


-
沉香是瑞香科(Thymelaeaceae)沉香属(Aquilaria)植物含树脂的木材[1],广泛用于香料、医药、文玩收藏等领域。作为传统中药,沉香具有行气止痛、温中止呕、纳气平喘的功效[2],可用于治疗胃肠道疾病、咳嗽、风湿病及各种疼痛[3 − 5],现代药理学研究表明沉香提取物具有镇痛、镇静、抗菌、抗炎等广泛药理作用[6]。研究者发现,沉香中倍半萜和色酮类化合物的含量分别为沉香总量的25.6%和52.0%,特别是倍半萜类成分,其具有多种药理活性,因而被视为发挥药效的主要成分[7]。因此,在研究白木香的结香机制时,所含的倍半萜类成分及其生物合成途径一直是研究者重点关注的方向。
倍半萜合酶(terpenoid synthases, TPSs)是倍半萜生物合成途径的最后一类关键酶,可以催化底物FPP发生环化反应,最终形成一系列的倍半萜类化合物[8]。研究表明,植物倍半萜合成具有一定的时序性,白木香会在不同生长期或不同响应诱导时期合成不同的倍半萜成分,且受不同AsTPSs的催化[9]。此外,同一种倍半萜合酶可以合成一种或多种倍半萜,如As-SesTPS蛋白以FPP为底物可以生成橙花叔醇、反-(+)-橙花叔醇、β-红没药烯和蛇麻烯四种倍半萜[10 − 11]。并有研究证明过量表达倍半萜合酶基因的转基因植株能够合成积累较多的倍半萜[8,12]。
近年来,许多香农在野外陆续发现了一类不同于普通白木香树种的野生种质,将其芽条嫁接到普通树种并经过伤害诱导后可结出数量较多、质量较高的沉香,因所结沉香的性状及成分组成与古籍记载的“奇楠”非常相似,因而这类种质又被称为奇楠种质[13]。为缓解沉香资源匮乏的现状,由野生奇楠树嫁接扩繁而来的“结香早、易结香、品质好”的人工奇楠迅速崛起,在广东、广西、海南等地被广泛引种栽培[14]。目前,许多白木香倍半萜合酶基因已被成功克隆,如AsSS4[8]、TPS9、TPS12[15]等,然而对奇楠种质中倍半萜合酶基因的研究仍有不足。本研究从普通白木香和奇楠种质的转录组数据中挖掘出一条与奇楠种质结香过程关系密切的倍半萜合酶基因CNTPS18。以奇楠种质为研究对象,克隆出CNTPS18的完整CDS序列,利用生物信息学方法分析了CNTPS18的序列信息、蛋白理化性质及进化关系等;并采用qRT-PCR技术对CNTPS18基因进行伤害诱导表达模式分析。为后续研究 CNTPS18在催化倍半萜成分生物合成中的功能提供依据。
-
本研究的植物材料为中国医学科学院药用植物研究所海南分所(N20o 01',E110o 25',)的海口试验地中培育的白木香种质(包括奇楠种质与普通种质),经魏建和研究员鉴定为白木香种质。EASYspin Plus Plant RNA快速提取试剂盒(北京艾德莱生物);琼脂糖凝胶DNA回收试剂盒(北京天根生化);TransScript One-step gDNA Removal and cDNA Synthesis SuperMix Kit,pEASY Blunt Simple Cloning Kit,pEASY-T1 Cloning Vector,TransStart Top Green qPCR SuperMix,6×DNA Loading Buffer,2×Ex Taq PCR Super Mix,DNA Marker 2000,DNA Marker 10000(北京全式金生物);GoldView I(10 000×)(北京索莱宝科技);Agarose(西班牙Biowest公司);TAE溶液(50×)(合肥兰杰柯科技);硫酸卡那霉素,IPTG,X-Gal(上海麦克林生化);Yeast Extract(美国赛默飞世尔);无水乙醇(广东西陇化工);化学试剂为分析纯,引物委托生工生物合成(表1)。
表 1 基因克隆与qRT-PCR检测引物序列
Table 1. Primers sequences for gene cloning and qRT-PCR detection.
引物类型
Primer types引物名称
Primer ID引物序列
Primer sequences(5' to 3')Full-length
CDS
cloningCNTPS18-
cloningF: ATGGCGGCAGAGACGAATC
R: TTAATCAAGAGGGAGTTGGTGAACM13 F: TGTAAAACGACGGCCAGT
R: CAGGAAACAGCTATGACCCNTPS18
qRT-PCRCNTPS18-
PCRF: GCCTTGGGAACAACTGGACT
R: CCTCATGCAAGCTCAACAGCReference
geneGAPDH F: CTGGTATGGCATTCCGTGTA
R:AACCACATCCTCTTCGGTGTA -
取1 mL裂解液RLT和100 μL PLANTaid至装有样品粉末的2 mL RNase free离心管内,立即剧烈振荡混匀。混合物在冰上静置15 min左右,其间颠倒2~3次,充分裂解;裂解充分后,4 ℃条件下,
13000 r·min−1离心10 min;小心吸取上清液加到新的离心管中,加入上清液一半体积的无水乙醇,吹打混匀;取700 µL的混合液至DNA消除柱中,4 ℃条件下,13000 r·min−1空离心1 min,将DNA消除柱转入新的离心管中,加入500 μL RLTplus,13000 r·min−1离心2 min,离心1 min,去废液;4 ℃条件下,13000 r·min−1收集滤液;加入250 μL无水乙醇,并吹打混匀,将混合液加入吸附柱RA中,4 ℃,13000 r·min−1离心1 min,去废液;再加入700 µL去蛋白液Rwl和500 µL漂洗液清洗吸附柱RA,最后加入85 ℃预热的RNase-free H2O,室温放置1 min后,4 ℃,13000 r·min−1离心1 min,重复一遍此步骤即得到总RNA溶液。用1%(w/w)琼脂糖凝胶,1×TAE稀释液为缓冲液,90 V电压下电泳20~25 min,紫外灯下观察RNA的条带完整性。利用分光光度计检测RNA的浓度及质量,当2.2≥OD260/280≥2.0,OD230/260≥1时,说明样品RNA合格。 -
奇楠种质中提取的总RNA样品通过TransScript反转录试剂盒合成第一链cDNA,20 μL反应体系如下:1 μg总RNA,10 μL 2×RT Reaction buffer,1 μL gDNA Remover,1 μL oligo dT Primer mix,1 μL Starscript II RT Mix,补充ddH2O至20 μL。整个体系于42 ℃反应15 min,85 ℃热激5 s失活酶,4 ℃冷却。cDNA保存于−20 ℃备用。
-
根据白木香与奇楠种质转录组测序数据得到的白木香AsTPS18基因全长序列信息,利用Primer Premier 6软件设计引物进行实验。以上述反转录得到的cDNA为模板,以特异性引物CNTPS18-cloning为引物,进行PCR扩增。20 μL反应体系如下:模板cDNA 1 μL,上游引物和下游引物各1 μL,2×Taq PCR Master Mix 10 μL,加ddH2O至20 μL。反应程序为95 ℃预变性5 min,95 ℃变性30 s,55 ℃退火30 s,72 ℃延伸30 s,以上程序进行35个循环,72 ℃延伸5 min。PCR扩增得到的目的片段经1%(w/w)琼脂糖凝胶电泳检测后,切胶回收,连接至T载体,转化至大肠杆菌Trans T1感受态细胞中,利用蓝白斑筛选转化成功的菌体, 挑选阳性单克隆复苏后在37 ℃摇床过夜培养, 经菌落PCR验证后送菌液测序,测序工作由武汉天一华煜基因科技有限公司完成。
-
利用软件DNAman对氨基酸序列进行ORF (Open Rending Frame) 预测;使用在线软件Expasy ProtParam和Expasy ProtScale (
https://web.expasy.org/cgi-bin/protparam/ )分析预测蛋白质的基本理化性质;使用在线网站InterPro (https://www.ebi.ac.uk/interpro )分析氨基酸序列的保守结构域;使用在线分析软件Sopma对蛋白质进行二级结构预测;使用在线分析软件SWISS-MODEL进行蛋白质的三维结构预测;使用在线分析软件SignalP-6.0 Server (https://services.healthtech.dtu.dk/services/SignalP-6.0/ )分析相应信号肽;使用在线软件TMHMM 2.0对蛋白质进行跨膜结构预测;使用在线网站Wolf Psort (https://wolfpsort.hgc.jp/ )分析蛋白质的亚细胞定位;使用在线软件NetPhos3.1预测蛋白质的潜在磷酸化位点;使用软件DNAman对氨基酸序列进行多序列比对,并用MEGE 11软件建立进化树以分析不同物种间的进化关系。 -
选取奇楠种质3年生生长良好、直径为(1.5±0.2) cm的侧枝, 5 mm钻头进行钻孔伤害处理,钻孔贯通茎干,孔与孔间距2 cm。在钻孔处理后的0、0.25、1、7、15和30 d收取样本,每个时间点取3个生物学重复,从茎干剪取包括3个孔的长8 cm的茎干,剥皮后迅速浸入液氮中,−80 ℃保存备用。从奇楠种质样品中提取总RNA并反转录合成第一链cDNA。以上述样品cDNA为模板,选择表达量相对稳定的管家基因GAPDH基因为内参基因,0 d样本为对照,在实时荧光定量PCR仪(LC 96,Roch)进行qRT-PCR检测,引物信息见表1。预计GAPDH基因扩增片段大小在200 bp左右,目的基因 CNTPS18扩增片段大小为250 bp左右。反应程序为95 ℃预变性10 min;95 ℃变性10 s,58 ℃退火15 s,72 ℃延伸20 s,40个循环后利用熔解曲线检验扩增产物的特异性。采用2−ΔΔCt法计算CNTPS18基因相对表达水平[16],并使用t检验比较数据间的差异。
-
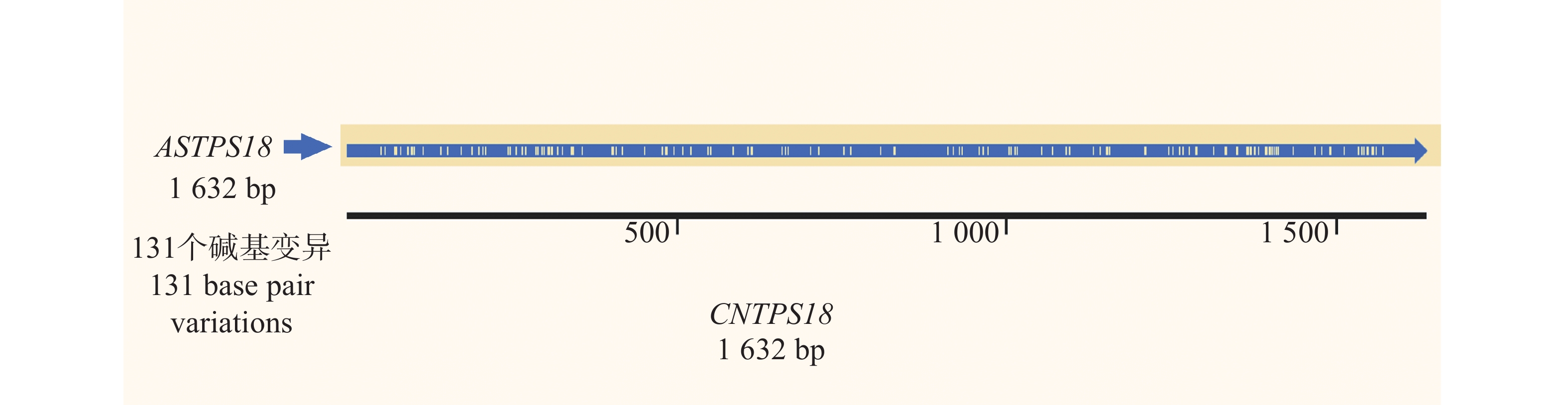
从白木香基因组中提取得到AsTPS18基因序列,依据该序列在奇楠种质中进行克隆,成功扩增出一条长为
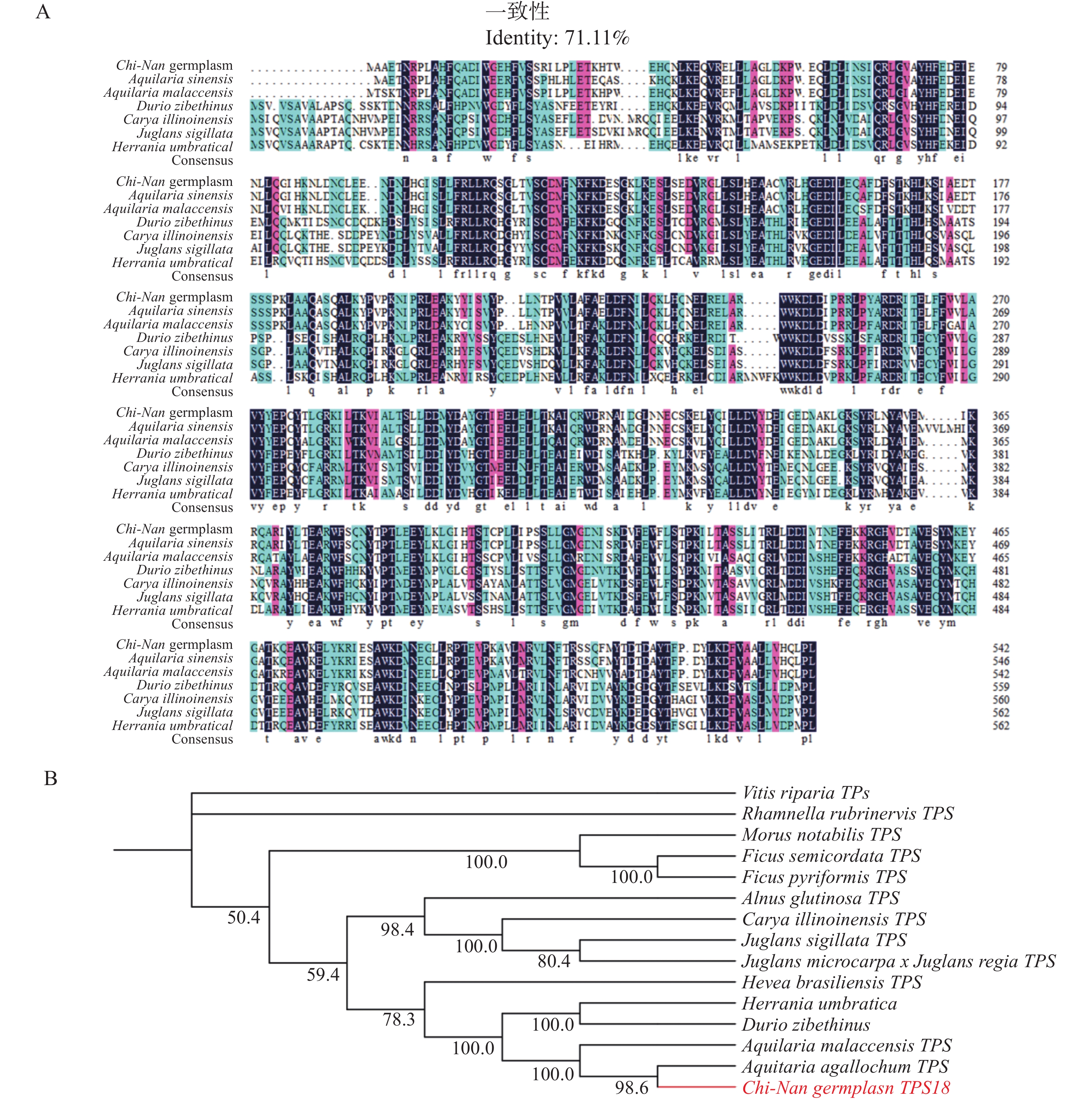
1632 bp的基因序列(基因号:PV764648)(图1),经测序验证确认序列与白木香中AsTPS18基因序列一致,一致率为92.98%。序列对比后发现,CNTPS18相较于普通白木香种质中的TPS18序列,共有131个单碱基变异,其中,59个为同义突变,62个为非同义突变,这些突变有可能导致CNTPS18与AsTPS18编码蛋白的酶活性和催化产物的不同(图2)。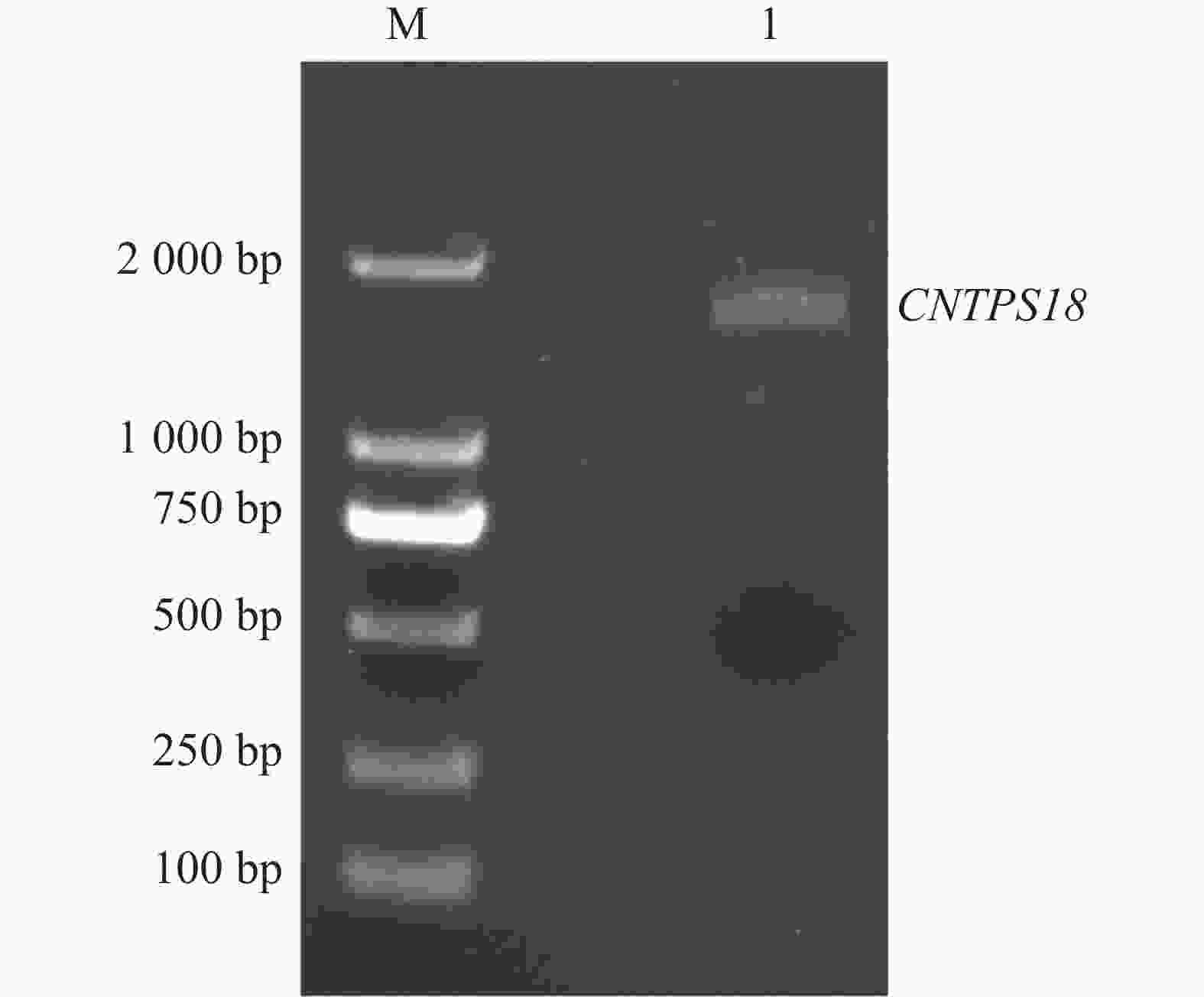
图 1 CNTPS18全长序列电泳图
Figure 1. Electropherogram of the full-length sequence of CNTPS18
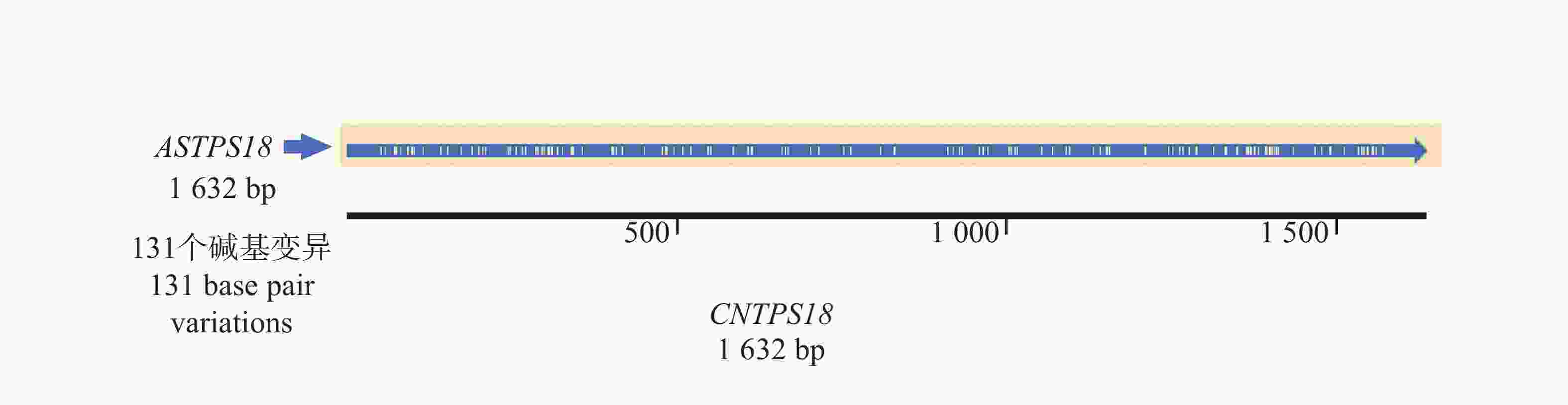
图 2 CNTPS18与普通白木香AsTPS18全长序列对比
Figure 2. Comparison of the full-length sequence of CNTPS18 with that of ASTPS18 of ordinary Aquilaria sinensis
-
ORF预测显示CNTPS18序列是一个完整的开放阅读框,包含543个氨基酸(图3)。蛋白质理化性质分析表明,该蛋白分子式为C
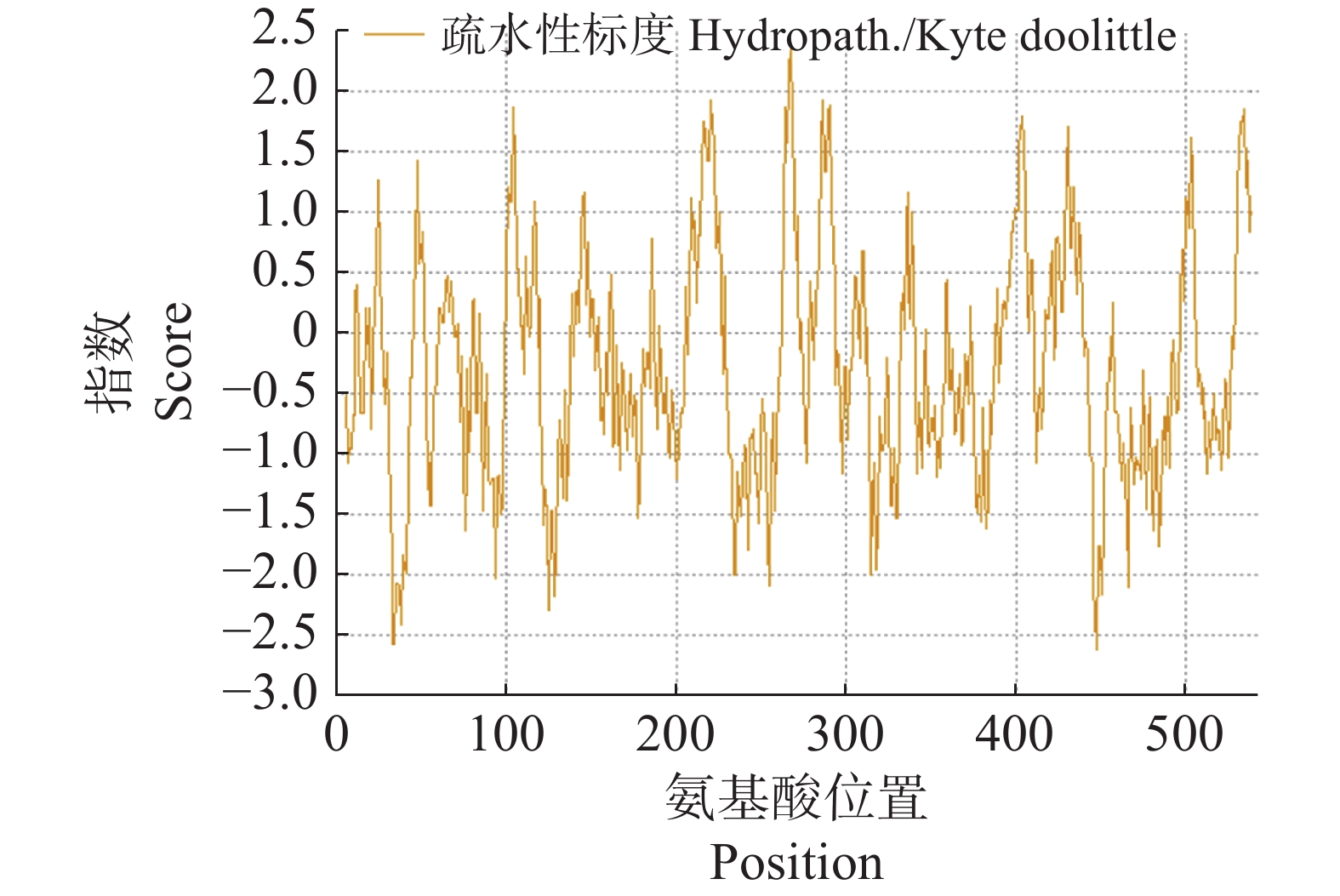
2843 H4447 N741O832S17,蛋白质的分子质量为62.865 KDa,等电点为5.36;蛋白带有81个负电荷残基和63个正电荷残基,在大肠杆菌中的半衰期大于10 h;蛋白不稳定系数为41.93,属于不稳定性蛋白。Expasy ProtScale预测显示,该蛋白的亲疏水指数范围为−2.622至2.356,总平均亲水性为−0.292,序列在第267位表现出较强的疏水性,在第448位表现出较强的亲水性,属于亲水性蛋白(图4)。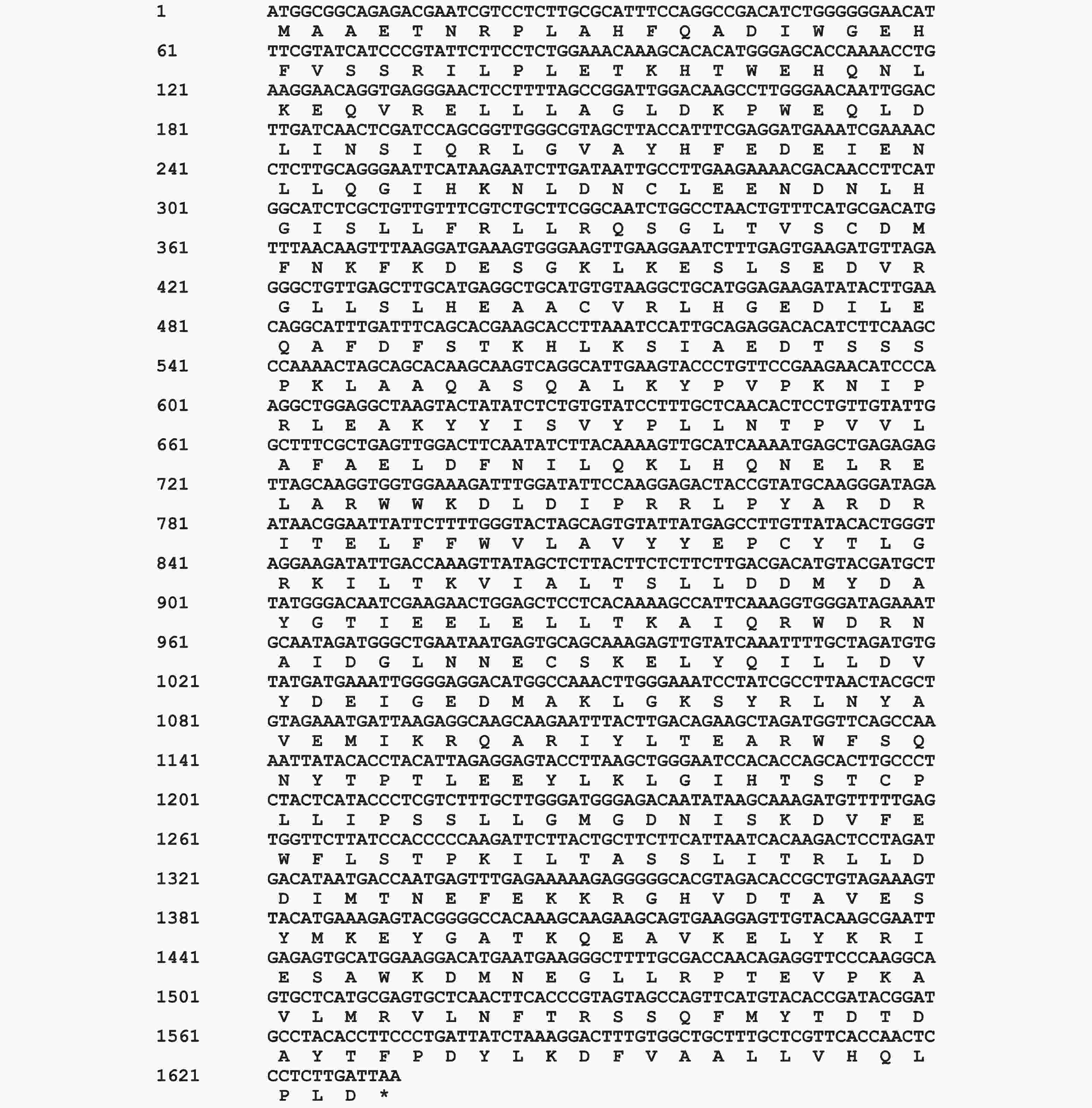
图 3 CNTPS18基因cDNA全长及预测的氨基酸序列
Figure 3. The sequences of CNTPS18 cDNA full-length and predicted amino acid
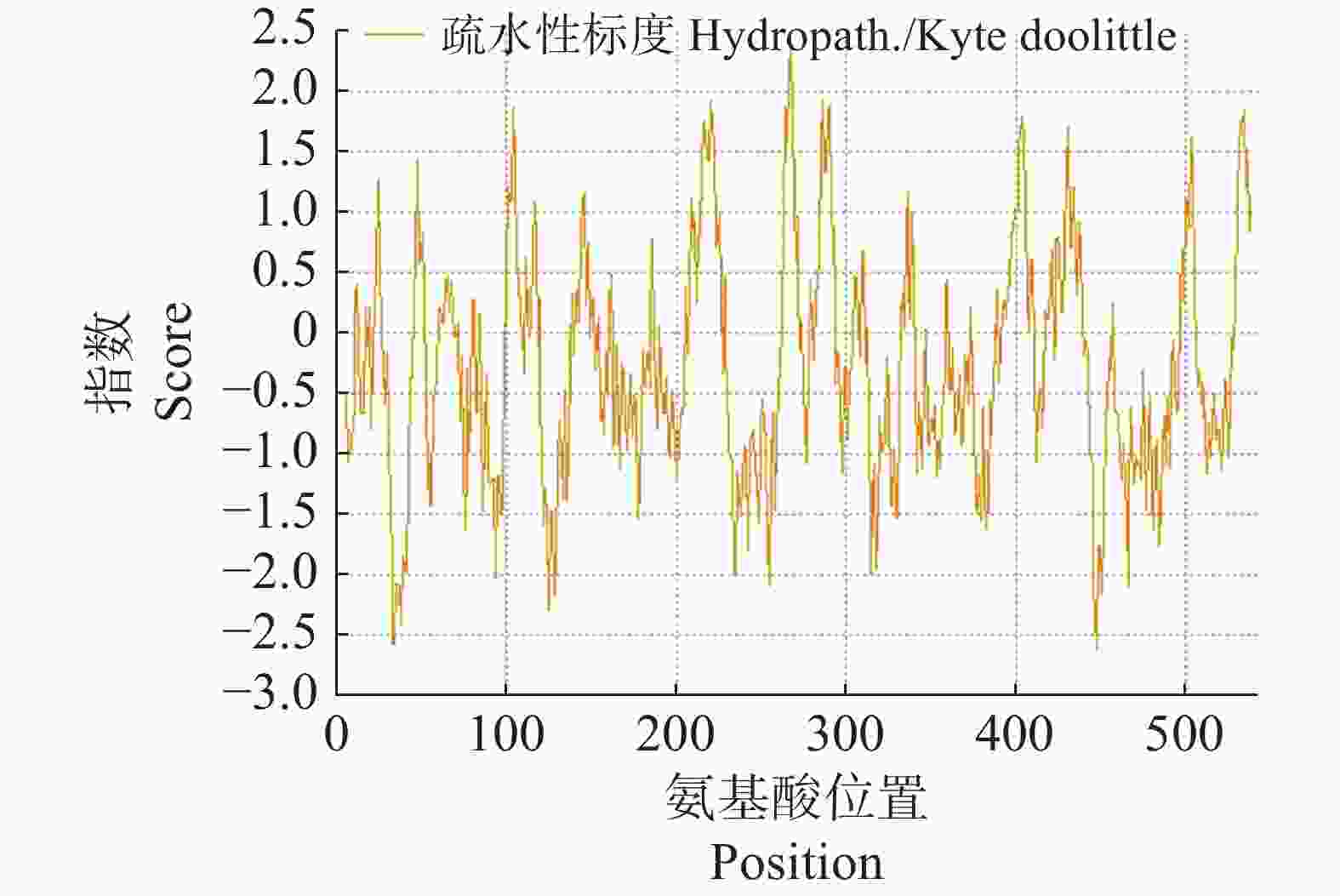
图 4 CNTPS18编码蛋白亲疏水性分析
Figure 4. The hydrophilic-hydrophobicity analysis of CNTPS18 encoded protein
-
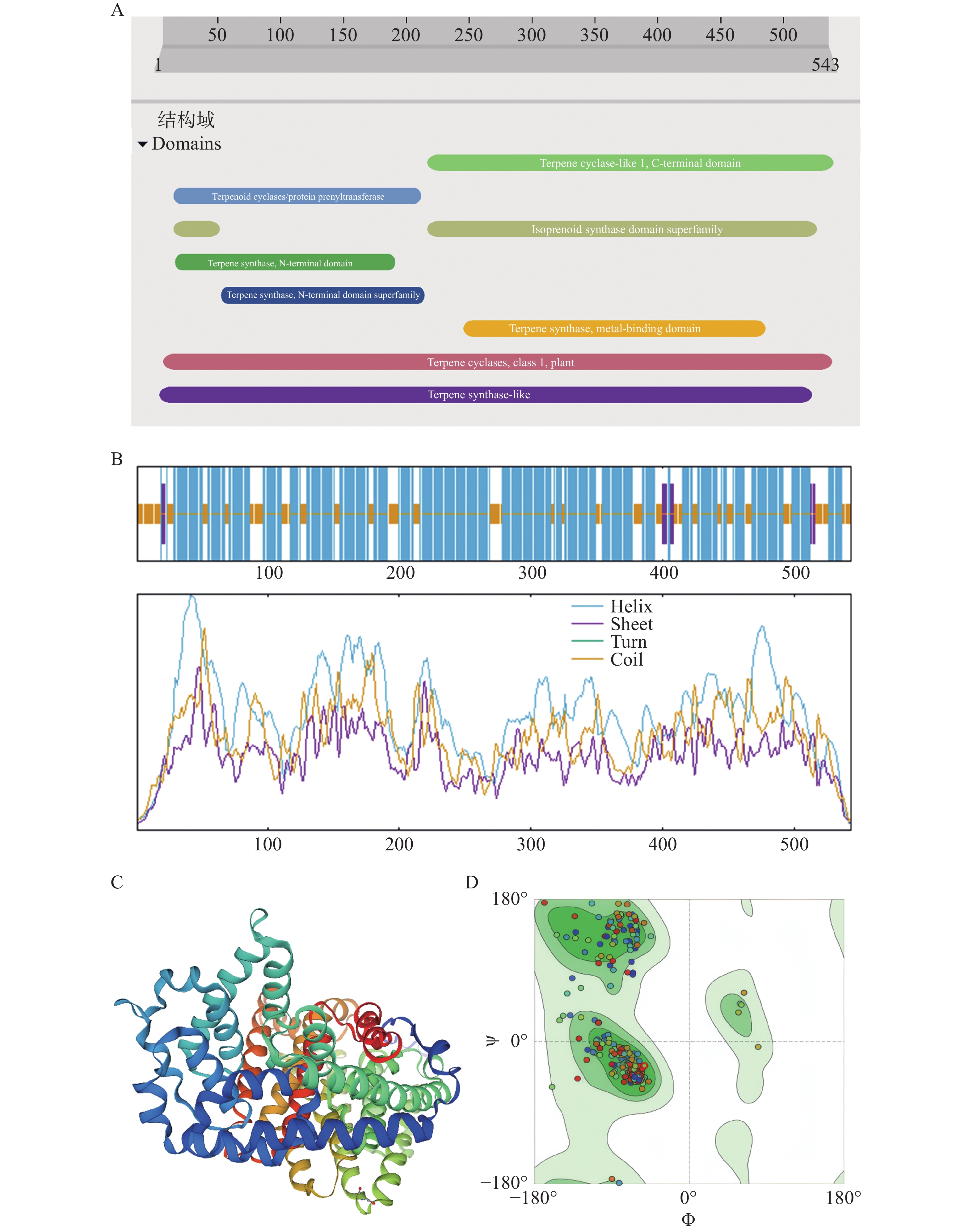
InterProScan预测该蛋白的保守结构域有Terpenoid cyclases/protein prenyltransferase(15~212),Terpene synthase, N-terminal domain(16~191),Isoprenoid synthase domain superfamily(15~52,216~526),Terpene synthase, N-terminal domain superfamily(53~215),Terpene synthase family,metal binding domain(246~486),Terpene cyclase-like 1, C-terminal domain(217~540), Terpene synthase-like(4~523), Terpene cyclases, class 1, plant(7~539 )(图5−A),表明该蛋白是一个高度保守的酶蛋白,证实该蛋白属于萜烯合成酶/环化酶家族。Sopma预测该蛋白二级结构主要含有α螺旋、无规则卷曲、延伸链。其中,α螺旋含量最高,有70.53%;无规则卷曲次之,有27.07%。α螺旋和无规则卷曲在整个氨基酸序列都有分布,延伸链穿插在序列的前后半段部分(图5−B)。CNTPS18编码蛋白的三级结构结果显示,CNTPS18蛋白与A0A411G8M5.1.A蛋白Vetispiradiene synthase一致性85.27%,GMQE值为0.94(图5−C),建模可信度较高(图5−D)。
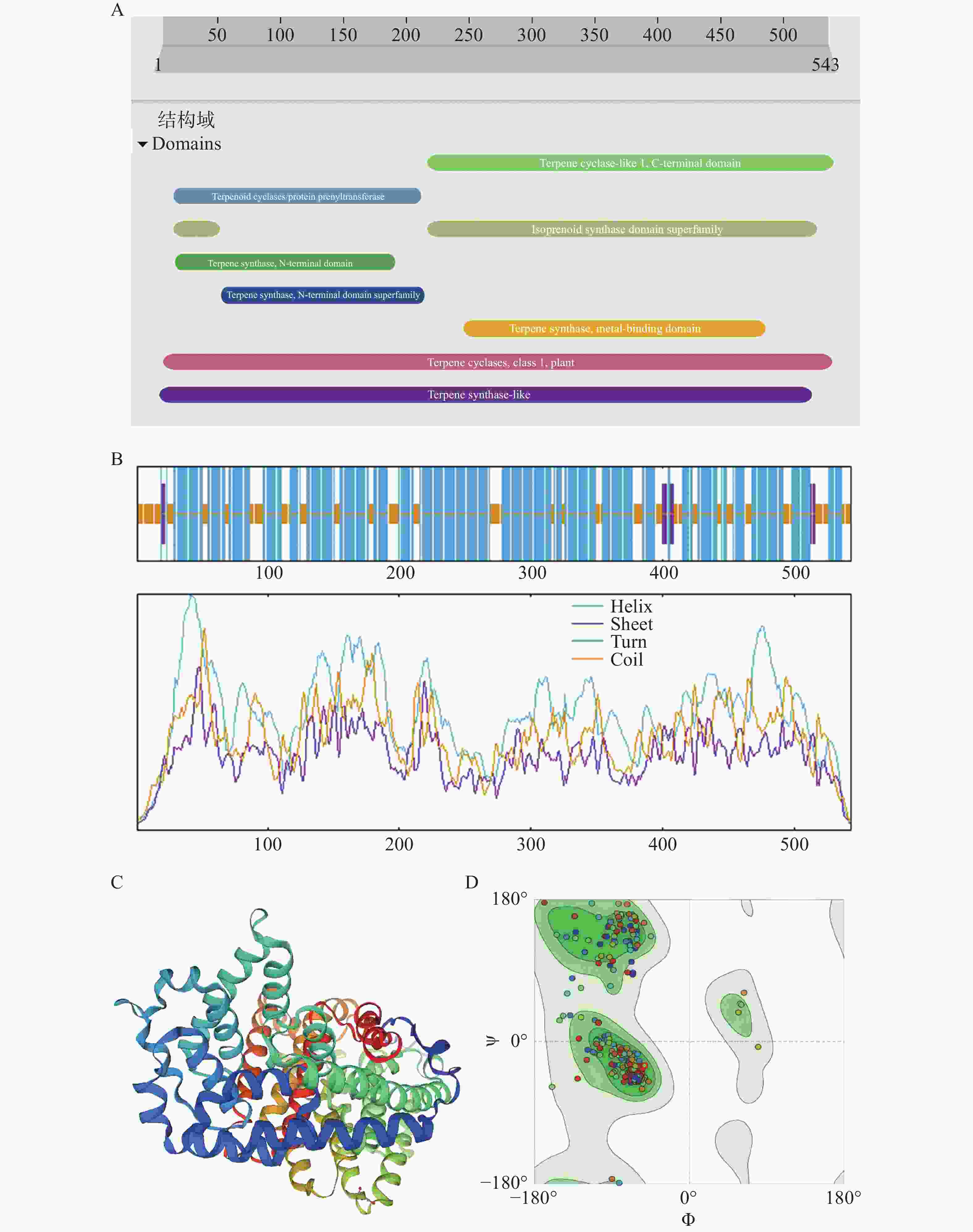
图 5 CNTPS18编码蛋白的保守结构域与结构预测
Figure 5. Conserved structural domains and structure prediction of CNTPS18 encoded protein
-
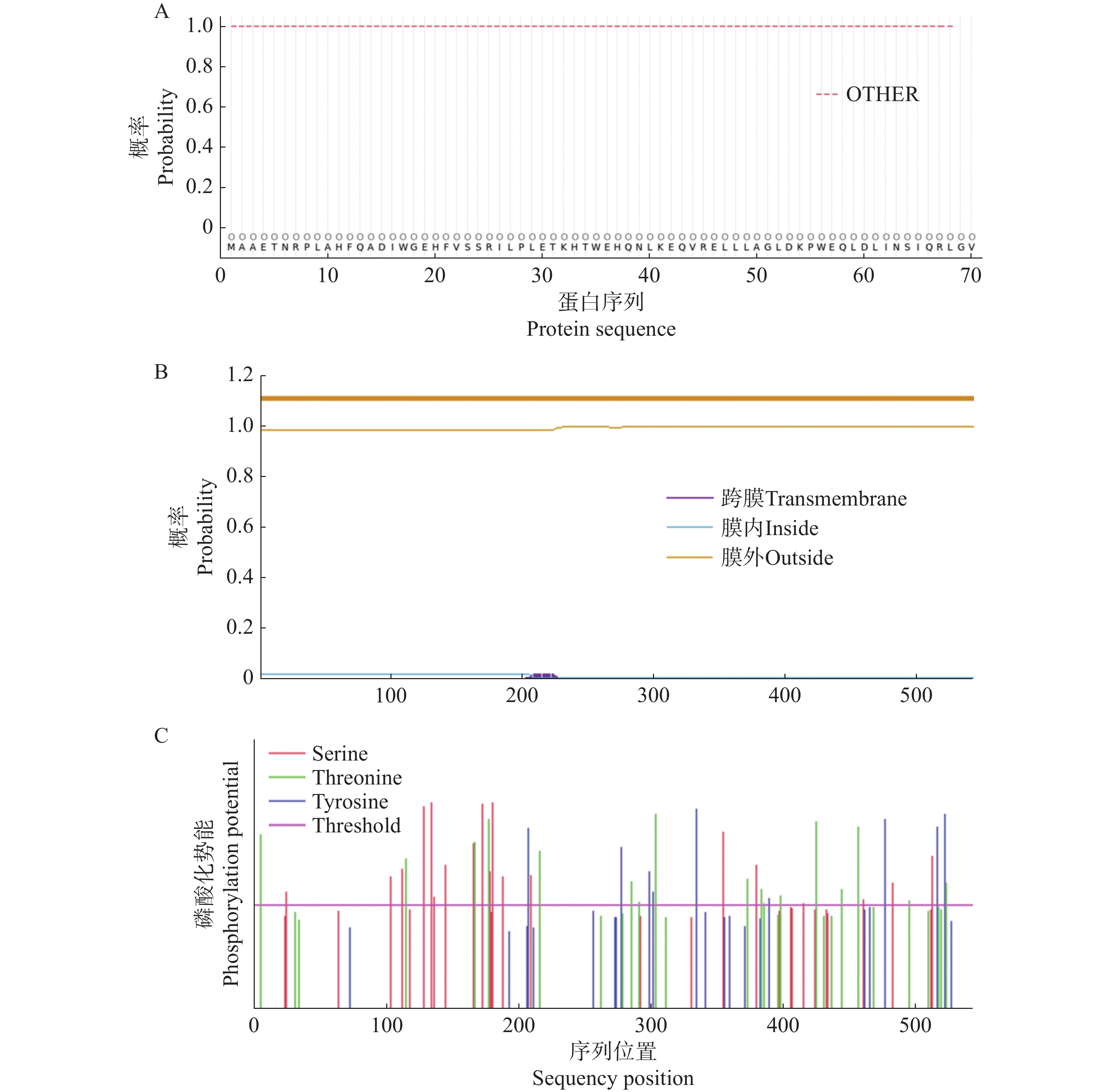
信号肽预测结果表明,CNTPS18蛋白不存在潜在的信号肽剪切位点,为非分泌蛋白(图6−A);跨膜结构预测结果表明,该蛋白无跨膜结构(图6−B);亚细胞定位预测该蛋白主要分布在细胞质基质(cyto=9)中,可能在细胞核(nucl=3)中也有少量分布,推测其可能为胞内蛋白。磷酸化位点在线预测显示 CNTPS18蛋白具有9个酪氨酸(Tyr)、17个苏氨酸(Thr)和19个丝氨酸(Ser)潜在磷酸化位点(图6−C)。
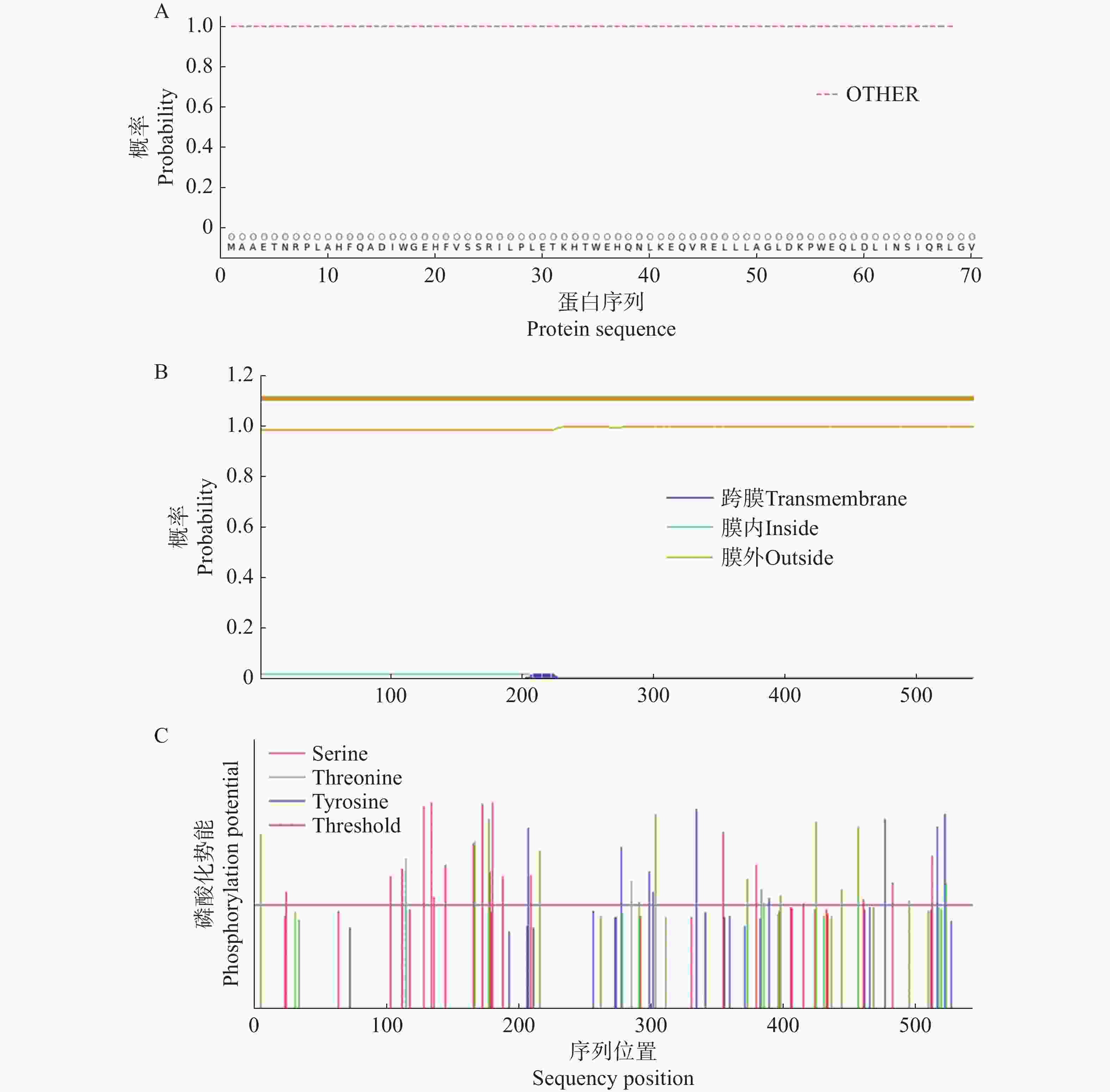
图 6 CNTPS18编码蛋白的信号肽、跨膜结构及磷酸化位点
Figure 6. The signal peptide, transmembrane structure and phosphorylation site of CNTPS18 encoded protein
-
将CNTPS18氨基酸序列进行多序列比对(图7−A),结果发现,CNTPS18与马来沉香(Aquilaria malaccensis)、榴莲(Durio zibethinus)、美国山核桃(Carya illinoinensis)、泡核桃(Juglans sigillata)和哥伦比亚锦葵(Herrania umbratical)等物种相似,其序列一致性均在71.11%以上。系统发育树显示,CNTPS18与马来沉香(Aquilaria malaccensis)和Aquilaria agallochum的倍半萜合酶、榴莲(Durio zibethinus)和哥伦比亚锦葵(Herrania umbratica)的反式大根香叶烯D合酶、巴西橡胶树(Hevea brasiliensis)的萜类合酶聚为一支,可见这些酶蛋白高度同源,亲缘关系更近。
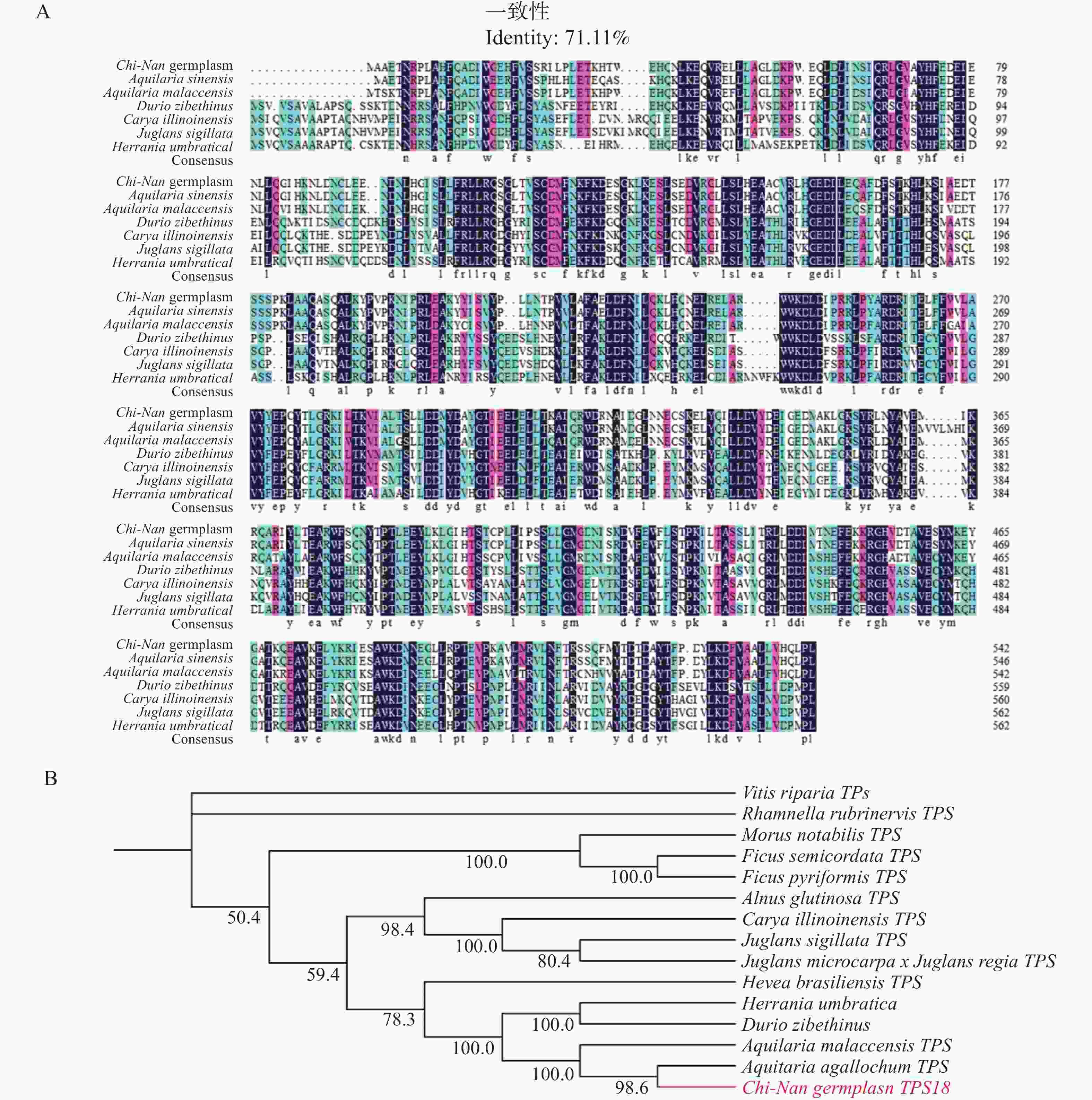
图 7 CNTPS18的氨基酸多序列比对和系统进化树图谱
Figure 7. Aligment of amino acid sequences and phylogenetic tree of CNTPS18
-
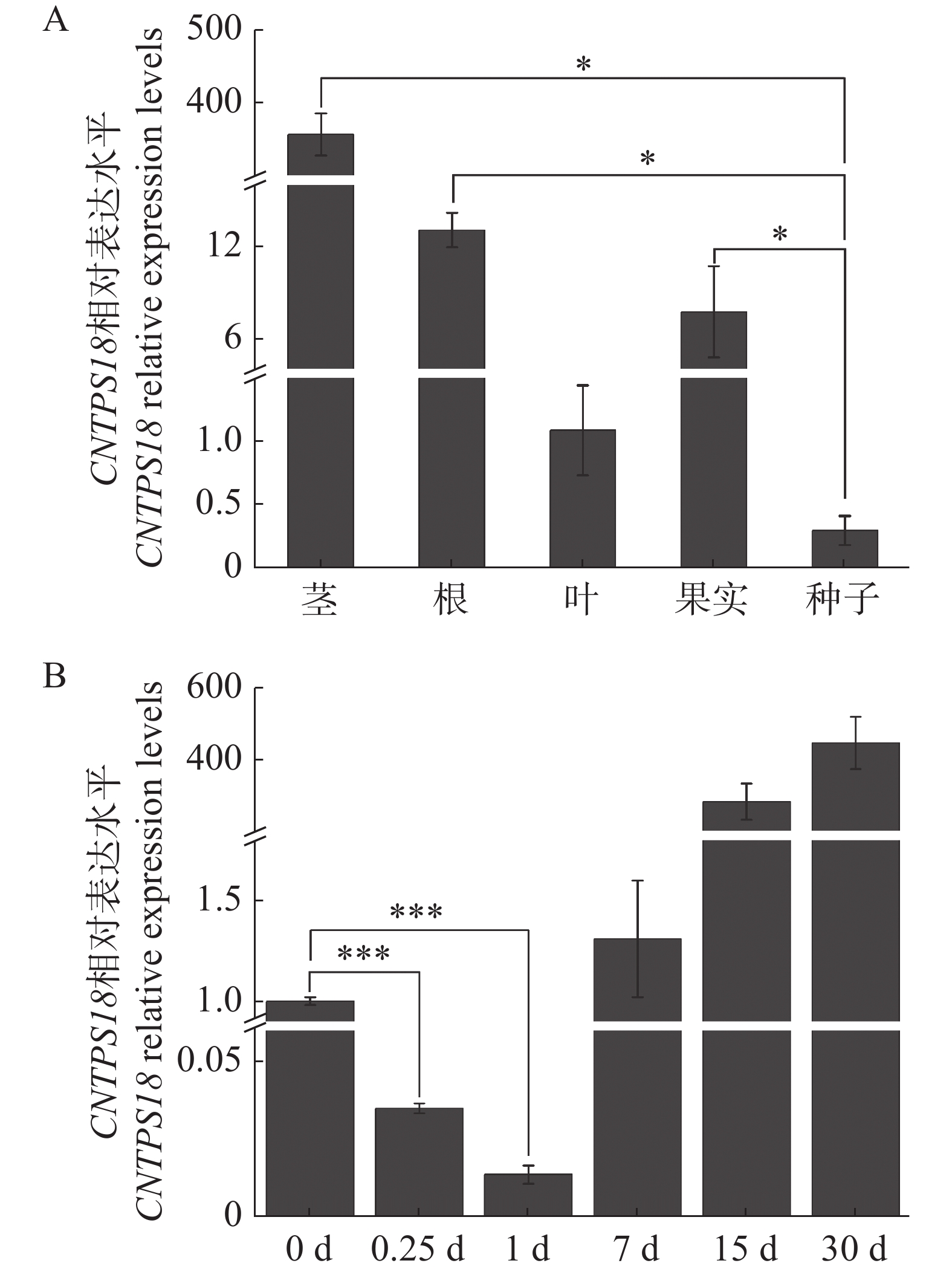
实时荧光定量PCR技术检测CNTPS18在奇楠种质和普通白木香伤害诱导表达特征。研究发现,CNTPS18在所有组织中都有表达,但表达水平各不相同。CNTPS18在茎中的表达水平最高,其次是根和果实,最低的是叶和种子(图8−A),这表明CNTPS18可能在沉香形成过程中起到了关键作用。通过检测奇楠种质伤害诱导后CNTPS18的表达水平,结果表明,CNTPS18在奇楠种质伤害诱导结香15 d后高度表达,协助奇楠种质稳定积累次生代谢产物抵御伤害胁迫(图8−B)。
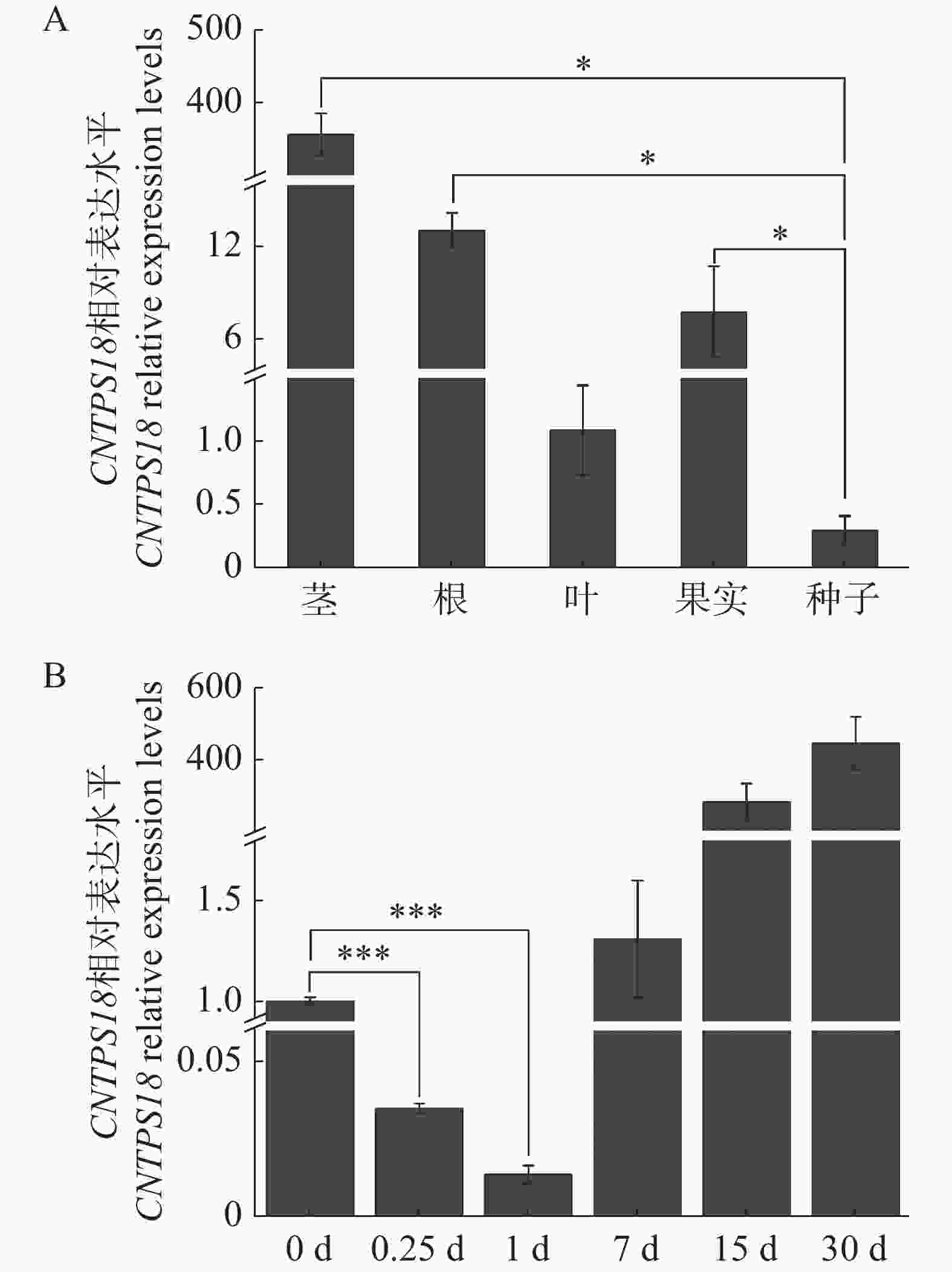
图 8 CNTPS18的组织特异性和伤害诱导表达
Figure 8. Tissue specific expression and injured induction expression characteristic of CNTPS18.
-
倍半萜是植物的一种重要的次生代谢产物,可以协助植物抵御生物或非生物的外界胁迫[17]。而在整个倍半萜类成分的生物合成途径中,倍半萜合酶的参与至关重要[15]。国内外研究者往往聚焦于农作物、水果和药用植物中倍半萜合酶基因的分子克隆和功能表征,以探索其在植物逆境胁迫下的功能和作用机制[17]。
一般认为,植物萜类合酶分为Tpsa-Tpsg7个亚家族[18]。倍半萜合酶主要属于Tpsa亚族,其他亚族较少分布。Li等[19]对白木香中的TPS基因家族进行了鉴定,共获得了分布在5个亚组中的26条TPS基因,其中有16个成员分布在TPSa,占家族成员的半数以上,这表明沉香中萜类成分主要以倍半萜为主。因此,本研究克隆了一条奇楠种质结香过程中重要的倍半萜合酶CNTPS18。该蛋白的理化性质分析表明其为胞内酶蛋白,在细胞质基质中发挥作用,与沉香木质部细胞内充满油脂的显微观察现象互相印证。保守结构域与系统发育树分析显示,CNTPS18蛋白序列高度保守,其与多种同源的植物倍半萜合酶属于Tpsa亚族,序列一致性很高;同时结构预测结果表明其与一种螺环二烯合成酶Vetispiradiene synthase结构高度吻合,推测其具有环化萜烯化合物的功能,主要合成双环倍半萜。另外,相比于AsTPS18,CNTPS18序列具有62个非同义突变,可能导致了TPS18蛋白在不同种质间的活性差异,但其保守结构域与三级结构等预测结果并未表现出差异,后续可以通过体外酶催化等实验,结合倍半萜生物合成通路,进一步探究奇楠种质产生高品质沉香过程中的分子调控机制。
萜类化合物在植物生长、发育和胁迫响应等生理过程中具有关键作用,尤其是其在植物对生物与非生物胁迫的防御机制的广泛参与,是植物对抗逆境胁迫的常用手段[20]。Yu等[21]在铁皮石斛中发现了涉及4个亚科(TPS-a,TPS-b,TPS-c和TPS-e/f)的34个DoTPS基因,其中大多数基因主要在花中表达,其次是根和茎。受到寒冷和渗透胁迫后,大多数DoTPS基因的表达均得到增强。Zhao等[22]鉴定并命名了153个小麦的TaTPS基因,转录组得出的表达谱表明,小麦对害虫(如蚜虫)的防御反应可以启动TaTPSs基因过表达,且主要通过激素途径。奇楠种质作为白木香一种新化学型种质,因其结香早、结香快、结香质量高的特性而被种植者青睐,其更优良结香性能的分子作用机制也逐渐被关注和挖掘。目前的研究已经证实,沉香特征成分生物合成相关通路的基因表达在高质量奇楠沉香形成过程中具有重要作用[23]。本研究通过检测奇楠种质各组织中CNTPS18的表达水平,并对伤害诱导30 d的奇楠种质进行qRT-PCR检测,发现CNTPS18在沉香形成部位(根和茎)高度表达,显著超过其他部位组织,且与其他作物或药用植物的表达模式相近,主要在防御反应中后期(15 d之后)开始大量积累倍半萜类物质,推测其在沉香形成过程中发挥重要作用。
本研究通过对奇楠种质CNTPS18和普通白木香种质AsTPS18基因的序列比对和生物信息学分析,为后期研究奇楠种质结香过程中的转录因子调控机制,解析倍半萜合酶基因的功能奠定基础。同时,本研究结果将为深入研究奇楠种质与普通白木香结香性能差异调控机制,以及今后高品质沉香的生产提供理论依据。
白木香奇楠种质CNTPS18基因克隆及伤害诱导表达分析
DOI: 10.15886/j.cnki.rdswxb.20250078
 CSTR: 32425.14.j.cnki.rdswxb.20250078
CSTR: 32425.14.j.cnki.rdswxb.20250078
Gene cloning and injury-induced expression pattern analysis of CNTPS18 gene in Chi-Nan germplasms (Aquilaria sinensis)
-
摘要: 奇楠种质是白木香[Aquilaria sinensis (Lour.) Spreng.]的一种新化学型种质,具有结香快和质量高的优良特性。倍半萜合酶是沉香特征成分倍半萜生物合成的关键酶,为了探究倍半萜合酶基因在高品质奇楠沉香形成过程中的作用,本研究成功克隆了奇楠种质中重要的倍半萜合酶基因CNTPS18,对其序列进行了生物信息学分析并检测了CNTPS18基因在奇楠种质受到伤害诱导后的表达模式。研究结果显示,该基因CDS全长
1632 bp,其编码蛋白主要分布在细胞质基质中,无跨膜结构,具有不稳定性和亲水性,属于非分泌蛋白;CNTPS18蛋白具有Terpene synthase-like、Terpene cyclases、class 1、plant等8个倍半萜合酶的保守结构域,三级结构与Vetispiradiene synthase的一致性很高; CNTPS18在进化上与同属的马来沉香和Aquilaria agallochum的TPS基因最为接近,与木棉亚科的榴莲和锦葵亚科的哥伦比亚锦葵处于同一分支,显示出同为锦葵目下的亲缘关系;CNTPS18在茎中的表达水平最高,其次是根和果实,最低的是叶和种子。同时,CNTPS18在奇楠种质伤害诱导结香15 d以后高度表达,推测其协助奇楠种质稳定积累次生代谢产物以抵御伤害胁迫。本研究的开展为探索奇楠种质结香机制和进一步分析CNTPSs的功能提供了一定的分子生物学依据。Abstract: As a new chemotypic germplasm of Aquilaria sinensis (Lour.) Spreng., Chi-Nan germplasm has the excellent characteristics of the faster agarwood formation and higher quality of agarwood formation. In this study we successfully cloned the CNTPS18, an important sesquiterpene synthase (TPS) gene in the Chi-Nan germplasm, and analyzed its sequence bioinformatically and detected the expression pattern of the CNTPS18 gene in Chi-Nan germplasm within induction by injury. The results showed that the full-length sequence of the CDS of this gene is1632 bp, and that its encoded protein is mainly distributed in the cytoplasmic matrix, has no transmembrane structure, is unstable and hydrophilic, and belongs to non-secretory protein. The CNTPS18 gene possessed the conserved structural domains of eight sesquiterpene synthases such as terpene synthase-like, terpene cyclases, class 1, plant, and the tertiary structure is in high identity with vetispiradiene synthase; the CNTPS18 is evolutionarily closest to the TPS genes from the same genus of Aquilaria (A. malaccensis and A. agallochum), and is in the same branch with Durio zibethinus of Bomboideae and Herrania umbratica of Malvaceae, indicating a kinship under the same malvales order; CNTPS18 was expressed at the highest level in stems, followed by roots and fruits, and at the lowest level in leaves and seeds. Meanwhile, CNTPS18 was highly expressed in Chi-Nan germplasm after 15 d of injury induction, which assisted Chi-Nan germplasm in stabilizing the accumulation of secondary metabolites against injury stress. The present study provides some molecular biological basis for exploring the mechanism of agarwood formation in the Chi-Nan germplasm and further analyzing the function of CNTPSs.-
Key words:
- Aquilaria sinensis /
- Chi-Nan germplasm /
- CNTPS18 /
- gene cloning /
- gene expression
-
图 2 CNTPS18与普通白木香AsTPS18全长序列对比
Fig. 2 Comparison of the full-length sequence of CNTPS18 with that of ASTPS18 of ordinary Aquilaria sinensis
图 3 CNTPS18基因cDNA全长及预测的氨基酸序列
Fig. 3 The sequences of CNTPS18 cDNA full-length and predicted amino acid
图 4 CNTPS18编码蛋白亲疏水性分析
Fig. 4 The hydrophilic-hydrophobicity analysis of CNTPS18 encoded protein
图 5 CNTPS18编码蛋白的保守结构域与结构预测
A. CNTPS18编码蛋白的保守结构域;B. CNTPS18编码蛋白二级结构;C. CNTPS18编码蛋白的三维空间结构;D. 拉式图。
Fig. 5 Conserved structural domains and structure prediction of CNTPS18 encoded protein
A. Conserved structural domains prediction of CNTPS18 encoded protein; B. Predicted CNTPS18 encoded protein Secondary Structure; C. Predicted three-dimensional spatial structure of CNTPS18 encoded protein;D. Ramachandran Plots.
图 6 CNTPS18编码蛋白的信号肽、跨膜结构及磷酸化位点
A. CNTPS18编码蛋白信号肽预测;B. CNTPS18编码蛋白跨膜结构预测;C. CNTPS18编码蛋白磷酸位点预测。
Fig. 6 The signal peptide, transmembrane structure and phosphorylation site of CNTPS18 encoded protein
A. Signal peptide prediction of CNTPS18 encoded protein; B. Transmembrane structure prediction of CNTPS18 encoded protein; C. Phosphoric acid site predictionprediction of CNTPS18 encoded protein.
图 7 CNTPS18的氨基酸多序列比对和系统进化树图谱
A. CNTPS18 蛋白氨基酸序列的多序列比对; B. 基于CNTPS18 蛋白氨基酸序列及其他同源序列构建的系统发育树。
Fig. 7 Aligment of amino acid sequences and phylogenetic tree of CNTPS18
A. Multiple alignment of amino acid sequences of CNTPS18 protein. B. The phylogenetic tree based on the amino acid sequence of CNTPS18 and other homologues sequences.
图 8 CNTPS18的组织特异性和伤害诱导表达
注:A.CNTPS18在奇楠种质根、茎、叶、果实和种子中的表达水平;B. CNTPS18在奇楠种质伤害诱导30 d内的表达水平。
Fig. 8 Tissue specific expression and injured induction expression characteristic of CNTPS18.
Note: A.The relative expression levels of CNTPS18 in roots, stems, leaves, fruits and seeds of Chi-Nan germplasm: B. wound-induced expression levels of CNTPS18 in Chi-Nan germplasm in 30 d.
表 1 基因克隆与qRT-PCR检测引物序列
Table 1 Primers sequences for gene cloning and qRT-PCR detection.
引物类型
Primer types引物名称
Primer ID引物序列
Primer sequences(5' to 3')Full-length
CDS
cloningCNTPS18-
cloningF: ATGGCGGCAGAGACGAATC
R: TTAATCAAGAGGGAGTTGGTGAACM13 F: TGTAAAACGACGGCCAGT
R: CAGGAAACAGCTATGACCCNTPS18
qRT-PCRCNTPS18-
PCRF: GCCTTGGGAACAACTGGACT
R: CCTCATGCAAGCTCAACAGCReference
geneGAPDH F: CTGGTATGGCATTCCGTGTA
R:AACCACATCCTCTTCGGTGTA 下载: 导出CSV
下载: 导出CSV
-
[1] CHEN X Y, LIU Y Y, YANG Y, et al. Trunk surface agarwood-inducing technique with Rigidoporus vinctus: an efficient novel method for agarwood production[J]. PLoS One, 2018, 13(6): e0198111. doi: 10.1371/journal.pone.0198111 [2] 国家药典委员会. 中华人民共和国药典-一部: 2020年版[M]. 北京: 中国医药科技出版社, 2020, 192. [3] WANG S, YU Z X, WANG C H, et al. Chemical constituents and pharmacological activity of agarwood and Aquilaria plants[J]. Molecules, 2018, 23(2): 342. doi: 10.3390/molecules23020342 [4] WANG C H, WANG S, PENG D Q, et al. Agarwood extract mitigates intestinal injury in fluorouracil-induced mice[J]. Biological and Pharmaceutical Bulletin, 2019, 42(7): 1112 − 1119. doi: 10.1248/bpb.b18-00805 [5] WANG C H, PENG D Q, LIU Y Y, et al. Agarwood alcohol extract ameliorates isoproterenol-induced myocardial ischemia by inhibiting oxidation and apoptosis[J]. Cardiology Research and Practice, 2020, 2020: 3640815. doi: 10.1155/2020/3640815 [6] 白发平, 靳若宁, 唐硕, 等. 中药沉香化学成分、药理作用及其应用研究进展[J]. 中国野生植物资源, 2022, 41(12): 61 − 66. doi: 10.3969/j.issn.1006-9690.2022.12.012 [7] 彭德乾, 王灿红, 刘洋洋, 等. 沉香的化学成分及其药理活性的研究进展[J]. 中国现代应用药学, 2021, 38(3): 358 − 365. doi: 10.13748/j.cnki.issn1007-7693.2021.03.019 [8] 梁良, 郭庆梅, 张争, 等. 白木香倍半萜合酶基因AsSS4的克隆、原核表达与功能鉴定[J]. 药学学报, 2014, 49(12): 1724 − 1729. doi: 10.16438/j.0513-4870.2014.12.008 [9] 吕菲菲, 孙佩文, 刘培卫, 等. 22个白木香倍半萜合酶(AsTPS)基因的健康与伤害诱导表达特性研究[J]. 中国现代中药, 2017, 19(8): 1076 − 1082. doi: 10.13313/j.issn.1673-4890.2017.8.005 [10] 何欣, 叶伟, 高晓霞, 等. 白木香倍半萜合成酶As-SesTPS1基因的克隆、生物信息学和表达分析[J]. 中草药, 2015,46(5): 733−739. [11] YE W, HE X, WU H Q, et al. Identification and characterization of a novel sesquiterpene synthase from Aquilaria sinensis: an important gene for agarwood formation[J]. International Journal of Biological Macromolecules, 2018, 108: 884 − 892. doi: 10.1016/j.ijbiomac.2017.10.183 [12] NAGEGOWDA D A. Plant volatile terpenoid metabolism: biosynthetic genes, transcriptional regulation and subcellular compartmentation[J]. FEBS Letters, 2010, 584(14): 2965 − 2973. doi: 10.1016/j.febslet.2010.05.045 [13] 郑卓江, 刘小金, 张启雷, 等. 13个奇楠沉香品系在广东潮州的早期生长分析[J]. 林业与环境科学, 2023, 39(2): 54 − 58. doi: 10.3969/j.issn.1006-4427.2023.02.008 [14] 杨云, 刘培卫, 康勇, 等. 中国奇楠沉香种质资源与代表性种质特征分析[J]. 福建林业科技, 2023, 50(1): 100 − 106 [15] YU C C, GAO S X, RONG M, et al. Identification and characterization of novel sesquiterpene synthases TPS9 and TPS12 from Aquilaria sinensis[J]. PeerJ, 2023, 11: e15818. doi: 10.7717/peerj.15818 [16] HABASH S S, KÖNEN P P, LOESCHCKE A, et al. The plant sesquiterpene nootkatone efficiently reduces Heterodera schachtii parasitism by activating plant defense[J]. International Journal of Molecular Sciences, 2020, 21(24): 9627. doi: 10.3390/ijms21249627 [17] LIU G H, YANG M, FU J Y. Identification and characterization of two sesquiterpene synthase genes involved in volatile-mediated defense in tea plant (Camellia sinensis)[J]. Plant Physiology and Biochemistry, 2020, 155: 650 − 657. doi: 10.1016/j.plaphy.2020.08.004 [18] JIA Q D, BROWN R, KÖLLNER T G, et al. Origin and early evolution of the plant terpene synthase family[J]. Proceedings of the National Academy of Sciences of the United States of America, 2022, 119(15): e2100361119. doi: 10.1073/pnas.2100361119 [19] LI R S, ZHU J H, GUO D, et al. Genome-wide identification and expression analysis of terpene synthase gene family in Aquilaria sinensis[J]. Plant Physiology and Biochemistry, 2021, 164: 185 − 194. doi: 10.1016/j.plaphy.2021.04.028 [20] LI H L, ZHANG X L, YANG Q L, et al. Genome-wide identification and tissue expression pattern analysis of TPS gene family in soybean (Glycine max)[J]. Frontiers in Plant Science, 2024, 15: 1487092. doi: 10.3389/fpls.2024.1487092 [21] YU Z M, ZHAO C H, ZHANG G H, et al. Genome-wide identification and expression profile of TPS gene family in Dendrobium officinale and the role of DoTPS10 in linalool biosynthesis[J]. International Journal of Molecular Sciences, 2020, 21(15): 5419. doi: 10.3390/ijms21155419 [22] ZHAO L, ZHAO X J, FRANCIS F, et al. Genome-wide identification and characterization of the TPS gene family in wheat (Triticum aestivum L. ) and expression analysis in response to aphid damage[J]. Acta Physiologiae Plantarum, 2021, 43(4): 64. doi: 10.1007/s11738-021-03236-y [23] SUN P W, LV F F, YANG Y, et al. Comparative transcriptome analysis reveals the differences in wound-induced agarwood formation between Chi-Nan and ordinary germplasm of Aquilaria sinensis[J]. Heliyon, 2024, 10(16): e35874. doi: 10.1016/j.heliyon.2024.e35874 -

 点击查看大图
点击查看大图
计量
- 文章访问数: 1167
- HTML全文浏览量: 467
- PDF下载量: 28
- 被引次数: 0
