

 下载:
下载:
-
水角(Hydrocera triflora(L.)Wight & Arn)属于凤仙花科水角属,是该属单种的多年生水生草本植物[1]。水角主要生长在沼泽地、湖边、水稻田中,在印度、斯里兰卡、泰国、中国等国家分布。水角的花左右对称,呈粉红色,花期长达9个月,具有较高的观赏价值。在中国,水角主要分布在海南省海口市的羊山湿地[2]。根据植物智的网站(
http://www.iplant.cn )和《中国生物多样性红色名录—高等植物卷》公布的信息,水角已处于地区绝灭状态(regional extinction, RE)。水角耐旱性差是导致其濒危的重要因素,在水位降低、泥土含水量减少的情况下,其极易失水枯萎死亡[2]。遗传转化技术是植物遗传改良的有效手段,是解决农业生产中存在问题有效的技术方法。基于植物组织培养的根癌农杆菌(Agrobacterium tumefaciens)介导的遗传转化方法被广泛应用,并在玉米(Zea mays)[3]、水稻(Oryza sativa)[4]、高粱(Sorghum bicolor)[5]等植物中取得良好的效果。但该方法依赖于植物的高效、成熟的再生体系,同时存在转化率低、耗时长、实验成本高等缺点,严重制约了一些植物的性状改良及重要基因功能的研究进展。近年来,发根农杆菌介导转化产生的转基因毛状根与野生型地上部组成的转基因复合植物的技术得到了发展。该技术不依赖植物组织培养,具有操作简单、非基因型依赖、转化率高、耗时短和成本低等优势,逐渐被广泛应用于植物性状改良和基因功能研究。Aggarwal等[6]建立了鹰嘴豆(Cicer arietinum)基因型依赖程度低的发根农杆菌介导的转化体系,发现在过表达AtTT2基因的转基因鹰嘴豆复合植株的毛状根中,原花青素水平得到显著提高。Chen等[7]用发根农杆菌侵染大豆(Glycine max)种子下胚轴,获得转基因大豆复合植株,在该转基因复合植株的毛状根中验证了Pi转运蛋白基因GmPT7的功能。Fan等[8]建立了大豆一步发根农杆菌介导转化方法,通过CRISPR/Cas9基因编辑系统在转基因的毛状根中验证了Rfg1基因功能,同时还在转基因的毛状根中验证了YAO启动子的功能。Wang等[9]建立了柱花草发根农杆菌介导转化技术,在转基因柱花草复合植株的毛状根中证实了SgEXPB1基因在磷缺乏的条件下能促进根的生长。
海藻糖是由2个葡萄糖分子通过1,1-糖苷键而成的非还原性双糖[10],并广泛存在于植物、动物、真菌、细菌及藻类当中[11]。海藻糖对提高植物耐旱性、耐寒性等非生物胁迫起到重要作用[12 − 13]。在海藻糖生物合成过程中,海藻糖-6-磷酸合成酶(trehalose-6-phosphate synthase, TPS)是一个调控其生物合成的关键酶[12 − 13]。绝大部分植物的海藻糖含量极低,但可通过调控TPS基因的表达从而达到提高其海藻糖含量的目的。过表达拟南芥AtTPS基因的转基因烟草(Nicotiana tabacum)其海藻糖含量得到显著提高,具有较强的耐受盐胁迫能力[14]。过表达水稻OsTPS1的转基因水稻的海藻糖含量显著高于野生型,其耐寒、耐盐、耐干旱的能力也得到改善[15]。过表达木薯MeTPS1的转基因烟草积累了更高水平的海藻糖,其耐旱能力也得到明显提高[16]。由此可知,提高植物TPS基因表达水平可以增强其耐旱、耐寒等非生物的胁迫的能力。
目前尚未有水角的遗传转化研究报道。本研究拟应用发根农杆菌介导法转化水角,建立水角转基因技术体系,同时构建过表达MeTPS6基因的载体转化水角,以期获得耐旱能力强的转基因水角新种质。本研究的实施为水角的性状改良和基因功能验证提供基础,也为其他水生植物高效转化体系的建立提供技术借鉴。
-
于2023年4月份在海南省海口市羊山地区采集生长健康且健壮的水角的幼嫩枝条。
-
将幼嫩的枝条剪成长约7 cm的插穗,顶端切口平切或保留顶芽,下端切成45°斜口;保留插穗上部叶片2 ~ 4片。每片叶子剪去1/2 ~ 2/3。
-
双元表达载体pCambia1301由中国热带农业科学院热带生物技术研究所言普博士提供。pCambia1301含有由CaMV35S启动子驱动β-葡糖苷酸酶(GUS)基因。参考已报道的热击法[17]将pCambia1301分别转入MSU440、C58C1和K599 等3种发根农杆菌菌株,具体的操作方法如下:1) 取1 μg pCambia1301于100 μL发根农杆菌感受态细胞中,轻轻混匀;2) 菌液置于冰上先预处理5 min,再置于液氮处理5 min,37 ℃水浴处理5 min,冰上静置5 min,加入700 μL TY液体培养基28 ℃ 200 r·min−1振荡培养2 h;3) 将菌液涂布于含50 mg·L−1卡那霉素(Kana)和50 mg·L−1 链霉素(Strep)的TY固体培养基上,28 ℃培养2 d;4) 将单克隆菌落接种于含有相应抗生素的TY液体培养基中,28 ℃ 200 r·min−1振荡培养过夜,PCR检测阳性克隆,获得含GUS基因的发根农杆菌菌株,用于下面的水角遗传转化实验。
-
采用土培扦插转化法和水培扦插转化法2种转化方式对水角插穗进行转化处理。土培扦插转化法的操作步骤如下:1) 在水角插穗下端斜面处涂布上含有GUS目的基因的K599菌体,移植于湿润的无菌土壤基质:m(蛭石)∶m(泥炭土)∶m(椰糠)=1∶1∶1,再向移植好的插穗浇灌1 mL 相应的发根农杆菌发酵液;2) 保持环境相对湿度90%,控制温度28 ℃,遮光度70%,培养15 d后,观察转化效果。水培扦插转化法的操作步骤:1) 对水角插穗下端斜面处蘸上含有GUS目的基因的K599菌体;2) 在含有50 mg·L−1 Kana、50 mg·L−1 Strep TY液体培养基中加入1%(v/v)的活化的带有GUS基因载体的K599菌株,28 ℃振荡培养至OD600为0.8,
4000 r·min−1离心5 min,收集菌体;3) 将菌体稀释于清水中,直至OD600为0.1;4) 将插穗置于含有菌体稀释液的水培育苗箱中,确保约1/3插穗的下部浸于溶液中,培养温度为28 ℃,培养15 d后,观察转化效果。为了比较这2种转化方法的效果,对获得的水角植株的毛状根进行GUS组织化学染色,来确定阳性的转基因水角复合植株,并统计转化率。为了数据的准确性,统一用带顶芽的水角插穗作为转化材料。每个处理侵染10个插穗,每个处理重复3次。根据GUS检测结果来统计转化率:
$$\begin{split} \text{转化率}(\text{%})=&(\text{阳性转基因植株数量/}\\&\text{检测的植株总数})\times 100 。 \end{split} $$ (1) 比较这2种转化方法的转化效果。作Student t-test检验,P< 0.05为显著差异。
-
用上面制备好的带GUS目的基因载体的MSU440、C58C1和K599 等3种发根农杆菌菌液,采用水培扦插转化法分别转化带顶芽的水角插穗。每个处理侵染10个插穗,每个处理重复3次。根据GUS检测结果来统计各处理转化率,比较各菌株的转化效果。作ANOVA方差分析, P< 0.05为显著差异。
-
用前面制备好的带GUS目的基因载体的K599菌液,采用水培扦插转化法分别处理带顶芽的和没顶芽的水角插穗。每个处理侵染10个插穗,每个处理重复3次。根据GUS检测结果来统计2种类型插穗的转化率,比较它们的转化效果。作Student t-test检验,P< 0.05为显著差异。
-
GUS组织化学染色应用商业化的GUS试剂盒(北京,中科瑞泰),实验操作按试剂盒说明书进行。剪取水角复合植株的毛状根,用dH2O清洗干净,滤纸吸干根上的水珠后置于含GUS染色液的离心管中,离心管用锡箔纸包扎好后于50 r·min−1振动培养30 min,将材料转至无水乙醇中脱色,体视显微镜下观察染色情况。
-
采用高效植物基因组DNA提取试剂盒(北京,天根)提取样品DNA。剪取100 mg复合植株的根部,用清水清洗3遍,无菌滤纸吸干水珠后置于研砵中,加入液氮进行研磨,再根据试剂盒说明书进行DNA提取。设计GUS基因引物,GUS-F:5'-CTGGGTGGACGATATCACCG-3',GUS-R: 5'-GCGAAATATTCCCGTGCACC-3'。1%(m/v)凝胶电泳检测PCR扩增产物。
-
步骤1)海藻糖-6-磷酸合成酶基因MeTPS6的克隆。取盆栽2个月的木薯品种Ku50的根部,采用RNAprep Pure植物总RNA提取试剂盒(北京,天根)提取木薯根的总RNA,再用TransScript一步法gDNA去除及cDNA合成试剂盒(北京,全式金)反转录合成一链cDNA。从GenBank中查找到木薯的海藻糖-6-磷酸合成酶基因MeTPS6序列(登录号:MF684374.1),设计MeTPS6引物(MeTPS6-1-F: 5'-ATGGTGTCAAGGTCATACTC-3', MeTPS6-1-R: 5'-CACTGCAACTGTTTGTTCTG-3'),以木薯根cDNA为模板DNA扩增MeTPS6。采用pMD19-T载体(北京,宝生物)对PCR扩增产物进行T载体克隆,将克隆好的T载体送至华大基因测序,确保MeTPS6序列正确。
步骤2)构建海藻糖-6-磷酸合成酶MeTPS6基因过表达载体。参考所报道的Nimble Cloning(NC分子克隆)技术[18]构建海藻糖-6-磷酸合成酶基因MeTPS6过表达载体。以上面构建好的含有MeTPS6基因的T载体质粒为模板,对MeTPS6基因进行扩增,所用的引物为:MeTPS6-2-F: 5'- agtggtctctgtccagtcctATGGTGTCAAGGTCATACTC-3',MeTPS6-2-R: 5'-ggtctcagcagaccacaagtCACTGCAACTGTTTGTTCTG-3'(小写字母为NC框接头序列)。对PCR扩增产物进行1%(m/v)凝胶电泳,用凝胶回收试剂盒(南京,诺唯赞)纯化回收目的条带。采用NC克隆试剂盒(海南,壹田)将纯化回收后的PCR产物与带有NC克隆框的载体pNC-Green-SubC(由中国热带农业科学院热带生物技术研究所言普博士馈赠)进行NC克隆反应。将反应产物通过热击法导入感受态大肠杆菌E.coli TOP10菌株中,菌液涂布在含有50 mg·L−1 Kana的LB固体培养基上,转化平板于37 ℃过夜培养后,挑选阳性克隆于含50 mg·L−1 Kana的液体LB培养上37 ℃振荡培养16 h,用菌液作模板扩增MeTPS6基因,PCR扩增产物送至华大基因测序,确定序列正确的阳性克隆为构建成功的新载体,命名为pGreen-35s:MeTPS6-EGFP。
-
参照前面所建立的优化的转基因水角复合植株技术体系进行MeTPS6基因过表达载体转化水角研究,具体操作如下:1) 采用热击法将载体pGreen-35s:MeTPS6-EGFP导入感受态发根农杆菌K599中。2) 对带有顶芽的水角插穗下端斜面处涂上携带pGreen-35s:MeTPS6-EGFP的发根农杆菌K599菌株。3) 在含有50 mg·L−1 Kana、50 mg·L−1 Strep TY液体培养基中加入1%(v/v)的活化的携带目的基因的K599的菌液,28 ℃振荡培养至OD600值为0.8,离心收集菌体。4) 用清水稀释菌体至OD600值为0.1,将下端斜面处已涂抹发根农杆菌的插穗置于含有菌体稀释液的水培育苗箱中,28 ℃培养40 d。
-
将手持荧光电筒(LUYOR-3415RG,美国)的发射光波调至绿色荧光波长500 nm,在黑暗条件下照射pGreen-35s:MeTPS6-EGFP转化的水角植株,检测阳性转基因植株。用高效植物基因组DNA提取试剂盒分别提取阳性转基因植株的根部基因组DNA,设计EGFP基因引物(EGFP-F:5'-AGGACGACGGCAACTACAAG-3', EGFP-R:5'-TCTCGTTGGGGTCTTTGCTC-3'),以根部基因组DNA为模板进行PCR扩增,1%(m/v)凝胶电泳检测PCR产物。
-
配制好3%(m/v)的polyethylene glycol-6000(PEG-6000)水溶液,将过表达MeTPS6基因的转基因和非转基因水角植株的根部浸泡于装有3%(m/v)PEG-6000的水溶液玻璃瓶中,28 ℃自然光照条件培养,观察它们的耐旱性。
-
参考Han等[16]报道的方法测定转基因水角复合植株海藻糖含量。剪取转基因植株的根1 g,用液氮研磨至粉末,加入15 mL 80%(v/v)乙醇,在80 ℃水浴中处理30 min,
6000 r·min−1离心10 min,取上清。再向沉淀物中加入15 mL 80%(v/v)乙醇,重复抽提2次。将重复抽提的上清与前面的上清合并,80 ℃烘干乙醇,加无离子水定容至5 mL,用0.22 um滤膜过滤滤液。准备好0.1、0.2、0.5、1.0、2.0、4.0 mg·L−1的海藻糖标准液。将样品进样至高效液相色谱仪(Waters e2695)中,每次进样10 uL,采用的流动相为乙腈∶水(含0.1%(m/v)的NH4OH)(70∶30),流速1.0 mL·min−1,柱温为25 ℃,xbridge-NH2为色谱柱。检测器的条件是氮气流速2 L·min−1,漂移管温度85 ℃。 -
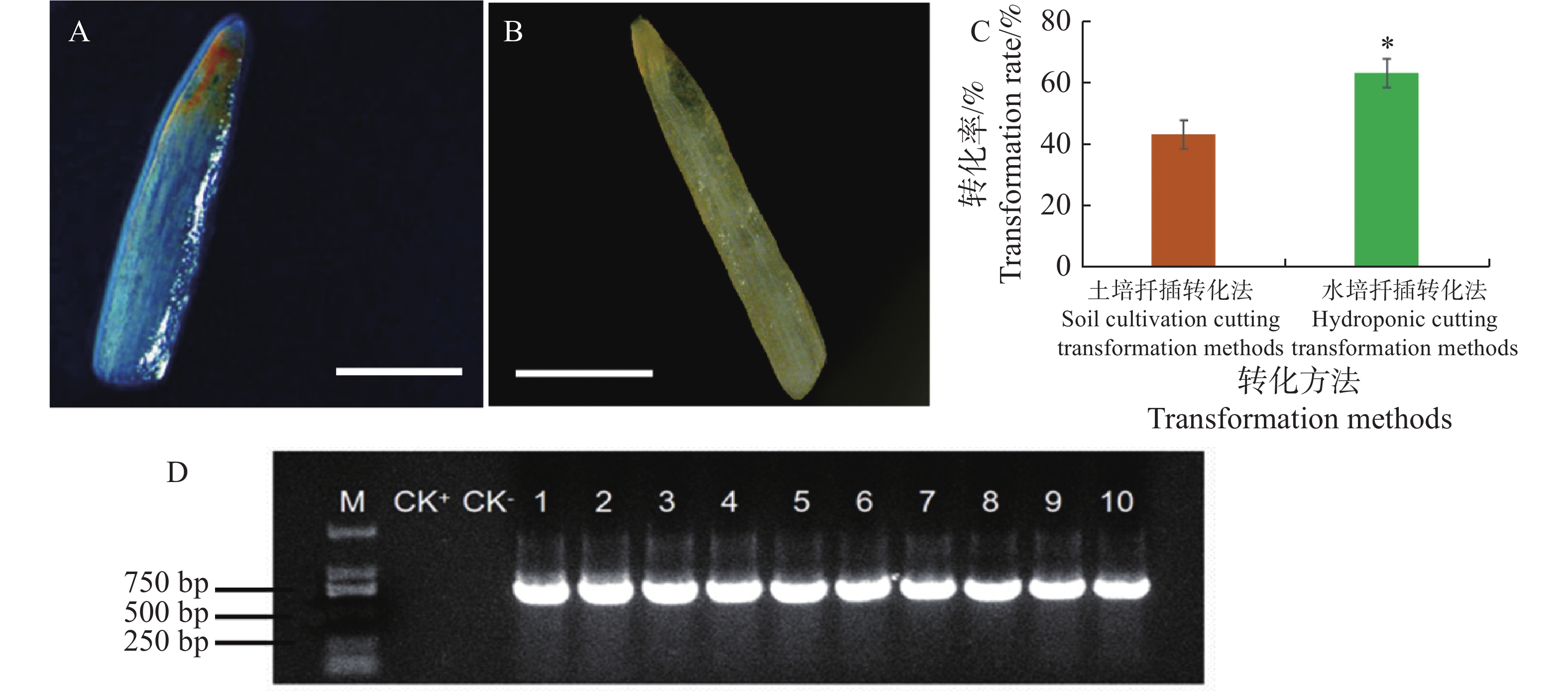
采用土培扦插转化法和水培扦插转化法对水角插穗进行转化处理,结果发现这2种方法均能获得具有毛状根的植株(图1)。在土培扦插转化法中,在插穗斜面处涂布K599菌体,将插穗移植于湿润的含蛭石、泥炭土、椰糠的基质中,再浇上K599菌液,培养15 d后插穗基部长出毛状根(图1−A)。在水培扦插转化法中,插穗斜面处预先蘸上K599菌体,再插入K599菌液稀释液,培养插15 d后,插穗基部也长出毛状根(图1−B)。
图 1 发根农杆菌介导的水角转基因体系的建立
Figure 1. Establishment of Agrobacterium rhizogenes-mediated transformation system for Hydrocera triflora
GUS组织化学染色结果显示,阳性转基因植株的毛状根显示蓝色(图2−A),而阴性非转基因植株的毛状根颜色没有任何变化(图2−B)。对这2种转化方法的转化率进行分析,结果显示土培扦插转化法的转化率为43.3%,而水培扦插转化法的转化率为63.3%,后者显著高于前者(图2−C)。随机选取GUS检测阳性的植株10株,提取其根部基因组DNA,PCR检测,结果发现均能扩增出GUS基因片段的目的条带,目的条带大小为732 bp(图2−D),因此再一步证实GUS检测阳性植株为转基因水角复合植株。上述结果表明水培扦插转化法优于土培扦插转化法。
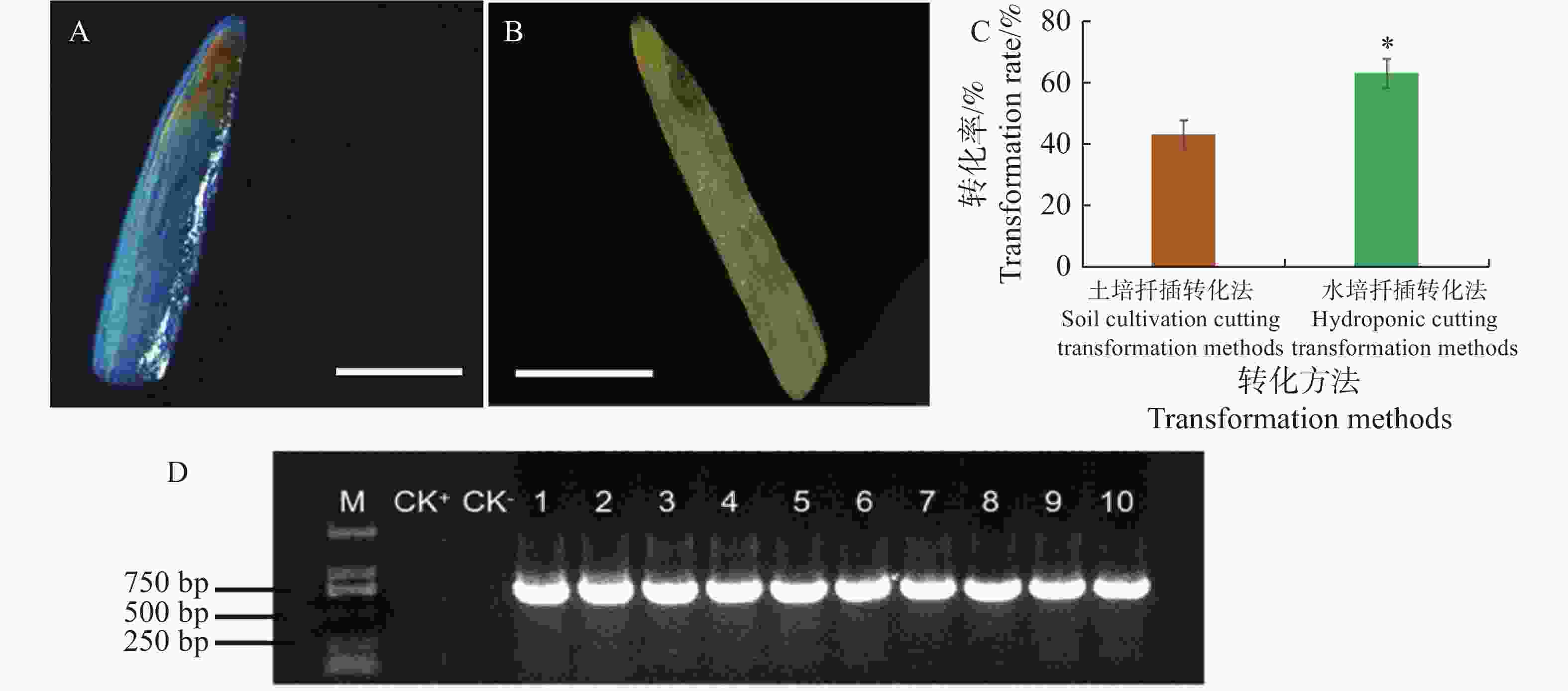
图 2 图2 土培扦插转化法与水培扦插转化法的转化率比较
Figure 2. Comparison of transformation rates between soil cultivation and hydroponic cutting transformation methods
-
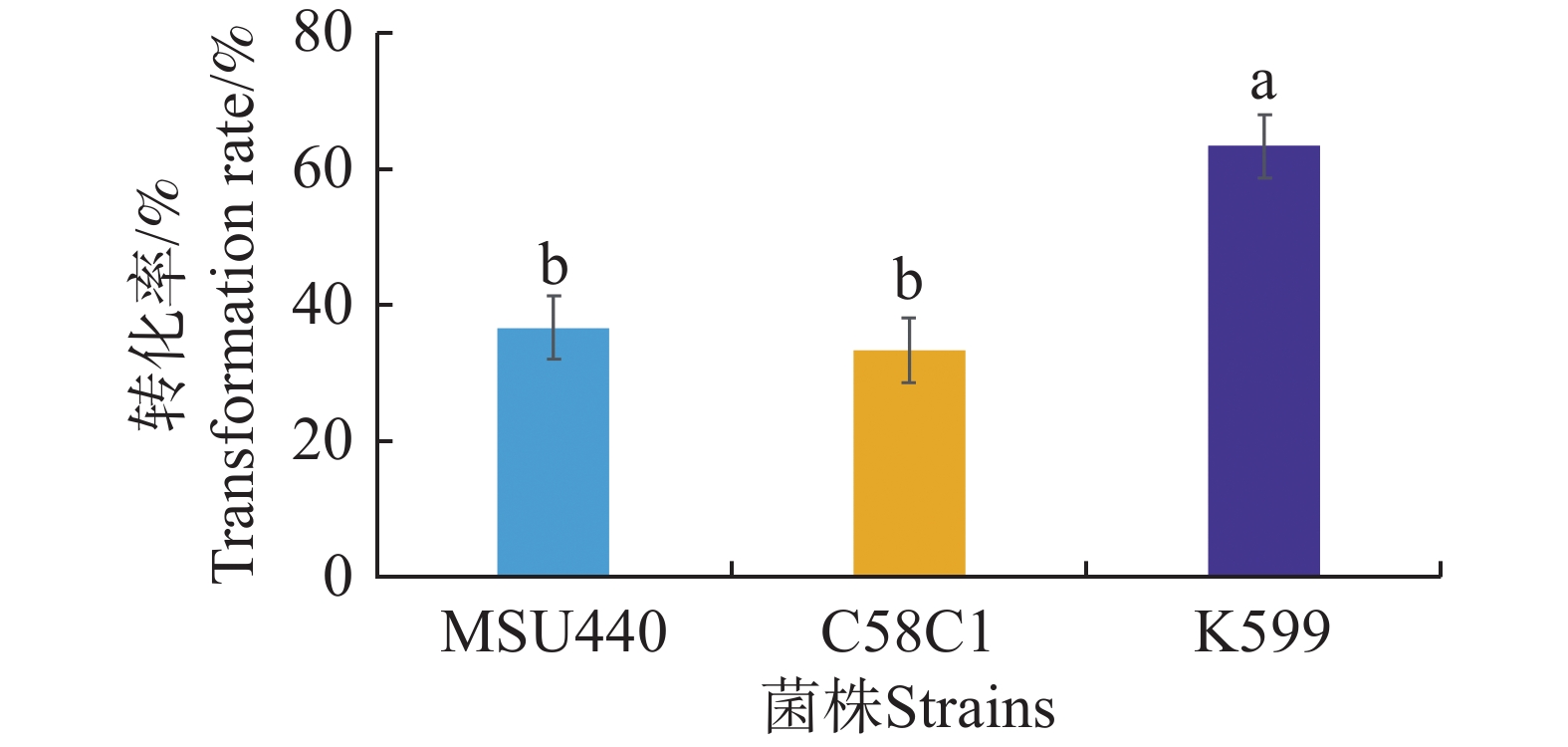
为了进一步优化发根农杆菌介导的水角转基因技术体系,采用水培扦插转化法实验MSU440、C58C1、K599等3种发根农杆菌对带顶芽的水角插穗的转化效果。这3种菌株均携带载体pCambia1301。对各处理植株的毛状根进行GUS组织染色检测,根据检测结果统计转化率。结果表明,菌株MSU440和C58C1转化率最低,分别为36.7%和33.3%;菌株K599的转化率显著高于另两个菌株,达到63.3%(图3)。因此,在发根农杆菌介导的水角转基因技术体系中选用菌株K599转化效果最佳。
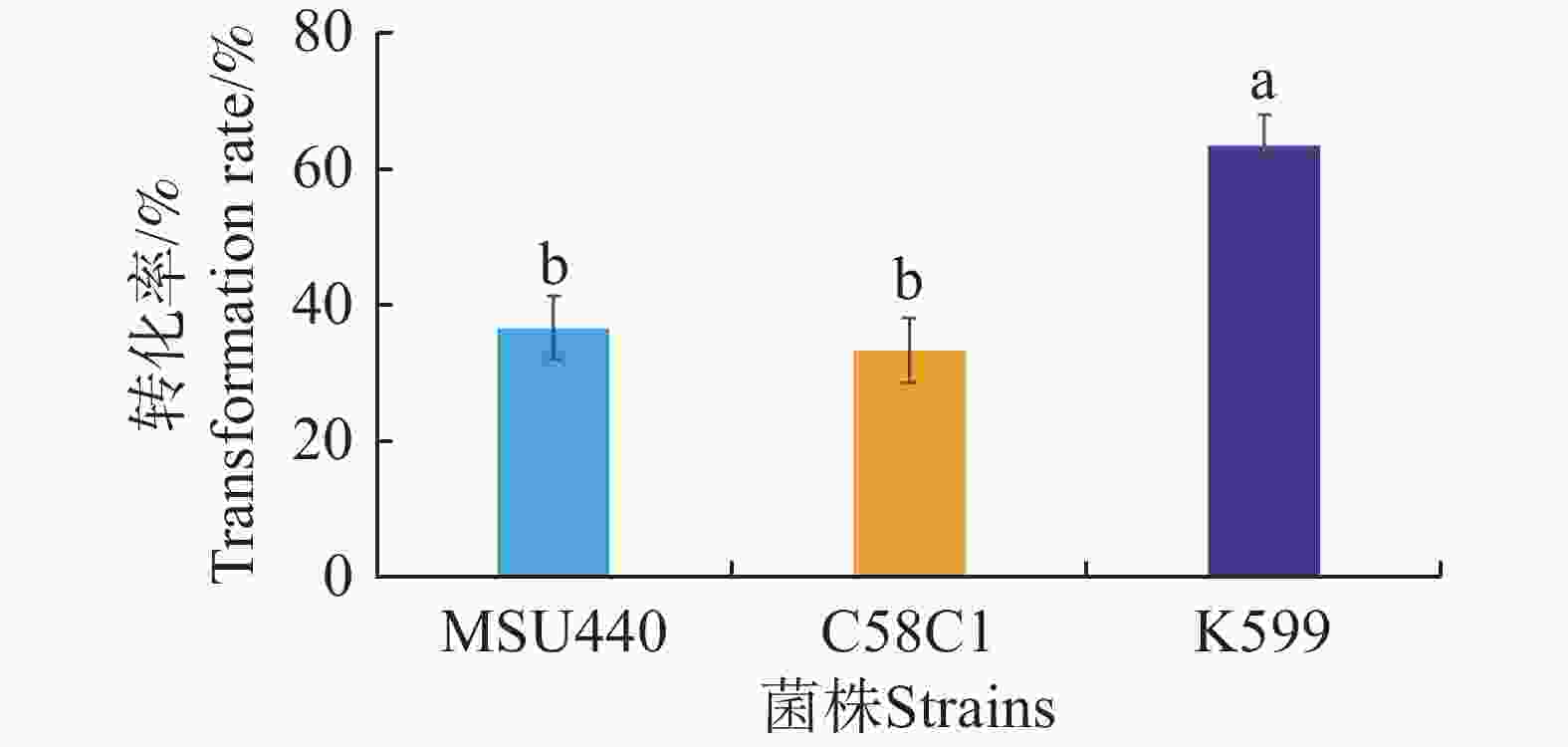
图 3 不同发根农杆菌株对水角转化效率的影响
Figure 3. The effect of different Agrobacterium rhizogenes strains on Hydrocera triflora transformation efficiency
-
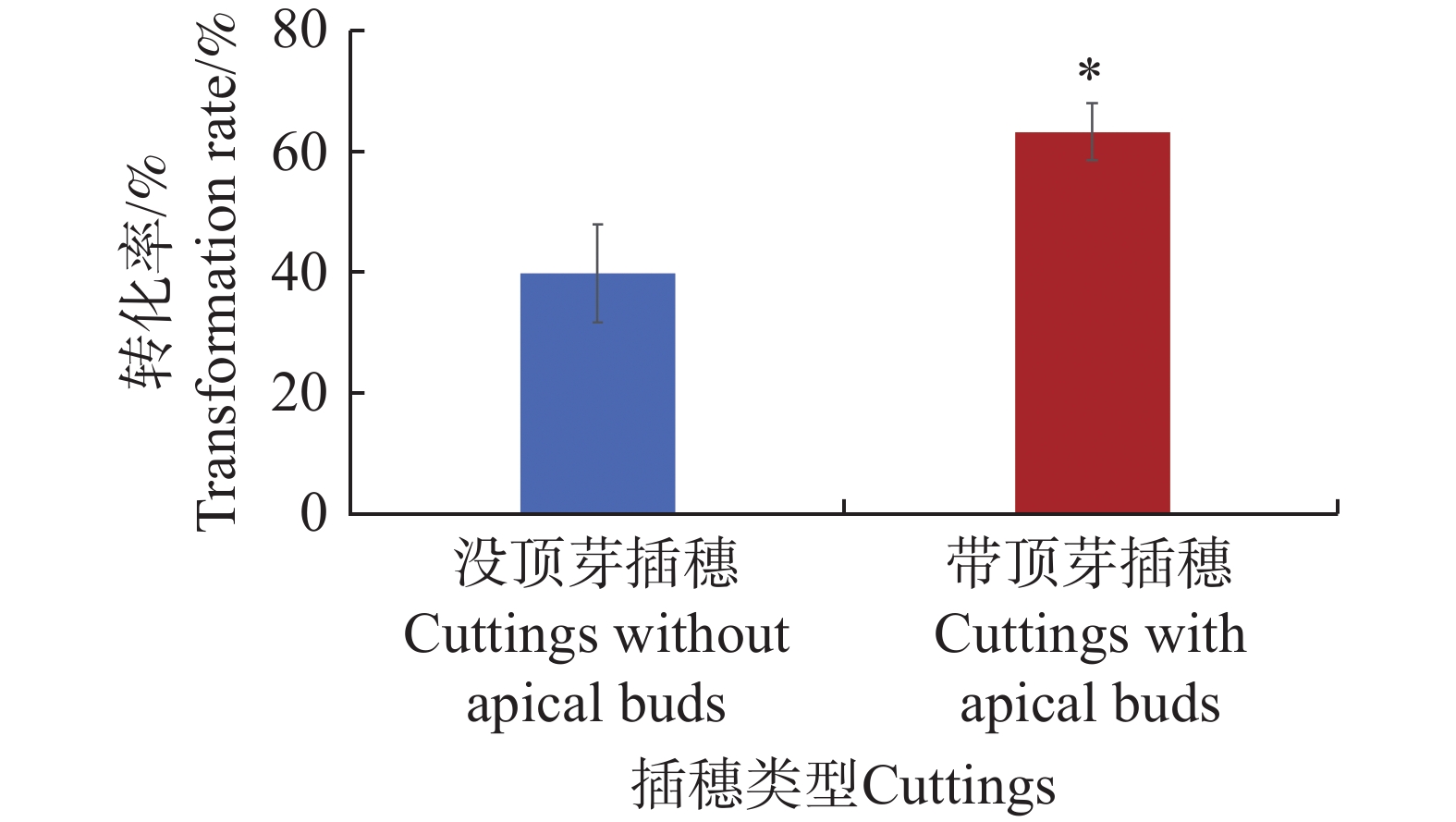
水角的插穗存在2种类型,一种是带顶芽的,另一种是没顶芽的。为了验证这2种类型的插穗是否对水角转化率存在影响,从而更进一步优化发根农杆菌介导的水角转基因技术体系,采用水培扦插转化法,并选用携带了表达载体pCambia1301的发根农杆菌K599菌株来进行该研究。结果表明,没顶芽的插穗转化率为40.0%;而带顶芽的插穗转化率高达63.3%,显著高于没顶芽的插穗(图4)。这些结果表明,带顶芽的插穗的转化率优于没顶芽的插穗。
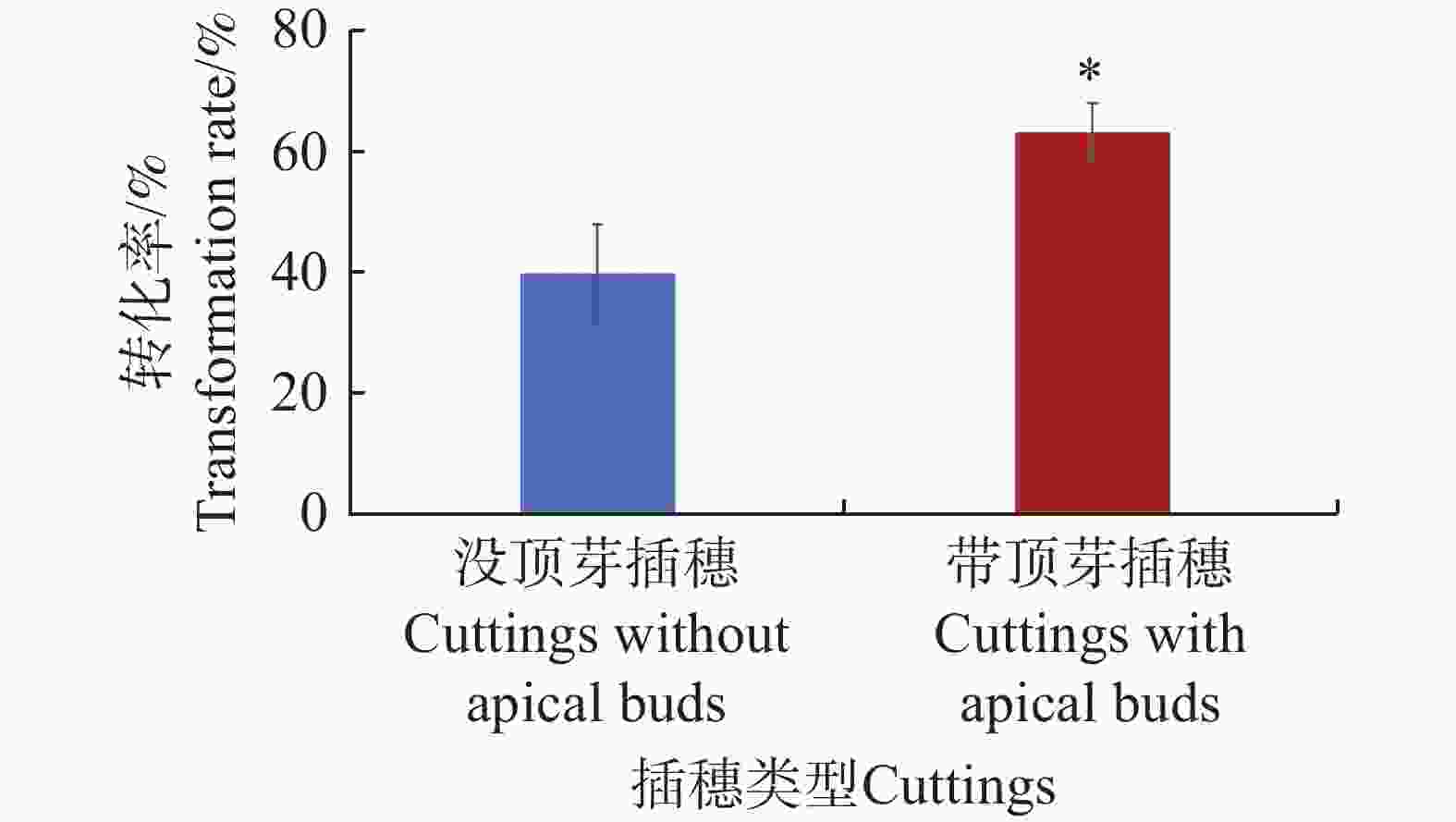
图 4 不同插穗类型对水角转化效率的影响
Figure 4. The effect of different types of cuttings on Hydrocera triflora transformation efficiency
-
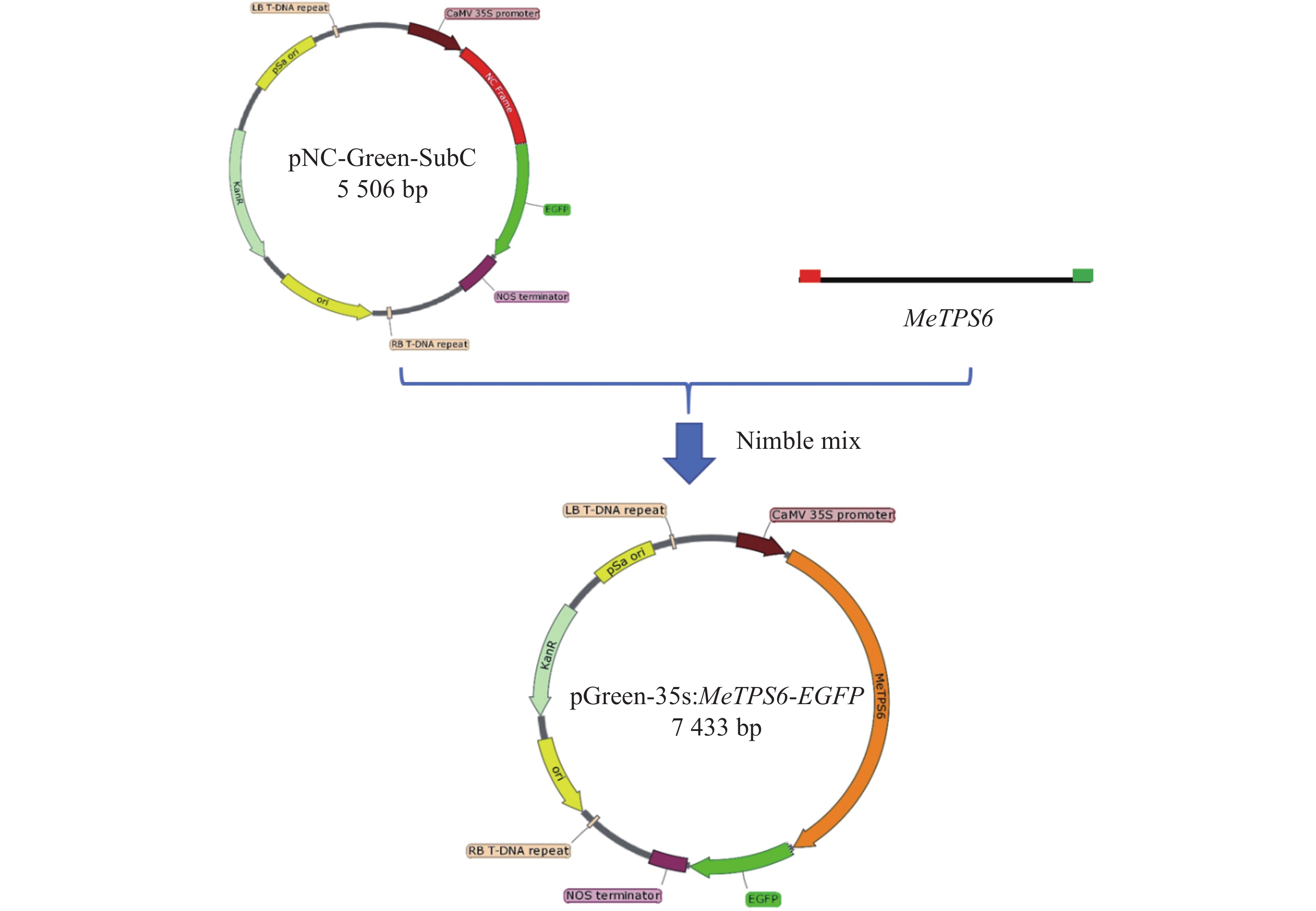
NC克隆技术是一种简单高效的分子克隆技术[18]。基于该技术逐渐被广泛应用,NC克隆技术的开发者随后开发一套基于Nimble Cloning的植物表达载体,其中包括pNC-Green-SubC[19] 。植物表达载体pNC-Green-SubC中的CaMV 35s启动子连接NC克隆框和标记基因EGFP。从木薯根部扩增到不带终止密码子的MeTPS6基因序列,长度共计
2562 bp,T载体克隆该基因序列,以此T载体质粒为模板,用带NC克隆框接头的引物MeTPS6-2扩增MeTPS6基因序列,获得两端带NC克隆框接头的MeTPS6基因序列。将MeTPS6基因序列与pNC-Green-SubC载体进行NC克隆反应,获得新的载体pGreen-35s:MeTPS6-EGFP(图5)。在新载体中,CaMV 35s启动子驱动MeTPS6和EGFP进行融合表达。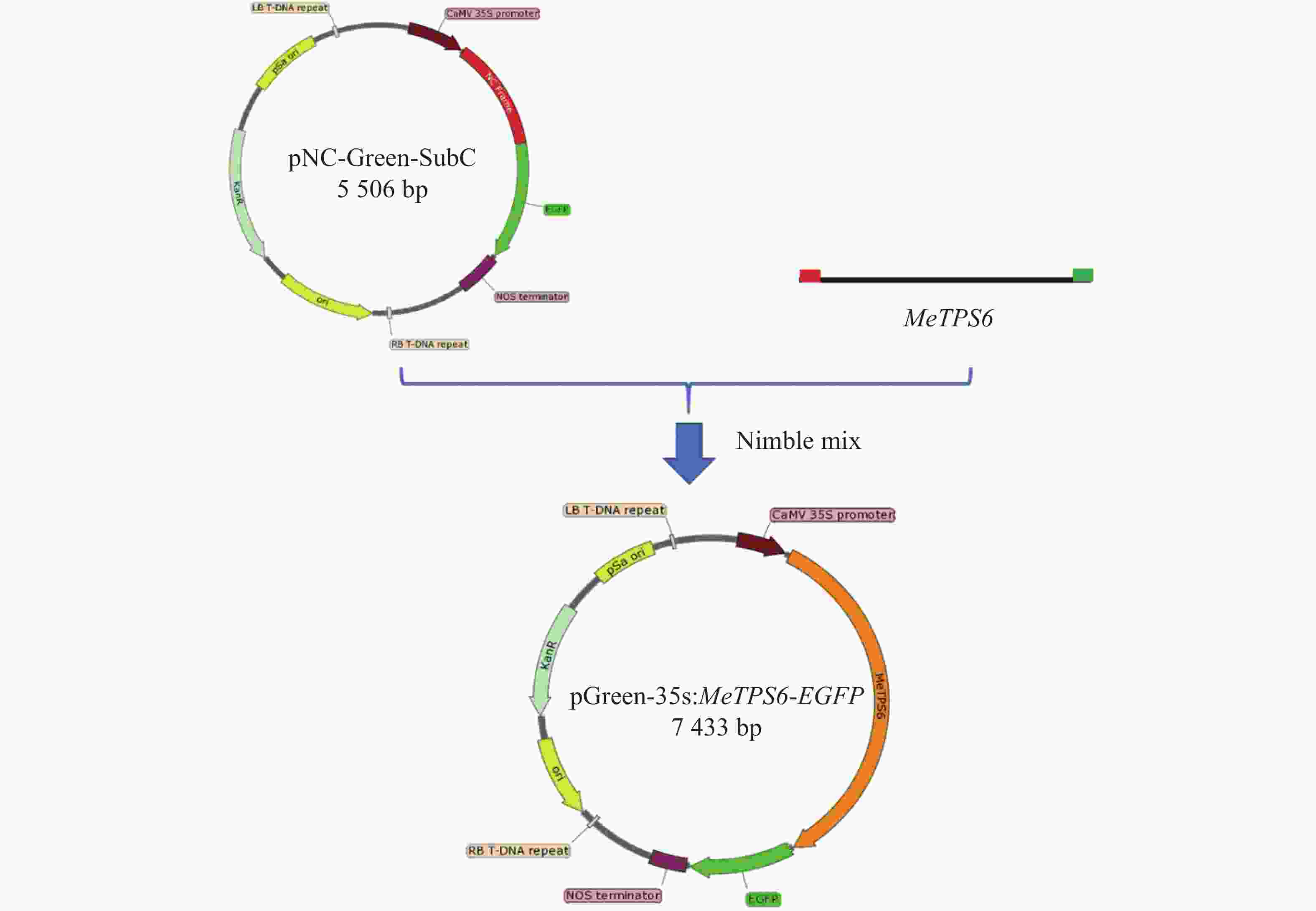
图 5 表达载体pGreen-35s:MeTPS6-EGFP的构建
Figure 5. Construction of pGreen-35s:MeTPS6-EGFP
-
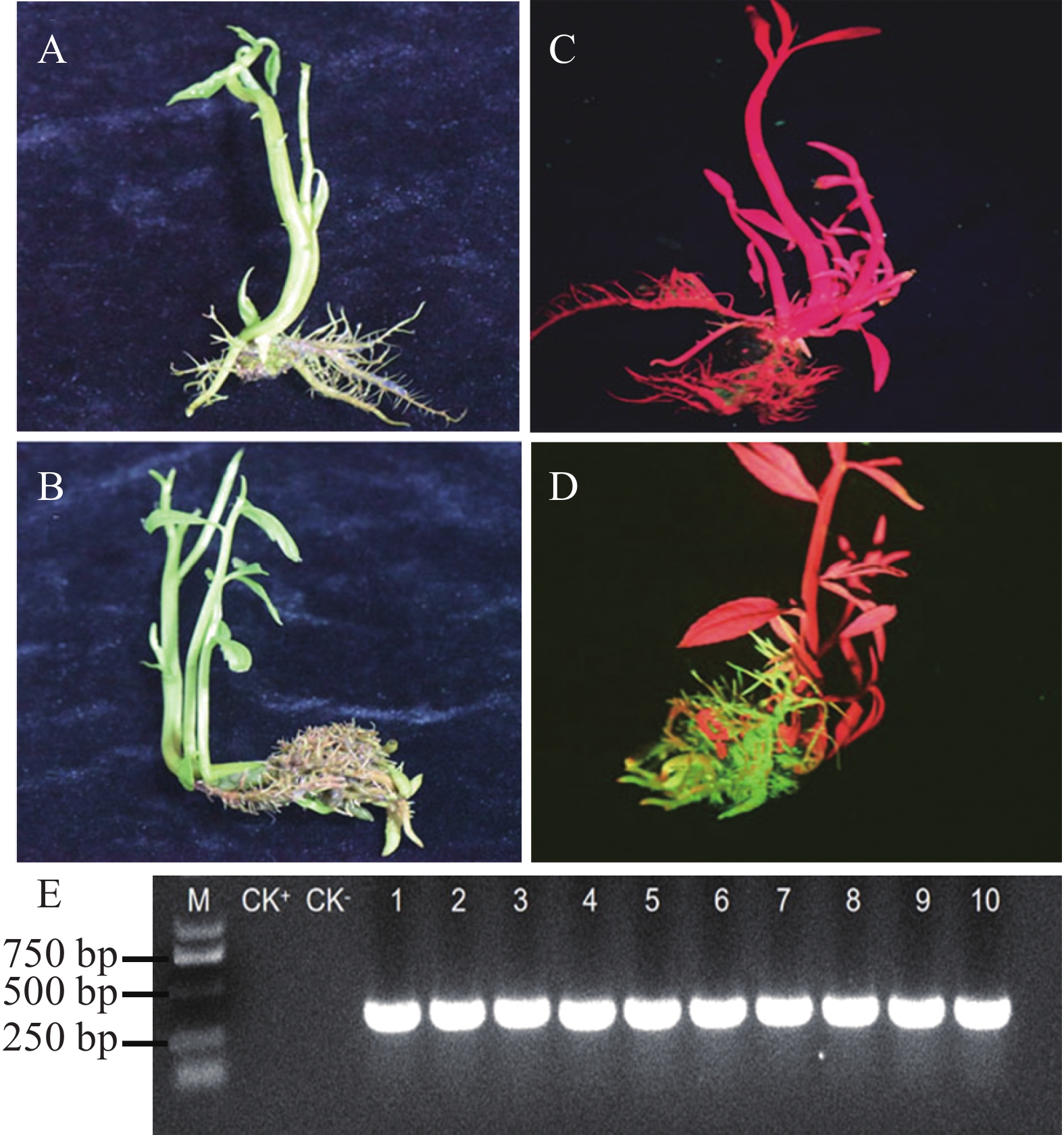
本研究构建了MeTPS6基因过表达载体pGreen-35s:MeTPS6-EGFP,应用所建立的发根农杆菌介导的转基因水角复合植株技术体系转化至水角植株中,获得了转基因水角复合植株。非转基因与转基因的水角植株在形态上没有表现差异(图6−A、B)。对转基因植株进行绿色荧光检测,结果发现在发射光波长500 nm照射下,非转基因植株的根、茎、叶均显示红色(图6−C);阳性转基因复合植株的毛状根显示绿色,而其茎和叶均显示红色(图6−D)。随机选取10株经荧光检测为阳性转基因的复合植株,分别提取它们的毛状根DNA,经PCR检测,它们均能扩增出EGFP基因片段(图6−E),进一步证实它们是转基因复合植株。
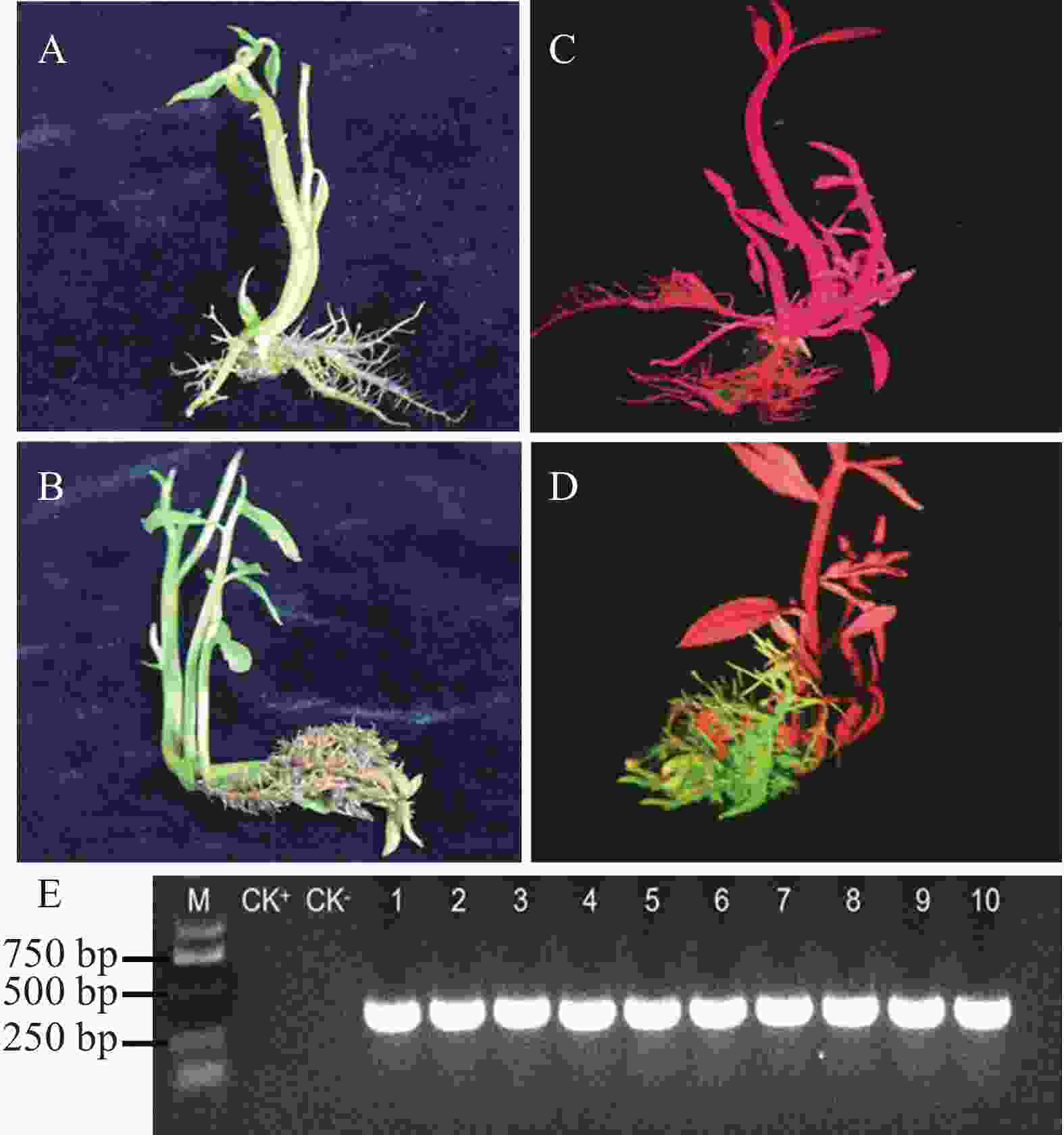
图 6 发根农杆菌介导表达载体pGreen-35s:MeTPS6-EGFP转化水角
Figure 6. pGreen-35s:MeTPS6-EGFP transformed Hydrocera triflora using Agrobacterium rhizogenes mediated method
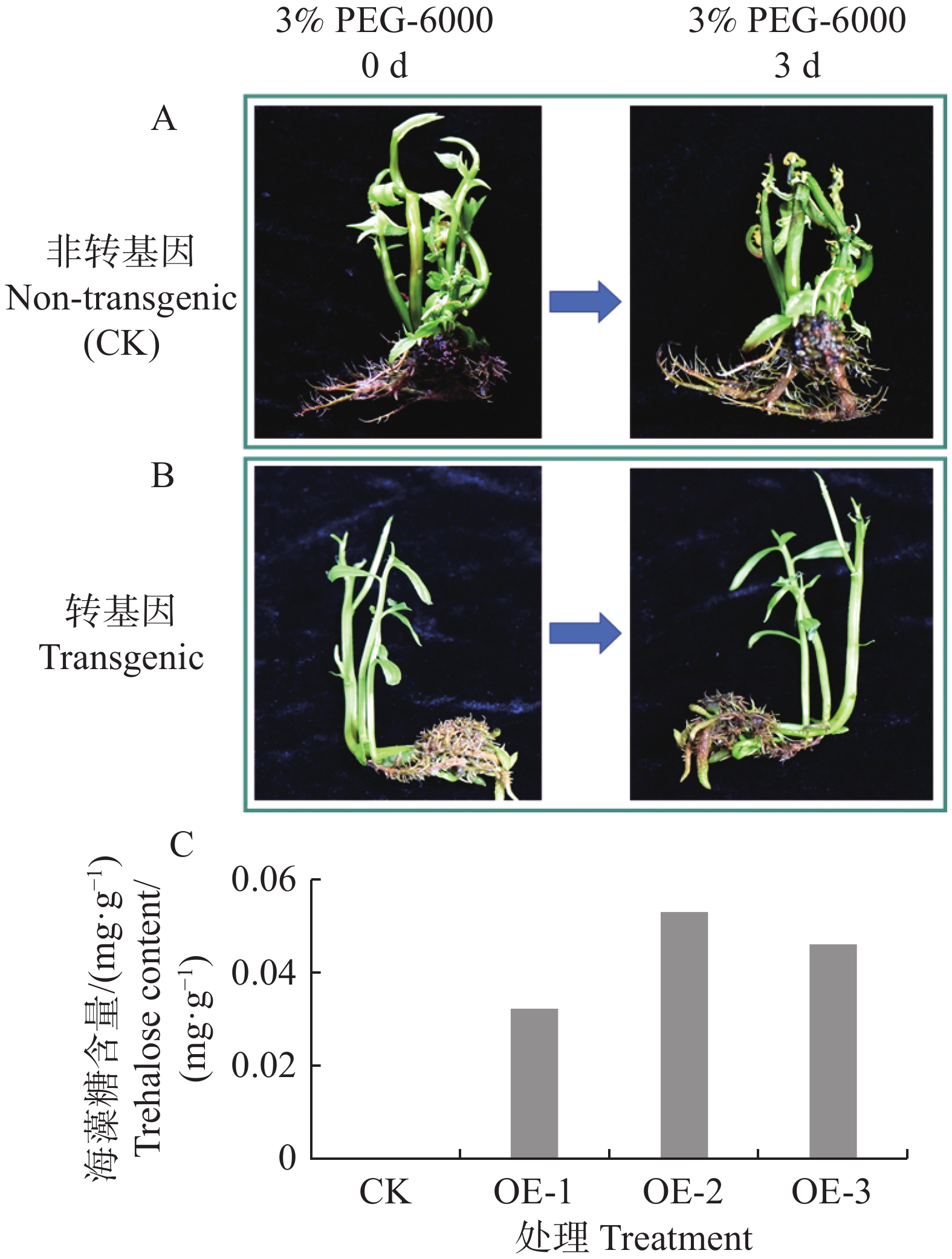
将转基因水角复合植株和非转基因水角植株的根部浸泡于3%(m/v)PEG-6000溶液中,培养3 d后,非转基因水角植株叶片出现萎蔫(图7−A),转基因水角复合植株生长正常(图7−B)。选取3株转基因植株检测其毛状根的海藻糖含量,发现它们的海藻糖含量在0.032~0.053 mg·g−1 (图7−C);而对照的根部由于海藻糖含量太低,超出仪器检测的灵敏度范围,因此没有检测到海藻糖含量。这些结果表明转基因水角复合植株的耐旱能力得到显著改善。
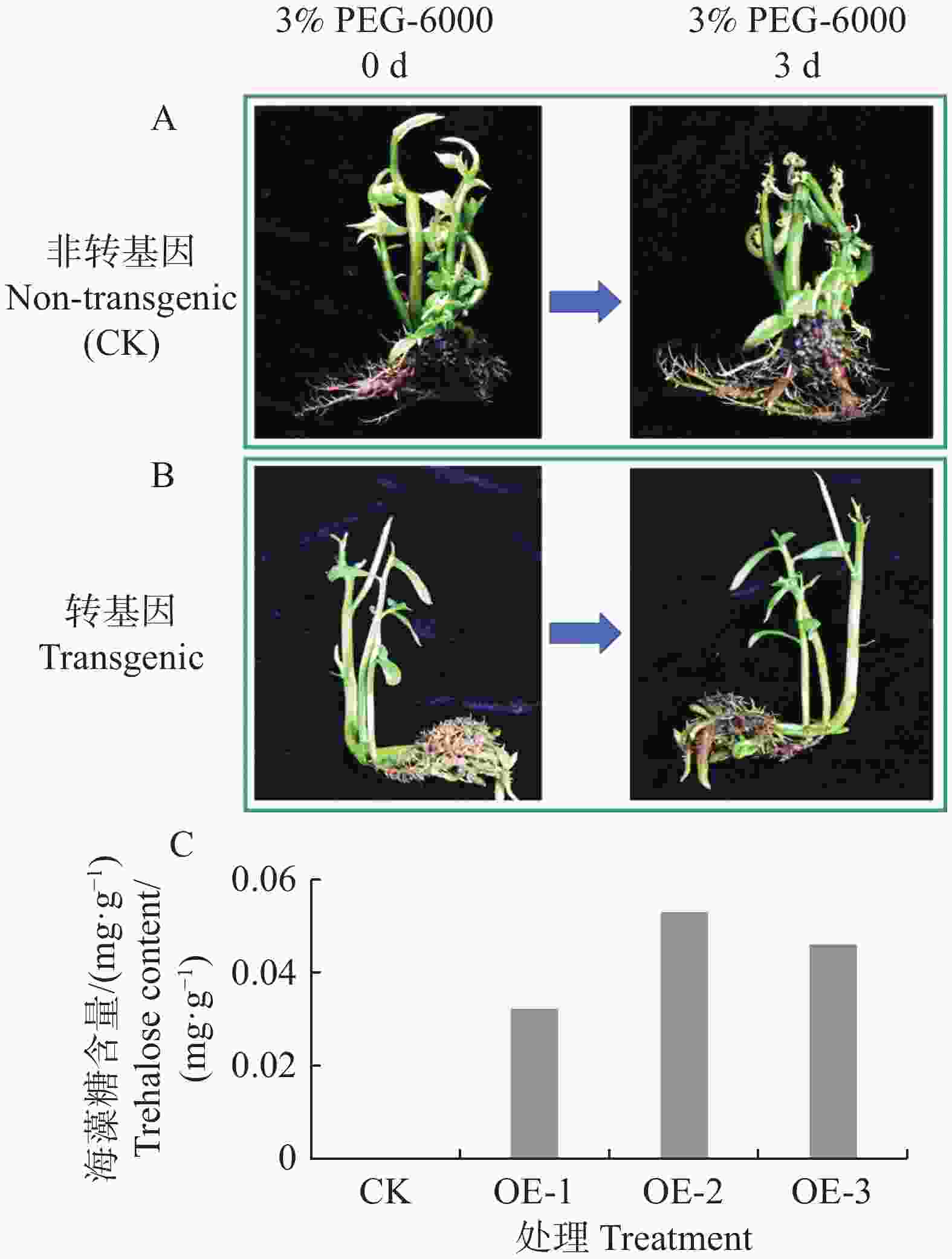
图 7 转基因水角复合植株耐旱性检测
Figure 7. Drought tolerance testing of transgenic Hydrocera triflora
-
有关水角组织培养的报道极少。王景飞等[20]以水角的种子和茎段为外植体,通过优化培养基组分来诱导这些外植体上的芽点形成不定芽和丛生芽,获得水角再生植株。这种不经过愈伤组织获得再生植株的组织培养方法不能应用于根瘤农杆菌介导的遗传转化技术上。目前发展出不依赖组织培养的发根农杆菌介导的遗传转化方法,这些研究均是通过携带目的基因的发根农杆菌侵染种子下胚轴,获得由转基因毛状根和野生型茎叶组成的转基因复合植株,这些转基因复合植株可用来改良性状和研究相关基因的功能[6 − 9]。这种遗传转化方法具有操作简单、转化率高、不依赖组织培养和基因型等优势。为此本研究借鉴该技术来尝试建立水角的高效、便捷、不依赖组织培养的转基因技术体系。由于水角种子萌发率极低,本研究选用水角插穗来代替种子幼苗作为农杆菌侵染外植体,并成功建立了基于发根农杆菌介导的水角转基因技术体系。本研究的结果为其他水生植物以及极难获得种子或种子萌发率低的植物的性状改良和基因功能验证提供了技术参考。
不同的发根农杆菌菌株对植物转化率存在一定的影响。在柱花草遗传转化研究中,菌株K599和Arqual的转化率达95%,均显著高于MSU440菌株,而ArA4和Ar1193菌株不能诱导产生转基因毛状根[9]。本研究也得到相似的结果,即菌株K599的转化率达到63.3%,显著高于菌株SU440和C58C1。在鹰嘴豆[6]、大豆[21]等植物的发根农杆菌介导转化研究中,菌株K599被证明是最合适的转化菌株。因此菌株K599可能适合多种植物的转化。
发根农杆菌能够在无激素促进、无抗性筛选的条件下诱导植物被侵染部位产生毛状根[22],因此被广泛应用于基因功能验证、次生代谢产物等方面的研究[6,9,23]。发根农杆菌诱导被侵染部位产生的毛状根其阳性率在不同的物种中存在差异。Wang等[9]发现柱花草阳性复合转基因植株的毛状根阳性率达到100%。而Fan等[8]发现大豆阳性复合转基因植株的毛状根阳性率与转化方法相关,其中二步转化法最高,为53.85%。在本研究中,水角阳性复合转基因植株的毛状根阳性率达到100%,与柱花草的相似。
海藻糖-6-磷酸合成酶是海藻糖生物合成代谢的关键酶,在植物干旱等非生物胁迫中起到重要作用[11,24]。MeTPS6基因主要在木薯的根中表达,且参与了木薯的抗旱等胁迫响应[25]。本研究构建了MeTPS6基因过表达载体,并转化至水角的毛状根中,获得转基因复合植株。目前所报道的植物耐旱实验基本上采用PEG-6000溶液处理来作为模拟干旱胁迫处理 [16,26 − 27]。本研究也采用PEG-6000溶液处理过表达MeTPS6基因的转基因复合植株,培养3 d后发现转基因复合植株生长正常,而对照出现萎蔫,说明转基因复合植株的耐干旱胁迫能力得到明显提升。
本研究采用NC分子克隆[18]这一新的克隆技术来构建植物表达载体,与当前广泛应用的Gibson assembly[28]和Gateway cloning[29]分子克隆技术相比,具有如下优势:1) Gibson assembly需要将目标载体质粒酶解线性化后,目的基因才能克隆至目标载体上,而NC不需要将目标载体质粒酶解线性化,目的基因片段可直接克隆至环状的质粒上;2) Gibson assembly的插入片段限制于PCR产物,Gateway cloning需要入门克隆,而NC比较灵活,可以直接使用PCR产物或者入门克隆;3) Gateway cloning在重组位点上会留下一个25 bp的缝,而NC能进行完全无缝克隆;4) 多片段组装时,Gateway cloning的克隆效果极差,而NC的克隆效果不受影响。此外,研究者还开发出一套基于Nimble Cloning的植物表达载体[19],可用于基因异位表达、基因沉默、亚细胞定位、蛋白互作、启动子活、CRISPR/Cas9基因编辑等研究。
-
首次建立了高效、不依赖组织培养的水角遗传转化体系。对该转化体系进行优化研究,以水角插穗为发根农杆菌侵染外植体,发现水培扦插转化法的转化率显著高于土培扦插转化法;发根农杆菌菌株K599转化率最高;选用带顶芽插穗作为侵染外植体,其转化效果优于无顶芽插穗。经优化的水角转化体系其转化率高达63.3%。此外,构建了过表达海藻糖-6-磷酸合成酶MeTPS6基因的载体pGreen-35s:MeTPS6-EGFP,通过转化获得了过表达MeTPS6转基因水角复合植株。3%(m/v)PEG-6000溶液处理3d后,发现转基因植株生长正常,而对照出现萎蔫现象;转基因植株毛状根的海藻糖含量为0.032~0.053 mg·g−1 ,显著高于对照,表明过表达MeTPS6转基因水角复合植株的耐旱能力得到显著改善。
Agrobacterium rhizogenes-mediated transformation system for Hydrocera triflora
-
摘要: 水角(Hydrocera triflora)是重要的濒危水生植物,耐旱性差是导致其濒危的重要因素。本研究拟建立水角遗传转化体系,并对其进行遗传改良,以期减轻其濒危程度。首次建立并优化了发根农杆菌介导的水角遗传转化体系。在优化该转化体系研究中,发现水培扦插转化法较优,菌株K599转化率最高,带顶芽插穗作侵染外植体转化效果较优。优化的转化体系转化率高达63.3%。构建了过表达海藻糖合成酶MeTPS6基因的载体并获得其转基因植株。3%(m/m) polyethylene glycol-6000(PEG-6000)溶液处理3 d,发现转基因植株生长正常,而对照叶片出现萎蔫。经检测转基因植株毛状根的海藻糖含量显著高于对照。这些结果表明过表达MeTPS6转基因水角的耐旱能力得到显著改善。本研究结果为水角的性状改良和基因功能验证提供基础,也为其他水生植物高效转化体系的建立提供技术借鉴。Abstract: Hydrocera triflora is an important aquatic plant that is in an endangered state. Its poor drought resistance is the main factor leading to its endangerment. An attempt was made to build a genetic transformation system for and to improve its traits via this transformation system. H. triflora Agrobacterium rhizogenes-mediated transformation system was successfully established and optimized for the first time. The results show that the transformation rate of hydroponic cutting transformation method was significantly higher than that of soil cultivation cutting transformation method. Agrobacterium rhizogenes K599 strain had the highest transformation rate. The cuttings with apical buds as infection explants had a higher transformation rate than those without apical buds. The transformation rate of the optimized H. triflora transformation system reached 63.3%. Meanwhile, pGreen-35s:MeTPS6-EGFP that overexpresses trehalose-6-phosphate synthase MeTPS6 was constructed by Nimble Cloning technology. The transgenic composite H. triflora were obtained. After 3 days of treatment with 3% PEG-6000 solution, it was found that the transgenic plants grew normally, while the control leaves showed wilting. The trehalose content in the hairy roots of transgenic plants was significantly higher than that of the control. These results indicate that overexpressing MeTPS6 in transgenic composite H. triflora significantly improves their drought stress tolerance.
-
图 2 图2 土培扦插转化法与水培扦插转化法的转化率比较
A. 阳性转基因植株毛状根GUS染色;B. 非转基因植株毛状根GUS染色;C. 土培扦插转化法与水培扦插转化法的转化率比较;D:阳性毛状根PCR检测。“*”表示显著差异(P<0.05)。CK+,阳性对照,以空载体的K599为模板的PCR扩增产物;CK-,阴性对照,以非转基因水角植株根部DNA为模板的PCR扩增产物;扩增的目的基因为GUS基因,目的条带为732 bp。标尺=1 mm。
Fig. 2 Comparison of transformation rates between soil cultivation and hydroponic cutting transformation methods
A. Positive transgenic H. triflora hairy root GUS staining; B. Non-transgenic H. triflora hairy root GUS staining; C. Comparison of transformation rates between soil cultivation and hydroponic cutting transformation methods; D. Positive hairy root PCR detection.“*” indicates significant differences(P<0.05); CK+: Positive control, PCR amplification using empty vector K599 as template; CK−: Negative control, PCR amplification using non-transgenic plant root DNA as template. The target gene is GUS gene, with a target band of 732 bp. Scales: 1 mm.
图 3 不同发根农杆菌株对水角转化效率的影响
不同字母表示显著差异(P< 0.05)。
Fig. 3 The effect of different Agrobacterium rhizogenes strains on Hydrocera triflora transformation efficiency
Different letters indicate significant differences(P<0.05).
图 4 不同插穗类型对水角转化效率的影响
“*”表示显著差异(P< 0.05)。
Fig. 4 The effect of different types of cuttings on Hydrocera triflora transformation efficiency
"*" indicates a significant difference(P<0.05).
图 6 发根农杆菌介导表达载体pGreen-35s:MeTPS6-EGFP转化水角
A. 可见光照下非转基因水角植株;B. 可见光照下转基因水角复合植株;C. 荧光光照下非转基因水角植株;D. 荧光光照下转基因水角复合植株;E. 转基因植株毛状根PCR检测。CK+. 阳性对照,以空载体的K599为模板的PCR扩增产物;CK−. 阴性对照,以非转基因水角植株根部DNA为模板的PCR扩增产物。
Fig. 6 pGreen-35s:MeTPS6-EGFP transformed Hydrocera triflora using Agrobacterium rhizogenes mediated method
A. Non-transgenic H. triflora under visible light; B. Transgenic H. triflora under visible light; C. Non-transgenic H. triflora under fluorescent light; D. Transgenic H. triflora under fluorescent light; E. PCR detection of hairy roots in transgenic plants. CK+. Positive control, PCR amplification using empty vector K599 as template; CK−. Negative control, PCR amplification using non-transgenic plant root DNA as template.
图 7 转基因水角复合植株耐旱性检测
A. 非转基因水角植株在3% PEG-6000溶液处理3 d后叶片出现萎蔫;B. 转基因水角复合植株在3% PEG-6000溶液处理3 d后生长正常;C. 转基因水角复合植株毛状根海藻糖的检测。
Fig. 7 Drought tolerance testing of transgenic Hydrocera triflora
A. Non-transgenic H. triflora showed wilting of leaves after treated with 3% PEG-6000 solution for 3 days; B. Transgenic H. triflora composite plants grew normally after treated with 3% PEG-6000 solution for 3 days; C. Detection of trehalose in hairy roots of transgenic H. triflora composite plants.
-
[1] 中国科学院中国植物志编辑委员会. 中国植物志-第一卷-总论[M]. 北京: 科学出版社, 2004: 219. [2] 王景飞, 吕德任, 黄赛, 等. 海南省濒危水生植物水角的资源现状及调查分析[J]. 中国园艺文摘, 2017, 33(12): 67 − 69. doi: 10.3969/j.issn.1672-0873.2017.12.027 [3] FRAME B R, SHOU H X, CHIKWAMBA R K, et al. Agrobacterium tumefaciens-mediated transformation of maize embryos using a standard binary vector system[J]. Plant Physiology, 2002, 129(1): 13 − 22. doi: 10.1104/pp.000653 [4] NISHIMURA A, AICHI I, MATSUOKA M. A protocol for Agrobacterium-mediated transformation in rice[J]. Nature Protocols, 2006, 1(6): 2796 − 2802. doi: 10.1038/nprot.2006.469 [5] GUREL S, GUREL E, MILLER T I, et al. Agrobacterium-mediated transformation of Sorghum bicolor using immature embryos[M]//DUNWELL J M, WETTEN A C. Transgenic plants: methods and protocols. 2nd ed. New York: Humana Press, 2012: 109-122. [6] AGGARWAL P R, NAG P, CHOUDHARY P, et al. Genotype-independent Agrobacterium rhizogenes-mediated root transformation of chickpea: a rapid and efficient method for reverse genetics studies[J]. Plant Methods, 2018, 14(1): 55. doi: 10.1186/s13007-018-0315-6 [7] CHEN L Y, QIN L, ZHOU L L, et al. A nodule-localized phosphate transporter GmPT7 plays an important role in enhancing symbiotic N2 fixation and yield in soybean[J]. New Phytologist, 2019, 221(4): 2013 − 2025. doi: 10.1111/nph.15541 [8] FAN Y L, ZHANG X H, ZHONG L J, et al. One-step generation of composite soybean plants with transgenic roots by Agrobacterium rhizogenes-mediated transformation[J]. BMC Plant Biology, 2020, 20(1): 208. doi: 10.1186/s12870-020-02421-4 [9] WANG L J, WANG W Q, MIAO Y, et al. Development of transgenic composite Stylosanthes plants to study root growth regulated by a β-expansin gene, SgEXPB1, under phosphorus deficiency[J]. Plant Cell Reports, 2023, 42(3): 575 − 585. doi: 10.1007/s00299-023-02978-y [10] 张雯, 王宇斐, 郭延平. 高等植物6-磷酸海藻糖信号调控研究进展[J]. 植物生理学报, 2016, 52(4): 394 − 400. doi: 10.13592/j.cnki.ppj.2016.0014 [11] 丁泽红, 付莉莉, 吴春来, 等. 木薯MeTPS1基因克隆、表达及生物信息学分析[J]. 江苏农业科学, 2018, 46(9): 28 − 33. doi: 10.15889/j.issn.1002-1302.2018.09.006 [12] GARG A K, KIM J K, OWENS T G, et al. Trehalose accumulation in rice plants confers high tolerance levels to different abiotic stresses[J]. Proceedings of the National Academy of Sciences of the United States of America, 2002, 99(25): 15898 − 15903. [13] 巩涛, 刘德海, 王继雯, 等. 海藻糖合成途径及分子生物学研究进展[J]. 中国农学通报, 2016, 32(14): 62 − 67. doi: 10.3969/j.issn.1009-4229.2016.08.008 [14] 郭蓓, 胡磊, 何欣, 等. 海藻糖-6-磷酸合成酶转基因烟草提高耐盐性的研究[J]. 植物学通报, 2008, 25(1): 41 − 49. doi: 10.3969/j.issn.1674-3466.2008.01.006 [15] LI H W, ZANG B S, DENG X W, et al. Overexpression of the trehalose-6-phosphate synthase gene OsTPS1 enhances abiotic stress tolerance in rice[J]. Planta, 2011, 234(5): 1007 − 1018. doi: 10.1007/s00425-011-1458-0 [16] HAN B Y, FU L L, ZHANG D, et al. Interspecies and intraspecies analysis of trehalose contents and the biosynthesis pathway gene family reveals crucial roles of trehalose in osmotic-stress tolerance in cassava[J]. International Journal of Molecular Sciences, 2016, 17(7): 1077. doi: 10.3390/ijms17071077 [17] LI X X, ZHENG J K, YANG Y Q, et al. INCREASING NODULE SIZE1 expression is required for normal rhizobial symbiosis and nodule development[J]. Plant Physiology, 2018, 178(3): 1233 − 1248. doi: 10.1104/pp.18.01018 [18] YAN P, ZENG Y J, SHEN W T, et al. Nimble cloning: a simple, versatile, and efficient system for standardized molecular cloning[J]. Frontiers in Bioengineering and Biotechnology, 2020, 7: 460. doi: 10.3389/fbioe.2019.00460 [19] YAN P, TUO D, SHEN W T, et al. A Nimble Cloning-compatible vector system for high-throughput gene functional analysis in plants[J]. Plant Communications, 2023, 4(2): 100471. doi: 10.1016/j.xplc.2022.100471 [20] 王景飞, 符瑞侃, 任军方, 等. 濒危水生植物水角的离体培养技术[J]. 江苏农业科学, 2019, 47(3): 110 − 113. doi: 10.15889/j.issn.1002-1302.2019.03.027 [21] GUO Z L, CAO H R, ZHAO J, et al. A natural uORF variant confers phosphorus acquisition diversity in soybean[J]. Nature Communications, 2022, 13(1): 3796. doi: 10.1038/s41467-022-31555-2 [22] 梅错, 刘志鹏. 发根农杆菌介导的箭筈豌豆毛状根遗传转化体系的建立[J]. 中国草地学报, 2020, 42(5): 1 − 7. doi: 10.16742/j.zgcdxb.20190305 [23] FATHI R, MOHEBODINI M, CHAMANI E. High-efficiency Agrobacterium rhizogenes-mediated genetic transformation in Cichorium intybus L. via removing macronutrients[J]. Industrial Crops and Products, 2019, 128: 572 − 580. doi: 10.1016/j.indcrop.2018.11.050 [24] 李莉, 黄群策, 秦广雍. 海藻糖在提高植物抗逆性方面的研究进展[J]. 生物学通报, 2003, 38(6): 6 − 7. doi: 10.3969/j.issn.0006-3193.2003.06.003 [25] 丁泽红, 铁韦韦, 付莉莉, 等. 木薯海藻糖合成酶基因MeTPS6克隆及其在非生物胁迫下的表达分析[J]. 南方农业学报, 2017, 48(11): 1923 − 1929. doi: 10.3969/j.issn.2095-1191.2017.11.01 [26] 罗颖洁, 武建新, 文昭竹, 等. 白三叶萌发期耐旱性评价中PEG6000适宜浓度筛选[J]. 湖南生态科学学报, 2018, 5(4): 6 − 11. doi: 10.3969/j.issn.2095-7300.2018.04.002 [27] 李静静, 任永哲, 白露, 等. PEG-6000模拟干旱胁迫下不同基因型小麦品种萌发期抗旱性的综合鉴定[J]. 河南农业大学学报, 2020, 54(3): 368 − 377. doi: 10.16445/j.cnki.1000-2340.20200622.001 [28] GIBSON D G, YOUNG L, CHUANG R Y, et al. Enzymatic assembly of DNA molecules up to several hundred kilobases[J]. Nature Methods, 2009, 6(5): 343 − 345. doi: 10.1038/nmeth.1318 [29] HARTLEY J L, TEMPLE G F, BRASCH M A. DNA cloning using in vitro site-specific recombination[J]. Genome Research, 2000, 10(11): 1788 − 1795. doi: 10.1101/gr.143000 -

 点击查看大图
点击查看大图

计量
- 文章访问数: 1005
- HTML全文浏览量: 338
- PDF下载量: 21
- 被引次数: 0

