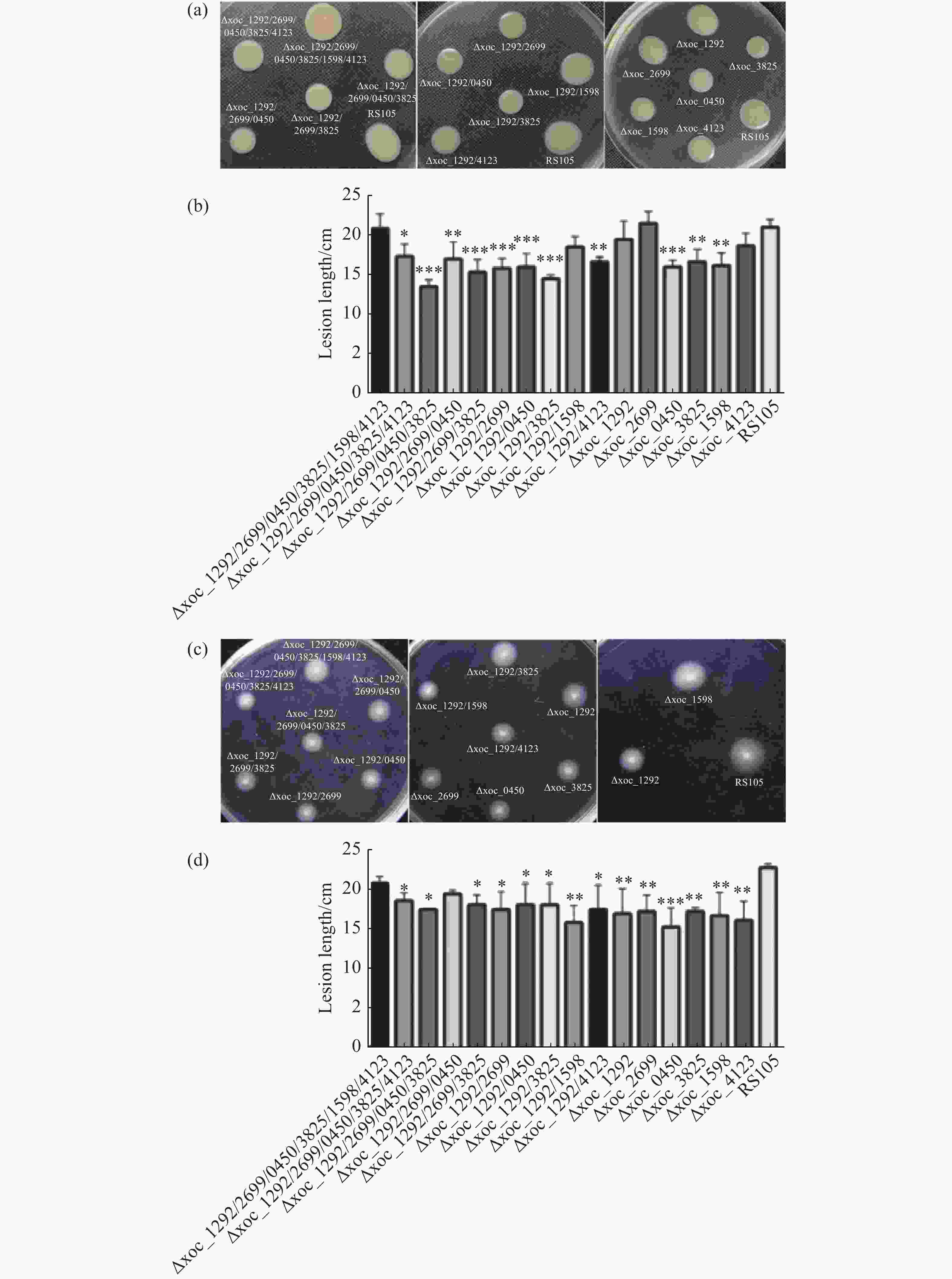
图 6 Xoc乙酰转移酶突变体的运动能力
在泳动性方面,Δxoc_0450平均生长直径为18.3mm,与野生型RS105的平均生长直径为27.3mm相比差异达到极显著水平。此外,Δ5、Δ4、Δxoc_1292/2699/3825、Δxoc_1292/2699、Δxoc_1292/0450、Δxoc_1292/3825、Δxoc_1292/1598、Δxoc_1292/4123、Δxoc_1292、Δxoc_2699、Δxoc_3825、Δxoc_1598、Δxoc_4123共13个突变体菌落生长直径与野生型菌株相比差异达到显著水平。剩余突变体Δ6、Δxoc_1292/2699/0450生长直径与野生型菌株没有显著性差异(图6-c、6-d)。
以上实验结果表明Xoc中乙酰转移酶调控由菌毛与鞭毛介导的运动。此外,在单突突变体中Δxoc_0450、Δxoc_3825和Δxoc_1598的蠕动能力与泳动能力均下降,说明这3个乙酰转移酶可能同时参与调控细菌蠕动性和泳动性。但与预期不符的是,Δ6的蠕动性与泳动性均与野生型菌株相近,具体原因还需进一步研究。
-
乙酰化修饰是细菌细胞功能调控的重要方式之一,在致病力,基础代谢等方面发挥重要作用,如通过乙酰化修饰调节细菌体内多种酶活性[22]。GNAT乙酰转移酶超家族是迄今为止发现的最大乙酰转移酶超家族,广泛分布于原核及真核生物中。GNAT在细菌中具有多种作用:调控细菌生物被膜形成,枯草芽孢杆菌中多种乙酰转移酶对细菌生物被膜形成发挥重要作用[23];调节细菌转录过程,硫叶菌中的Pat同源蛋白可以乙酰化Alba降低其DNA结合能力,减少Alba的转录抑制作用[24];调节细菌代谢通量,大肠杆菌中Pat可以通过乙酰化不同底物来调节细菌对不同碳源的利用[25];调节细菌毒力,大肠杆菌中的毒素激活酰基转移酶TAAT,参与调控成孔毒素的分泌[26]。
在本研究中笔者发现,这些乙酰转移酶对于Xoc的致病力和营养缺陷情况下的生长非常重要,同时对Xoc胞外酶活性和运动性也有调控作用。这与在其他细菌中的乙酰转移酶的功能类似。如分支杆菌中的Eis参与调控细菌致病力,大肠杆菌YfiQ通过乙酰化RcsB蛋白调节细菌分裂、生物被膜和鞭毛合成等[27]。但Xoc致病力下降与细菌生长、运动性和胞外酶活性是否有直接关联还没有证据。研究中发现,Δxoc_1598的生长和胞外酶活性均与野生型菌株相近,而运动性却弱于野生型,同时致病力大幅下降,这表明乙酰转移酶在Xoc中可能形成了一个复杂的调控网络。更重要的是,Δxoc_1292生长表型与Δxoc_1292/1598生长表型一致,都明显弱于野生型,这说明Xoc_1292可能位于Xoc_1598信号下游,参与细菌的生长调控。与预期相反的是,敲掉全部6个GNAT类乙酰转移酶的突变体生长和胞外淀粉酶活性却接近于野生型菌株。细菌中乙酰化有酶促(乙酰转移酶参与)和非酶催化两种。在真核细胞,特别是线粒体中,AcCoA(乙酰辅酶A)可非酶乙酰化蛋白质[28]。据此推测乙酰转移酶全部缺失后,细菌内可能以非酶乙酰化途径为主,用以替代乙酰转移酶的功能。但Δ6在致病力及胞外蛋白酶活性方面仍然弱于野生型,这说明非酶乙酰化并不能完全替代GNAT类乙酰转移酶的功能,可能还有其他类别的乙酰转移酶参与Xoc生长和致病力的调控。但是,同源分析并未在Xoc中鉴定到其他类别的乙酰转移酶。此外,非酶乙酰化反应可分为两种途径:乙酰辅酶A(AcCoA)依赖型和乙酰磷酸(AcP)依赖型[24]。细菌乙酰化位点附近存在酸性残基富集,这种富集可能是非酶乙酰化催化机制的组成部分[29]。由于AcCoA是细菌生存所必需的,同时AcP也是多种细菌中乙酰化修饰的主要乙酰基供体。因此,非酶乙酰化很难在体内进行验证,这也是目前细菌体内酶促乙酰化修饰难以精确定量的原因。尽管如此,本研究发现乙酰转移酶在Xoc生理生化功能调控中起到重要的作用,但其具体的调控机制和靶标蛋白还不清楚,需要进一步研究。
GNAT acetyltransferase regulates the growth and virulence of Xanthomonas oryzae pv. oryzicola
-
摘要: 为探究GNAT乙酰转移酶在稻黄单胞菌稻生致病变种(Xanthomonas oryzae pv. oryzicola,Xoc)中的功能,本研究首先对GNAT家族蛋白进行了结构域分析,随后构建了GNAT编码基因的单突及多突突变体,并比较了野生型菌株和这些突变菌株间的生长速度、胞外酶活性和致病力的差异。结果发现:在营养缺乏条件下,除xoc_1598突变体和敲除全部GNAT类乙酰转移酶基因的6突突变体外,其他所有突变体的生长都弱于野生型,说明此类乙酰转移酶调控Xoc的生长。此外,所有GNAT类乙酰转移酶突变都导致Xoc致病力下降。同时还发现此类乙酰转移酶对运动性、胞外蛋白酶和淀粉酶活性也有调控作用。实验结果表明,乙酰化修饰是Xoc生长和致病力的重要调节机制。Abstract: To investigate the function of GNAT acetyltransferases in Xanthomonas oryzae pv. oryzicola (Xoc), a pathogen of rice bacterial leaf streak, we performed domain analysis of GNAT family proteins in Xoc, and then constructed the single and multiple mutants of these GNAT genes. Finally, we compared the growth rates, extracellular enzyme activities, and virulence of these mutants and the wild type Xoc strains. The results showed that GNAT acetyltransferases regulated Xoc growth in nutrient deficiency media. All mutants except Δxoc_1598 and Δ6 grew more slowly than the wild type, and had lower ability of infection than the wild type strain. We also found that mutation in acetyltransferases could alter motility and the activities of the extracellular protease and amylase. All the results showed that protein acetylation is an important regulatory mechanism for Xoc growth and virulence as well as the virulence-related factors′ production.
-
Key words:
- Xanthomonas oryzae pv. oryzicola /
- acetyltransferase /
- extracellular enzyme /
- virulence
-
图 2 Xoc乙酰转移酶突变体的PCR验证
(a).Δ6基因组PCR产物电泳图。(b). Δ5基因组PCR产物电泳图。(c).Δ4基因组PCR产物电泳图。(d).Δxoc_1292/2699/3825基因组PCR产物电泳图。(e).Δxoc_1292/2699/0450基因组PCR产物电泳图。(f).Δxoc_1292/2699基因组PCR产物电泳图。(g).Δxoc_1292/0450基因组PCR产物电泳图。(h).Δxoc_1292/3825基因组PCR产物电泳图。(i).突变体Δxoc_1292/1598基因组PCR产物电泳图。(j).突变体Δxoc_1292/4123基因组PCR产物电泳图。(k).野生型及单突突变体基因组PCR产物电泳图。基因组标注在图上面,引物标注在下面。M:DL 5 000 maker。Negative:以ddH2O为模板的PCR产物;WT:以RS105基因组为模板的PCR产物。
图 4 Xoc乙酰转移酶突变体的致病力
(a).突变体与野生型菌株接种水稻后的病斑特征;(b).病斑长度;*:P<0.05,**:P<0.01,***:P<0.001;每个菌株设3个生物学重复,下同。
图 5 Xoc乙酰转移酶突变体的胞外淀粉酶和蛋白酶活性
(a).待测菌株在包含2%脱脂奶粉的PSA培养基上的生长情况;(c).待测菌株在包含0.1%可溶性淀粉的PSA培养基上的生长情况;(b)、(d).菌株胞外蛋白酶和淀粉酶活性统计。D:水解产物直径,d:菌落直径。
图 6 Xoc乙酰转移酶突变体的运动能力
(a).待测菌株在NY1平板上的菌落特征;(c).待测菌株在NY2平板上的菌落特征;(b)、(d).菌落直径统计;使用SPSS对菌落直径进行显著性分析,所有突变体与野生型菌株进行对比,*: P<0.05, **: P<0.01, ***: P<0.001。每个菌株设3个生物学重复。
表 1 引物序列
名称 序列 xocmindFF GACGAAGCTTCAGCCAGAAGTAGCTGGCAC xocmindFR ACACGGATCCATCGAGCTCGTGCTCGCGCAC xoc1292ddRF ACACGGATCCGTGTCGGTCGCGCGTTCTATG xoc1292ddRR GACGTCTAGACACGTCCGGCACCTGCGACAG xoc2699ddFF ACTGAAGCTTGATCAATACCTCGGTCGGTC xoc2699ddFR CTGCTCTAGACATGTGGTCAGTCGATGTAG xoc2699ddRF CTGCTCTAGACCTCCTTCTGATGAAAACCC xoc2699ddRR CTACGAATTCCATCTTGCCGAAGAATCCCA xoc0450ddFF ACTGAAGCTTCGGTGGTCCCGTATCCGAAG xoc0450ddFR CTGCTCTAGAGTGGTACGTACTCATCGGAC xoc0450ddRF CTGCTCTAGAGGACGATGCGACCATGATGG xoc0450ddRR CTACGAATTCGACGGATAGTTGTCGGTCAG xoc3825ddFF ACTGAAGCTTCAGAGCTGCCATGAGCGTGT xoc3825ddFR CTGCTCTAGAGGCACGTTCAATGCGCTCAC xoc3825ddRF CTGCTCTAGACGTTGGTGATGGCGATCGAG xoc3825ddRR CTACGAATTCCGTTAGTCGTTGCAACCCGT xoc1598ddFF ACTGAAGCTTCTCAAGAAGCATCTCGGCGA xoc1598ddFR CTGCTCTAGATGGCCATGACCGGTGTTCCT xoc1598ddRF CTGCTCTAGATGGAGCTGGATGCCTGAGGT xoc1598ddRR CTACGAATTCGAAGGTAGGCGACGGCATCA xoc4123ddFF ACTGAAGCTTGGTAGGCGTCACGAACAGAT xoc4123ddFR CTGCTCTAGAGTGCATTGGGTCCGAGAAAC xoc4123ddRF CTGCTCTAGAGACTAGGTGCGGTGACCAAG xoc4123ddRR CTACGAATTCCCTTGACCATGACCTTGGTC 注:下划线表示酶切位点。  下载: 导出CSV
下载: 导出CSV
-
[1] AWODERV V A, BANGURA N, JOHN V T. Incidence, distribution and severity of bacterial diseases on rice in West Africa[J]. Tropical Pest Management, 1991, 37(2): 113 − 117. [2] NIñO-LIU D O, RONALD P C, BOGDANOVE A J. Xanthomonas oryzae pathovars: model pathogens of a model crop[J]. Molecular Plant Pathology, 2006, 7(5): 303 − 324. doi: 10.1111/j.1364-3703.2006.00344.x [3] BüTTNER D, BONAS U. Regulation and secretion of Xanthomonas virulence factors[J]. FEMS Microbiology Reviews, 2010, 34(2): 107 − 133. doi: 10.1111/j.1574-6976.2009.00192.x [4] PFEILMEIER S, CALY D L, MALONE J G. Bacterial pathogenesis of plants: future challenges from a microbial perspective: Challenges in bacterial molecular plant pathology[J]. Molecular Plant Pathology, 2016, 17(8): 1298 − 1313. doi: 10.1111/mpp.12427 [5] HENTCHEL K L, ESCALANTE-SEMERENA J C. Acylation of biomolecules in prokaryotes: a widespread strategy for the control of biological function and metabolic stress[J]. Microbiology and Molecular Biology Reviews, 2015, 79(3): 321 − 346. doi: 10.1128/MMBR.00020-15 [6] XIE L, ZENG J, LUO H, et al. The roles of bacterial GCN5-related N-acetyltransferases[J]. Critical Reviews™ in Eukaryotic Gene Expression, 2014, 24(1): 77-87. [7] CHRISTENSEN D G, BAUMGARTNER J T, XIE X, et al. Mechanisms, detection, and relevance of protein acetylation in prokaryotes[J]. mBio, 2019, 10(2): e02708 − e02718. [8] LIU W, TAN Y, CAO S, et al. Protein acetylation mediated by YfiQ and CobB is involved in the virulence and stress response of Yersinia pestis[J]. Infection and Immunity, 2018, 86(6): e00224 − e00218. [9] YOSHIKAWA A, ISONO S, SHEBACK A, et al. Cloning and nucleotide sequencing of the genes rimI and rimJ which encode enzymes acetylating ribosomal proteins S18 and S5 of Escherichia coli K12[J]. Molecular & General Genetics MGG, 1987, 209(3): 481 − 488. doi: 10.1007/BF00331153 [10] PATHAK D, BHAT A H, SAPEHIA V, et al. Biochemical evidence for relaxed substrate specificity of Nα-acetyltransferase (Rv3420c/rimI) of Mycobacterium tuberculosis[J]. Scientific Reports, 2016, 6(1): 28892. doi: 10.1038/srep28892 [11] LI H, YU C, CHEN H, et al. PXO00987, a putative acetyltransferase, is required for flagellin glycosylation, and regulates flagellar motility, exopolysaccharide production, and biofilm formation in Xanthomonas oryzae pv. oryzae[J]. Microbial Pathogenesis, 2015, 85: 50 − 57. doi: 10.1016/j.micpath.2015.06.001 [12] CHEONG M S, KIRIK A, KIM J G, et al. AvrBsT acetylates Arabidopsis ACIP1, a protein that associates with microtubules and is required for immunity[J]. PLoS pathogens, 2014, 10(2): e1003952. doi: 10.1371/journal.ppat.1003952 [13] HAN S W, HWANG B K. Molecular functions of Xanthomonas type III effector AvrBsT and its plant interactors in cell death and defense signaling[J]. Planta, 2017, 245(2): 237 − 253. doi: 10.1007/s00425-016-2628-x [14] SCHäFER A, TAUCH A, JäGER W, et al. Small mobilizable multi-purpose cloning vectors derived from the Escherichia coli plasmids pK18 and pK19: selection of defined deletions in the chromosome of Corynebacterium glutamicum[J]. Gene, 1994, 145(1): 69 − 73. doi: 10.1016/0378-1119(94)90324-7 [15] KUMAR S, STECHER G, TAMURA K. MEGA7: molecular evolutionary genetics analysis version 7.0 for bigger datasets[J]. Molecular Biology and Evolution, 2016, 33(7): 1870 − 1874. doi: 10.1093/molbev/msab120 [16] CHEN C, CHEN H, ZHANG Y, et al. TBtools: an integrative toolkit developed for interactive analyses of big biological data[J]. Molecular plant, 2020, 13(8): 1194 − 1202. doi: 10.1016/j.molp.2020.06.009 [17] 赵严, 罗登杰, 何圣贤等. 水稻细菌性条斑病4种接种方法的比较[J]. 亚热带农业研究, 2018, 14(4): 242 − 246. doi: 10.13321/j.cnki.subtrop.agric.res.2018.04.005 [18] 韦梅良. 用野油菜黄单胞菌插入突变体库筛选与胞外蛋白酶产生相关的基因[D]. 南宁:广西大学, 2002. [19] ZHANG Z C, ZHAO M, XU L D, et al. Genome-wide screening for novel candidate virulence related response regulator genes in Xanthomonas oryzae pv. oryzicola[J]. Frontiers in Microbiology, 2018, 9: 1789. doi: 10.3389/fmicb.2018.01789 [20] HENTCHEL K L, ESCALANTE-SEMERENA J C. In Salmonella enterica, the Gcn5-related acetyltransferase MddA (formerly YncA) acetylates methionine sulfoximine and methionine sulfone, blocking their toxic effects[J]. Journal of bacteriology, 2015, 197(2): 314 − 325. doi: 10.1128/JB.02311-14 [21] YU M, DE CARVALHO L P S, SUN G, et al. Activity-based substrate profiling for Gcn5-related N-acetyltransferases: the use of chloroacetyl-coenzyme a to identify protein substrates[J]. Journal of the American Chemical Society, 2006, 128(48): 15356 − 15357. doi: 10.1021/ja066298w [22] NAKAYASU E S, BURNET M C, WALUKIEWICZ H E, et al. Ancient regulatory role of lysine acetylation in central metabolism[J]. mBio, 2017, 8(6): e01817 − e01894. [23] REVERDY A, CHEN Y, HUNTER E, et al. Protein lysine acetylation plays a regulatory role in Bacillus subtilis multicellularity[J]. PLoS One, 2018, 13(9): e0204687. doi: 10.1371/journal.pone.0204687 [24] MARSH V L, PEAK-CHEW S Y, BELL S D. Sir2 and the acetyltransferase, Pat, regulate the archaeal chromatin protein, Alba[J]. The Journal of Biological Chemistry, 2005, 280(22): 21122 − 21128. doi: 10.1074/jbc.M501280200 [25] WANG Q, ZHANG Y, YANG C, et al. Acetylation of metabolic enzymes coordinates carbon source utilization and metabolic flux[J]. Science, 2010, 327(5968): 1004 − 1007. doi: 10.1126/science.1179687 [26] GREENE N P, CROW A, HUGHES C, et al. Structure of a bacterial toxin-activating acyltransferase[J]. Proceedings of the National Academy of Sciences of the United States of America, 2015, 112(23): e3058−e3066. [27] WOLFE A J. Bacterial protein acetylation: new discoveries unanswered questions[J]. Current Genetics, 2016, 62(2): 335 − 341. doi: 10.1007/s00294-015-0552-4 [28] HOSP F, LASSOWSKAT I, SANTORO V, et al. Lysine acetylation in mitochondria: From inventory to function[J]. Mitochondrion, 2017, 33: 58 − 71. doi: 10.1016/j.mito.2016.07.012 [29] CARABETTA V J, CRISTEA I M. Regulation, function, and detection of protein acetylation in bacteria[J]. Journal of bacteriology, 2017, 199(16): e00107 − e00117. -

 点击查看大图
点击查看大图
计量
- 文章访问数: 319
- HTML全文浏览量: 255
- PDF下载量: 11
- 被引次数: 0
