

-
水稻(Oryza sativa L.)是我国乃至世界上最重要的粮食作物之一,对保障我国粮食安全意义重大[1]。目前海南岛稻田以双季稻耕作模式为主,长期的连作和依赖化学肥料,降低了土壤微生物活性和土壤养分的转化,这导致作物减产和土传病害增加,不利于农业生产的可持续发展[2-3]。稻田采用水旱轮作是克服水稻连作障碍的有效措施[4]。轮作是指在同一块土地上有计划地按顺序种植不同种类作物的复种方式[5]。水旱轮作能够显著提高土壤有机质的矿化速率,使土壤有机物质短期内快速下降,导致团聚体裂解和重组,促进养分的周转和循环利用[6]。土壤团聚体是土壤结构的基础[7],它在土壤中具有保证和协调土壤中的水肥气热、影响土壤微生物活性、维持和稳定土壤疏松熟化具有重要意义[8],但团聚体的形成和稳定性受生物和非生物因素的影响较多[9]。在非生物因素中,轮作能够显著增加表层土壤水稳性大团聚体的数量,从而促进土壤团聚体形成和稳定[10]。在生物因素中,土壤微生物通过代谢活动释放的代谢产物和酶类物质来影响土壤有机质及土壤团聚过程[11]。稻豆轮作能够缓解土壤盐渍化、有效提高土壤大团聚体的比例,增加团聚体的稳定性,提高土壤有机碳的含量,增加土壤微生物多样性,对实现农业可持续发展具有重要意义[12-14]。目前,关于稻豆轮作对土壤理化性质和微生物群落结构的影响研究报道较少。为此,以水稻连作和稻豆轮作为研究对象,结合土壤理化性质和微生物的分析方法,探究不同施肥处理对土壤的综合影响,以期为稻豆轮作的实际应用提供科学依据。
-
试验地为海南省东方市农乐南繁科技有限公司田间实验基地(18°87′ N,108°70′E),属热带季风海洋性气候,年平均降水量为950 mm,年平均蒸发量2 300 mm,土壤类型为水稻土。试验布置前耕层土壤基础理化性质为pH 5.4,土壤全氮0.43 g·kg−1,有机质1.34 %,有效磷28.51 mg·kg−1、速效钾84.98 mg·kg−1。
-
试验开始于2021年7月,试验共设2个处理:水稻连作(稻-稻-稻)处理(RRR)和水稻-大豆-水稻轮作(稻-豆-稻)处理(RSR);每个处理4次重复,共8个小区。水稻季氮、磷、钾肥用量分别为180、90、120 kg·hm−2,磷肥和钾肥作为底肥一次性施入,氮肥分别在播前、分蘖期、抽穗期按比例40∶35∶25施入;大豆季氮、磷、钾肥用量为150、150 、150 kg·hm−2 。
供试水稻品种为‘寿乡一号’,大豆品种为‘闽豆64’,由海南农乐南繁科技有限公司提供。水稻连作播种日期:2021年7—11月;收获日期:2022年2—6月、2022年8—11月;水稻播种量为45 kg·hm−2,株行距为25 cm×25 cm。稻豆轮作播种日期:2021年7—11月,收获日期:2021年12月—2022年3月、2022年8—11月;水稻播种量为90 kg·hm−2,株行距为20 cm×25 cm,播种深度为5 cm。试验周期为1 a,栽培和植保等田间管理与农户保持一致。
-
采用5点取样法采集RRR和RSR处理种植前后0~30 cm耕层土壤,过2 mm筛并将土壤样品分为3个部分,一部分保存于−80 ℃冰箱中,用于测定微生物多样性;一部分存于4 ℃冰箱中,用于测定土壤微生物量碳氮及有机碳组分;剩余部分风干,用于测定土壤理化性质及土壤团聚体。
-
土壤微生物生物量碳采用氯仿熏蒸-K2SO4浸提法测定[15];水溶性碳(WSC)和热水可氧化碳(HWEC)含量采用振荡浸提分光光度法测定[16];测土壤pH时按土水质量比1∶2.5配样,pH计测定[17];土壤有机碳采用外加热重铬酸钾氧化法容量测定;全氮用凯氏定氮法测定;有效磷采用0.5 mol·L−1碳酸氢钠法测定;速效钾采用中性乙酸浸提法测定[18];土壤团聚体稳定性采用湿筛法测定[19]。
-
根据DNeasy Power Soil Kit(QIAGEN)试剂盒说明书对土壤总DNA进行提取,利用Novaseq 6000 PE250平台对提取的DNA质量浓度和纯度进行检测;对细菌16S rDNA V4+V5区和真菌ITS1F和2R区域进行扩增。对构建好的文库进行检验、测序。对测序结果进行操作单位聚类与分析、Alpha多样性分析、Beta多样性分析及不同分类水平的群落结构分析。
-
于2021−11−08、2022−03−15、2022−11−25测定RRR和RSR处理水稻成熟期的产量。植株全氮含量采用凯氏定氮法测定[18],并计算周年产量、吸氮量和氮肥利用率。计算如下:(1)产量=每季产量之和;(2)吸氮量=每季(秸秆+籽粒)含氮量×产量;(3)氮肥利用率=吸氮量×氮肥投入量。
-
采用Excel 2016和IBM SPSS 23.0软件对数据进行统计分析,采用单因素(one-way ANOVA)进行方差分析,Duncan法进行多重比较(P<0.05),利用软件Origin Pro 2021作图。图标中数据为平均值±标准差。
-
如表1所示, RRR处理与RSR处理相比,水稻周年产量、周年吸氮量、氮素利用效率和下季水稻产量产生了显著变化。RSR处理的周年产量为7869.2 kg·hm−2,与RRR处理相比降低了17.3%。RSR处理周年吸氮量为274.2 kg·hm−2,氮素利用率为83.1%,与RRR处理相比分别显著提高了56.2%和70.3%。RSR处理的第二季水稻产量为5865.4 kg·hm−2,与RRR处理相比提高了23.5%。
表 1 稻稻连作和稻豆轮作周年产量、周年吸氮量、氮素利用率和第二季水稻产量
处理 氮投入量/(kg·hm−2) 周年产量 /(kg·hm−2) 周年吸氮量/(kg·hm−2) 氮素利用率/% 第二季水稻产量/(kg·hm−2) RRR 360 9520.6±115.5a 175.6±12.6b 48.8±3.47b 4748.5±26.3b RSR 330 7869.2±367.8b 274.2±8.10a 83.1±2.46a 5865.4±36.1a 注: 小写字母表示不同处理之间的显著差异(P <0.05)。下同 -
如表2所示,与RRR处理相比,RSR处理土壤理化性质产生了显著变化。RSR处理土壤的全氮含量为0.57 g·kg−1,有效磷含量为38.4 mg·kg−1,与RRR处理相比分别显著提高了16.6%和12.6%。土壤速效钾和有机质含量为109.1 mg·kg−1和15.7 g·kg−1,与RRR处理相比分别显著提高了14.3%和26.6%。RSR处理土壤pH为5.14,与RRR处理相比降低了0.51。
表 2 不同处理对土壤理化性质的影响
处理 有机质/(g·kg−1) 全氮/(g·kg−1) 有效磷/(mg·kg−1) 速效钾/(mg·kg−1) pH RRR 12.4±0.23b 0.49±0.06a 34.1±1.38b 95.4±0.98b 5.65±0.2a RSR 15.7±0.46a 0.57±0.03a 38.4±4.79a 109.1±9.44a 5.14±0.1b -
土壤团聚体作为土壤的基本结构之一,其组成和稳定性影响土壤养分在团聚体中的分布。不同粒级团聚体的组成是鉴定土壤肥力状况的指标之一。土壤团聚体的组成主要受不同粒径分布和轮作模式影响。如表3所示,轮作处理土壤水稳性平均质量直径(MWD)和几何平均直径为0.32 和0.906 mm,与RRR处理相比,上升了0.01 和下降了0.001 mm,大团聚体比例( R >0.25)为0.24 mm,下降了0.01 mm。
表 3 不同处理对土壤水稳性团聚体的影响
mm 处理 平均质量直径(MWD) 几何平均直径(GMD) 大团聚体比例(R>0.25) RRR 0.31±0.01a 0.907±0.0001a 0.23±0.04a RSR 0.32±0.01a 0.906±0.0001a 0.24±0.03a -
土壤碳组分能直观反映土壤碳动态,常与养分循环、团聚体稳定性等土壤功能联系在一起。同时土壤碳组分也是土壤健康的重要组成部分,与土壤养分有效性及微生物活性密切相关。土壤碳组分中除土壤有机碳(SOC)外,微生物生物量碳(MBC)、热水可提取碳(HWEC)、水溶性碳(WSC)也是碳组分的重要组成部分,是反映土壤健康的重要指标。如表4所示,RRR处理与RSR处理相比,土壤有机碳组分产生了显著性变化。RSR处理的热水可提取碳(HWEC)、水溶性碳(WSC)含量分别为43.7、10.3 mg·kg−1,与RRR处理相比分别显著提高了35.7%和18.9%。RSR处理微生物生物量碳(MBC)和土壤有机碳(SOC)的含量分别为34.8 和1.79 g·kg−1,与RRR处理相比,显著提高38%和26.9%。
表 4 不同处理对土壤有机碳组分的影响
处理 土壤有机碳(SOC)/
(g·kg−1)热水可提取碳(HWEC)/
(mg·kg−1)水溶性碳(WSC)/
(mg·kg−1)微生物生物量碳(MBC)/
(mg·kg−1)RRR 1.41±0.03b 32.2±0.74b 8.66±1.12b 25.2±0.60b RSR 1.79±0.05a 43.7±2.14a 10.3±0.04a 34.8±2.49a -
Ace值是用来估计群落中含有OTU数目的指数,使用Chao1算法估算群落中所含OTU数目,其中Ace和Chao1的数值越大代表样本中所含物种越多;Shannon和Simpson指数用于评估样本中物种组成的多样性,数值越大代表该环境中的多样性越高。如表5所示,RRR处理与RSR处理相比,土壤微生物丰度和多样性产生了显著性变化。RSR处理下土壤细菌的ACE指数、Chao1指数和Shannon指数,与RRR处理相比,分别显著提高了40.2%、40.3%、5.7%,对Simpson指数则无显著影响。RSR处理与RRR处理相比,土壤真菌Shannon指数和Simpson指数,显著提高了20.1%和16.5%。稻豆轮作没有对土壤真菌Ace指数和Chao1指数产生显著性影响。
表 5 不同处理对土壤微生物α多样性的影响
种植方式 微生物 ACE指数 Chao1 指数 Shannon指数 Simpson指数 RRR 细菌 Bacteria 1390.8±35.2b 1386.9±34.5b 9.33±0.13b 0.99±0.0006a RSR 细菌 Bacteria 1949.6±142.1a 1945.9±142.5a 9.86±0.14a 0.98±0.0003a RRR 真菌 Fungus 265.7±44.5a 265.5±44.4a 4.03±0.33b 0.79±0.04b RSR 真菌 Fungus 328.5±42.4a 328.5±42.4a 4.84±0.25a 0.92±0.02a -
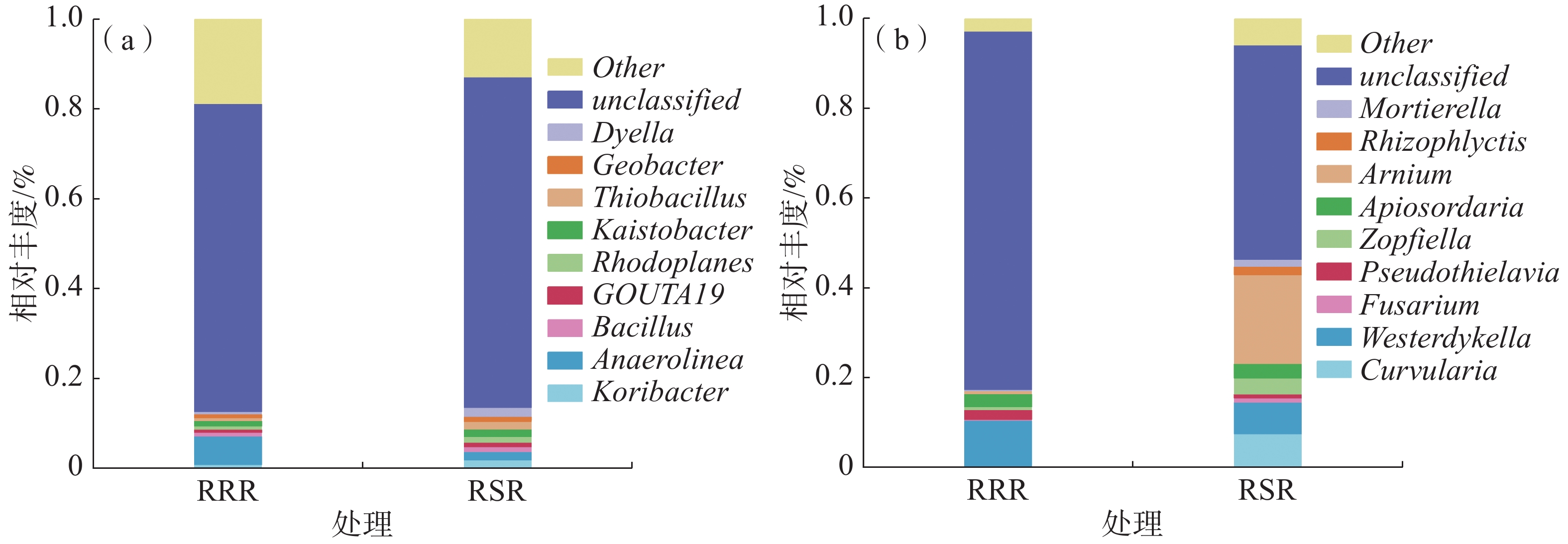
不同耕作条件下土壤共有11个优势细菌属(选取相对丰度前10位,其他属为other)。如图1-a所示,土壤细菌属水平的群落结构基本一致。其中,轮作处理下的优势细菌属为Koribacter菌属(平均相对丰度为1.7%)、红游动菌属(Rhodoplanes 平均相对丰度为1.3%)、硫杆菌(Thiobacillus 平均相对丰度为1.6%)、戴氏菌属(Dyella 平均相对丰度为1.9%),这4个优势菌属累计相对丰度为6.5%。与RRR处理相比,RSR处理Koribacter菌属、红游动菌属(Rhodoplanes)、硫杆菌属(Thiobacillus)、戴氏菌属(Dyella)的相对丰度分别显著提高了35.4%、91.4%、161.8%、275.5%,Anaerolinea菌属的相对丰度显著降低了69.9%。

图 1 不同处理对细菌优势菌属(a)和真菌优势菌属(b)的影响
不同耕作条件下土壤共有11个优势真菌属(选取相对丰度前10位,其他属other)。如图1-b所示,土壤真菌属水平的群落结构基本一致。 其中,轮作处理下的优势真菌属为小被孢霉属(Mortierella 平均相对丰度1.5%)、柄孢壳属(Zopfiella 平均相对丰度3.5%)、镰刀菌属(Fusarium 平均相对丰度1%)、Apiosordaria属(Apiosordaria 平均相对丰度1.5%),这4个优势菌属累计相对丰度为1.9%。与RRR处理相比,RSR处理小被孢霉属(Mortierella)、柄孢壳属(Zopfiella)、镰刀菌属(Fusarium)、Apiosordaria属的相对丰度分别提高了690.6%、428.7%、398.4%、11.3%。Westerdykella菌属和伪硫杆菌 (Pseudothielavia)的相对丰度分别显著降低31.9%和57.4%。
-
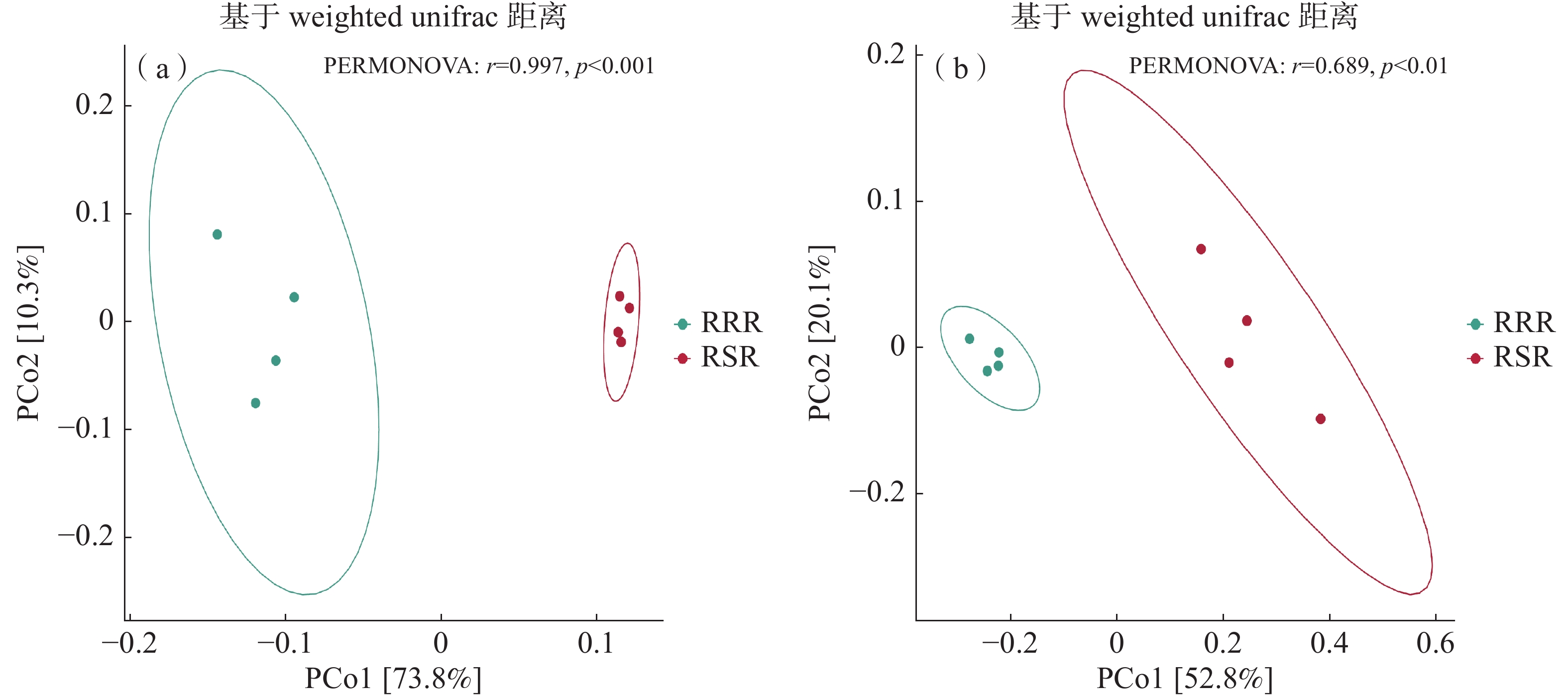
采用基于Bray-Curtis距离矩阵的主坐标分析(PCoA)来分析不同处理下的土壤细菌群落结构差异。PCoA结果的分析表明(图2-a),前2个主成分解释了84.1%的细菌群落变异;结合PERMANOVA分析结果表明,不同耕作模式下细菌OTU数目在两轴间存在显著差异,说明RSR处理对土壤细菌群落结构产生了显著性影响。PCoA结果的分析表明(图2-b),前2个主成分解释了72.9%的真菌群落变异;结合PERMANOVA分析的结果表明,不同耕作措施下真菌OTU数目在两轴间存在显著差异,说明RSR处理对土壤真菌群落结构产生了显著性影响。
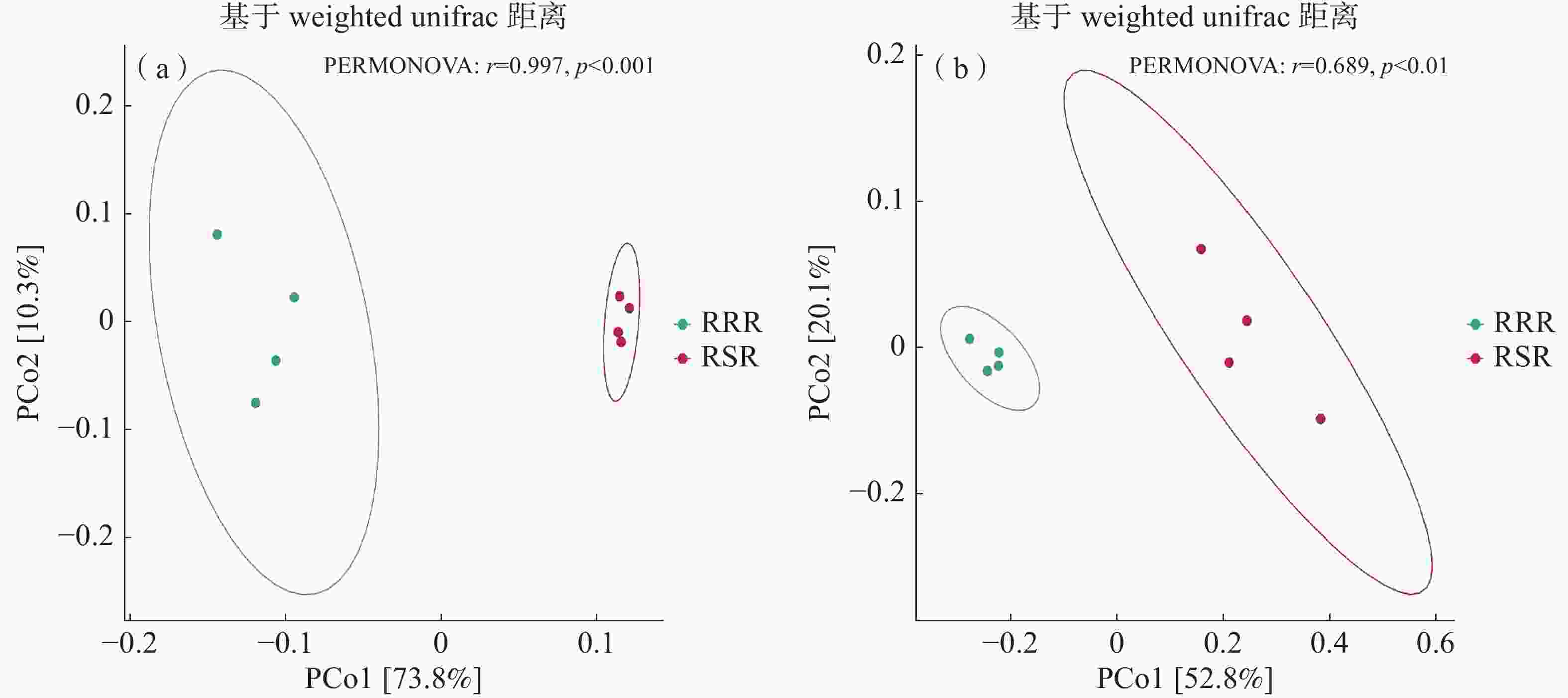
图 2 不同处理下土壤细菌(a)和真菌(b)群落结构PCoA分析
-
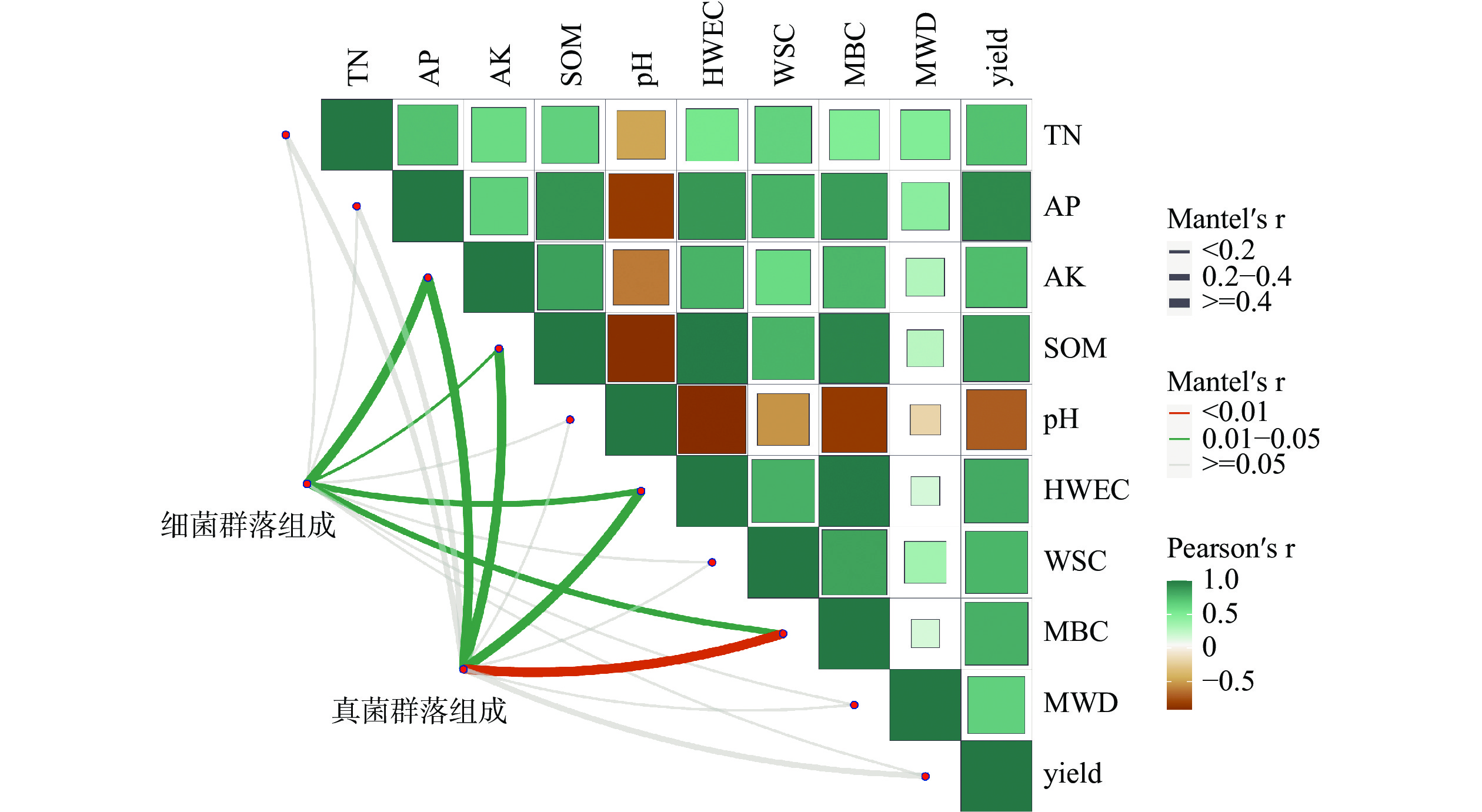
土壤细菌群落(图3)与土壤速效钾(r=0.49)、有机质(r=0.19)、HWEC (r=0.25)、MBC (r=0.33)呈显著相关性(P<0.05)。说明土壤速效钾、有机质、HWEC和MBC对土壤细菌产生了显著影响,而与土壤全氮、有效磷、pH、WSC、MWD、产量无明显差异(P>0.05)。这说明土壤细菌主要受土壤速效钾、有机质、HWEC、MBC指标变化而改变。
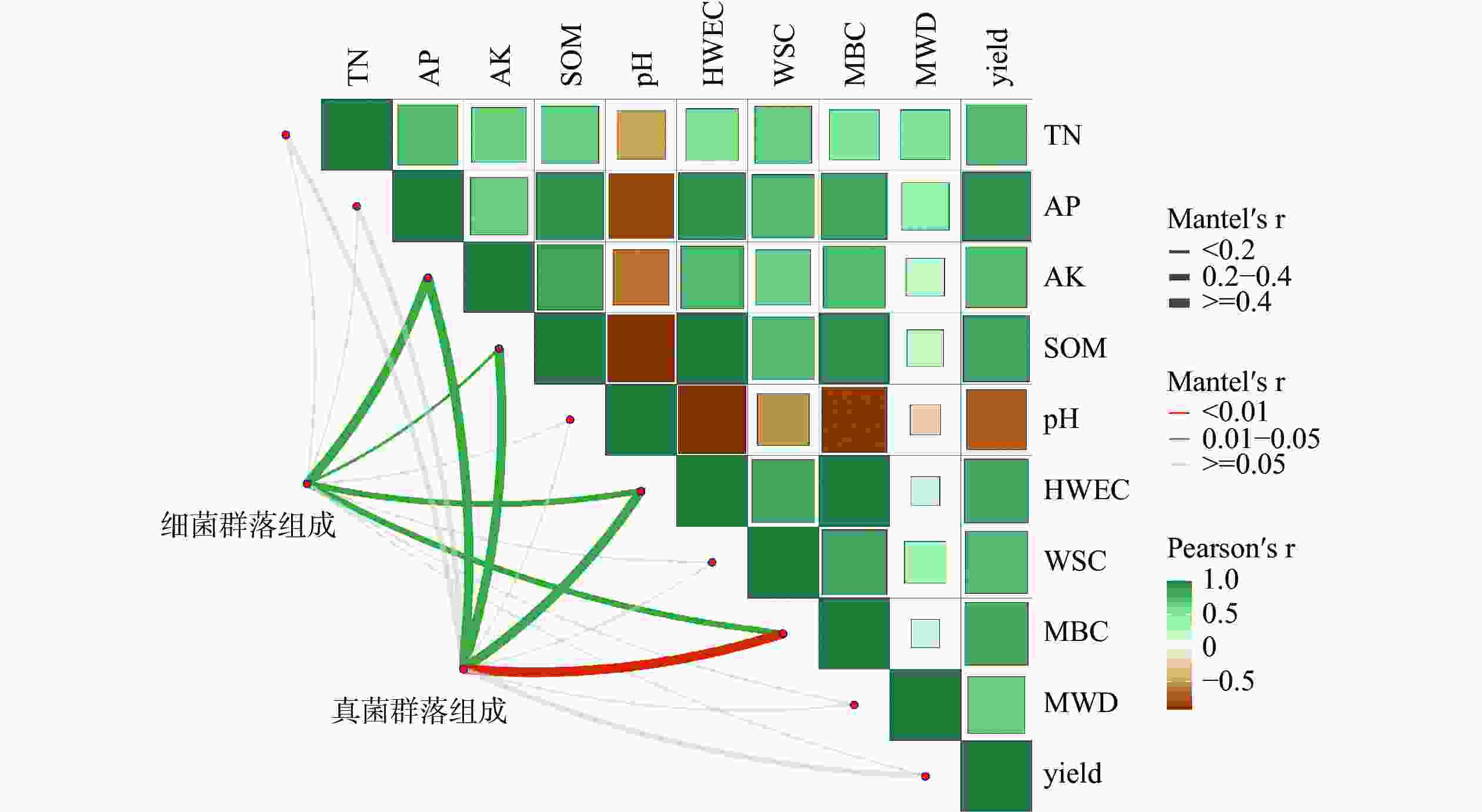
图 3 土壤细菌、真菌群落组成与土壤性质的关系
土壤真菌群落(图3)与土壤速效钾(r=0.60)、有机质(r=0.56)、HWEC(r=0.58)呈显著相关性(P<0.05),与MBC(r=0.59)呈极显著相关性(P<0.01)。这说明土壤真菌群落主要会随着土壤速效钾、有机质、HWEC和MBC的变化而发生改变。
-
土壤养分含量和作物产量是衡量土壤能否可持续生产的主要指标,而土壤养分是提高轮作系统生物量的主要因素[20-21]。与水稻连作相比,RSR处理水稻周年产量下降了17.3%。RSR处理周年吸氮量和氮素利用率,与RRR相比提高了56.2%和70.3%。RSR处理第二季水稻产量,与RRR处理相比提高了23.5%。因为轮作处理中大豆的单产较低,从而使轮作周年产量下降,而同时大豆籽粒含氮量较高,使RSR处理周年氮产量得到提高。轮作影响土壤团粒结构,改善了土壤养分供给能力,促进植株的干物质积累与营养元素含量,从而提高了下茬水稻产量[21-23]。
-
本研究结果表明,稻豆轮作后pH降低了0.51,这是因为大豆根系分泌物中的大量有机酸分子能够增加土壤中的H+,使根际土壤酸化,从而改变了土壤的pH[24]。此外,稻豆轮作增加了土壤氮、有效磷、速效钾的含量。这是由于轮作改变了土壤微生物群落结构与多样性,使得土壤胶结物质所固定的养分元素在微生物的介导下得到释放,提高了土壤养分的有效性[25-26]。
土壤团聚体是土壤结构的基础,土壤团聚体平均质量直径、几何平均直径和大团聚体比例是表示土壤团聚体稳定性的关键指标,它们的值越高,表征土壤团聚度越好,团聚体结构越稳定[19,25]。轮作下土壤微团聚体比例没有显著变化,虽然轮作促进了土壤微生物、植物等生物因素的变化,但由于轮作时间较短,对土壤团聚体没有产生显著影响[26]。
-
本研究结果表明,稻豆轮作使HWEC、WSC、MBC和SOC含量分别显著提高了35.7%、18.9%、38%和26.9%。众多研究指出,种植制度和耕作栽培方式的不同,轮作明显提高了土壤中有机碳,增加了氮素供应来提高微生物对其利用率,也可能因为激发效应降低土壤碳组分含量,从而改变土壤中有机碳组分,这与前人研究结果一致[27-29]。
-
微生物是土壤与作物根际养分转化的主要过程之一[5],它生长所需要的营养元素和碳源来自于土壤,同时微生物又能够分解土壤中的有机碳/氮,使土壤碳/氮含量提高[30]。本研究结果表明,稻豆轮作提高了土壤细菌和真菌的丰富度与多样性。轮作通过改变微生物群落结构,从而影响了土壤的养分形态及产量[31]。细菌和真菌PCoA分析的结果表明,水稻连作和稻豆轮作后土壤OTU数目存在显著性差异,说明稻豆轮作显著影响土壤微生物群落结构,与张河庆等[32]的研究结果一致。一方面轮作改变了土壤微生物的生态系统,另一方面不同作物类型释放的根系分泌物,改变了土壤的环境条件,从而对土壤微生物物种和丰度产生影响,改变了土壤微生物群落的结构[5,33]。
随着轮作的进行,土壤理化指标会随着细菌群落的改变而发生变化。一方面,水旱轮作改善了土壤通气性;另一方面不同作物的根系分泌物改变了土壤的环境条件,进而对微生物活性产生了影响,间接提高土壤养分和有机碳含量,从而使土壤细菌群落也发生了改变。土壤真菌主要可以分解土壤中的植物残体和纤维素、半木纤维素、木质素等难降解有机物,在降解过程中释放养分,促进植物生长,进而影响土壤真菌[34]。土壤真菌群落对土壤全氮、有效磷、pH、WSC、MWD、产量无明显差异(P>0.05),这可能是不同作物对土壤的利用方式和对土壤性质需求不同所致[31,35]。
-
稻豆轮作能够显著提高土壤全氮、有效磷、速效钾、HWEC、SOC等,对土壤WSC和MBC影响不显著;此外稻豆轮作能够显著提高土壤细菌和真菌的丰富度、多样性与群落结构。不同耕作措施的土壤细菌和真菌的群落组成具有较大差异,说明不同耕作模式能够对土壤细菌和真菌群落组成有显著影响。土壤细菌和真菌群落主要与土壤速效钾、有机质、HWEC、MBC (P<0.01)有关。这说明稻豆轮作通过影响植物和微生物的生长代谢,使土壤养分含量和土壤微生物发生改变。本研究结果有助于理解稻豆轮作下土壤理化性质与土壤微生物群落的变化,为不同耕作模式的实际应用提供理论支撑,然而关于稻豆轮作的遗留效应还有待进一步研究。
稻豆轮作对土壤理化性质及微生物群落的影响
DOI: 10.15886/j.cnki.rdswxb.20230036
 CSTR: 32425.14.j.cnki.rdswxb.20230036
CSTR: 32425.14.j.cnki.rdswxb.20230036
Effects of rice-soybean-rice rotation on soil physicochemical properties and microbial community
-
摘要: 为了研究稻豆轮作对土壤理化性质和微生物群落结构的影响,采用常规土壤理化分析和微生物分析方法,以在海南岛东方市实施的稻-稻-稻(RRR)连作和稻-豆-稻(RSR)轮作下的土壤为研究对象,比较了水稻连作和稻豆轮作下的土壤理化性质、碳组分和微生物群落结构的差异。结果显示,RSR处理的土壤有机质、全氮含量、有效磷、速效钾、水稻产量,均高于RRR处理,说明稻豆轮作显著提高了土壤全氮、速效钾含量并降低了pH值;土壤微生物分析结果表明,RSR处理的土壤细菌ACE、Chao1和Shannon指数值比RRR处理提高了40.2%、40.3%、5.7%;土壤微生物分析结果还表明,RSR处理的土壤真菌Shannon和Simpson指数比RRR处理提高了20.1%和16.5%。土壤微生物分析结果说明,稻豆轮作对土壤微生物群落组成产生了显著影响。土壤细菌和真菌群落组成与土壤有机质、速效钾、pH、热水可提取碳(HWEC)、微生物生物量碳(MBC)呈极显著(P<0.01)相关。本研究结果表明,稻豆轮作显著提高了土壤微生物群落结构,促进了有机质分解、提高了磷和速效钾的有效性,从而提高了作物产量。Abstract: Paddy rice (Oryza sativa L.) is mainly grown thrice a year in Hainan Island. However, long-term continuous cropping and excessive nutrient input will reduce soil microbial activity and soil nutrient conversion, which is not conducive to the sustainable development of agricultural production. The soil under rice-rice-rice (RRR) continuous cropping and rice-soybean-rice (RSR) rotation systems in Dong fang City, Hainan Island was determined by using conventional soil physical and chemical analysis and microbial analysis methods, and the soil physical and chemical properties, carbon components and microbial community structure under the rice continuous cropping and the rice-soybean-rice rotation were compared to analyze the effects of the RRR and RSR on the soil physical and chemical properties and microbial community structure. The results showed that soil organic matter, total nitrogen content, available phosphorus, available potassium and rice yield under the RSR treatment were all higher than those under the RRR treatment, indicating that the RSR treatment significantly increased soil total nitrogen and available potassium content and decreased soil pH value. The soil microbial analysis showed that ACE, Chao1 and Shannon indices of the soil bacteria under the RSR treatment increased by 40.2%, 40.3% and 5.7%, respectively as compared with those under the RRR treatment. The soil microbial analysis also showed that the Shannon and Simpson indices of the soil fungi under the RSR treatment increased by 20.1% and 16.5%, respectively as compared with those under the RRR treatment. The soil microbial analysis showed that the RSR rotation had a significant effect on soil microbial community composition. The soil bacterial and fungal community compositions were significantly correlated with soil organic matter, available potassium, pH, hot water extractable carbon (HWEC) and microbial biomass carbon (MBC) (P>0.01). All the results showed that the RSR rotation significantly improved the soil microbial community structure, promoted the decomposition of organic matter, improved the availability of phosphorus and available potassium, and thus improved the crop yield.
-
Key words:
- rotation /
- soil organic matter /
- soil nutrition content /
- soil aggregate /
- soil microbe
-
表 1 稻稻连作和稻豆轮作周年产量、周年吸氮量、氮素利用率和第二季水稻产量
处理 氮投入量/(kg·hm−2) 周年产量 /(kg·hm−2) 周年吸氮量/(kg·hm−2) 氮素利用率/% 第二季水稻产量/(kg·hm−2) RRR 360 9520.6±115.5a 175.6±12.6b 48.8±3.47b 4748.5±26.3b RSR 330 7869.2±367.8b 274.2±8.10a 83.1±2.46a 5865.4±36.1a 注: 小写字母表示不同处理之间的显著差异(P <0.05)。下同  下载: 导出CSV
下载: 导出CSV
表 2 不同处理对土壤理化性质的影响
处理 有机质/(g·kg−1) 全氮/(g·kg−1) 有效磷/(mg·kg−1) 速效钾/(mg·kg−1) pH RRR 12.4±0.23b 0.49±0.06a 34.1±1.38b 95.4±0.98b 5.65±0.2a RSR 15.7±0.46a 0.57±0.03a 38.4±4.79a 109.1±9.44a 5.14±0.1b
下载: 导出CSV
表 3 不同处理对土壤水稳性团聚体的影响
mm 处理 平均质量直径(MWD) 几何平均直径(GMD) 大团聚体比例(R>0.25) RRR 0.31±0.01a 0.907±0.0001a 0.23±0.04a RSR 0.32±0.01a 0.906±0.0001a 0.24±0.03a
下载: 导出CSV
表 4 不同处理对土壤有机碳组分的影响
处理 土壤有机碳(SOC)/
(g·kg−1)热水可提取碳(HWEC)/
(mg·kg−1)水溶性碳(WSC)/
(mg·kg−1)微生物生物量碳(MBC)/
(mg·kg−1)RRR 1.41±0.03b 32.2±0.74b 8.66±1.12b 25.2±0.60b RSR 1.79±0.05a 43.7±2.14a 10.3±0.04a 34.8±2.49a
下载: 导出CSV
表 5 不同处理对土壤微生物α多样性的影响
种植方式 微生物 ACE指数 Chao1 指数 Shannon指数 Simpson指数 RRR 细菌 Bacteria 1390.8±35.2b 1386.9±34.5b 9.33±0.13b 0.99±0.0006a RSR 细菌 Bacteria 1949.6±142.1a 1945.9±142.5a 9.86±0.14a 0.98±0.0003a RRR 真菌 Fungus 265.7±44.5a 265.5±44.4a 4.03±0.33b 0.79±0.04b RSR 真菌 Fungus 328.5±42.4a 328.5±42.4a 4.84±0.25a 0.92±0.02a
下载: 导出CSV
-
[1] 岑新杰, 严小微, 唐清杰, 等. 近年来海南水稻生产情况调研及发展建议[J]. 上海农业科技, 2020(6): 18 − 19. [2] CHEN Z, TI J S, CHEN F. Soil aggregates response to tillage and residue management in a double paddy rice soil of the Southern China[J]. Nutrient Cycling in Agroecosystems, 2017, 109(2): 103 − 114. doi: 10.1007/s10705-017-9864-8 [3] 孙子欣, 蔡柏岩. 连作对土壤微生物菌群影响及修复研究进展[J]. 作物杂志, 2022(6): 7 − 13. [4] CHEN S, ZHENG X, WANG D, et al. Effect of long-term paddy-upland yearly rotations on rice (Oryza sativa) yield, soil properties, and bacteria community diversity[J]. The Scientific World Journal, 2012, 2012: 279641. [5] 杨敬林, 李琳, 胡德勇. 长期稻-稻-油菜轮作对土壤理化性质及微生物群落结构的影响[J]. 河南农业科学, 2017, 46(9): 45 − 49. doi: 10.15933/j.cnki.1004-3268.2017.09.008 [6] MIZUHIKO N I S H I D A. Decline in fertility of paddy soils induced by paddy rice and upland soybean rotation, and measures against the decline[J]. Japan Agricultural Research Quarterly:JARQ, 2016, 50(2): 87 − 94. doi: 10.6090/jarq.50.87 [7] 李娜, 韩晓增, 尤孟阳, 等. 土壤团聚体与微生物相互作用研究[J]. 生态环境学报, 2013, 22(9): 1625 − 1632. doi: 10.3969/j.issn.1674-5906.2013.09.027 [8] DOMŻŁ H, HODARA J, SŁOWIŃSKA-JURKIEWICZ A, et al. The effects of agricultural use on the structure and physical properties of three soil types[J]. Soil and Tillage Research, 1993, 27(1/2/3/4): 365 − 382. [9] 刘亚龙, 王萍, 汪景宽. 土壤团聚体的形成和稳定机制: 研究进展与展望[J]. 土壤学报, 2023, 60(3): 627 − 643. [10] 白怡婧, 刘彦伶, 李渝, 等. 长期不同轮作模式对黄壤团聚体组成及有机碳的影响[J]. 土壤, 2021, 53(1): 161 − 167. [11] 沈晓琳, 王丽丽, 赵建宁, 等. 耕作方式对潮土土壤团聚体微生物群落结构的影响[J]. 应用生态学报, 2021, 32(8): 2713 − 2721. [12] 卢欣欣, 邢茂玉, 姚陆铭, 等. 扁豆-水稻轮作对土壤环境影响的比较分析[J]. 上海农业学报, 2016, 32(5): 92 − 96. [13] 吴玉红, 王吕, 崔月贞, 等. 轮作模式及秸秆还田对水稻产量、稻米品质及土壤肥力的影响[J]. 植物营养与肥料学报, 2021, 27(11): 1926 − 1937. [14] 徐光辉, 王洋, 王继红, 等. 休耕轮作对农田土壤微生物量碳的影响[J]. 土壤通报, 2018, 49(4): 897 − 901. [15] JOERGENSEN R G. The fumigation-extraction method to estimate soil microbial biomass: Calibration of the kEC value[J]. Soil Biology and Biochemistry, 1996, 28(1): 25 − 31. doi: 10.1016/0038-0717(95)00102-6 [16] GHANI A, DEXTER M, PERROTT K W. Hot-water extractable carbon in soils: a sensitive measurement for determining impacts of fertilisation, grazing and cultivation[J]. Soil Biology and Biochemistry, 2003, 35(9): 1231 − 1243. doi: 10.1016/S0038-0717(03)00186-X [17] 黄清扬, 徐仁扣, 俞元春. 不同产地油菜秸秆制备的生物质炭对红壤酸度和土壤pH缓冲容量的影响[J]. 土壤通报, 2022, 53(4): 821 − 827. [18] 鲍士旦. 土壤农化分析 [M]. 3版. 北京: 中国农业出版社, 2000: 268−270. [19] 蒋雪洋, 张前前, 沈浩杰, 等. 生物质炭对稻田土壤团聚体稳定性和微生物群落的影响[J]. 土壤学报, 2021, 58(6): 1564 − 1573. [20] 贾雪杰, 游明鸿, 李达旭, 等. 减量施肥对金花菜-水稻轮作系统中产量和土壤养分的影响[J]. 草地学报, 2023, 31(3): 876 − 883. [21] 朱兆良, 金继运. 保障我国粮食安全的肥料问题[J]. 植物营养与肥料学报, 2013, 19(2): 259 − 273. [22] DICK W A, THAVAMANI B, CONLEY S, et al. Prediction of β-glucosidase and β-glucosaminidase activities, soil organic C, and amino sugar N in a diverse population of soils using near infrared reflectance spectroscopy[J]. Soil Biology and Biochemistry, 2013, 56: 99 − 104. doi: 10.1016/j.soilbio.2012.04.003 [23] GüNAL E, ERDEM H, DEMIRBAŞ A. Effects of three biochar types on activity of β-glucosidase enzyme in two agricultural soils of different textures[J]. Archives of Agronomy and Soil Science, 2018, 64(14): 1963 − 1974. doi: 10.1080/03650340.2018.1471205 [24] 于崧, 杨贺麟, 朱雪天, 等. 盐碱胁迫对芸豆根系分泌物组分及含量的影响[J]. 北方园艺, 2021(17): 90 − 96. [25] 陈晓东, 吴景贵, 范围, 等. 有机物料对原生盐碱土微团聚体特征及稳定性的影响[J]. 水土保持学报, 2020, 34(2): 201 − 207. [26] 温云杰. 有机培肥过程中有机碳对红壤旱地团聚体形成的影响及其分子机制 [D]. 杨凌: 西北农林科技大学, 2021. [27] SIX J, CALLEWAERT P, LENDERS S, et al. Measuring and understanding carbon storage in afforested soils by physical fractionation[J]. Soil Science Society of America Journal, 2002, 66(6): 1981 − 1987. doi: 10.2136/sssaj2002.1981 [28] DUAN Y, CHEN L, ZHANG J, et al. Long-term fertilisation reveals close associations between soil organic carbon composition and microbial traits at aggregate scales[J]. Agriculture, Ecosystems & Environment, 2021, 306: 107169. [29] YANG F, TIAN J, MEERSMANS J, et al. Functional soil organic matter fractions in response to long-term fertilization in upland and paddy systems in South China[J]. CATENA, 2018, 162: 270 − 277. doi: 10.1016/j.catena.2017.11.004 [30] 吴愉萍. 基于磷脂脂肪酸(PLFA)分析技术的土壤微生物群落结构多样性的研究 [D]. 杭州: 浙江大学, 2009. [31] 姚钦, 许艳丽, 宋洁, 等. 不同种植方式下大豆田土壤微生物磷脂脂肪酸特征[J]. 大豆科学, 2015, 34(3): 442 − 448. [32] 张河庆, 吴婕, 韩帅, 等. 4种周年轮作模式对耕作层土壤微生物的影响[J]. 中国农学通报, 2022, 38(20): 73 − 80. [33] 王秀呈. 稻—稻—绿肥长期轮作对水稻土壤及根系细菌群落的影响 [D]. 北京: 中国农业科学院, 2015. [34] 张翰林, 白娜玲, 郑宪清, 等. 秸秆还田与施肥方式对稻麦轮作土壤细菌和真菌群落结构与多样性的影响[J]. 中国生态农业学报(中英文), 2021, 29(3): 531 − 539. [35] 路妍, 高健, 康文钦, 等. 不同轮作模式下高发小麦赤霉病土壤真菌多样性变化[J]. 华北农学报, 2022, 37(6): 173 − 181. -

 点击查看大图
点击查看大图
图(3) / 表 (5)
计量
- 文章访问数: 2362
- HTML全文浏览量: 916
- PDF下载量: 55
- 被引次数: 0
