





-
鱼类骨骼肌生长发育是一个由多种转录因子参与调控的复杂过程,其中生肌调节因子(myogenic regulatory factors,MRFs)发挥至关重要的调控作用[1]。MRFs基因家族主要包括MyoD、MyoG、Myf5和MRF4(Myf6)[2],其均编码一个高度保守的碱性螺旋-环-螺旋结构域(basic helix-loop-helix,bHLH)。bHLH结构域中HelixⅠ和HelixⅡ与由E2A基因编码的E12或E47蛋白结合形成异二聚体复合物,而basic碱性区域识别E-box(一种包含CANNTG的DNA序列)中的启动子进而与其特异性结合,从而激活骨骼肌特异性的基因表达[3-4]。因此,MRFs基因家族调控着动物体骨骼肌发育整个过程,包括肌细胞的增殖、分化及肌纤维的形成,以及个体出生后骨骼肌功能的完善[5-6]。
Chen 等[7]在斑马鱼(Danio rerio)的研究中发现,Myf5 在准体节中胚层(presomitic mesoderm,PSM)检测到表达,同时在 PSM 中未检测到其他生肌调节因子的表达,表明 Myf5 是肌发生的关键调控因子。同样的结果也出现在鲤鱼(Cyprinus carpio)[8]、牙鲆(Paralichthys olivaceus)[9]中。研究显示,在虹鳟(Oncorhynchus mykiss)和大西洋鳕鱼(Gadus morhua)的胚胎发育过程中,MyoD 基因在体节出现到原肠胚中期均检测到表达,说明MyoD在肌肉发生过程中起着重要作用[10-11]。Ganassi等[12]研究表明,MyoG基因敲除可显著抑制斑马鱼胚胎期单核肌纤维生成及肌节发育,表明其在胚胎发生过程中对肌肉分化的关键调控作用。Hinits 等[13]在斑马鱼中分离到的 MRF4 基因亚型,并在终末分化的肌肉纤维中大量表达,表明 MRF4 参与肌纤维分化过程。Schnapp等[14]通过基因敲除实验发现,MRF4能够在 MyoD 和 Myf5 基因敲除的斑马鱼中激活 MyoD 促进骨骼肌的形成。目前,生肌调节因子已在部分鱼类中被克隆。例如,鳜鱼(siniperca chuatsi)和达氏鲟(Acipenser dabryanus)中发现4种MRFs因子[15-16];MyoD 在牙鲆、黄尾鰤(Seriola lalandi)、金头鲷(Sparus aurata)和红鳍东方鲀(Takifugu rubripes)中被克隆[17-20]; MyoG分别在赤眼鳟(Squaliobarbus curriculus)、刀鲚(Coilia nasus)等中被克隆[21-22];MRF4在尼罗罗非鱼(Oreochromis niloticus)中被克隆[23]。
鞍带石斑鱼(Epinephelus lanceolatu)俗称龙趸、龙胆石斑鱼,隶属鲈形目(Perciformes)、鮨科(Serranidae)、石斑鱼属(Epinephelus),最大体长可达2 m,体质量可达400 kg,为体型最大的珊瑚礁鱼类[24]。鞍带石斑鱼是中国海南省重要的海水养殖品种,由于其生长迅速的特点,常用于与其他石斑鱼如棕点石斑鱼[25]、云纹石斑鱼[26]、驼背鲈[27]等进行杂交。本研究通过克隆鞍带石斑鱼生肌调节因子,获得其基因序列,并对其氨基酸序列进行同源性对比及进化树分析,为进一步探究生肌调节因子对骨骼肌生长发育的调控提供理论基础。
-
鞍带石斑鱼[(20.00±0.07) g,3月龄]购自海南省万宁市一养殖场。在实验开始前,将鱼置于室温(28 ℃)下的循环水槽中驯化两周。随后,挑选4尾健康的鞍带石斑鱼,用间氨基苯甲酸乙酯甲磺酸(MS-222)对其进行麻醉,在无菌操作下解剖取出鞍带石斑鱼7种不同的器官式组织,包括骨骼肌、肝脏、肾脏、肠道、胃、心脏和脂肪组织,液氮速冻后于−80 ℃冰箱中保存。
GoScript™ Reverse Transcription System试剂盒购自美国Promeg公司;FastPure® Gel DNA Extraction Mini Kit购自南京诺唯赞生物科技有限公司;DH5α感受态细胞购自上海唯地生物技术有限公司;Gold ViewI型核酸染色剂购自北京索莱宝科技有限公司;RNAiso Plus、GoScript™ Reverse Transcription System试剂盒、SMARTer® RACE5′/3′kit试剂盒、pMD19-T Vector、PrimeScript™ RT reagent kit with gDNA Eraser试剂盒、TB Green® Premix Ex Taq™ Ⅱ(Tli RNaseH Plus)均购自宝日医生物技术(北京)有限公司。
-
NCBI数据库获得斜带石斑鱼(Epinephelus coioides)、斑马鱼、鲤鱼生肌调节因子cDNA序列,根据序列保守区域设计简并引物来扩增鞍带石斑鱼MRFs基因家族中间片段。根据中间片段序列设计用于扩增3′RACE(3′-F1、3′-F2)和5′RACE(5′-R1、5′-R1)的特异性引物。将中间片段、3′RACE和5′RACE的cDNA序列拼接,设计用于验证鞍带石斑鱼生肌调节因子ORF框的特异性引物(表1),并根据克隆得到的鞍带石斑鱼5种生肌调节因子基因序列,设计用于组织分布的荧光定量PCR引物(表2)。引物由生工生物工程(上海)股份有限公司合成。
表 1 基因克隆所用引物序列
Table 1. Primer sequences for gene cloning
引物 Primer 序列(3′-5′ ) Sequence 中间片段引物序列(5′-3′)
Middle fragment primer sequences (5′-3′)MyoD1 F1 ATGCACTTCTTCGAGGAC MyoD1 R1 TGTTGTCGGTGGAGATTC MyoD2 F1 CTCTGCTGACGACCTCTA MyoD2 R1 TTACTGCTGCTGGAATCG MyoG F1 TACGACCAATCCACCTATCA MyoG R1 TCCGCCTGCTGTAGAGAT MRF4 F1 CGCTATCTGGAAGAAGGG MRF4 R1 CTCTCTGGTTTGTCATTGC Myf5 F1 ACGTCTTCTCACCATCCCAG Myf5 R1 AGAAGGTGGCCATGTCTCTC 3′RACE所用引物序列(5′-3′)
3′RACE primer sequences (5′-3′)MyoD1 3′-F1 GTCCTCCCTCCTGCACCTCCATCAC MyoD1 3′-F2 GAGCACTACAGCGGGGACTCAGACG MyoD2 3′-F1 CCCCGCTCCAACTGCTCCGATGG MyoD2 3′-F2 GCAGCCACACAGACGATTCCAGCAGC MyoG 3′-F1 CCCAACCCGACCCAACCCAGAGTGT MyoG 3′-F2 CATGCGTGCCCTGACCTCCATCGTG MRF4 3′-F1 CCAACCCCAACCAGAGGCTACCCAA MRF4 3′-F2 CGACCTCTGCCGACCATTCCACTGC Myf5 3′-F1 CGCAACGCCATCCAGTACATCGAGAGC Myf5 3′-F2 TCTGAATGCAAACTACAGCGGCGGAT 5′RACE所用引物序列(5′-3′)
5′RACE primer sequences (5′-3′)MyoD1 5′-R1 CGTGGTGATGGAGGTGCAGGAGGGA MyoD1 5′-R2 ATGGAGGTGCAGGAGGGAGG MyoD2 5′-R1 CTTACCAAGCCGCCGCCTTTCTCGC MyoD2 5′-R2 CTTTCTCGCATCGTCGCTGCTTTCC MyoG 5′-R1 CTTGGGCAGCCTCTGGTTTGGGTTC MyoG 5′-R2 GACGCCTCTTCTCCCTCATCGTGGC MRF4 5′-R1 TGGGCGCCGACTTCCTCTTGCAGAT MRF4 5′-R2 CATGAGGCACTGGCCGTCGCAG Myf5 5′-R1 TGCAGGCTCTCGATGTACTGGATGGCG Myf5 5′-R2 CTGGCTGGGGTTGGCTGAGGTGC 全长验证引物序列(5′-3′)
Full-length validation primer sequences (5′-3′)MyoD1 Full-F ACGCTTCTGTCTCTGTCACTGGAT MyoD1 Full-R ATGGTAAAGTCCCAGGTGTGAAGG MyoD2 Full-F CCCCGCTCCAACTGCTCCGATGG MyoD2 Full-R GCAGCCACACAGACGATTCCAGCAGC MyoG Full-F CCGAACACCACATACAGTAGAGC MyoG Full-R TGGAGGGTCTACTTGGGAAT MRF4 Full-F GGGCGCAACAATATGATGGACCTTT MRF4 Full-R GAATATAGTGGAAATAATTTAGGGA Myf5 Full-F ATGGAYGTCTTYTCVCCVTCCCA Myf5 Full-R TGGCGTTCGGTAAAAGCAG 接头引物序列(5′-3′)
Adapter primer sequences (5′-3′)3′ CDS AAGCAGTGGTATCAACGCAGAGTACTT
TTTTTTTTTTTTTTTTTTTTTTTTTTTTVN5′ CDS TTTTTTTTTTTTTTTTTTTTTTTTTVN NUPA AAGCAGTGGTATCAACGCAGAGT UPMA-S CTAATACGACTCACTATAGGGC UPMA-L CTAATACGACTCACTATAGGGCAAGCAG
TGGTATCAACGCAGAGT表 2 实时荧光定量PCR检测5种基因组织分布所用引物序列
Table 2. Nucleotide sequences of the primers used for Real-time PCR
引物 Primer 序列3′-5′ Sequence MyoD1 RT-F GGCATGACGGATTTCAACGG MyoD1 RT-R CTGCTGTTGTCGGTGGAGAT MyoD2 RT-F ATCATCACCACCACCACGTC MyoD2 RT-R TTCAGGGTCTCAAAGGCGTC MyoG RT-F CACGATGAGGGAGAAGAGGC MyoG RT-R TGCTGGTTGAGGGAAGACAC MRF4 RT-F CAGTGTGGCCAGTCATGAGT MRF4 RT-R TCTTCGGTGGTGATGCTGTC Myf5 RT-F GAGGAGGACGAGCATGTGAG Myf5 RT-R CTCGATGTACTGGATGGCGT EF1-α RT-F AGGGATGGAAGATTGAGCGC EF1-α RT-R CGTACCGGGCTTCAGGATAC -
使用Trizol试剂(Invitrogen,美国)提取总RNA,1.0%的琼脂糖凝胶电泳检测RNA质量,并在NanoDrop ND−1000分光光度计(NanoDrop,威尔明顿,特拉华州)上进行产率测定。RNA经脱氧核糖核酸去除酶(Takara,日本)去除DNA后,中间片段cDNA模板制备和3′端cDNA模板制备通过GoScript™ Reverse Transcription System试剂盒(Promega,美国)进行反转录;5′端cDNA模板制备采用SMARTer® RACE5′/3′kit(Takara,日本)试剂盒反转录合成;不同组织分布cDNA模板制备采用PrimeScript™ RT reagent kit with gDNA Eraser(Perfect Real Time)(Takara,日本)试剂盒进行反转录合成。
-
以所获得中间片段cDNA为模板,以引物 F1 和 R1 进行第1轮 PCR 扩增,PCR反应条件如下:94 ℃ 5 min;94 ℃ 30 s,TM值30 s,72 ℃ 60 s,35循环;72 ℃ 5 min;4 ℃ 保持。
-
以3' 端cDNA为模板,分别以鞍带石斑鱼MRFs基因家族基因正义引物3'-F1和反义接头引物UPMA(UPMA-L:UPMA-S为1﹕5混合使用)进行第1轮PCR扩增;将第1轮PCR产物稀释100倍,并取2 μL作为第2轮PCR扩增模板,以正义引物3'-F2和反义接头引物NUPA进行第2轮嵌套PCR扩增,第1轮PCR反应条件:94 ℃ 5 min;94 ℃ 30 s,TM值+5 ℃(每循环降落0.5 ℃) 30 s,72 ℃ 60 s,20循环;94 ℃ 30 s,TM值 30 s,72 ℃ 60 s,20循环;72 ℃ 7 min,4 ℃ 保持。第2轮PCR:94 ℃ 5 min;94 ℃ 30 s,TM值30 s,72 ℃ 60 s,40循环;72 ℃ 10 min,4 ℃ 保持。
-
以5'端cDNA为模板,分别以鞍带石斑鱼MRFs基因家族基因正义引物5'-R1和反义接头引物UPMA进行第1轮PCR扩增;将第1轮PCR产物稀释10倍,并分别取2 μL作为第2轮PCR扩增模板,分别以鞍带石斑鱼MRFs基因家族的正义引物3'-R2和反义接头引物NUPA进行第2轮嵌套PCR扩增,PCR反应条件同1.4.2。
-
PCR产物用1%琼脂糖凝胶电泳检测,并根据FastPure® Gel DNA Extraction Mini Kit(诺唯赞,中国)说明书切取含有目的片段凝胶,胶回收纯化PCR产物。
-
回收产物连接于pMD19-T Vector,转化到大肠杆菌(DH5α)感受态细胞中,涂在含氨苄青霉素LA平板上,挑取白色单菌落培养,进行PCR阳性克隆检测,分别取100 μL菌液送至广州天一辉远公司测序。
-
根据测序结果,将MRFs基因家族cDNA序列拼接以获得基因全长序列,利用在线ORF Finder对5个基因ORF进行预测,并在其序列两端非编码区设计特异性引物(MyoD1 Full-F,MyoD1 Full-R;MyoD2 Full-F,MyoD2 Full-R;MyoG Full-F,MyoG Full-R;MRF4 Full-F,MRF4 Full-R;Myf5 Full-F,Myf5 Full-R)进行PCR全长扩增反应,以进行全长验证。
-
鞍带石斑鱼MyoD1、MyoD2、MyoG、MRF4、Myf5 5个基因cDNA序列使用ORF Finder(
http://www.bioinformatics.org/sms2/orf_find.html )预测基因编码区;核苷酸和氨基酸序列同源性比对使用NCBI(https://www.ncbi.nlm.nih.gov/ )blast程序进行;核酸序列翻译成基酸序列通过Expasy(https://web.expasy.org/translate/ )完成;采用SWISS-MODEL(https://swissmodel.expasy.org/ )对蛋白质三级结构进行预测分析;多重氨基酸序列比对图谱使用 Clustal X和 ESPript v3.0构建;在NCBI数据库上进行比对,获得MRFs基因同源序列。利用MEGA v7.0软件中的ClustalW算法对MRFs基因的氨基酸序列进行多重序列比对,并采用Neighbor-Joining(NJ)法构建系统进化树,通过自引导检验(bootstrap)获得系统分支的置信度(重复次数为1 000)。 -
鞍带石斑鱼骨骼肌、脑、心脏、脂肪、胃等组织cDNA进行实时荧光定量 PCR 反应,根据TB Green® Premix Ex TagTM Ⅱ(Takara,日本)试剂盒说明书进行操作。实时荧光定量在定量热循环仪(Roche LightCycler® 480,瑞士)中进行。扩增体系为20 μL,其中包含10 μL 2×Taq PCR MasterMix Ⅱ反应混合液,0.5 μL的正反引物,7 μL无核酶水和2 μL cDNA模板,反应程序为95 ℃ 30 s预变性,95 ℃ 5 s,55 ℃ 30 s,72 ℃ 30 s,共40循环,95 ℃ 5 s,65 ℃ 60 s绘制溶解曲线; 50 ℃ 30 s冷却。以EF1-α 作为内参基因。用5种浓度稀释不同倍数的cDNA样品制作标准曲线(每个cDNA样品设置3次重复试验),根据公式E=10(−1/slope)−1分析扩增效率。目的基因的相对表达水平采用R=2−ΔΔCt法进行计算[28]。
-
本研究将克隆得到的基因序列上传已上传至 GenBank,登录号分别为MW762676、MW762677、MW762678、MW762679、MW762680。
-
实验数据均以平均值±标准差表示,并进行了方差齐性检验和正态性检验。使用 SPSS v22.0 软件对数据进行单因素方差分析(one-way ANOVA),使用 Duncan’s 进行多重比较分析,以 P<0.05作为差异显著性判断标准。
-
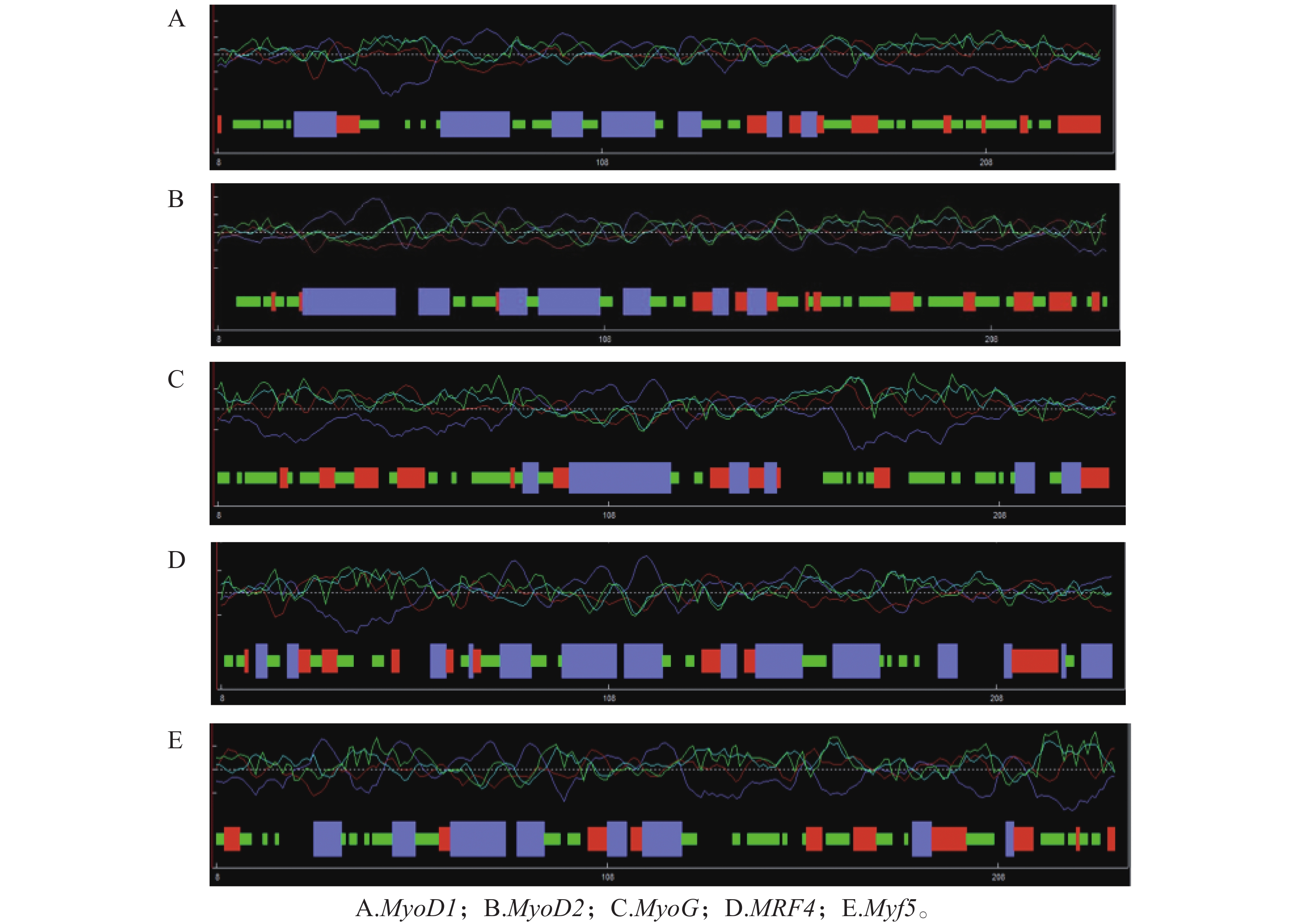
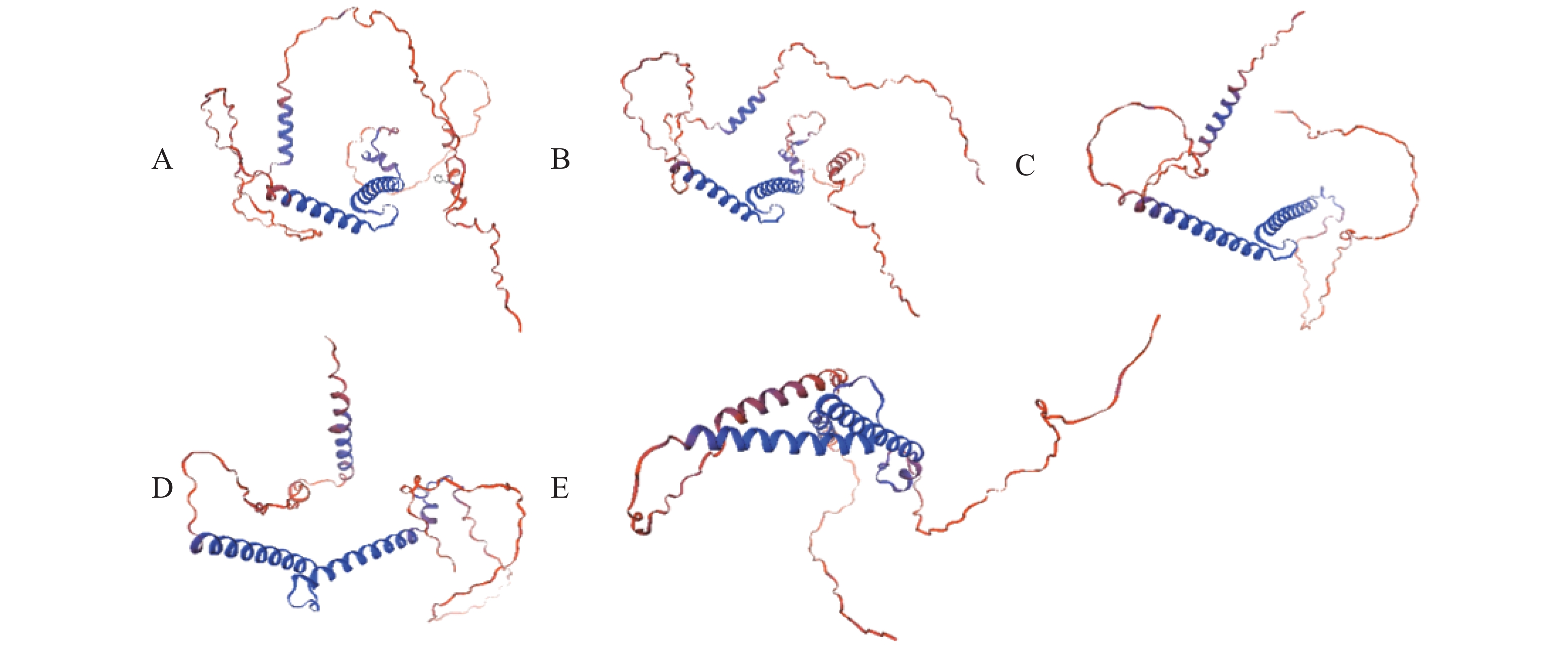
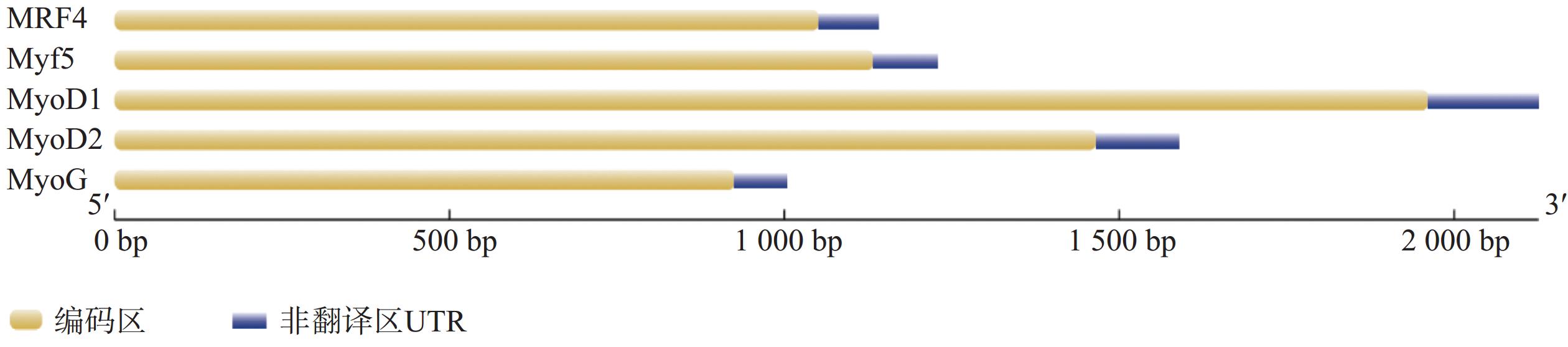
克隆获得鞍带石斑鱼MyoD1、MyoD2、MyoG、MRF4和Myf5 cDNA序列长度分别为1 962、1 466、926、1 052和1 133 bp,开放阅读框(ORF)分别为891、810、753、714 和726 bp,分别编码297、270、251、238和242个氨基酸。GenBank登录号分别为MW762676、MW762677、MW762678、MW762679和MW762680。结构分析表明,鞍带石斑鱼MRFs家族5个基因均由编码区(CDS)和非翻译区(UTR)组成(图1)。采用Garnier-Gibrat-Robson方法预测其二级结构,结果显示MyoD1、MyoD2、MyoG、MRF4和Myf5均包含多个α−螺旋和β−折叠结构域(图2)。三级结构预测进一步表明其均呈现典型的螺旋−环−螺旋(HLH)结构(图3)。
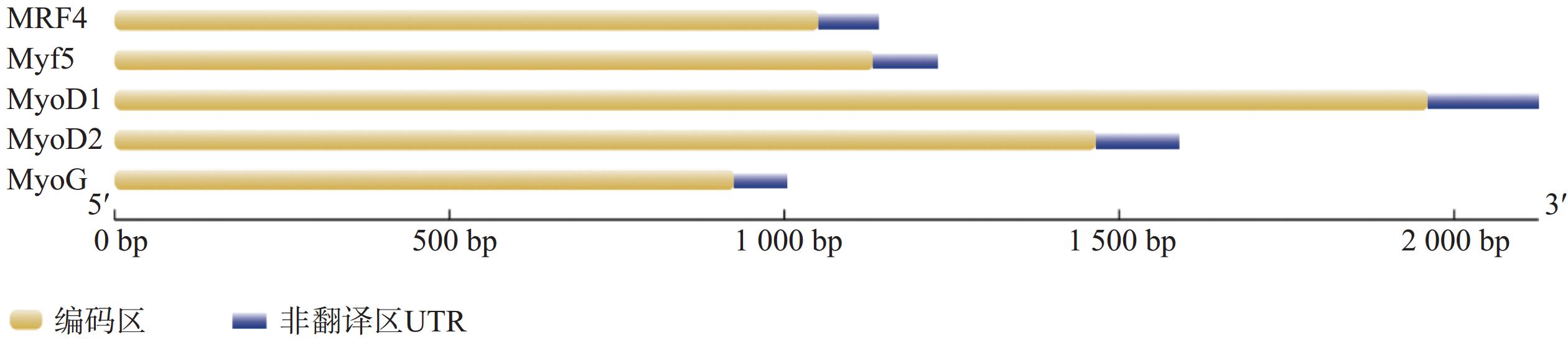
图 1 鞍带石斑鱼MRFs家族基因的结构
Figure 1. Gene structure of the Epinephelus lanceolatus MRFs family genes
图 2 鞍带石斑鱼MRFs家族蛋白二级结构预测
Figure 2. Prediction of the secondary structure of MRFs protein in Epinephelus lanceolatus
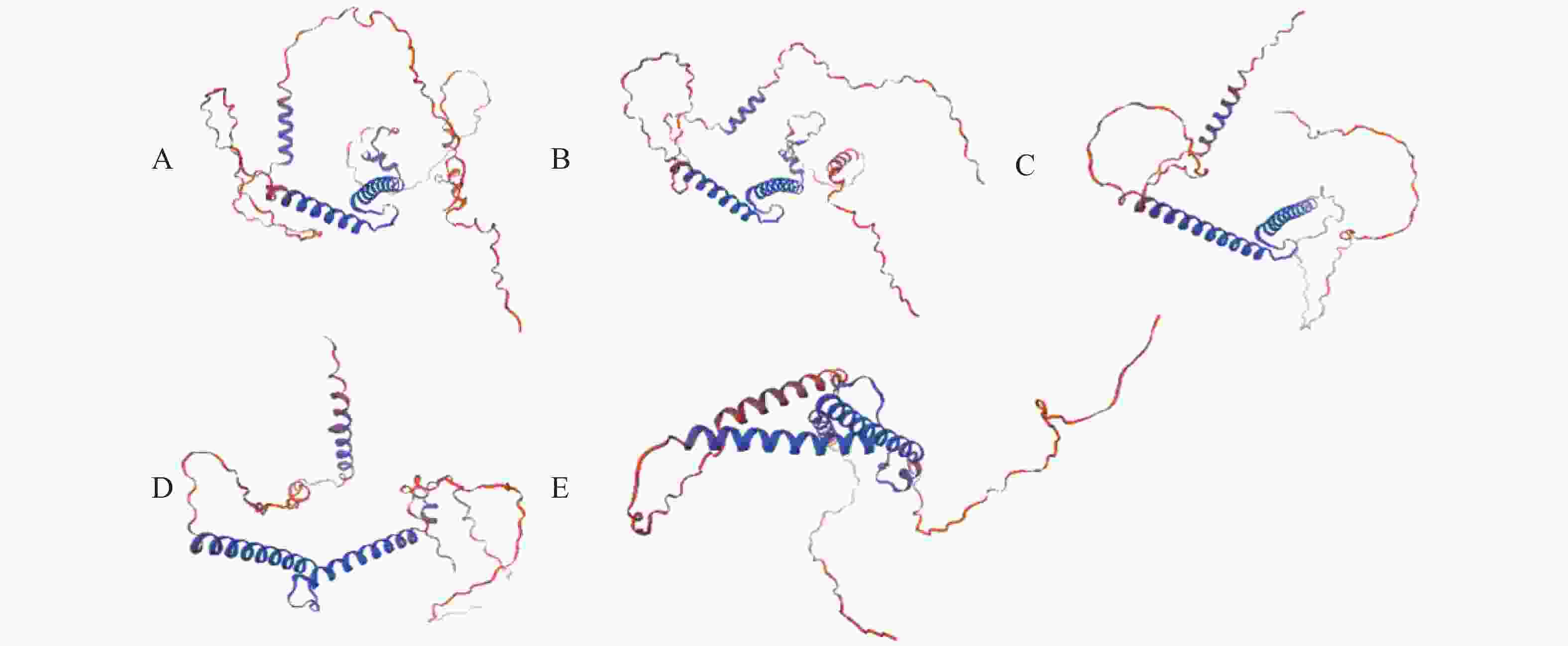
图 3 鞍带石斑鱼MRFs家族蛋白质的三级结构
Figure 3. Tertiary structure of MRFs family proteins in Epinephelus lanceolatus
-
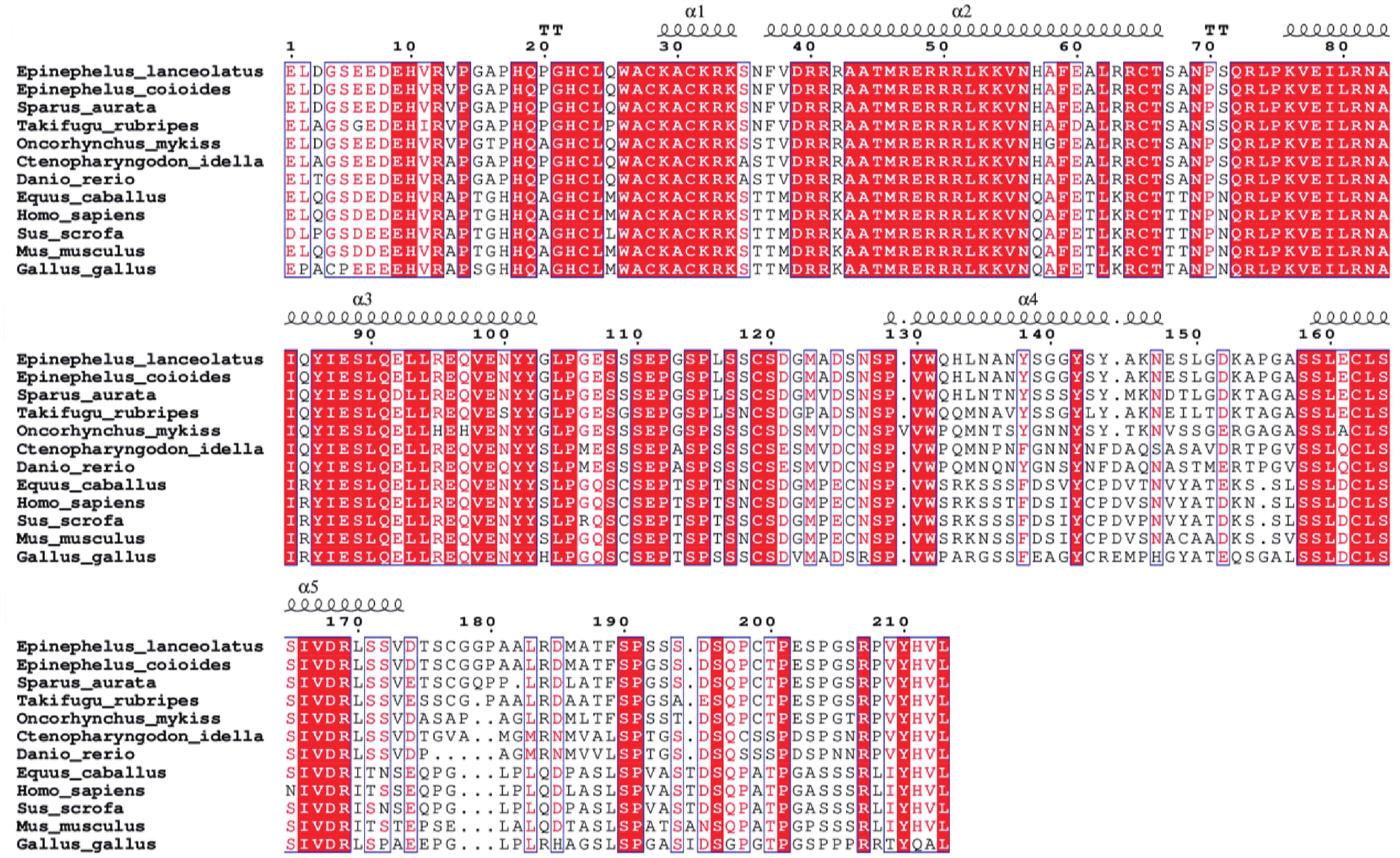
采用DNAMAN软件对鞍带石斑鱼5种生肌调节因子氨基酸序列进行比对,结果表明其具有60个左右氨基酸组成的保守碱性螺旋−环−螺旋(basic helix-loop-helix,bHLH)结构域(图4)。

图 4 鞍带石斑鱼5种MRFs氨基酸序列比对
Figure 4. Multiple alignments of the five MRFs amino acid sequences
-
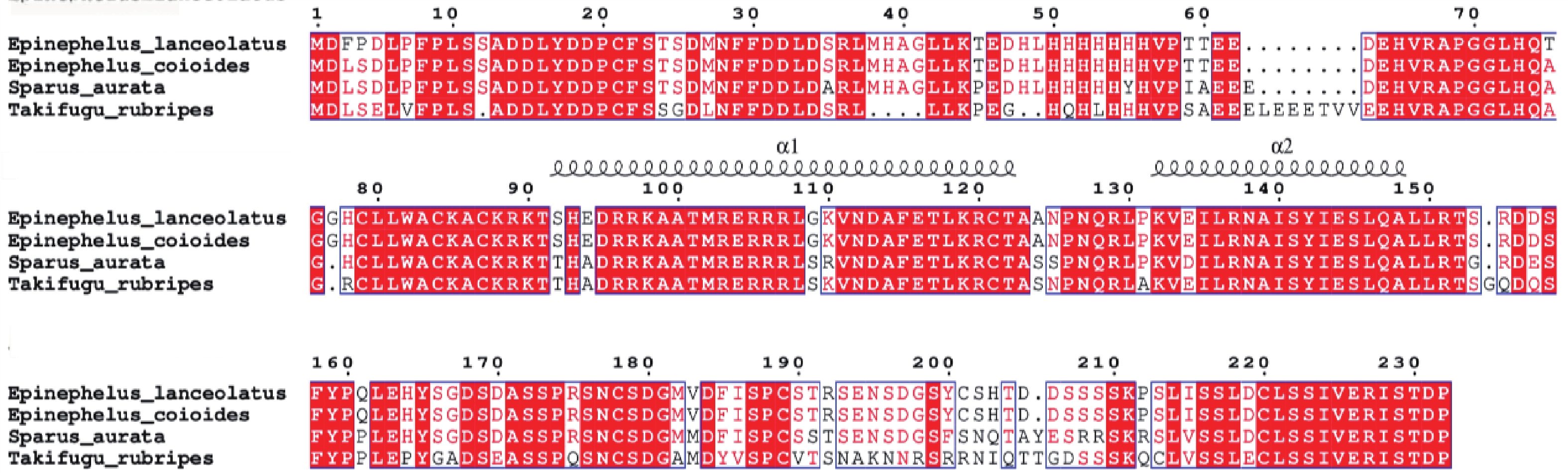
多重序列比对结果表明,鞍带石斑鱼MyoD1与金头鲷同源性最高,为92.93%,其次是草鱼(68.54%),与哺乳类同源性在54.49%~56.12%,与鸟类为64.47%(图5)。

图 5 鞍带石斑鱼MyoD1与其他物种的MyoD1编码区氨基酸序列之间的多重序列比对图
Figure 5. Multiple sequence alignment of the MyoD1 amino acid sequences between Epinephelus lanceolatus and other species
-
多重序列比对结果表明,鞍带石斑鱼MyoD2与斜带石斑鱼同源性最高(98.51%),其次是金头鲷(85.14%),与红鳍东方鲀(70.00%)最低(图6)。
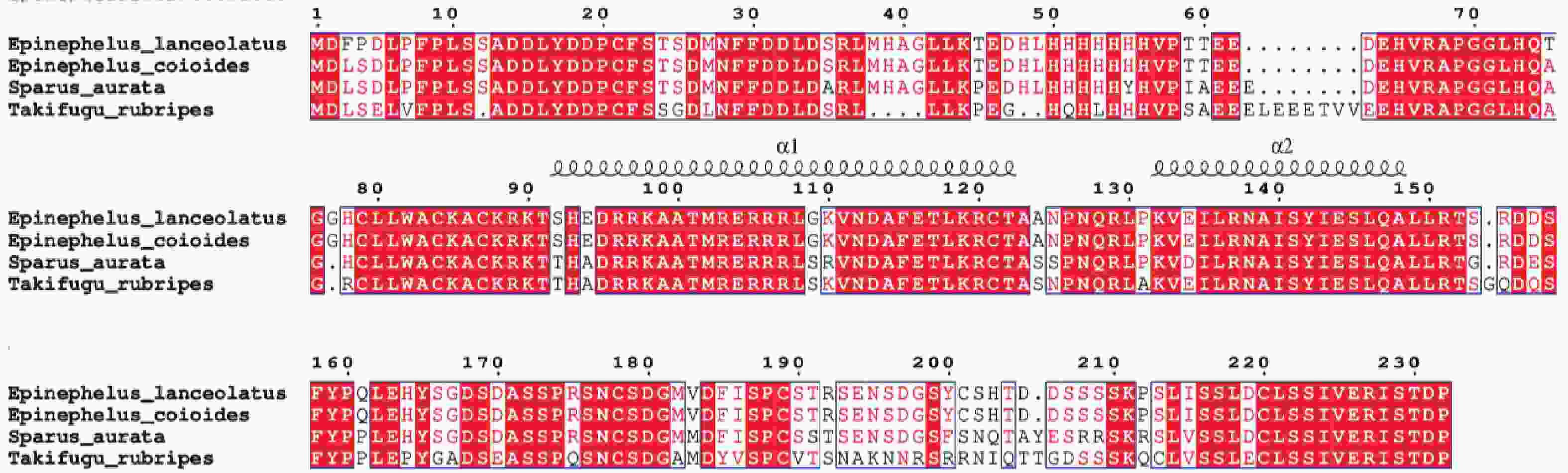
图 6 鞍带石斑鱼MyoD2与其他物种的MyoD2编码区氨基酸序列之间的多重序列比对图
Figure 6. Multiple sequence alignment of the MyoD2 amino acid sequences between Epinephelus lanceolatus and other species
-
多重序列比对结果表明,鞍带石斑鱼MyoG与斜带石斑鱼和金头鲷同源性最高,同源性分别为98.80%、94.40%。与哺乳类(50.20%~51.39%)和鸟类(56.18%)同源性较低(图7)。
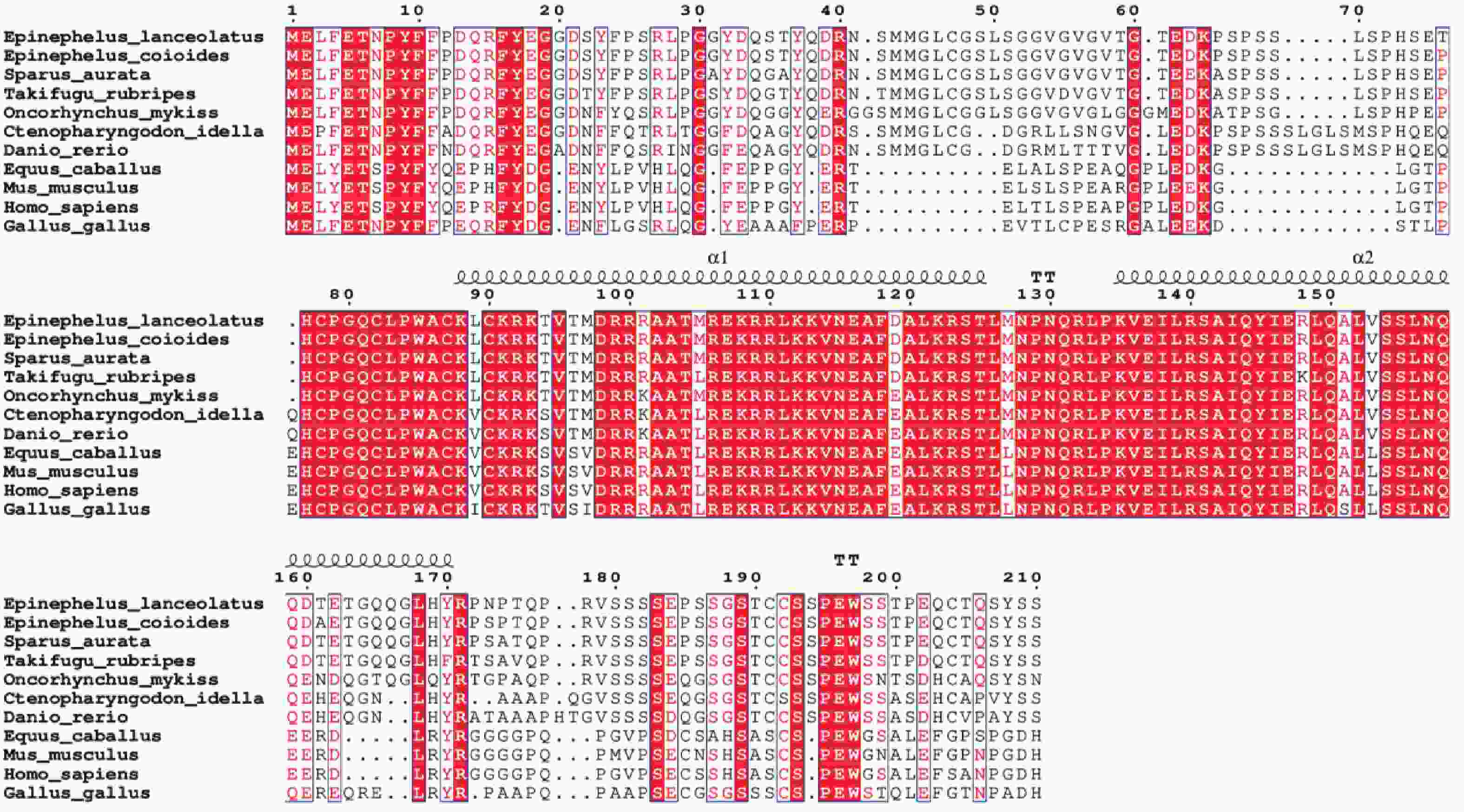
图 7 鞍带石斑鱼MyoG与其他物种的MyoG编码区氨基酸序列之间的多重序列比对图
Figure 7. Multiple sequence alignment of the MyoG amino acid sequences between Epinephelus lanceolatus and other species
-
多重序列比对结果表明,鞍带石斑鱼MRF4与斜带石斑鱼同源性最高,为97.47%,其次是金头鲷(82.92%)和红鳍东方鲀(81.40%),与哺乳类(50.20%~51.39%)和鸟类(56.18%)同源性最低(图8)。
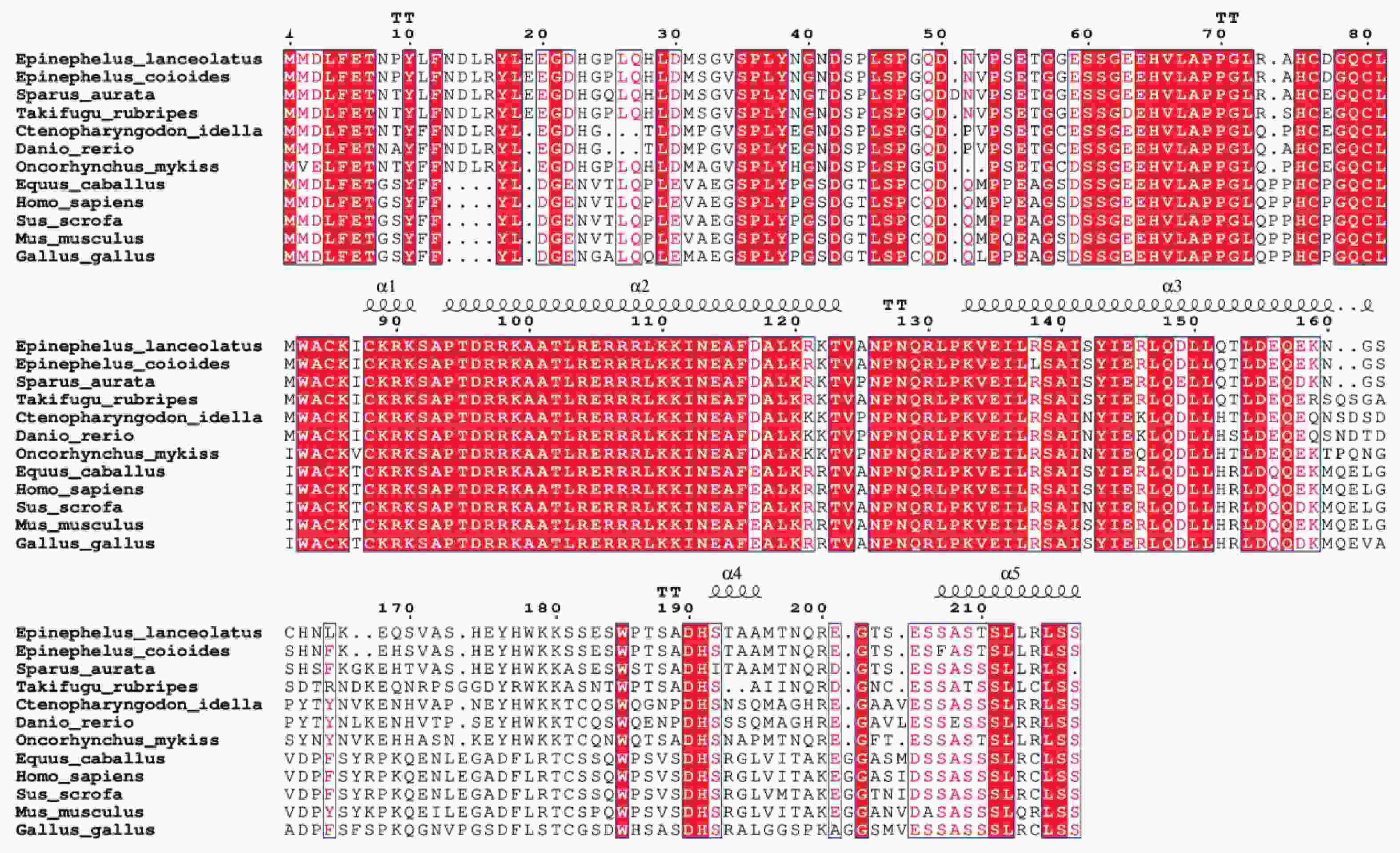
图 8 鞍带石斑鱼MRF4与其他物种的MRF4编码区氨基酸序列之间的多重序列比对图
Figure 8. Multiple sequence alignment of the MRF4 amino acid sequences between Epinephelus lanceolatus and other species
-
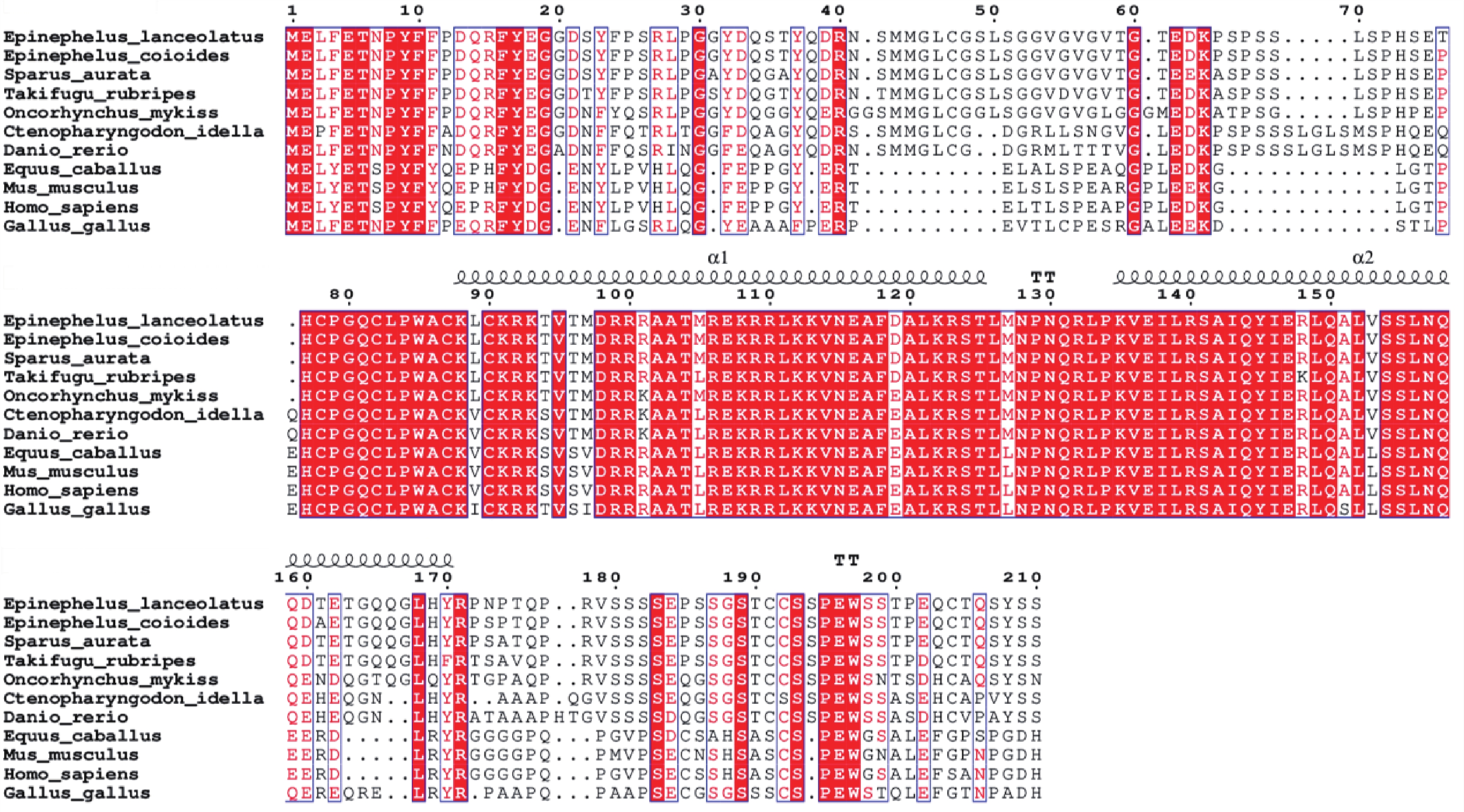
多重序列比对结果表明,鞍带石斑鱼Myf5与斜带石斑鱼同源性最高,为99.59%,其次是金头鲷(91.29%),与红鳍东方鲀、虹鳟及草鱼同源性分别为87.97%、81.82%和75.21%,与哺乳类(56.20%~56.59%)和鸟类(57.09%)同源性较低(图9)。
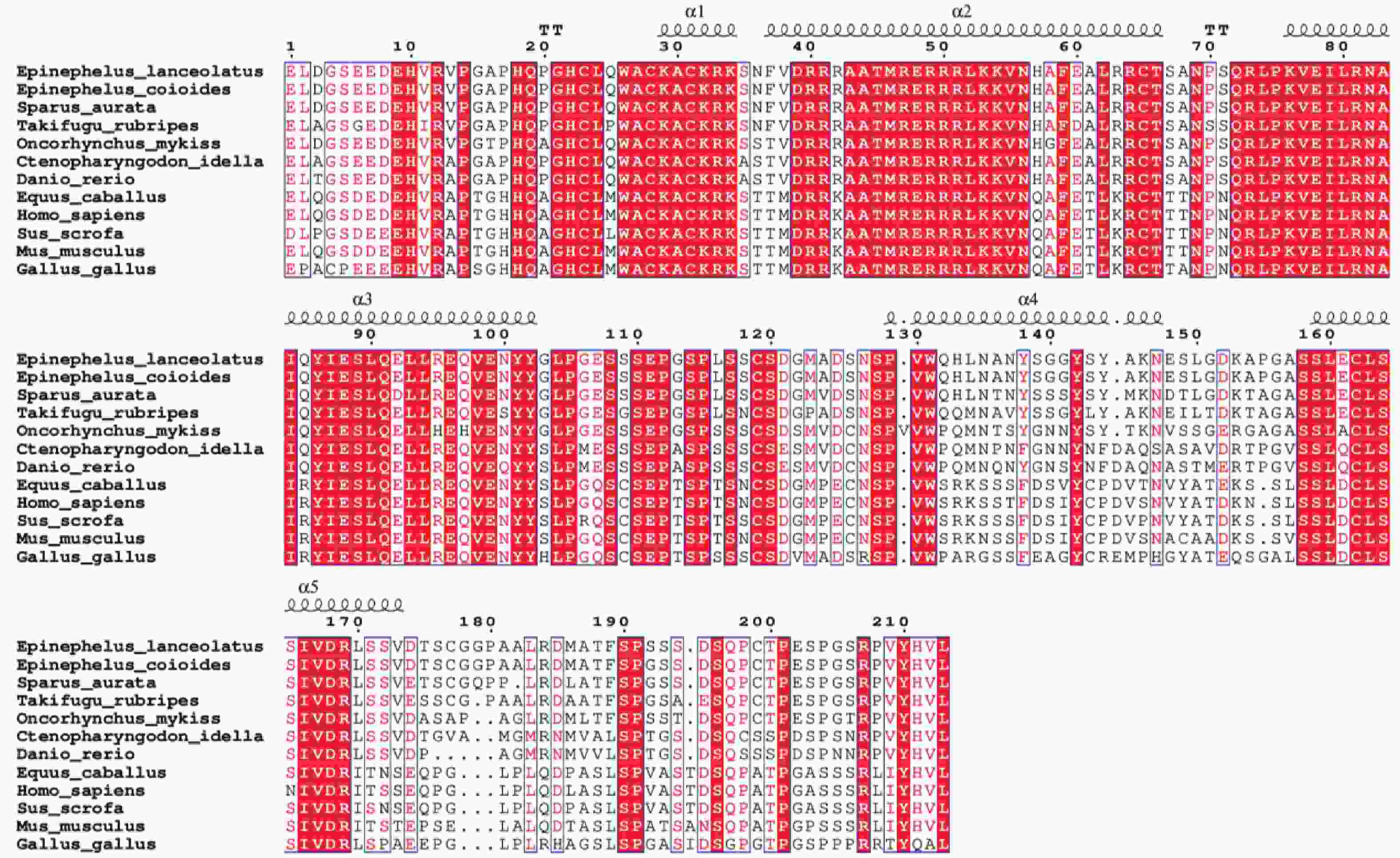
图 9 鞍带石斑鱼Myf5与其他物种的Myf5编码区氨基酸序列之间的多重序列比对图
Figure 9. Multiple sequence alignment of the Myf5 amino acid sequences between Epinephelus lanceolatus and other species
-
采用软件MEGA v7.0基于Neighbor-Joining(N-J)法构建了包括鞍带石斑鱼、人、小鼠、红原鸡、草鱼、斑马鱼等11种物种的系统进化树。结果显示,5种生肌调节因子(MRFs)明显分为两支,其中MyoD1、MyoD2与Myf5聚为一支,MRF4与MyoG聚为另一支。在系统进化过程中,每个基因均明显划分为鱼类、哺乳类和鸟类3大分支。在鱼类分支中,鞍带石斑鱼MRFs与斜带石斑鱼亲缘关系最近,二者聚成簇后与金头鲷聚在一起,再与红鳍东方鲀形成一小支,且各节点具有较高的自展值支持;与草鱼、斑马鱼、虹鳟的进化关系较远(图10)。

图 10 基于不同物种MRFs氨基酸序列构建的系统发育进化树(NJ法)
Figure 10. Phylogenetic tree based on MRFs amino acid sequences of different species (Neighbor-Joining method)
-
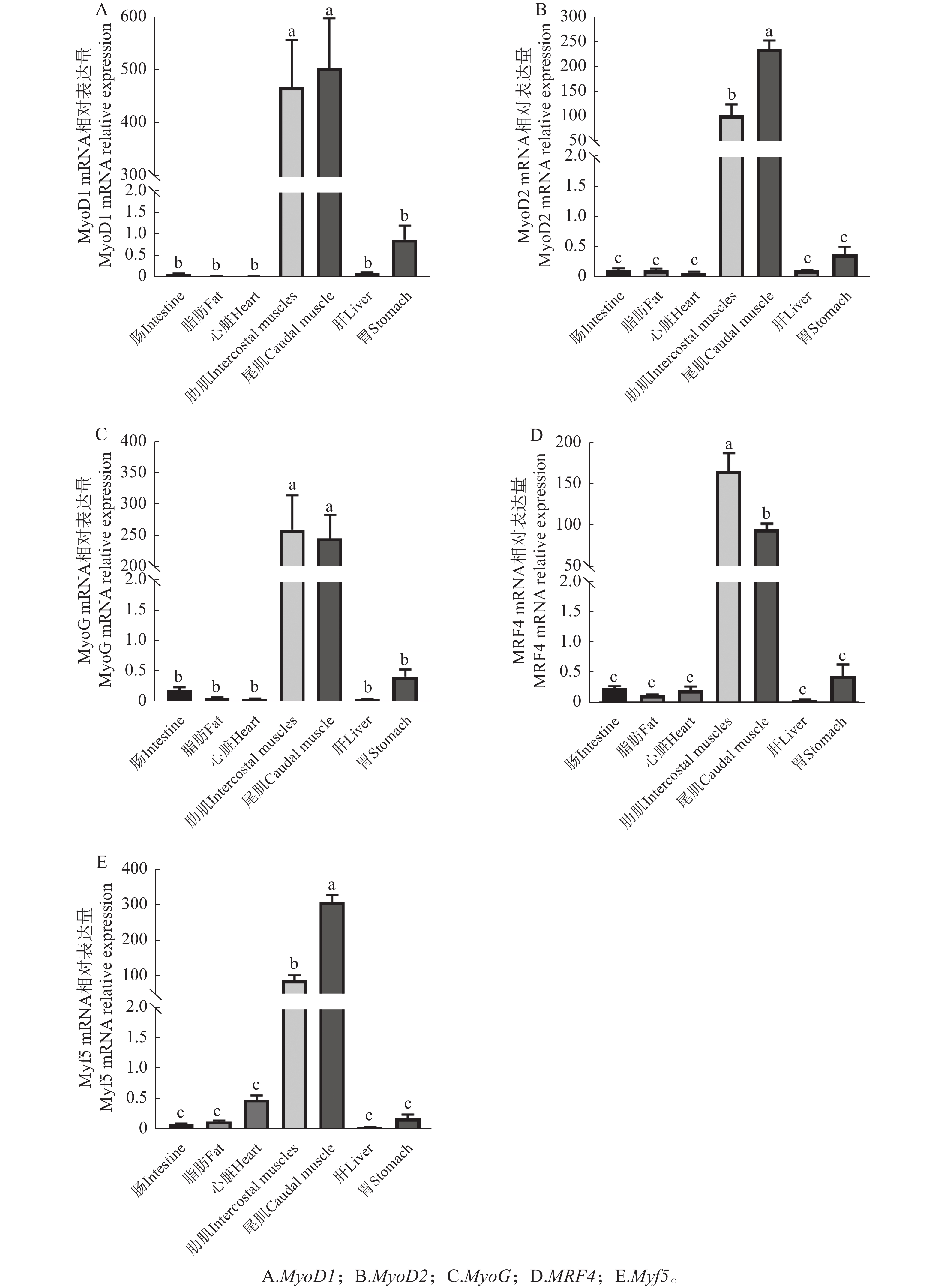
如图11-A所示,MyoD1基因在肠道、胃、肝脏和骨骼肌中均有表达,其中骨骼肌表达量最高,且尾肌表达量略高于肋肌,但两者无显著性差异(P>0.05);在心脏和脂肪组织中几乎检测不到MyoD1表达。MyoD2基因在以上6种器官式组织中均有表达,骨骼肌中表达量最高,且尾肌表达量显著高于肋肌(P<0.05),心脏中表达量最低(图11-B)。MyoG基因在6种组织中均有表达,骨骼肌中表达量最高,肋肌表达量略高于尾肌,但两者无显著性差异(P>0.05)(图11-C)。MRF4基因在肠道、胃、心脏、骨骼肌和脂肪中均有表达,骨骼肌中表达量最高,且肋肌表达量显著高于尾肌(P<0.05),而在肝脏中几乎检测不到MRF4表达(图11-D)。Myf5基因在除肝脏外的5种组织中均有表达,骨骼肌中表达量最高,且肋肌与尾肌表达差异与MyoD2结果一致(图11-E)。总体而言,5种生肌调节因子mRNA在骨骼肌表达量均显著高于其他组织(P<0.000 1)。
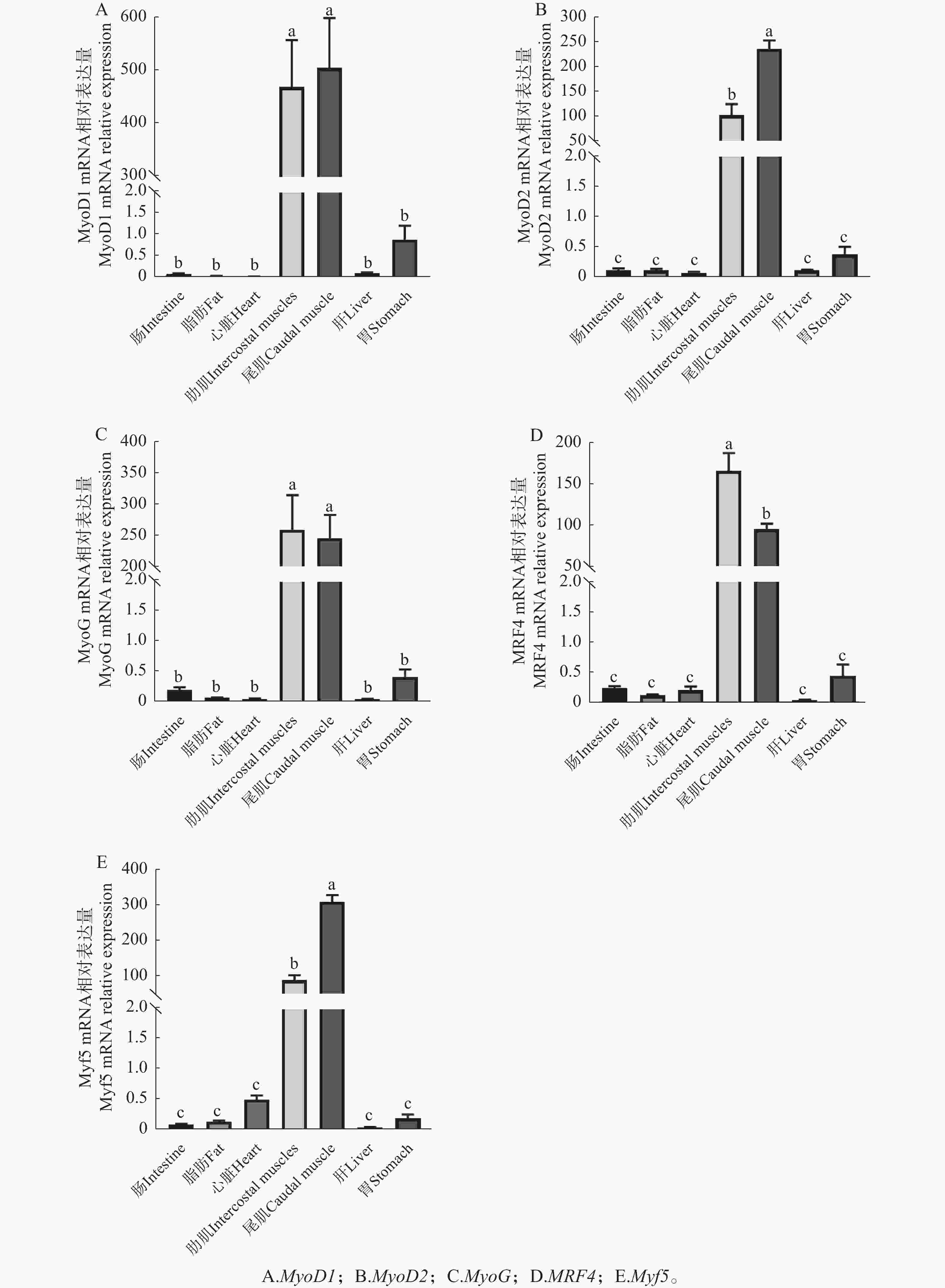
图 11 鞍带石斑鱼5种生肌调节因子在不同组织的相对表达情况
Figure 11. Relative expression levels of 5 MRFs in different tissues of Epinephelus lanceolatus
-
本研究成功克隆了鞍带石斑鱼MyoD1、MyoD2、MyoG、MRF4和Myf5 cDNA序列,其长度分别为1 962、1 466、926、1 052和
1133 bp,分别编码297、270、251、238和242个氨基酸。蛋白结构预测分析表明,鞍带石斑鱼5种MRFs均具有高度保守的碱性螺旋-环-螺旋结构域(bHLH),该区域由60个左右的氨基酸残基组成,其中碱性区域(basic)识别并结合E-box启动子,而HLH区域与E蛋白形成异二聚体复合物,从而激活肌肉特异性基因转录[3]。蛋白结构预测还显示,Myf5、MyoD1和MyoD2均属于Myf5基因超家族,且Myf5与MyoD在功能上具有相似性,均参与肌卫星细胞增殖及肌纤维增生调控。Kassar-Duchossoy等[29]研究发现,Myf5或MyoD单基因敲除小鼠的骨骼肌发育基本正常,但双基因敲除小鼠则完全丧失成肌能力,表明二者在肌生成中具有功能互补性。Relaix等[30]证实,Pax3和Pax7通过结合Myf5和MyoD启动子中的保守E-box和Pax结合位点,直接激活其表达。MyoD作为“主调控因子”的功能依赖于其启动子中的E-box元件(CANNTG),这一特征在Myf5中同样保守。Tapscott等[31]首次提出,E-box是肌源性分化的核心调控模块,且Myf5与MyoD共享这一元件的结合特性。这些研究表明,Myf5与MyoD在肌卫星细胞增殖中的冗余功能及调控机制的保守性,进一步支持了二者由同一个祖先基因复制进化而来的假说。同源性分析结果表明,鞍带石斑鱼MRFs氨基酸序列与其他鱼类的同源性较高,为68.54%~99.59%。特别是与鲈形目的同源性最高,为82.92%~99.59%;而与鸟类和哺乳类的同源性相对较低,为50.20%~64.47%,这证明本研究所得,鞍带石斑鱼MRFs序列所编码的蛋白质与各物种的基因编码的蛋白质具有相似的功能,在鱼类中的高保守性同时也揭示了该蛋白质功能的高度保守性。结合系统进化树结果,鞍带石斑鱼与同属石斑鱼属的斜带石斑鱼聚在同一分支,并与鲈形目鱼类形成一簇,说明亲缘关系最近;而与哺乳类、鸟类的距离较远,说明与陆生动物亲缘性较远。与Chen等[32]基于线粒体构建的进化树结果相符。鱼类MRFs与鸟类和哺乳动物的同源性存在较大差异,推测与物种进化、亲缘关系以及所含的氨基酸残基数目不同有关。
根据结构和功能的不同,MRFs基因家族可分为初级MRFs(Myf5和MyoD)和次级MRFs(MyoG和Myf6)。初级MRFs同源性较高,功能互补,主要调控体节细胞向生肌细胞系的定向分化;次级MRFs则促进成肌细胞增殖并分化为肌细胞,最终形成肌纤维[33]。在鱼类胚胎发育中,MyoD和Myf5表达通常早于MyoG和MRF4。例如,在牙鲆胚胎发育过程中,Myf5和MyoD在受精26 h后即可检测到,而MyoG的表达则首次出现在受精32.5 h后[34]。类似结果在鲤鱼[8]和虹鳟[35]中也有报道。然而,不同鱼类间MRFs表达模式也存在差异。例如,牙鲆MyoG在胚胎发育形成3个体节时首次表达,其表达水平持续上调至30个体节后逐渐下调[34];而大西洋鲱(Clupea harengus)MyoG在胚胎发育形成34个体节前仅短暂表达,并在MyoD开始表达前从体节中消失[36]。这些研究表明,MRFs基因表达时序在鱼类中具有保守性,但也存在物种特异性差异。而进化分析结果同样显示,鞍带石斑鱼MRFs 基因家族明显分为两支,其中MyoD与Myf5聚成一支,而MyoG与MRF4聚成另一支,说明MyoD与Myf5同源性较高,可能与其在肌肉生长发育调控中的功能一致。
本研究发现,鞍带石斑鱼MRFs基因在骨骼肌中呈现高表达,这与在其他鱼类中的研究结果一致。例如,MyoD在斑马鱼胚胎体节及成体骨骼肌卫星细胞中特异性高表达,其敲除可导致体节肌细胞分化完全停滞,证实MyoD是斑马鱼骨骼肌发育的核心调控因子[37]。在鲶鱼(Silurus asotus)中,MyoG在骨骼肌快肌纤维分化阶段特异性高表达,通过激活肌球蛋白重链(MyHC)及肌钙蛋白编码基因,直接驱动肌纤维增粗[38]。松江鲈(Trachidermus fasciatus)的MRF4在骨骼肌中高表达,MRF4不仅维持成熟肌纤维功能稳态,还在肌肉再生过程中被激活,参与修复调控[39]。此外,虹鳟的Myf5主要在骨骼肌中表达,Myf5在肌肉前体细胞分化和胚胎肌肉发育中发挥主导作用[40]。这些结果表明,MRFs家族基因在鱼类骨骼肌中的高表达与其在肌肉生成和分化中的核心功能密切相关。此外,鞍带石斑鱼MRFs基因在其他器官是式组织中也检测到微量表达,推测其可能不仅参与肌肉分化,还在其他器官组织中发挥潜在作用。类似现象在其他鱼类中也有报道,如虹鳟Myf5在肝脏、肠、心脏、鳃和脑中有少量表达[40],达氏鲟的MRFs基因在心脏、脑、肝、脾和肾中也有表达[16]。这些发现进一步支持了MRFs基因在鱼类多种器官式组织中的多功能性。研究表明,鱼类早期生长发育的各个阶段均受到MRFs基因表达的影响。例如,在牙鲆中,MyoD在胚胎发生的初始阶段即在前体肌肉细胞中检测到表达[19];在鳜鱼中,Myf5基因在原肠胚期这一早期发育阶段即开始表达[41];此外,Johansen等[42]研究发现,虹鳟在不同生长阶段中,骨骼肌内的MyoD、Myf5、MyoG和MRF4表达水平存在显著差异。基于上述研究结果可以推断,MRFs在不同发育阶段的差异性表达可能与其调控肌肉生长的分子机制密切相关。
综上所述,本研究成功克隆得到鞍带石斑鱼5种生肌调节因子MyoD(MyoD1和MyoD2)、MyoG、MRF4、Myf5 cDNA 全长序列,并通过生物信息学软件对5种生肌调节因子 CDS区、氨基酸序列、蛋白结构、进化地位、亲缘关系以及不同组织中相对表达等进行了分析。鞍带石斑鱼5种生肌调节因子都具有高度保守的碱性螺旋−环−螺旋结构域(basic helix-loop-helix,bHLH),同时每个基因均与鲈形目鱼类分为一支,且5种生肌调节因子皆在骨骼肌中表达丰度较高,而在肝脏、心脏、肠道等其他组织中表达量较低。研究结果为进一步探究生肌调节因子在鞍带石斑鱼骨骼肌生长发育的作用机制奠定了基础。
鞍带石斑鱼MRFs基因cDNA克隆及组织表达分析
DOI: 10.15886/j.cnki.rdswxb.20250030
 CSTR: 32425.14.j.cnki.rdswxb.20250030
CSTR: 32425.14.j.cnki.rdswxb.20250030
Gene cloning and expression analysis of MRFs gene cDNA in the tissues of giant grouper Epinephelus lanceolatus
-
摘要: 生肌调节因子(MRFs)在动物体骨骼肌生长发育过程中发挥着重要作用。本研究通过RT-PCR和RACE技术克隆得到鞍带石斑鱼5种生肌调节因子(MRFs)cDNA全长序列,包括MyoD1(1 962 bp)、MyoD2(1 466 bp)、MyoG(926 bp)、MRF4(1 052 bp)和Myf5(1 133 bp),分别编码297、270、251、238和242个氨基酸。通过氨基酸序列比对分析发现,5种MRFs均含有由60个氨基酸组成的碱性螺旋−环−螺旋(bHLH)保守结构域。进化树分析显示,MRFs分为两支,其中MyoD(MyoD1和MyoD2)与Myf5聚为一支,MRF4与MyoG聚为另一支,且各基因均与鲈形目鱼类聚为一支。实时荧光定量PCR结果显示,5种MRFs在骨骼肌中表达量较高,而在肝脏、心脏和肠道等组织中表达较低。该研究为揭示MRFs在鞍带石斑鱼骨骼肌生长发育中的作用机制奠定了基础。Abstract: Myogenic regulatory factors (MRFs) are critical regulators of skeletal muscle growth and development in animals. The full-length cDNA sequences of five MRFs from the giant grouper (Epinephelus lanceolatus) were cloned using RT-PCR and RAC, including MyoD1 (1,962 bp), MyoD2 (1,466 bp), MyoG (926 bp), MRF4 (1,052 bp), and Myf5 (1,133 bp), encoding 297, 270, 251, 238, and 242 amino acids, respectively. Amino acid sequence alignment revealed that all five MRFs contain a conserved basic helix-loop-helix (bHLH) domain consisting of 60 amino acids. Phylogenetic analysis demonstrated that the MRF genes are clustered into two distinct branches: MyoD (MyoD1 and MyoD2) groups with Myf5 in one clade, while MRF4 and MyoG form the other. All genes showed closest evolutionary relationships with Perciformes homologs. Real-time quantitative PCR analysis revealed predominant expression of these MRFs in skeletal muscle, with significantly lower expression levels in the liver, heart, and intestine. These findings provide foundation for elucidating the regulatory mechanisms of MRFs in skeletal muscle development of the giant grouper.
-
Key words:
- Epinephelus lanceolatus /
- myogenic regulatory factors /
- cDNA cloning /
- gene expression
-
图 2 鞍带石斑鱼MRFs家族蛋白二级结构预测
注:蓝色代表α−螺旋区域,红色代表β−折叠区域,绿色代表转角区域,空缺代表无规则卷曲区域。
Fig. 2 Prediction of the secondary structure of MRFs protein in Epinephelus lanceolatus
Note: Blue indicate α−helix area; red indicate β−sheet area; green indicate β−turn area; vacancy indicate random coil area.
图 3 鞍带石斑鱼MRFs家族蛋白质的三级结构
Fig. 3 Tertiary structure of MRFs family proteins in Epinephelus lanceolatus
A.MyoD1;B.MyoD2;C.MyoG;D.MRF4;E.Myf5。
图 4 鞍带石斑鱼5种MRFs氨基酸序列比对
注:bHLH结构域用下划线标记并字体加粗注明;“−”代表插入缺失;“*”代表氨基酸完全一致;“.”代表有2种氨基酸。
Fig. 4 Multiple alignments of the five MRFs amino acid sequences
Note: The bHLH domain is underlined and bold typed; "−" represents indel; "*" means identical amino acids; "." means two amino acids.
图 5 鞍带石斑鱼MyoD1与其他物种的MyoD1编码区氨基酸序列之间的多重序列比对图
注:MyoD1的二级结构元素显示在排列上方;α−螺旋表示为螺旋。
Fig. 5 Multiple sequence alignment of the MyoD1 amino acid sequences between Epinephelus lanceolatus and other species
Note: The secondary structural elements of MyoD1 are showed above the arrangement,α−helix is represented as a helix.
图 6 鞍带石斑鱼MyoD2与其他物种的MyoD2编码区氨基酸序列之间的多重序列比对图
注:MyoD2的二级结构元素显示在排列上方;α−螺旋表示为螺旋。
Fig. 6 Multiple sequence alignment of the MyoD2 amino acid sequences between Epinephelus lanceolatus and other species
Note: The secondary structural elements of MyoD2 are showed above the arrangement; α−helix is represented as a helix.
图 7 鞍带石斑鱼MyoG与其他物种的MyoG编码区氨基酸序列之间的多重序列比对图
注:MyoG的二级结构元素显示在排列上方;α−螺旋表示为螺旋;β−转角表示为 TT。
Fig. 7 Multiple sequence alignment of the MyoG amino acid sequences between Epinephelus lanceolatus and other species
Note: The secondary structural elements of MyoG are showed above the arrangement; α−helix is represented as a helix; β−turn is denoted as TT.
图 8 鞍带石斑鱼MRF4与其他物种的MRF4编码区氨基酸序列之间的多重序列比对图
注:MRF4的二级结构元素显示在排列上方;α−螺旋表示为螺旋;β−转角表示为 TT。
Fig. 8 Multiple sequence alignment of the MRF4 amino acid sequences between Epinephelus lanceolatus and other species
Note: The secondary structural elements of MRF4 are showed above the arrangement; α−helix is represented as a helix; β−turn is denoted as TT.
图 9 鞍带石斑鱼Myf5与其他物种的Myf5编码区氨基酸序列之间的多重序列比对图
注:Myf5的二级结构元素显示在排列上方;α−螺旋表示为螺旋;β−转角表示为 TT。
Fig. 9 Multiple sequence alignment of the Myf5 amino acid sequences between Epinephelus lanceolatus and other species
Note: The secondary structural elements of Myf5 are showed above the arrangement; α−helix is represented as a helix; β−turn is denoted as TT.
图 10 基于不同物种MRFs氨基酸序列构建的系统发育进化树(NJ法)
Fig. 10 Phylogenetic tree based on MRFs amino acid sequences of different species (Neighbor-Joining method)
图 11 鞍带石斑鱼5种生肌调节因子在不同组织的相对表达情况
注:EF1-α作为内参基因。数值以平均值±标准差(n=6)表示;不同字母表示5种生肌调节因子在不同组织间具有显著性差异。
Fig. 11 Relative expression levels of 5 MRFs in different tissues of Epinephelus lanceolatus
Note: EF1-α serves as the reference gene. Values are expressed as mean ± SE (n = 6); different letters indicate significant differences in the five MRFs among different tissues.
表 1 基因克隆所用引物序列
Table 1 Primer sequences for gene cloning
引物 Primer 序列(3′-5′ ) Sequence 中间片段引物序列(5′-3′)
Middle fragment primer sequences (5′-3′)MyoD1 F1 ATGCACTTCTTCGAGGAC MyoD1 R1 TGTTGTCGGTGGAGATTC MyoD2 F1 CTCTGCTGACGACCTCTA MyoD2 R1 TTACTGCTGCTGGAATCG MyoG F1 TACGACCAATCCACCTATCA MyoG R1 TCCGCCTGCTGTAGAGAT MRF4 F1 CGCTATCTGGAAGAAGGG MRF4 R1 CTCTCTGGTTTGTCATTGC Myf5 F1 ACGTCTTCTCACCATCCCAG Myf5 R1 AGAAGGTGGCCATGTCTCTC 3′RACE所用引物序列(5′-3′)
3′RACE primer sequences (5′-3′)MyoD1 3′-F1 GTCCTCCCTCCTGCACCTCCATCAC MyoD1 3′-F2 GAGCACTACAGCGGGGACTCAGACG MyoD2 3′-F1 CCCCGCTCCAACTGCTCCGATGG MyoD2 3′-F2 GCAGCCACACAGACGATTCCAGCAGC MyoG 3′-F1 CCCAACCCGACCCAACCCAGAGTGT MyoG 3′-F2 CATGCGTGCCCTGACCTCCATCGTG MRF4 3′-F1 CCAACCCCAACCAGAGGCTACCCAA MRF4 3′-F2 CGACCTCTGCCGACCATTCCACTGC Myf5 3′-F1 CGCAACGCCATCCAGTACATCGAGAGC Myf5 3′-F2 TCTGAATGCAAACTACAGCGGCGGAT 5′RACE所用引物序列(5′-3′)
5′RACE primer sequences (5′-3′)MyoD1 5′-R1 CGTGGTGATGGAGGTGCAGGAGGGA MyoD1 5′-R2 ATGGAGGTGCAGGAGGGAGG MyoD2 5′-R1 CTTACCAAGCCGCCGCCTTTCTCGC MyoD2 5′-R2 CTTTCTCGCATCGTCGCTGCTTTCC MyoG 5′-R1 CTTGGGCAGCCTCTGGTTTGGGTTC MyoG 5′-R2 GACGCCTCTTCTCCCTCATCGTGGC MRF4 5′-R1 TGGGCGCCGACTTCCTCTTGCAGAT MRF4 5′-R2 CATGAGGCACTGGCCGTCGCAG Myf5 5′-R1 TGCAGGCTCTCGATGTACTGGATGGCG Myf5 5′-R2 CTGGCTGGGGTTGGCTGAGGTGC 全长验证引物序列(5′-3′)
Full-length validation primer sequences (5′-3′)MyoD1 Full-F ACGCTTCTGTCTCTGTCACTGGAT MyoD1 Full-R ATGGTAAAGTCCCAGGTGTGAAGG MyoD2 Full-F CCCCGCTCCAACTGCTCCGATGG MyoD2 Full-R GCAGCCACACAGACGATTCCAGCAGC MyoG Full-F CCGAACACCACATACAGTAGAGC MyoG Full-R TGGAGGGTCTACTTGGGAAT MRF4 Full-F GGGCGCAACAATATGATGGACCTTT MRF4 Full-R GAATATAGTGGAAATAATTTAGGGA Myf5 Full-F ATGGAYGTCTTYTCVCCVTCCCA Myf5 Full-R TGGCGTTCGGTAAAAGCAG 接头引物序列(5′-3′)
Adapter primer sequences (5′-3′)3′ CDS AAGCAGTGGTATCAACGCAGAGTACTT
TTTTTTTTTTTTTTTTTTTTTTTTTTTTVN5′ CDS TTTTTTTTTTTTTTTTTTTTTTTTTVN NUPA AAGCAGTGGTATCAACGCAGAGT UPMA-S CTAATACGACTCACTATAGGGC UPMA-L CTAATACGACTCACTATAGGGCAAGCAG
TGGTATCAACGCAGAGT 下载: 导出CSV
下载: 导出CSV
表 2 实时荧光定量PCR检测5种基因组织分布所用引物序列
Table 2 Nucleotide sequences of the primers used for Real-time PCR
引物 Primer 序列3′-5′ Sequence MyoD1 RT-F GGCATGACGGATTTCAACGG MyoD1 RT-R CTGCTGTTGTCGGTGGAGAT MyoD2 RT-F ATCATCACCACCACCACGTC MyoD2 RT-R TTCAGGGTCTCAAAGGCGTC MyoG RT-F CACGATGAGGGAGAAGAGGC MyoG RT-R TGCTGGTTGAGGGAAGACAC MRF4 RT-F CAGTGTGGCCAGTCATGAGT MRF4 RT-R TCTTCGGTGGTGATGCTGTC Myf5 RT-F GAGGAGGACGAGCATGTGAG Myf5 RT-R CTCGATGTACTGGATGGCGT EF1-α RT-F AGGGATGGAAGATTGAGCGC EF1-α RT-R CGTACCGGGCTTCAGGATAC
下载: 导出CSV
-
[1] Rajesh M, Kamalam B S, Ciji A, et al. Molecular characterisation and transcriptional regulation of muscle growth regulatory factors myogenin and myogenic factor 6 in the Trans-Himalayan cyprinid fish Schizothorax richardsonii [J]. Comparative Biochemistry and Physiology Part A: Molecular & Integrative Physiology, 2019, 231: 188−200. [2] Zammit P S. Function of the myogenic regulatory factors Myf5, MyoD, Myogenin and MRF4 in skeletal muscle, satellite cells and regenerative myogenesis [J]. Seminars in Cell & Developmental Biology, 2017, 72: 19−32. [3] Tapscott S J. The circuitry of a master switch: Myod and the regulation of skeletal muscle gene transcription [J]. Development, 2005, 132(12): 2685−2695. https://doi.org/10.1242/dev.01874 doi: 10.1242/dev.01874 [4] Buckingham M. Molecular biology of muscle development [J]. Cell, 1994, 78(1): 15−21. https://doi.org/10.1016/0092-8674(94)90568-1 doi: 10.1016/0092-8674(94)90568-1 [5] Pas M F, Visscher A H. Genetic regulation of meat production by embryonic muscle formation - a review [J]. Zeitschrift Fur Tierzuchtung und Zuchtungsbiologie, 1994, 111(1/2/3/4/5/6): 404−412. [6] Watabe S. Myogenic regulatory factors and muscle differentiation during ontogeny in fish [J]. Journal of Fish Biology, 2005, 55: 1−18. [7] Chen Y H, Lee W C, Liu C F, et al. Molecular structure, dynamic expression, and promoter analysis of zebrafish (Danio rerio) myf-5 gene [J]. Genesis, 2001, 29(1): 22−35. [8] Cole N J, Hall T E, Martin C I, et al. Temperature and the expression of myogenic regulatory factors (MRFs) and myosin heavy chain isoforms during embryogenesis in the common carp Cyprinus carpio L.[J]. Journal of Experimental Biology, 2004, 207(Pt 24): 4239−4248. [9] Tan X, Zhang Y, Zhang P J, et al. Molecular structure and expression patterns of flounder (Paralichthys olivaceus) Myf-5, a myogenic regulatory factor [J]. Comparative Biochemistry and Physiology Part B: Biochemistry and Molecular Biology, 2006, 145(2): 204−213. https://doi.org/10.1016/j.cbpb.2006.07.008 doi: 10.1016/j.cbpb.2006.07.008 [10] Delalande J M, Rescan P Y. Differential expression of two nonallelic MyoD genes in developing and adult myotomal musculature of the trout (Oncorhynchus mykiss)[J]. Development Genes and Evolution, 1999, 209(7): 432−437. https://doi.org/10.1007/s004270050274 doi: 10.1007/s004270050274 [11] Hall T E, Cole N J, Johnston I A. Temperature and the expression of seven muscle-specific protein genes during embryogenesis in the Atlantic cod Gadus morhua L. [J]. Journal of Experimental Biology, 2003, 206(Pt 18): 3187−3200. [12] Ganassi M, Badodi S, Quiroga H P O, et al. Myogenin promotes myocyte fusion to balance fibre number and size [J]. Nature Communications, 2018, 9(1): 4232. https://doi.org/10.1038/s41467-018-06583-6 doi: 10.1038/s41467-018-06583-6 [13] Hinits Y, Osborn D P S, Carvajal J J, et al. Mrf4 (myf6) is dynamically expressed in differentiated zebrafish skeletal muscle [J]. Gene Expression Patterns, 2007, 7(7): 738−745. https://doi.org/10.1016/j.modgep.2007.06.003 doi: 10.1016/j.modgep.2007.06.003 [14] Schnapp E, Pistocchi A S, Karampetsou E, et al. Induced early expression of mrf4 but not myog rescues myogenesis in the myod/myf5 double-morphant zebrafish embryo [J]. Journal of Cell Science, 2009, 122(Pt 4): 481−488. [15] 李虹辉. 翘嘴鳜(Siniperca chuatsi)生肌调节因子MRF4和Myf5的克隆及其表达研究[D]. 南宁: 广西大学, 2014. [16] Wu X Y, Lai J S, Chen Y Y, et al. Characterization of MRF genes and their tissue distributions and analysis of the effects of starvation and refeeding on the expression of these genes in Acipenser dabryanus muscle [J]. Comparative Biochemistry and Physiology Part B: Biochemistry and Molecular Biology, 2021, 256: 110648. [17] Fernandes J M O, Kinghorn J R, Johnston I A. Differential regulation of multiple alternatively spliced transcripts of MyoD [J]. Gene, 2007, 391(1/2): 178−185. [18] Tan X, Du S J. Differential expression of two MyoD genes in fast and slow muscles of gilthead seabream (Sparus aurata) [J]. Development Genes and Evolution, 2002, 212(5): 207−217. https://doi.org/10.1007/s00427-002-0224-5 doi: 10.1007/s00427-002-0224-5 [19] Zhang Y, Tan X, Zhang P J, et al. Characterization of muscle-regulatory gene, MyoD, from flounder (Paralichthys olivaceus) and analysis of its expression patterns during embryogenesis [J]. Marine Biotechnology, 2006, 8(2): 139−148. [20] Shi B, Sun R, Liu X, et al. Cloning, phylogenetic and expression analysis of two MyoDs in yellowtail kingfish (Seriola lalandi)[J]. General and Comparative Endocrinology, 2024, 347: 114422. [21] 张燕萍, 邵芳, 周峰, 等. 赤眼鳟MyoG基因的克隆与组织表达分析[J]. 南京农业大学学报, 2014, 37(5): 117−122. https://doi.org/10.7685/j.issn.1000-230.2014.05.019 doi: 10.7685/j.issn.1000-230.2014.05.019 [22] 朱春月, 杨培民, 刘忠航, 等. 刀鲚MyoG基因克隆和组织表达分析[J]. 天津农业科学, 2024, 30(10): 33−39. https://doi.org/10.3969/j.issn.1006-6500.2024.10.006 doi: 10.3969/j.issn.1006-6500.2024.10.006 [23] Sukhan Z P, Cho Y, Hossen S, et al. Molecular characterization, expression analysis, and CRISPR/Cas9 mediated gene disruption of myogenic regulatory factor 4 (MRF4) in Nile Tilapia [J]. Current Issues in Molecular Biology, 2024, 46(12): 13725−13745. [24] Palma P, Takemura A, Libunao G X, et al. Reproductive development of the threatened giant grouper Epinephelus lanceolatus [J]. Aquaculture, 2019, 509: 1−7. [25] 陈超, 孔祥迪, 李炎璐, 等. 棕点石斑鱼(♀)×鞍带石斑鱼(♂)杂交子代胚胎及仔稚幼鱼发育的跟踪观察[J]. 渔业科学进展, 2014, 35(5): 135−144. https://doi.org/10.11758/yykxjz.20140520 doi: 10.11758/yykxjz.20140520 [26] 吴水清, 郑乐云, 黄种持, 等. 云纹石斑鱼(♀)×鞍带石斑鱼(♂)杂交子代胚胎发育及仔稚幼鱼形态观察[J]. 渔业研究, 2016, 38(1): 27−35. [27] 刘玲, 陈超, 李炎璐, 等. 短期温度胁迫对驼背鲈(♀)×鞍带石斑鱼(♂)杂交子代幼鱼抗氧化及消化酶活性的影响[J]. 渔业科学进展, 2018, 39(2): 59−66. [28] Livak K J, Schmittgen T D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method [J]. Methods, 2001, 25(4): 402−408. https://doi.org/10.1006/meth.2001.1262 doi: 10.1006/meth.2001.1262 [29] Kassar-Duchossoy L, Gayraud-Morel B, Gomès D, et al. Mrf4 determines skeletal muscle identity in Myf5: Myod double-mutant mice [J]. Nature, 2004, 431(7007): 466−471. https://doi.org/10.1038/nature02876 doi: 10.1038/nature02876 [30] Relaix F, Rocancourt D, Mansouri A, et al. A Pax3/Pax7-dependent population of skeletal muscle progenitor cells [J]. Nature, 2005, 435(7044): 948−953. https://doi.org/10.1038/nature03594 doi: 10.1038/nature03594 [31] Tapscott S J, Davis R L, Thayer M J, et al. MyoD1: a nuclear phosphoprotein requiring a Myc homology region to convert fibroblasts to myoblasts [J]. Science, 1988, 242(4877): 405−411. https://doi.org/10.1126/science.3175662 doi: 10.1126/science.3175662 [32] Chen D X, Chu W Y, Liu X L, et al. Phylogenetic studies of three sinipercid fishes (Perciformes: Sinipercidae) based on complete mitochondrial DNA sequences [J]. Mitochondrial DNA, 2012, 23(2): 70−76. https://doi.org/10.3109/19401736.2011.653799 doi: 10.3109/19401736.2011.653799 [33] Perry R L, Rudnick M A. Molecular mechanisms regulating myogenic determination and differentiation [J]. Frontiers in Bioscience, 2000, 5: D750−D767. https://doi.org/10.2741/Perry doi: 10.2741/Perry [34] Xu P, Tan X, Zhang Y, et al. Cloning and expression analysis of myogenin from flounder (Paralichthys olivaceus) and promoter analysis of muscle-specific expression [J]. Comparative Biochemistry and Physiology Part B: Biochemistry and Molecular Biology, 2007, 147(1): 135−145. https://doi.org/10.1016/j.cbpb.2007.01.012 doi: 10.1016/j.cbpb.2007.01.012 [35] Xie S Q, Mason P S, Stickland N C, et al. Lower environmental temperature delays and prolongs myogenic regulatory factor expression and muscle differentiation in rainbow trout (Onchrhynchus mykiss) embryos [J]. Differentiation, 2001, 68(2/3): 106−114. [36] Temple G K, Cole N J, Johnston I A. Embryonic temperature and the relative timing of muscle-specific genes during development in herring (Clupea harengus L.)[J]. Journal of Experimental Biology, 2001, 204(Pt 21): 3629−3637. [37] Hinits Y, Williams V C, Sweetman D, et al. Defective cranial skeletal development, larval lethality and haploinsufficiency in Myod mutant zebrafish [J]. Developmental Biology, 2011, 358(1): 102−112. https://doi.org/10.1016/j.ydbio.2011.07.015 doi: 10.1016/j.ydbio.2011.07.015 [38] Gregory D J, Waldbieser G C, Bosworth B G. Cloning and characterization of myogenic regulatory genes in three Ictalurid species [J]. Animal Genetics, 2004, 35(6): 425−430. https://doi.org/10.1111/j.1365-2052.2004.01193.x doi: 10.1111/j.1365-2052.2004.01193.x [39] 杨磊, 李风鸣, 蔡宝平, 等. 松江鲈MyoG基因cDNA的克隆与序列分析[J]. 江苏农业科学, 2013, 41(1): 16−21. https://doi.org/10.3969/j.issn.1002-1302.2013.01.005 doi: 10.3969/j.issn.1002-1302.2013.01.005 [40] Johansen K A, Overturf K. Sequence, conservation, and quantitative expression of rainbow trout Myf5 [J]. Comparative Biochemistry and Physiology Part B: Biochemistry and Molecular Biology, 2005, 140(4): 533−541. https://doi.org/10.1016/j.cbpc.2004.11.016 doi: 10.1016/j.cbpc.2004.11.016 [41] Zhu X, Li Y L, Liu L, et al. Molecular characterization of Myf5 and comparative expression patterns of myogenic regulatory factors in Siniperca chuatsi [J]. Gene Expression Patterns, 2016, 20(1): 1−10. https://doi.org/10.1016/j.gep.2015.10.003 doi: 10.1016/j.gep.2015.10.003 [42] Johansen K A, Overturf K. Quantitative expression analysis of genes affecting muscle growth during development of rainbow trout (Oncorhynchus mykiss)[J]. Marine Biotechnology, 2005, 7(6): 576−587. https://doi.org/10.1007/s10126-004-5133-3 doi: 10.1007/s10126-004-5133-3 -

 点击查看大图
点击查看大图
计量
- 文章访问数: 1018
- HTML全文浏览量: 580
- PDF下载量: 68
- 被引次数: 0
