




-
据考证,辣椒有着六千多年的栽培史[1],于四百多年前被传入中国[2],其从观赏植物开始发展至今,已在人们的饮食生活中占据重要地位,并在医用、工业等方面也被广泛应用。辣椒五大栽培种之一的一年生辣椒(Capsicum annuum)有着最长的栽培历史,是分化最多、栽培最广的一个种[3]。然而低温限制了植物的地理分布,并显著降低作物产量[4]。在低温胁迫下,植物膜平衡被打破,细胞膜透性提高,钙离子从外界进入细胞质,导致内部代谢失衡,活性氧自由基大量累积,叶绿体等结构遭破坏[5 − 7]。辣椒适宜栽培温度15~34 ℃,对低温敏感,在低温胁迫下,辣椒生长发育缓慢,种子萌发时间延长,种子发芽率下降,茎径、叶片增速减缓并降低产量和质量等[8 − 10]。已有研究发现MYB(v-myb avian myeloblastosis viral oncogene homolog)作为植物中一大转录因子家族,对植株在低温环境下的耐受能力有着重要的调控作用[11 − 13]。但目前对于提高辣椒耐低温胁迫能力的研究大多通过外源物的施加,在MYB转录因子调控辣椒耐低温能力方面的研究较少[14 − 16]。故根据团队前期比较组学结果,本研究从响应胁迫的MYB转录因子中,克隆出CaMYB44基因,并通过生物信息学分析,运用实时荧光定量反转录·聚合酶链反应(real time fluorescent quantitative reverse transcription-polymerase chain reaction, RT-qPCR)技术,检测低温胁迫前后辣椒叶片中CaMYB44表达情况,为后续进一步探索该转录因子调控功能提供基础数据。
-
实验材料为本团队保存的辣椒种质HNUCA54,该种质在强日照下,植株出现生长停滞、新叶枯死等现象,大田栽培期间,对肥害、缺素敏感性高,植株萎蔫,叶片发黄,较其他种质表现出更低的耐受性。植株种植于海南大学实验基地1号大棚(20°3′38″N,110°19′8″E)。亚细胞定位所用材料为本氏烟草,种植于海南大学园艺学院植物培养室,光周期16 h/8 h。RNA提取所采用材料为辣椒植株嫩叶片,用液氮研磨后,使用诺唯赞RNA提取试剂盒(编号RC411-01),进行RNA提取,存于−80 ℃。用诺唯赞cDNA反转录试剂盒(编号R312-02),反转录RNA,并将所得cDNA于−20 ℃存储备用。
-
结合团队先前保存的辣椒转录组数据[17],参考一年生辣椒基因组(
https://www.ncbi.nlm.nih.gov/datasets/taxonomy/4072/ )[18],根据目标基因ID号(XM_016713322.2),下载相应序列信息。利用Primer5.0软件设计基因全长克隆引物:CaMYB44-F,5′-ATGGCGGCGAGGAAAGAT-3′,CaMYB44-R,5′-TCACTCAATTTTGCTGATTCC-3′。以HNUCA54叶片cDNA为模板,使用2×Phanta@Max Master Mix酶PCR扩增序列全长,凝胶电泳后切取正确条带,用诺唯赞的产物纯化试剂盒(编号DC301-01)回收PCR产物,连接TA克隆载体,转化感受态大肠杆菌DH5α。挑取单菌落进行菌液PCR检测,将条带正确且明亮的菌株送海南楠山生物技术有限公司进行测序分析。 -
使用ProtParam软件进行蛋白理化性质分析,使用SignalP、SOPMA、PONDR、TMHMM、Swiss-model等在线网页分析蛋白结构特征。使用DNAMAN、TBtools、Notepad和MEGAX等软件用于多序列比对、蛋白质翻译以及系统发育树构建等数据分析工作。
-
使用cNLS Mapper在线网页对CaMYB44蛋白预测其定位结果,并实验验证。从pCAMBIA1300载体选取酶切位点(Sal I和Kpn I)结合基因序列设计同源臂引物:CaMYB44-F,5′-gttgatacatatgcccgtcgacATGGCGGCGAGGAAAGAT-3′,CaMYB44-R,5′-gctcaccatggatccggtaccCTCAATTTTGCTGATTCC-3′。利用同源重组技术构建pCAMBIA1300-CaMYB44-GFP荧光表达载体,经转化感受态大肠杆菌DH5α,并测序正确后,转入感受态农杆菌GV3101中,用注射针头侵染已生长5周的烟草叶片背面,放置于24 ℃黑暗环境2~3 d,利用激光扫描共聚焦显微镜观察植物叶片背面荧光情况。
-
使用软件Primer5.0从目标基因序列中选取200 bp左右特异片段设计引物:CaMYB44-qF,5′-TTCGGGTCTGGATCTCCCTC-3′;CaMYB44-qR,5′-GTCGGGTACAGATGGAGCAG-3′。内参基因(登录号AY486137.1)引物:Actin-qF,5′-G-CTGGAGGTGTATTTTTGGT-3′;Actin-qR,5′-ATTTC-TGGTACGAGCGGAT-3′。采集辣椒植株在4 ℃处理前,以及4 ℃处理24 、48 、72 h后的叶片,每个时期设置3个生物学重复。提取各时期叶片RNA反转录为cDNA,并以各时期cDNA为模板,设置3个技术性重复,进行RT-qPCR检测,本研究选用2-△△Ct法分析实验数据。
-
根据基因ID,从NCBI官网(
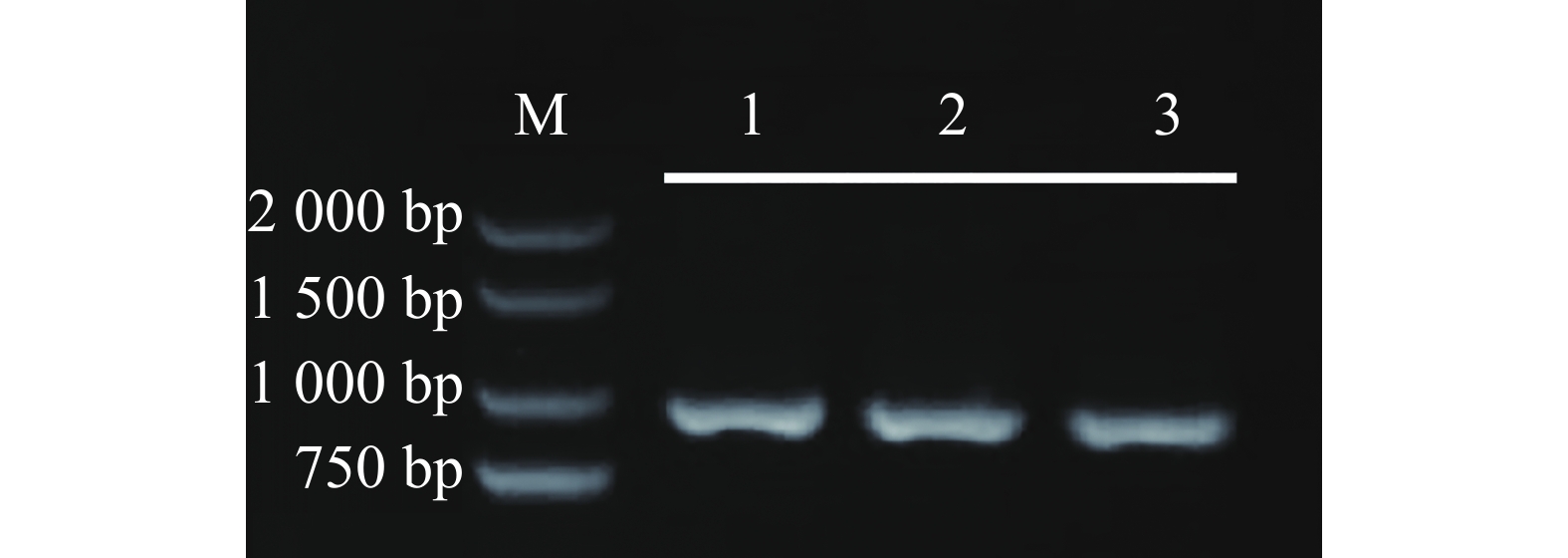
https://www.ncbi.nlm.nih.gov/nuccore/XM_016713322.2/ )下载目标基因CDS序列(981 bp),设计引物对CaMYB44-F和CaMYB44-R,反转录·聚合酶链反应(reverse transcription-polymerase chain reaction,RT-PCR)后,琼脂糖凝胶电泳检测(图1),回收目的条带,利用TA载体进行克隆,转化大肠杆菌,阳性单克隆序列测序结果分析,HNUCA54中的CaMYB44 cDNA与NCBI中参考序列信息一致(图2),提示CaMYB44基因种内保守性强。
图 1 辣椒CaMYB44基因全长的PCR扩增图
Figure 1. The PCR amplification of the full length of CaMYB44 gene in pepper

图 2 CaMYB44 CDS序列与克隆测序序列比对图
Figure 2. Sequence alignment diagram of CaMYB44 CDS sequence and sequence of clone sequencing
-
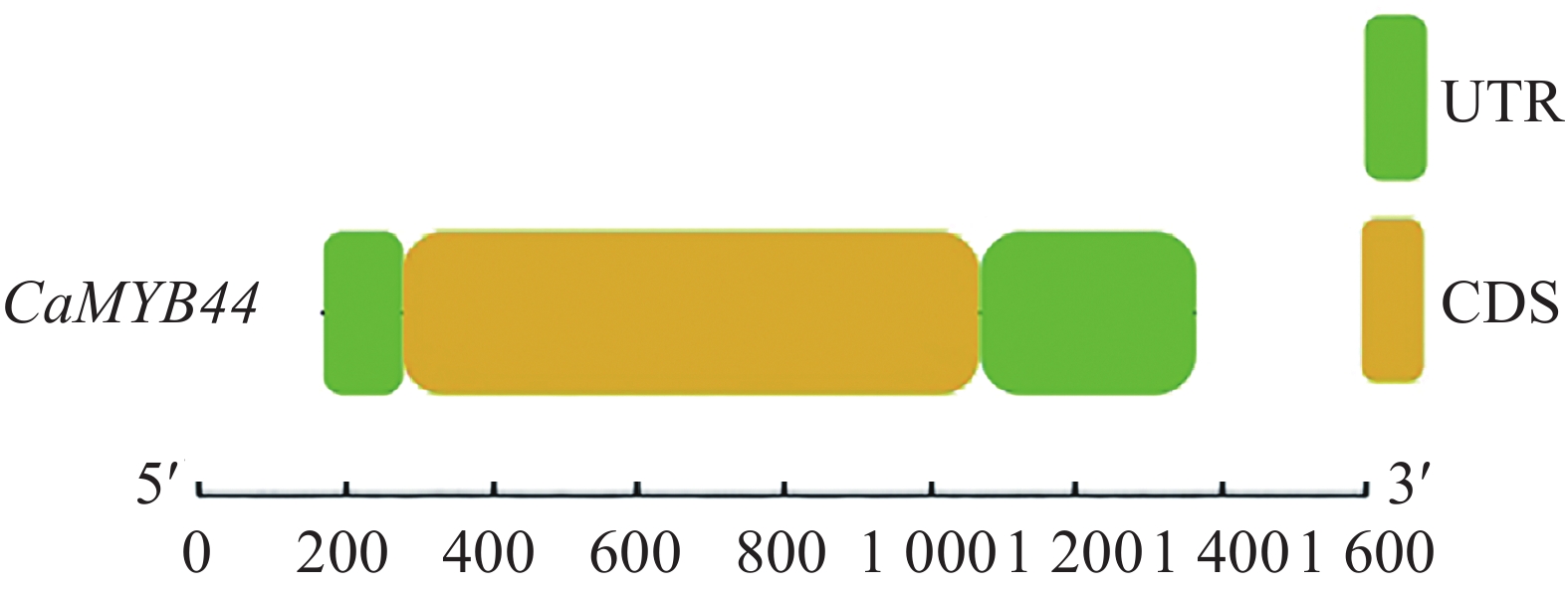
在美国国家生物技术信息中心(National Center for Biotechnology Information,NCBI )中下载 CaMYB44参考序列注释信息,cDNA和基因组序列比对表明,CaMYB44基因无内含子,且由图3可看出基因组序列1~134 bp为5′-UTR区,1 116~1 482 bp为3′-UTR区,共编码326个氨基酸(图4)。使用ProtParam软件分析其蛋白的理化性质,碱性氨基酸残基数多于酸性氨基酸残基数,为碱性蛋白质;不稳定系数大于40,为不稳定蛋白;总平均亲水性小于0为亲水蛋白(表1)。
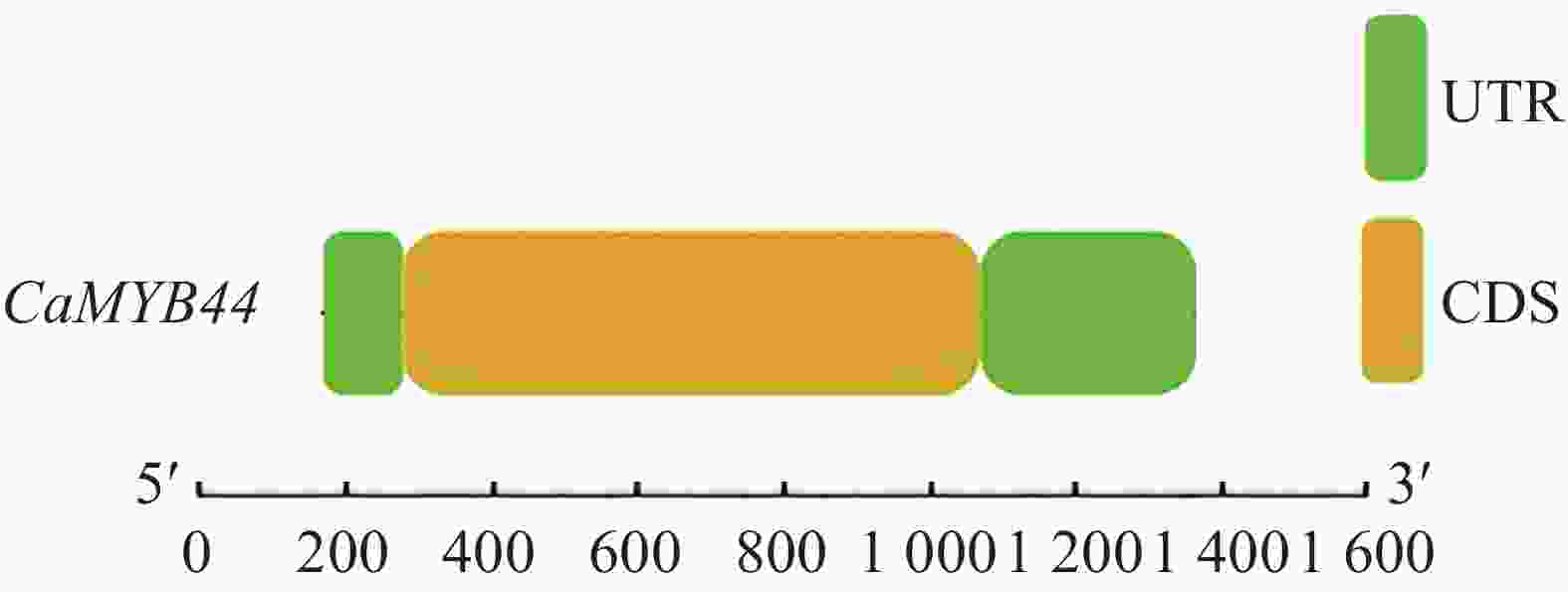
图 3 CaMYB44基因结构图
Figure 3. Gene structure of CaMYB44

图 4 CaMYB44基因编码序列及其氨基酸序列
Figure 4. Coding sequence of CaMYB44 gene and its amino acid sequence
表 1 CaMYB44蛋白理化性质预测
Table 1. Physicochemical properties prediction of CaMYB44 protein
指标 Index CaMYB44 指标 Index CaMYB44 分子式
Molecular formulaC 1577 H2491 N439O469S13碱性氨基酸残基总数/aa
Total number of basic amino acid residues/aa38 相对分子质量
Relative molecular mass35 521.57 半衰期/h
Half-life period/h30 氨基酸数目/aa
Number of amino acids/aa326 不稳定系数
Instability coefficient68.35 理论PI值
Isoelectric point9.31 脂肪系数
Aliphatic index66.10 酸性氨基酸残基总数/aa
Total number of acidic amino acid residues/aa29 总平均亲水性
Total average hydrophobic index−0.548 -
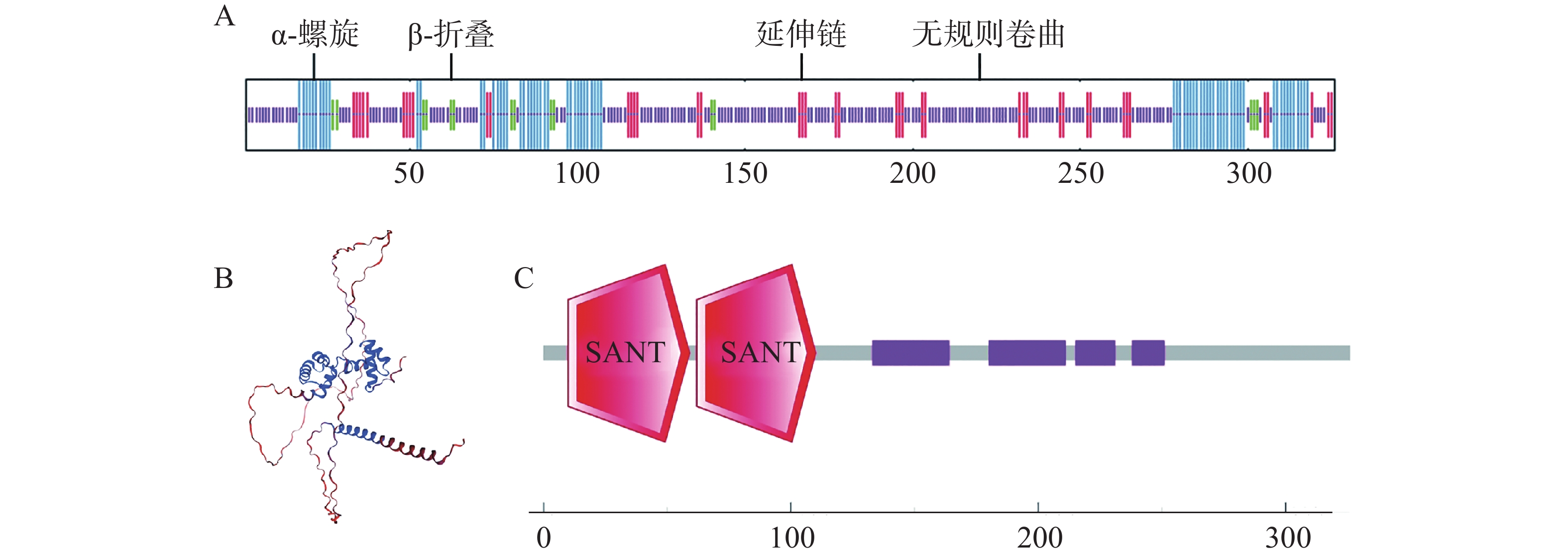
经SignalP在线预测,CaMYB44蛋白序列内不含信号肽,通过跨膜区分析(TMHMM)对氨基酸序列进行预测,可见其编码蛋白无跨膜结构,故CaMYB44非膜蛋白以及分泌蛋白。使用SOPMA在线网页分析CaMYB44蛋白二级结构,发现其结构以无规则卷曲为主,占59.82%,剩下α-螺旋、β-折叠、延伸链分别占22.39%、4.60%、13.19%(图5−A)。PONDR软件预测的无规则卷曲占比为59.82%。使用Swiss-model在线网页预测其三级结构,其三级结构图与二级结构的预测相一致(图5−B)。使用SMART在线网页进行保守结构域分析,可知CaMYB44转录因子属R2R3型MYB(图5−C)。
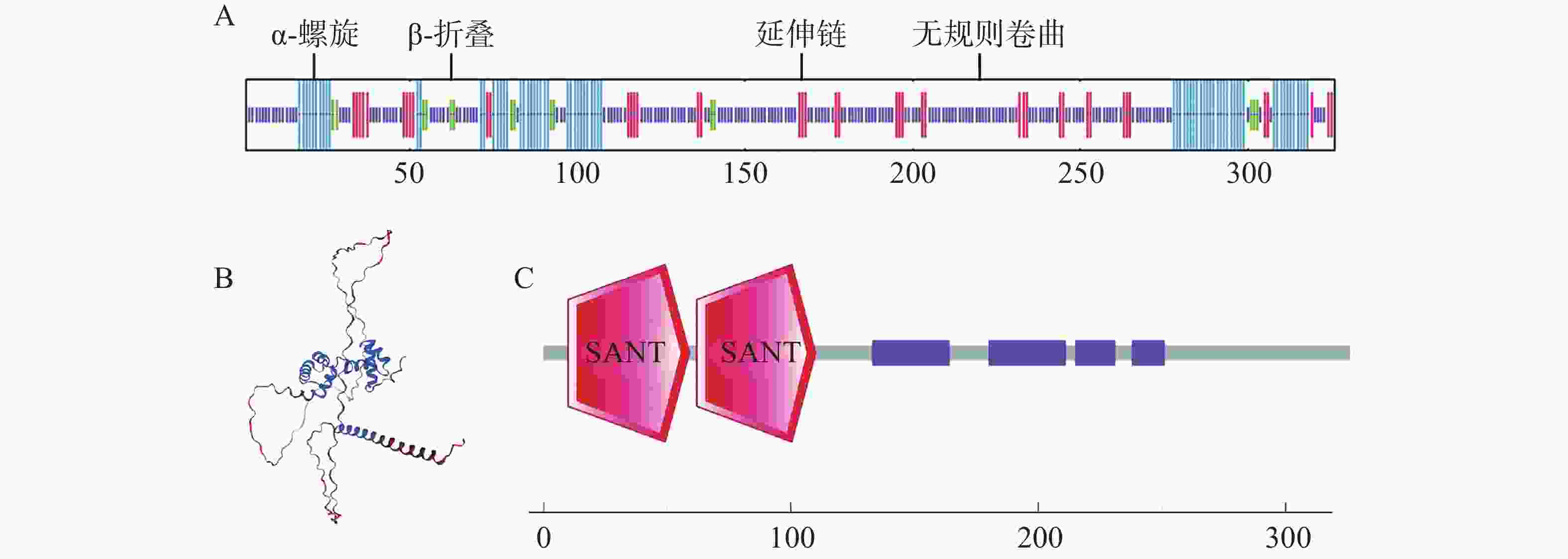
图 5 CaMYB44蛋白结构及保守结构域预测
Figure 5. Protein structure and conserved domain prediction of CaMYB44
-
使用TBtools软件将CaMYB44氨基酸序列与水稻、番茄、土豆、拟南芥、茄子、烟草和其他辣椒栽培种等基因组进行blast比对。从中选出同源性较好的基因,将其蛋白质序列与CaMYB44一起制作系统发育树(图6)。由系统发育树结果可得CaMYB44与番茄、马铃薯等茄科作物的MYB44、MYB44-like同源关系较近,处于同一分支,其次与拟南芥MYB70以及水稻的MYB77等同源关系也较好。
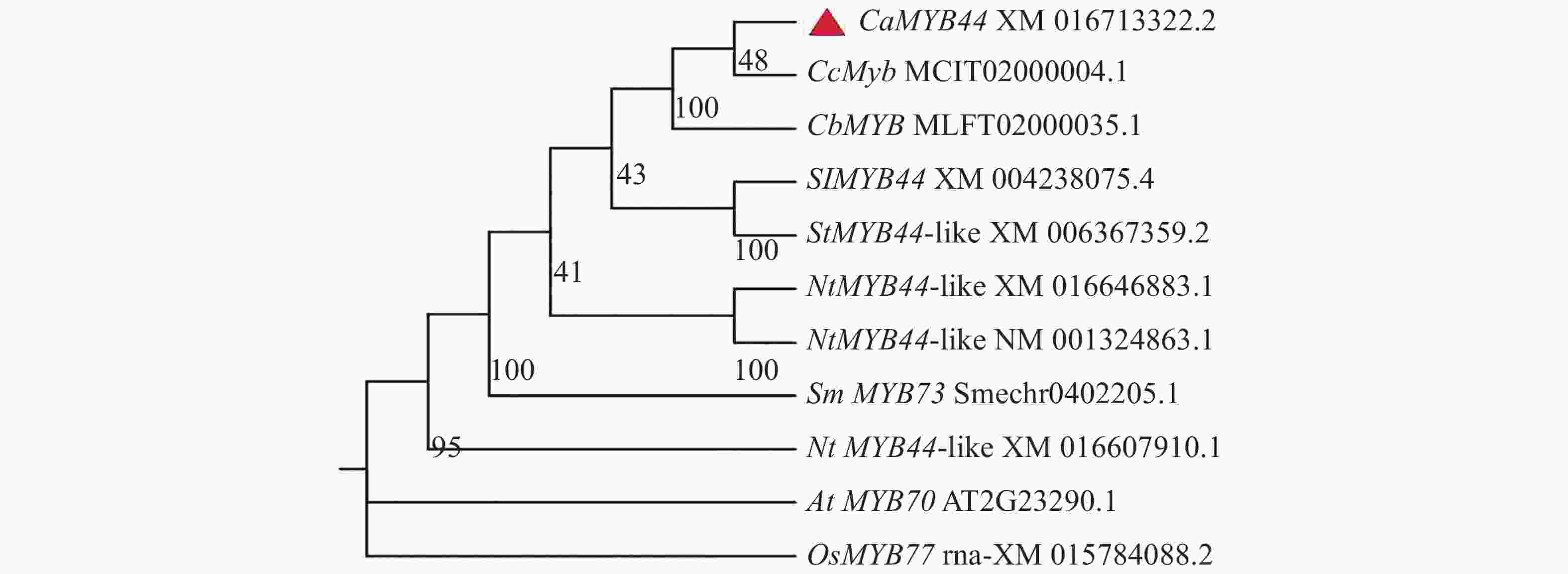
图 6 辣椒CaMYB44与其他物种MYB转录因子的进化关系
Figure 6. Evolutionary relationship between CaMYB44 and other MYB transcription factors in pepper
-
用TBtools软件提取CaMYB44基因上游2 000 bp序列,上传PlantCARE在线网页,预测基因启动子顺式作用元件(表2)。结果显示,CaMYB44启动子区域含有22个参与光响应的元件22个,参与脱落酸响应的元件8个,参与低温响应的LTR元件1个。
表 2 CaMYB44启动子顺式作用元件预测
Table 2. Prediction of cis-acting elements of CaMYB44 promoter
顺式作用元件
Cis-acting
element序列
Sequence功能
Function数量
QuantityLTR CCGAAA 参与低温响应
Participate in low temperature response1 CGTCA-motif CGTCA 参与茉莉酸甲酯响应
Involved in the response of methyl jasmonate3 ABRE ACGTG/CACGTG/GCCGCGTGGC/
CGTACGTGCA/AACCCGG参与脱落酸响应
Involvement in abscisic acid response8 TGACG-motif TGACG 参与茉莉酸甲酯响应
Involved in the response of methyl jasmonate3 Box 4 ATTAAT 参与光响应的保守DNA模块的一部分
Part of a conserved DNA module involved in light response5 G-box TACGTG/CACGTC/CACGAC/CACGTG/
GCCACGTGGA/CACGTT参与光响应
Participate in light response7 TCT-motif TCTTAC 光响应模块的一部分
Part of the light response module1 TGA-element AACGAC 参与生长素响应
Involvement in auxin response1 TCA-element CCATCTTTTT 参与水杨酸响应
Involved in salicylic acid response1 AE-box AGAAACAA 光响应模块的一部分
Part of the Light Response Module1 ATCT-motif AATCTAATCC 参与光响应的保守DNA模块的一部分
Part of a conserved DNA module involved in light response1 MBS CAACTG 参与干旱诱导的MYB结合位点
MYB binding sites involved in drought induction1 GATA-motif GATAGGG/GATAGGA 光响应模块的一部分
Part of the light response module1 GT1-motif GGTTAA 参与光响应
Participate in light response1 ACE CTAACGTATT 参与光响应的顺式作用元件
cis-acting elements involved in light response1 ATC-motif TGCTATCCA 参与光响应的保守DNA模块的一部分
Part of a conserved DNA module involved in light response1 Box Ⅱ CCACGTGGC 光响应元件的一部分
Part of the optical response element2 MRE AACCTAA 参与光响应的MYB结合位点
MYB binding sites involved in light response1 -
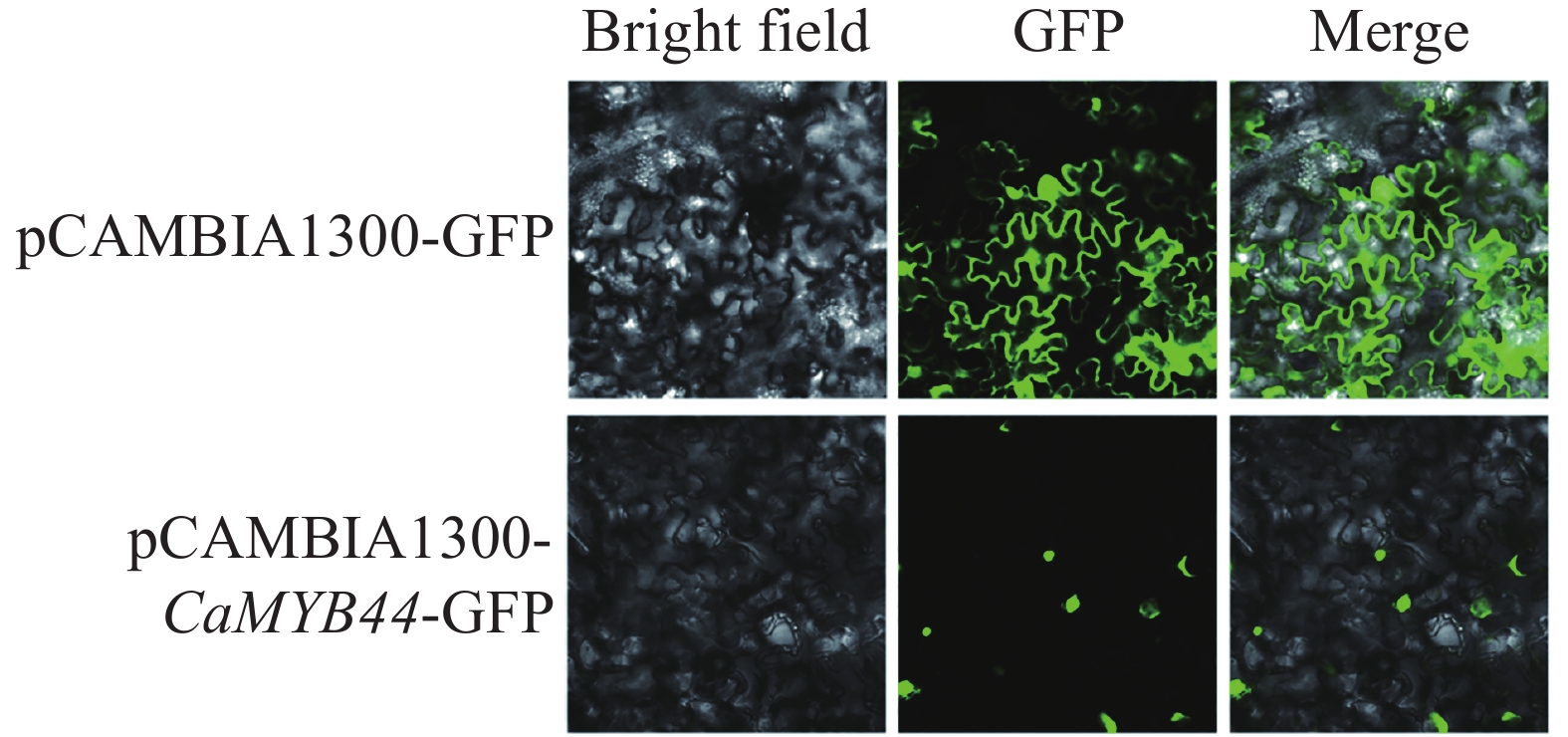
使用cNLS Mapper在线网页对CaMYB44蛋白进行预测,发现其存在核定位信号,预测该蛋白定位于核中。将CaMYB44序列用同源重组的方式,构建荧光表达载体pCAMBIA1300-MYB44-GFP,侵染烟草叶片,激光共聚焦显微镜检测荧光信号,观察发现CaMYB44蛋白定位在细胞核,进一步提示CaMYB44在细胞核发挥作用(图7)。
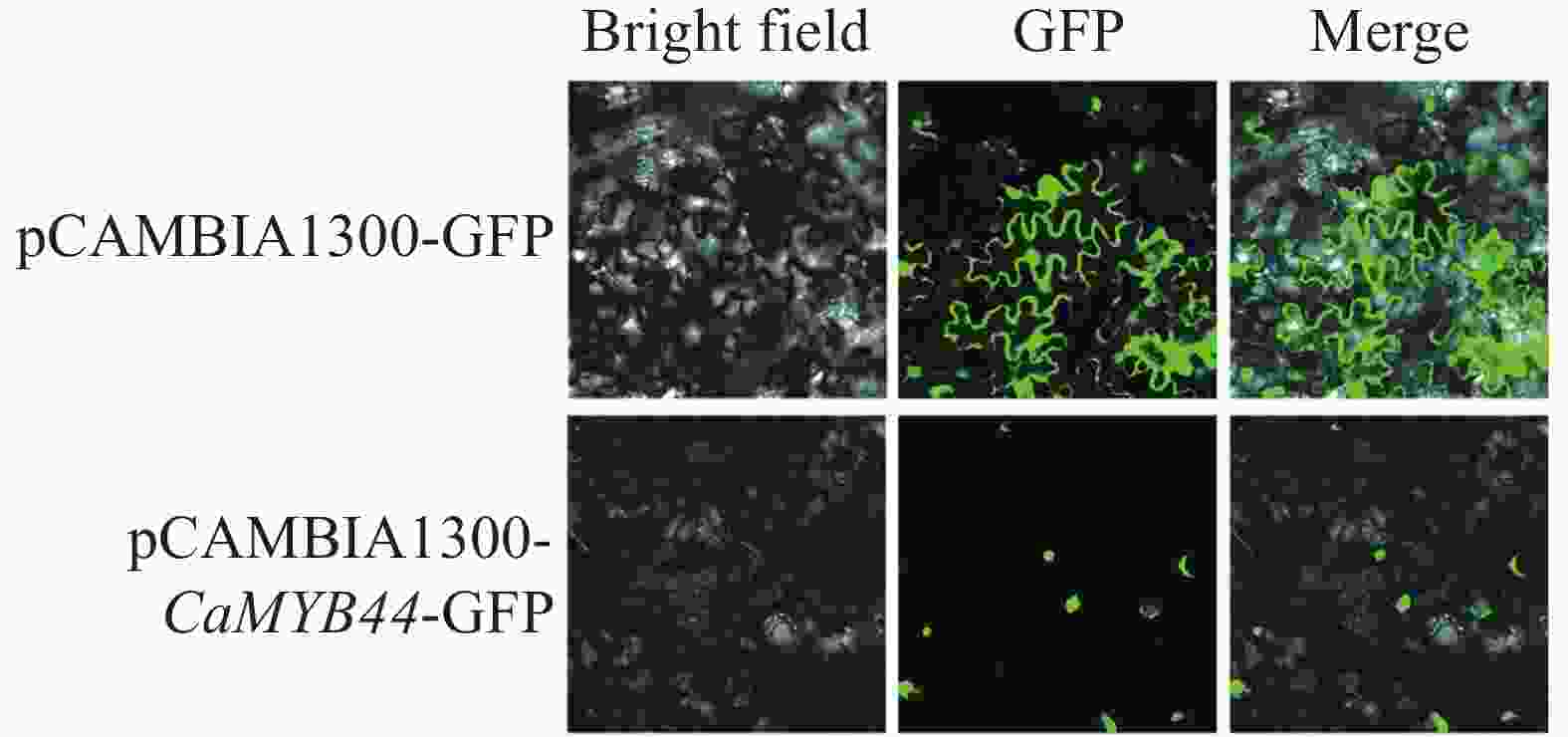
图 7 CaMYB44基因的亚细胞定位
Figure 7. Subcellular localization of CaMYB44 gene
-
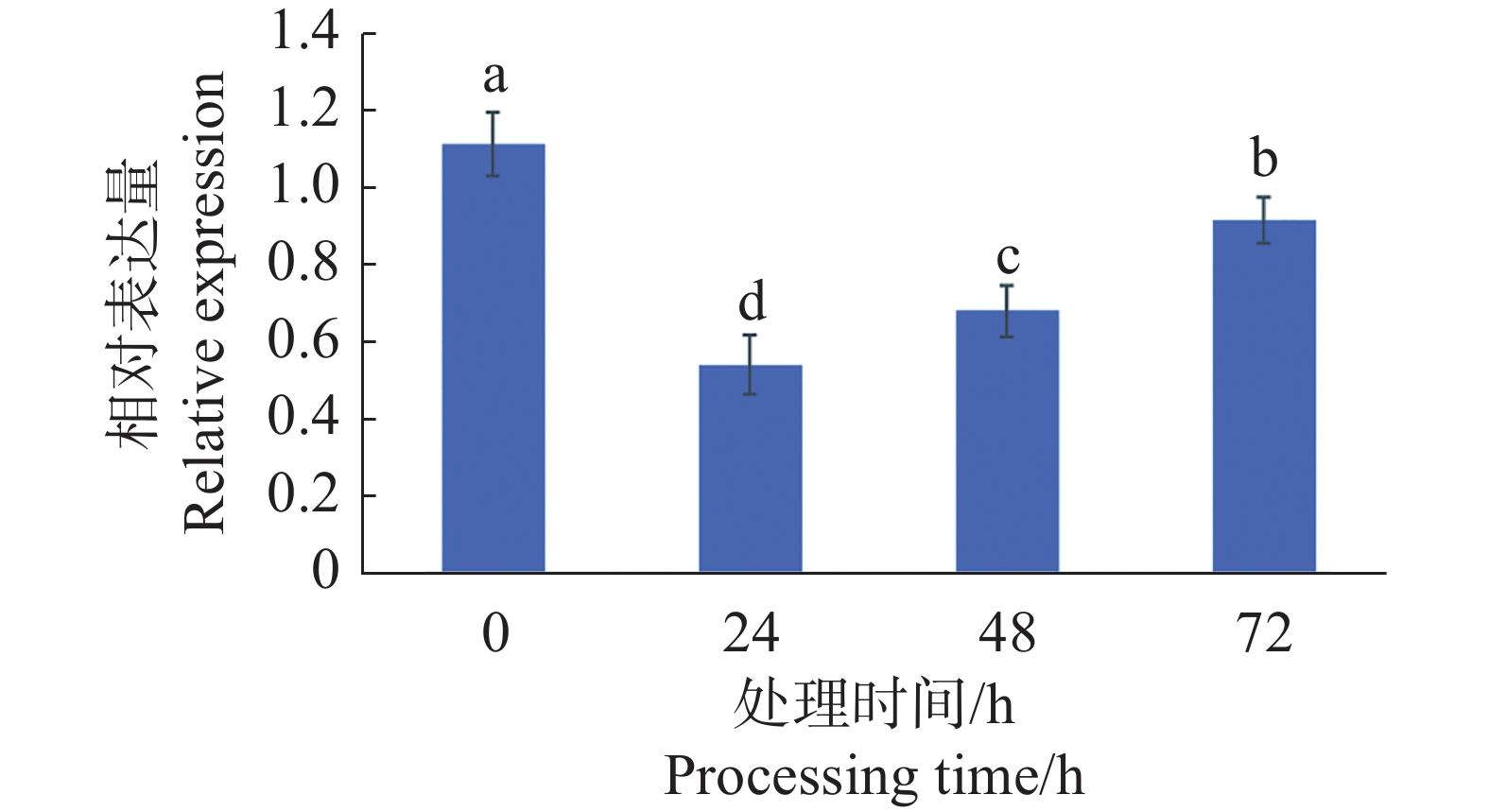
对4 ℃处理0、24、48、72 h的辣椒植株叶片cDNA进行RT-qPCR分析。可见CaMYB44在4 ℃低温胁迫处理24 h后,在辣椒叶片中的表达量急剧下调,后随着处理时间的增加,表达量呈缓慢上升的趋势(图8)。
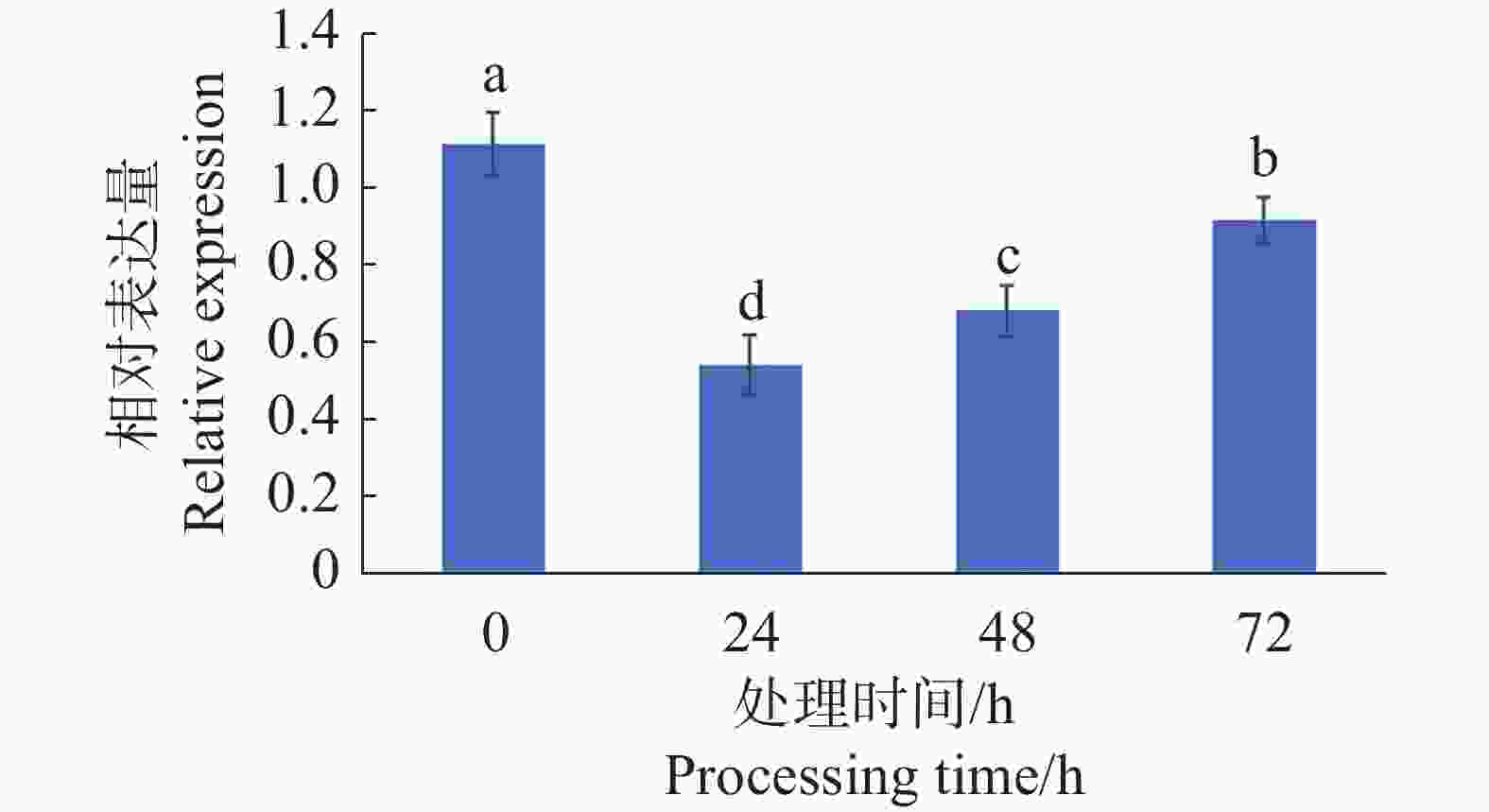
图 8 CaMYB44基因在低温处理下的表达变化
Figure 8. The expression changes of CaMYB44 gene under low temperature treatment
-
一年生辣椒在中国栽培广泛,具有极高的食用价值,而低温限制了其种植时段和区域,影响产量和质量。MYB作为在众多植物中广泛存在的转录因子,参与植物各种生理、生化过程,其中包括对逆境胁迫等基因的调控表达。本研究以本团队前期比较组学结果中响应外环境胁迫的大量MYB转录因子为基础,从中克隆了CaMYB44基因,其CDS序列与NCBI所得序列信息一致。通过生物信息学分析,CaMYB44属不稳定亲水性蛋白,且无信号肽与跨膜结构,故为非分泌蛋白。经预测发现,CaMYB44蛋白无规则卷曲占59.82%,占比较高,无规则卷曲是蛋白质的柔性连接区域,允许蛋白质进行必要的动态变化,并与其他蛋白质、核酸和小分子结合,调节蛋白功能、控制核膜稳态等,在细胞功能和调控中发挥重要作用[19 − 20]。亚细胞定位结果显示,CaMYB44定位于细胞核中,在细胞核发挥作用。
通过系统发育树分析CaMYB44与茄科物种MYB44、MYB44-like存在较高的同源关系,与拟南芥MYB70及水稻MYB77也存在较好的同源关系。有研究发现,在葡萄中过表达MYB44基因会使其植株表现出更严重的低温胁迫损伤[21]。Persak等[22]通过酵母双杂实验,发现拟南芥应激诱导的MYB44基因表达与丝裂原活化蛋白激酶进化枝MPK3相互作用,启动对多种非生物和生物胁迫的适应反应。AtMYB70与AtMYB44共处于S22亚族,在调控功能上可能相类似,Kagale等[23]也在转录组分析后推测AtMYB70通过负调控参与发育、激素和应激信号转导通路的基因,在多种生物学功能中发挥关键作用。故本研究团队推测CaMYB44转录因子可能同样在辣椒抵抗低温胁迫过程中发挥调控作用。
根据对CaMYB44基因启动子预测结果,LTR元件赋予低温反应基因的低温诱导活性,冬性甘蓝型油菜冷诱导基因BN115低温响应元件LTRE的核心五聚体突变,使得启动子低温调节的丧失[24]。经预测CaMYB44基因启动子区含有LTR基序CCGAAA,故推测该基因可能对低温胁迫环境产生响应。
从表2可见CaMYB44基因启动子区域除含LTR元件外还含有多个响应脱落酸(ABA)和光的元件。ABA可刺激植物适应胁迫环境,使其在生理、分子和发育进程等方面发生改变[25]。在大豆中,过表达受ABA诱导的GmZF1转基因拟南芥,在低温环境中,有更好的耐受性[26]。光照根据时长、亮度以及光质等多方面调控影响着植物的生长发育,同时影响植物在不同温度环境下的适应性,合适的光照刺激可提高植株对于低温胁迫的抗性[27]。低红光与远红光的光信号能够通过影响生物钟机制,提升拟南芥中CBF基因表达水平,这种由光质驱动的CBF表达增强,有效增强了植物的耐低温能力[28]。故可推测CaMYB44基因也可能受脱落酸或光照等途径诱导响应低温胁迫环境。
MYB44已在多个物种中被验证为植株低温胁迫响应的调控因子,例如在苹果愈伤组织中过表达MdMYB44基因,会使其低温耐受性显著降低[29];在黄瓜中,CsMYB44可通过转录激活PAs基因,减轻黄瓜果实冻害[30]。本研究中RT-qPCR结果显示,CaMYB44在4 ℃低温胁迫处理24 h后,表达显著下调,在48 h处理后又逐渐上调。与葡萄VaMYB44基因及红枣ZjCBL1基因在低温胁迫下的表达趋势相类似,低温处理后基因表达先下调后恢复原表达水平[21, 31]。故本研究团队推测CaMYB44基因可能在辣椒抵抗低温胁迫中发挥负调控作用。
一年生辣椒CaMYB44基因功能的初步解析
DOI: 10.15886/j.cnki.rdswxb.20240067
 CSTR: 32425.14.j.cnki.rdswxb.20240067
CSTR: 32425.14.j.cnki.rdswxb.20240067
Analysis of the function of the CaMYB44 gene in Capsicum annuum
-
摘要: MYB(v-myb avian myeloblastosis viral oncogene homolog)转录因子家族在植物应对环境胁迫中发挥重要作用。一年生辣椒(Capsicum annuum)是辣椒属种植范围最广的一个栽培种。通过分析前期研究得到的转录组数据,聚焦CaMYB44转录因子,并对其基因启动子顺式作用元件、蛋白结构、进化关系、亚细胞定位以及低温响应等进行生物信息学分析与初步实验验证。CaMYB44基因开放阅读框(Open Reading Frame,ORF)长度为981 bp,编码氨基酸为326 aa,不稳定的亲水性蛋白。经预测,CaMYB44蛋白上无信号肽以及跨膜结构域为非分泌蛋白,属于R2R3型MYB。系统发育树分析结果显示,CaMYB44基因与SmMYB73、NtMYB44-like、StMYB44-like和SlMYB44同源性较高,其次为AtMYB70、OsMYB77。顺式作用元件分析结果显示,CaMYB44基因启动子区包含1个低温响应元件以及多个脱落酸和光响应元件。亚细胞定位结果表明CaMYB44定位于细胞核。实时荧光定量反转录·聚合酶链反应(real time fluorescent quantitative reverse transcription-polymerase chain reaction,RT-qPCR)结果显示,CaMYB44的表达量在4 ℃低温胁迫24 h后下调,与其他低温胁迫负调控因子在处理下的表达趋势相似。研究结果为进一步解析一年生辣椒中耐低温胁迫转录因子提供了基础数据。Abstract: The MYB transcription factor family plays an important role in plant response to environmental stress. Capsicum annuum is the most widely cultivated species in the genus Capsicum. The transcriptome data obtained by our group were analyzed, based on which MYB44 transcription factor was selected for bioinformatics analysis of the cis-acting elements of the promoter, protein structure, evolutionary relationship. A preliminary experimental research was then conducted on subcellular localization and low temperature response. The open reading frame (ORF) of CaMYB44 gene is 981 bp in length, encoding an unstable hydrophilic protein with an amino acid of 326 aa. It is predicted that CaMYB44 protein has no signal peptide and transmembrane domain, is a non-secretory protein, and belongs to R2R3 MYB. Phylogenetic tree analysis showed that it had high homology with SmMYB73, NtMYB44-like, StMYB44-like and SlMYB44, followed by AtMYB70 and OsMYB77. Cis-acting element analysis showed that the promoter region contained a low temperature response element and multiple abscisic acid and light response elements. Subcellular localization results showed that CaMYB44 was localized in the nucleus. The RT-qPCR showed that the expression of CaMYB44 was down-regulated after 24 h of low temperature stress at 4 ℃, which was similar to the expression trend of other negative regulatory factors under low temperature stress. This paper provides basic data for further analysis of low temperature tolerance transcription factors in C. annuum.
-
Key words:
- Capsicum annuum /
- CaMYB44 /
- gene cloning /
- expression analysis
-
图 2 CaMYB44 CDS序列与克隆测序序列比对图
Fig. 2 Sequence alignment diagram of CaMYB44 CDS sequence and sequence of clone sequencing
图 4 CaMYB44基因编码序列及其氨基酸序列
Fig. 4 Coding sequence of CaMYB44 gene and its amino acid sequence
图 5 CaMYB44蛋白结构及保守结构域预测
A.蛋白质二级结构;B.蛋白质三级结构;C.保守结构域。
Fig. 5 Protein structure and conserved domain prediction of CaMYB44
A.Protein secondary structure;B.Protein tertiary structure;C.Conserved domain.
图 6 辣椒CaMYB44与其他物种MYB转录因子的进化关系
Fig. 6 Evolutionary relationship between CaMYB44 and other MYB transcription factors in pepper
图 8 CaMYB44基因在低温处理下的表达变化
不同小写字母表示差异显著(P < 0.05)。
Fig. 8 The expression changes of CaMYB44 gene under low temperature treatment
Different lowercase letters indicate significant difference at P < 0.05.
表 1 CaMYB44蛋白理化性质预测
Table 1 Physicochemical properties prediction of CaMYB44 protein
指标 Index CaMYB44 指标 Index CaMYB44 分子式
Molecular formulaC 1577 H2491 N439O469S13碱性氨基酸残基总数/aa
Total number of basic amino acid residues/aa38 相对分子质量
Relative molecular mass35 521.57 半衰期/h
Half-life period/h30 氨基酸数目/aa
Number of amino acids/aa326 不稳定系数
Instability coefficient68.35 理论PI值
Isoelectric point9.31 脂肪系数
Aliphatic index66.10 酸性氨基酸残基总数/aa
Total number of acidic amino acid residues/aa29 总平均亲水性
Total average hydrophobic index−0.548  下载: 导出CSV
下载: 导出CSV
表 2 CaMYB44启动子顺式作用元件预测
Table 2 Prediction of cis-acting elements of CaMYB44 promoter
顺式作用元件
Cis-acting
element序列
Sequence功能
Function数量
QuantityLTR CCGAAA 参与低温响应
Participate in low temperature response1 CGTCA-motif CGTCA 参与茉莉酸甲酯响应
Involved in the response of methyl jasmonate3 ABRE ACGTG/CACGTG/GCCGCGTGGC/
CGTACGTGCA/AACCCGG参与脱落酸响应
Involvement in abscisic acid response8 TGACG-motif TGACG 参与茉莉酸甲酯响应
Involved in the response of methyl jasmonate3 Box 4 ATTAAT 参与光响应的保守DNA模块的一部分
Part of a conserved DNA module involved in light response5 G-box TACGTG/CACGTC/CACGAC/CACGTG/
GCCACGTGGA/CACGTT参与光响应
Participate in light response7 TCT-motif TCTTAC 光响应模块的一部分
Part of the light response module1 TGA-element AACGAC 参与生长素响应
Involvement in auxin response1 TCA-element CCATCTTTTT 参与水杨酸响应
Involved in salicylic acid response1 AE-box AGAAACAA 光响应模块的一部分
Part of the Light Response Module1 ATCT-motif AATCTAATCC 参与光响应的保守DNA模块的一部分
Part of a conserved DNA module involved in light response1 MBS CAACTG 参与干旱诱导的MYB结合位点
MYB binding sites involved in drought induction1 GATA-motif GATAGGG/GATAGGA 光响应模块的一部分
Part of the light response module1 GT1-motif GGTTAA 参与光响应
Participate in light response1 ACE CTAACGTATT 参与光响应的顺式作用元件
cis-acting elements involved in light response1 ATC-motif TGCTATCCA 参与光响应的保守DNA模块的一部分
Part of a conserved DNA module involved in light response1 Box Ⅱ CCACGTGGC 光响应元件的一部分
Part of the optical response element2 MRE AACCTAA 参与光响应的MYB结合位点
MYB binding sites involved in light response1
下载: 导出CSV
-
[1] 邹学校, 朱凡. 辣椒的起源、进化与栽培历史[J]. 园艺学报, 2022, 49(6): 1371 − 1381. doi: 10.16420/j.issn.0513-353x.2021-0853 [2] 邹学校, 马艳青, 戴雄泽, 等. 辣椒在中国的传播与产业发展[J]. 园艺学报, 2020, 47(9): 1715 − 1726. doi: 10.16420/j.issn.0513-353x.2020-0103 [3] 李涛, 徐小万, 李颖, 等. 一年生辣椒(Cpsicum annuum L.)与中华辣椒(Cpsicum chinense Jacquin)DNA甲基化多样性分析[J]. 分子植物育种, 2014, 12(2): 306 − 315. doi: 10.13271/j.mpb.012.000306 [4] RIHAN H Z, AL-ISSAWI M, FULLER M P. Advances in physiological and molecular aspects of plant cold tolerance[J]. Journal of Plant Interactions, 2017, 12(1): 143 − 157. doi: 10.1080/17429145.2017.1308568 [5] Steponkus P L, Webb M S. Freeze-induced dehydration and membrane destabilization in plants[M]//Water and Life: Comparative Analysis of Water Relationships at the Organismic, Cellular, and Molecular Levels. Berlin, Heidelberg: Springer Berlin Heidelberg, 1992: 338−362. [6] ZHANG S, JIANG H, PENG S M, et al. Sex-related differences in morphological, physiological, and ultrastructural responses of Populus cathayana to chilling[J]. Journal of Experimental Botany, 2011, 62(2): 675 − 686. doi: 10.1093/jxb/erq306 [7] ROHDE P, HINCHA D K, HEYER A G. Heterosis in the freezing tolerance of crosses between two Arabidopsis thaliana accessions (Columbia‐0 and C24) that show differences in non‐acclimated and acclimated freezing tolerance[J]. The Plant Journal, 2004, 38(5): 790 − 799. doi: 10.1111/j.1365-313X.2004.02080.x [8] LIU Z B, SONG J S, MIAO W, et al. Comprehensive proteome and lysine acetylome analysis reveals the widespread involvement of acetylation in cold resistance of pepper (Capsicum annuum L.)[J]. Frontiers in Plant Science, 2021, 12: 730489. doi: 10.3389/fpls.2021.730489 [9] 邓惠如, 张素勤, 耿广东. 辣椒逆境响应基因研究现状及前景[J/OL]. 分子植物育种, (2023-11-09)[2024-04-12]. http://kns.cnki.net/kcms/detail/46.1068.S.20231108.1149.002.html. [10] 朱晨曦, 马艳青, 张竹青. 我国辣椒耐低温弱光的研究概况[J]. 安徽农业科学, 2013, 41(15): 6581 − 6583. doi: 10.3969/j.issn.0517-6611.2013.15.001 [11] YAO C Y, LI X G, LI Y M, et al. Overexpression of a Malus baccata MYB transcription factor gene MbMYB4 increases cold and drought tolerance in Arabidopsis thaliana[J]. International Journal of Molecular Sciences, 2022, 23(3): 1794. doi: 10.3390/ijms23031794 [12] MA X, YU Y N, JIA J H, et al. The pepper MYB transcription factor CaMYB306 accelerates fruit coloration and negatively regulates cold resistance[J]. Scientia Horticulturae, 2022, 295: 110892. doi: 10.1016/j.scienta.2022.110892 [13] DONG J, CAO L, ZHANG X Y, et al. An R2R3-MYB transcription factor RmMYB108 responds to chilling stress of Rosa multiflora and conferred cold tolerance of Arabidopsis[J]. Frontiers in Plant Science, 2021, 12: 696919. doi: 10.3389/fpls.2021.696919 [14] SONG X P, ZHU L, WANG D, et al. Molecular regulatory mechanism of exogenous hydrogen sulfide in alleviating low-temperature stress in pepper seedlings[J]. International Journal of Molecular Sciences, 2023, 24(22): 16337. doi: 10.3390/ijms242216337 [15] LI N H, PU K G, DING D X, et al. Foliar spraying of glycine betaine alleviated growth inhibition, photoinhibition, and oxidative stress in pepper (Capsicum annuum L.) seedlings under low temperatures combined with low light[J]. Plants, 2023, 12(13): 2563. doi: 10.3390/plants12132563 [16] ALTAF M A, SHU H Y, HAO Y Y, et al. Melatonin affects the photosynthetic performance of pepper (Capsicum annuum L.) seedlings under cold stress[J]. Antioxidants, 2022, 11(12): 2414. doi: 10.3390/antiox11122414 [17] ZHOU Y, MUMTAZ M A, ZHANG Y H, et al. Response of anthocyanin accumulation in pepper (Capsicum annuum) fruit to light days[J]. International Journal of Molecular Sciences, 2022, 23(15): 8357. doi: 10.3390/ijms23158357 [18] HULSE-KEMP A M, MAHESHWARI S, STOFFEL K, et al. Reference quality assembly of the 3.5-Gb genome of Capsicum annuum from a single linked-read library[J]. Horticulture Research, 2018, 5(1): 4. doi: 10.1038/s41438-017-0011-0 [19] HAUSMANN S, GEISER J, ALLEN G E, et al. Intrinsically disordered regions regulate RhlE RNA helicase functions in bacteria[J]. Nucleic Acids Research, 2024, 52(13): 7809 − 7824. doi: 10.1093/nar/gkae511 [20] HIRANO Y, SATO T, MIURA A, et al. Disordered region of nuclear membrane protein Bqt4 recruits phosphatidic acid to the nuclear envelope to maintain its structural integrity[J]. Journal of Biological Chemistry, 2024, 300(7): 107430. doi: 10.1016/j.jbc.2024.107430 [21] ZHANG H J, HU Y F, GU B, et al. VaMYB44 transcription factor from Chinese wild Vitis amurensis negatively regulates cold tolerance in transgenic Arabidopsis thaliana and V. vinifera[J]. Plant Cell Reports, 2022, 41(8): 1673 − 1691. doi: 10.1007/s00299-022-02883-w [22] PERSAK H, PITZSCHKE A. Tight interconnection and multi-level control of Arabidopsis MYB44 in MAPK cascade signalling[J]. PLoS One, 2013, 8(2): e57547. doi: 10.1371/journal.pone.0057547 [23] KAGALE S, LINKS M G, ROZWADOWSKI K. Genome-wide analysis of ethylene-responsive element binding factor-associated amphiphilic repression motif-containing transcriptional regulators in Arabidopsis[J]. Plant Physiology, 2010, 152(3): 1109 − 1134. doi: 10.1104/pp.109.151704 [24] JIANG C, IU B, SINGH J. Requirement of a CCGAC cis-acting element for cold induction of the BN115 gene from winter Brassica napus[J]. Plant Molecular Biology, 1996, 30(3): 679 − 684. doi: 10.1007/BF00049344 [25] TON J, FLORS V, MAUCH-MANI B. The multifaceted role of ABA in disease resistance[J]. Trends in Plant Science, 2009, 14(6): 310 − 317. doi: 10.1016/j.tplants.2009.03.006 [26] YU G H, JIANG L L, MA X F, et al. A soybean C2H2-type zinc finger gene GmZF1 enhanced cold tolerance in transgenic Arabidopsis[J]. PLoS One, 2014, 9(10): e109399. doi: 10.1371/journal.pone.0109399 [27] JIANG B C, SHI Y T, PENG Y, et al. Cold-induced CBF-PIF3 interaction enhances freezing tolerance by stabilizing the phyB thermosensor in Arabidopsis[J]. Molecular Plant, 2020, 13(6): 894 − 906. doi: 10.1016/j.molp.2020.04.006 [28] FRANKLIN K A, WHITELAM G C. Light-quality regulation of freezing tolerance in Arabidopsis thaliana[J]. Nature Genetics, 2007, 39(11): 1410 − 1413. doi: 10.1038/ng.2007.3 [29] WU R G, WANG Y, WU T, et al. Functional characterisation of MdMYB44 as a negative regulator in the response to cold and salt stress in apple calli[J]. The Journal of Horticultural Science and Biotechnology, 2018, 93(4): 347 − 355. doi: 10.1080/14620316.2017.1373038 [30] MADEBO M P, BOKHARY S U F, YOU W L, et al. Melatonin improves cold storage tolerance in cucumber via CsMYB44-mediated transcriptional activation of the polyamine biosynthesis gene family[J]. Postharvest Biology and Technology, 2024, 213: 112937. doi: 10.1016/j.postharvbio.2024.112937 [31] QI C F, WANG Q F, NIU Y H, et al. Characteristics of ZjCIPKs and ZjbHLH74-ZjCIPK5 regulated cold tolerance in jujube[J]. International Journal of Biological Macromolecules, 2024, 264: 130429. doi: 10.1016/j.ijbiomac.2024.130429 -

 点击查看大图
点击查看大图
计量
- 文章访问数: 1120
- HTML全文浏览量: 564
- PDF下载量: 11
- 被引次数: 0
