

-
吸血蚊虫是重要的媒介昆虫,分类上隶属于双翅目(Diptera)蚊科(Culicidae)。雌蚊以人或动物血为食,供其产卵,繁殖后代,在不同的宿主体表吸血的同时传播疟疾、乙型脑炎、登革热、黄热病、寨卡病毒病等疾病[1 − 2]。海南是重要病媒传染病乙脑和登革热的高发地区。1980—1995年对海南进行的多次媒介昆虫种类及传播疾病的调查结果显示,海南存在包括库蚊(Culex)、按蚊(Anopheles)、伊蚊(Aedes)、阿蚊(Armigeres)、曼蚊(Mansonia)等多种媒介蚊虫,它们传播的病原包括乙脑病毒(Japanese encephalitis virus)、登革病毒(Dengue virus)、盖塔病毒(Getah virus)、基孔肯雅病毒(Chikungunya virus)、罗斯河病毒(Ross River virus)、马雅罗病毒(Mayaro virus)和呼肠孤病毒科(Reoviridae)病毒等[3 − 10]。白纹伊蚊(Aedes albopictus)是登革病毒、基孔肯雅病毒、寨卡病毒的主要传播媒介。白纹伊蚊在海南省分布广泛。何昌华等[11]2012年在海南捕获白纹伊蚊928只,占捕获总量的2.29%。王晓花等[12]2012—2014年在海口市捕获白纹伊蚊186只,占捕获总量的1.2%。陈学文等[13]2017年在海口市捕获白纹伊蚊151只,占捕获总量的5.26%。范娜等[14]2017—2018年在海南省儋州市、澄迈县、文昌市、昌江黎族自治县、东方市、乐东黎族自治县共采集到白纹伊蚊35只,占捕获总量的0.23%。Li等[15]2018—2019年在海南省12个采样点捕获白纹伊蚊352只,占捕获总量的28.66%。
作为分子标记的线粒体DNA(mitochondrial DNA,mtDNA)细胞色素氧化酶C亚单位Ⅰ(Cytochrome C oxidase subunit Ⅰ,COⅠ)是有效鉴别昆虫的DNA条形码基因片段[16]。近年来,COⅠ基因序列作为 DNA 条形码已经广泛用于昆虫鉴定。同时由于COⅠ基因的母系遗传特征,COⅠ基因比核基因更易受遗传漂移的影响,为此COⅠ基因也被广泛用于昆虫种群的遗传多样性和遗传结构的历史原因和地理原因分析,使COⅠ基因成为研究分子进化遗传常用的分子标记物[16 − 18]。本研究通过扩增海南蚊虫的COⅠ基因,并利用COⅠ基因的序列来分析海南白纹伊蚊系统发育关系,为海南蚊虫的物种识别奠定基础,为了解白纹伊蚊生物多样性及对蚊媒疾病的预警提供依据。
-
于2022年10月—2024年3月采集实验蚊虫,采集地:1)海南省海口市中国热带农业科学院院区[海口,HK(19°59′09″ N, 110°19′26″ E)];2)琼海市嘉积镇长边园[琼海, QH(19°17′56″ N, 110°26′31″ E)];3)三亚市白鹭公园[三亚, SY(18°15′05″ N 109°30′58″ E)];4)儋州市两院实验农场十队[儋州,DZ(19°27′25″ N 109°40′14″ E)]。于夜间 20: 00 悬挂功夫小帅光催化诱蚊灯收集的蚊虫,次日早晨收集纱网内的虫体并浸泡在无水乙醇中保存、备用。
-
成蚊的形态学鉴定参考陆宝麟等[19 − 21]对成蚊的描述,同时参考生命条形码数据库(
http://v4.boldsystems.org/index.php ), 沃里德生物系统(http://www.wrbu.org/VecID_MQ.html )和蚊虫分类目录 (http://mosquito-taxonomic-inventory.info/valid-species-list )。 -
使用镊子夹取单只样本的成蚊蚊腿,加入多颗研磨珠,加入50 μL配制好的碱性裂解液,使用组织破碎仪破碎(频率:30/s,时间:2 min)。参考Mutebi等[22 − 23]方法,用 HotShot 法从研磨液中提取蚊虫基因组DNA,保存至−20 ℃。
-
以基因组DNA为模板,以COⅠ(Forward):GGTCAACAAATCATAAAGATATTGG,COⅠ(Reverse):TAAACTTCAGGGTGACCAAAAAATCA引物对COⅠ 基因进行PCR扩增[24 − 25]。扩增体系为 30 μL,其中DNA模板2 μL,TaqDNA聚合酶0.4 μL,上述引物各0.6 μL,10X 反应液3 μL,dNTPs 2.4 μL,ddH2O 21 μL。PCR反应条件:95 ℃ 预变性5 min;95 ℃变性30 s,45 ℃ 复性50 s,72 ℃延伸60 s,共30个循环;最后72 ℃延伸10 min。扩增反应结束后,取3 μL PCR产物进行琼脂糖凝胶电泳检测,PCR 产物经凝胶电泳检测后,切取特异性目标条带进行胶回收,回收产物送上海生工生物工程有限公司进行测序。
-
1)序列勘校:通过观察测序峰图中的测序峰是否为单一峰形,且单个峰的峰形是否完整,以确认测序是否成功,对测序前端和尾端数据进行校正,确保测序得到的碱基正确可靠。利用MEGA11对序列进行校正编辑并确保所有序列长度保持一致。将测序得到的序列与GenBank数据库收藏的序列进行同源比对,选择比对后同源性大于98%的序列进行后续分析。
2)序列分析:使用MEGA11对不同来源的COⅠ基因序列进行比对,以确定位点的变异情况。计算每个碱基(腺嘌呤、胞嘧啶、鸟嘌呤、胞腺嘧啶)的相对含量;计算碱基和密码子的使用频率。应用MEGA11邻接(neighbor-joining,NJ)算法来构建单倍型系统发育树,Bootstrap值设置为1 000。使用Dnasp6.0分析位点多态性进行种群遗传距离计算及单倍型、多样性分析、错配分布(mismatch distribution)分析。使用PopART 1.7 软件构建单倍型网络(median-joining network)图。使用Arlequin 3.1软件分析计算群体间遗传分化系数(Fst值)、基因流(Nm值)等。以及使用分子变异分析(analysis of molecular variance, AMOVA)分析种群内和种群间的遗传变异等。从GenBank 中下载中国(福建省、广东省、云南省)和越南、老挝、马来西亚、日本、韩国、意大利、美国和刚果的白纹伊蚊种群的单倍型序列进行群体间Fst和Nm值分析。
-
PCR扩增产物测序鉴定了516个样本,共检出8属43种吸血蚊虫。它们分别属于按蚊属(Anopheles)、库蚊属(Culex)、伊蚊属(Aedes)、阿蚊属(Armigeres)、巨蚊属(Toxorhynchites)、曼蚊属(Mansonia)、小蚊属(Mimomyia)和煞蚊属(Sabethes)(表1)。还有229个样本经鉴定不属于吸血蚊虫,主要为摇蚊(Chironomidae)及少量蕈蚊(Mycetophilidae)和大蚊(Tipulidae)(表2)。
表 1 PCR测序鉴定的吸血蚊虫种类、对应数量以及采样地点
Table 1. Blood-sucking mosquito species identified by sequencing, corresponding populations, and sampling locations
吸血蚊虫种类
Types of blood-sucking mosquitoes蚊虫数量 Number of mosquitoes 合计
Total总计
All海口Haikou 儋州Danzhou 三亚Sanya 琼海Qionghai 中华按蚊(An. sinensis) 4 2 1 33 278 顿尼按蚊(An. deaneorum) 4 赫坎按蚊(An. hyrcanus) 2 2 达氏按蚊(An. darlingi) 1 1 带棒按蚊(An. claviger) 2 埃文斯按蚊(An. evansae) 1 朝鲜按蚊(An. koreicus) 1 美彩按蚊(An. splendidus) 1 孟加拉按蚊(An. bengalensis) 1 吉普尔按蚊(An. jeyporiensis) 1 乌有按蚊(An. nili) 1 雷氏按蚊(An. lesteri) 1 棋斑按蚊(An. tessellatus) 1 白跗按蚊(An. albitarsis) 1 巴西按蚊(An. braziliensis) 1 里氏按蚊(An. rivulorum) 1 福拉蒂按蚊(An. forattinii) 1 瓜拉尼按蚊(An. guarani) 1 巴氏按蚊(An. Barberi) 1 簇角库蚊(C. lophoceraomyia) 2 63 白雪库蚊(C. gelidus) 6 黑点库蚊(C. nigropunctatus) 1 杂鳞库蚊(C. vishnui) 2 12 环纹库蚊(C. annulirostris) 1 C. usquatissimus 2 三带喙库蚊(C. tritaeniorhynchus) 8 尖音库蚊(C. pipiens) 2 白胸库蚊(C. pallidothorax) 2 环带库蚊(C. annulus) 6 9 二带喙库蚊(C. bitaeniorhynchus) 2 致倦库蚊(C. quinquefasciatus) 5 海滨库蚊(C. sitiens) 3 白纹伊蚊(Ae. albopictus) 82 27 31 19 173 刺扰伊蚊(Ae. vexans) 4 1 3 2 窄翅伊蚊(Ae. lineatopennis) 1 曼氏伊蚊(Ae. mcintoshi) 1 长丝伊蚊(Ae. longfilamentus) 1 伪白纹伊蚊(Ae. pseudalbopictus) 1 骚扰阿蚊(Armigeres subalbatus) 3 9 常型曼蚊(Mansonia uniformis) 1 吕宋小蚊(Mimomyia luzonensis) 2 巨蚊(Toxorhynchites ambionensis) 1 煞蚊(Sabethes glaucodaemo) 1 1 表 2 PCR测序鉴定的非吸血蚊虫种类及对应数量
Table 2. Non-blood-sucking mosquito species identified by sequencing and corresponding numbers
非吸血蚊虫种类
Non-blood-sucking mosquito species数量/只
Number/pc非吸血蚊虫种类
Non-blood-sucking mosquito species数量/只
Number/pc环带摇蚊Chironomus circumdatus 16 大蚊Limonia sp 3 克拉格摇蚊Ch. claggi 21 沼大蚊Limoniidae sp 2 粗钳摇蚊Ch. crassiforceps 1 台湾尼罗摇蚊Nilodorum tainanus 53 黄羽摇蚊Ch. flaviplumus 11 云斑多足摇蚊Polypedilum nubifer 1 爪哇摇蚊Ch. javanus 41 勾臂眼蕈蚊Sciara humeralis 1 纪伊摇蚊Ch. kiiensis 25 尖眼蕈蚊科Sciaridae sp. 12 纹翅摇蚊Ch. striatipennis 2 双突柱角蚊Styringomyia didyma 2 德永氏摇蚊Glyptotendipes tokunagai 4 台湾长足摇蚊Tanytarsus formosanus 14 毛跗球附器摇蚊Kiefferulus barbatitarsis 12 三角附器摇蚊Kiefferulus trigonum 6 按蚊属鉴定了19种,分别为中华按蚊(An. sinensis)、顿尼按蚊(An. deaneorum)、赫坎按蚊(An. hyrcanus)、达氏按蚊(An. darlingi)、带棒按蚊(An. claviger)、埃文斯按蚊(An. evansae)、朝鲜按蚊(An. koreicus)、美彩按蚊(An. splendidus)、孟加拉按蚊(An. bengalensis)、吉普尔按蚊(An. jeyporiensis)、乌有按蚊(An. nili)、雷氏按蚊(An. lesteri)、棋斑按蚊(An. tessellatus)、白跗按蚊(An. albitarsis)、巴西按蚊(An. Braziliensis)、里氏按蚊(An. Rivulorum)、福拉蒂按蚊(An. Forattinii)、瓜拉尼按蚊(An. Guarani)和巴氏按蚊(An. Barberi)。库蚊属鉴定了13种,分别为簇角库蚊(Culex Lophoceraomyia)、白雪库蚊(C. gelidus)、黑点库蚊(C. nigropunctatus)、杂鳞库蚊(C. vishnui)、环纹库蚊(C. annulirostris)、C. usquatissimus、三带喙库蚊(C. tritaeniorhynchus)、尖音库蚊(C. pipiens)、白胸库蚊(C. pallidothorax)、环带库蚊(C. annulus)、二带喙库蚊(C. bitaeniorhynchus)、致倦库蚊(C. quinquefasciatus)、海滨库蚊(C. sitiens)。杂鳞库蚊和环带库蚊占比较多(22.2%、23.8%)。其中,在琼海有数量较多的三带喙库蚊(12.6%)。三带喙库蚊是流行性乙型脑炎主要传播媒介,需提高警惕。本研究在海口检测到5只致倦库蚊,致倦库蚊也传播流行性乙型脑炎。伊蚊属中鉴定了6个种,分别为白纹伊蚊(Ae. albopictus)、刺扰伊蚊(Ae. vexans)、曼氏伊蚊(Ae. mcintoshi)、窄翅伊蚊(Ae. lineatopennis)、长丝伊蚊(Ae. longfilamentus)和伪白纹伊蚊(Ae. pseudalbopictus)。白纹伊蚊数量占总伊蚊比例最多(91.9%)。其中,海口、儋州和三亚白纹伊蚊数量占总白纹伊蚊比例较高(51.6%、16.9.6%、19.4%),琼海(11.9%)较低。骚扰阿蚊(Armigeres subalbatus)、吕宋小蚊(Mimomyia luzonensis)、巨蚊(Toxorhynchites ambionensis)、常型曼蚊(Mansonia uniformis)、煞蚊(Sabethes glaucodaemo)数量稀少,骚扰阿蚊仅在儋州发现,吕宋小蚊和煞蚊是首次在海南使用分子标记发现的新记录种(表1)。
-
经过测序得到白纹伊蚊COⅠ基因序列118条,去除头尾引物序列,有效序列长度为600 bp。同源比对发现白纹伊蚊COⅠ基因扩增片段同源率高达99%。分析显示,白纹伊蚊COⅠ基因序列的碱基组成为A:28.5%,T:36.8%,C:17.3%,G:16.0%。A+T含量(65.3%)大于G+C 含量(34.7%);该序列具有明显的碱基偏好,主要为A和T,特别是第三位碱基,A+T的含量达到86.0%(表3)。在此基础上,本研究团队统计分析白纹伊蚊COⅠ基因扩增片段的密码子的使用频率,扩增的COⅠ基因片段不编码丙氨酸和色氨酸。使用频率最高的为亮氨酸(Leu)(30.6%),其次为谷氨酸(11.0%),使用频率最低的密码子是天冬酰胺(0.6%)(表4)。
表 3 白纹伊蚊COⅠ基因序列不同位点脱氧核糖核苷酸组成
Table 3. Deoxyribonucleotide composition of different sites in the COⅠ gene of Ae. albopictus
% 位点(Site) A T C G A+T 第一位碱基(First base) 30.0 23.5 18.0 28.5 53.5 第二位碱基(Second base) 13.0 43.5 27.5 16.0 56.5 第三位碱基(Third base ) 42.5 43.5 6.5 3.5 86.0 平均含量(Average) 28.5 36.8 17.3 16.0 65.3 表 4 白纹伊蚊COⅠ基因编码氨基酸的使用频率
Table 4. Frequency of use of amino acids encoded by the COⅠ gene for Ae. albopictus
% Ala Cys Asp Glu Phe Gly His Ile Lys Leu 0.0 1.7 3.5 11.0 9.8 1.2 2.3 8.7 1.2 30.6 Met Asn Pro Gln Arg Ser Thr Val Trp Tyr 2.3 0.6 4.1 6.4 1.2 1.2 2.3 2.9 0.0 9.3 注:Ala:丙氨酸;Cys:半胱氨酸;Asp:天冬氨酸;Glu:谷氨酸;Phe:苯丙氨酸;Gly:甘氨酸;His:组氨酸;Ile:异亮氨酸;Lys:赖氨酸;Leu:亮氨酸;Met:甲硫氨酸;Asn:天冬酰胺;Pro:脯氨酸;Gln:谷氨酰胺;Arg:精氨酸;Ser:丝氨酸;Thr:苏氨酸;Val:缬氨酸;Trp:色氨酸;Tyr:酪氨酸 Note:Ala: alanine; Cys: cysteine; Asp: aspartine; Glu: glutamine; Phe: phenylalanine; Gly: glycine; His: histidine; Ile: isoleucine; Lys: lysine; Leu: leucine; Met: methionine; Asn: asparagine; Pro: proline; Gln: glutamine; Arg: arginine; Ser: serine; Thr: threonine; Val: valine; Trp: tryptophan; Tyr: tyrosine. -
使用Dnasp6.0对各组的白纹伊蚊COⅠ基因的群体遗传距离进行计算(表5)。各个种群组内遗传距离最大的为三亚种群(0.002 0),其次为海口(0.001 9)、琼海(0.001 3)和儋州(0.000 7)种群。各种群间遗传距离变化范围为 0.001 10~0.001 97,除了琼海种群和儋州种群(0.001 10)距离较近外,其他种群两两间距离差别不大(0.001 69~0.001 97)。这说明海南4个采样区域间白纹伊蚊遗传差异较小。
表 5 海南白纹伊蚊各地理种群COⅠ基因的遗传距离
Table 5. Genetic distances of COⅠ genes among geographic populations of Ae. albopictus genes in Hainan
种群Population 群内Within population 群间Between population 儋州Danzhou 海口Haikou 琼海Qionghai 儋州Danzhou 0.0007 海口Haikou 0.0019 0.00177 琼海Qionghai 0.0013 0.00110 0.00178 三亚Sanya 0.0020 0.00169 0.00197 0.00172 通过Dnasp6.0分析测序数据,共获得各组白纹伊蚊COⅠ基因单倍型23种(表6)。其中共享单倍型共5个,Hap_1为 4个地理种群共享(海口、琼海、三亚、儋州),Hap_3 为3个地理种群共享(海口、琼海、三亚),Hap_4、Hap_13为海口与琼海共享,Hap_10为海口和三亚共享。这说明海南的白纹伊蚊主要在Hap_1和Hap_3 2个位点发生基因交流。进而,通过MEGA 11邻接(Neighbor-Joining,NJ)算法构建单倍型系统发育树,对海南白纹伊蚊23个单倍型进行分析,结果(图1−A)显示,Hap_1、Hap_2、Hap_3、Hap_7、Hap_5、Hap_13、Hap_23、Hap_12和Hap_16聚为1大支。Hap_2、Hap_14、Hap_11和Hap_22,Hap_6、Hap_8、Hap_15、Hap_17、Hap_19和Hap_9,Hap_18、Hap_10和Hap_21则各聚为1支。通过 PopART 1.7 软件构建的23个单倍型网络(median-joining network)(图1−B)显示,Hap_1(被全部4个地理种群所共享)为优势单倍型,基本所有的单倍型都以Hap_1为中心,其他单倍型则可能从Hap_1演化而来。而Hap_3也成为一个小中心。各单倍型之间相差一个突变步骤(图1−B)。
表 6 海南白纹伊蚊各地理种群COⅠ基因的单倍型数量及分布
Table 6. Haplotype number and distribution of COⅠ gene in various geographical populations of Ae. albopictus in Hainan
单倍型Hapotape 种群Populations 儋州Danzhou 海口Haikou 琼海Qionghai 三亚Sanya Hap_1 8 37 11 11 Hap_2 1 0 0 0 Hap_3 0 6 4 6 Hap_4 0 1 1 0 Hap_5 0 1 0 0 Hap_6 0 1 0 0 Hap_7 0 2 0 0 Hap_8 0 2 0 0 Hap_9 0 1 0 0 Hap_10 0 2 0 1 Hap_11 0 2 0 0 Hap_12 0 2 0 0 Hap_13 0 2 1 0 Hap_14 0 2 0 0 Hap_15 0 1 0 0 Hap_16 0 2 0 0 Hap_17 0 1 0 0 Hap_18 0 1 0 0 Hap_19 0 0 1 0 Hap_20 0 0 1 0 Hap_21 0 0 0 4 Hap_22 0 0 0 1 Hap_23 0 0 0 1 合计 9 66 19 24 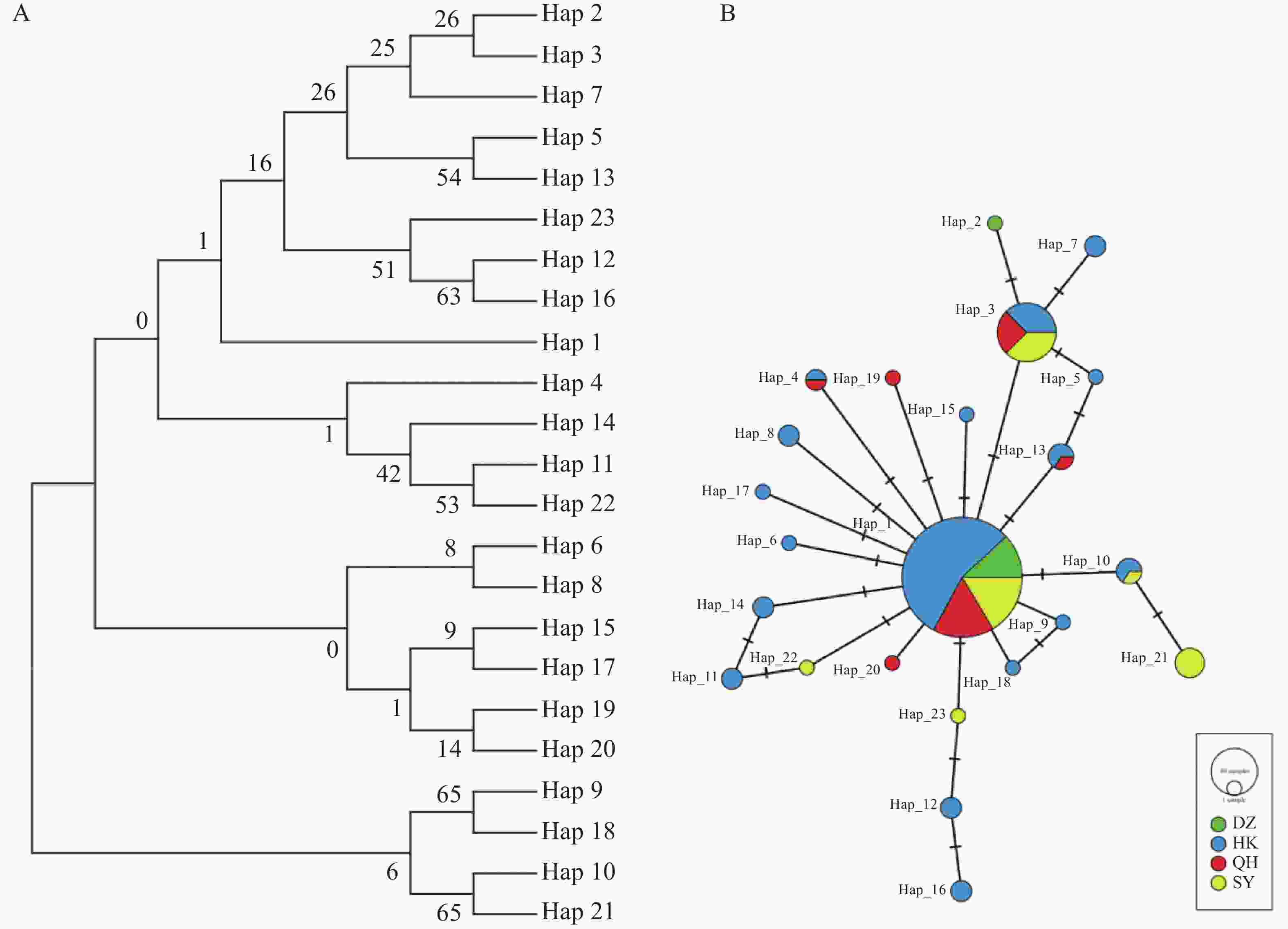
图 1 海南白纹伊蚊地理种群遗传结构和遗传多样性分析
Figure 1. Analysis of the genetic structure and genetic diversity of geographic populations of Ae. albopictus in Hainan
对海南白纹伊蚊遗传多样性分析发现三亚的白纹伊蚊单倍型多样性最高(0.73),核苷酸多样性也是最高为(0.001 99)。其次为海口(单倍型多样性0.68,核苷酸多样性0.001 91)。儋州的单倍型多样性(0.22)和核酸多样性(0.000 74)最低。虽然海口的单倍型数量(17)最多,单倍型多样性(0.68)很高,但有意思的是,三亚单倍体数量并不多,但单倍多样性和核酸多样性均是最高,说明三亚的遗传多样性较为丰富(表7)。
表 7 白纹伊蚊种群的遗传多样性
Table 7. Genetic diversity of Ae. albopictus populations
种群
Population样本量
(N)单倍型数量
Number of
haplotypes单倍型多
样性(Hd)核酸多
样性(Pi)儋州Danzhou 9 2 0.22 0.00074 海口Haikou 66 17 0.68 0.00191 琼海Qionghai 19 6 0.64 0.00129 三亚Sanya 24 6 0.73 0.00199 总计Total 118 23 0.66 0.00177 -
分子变异分析(Analysis of molecular variance, AMOVA)结果表明:海南4个种群间COⅠ基因的遗传差异占11.51%,而种群内部的差异占88.49%。使用软件Arlequin 3.1计算白纹伊蚊 COⅠ基因的种群间遗传分化系数Fst,根据Fst值计算基因流水平(1−Fst)/(4*Fst)(表8)。按照Fst<0.05为种群出现轻度分化,0.05<Fst<0.15为种群出现中度分化,0.15<Fst为种群出现高度分化[26]。结果显示,三亚种群和儋州种群(Fst=0.065,Nm=3.6)为中度分化,说明三亚种群和儋州种群基因有一定交流。其他种群两两之间(Fst<0.05,Nm>5.18)处于轻度分化, 其他种群基因交流频繁。同样,本研究团队还分析了海南白纹伊蚊种群与国内外其他地区种群的Fst值和Nm值(表9)。海南种群与福建种群(Fst = 0.65)、意大利种群(Fst = 0.61)、日本种群(Fst = 0.61)、韩国种群(Fst = 0.58)之间,遗传分化较大。而海南种群与云南种群(Fst = 0.27)、老挝种群(Fst = 0.22)、越南种群(Fst = 0.19)、马来西亚种群(Fst = 0.28)Fst 值较小,而Nm值较大(1.26−2.15)。表明海南种群与云南种群、老挝种群、越南种群、马来西亚种群Fst 值相对较小,但依然是高度分化,基因交流程度依然比较低。
表 8 海南白纹伊蚊种群间COⅠ基因遗传分化(Fst:下三角);基因流(Nm:上三角)
Table 8. Genetic differentiation of COⅠ genes among different populations (Fst: lower triangle); Gene flow (Nm: upper triangle)
Population 儋州Danzhou 海口Haikou 琼海Qionhai 三亚Sanya 儋州Danzhou 83.08 12.25 3.60 海口Haikou 0.003 62.25 5.18 琼海Qionghai 0.020 0.004** 5.85 三亚Sanya 0.065* 0.046* 0.041** 注:−,表示无穷大;*,表示p≤0.05;**,表示p≤0.01。 Note: −, denotes infinity; *, p≤0.05; **, p≤0.01. 表 9 白纹伊蚊不同地理种群之间的遗传分化(Fst;下三角)和基因流(Nm;上三角)
Table 9. Genetic differentiation (Fst; lower triangle) and gene flow (Nm; upper triangle) between different geographical populations of Ae.albopictus
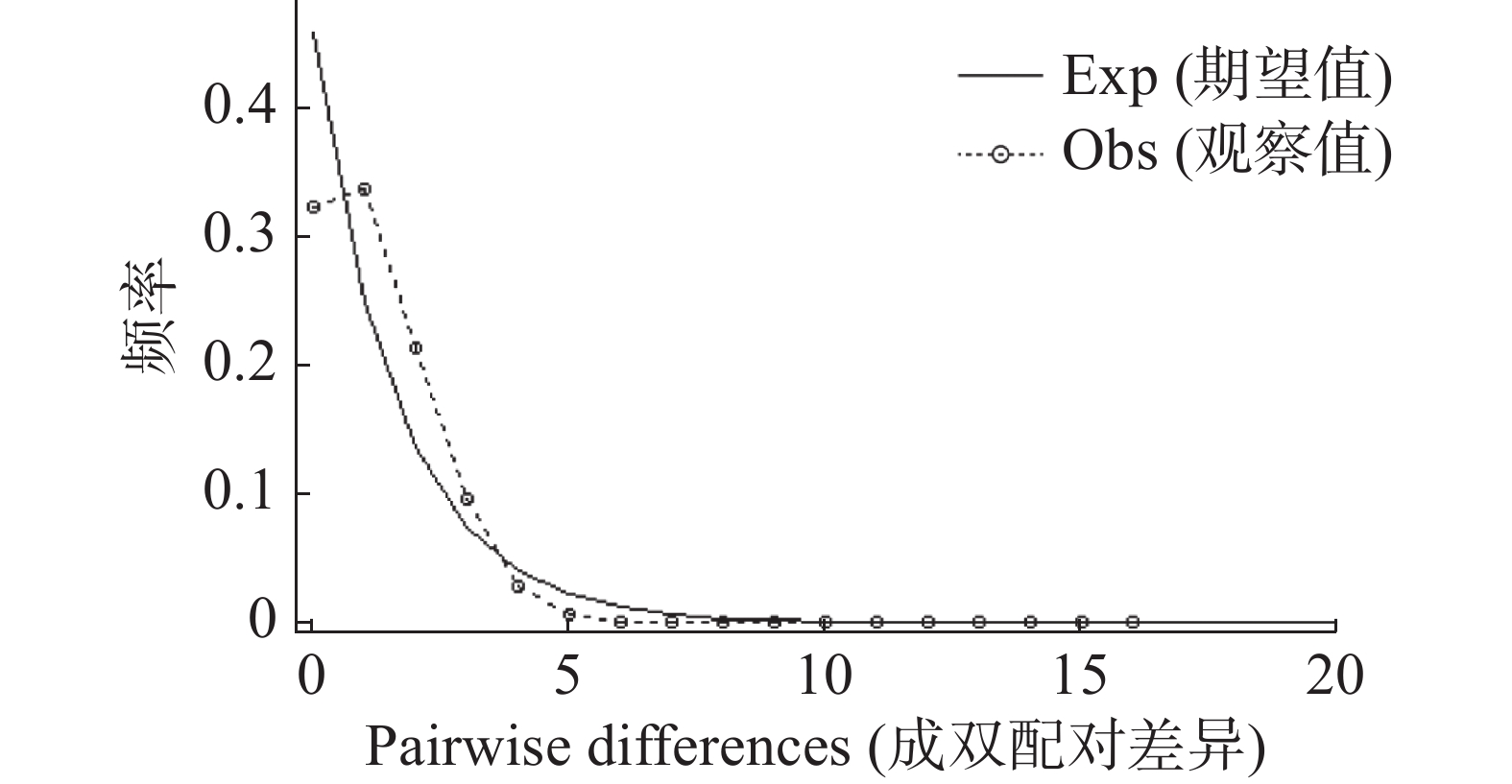
Country/Region HN FJ GD YN USA IT LO JP VN ML SK CN HN 0.27 0.45 1.37 0.59 0.32 1.75 0.32 2.15 1.26 0.36 FJ 0.65** 2.60 0.36 1.49 0.41 0.49 0.87 1.52 2.86 0.90 GD 0.52** 0.16* 1.22 11.83 5.21 0.89 5.98 3.37 3.11 2.67 YN 0.27** 0.58** 0.29** 1.63 0.60 6.32 0.46 73.05 3.23 0.78 USA 0.46** 0.25** 0.04 0.23** 4.55 1.28 3.98 4.06 1.96 2.77 IT 0.61** 0.55** 0.09** 0.46** 0.10** 0.70 2.18 2.50 2.91 2.12 LO 0.22** 0.51** 0.36** 0.02 0.28** 0.42** 0.64 9.88 1.81 0.67 JP 0.61** 0.36** 0.08 0.52** 0.11** 0.19** 0.44** 2.33 2.13 1.73 VN 0.19** 0.25** 0.13** 0.01 0.11** 0.17** 0.05** 0.18** 1.63 1.61 ML 0.28** 0.15* 0.14* 0.13 0.20** 0.15* 0.22** 0.19** 0.23** 2.00 SK 0.58** 0.36** 0.16** 0.39** 0.15** 0.19** 0.43** 0.22** 0.24** 0.20** 注:HN:中国海南(101个样本,Genbank登录号:PQ858251-PQ858367);FJ:中国福建(8个样本,KY971602.1-KY971596.1;KY971591.1);GD:中国广东(9个样本,KX886287.1-KX886295.1);YN:中国云南(9个样本,KY765501.1;KY765474.1-KY765467.1);USA:美国(66个样本,KC690896.1-KC690961.1);IT:意大利(9个样本,JX679373.1-JX679380.1);LO:老挝(46个样本,MN080765.1-MN080720.1);JP:日本(21个样本,AB738310.1;AB738308.1;AB738294.1;AB738241.1;AB738240.1;AB738211.1;AB738203.1;AB738197.1;AB738192.1;AB738179.1;AB738161.1;AB738143.1;AB738122.1-AB738121.1;AB738090.1;LC544010.1;LC544012.1;LC544014.1;LC544013.1;LC544009.1;LC544011.1);VN:越南(65个样本,MZ573376.1-MZ573312);ML:马来西亚(90个样本,MF148303.1-MF148249.1;KY982366.1-KY982334.1);SK:韩国(24个样本,MG871379.1-MG871402.1)。*p≤0.05,**p≤0.01。 如果错配分布的实际观察结果与期望值的某个假定模型(恒定或收缩)相吻合,则说明群体状态与期望的假定模型一致,反之,则说明背离期望值的假定模型,群体处于扩张状态(曲线为单峰)。由图2可见海南的白纹伊蚊种群的分布为单峰分布,与期望的分布不一致,说明海南白纹伊蚊种群经历过扩张。
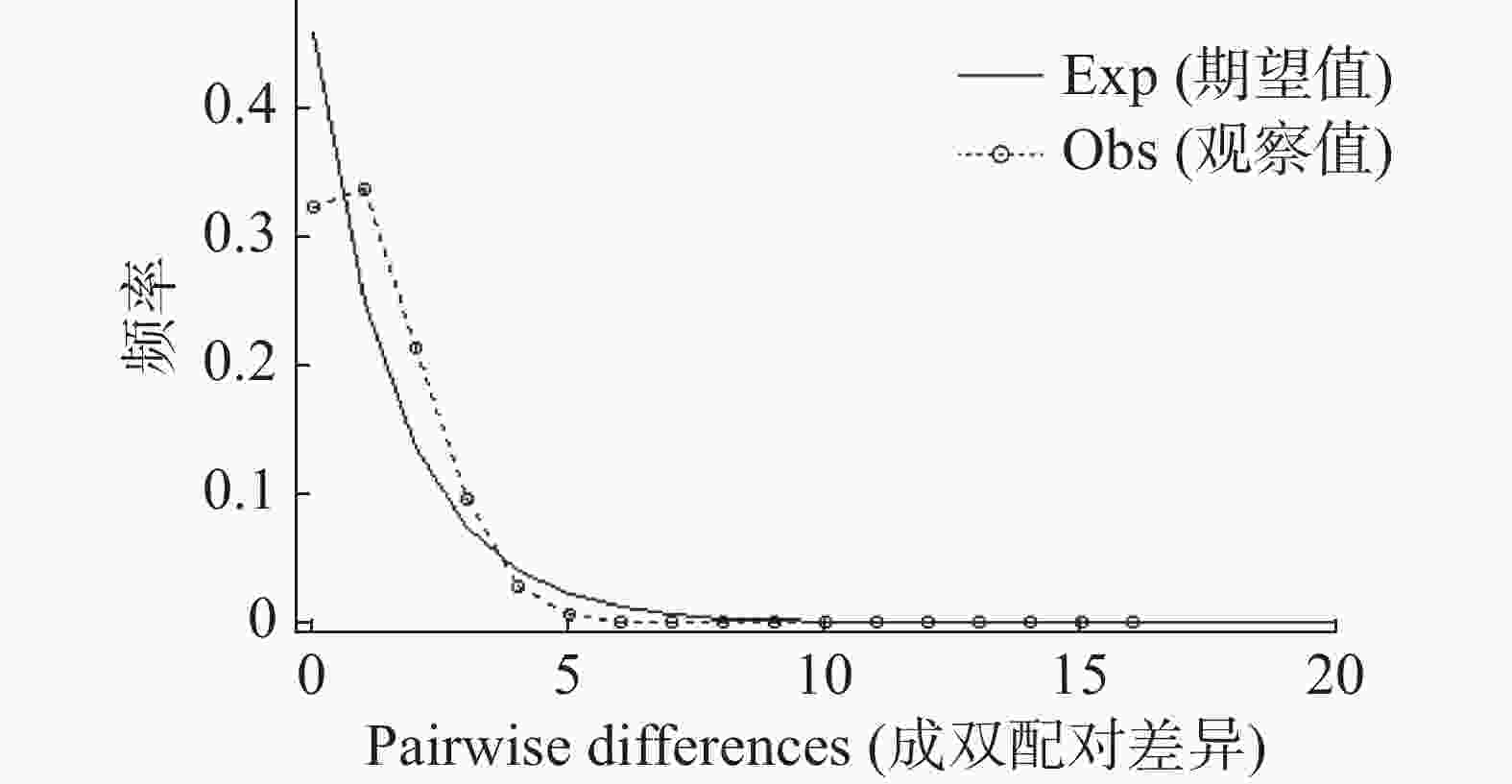
图 2 海南白纹伊蚊基于线粒体COⅠ基因的种群错配分析
Figure 2. Population mismatch analysis of Hainan Ae.albopictus based on mitochondrial COⅠ gene
-
海南岛是中国南方的一个热带岛屿,蚊种丰富,有面临蚊媒疾病暴发的风险。COⅠ基因是研究蚊虫遗传多样性和建立种内关系有价值且可靠的诊断工具。随着数据的丰富,COⅠ基因鉴定蚊虫种类越来越有效[27 − 28]。本研究从海南岛东西南北4个地理区域收集蚊虫并研究蚊虫的多样性和遗传分化情况,共捕获8属43种吸血蚊种,它们分别属于按蚊属(Anopheles)、库蚊属(Culex)、伊蚊属(Aedes)、阿蚊属(Armigeres)、巨蚊属(Toxorhynchites)、曼蚊属(Mansonia)、小蚊属(Mimomyia)和煞蚊属(Sabethes)。其中,主要蚊虫为按蚊属19种,库蚊属13种,伊蚊属6种。吕宋小蚊和煞蚊(S. glaucodaemo)是首次在海南使用分子标记发现。测序得到的COⅠ基因有效序列长度为600 bp,该短序列碱基组成为A:28.5%,T:38.2%,C:16.3%,G:16.0%。A+T含量(66.7%)大于 G+C 含量(32.3%)。碱基组成主要为A和T,特别是第三位碱基,A+T的含量达到86%,这个结果与其他学者对线粒体COⅠ基因研究结果相似[24, 29 − 30]。本研究统计了COⅠ基因编码蛋白的氨基酸组成,其编码18种氨基酸,其中,亮氨酸密码子的使用频率最高,该结果与对天牛中线粒体基因的研究结果相似[31]。通过分析海南4个地理种群的遗传距离发现,儋州、海口、三亚、琼海种群间遗传距离较小,种群两两间距离差别不大。这说明海南4个采样区域间白纹伊蚊遗传差异较小,与 Li 等[15]研究结果相似。
种群分化系数(Fst)又称为固定系数(Fixationindex),表示种群之间的遗传分化程度,范围在0~1之间,值越大表明种群的分化程度越高。基因流(Nm)表示基因在群体中的流动,Nm越大说明群体之间交流越频繁,Nm小于1时表明种群间基因交流极少,存在很大分化,当Nm的值大于1时可以抑制群体间的分化[32]。分子变异分析表明海南4个区域中,除了三亚种群和儋州种群(Fst=0.065,Nm=3.6)之间为中度分化,基因有一定交流外,其他种群两两之间(Fst<0.05,Nm>5.18)处于轻度分化,基因交流频繁。系统发育树中各单倍型的聚类也没有表现出明显的地域特征,这可能与海南地理隔离程度较低,不同地区之间存在一定较频繁基因交流相关。将海南种群与其他地区的种群进行分子变异分析发现,中国海南种群与中国福建种群、意大利种群、日本种群、韩国种群之间,遗传分化较大。研究还发现,中国海南白纹伊蚊种群与中国云南种群、老挝种群、越南种群、马来西亚种群Fst 值相对较小,有一定的基因交流。Fang等[33]研究发现,中国海南白纹伊蚊仅与中国云南种群表现出一定的基因流动,中国海南与中国广东或中国福建种群之间的基因交流受阻。本研究结果与Fang等[33]研究结果类似。Li等[15]研究结果显示中国海南种群与中国云南、老挝种群之间存在一定的基因流动。研究结果提醒:海南自由贸易港建成后,人员会频繁流动,蚊卵有可能会被东南亚国家或非洲务工人员携带入岛,进而可能会造成白纹伊蚊传播的登革热等疾病风险升高,故建议政府应做好相应应对措施,防范输入性病例的出现。通过使用DnaSP6.0分析中国海南白纹伊蚊进化过程,结果显示,海南白纹伊蚊的错配分布曲线呈现单峰泊松分布,海南白纹伊蚊种群可能发生过一次群体扩张,群体大小保持相对稳定[34 − 35]。
本研究丰富了海南蚊虫的分子鉴定的内容,为海南白纹伊蚊的遗传结构和种群动态提供了新的认识,为进一步研究该物种的生态学、病原体传播机制及防控策略提供了重要线索。研究发现儋州、海口、三亚、琼海各种群间遗传距离较小,种群两两之间大多数处于轻度分化,种群之间基因交流较频繁,各种群间的遗传多样性不高。这些使得海南不同地理种群的白纹伊蚊具有相似的媒介能力[36]。本研究还存在一些局限性,未来可以进一步完善和扩展,通过更全面的地理区域采样来了解海南白纹伊蚊的生态和遗传特征。
利用COⅠ基因鉴定海南省蚊种及白纹伊蚊种群的遗传多样性分析
DOI: 10.15886/j.cnki.rdswxb.20240179
 CSTR: 32425.14.j.cnki.rdswxb.20240179
CSTR: 32425.14.j.cnki.rdswxb.20240179
Identification of mosquito species in Hainan Province using COⅠ gene and genetic diversity analyses of Aedes albopictus populations
-
摘要: 为获得海南最新蚊子种类信息,分析登革热传播媒介白纹伊蚊的种群遗传多样性及地理种群间分化和基因流状况,本研究从海南省4个地理区域收集蚊虫,PCR扩增COⅠ基因并测序,通过Blast比较鉴定蚊种。使用MEGA11对不同来源白纹伊蚊COⅠ基因序列进行比对,计算碱基的相对含量,计算碱基和密码子的使用频率。应用邻接算法来构建单倍型系统发育树。使用Dnasp6.0分析位点多态性、种群遗传距离、单倍型和多样性分析,以及错配分布分析。使用PopART 1.7 软件构建单倍型网络图。使用Arlequin 3.1软件分析计算群体间遗传分化系数(Fst值)、基因流(Nm值)。使用分子变异分析(Analysis of molecular variance, AMOVA)法分析种群内和种群间的遗传变异。研究结果表明, 共捕获8属43种吸血蚊种,它们分别属于按蚊属(Anopheles)、库蚊属(Culex)、伊蚊属(Aedes)、阿蚊属(Armigeres)、巨蚊属(Toxorhynchites)、曼蚊属(Mansonia)、小蚊属(Mimomyia)和煞蚊属(Sabethes)。共鉴定了19种按蚊、13种库蚊、6种伊蚊,以及少量的骚扰阿蚊(Armigeres subalbatus)、吕宋小蚊(Mimomyia luzonensis)、巨蚊(Toxorhynchites ambionensis)、常型曼蚊(Mansonia uniformis)和煞蚊(Sabethes glaucodaemo)。研究发现白纹伊蚊三亚种群和儋州种群之间为中度分化,其他种群两两之间处于轻度分化,基因交流频繁。白纹伊蚊种群中的中国海南种群与中国云南种群和老挝种群、越南种群、马来西亚种群之间遗传分化较低,存在明显的基因交流。这揭示海南岛外的病毒有传进岛内的安全隐患,本研究结果对海南岛公共卫生具有重要的意义。
-
关键词:
- 白纹伊蚊 /
- 线粒体细胞色素c氧化酶亚基Ⅰ /
- 种群遗传多样性
Abstract: Obtaining up-to-date information on mosquito species in Hainan and analysing population genetic diversity and the status of differentiation and gene flow among geographic populations of the dengue vector Ae. albopictus. Mosquitoes were collected from four geographical areas in Hainan Province, the COⅠ gene was PCR amplified and sequenced, and mosquito species were identified by Blast comparison. The COⅠ gene sequences of Ae. albopictus from different sources were compared using MEGA11 to calculate the relative content of bases and the frequency of base and codon usage. Neighbour-joining algorithm was applied to construct the haplotype phylogenetic tree. Dnasp 6.0 was used to analyse locus polymorphisms, population genetic distances, haplotype and diversity analyses, and mismatch distribution analyses. Haplotype network diagrams were constructed using PopART 1.7 software. Arlequin 3.1 software was used to calculate the coefficient of genetic differentiation (Fst value) and gene flow (Nm value) between populations. Genetic variation within and between populations was analysed using Analysis of molecular variance (AMOVA). A total of 8 genera and 43 species of blood-sucking mosquitoes belonging to Anopheles, Culex, Aedes, Armigeres, Toxorhynchites, Mansonia, Mimomyia and Sabethes were captured. Ae. albopictus was found to be moderately differentiated between the Sanya and Danzhou populations, while the other populations were mildly differentiated, with frequent gene exchange. The genetic differentiation between Aedes albopictus populations in Hainan and populations in Yunnan, Laos, Vietnam and Malaysia was low, and there was obvious gene exchange. It reveals that the virus outside the island has a safety risk of being transmitted into the island, which is of great public health significance. -
图 2 海南白纹伊蚊基于线粒体COⅠ基因的种群错配分析
Fig. 2 Population mismatch analysis of Hainan Ae.albopictus based on mitochondrial COⅠ gene
表 1 PCR测序鉴定的吸血蚊虫种类、对应数量以及采样地点
Table 1 Blood-sucking mosquito species identified by sequencing, corresponding populations, and sampling locations
吸血蚊虫种类
Types of blood-sucking mosquitoes蚊虫数量 Number of mosquitoes 合计
Total总计
All海口Haikou 儋州Danzhou 三亚Sanya 琼海Qionghai 中华按蚊(An. sinensis) 4 2 1 33 278 顿尼按蚊(An. deaneorum) 4 赫坎按蚊(An. hyrcanus) 2 2 达氏按蚊(An. darlingi) 1 1 带棒按蚊(An. claviger) 2 埃文斯按蚊(An. evansae) 1 朝鲜按蚊(An. koreicus) 1 美彩按蚊(An. splendidus) 1 孟加拉按蚊(An. bengalensis) 1 吉普尔按蚊(An. jeyporiensis) 1 乌有按蚊(An. nili) 1 雷氏按蚊(An. lesteri) 1 棋斑按蚊(An. tessellatus) 1 白跗按蚊(An. albitarsis) 1 巴西按蚊(An. braziliensis) 1 里氏按蚊(An. rivulorum) 1 福拉蒂按蚊(An. forattinii) 1 瓜拉尼按蚊(An. guarani) 1 巴氏按蚊(An. Barberi) 1 簇角库蚊(C. lophoceraomyia) 2 63 白雪库蚊(C. gelidus) 6 黑点库蚊(C. nigropunctatus) 1 杂鳞库蚊(C. vishnui) 2 12 环纹库蚊(C. annulirostris) 1 C. usquatissimus 2 三带喙库蚊(C. tritaeniorhynchus) 8 尖音库蚊(C. pipiens) 2 白胸库蚊(C. pallidothorax) 2 环带库蚊(C. annulus) 6 9 二带喙库蚊(C. bitaeniorhynchus) 2 致倦库蚊(C. quinquefasciatus) 5 海滨库蚊(C. sitiens) 3 白纹伊蚊(Ae. albopictus) 82 27 31 19 173 刺扰伊蚊(Ae. vexans) 4 1 3 2 窄翅伊蚊(Ae. lineatopennis) 1 曼氏伊蚊(Ae. mcintoshi) 1 长丝伊蚊(Ae. longfilamentus) 1 伪白纹伊蚊(Ae. pseudalbopictus) 1 骚扰阿蚊(Armigeres subalbatus) 3 9 常型曼蚊(Mansonia uniformis) 1 吕宋小蚊(Mimomyia luzonensis) 2 巨蚊(Toxorhynchites ambionensis) 1 煞蚊(Sabethes glaucodaemo) 1 1  下载: 导出CSV
下载: 导出CSV
表 2 PCR测序鉴定的非吸血蚊虫种类及对应数量
Table 2 Non-blood-sucking mosquito species identified by sequencing and corresponding numbers
非吸血蚊虫种类
Non-blood-sucking mosquito species数量/只
Number/pc非吸血蚊虫种类
Non-blood-sucking mosquito species数量/只
Number/pc环带摇蚊Chironomus circumdatus 16 大蚊Limonia sp 3 克拉格摇蚊Ch. claggi 21 沼大蚊Limoniidae sp 2 粗钳摇蚊Ch. crassiforceps 1 台湾尼罗摇蚊Nilodorum tainanus 53 黄羽摇蚊Ch. flaviplumus 11 云斑多足摇蚊Polypedilum nubifer 1 爪哇摇蚊Ch. javanus 41 勾臂眼蕈蚊Sciara humeralis 1 纪伊摇蚊Ch. kiiensis 25 尖眼蕈蚊科Sciaridae sp. 12 纹翅摇蚊Ch. striatipennis 2 双突柱角蚊Styringomyia didyma 2 德永氏摇蚊Glyptotendipes tokunagai 4 台湾长足摇蚊Tanytarsus formosanus 14 毛跗球附器摇蚊Kiefferulus barbatitarsis 12 三角附器摇蚊Kiefferulus trigonum 6
下载: 导出CSV
表 3 白纹伊蚊COⅠ基因序列不同位点脱氧核糖核苷酸组成
Table 3 Deoxyribonucleotide composition of different sites in the COⅠ gene of Ae. albopictus
% 位点(Site) A T C G A+T 第一位碱基(First base) 30.0 23.5 18.0 28.5 53.5 第二位碱基(Second base) 13.0 43.5 27.5 16.0 56.5 第三位碱基(Third base ) 42.5 43.5 6.5 3.5 86.0 平均含量(Average) 28.5 36.8 17.3 16.0 65.3
下载: 导出CSV
表 4 白纹伊蚊COⅠ基因编码氨基酸的使用频率
Table 4 Frequency of use of amino acids encoded by the COⅠ gene for Ae. albopictus
% Ala Cys Asp Glu Phe Gly His Ile Lys Leu 0.0 1.7 3.5 11.0 9.8 1.2 2.3 8.7 1.2 30.6 Met Asn Pro Gln Arg Ser Thr Val Trp Tyr 2.3 0.6 4.1 6.4 1.2 1.2 2.3 2.9 0.0 9.3 注:Ala:丙氨酸;Cys:半胱氨酸;Asp:天冬氨酸;Glu:谷氨酸;Phe:苯丙氨酸;Gly:甘氨酸;His:组氨酸;Ile:异亮氨酸;Lys:赖氨酸;Leu:亮氨酸;Met:甲硫氨酸;Asn:天冬酰胺;Pro:脯氨酸;Gln:谷氨酰胺;Arg:精氨酸;Ser:丝氨酸;Thr:苏氨酸;Val:缬氨酸;Trp:色氨酸;Tyr:酪氨酸 Note:Ala: alanine; Cys: cysteine; Asp: aspartine; Glu: glutamine; Phe: phenylalanine; Gly: glycine; His: histidine; Ile: isoleucine; Lys: lysine; Leu: leucine; Met: methionine; Asn: asparagine; Pro: proline; Gln: glutamine; Arg: arginine; Ser: serine; Thr: threonine; Val: valine; Trp: tryptophan; Tyr: tyrosine.
下载: 导出CSV
表 5 海南白纹伊蚊各地理种群COⅠ基因的遗传距离
Table 5 Genetic distances of COⅠ genes among geographic populations of Ae. albopictus genes in Hainan
种群Population 群内Within population 群间Between population 儋州Danzhou 海口Haikou 琼海Qionghai 儋州Danzhou 0.0007 海口Haikou 0.0019 0.00177 琼海Qionghai 0.0013 0.00110 0.00178 三亚Sanya 0.0020 0.00169 0.00197 0.00172
下载: 导出CSV
表 6 海南白纹伊蚊各地理种群COⅠ基因的单倍型数量及分布
Table 6 Haplotype number and distribution of COⅠ gene in various geographical populations of Ae. albopictus in Hainan
单倍型Hapotape 种群Populations 儋州Danzhou 海口Haikou 琼海Qionghai 三亚Sanya Hap_1 8 37 11 11 Hap_2 1 0 0 0 Hap_3 0 6 4 6 Hap_4 0 1 1 0 Hap_5 0 1 0 0 Hap_6 0 1 0 0 Hap_7 0 2 0 0 Hap_8 0 2 0 0 Hap_9 0 1 0 0 Hap_10 0 2 0 1 Hap_11 0 2 0 0 Hap_12 0 2 0 0 Hap_13 0 2 1 0 Hap_14 0 2 0 0 Hap_15 0 1 0 0 Hap_16 0 2 0 0 Hap_17 0 1 0 0 Hap_18 0 1 0 0 Hap_19 0 0 1 0 Hap_20 0 0 1 0 Hap_21 0 0 0 4 Hap_22 0 0 0 1 Hap_23 0 0 0 1 合计 9 66 19 24
下载: 导出CSV
表 7 白纹伊蚊种群的遗传多样性
Table 7 Genetic diversity of Ae. albopictus populations
种群
Population样本量
(N)单倍型数量
Number of
haplotypes单倍型多
样性(Hd)核酸多
样性(Pi)儋州Danzhou 9 2 0.22 0.00074 海口Haikou 66 17 0.68 0.00191 琼海Qionghai 19 6 0.64 0.00129 三亚Sanya 24 6 0.73 0.00199 总计Total 118 23 0.66 0.00177
下载: 导出CSV
表 8 海南白纹伊蚊种群间COⅠ基因遗传分化(Fst:下三角);基因流(Nm:上三角)
Table 8 Genetic differentiation of COⅠ genes among different populations (Fst: lower triangle); Gene flow (Nm: upper triangle)
Population 儋州Danzhou 海口Haikou 琼海Qionhai 三亚Sanya 儋州Danzhou 83.08 12.25 3.60 海口Haikou 0.003 62.25 5.18 琼海Qionghai 0.020 0.004** 5.85 三亚Sanya 0.065* 0.046* 0.041** 注:−,表示无穷大;*,表示p≤0.05;**,表示p≤0.01。 Note: −, denotes infinity; *, p≤0.05; **, p≤0.01.
下载: 导出CSV
表 9 白纹伊蚊不同地理种群之间的遗传分化(Fst;下三角)和基因流(Nm;上三角)
Table 9 Genetic differentiation (Fst; lower triangle) and gene flow (Nm; upper triangle) between different geographical populations of Ae.albopictus
Country/Region HN FJ GD YN USA IT LO JP VN ML SK CN HN 0.27 0.45 1.37 0.59 0.32 1.75 0.32 2.15 1.26 0.36 FJ 0.65** 2.60 0.36 1.49 0.41 0.49 0.87 1.52 2.86 0.90 GD 0.52** 0.16* 1.22 11.83 5.21 0.89 5.98 3.37 3.11 2.67 YN 0.27** 0.58** 0.29** 1.63 0.60 6.32 0.46 73.05 3.23 0.78 USA 0.46** 0.25** 0.04 0.23** 4.55 1.28 3.98 4.06 1.96 2.77 IT 0.61** 0.55** 0.09** 0.46** 0.10** 0.70 2.18 2.50 2.91 2.12 LO 0.22** 0.51** 0.36** 0.02 0.28** 0.42** 0.64 9.88 1.81 0.67 JP 0.61** 0.36** 0.08 0.52** 0.11** 0.19** 0.44** 2.33 2.13 1.73 VN 0.19** 0.25** 0.13** 0.01 0.11** 0.17** 0.05** 0.18** 1.63 1.61 ML 0.28** 0.15* 0.14* 0.13 0.20** 0.15* 0.22** 0.19** 0.23** 2.00 SK 0.58** 0.36** 0.16** 0.39** 0.15** 0.19** 0.43** 0.22** 0.24** 0.20** 注:HN:中国海南(101个样本,Genbank登录号:PQ858251-PQ858367);FJ:中国福建(8个样本,KY971602.1-KY971596.1;KY971591.1);GD:中国广东(9个样本,KX886287.1-KX886295.1);YN:中国云南(9个样本,KY765501.1;KY765474.1-KY765467.1);USA:美国(66个样本,KC690896.1-KC690961.1);IT:意大利(9个样本,JX679373.1-JX679380.1);LO:老挝(46个样本,MN080765.1-MN080720.1);JP:日本(21个样本,AB738310.1;AB738308.1;AB738294.1;AB738241.1;AB738240.1;AB738211.1;AB738203.1;AB738197.1;AB738192.1;AB738179.1;AB738161.1;AB738143.1;AB738122.1-AB738121.1;AB738090.1;LC544010.1;LC544012.1;LC544014.1;LC544013.1;LC544009.1;LC544011.1);VN:越南(65个样本,MZ573376.1-MZ573312);ML:马来西亚(90个样本,MF148303.1-MF148249.1;KY982366.1-KY982334.1);SK:韩国(24个样本,MG871379.1-MG871402.1)。*p≤0.05,**p≤0.01。
下载: 导出CSV
-
[1] KHOSHDEL-NEZAMIHA F, VATANDOOST H, OSHAGHI M A, et al. Molecular characterization of mosquitoes (Diptera: Culicidae) in northwestern Iran by using rDNA-ITS2[J]. Japanese Journal of Infectious Diseases, 2016, 69(4): 319 − 322. doi: 10.1023/A:1026530522612 [2] AJAMMA Y U, VILLINGER J, OMONDI D, et al. Composition and genetic diversity of mosquitoes (Diptera: Culicidae) on islands and mainland shores of Kenya's Lakes Victoria and Baringo[J]. Journal of Medical Entomology, 2016, 53(6): 1348 − 1363. doi: 10.1093/jme/tjw102 [3] 郑雅匀, 王志玉, 梁国栋. 海南省虫媒病毒调查研究状况[J]. 中国媒介生物学及控制杂志, 2011, 22(6): 607 − 610. [4] 游志勇, 王逸民, 赵子江, 等. 海南岛两个虫媒病毒分离物的初步鉴定[J]. 病毒学报, 1988, 4(1): 16 − 22. [5] 周俊梅, 江丽芳, 高阳, 等. 登革2型病毒海南分离株全长E基因的测定与分析[J]. 中华微生物学和免疫学杂志, 2003, 23(4): 304 − 306. doi: 10.3760/j:issn:0254-5101.2003.04.021 [6] ZHAI Y G, WANG H Y, SUN X H, et al. Complete sequence characterization of isolates of Getah virus (genus Alphavirus, family Togaviridae) from China[J]. Journal of General Virology, 2008, 89(6): 1446 − 1456. doi: 10.1099/vir.0.83607-0 [7] 董必军, 陈文州, 李秀维, 等. 首次从海南岛蚊虫和蝙蝠中分离出两株基孔肯雅病毒[J]. 中国媒介生物学及控制杂志, 1993, 4(3): 205 − 208. [8] 赵春生, 蒋廉华, 余兴龙, 等. 从海南省蝙蝠脑中分离出1株罗斯河病毒及其血清抗体调查[J]. 中国兽医学报, 1997, 17(3): 241 − 243. doi: 10.16303/j.cnki.1005-4545.1997.03.010 [9] 游志勇, 王逸民, 赵治国, 等. 海南省两株环状病毒的分离与鉴定[J]. 病毒学报, 1990, 6(3): 272 − 276. [10] LIU H, GAO X Y, LIANG G D. Newly recognized mosquito-associated viruses in mainland China, in the last two decades[J]. Virology Journal, 2011, 8(1): 68. doi: 10.1186/1743-422X-8-68 [11] 何昌华, 赵伟, 王善青, 等. 海南省城区2012年蚊虫密度及季节消长分析[J]. 中国媒介生物学及控制杂志, 2014, 25(1): 15 − 17. doi: 10.11853/j.issn.1003.4692.2014.01.004 [12] 王晓花, 杨新艳, 赵伟, 等. 海口市2012-2014年蚊虫密度监测结果分析[J]. 中国媒介生物学及控制杂志, 2015, 26(4): 424 − 426. doi: 10.11853/j.issn.1003.4692.2015.04.027 [13] 陈学文, 林怡, 林春燕, 等. 海口市2017年病媒生物密度监测[J]. 中国热带医学, 2018, 18(8): 795 − 798. doi: 10.13604/j.cnki.46-1064/r.2018.08.12 [14] 范娜, 孙定炜, 程睿, 等. 海南省2017—2018年虫媒病毒的分离与鉴定[J]. 中华流行病学杂志, 2020, 41(2): 236 − 243. doi: 10.3760/cma.j.issn.0254-6450.2020.02.018 [15] LI S P, JIANG F, LU H, et al. Mosquito diversity and population genetic structure of six mosquito species from Hainan island[J]. Frontiers in Genetics, 2020, 11: 602863. doi: 10.3389/fgene.2020.602863 [16] 胡坤敏, 陈韶红, 艾琳, 等. 豫皖闽浙4省溪蟹并殖吸虫囊蚴核糖体ITS2和线粒体CO1基因序列分析[J]. 中国寄生虫学与寄生虫病杂志, 2020, 38(1): 87 − 94. doi: 10.12140/j.issn.1000-7423.2020.01.013 [17] 赵健, 胡劭骥, 和秋菊, 等. 基于线粒体COⅠ和ND1基因分析三尾凤蝶遗传多样性[J]. 福建农林大学学报(自然科学版), 2017, 46(4): 387 − 391. doi: 10.13323/j.cnki.j.fafu(nat.sci.).2017.04.004 [18] 周慧, 李迪强, 张于光, 等. 藏羚羊mtDNAD-loop区遗传多样性研究[J]. 遗传, 2006, 28: 299 − 305. doi: 10.16288/j.yczz.2006.03.009 [19] 陆宝麟. 中国动物志, 昆虫纲, 双翅目, 蚊科(上卷)第8卷[M]. 北京: 科学出版社, 1997: 1−904. [20] 陆宝麟. 中国动物志, 昆虫纲, 双翅目, 蚊科(下卷)第9卷[M]. 北京: 科学出版社, 1997: 1−190. [21] 陆宝麟, 吴厚永. 中国重要医学昆虫分类与鉴别[M]. 郑州: 河南科学技术出版社, 2003: 1-800. [22] MUTEBI J P, BARRETT A D T. The epidemiology of yellow fever in Africa[J]. Microbes and Infection, 2002, 4(14): 1459 − 1468. doi: 10.1016/s1286-4579(02)00028-x [23] DICK G W A, KITCHEN S F, HADDOW A J. Zika virus (I). Isolations and serological specificity[J]. Transactions of the Royal Society of Tropical Medicine and Hygiene, 1952, 46(2): 509 − 520. doi: 10.1016/0035-9203(52)90042-4 [24] 郑梓豪, 吴珊珊, 魏勇, 等. 基于线粒体COⅠ基因分析广州市15个白纹伊蚊种群的遗传多样性[J]. 中国人兽共患病学报, 2021, 37(11): 985 − 994. doi: 10.3969/j.issn.1002-2694.2021.00.141 [25] FOLMER O F, BLACK M, HOEH W R, et al. DNA primers for amplification of mitochondrial cytochrome c oxidase subunit I from diverse metazoan invertebrates[J]. Molecular Marine Biology and Biotechnology, 1994, 3(5): 294 − 299. [26] 郭颂, 凌锋, 王金娜, 等. 浙江省不同地理株白纹伊蚊mtDNA-COⅠ基因多态性研究[J]. 中国人兽共患病学报, 2016, 32(2): 133 − 136. doi: 10.3969/j.issn.1002-2694.2016.02.007 [27] TORRES-GUTIERREZ C, BERGO E S, EMERSON K J, et al. Mitochondrial COⅠ gene as a tool in the taxonomy of mosquitoes Culex subgenus Melanoconion[J]. Acta Tropica, 2016, 164: 137 − 149. doi: 10.1016/j.actatropica.2016.09.007 [28] WANG G, LI C X, GUO X X, et al. Identifying the main mosquito species in China based on DNA barcoding[J]. PLoS One, 2012, 7(10): e47051. doi: 10.1371/journal.pone.0047051 [29] 李大命, 张彤晴, 唐晟凯, 等. 基于线粒体COⅠ序列的洪泽湖河蚬(Corbicula fluminea)遗传多样性和种群结构分析[J]. 海洋与湖沼, 2015, 46(6): 1339 − 1346. doi: 10.11693/hyhz20141200353 [30] 孙鹏, 尹飞, 施兆鸿, 等. 基于COⅠ部分序列探讨南海鲳属鱼类系统进化与种群遗传结构[J]. 海洋科学, 2012, 36(6): 15 − 21. [31] 王焓谕. 六种天牛线粒体全基因组测定和分析[D]. 西安: 陕西师范大学, 2018. [32] 郑宇婷, 姜进勇, 周红宁, 等. 利用ssr标记浅析云南登革热重点地区埃及伊蚊种群遗传特征[J]. 中国人兽共患病学报, 2021, 37(2): 176 − 182. doi: 10.3969/j.issn.1002-2694.2021.00.014 [33] FANG Y, ZHANG J, WU R, et al. Genetic polymorphism study on Aedes albopictus of different geographical regions based on DNA barcoding[J]. BioMed Research International, 2018, 2018: 1501430. doi: 10.1155/2018/1501430 [34] 刘雪莹, 韩玉立, 司静, 等. 基于ITS序列的灵芝遗传多样性与群体遗传分化研究[J]. 菌物学报, 2018, 37(5): 565 − 575. doi: 10.13346/j.mycosystema.180019 [35] 李大命, 唐晟凯, 刘燕山, 等. 基于Cytb基因的江苏省大银鱼种群遗传多样性和遗传结构分析[J]. 上海海洋大学学报, 2021, 30(3): 416 − 425. doi: 10.12024/jsou.20190702720 [36] 张瑞玲, 姚广琴, 潘晓倩, 等. 不同地理种群白纹伊蚊线粒体基因COⅠ的遗传多样性分析[J]. 中国人兽共患病学报, 2017, 33(4): 316 − 320. doi: 10.3969/j.issn.1002-2694.2017.04.005 -

 点击查看大图
点击查看大图
计量
- 文章访问数: 1209
- HTML全文浏览量: 518
- PDF下载量: 16
- 被引次数: 0
