

 下载:
下载:
-
巴西橡胶树(Hevea brasiliensis)是天然橡胶最主要的商业来源。天然橡胶是在橡胶树乳管细胞中橡胶粒子上合成的一种重要次生代谢产物,以甲羟戊酸(mevalonic acid,MVA)途径或丙酮酸/磷酸甘油醛(methylerythritol phosphate,MEP)途径生成的异戊烯基焦磷酸(IPP)为前体,经过一系列的酶促反应,形成以顺−1,4−聚异戊二烯为主要成分的天然高分子化合物[1]。早期对天然橡胶生物合成的研究主要集中在对合成途径中酶的特性分析、基因克隆、表达和功能分析[2]。Tang等[3]在橡胶树基因组中鉴定了20个基因家族共89个橡胶生物合成相关基因。转录因子是调节次生代谢产物的生物合成途径的重要基因[4],一些调控橡胶生物合成途径的转录因子也被相继鉴定[5-14],但对橡胶生物合成的调控机制仍缺乏深入了解。
bZIP转录因子家族是植物中规模最大、种类最丰富的转录因子家族之一。bZIP转录因子具有60~80个氨基酸组成的高度保守的bZIP结构域,由1个位于结构域N端保守的碱性区域,和1个位于C端的亮氨酸拉链区域组成[15]。碱性区域由16~20个氨基酸残基组成,包含1个核定位信号,和1个固定的N-x7-R/K基序,可以识别并特异结合启动子中的特定序列[16]。亮氨酸拉链区域由1个或多个7肽重复单元,每个单元的末位为亮氨酸或疏水氨基酸,该区域参与bZIP转录因子同源或异源二聚体的形成[17-18]。植物bZIP蛋白主要结合ACGT为核心序列的顺式作用元件,如ABRE、G-box、C-box和A-box等[19],调节下游基因表达,在植物生长发育、逆境胁迫应答及次生代谢产物生物合成过程中发挥重要作用。目前,在拟南芥(Arabidopsis thaliana)[16]、大豆(Glycine max)[20]、番茄(Solanum lycopersicum)[21]、烟草(Nicotiana tabacum)[22]、棉花(Gossypium hirsutum)[23]、木薯(Manihot esculenta)[24]等许多植物鉴定到bZIP家族成员。研究表明,bZIP转录因子广泛参与植物次生代谢产物生物合成的调控。青蒿AabZIP1激活下游ADS和CYP71AV1的表达,调控青蒿素的生物合成[25]。短角蒲公英TbbZIP1通过促进TbSRPP的表达,调节天然橡胶生物合成[26]。喜树(Camptotheca acuminata)bZIP转录因子CaLMF负调控喜树碱合成相关基因CaTDC1、CaG8O、CaCYC1和Ca7DLS表达,抑制喜树碱的生物合成[27]。橡胶树bZIP转录因子HbTGA1调控HbHMGS2、HbHMGR2、HbCPT6、HbCPT8和HbSRPP2等多个橡胶生物合成基因转录,参与天然橡胶生物合成调控[28]。
本研究前期在橡胶树胶乳cDNA文库中筛选到与法尼基焦磷酸合酶、橡胶转移酶、小橡胶粒子蛋白和橡胶延长因子等橡胶生物合成相关酶和蛋白基因启动子结合的转录因子HbbZIP74,推测其可能参与天然橡胶生物合成调控。本研究通过PCR克隆了HbbZIP74基因,并进行生物信息学分析,构建pET28-HbbZIP74原核表达载体,在大肠杆菌中异源表达,优化诱导表达条件并成功纯化重组蛋白,旨在为下一步研究HbbZIP74在天然橡胶生物合成中的功能奠定基础。
-
实验材料为7年生未开割巴西橡胶树无性系‘热研7-33-97’组培苗植株,种植于中国热带农业科学院试验场。茉莉酸、脱落酸、乙烯和水杨酸处理参照Guo等[9]的方法。橡胶树根、树皮、叶和花等组织同时采样后立即用液氮研磨或保存在−80 ℃。胶乳用液氮速冻后收集到离心管,再放入液氮中,用于RNA的提取。大肠杆菌(Escherichia coli)DH5α和BL21(DE3)菌株分别购买于天根生物科技有限公司(北京)和上海唯地生物技术有限公司,蛋白表达载体pET28a由实验室保存,DNA Marker、pMD19T载体、高保真酶PrimeSTAR和T4 DNA连接酶购自Takara公司(大连),质粒提取试剂盒、RNA抽提试剂盒、DNA凝胶回收试剂盒来自FOREGENE公司(成都),反转录试剂盒和荧光定量试剂来自NOVA公司(苏州)。引物合成和测序由中美泰和生物技术有限公司完成。
-
根据HbbZIP74基因(基因编号:LOC110658634)在 NCBI中橡胶树参考序列XM_021816336.2,使用SnapGene软件设计基因特异性扩增引物(正向引物SPF: 5′-ATGGCATCCTCCAGTGGAAA-3′,反向引物SPR: 5′-ATACCGAAAGAAATCTGGAG-3′)。以橡胶树胶乳cDNA 为模板扩增目的序列,PCR扩增体系:2 μL模板、上下游引物各1 μL、25 μL 2×PrimeSTAR HS(Premix)、ddH2O 22 μL。PCR反应程序为:95 ℃预变性5 min;95 ℃变性30 s,58 ℃退火40 s,72 ℃延伸40 s,38个循环;72 ℃延伸10 min。PCR产物经挖胶回收纯化后克隆到pMD19T载体中,pMD19T-HbbZIP74经PCR(正向引物M13-F: 5′-TGTAAAACGACGGCCAGT-3′,反向引物M13-R: 5′-CAGGAAACAGCTATGACC-3′)扩增筛选阳性克隆,将阳性克隆抽提质粒后测序。
-
在NCBI网站查找HbbZIP74基因编码的氨基酸序列,利用ExPASy-ProtParam tool(
http://web.expasy.org/protparam/ )在线软件分析HbbZIP74蛋白质分子质量、等电点、稳定性和平均亲水系数;采用SMART在线软件(http://smart.embl-heidelberg.de/ )预测蛋白保守结构域;采用Plant-mPLoc在线工具(https://www.ncbiwww.csbio.sjtu.edu.cn/cgi-bin/PlantmPLoc.cgi )进行蛋白亚细胞定位预测;分别采用SOPMA(https://npsa.lyon.inserm.fr/cgi-bin/npsa_automat.pl?page=/NPSA/npsa_sopma.html )和SWISS-MODEL(https://swissmodel.expasy.org/ )分析HbbZIP74蛋白的二级结构和三级结构。利用Clustal在线网站(https://www.genome.jp/tools-bin/clustal )进行多序列比对,再通过ESPript网站(https://espript.ibcp.fr/ESPript/cgi-bin/ESPript.cgi )将序列比对结果可视化;在拟南芥中bZIP的各基因亚家族各选取1~2个bZIP转录因子,将HbbZIP74蛋白序列通过Clustal进行多序列比对,并用MEGA构建进化树。 -
RNA的提取采用FOREGENE公司(成都)的植物总RNA提取试剂盒(多糖多酚含量高的样本),并参照说明书操作。cDNA第一链的合成使用NOVA公司(苏州)的去基因组与逆转录一管化三代预混液试剂盒,按照说明书进行操作。
-
采用NOVA公司(苏州)的Taq SYBR® Green qPCR Premix(Universal)试剂盒,使用BIO-RADCFX96 Real-time PCR系统(Bio-Rad,USA),进行qRT-PCR分析。HbbZIP74的引物为:qbZIP74-F: 5′-GGGCACAGATGATGGAACTTA-3′;qbZIP74-R: 5′-CAGCAGCAACAGCACTTATTT-3′。采用Actin为内参基因,引物为:Actin-F: 5′-CAGTGGTCGTACAACTGGTAT-3′;Actin-R: 5′-ATCCTCCAATCCAGACACTGT-3′。qRT-PCR的反应体系(20 μL):1 μL模板、正反向引物(10 μmol·L−1)各1 μL、10 μL Taq SYBR® Green qPCR Premix、ddH2O 7 μL。qRT-PCR反应程序为:95 ℃预变性30 s;95 ℃变性10 s,58 ℃退火10 s,72 ℃延伸30 s,40个循环。每个样品进行3次生物学重复和3次技术重复实验,结果采用2−ΔΔCt法分析基因的相对表达量,使用SPSS分析样本间的差异显著性。
-
根据pET28a载体图谱和HbbZIP74的CDS序列设计正向引物F: 5′-TCGCGGATCCGAATTCATGGCATCCTCCAGTGGAAAT-3′,反向引物R: 5′-GGTGGTGGTGCTCGAGATACCGAAAGAAATCTGGAGAAGCC-3′,以构建成功的pMD19T-HbbZIP74的质粒为模板,通过PCR扩增目的片段并挖胶回收纯化。使用同源重组的方法将扩增产物片段连接到已经酶切完成的pET28a载体上,转化后大肠杆菌DH5α,经过PCR验证(正向引物T7-F: 5′-TAATACGACTCACTATAGGG-3′,反向引物T7-R: 5′-GCTAGTTATTGCTCAGCGG-3′),挑选具有单一明亮条带的菌落进行质粒提取后,将测序验证的重组表达载体命名为pET28-HbbZIP74。
-
将pET28-HbbZIP74质粒转入BL21(DE3)菌株中,挑取单菌落接种于含有50 mg·L−1卡那霉素的LB液体培养基中,37 ℃,200 r·min−1,培养16 h。按菌体体积1︰100的比例将菌液接种于含有50 mg·L−1 卡那霉素的5 mL LB液体培养基,在37 ℃,180 r·min−1培养至OD600为0.4~0.6,IPTG的终浓度分别为0.1、0.3、0.5、1.0和2.0 mmol·L−1,然后37 ℃振荡培养5 h;在1 mmol·L−1 IPTG浓度下,分别16、20、28和37 ℃振荡培养5 h;在1 mmol·L−1 IPTG、37 ℃条件下,分别诱导1、2、3、4和5 h。对不同条件的诱导样品进行SDS-PAGE电泳,确定最佳诱导表达条件。
-
根据上述摸索的最佳诱导表达条件,将诱导的200 mL菌液8 000 r·min−1离心5 min后,用PBS缓冲液重悬后离心,再用30 mL洗涤液(体积分数为1%的 TritonX-100,500 mmol·L−1 NaCl,2 mol·L−1脲,50 mmol·L−1 Tris-HCl pH7.5,1 mmol·L−1 EDTA)重悬,加入终浓度(质量浓度)为1.5 g·L−1的溶菌酶,在冰上摇过夜。冰上超声破碎细胞,4 ℃,9 000 r·min−1离心15 min,用洗涤液重悬,重复2次。再用裂解液(50 mmol·L−1 Tris-HCl pH8.5,8 mol·L−1脲,1 mmol·L−1 DTT)按称重溶解为10~20 g·L−1重悬,静置30 min至沉淀溶解。用0.45 μm水系滤膜过滤裂解液后上镍柱进行纯化,依次用洗脱液(8 mol·L−1脲,50 mmol·L−1 Tris-HCl pH8.5,1 mmol·L−1 DTT),10 mmol·L−1咪唑洗脱液,50 mmol·L−1咪唑洗脱液,100 mmol·L−1咪唑洗脱液进行洗脱后,对收集的样品进行SDS-PAGE电泳。洗脱液装入透析袋,置于复性液(50 mmol·L−1 Tris-HCl,1 mmol·L−1 EDTA,2.2 mmol·L−1 GSH,1.1 mmol·L−1 GSSG,体积分数5% 甘油,pH8.5)中,4 ℃静置透析过夜。再将透析袋放入新的透析液(50 mmol·L−1 Tris-HCl,1 mmol·L−1 EDTA,pH8.5)中透析,换液3次。透析完后收集透析袋中液体,并分装存于−20 ℃冰箱。
-
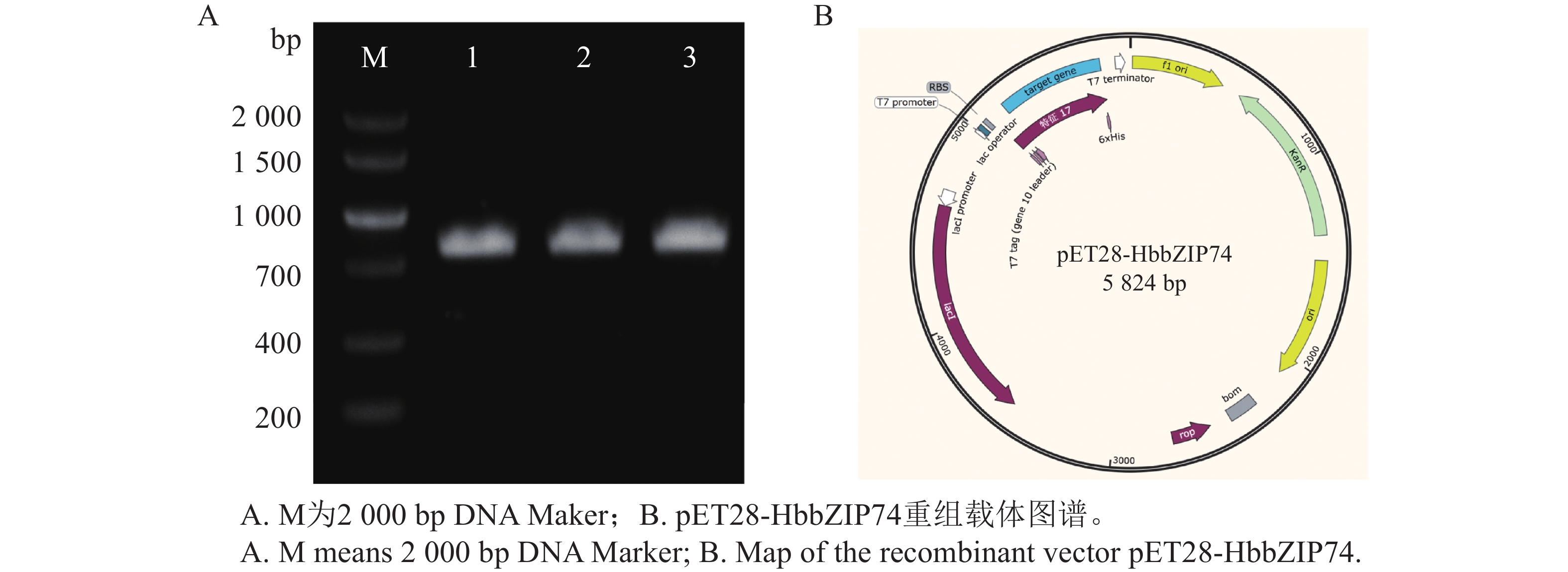
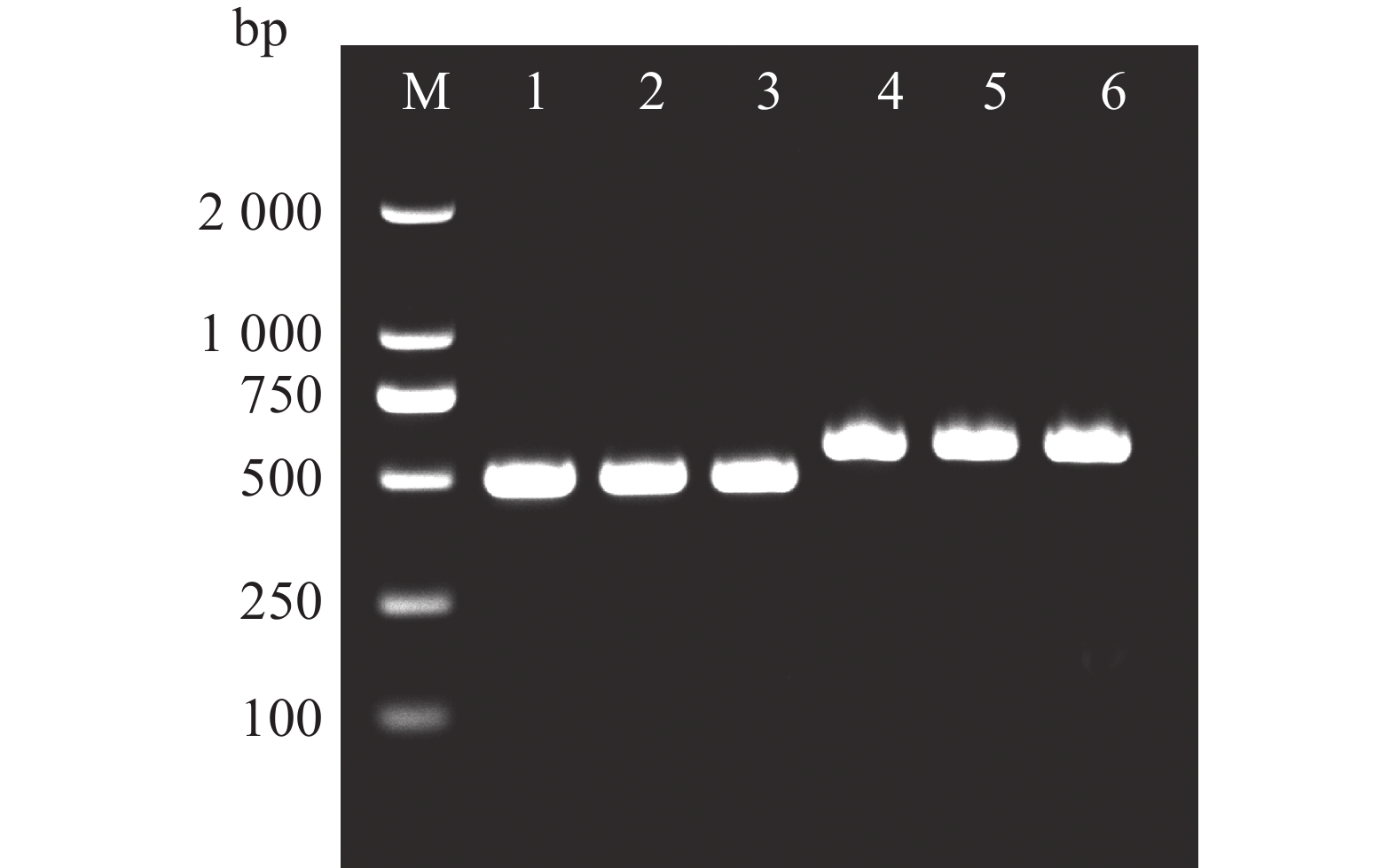
以橡胶树胶乳cDNA为模板,通过特异引物SPF和SPR扩增,得到长度约为500 bp的片段(图1),条带清晰且单一,与预期HbbZIP74 ORF长度相符。将目的片段连接至pMD19T载体后转化到DH5α菌株中,经菌液PCR筛选获得阳性克隆(图1),抽提质粒经测序,确定扩增片段为486 bp,与NCBI数据库中HbbZIP74(登录号:XM_021816336.2)的ORF序列完全一致。重组质粒命名为pMD19T-HbbZIP74。
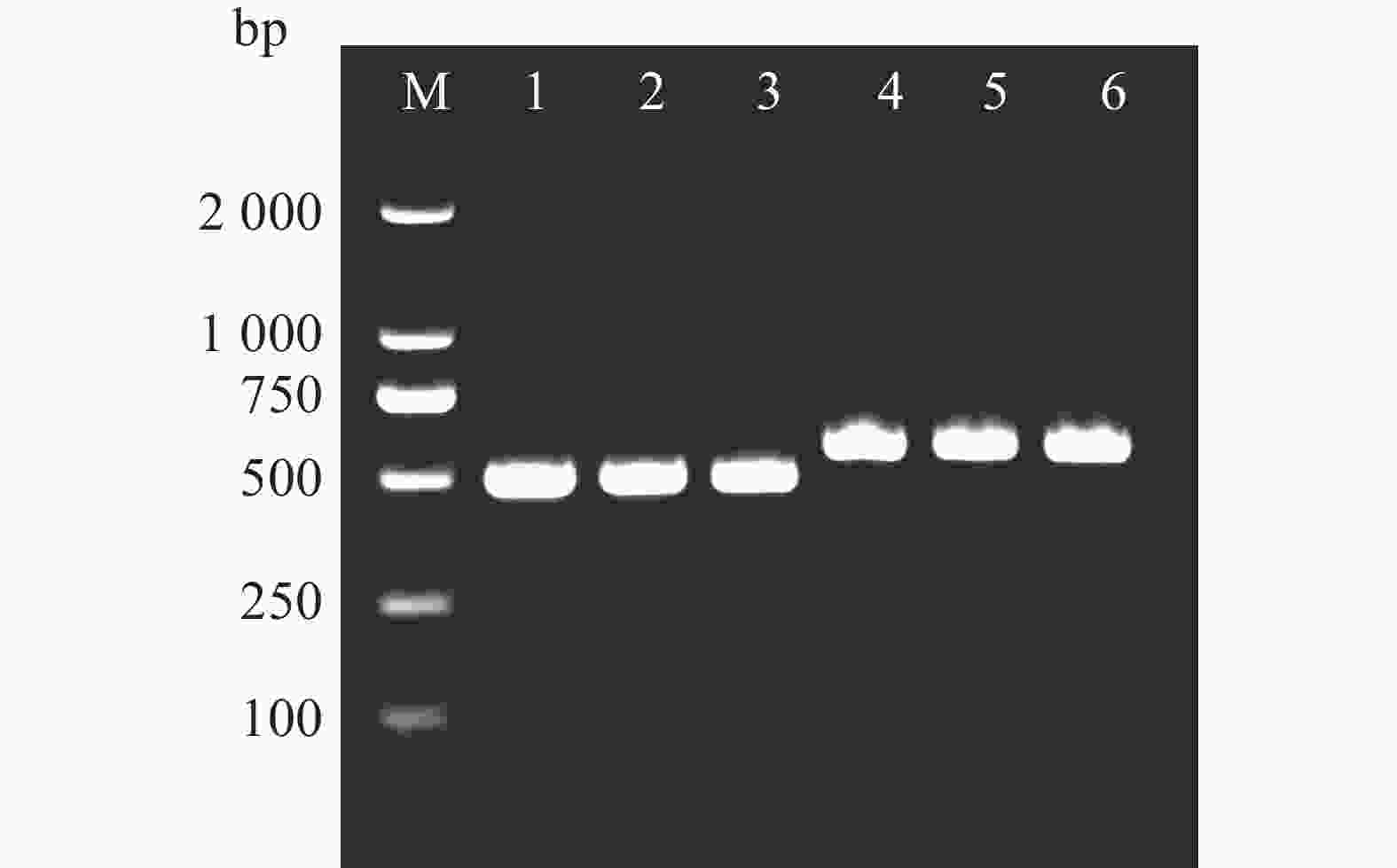
图 1 HbbZIP74全长克隆
Figure 1. Full-length cloning of HbbZIP74
-
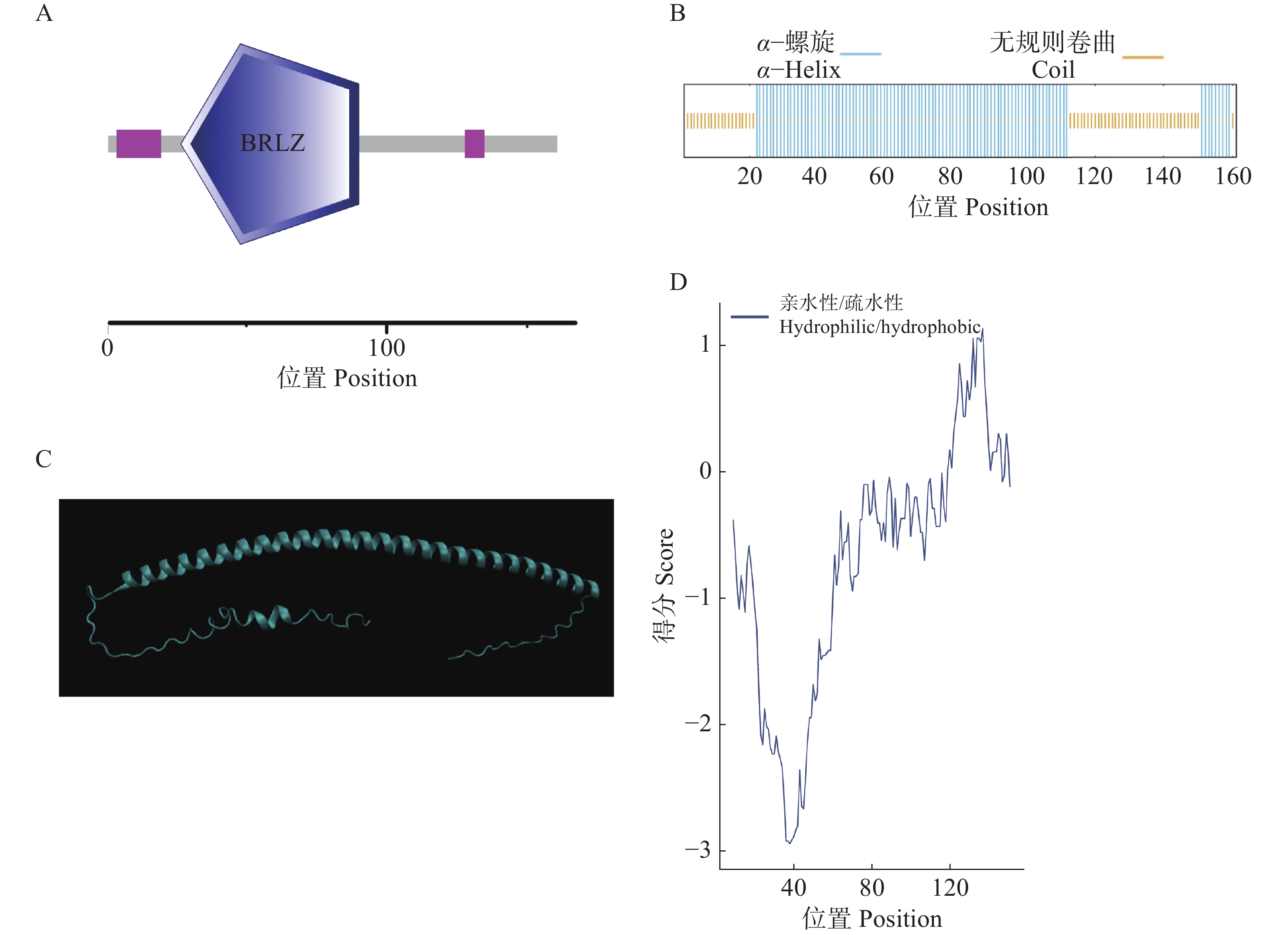
HbbZIP74的ORF全长486 bp,共编码161个氨基酸;相对分子质量为18.28 kDa;理论等电点为6.51;不稳定指数为54.54,平均亲水系数为−0.673,表明该蛋白为不稳定亲水蛋白。该蛋白包含一个碱性亮氨酸拉链结构域,位于第26~90个氨基酸(图2-A)。HbbZIP74蛋白二级结构预测结果显示,100个氨基酸(62.11%)参与形成α−螺旋,61个氨基酸(37.89%)参与形成无规则卷曲(图2-B);三级结构预测结果显示,HbbZIP74蛋白主要由α−螺旋和形成无规则卷曲组成(图2-C);与二级结构预测结果相符,约在40 kDa前为亲水区域,41~130 kDa左右为疏水区域(图2-D),HbbZIP74蛋白亚细胞定位预测位于细胞核。
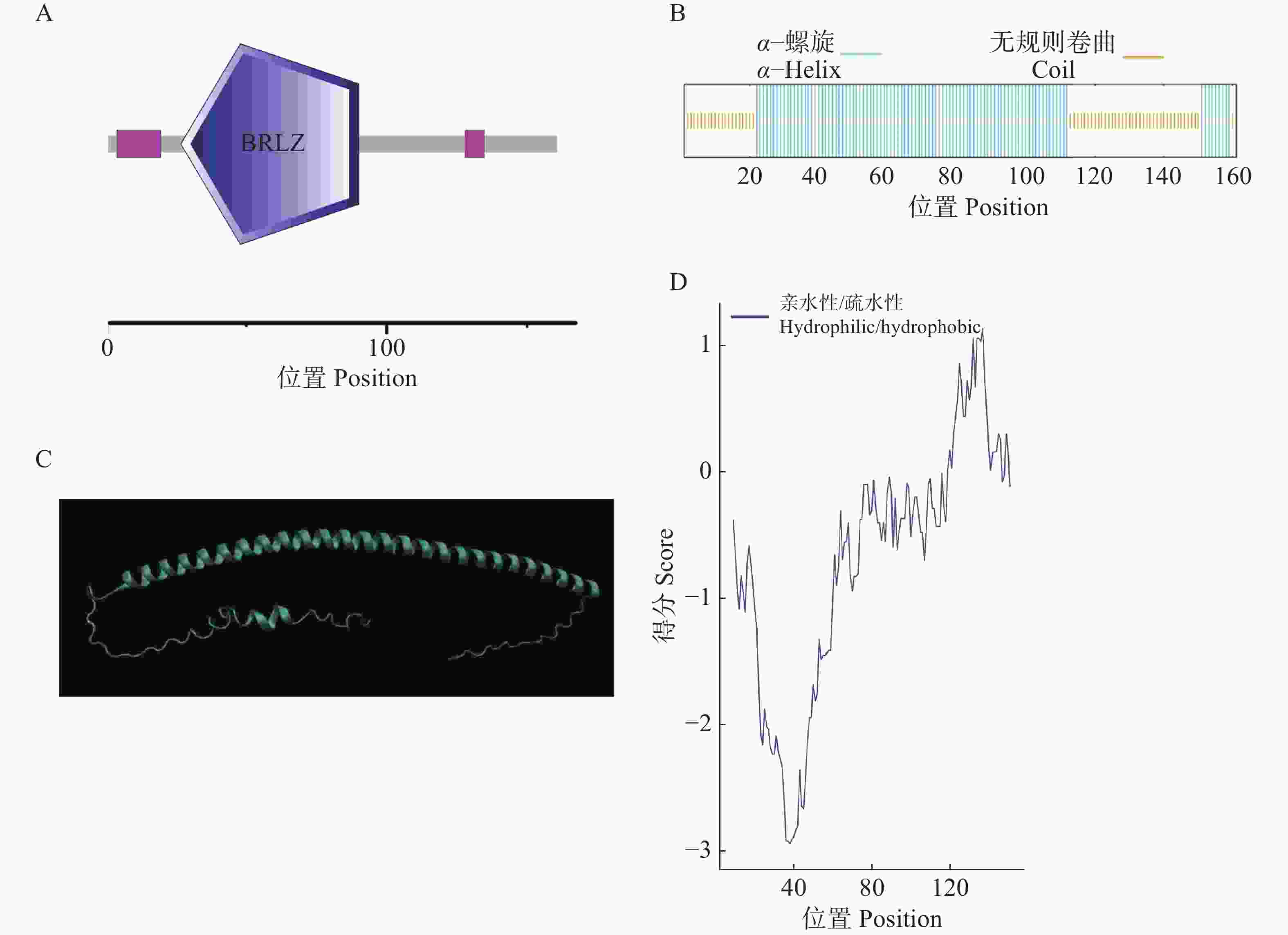
图 2 HbbZIP74蛋白结构分析
Figure 2. Structural analysis of HbbZIP74 protein
-
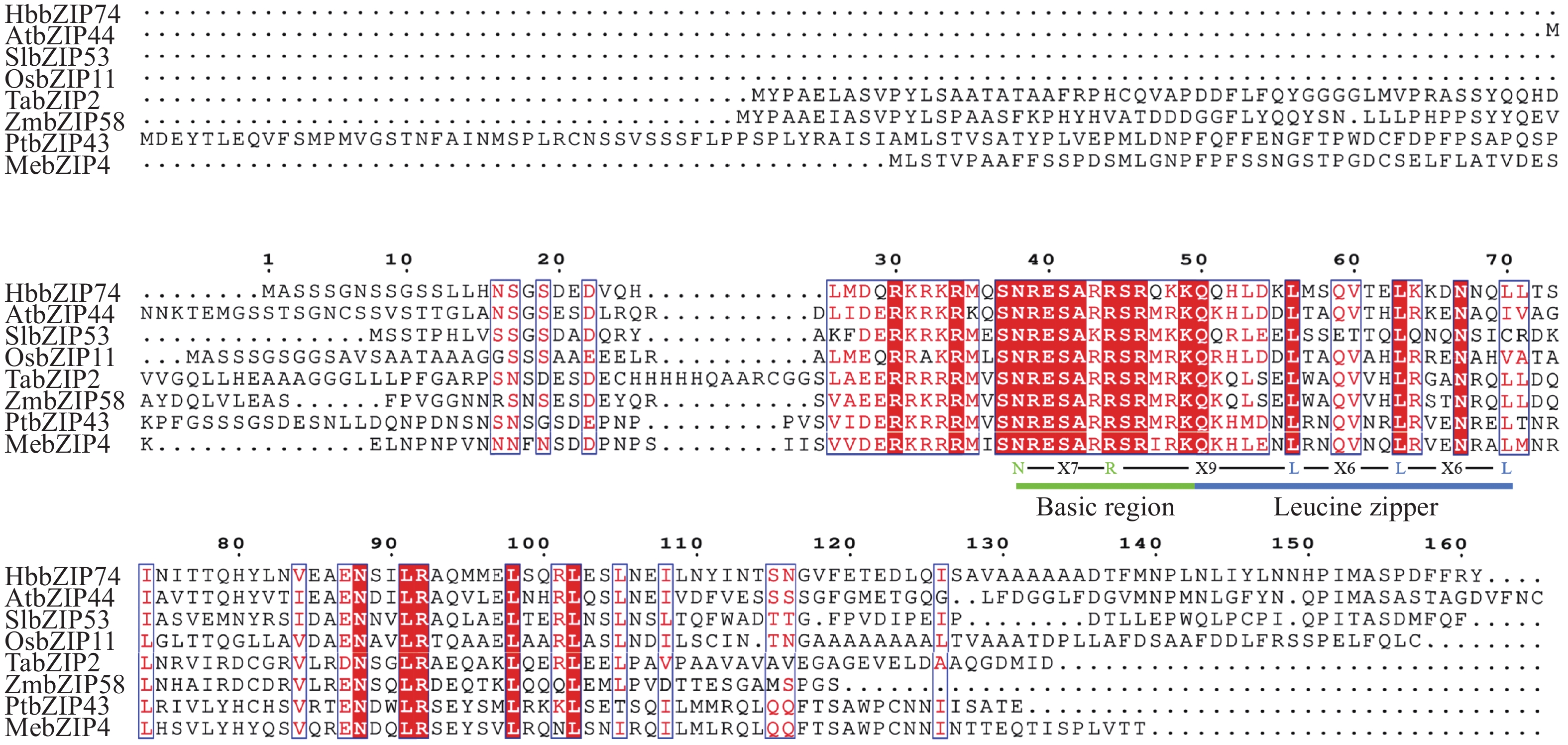
将橡胶树HbbZIP74(XP_021672028.1)氨基酸序列与拟南芥AtbZIP44(NP_177672.2)、小麦TabZIP2(XP_044406621.1)、水稻OsbZIP11(XP_015622690.1)、西红柿SlbZIP53(XP_004240572.1)、玉米ZmbZIp58(AQK74499.1)、毛果杨PtbZIP43(XP_024461072.1)和木薯MebZIP4(XP_021631325.1)的氨基酸序列进行多序列比对分析,氨基酸序列相似性分别为52.53%、34.43%、42.57%、42.22%、36.11%、31.75%和33.33%。比对结果显示,每个蛋白都含有N-x7-R/K基序,以及2~3个不等的x6-L的7肽重复序列(图3),表明HbbZIP74属于bZIP基因家族。
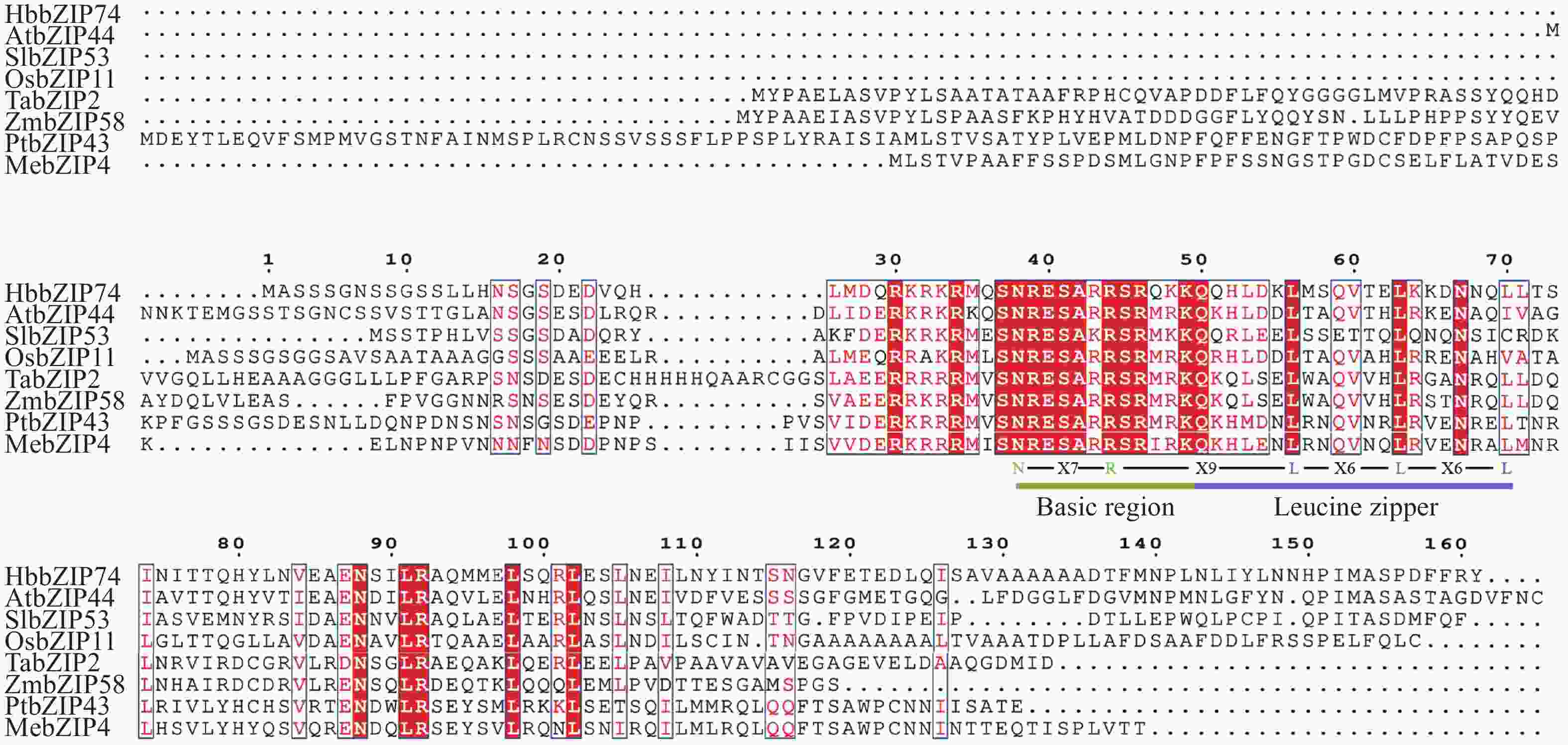
图 3 HbbZIP74与其他物种氨基酸序列同源比对
Figure 3. Homologous alignment of HbbZIP74 amino acid sequences with other species
-
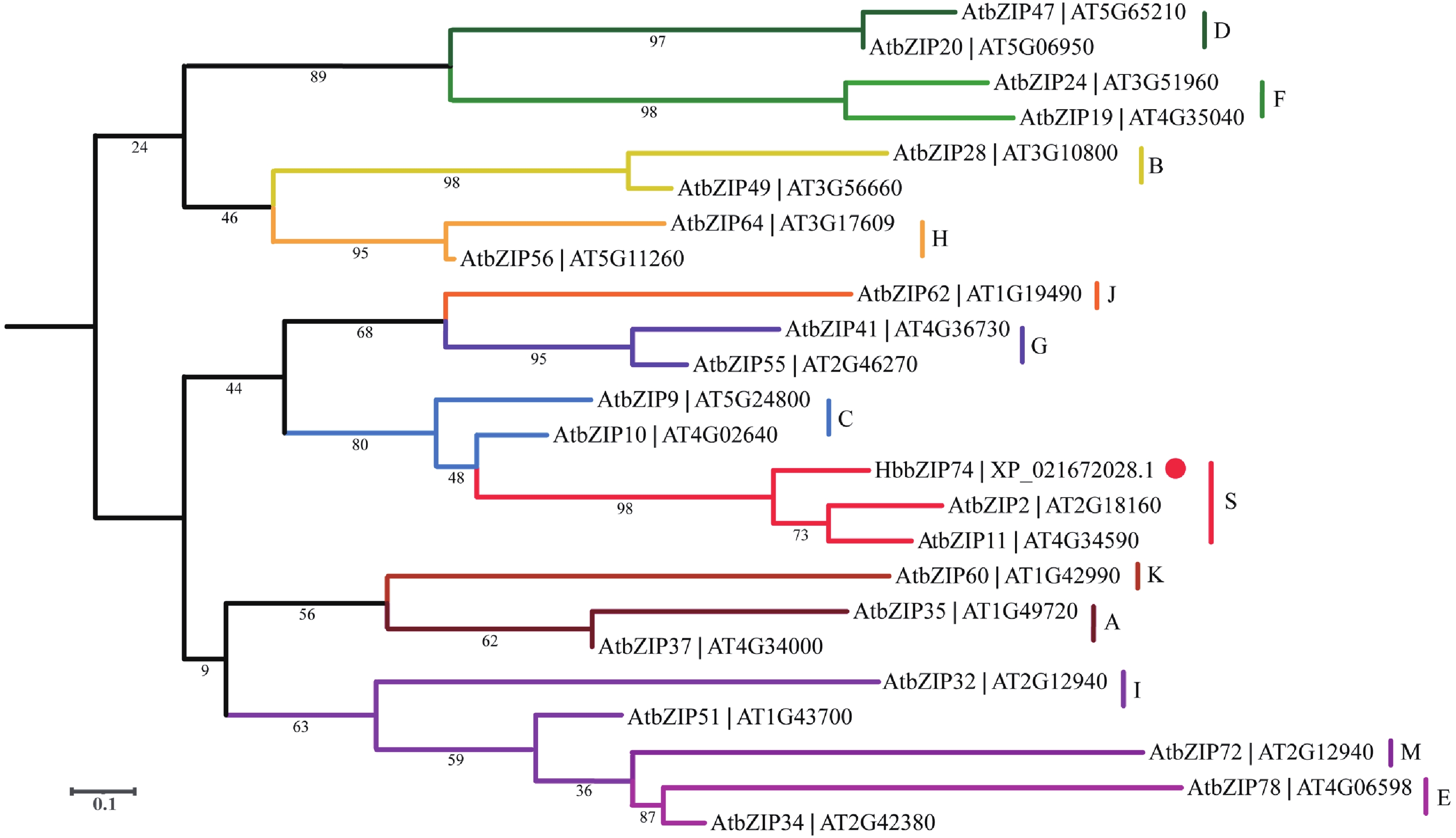
将拟南芥中的多个bZIP氨基酸序列与HbbZIP74进行序列比对,并构建进化树。结果发现,拟南芥bZIP依然被分为A、B、C、D、E、F、G、H、I、J、K、M和S等共13个亚家族,HbbZIP74与AtbZIP2和AtbZIP11处于一个分支,属于S亚家族(图4)。
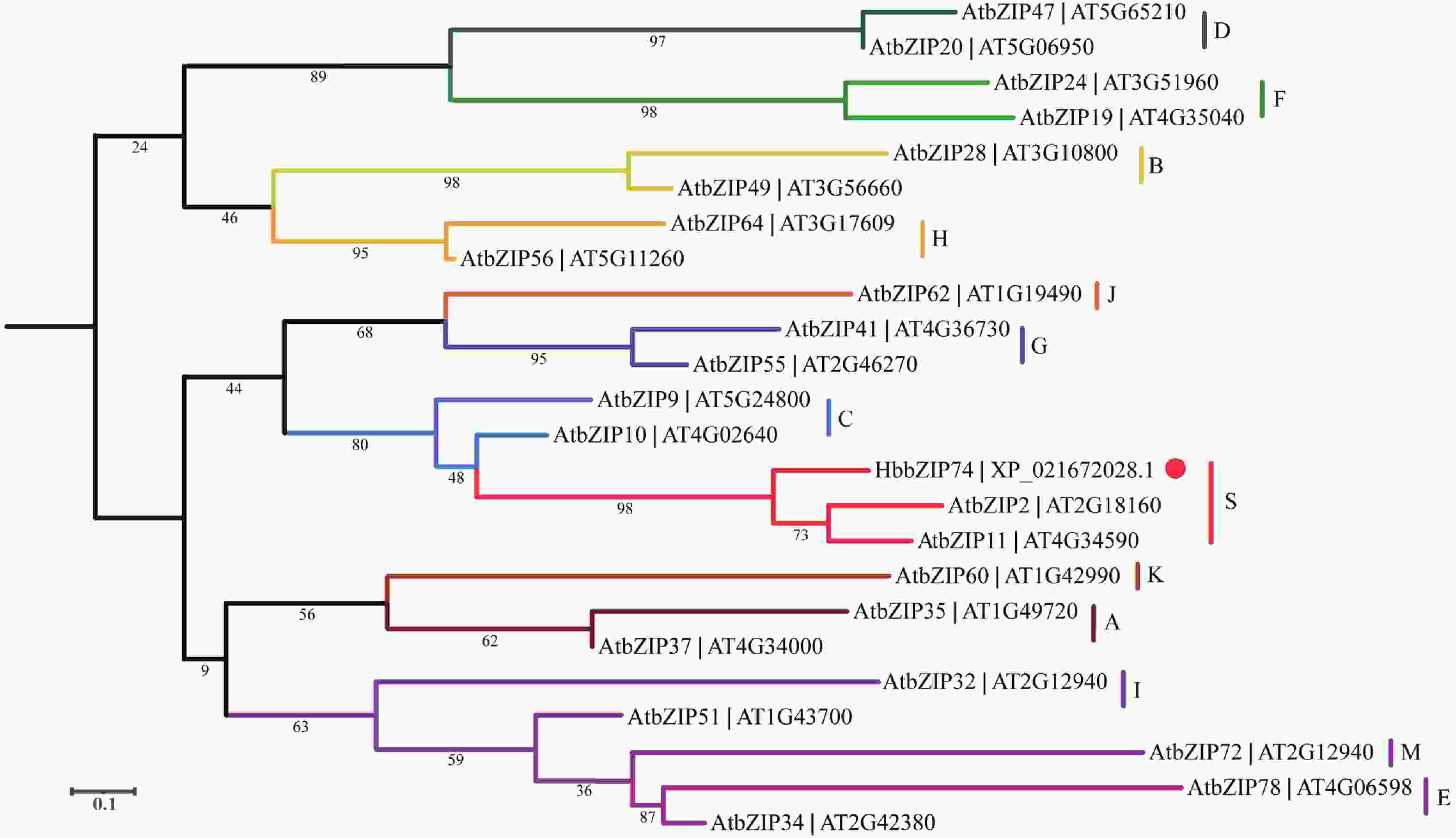
图 4 HbbZIP74系统进化树
Figure 4. Phylogenetic tree of HbbZIP74
-
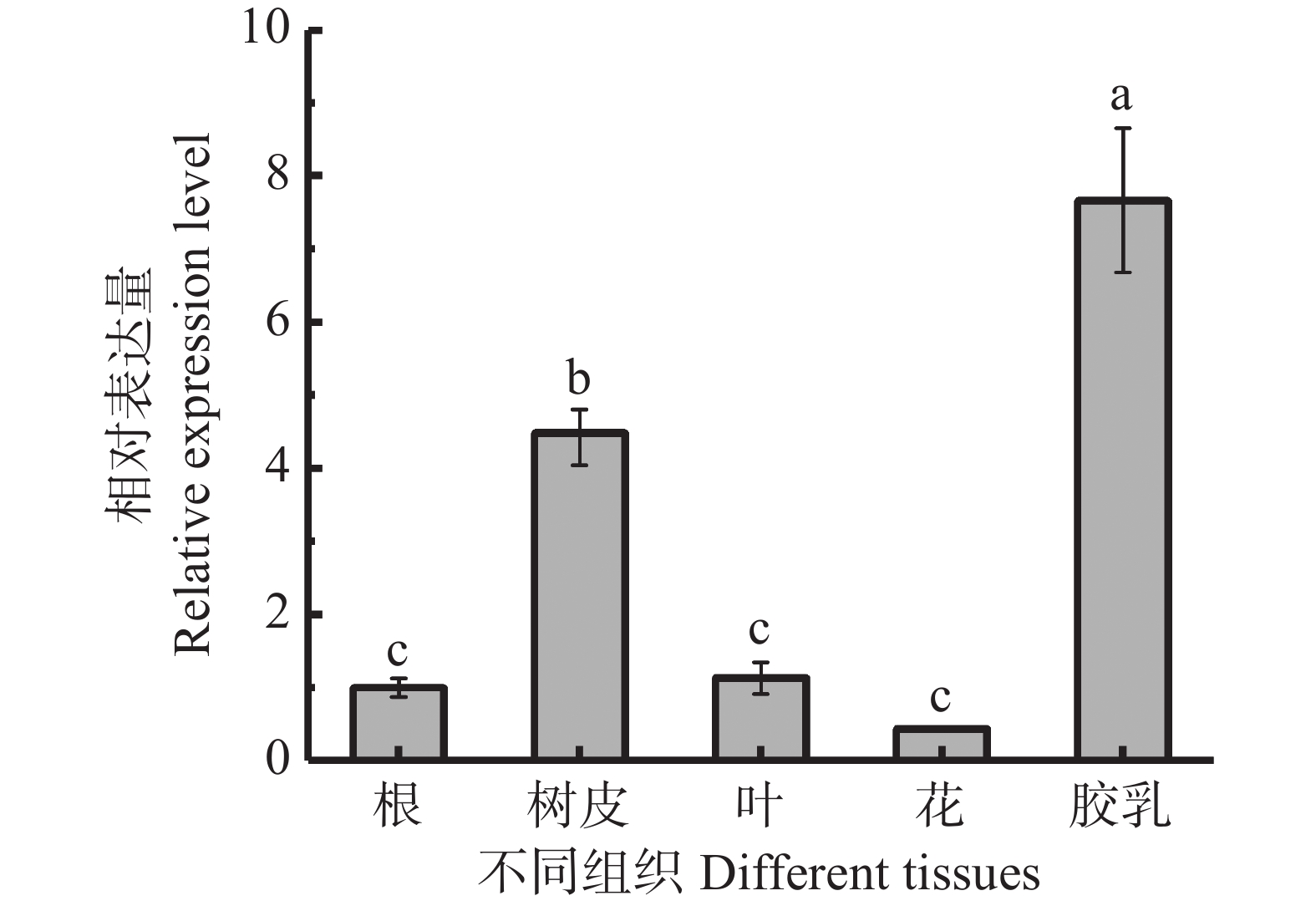
利用qRT-PCR检测了HbbZIP74在橡胶树根、树皮、叶片、花和胶乳等组织的表达情况。从图5可知,HbbZIP74在胶乳中的表达水平最高,树皮中次之,分别为根中表达量的7.66和4.48倍,在叶中的表达量和根相当,在花中的表达量最低。
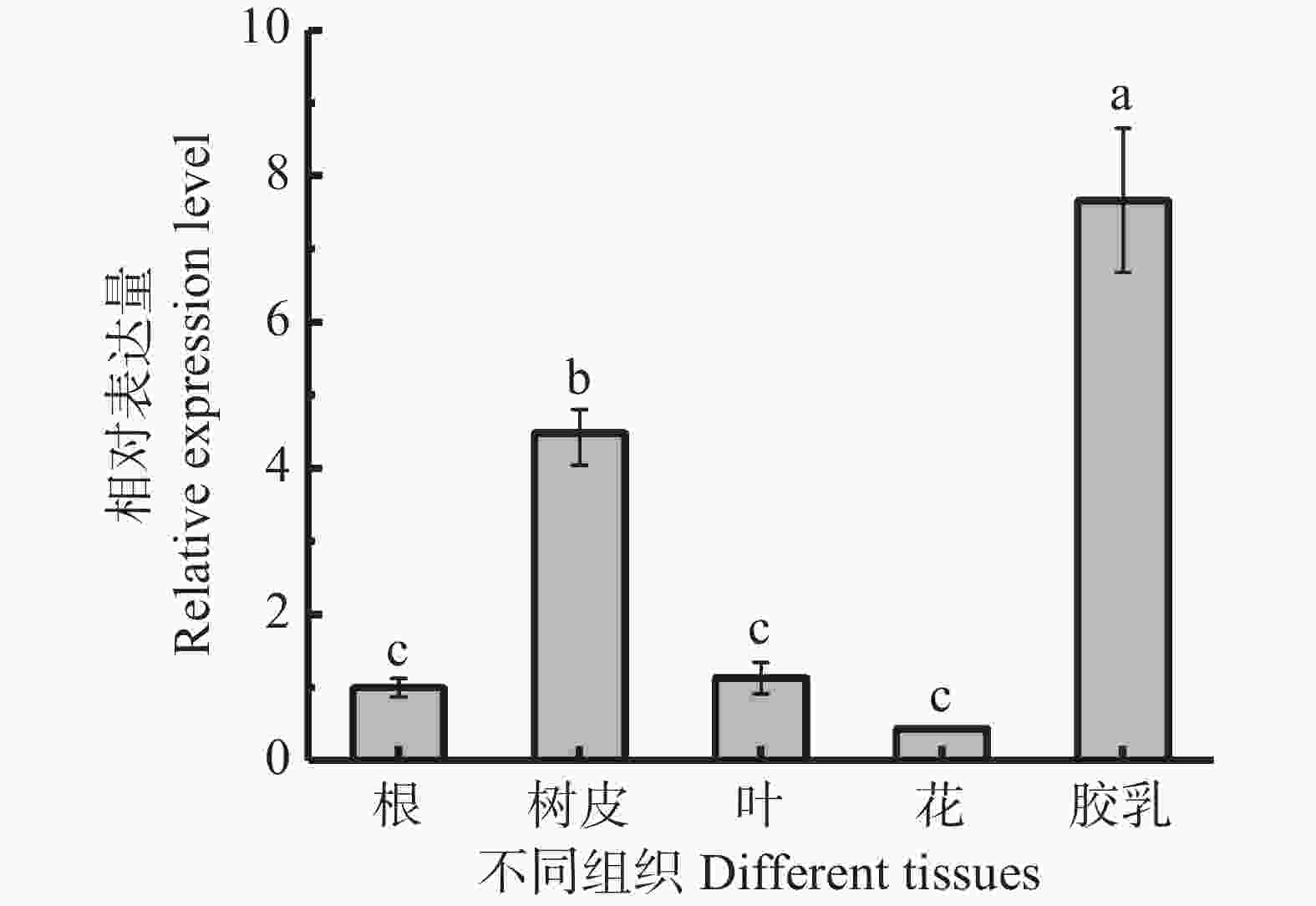
图 5 橡胶树不同组织HbbZIP74表达情况
Figure 5. Expression of HbbZIP74 in different tissues of the rubber tree
-
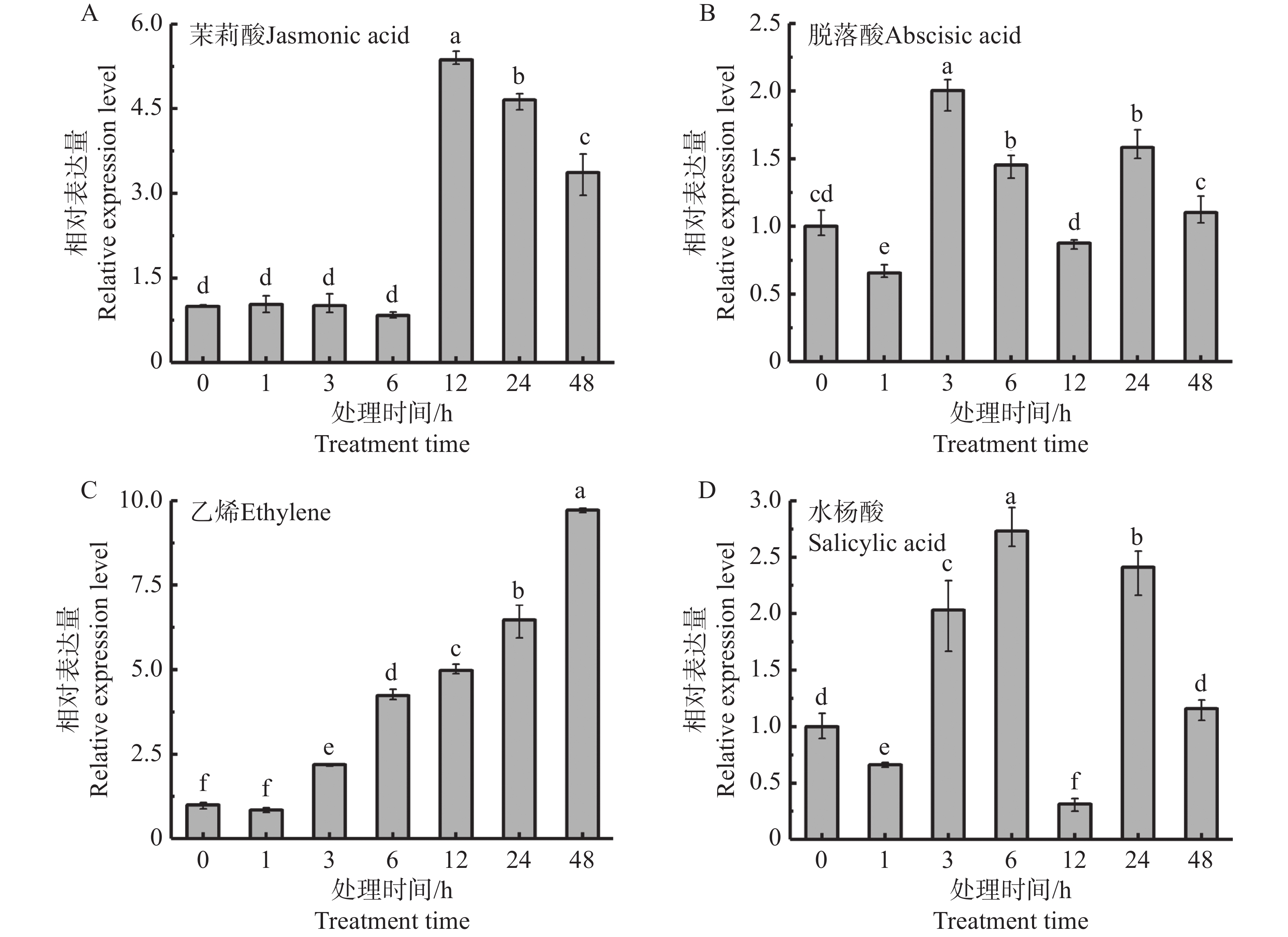
激素处理在天然橡胶的生物合成中起着重要的调控作用,因此检测了茉莉酸、脱落酸、乙烯和水杨酸处理的胶乳中HbbZIP74的表达情况。qRT-PCR结果显示,4种激素处理均能诱导HbbZIP74的表达(图6)。茉莉酸在处理6 h内,HbbZIP74的表达水平基本没有变化,12 h的表达量达到最高值,是正常表达水平的5.36倍(图6-A),脱落酸、乙烯和水杨酸在处理3 h均使HbbZIP74的表达超过正常水平的2倍(图6-B—D)。
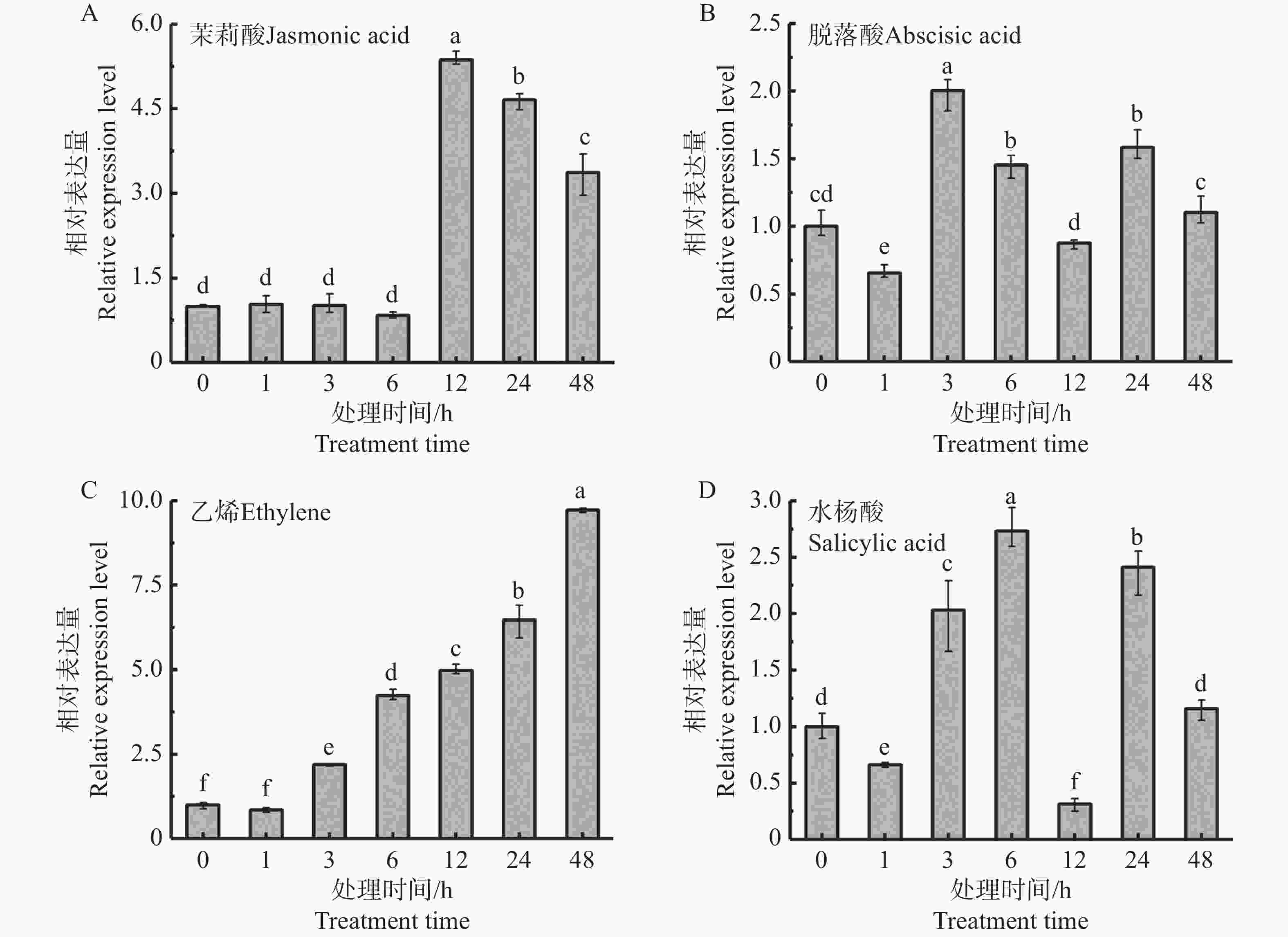
图 6 激素处理对胶乳中HbbZIP74表达的影响
Figure 6. Expressions of HbbZIP74 in latex under different hormone treatments
-
以pMD19T-HbbZIP74质粒为模板,使用引物NCF和NCR进行PCR扩增得到目标片段,通过同源重组将目标片段连接到线性化后的pET28a载体上,转化大肠杆菌。通过菌液PCR检测获得阳性克隆,结果如图7-A所示。阳性单克隆提取质粒,经测序与HbbZIP74的CDS序列一致,表明HbbZIP74蛋白原核表达载体构建成功,重组质粒命名为pET28-HbbZIP74,重组载体如图7-B所示。
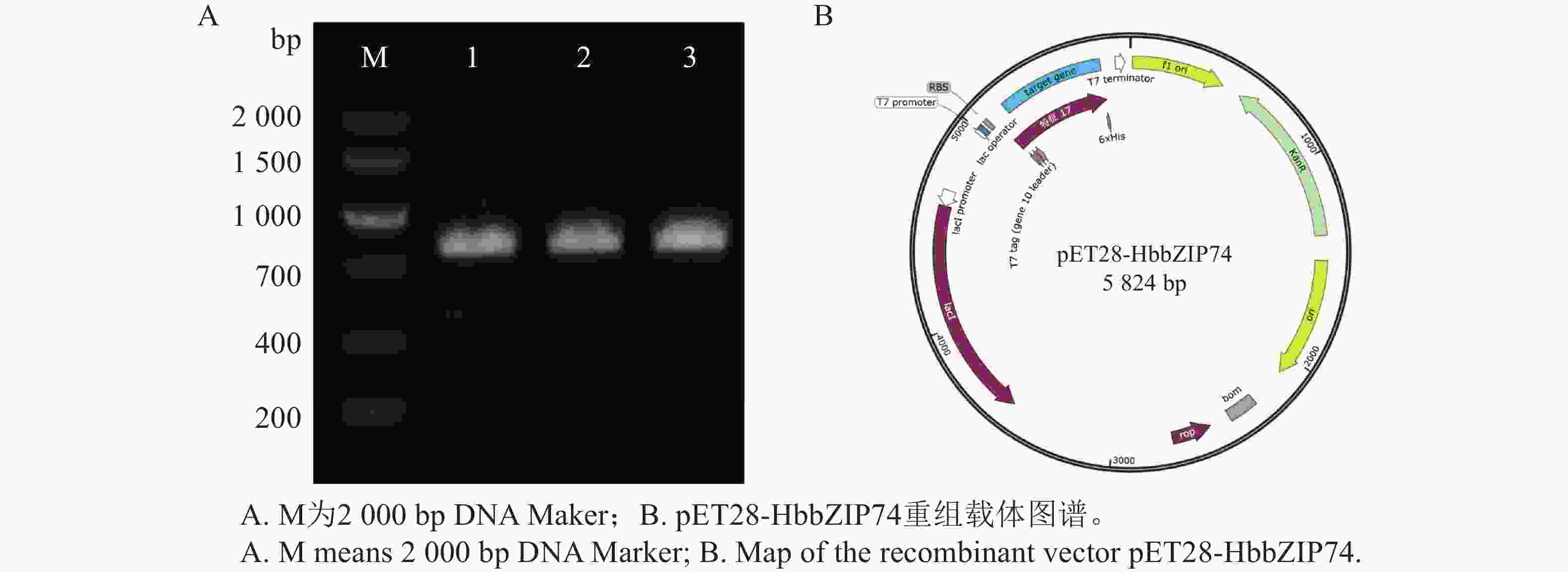
图 7 重组载体pET28-HbbZIP74的构建
Figure 7. Construction of recombinant vector pET28-HbbZIP74
-
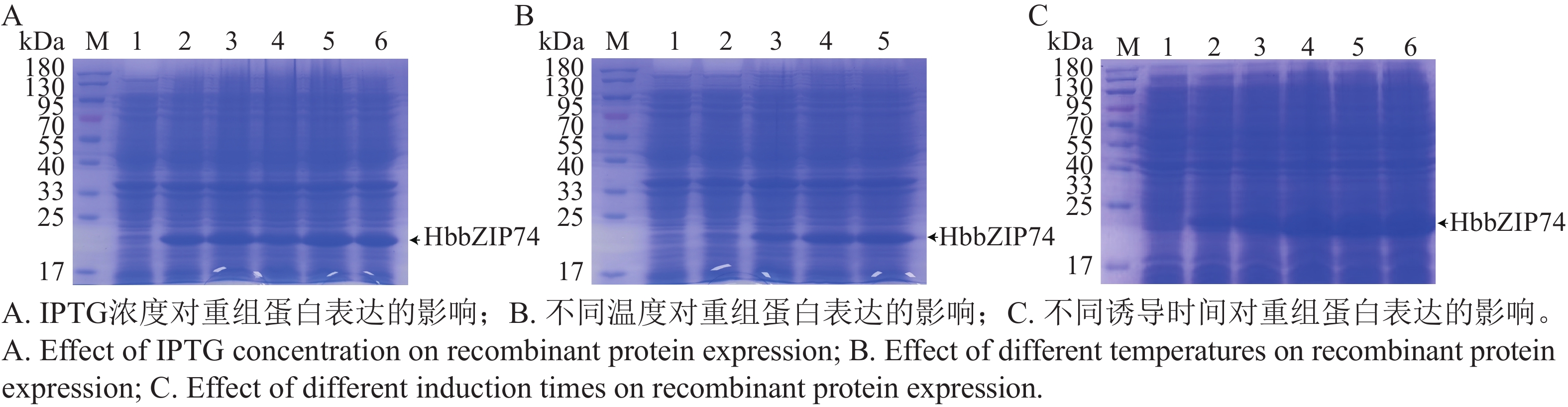
将pET28-HbbZIP74重组质粒转入BL21(DE3)菌株中,37 ℃扩大培养至OD600为0.6时,分别加入终浓度为0.1、0.3、0.5、1.0、2.0 mmol·L−1 IPTG后,37 ℃诱导5 h,SDS-PAGE蛋白电泳检测结果如图8-A所示,添加IPTG的菌液均诱导出分子质量约22 kDa蛋白,与预测的HbbZIP74-His重组蛋白分子质量大小一致,表明HbbZIP74-His蛋白在大肠杆菌BL21中成功诱导表达。泳道5和6诱导蛋白量相当且明显高于泳道2~4,说明1.0 mmol·L−1 IPTG为诱导重组蛋白表达的最适浓度。
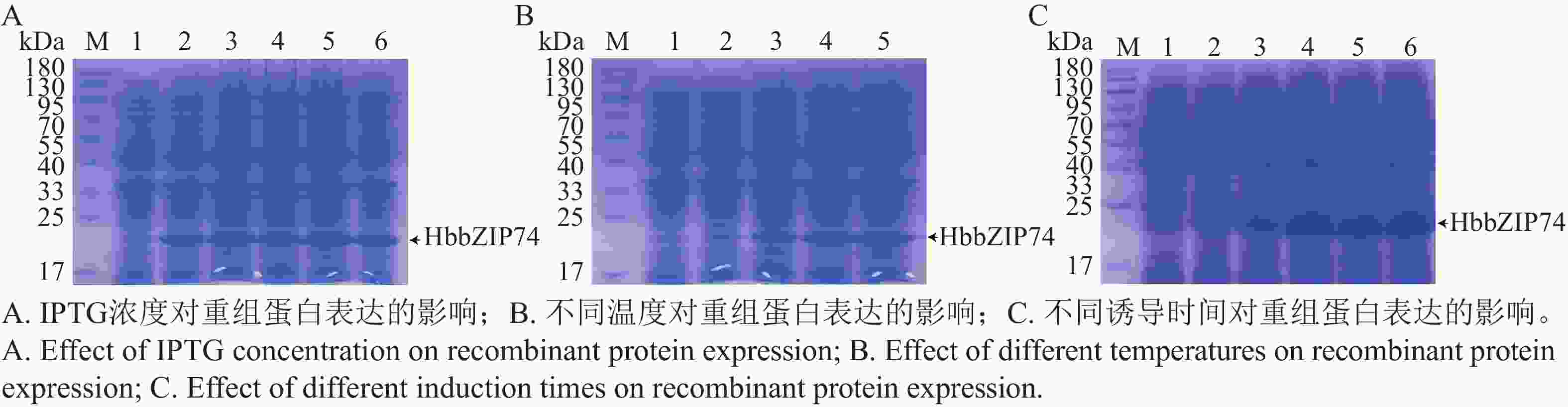
图 8 HbbZIP74蛋白诱导表达条件筛选
Figure 8. Screening of conditions for expression of HbbZIP74 protein by inducing
将OD600为0.6的菌液加入1 mmol·L−1终浓度的IPTG,分别在16、20、28、37 ℃诱导5 h,SDS-PAGE蛋白电泳检测结果(图8-B)表明,泳道2蛋白量最少,泳道5诱导蛋白表达量最高,表明在37 ℃重组蛋白表达效果最佳。
将OD600为0.6的菌液加入1 mmol·L−1终浓度的IPTG,在37 ℃分别诱导1、2、3、4、5 h,SDS-PAGE蛋白电泳检测结果(图8-C)表明,蛋白在1 h就检测到了表达,在3 h的表达量达到最大,继续增加诱导时间蛋白表达量没有明显增多。因此,确定HbbZIP74-His重组蛋白的最佳诱导条件为在37 ℃使用终浓度1.0 mmol·L−1 IPTG诱导3 h。
-
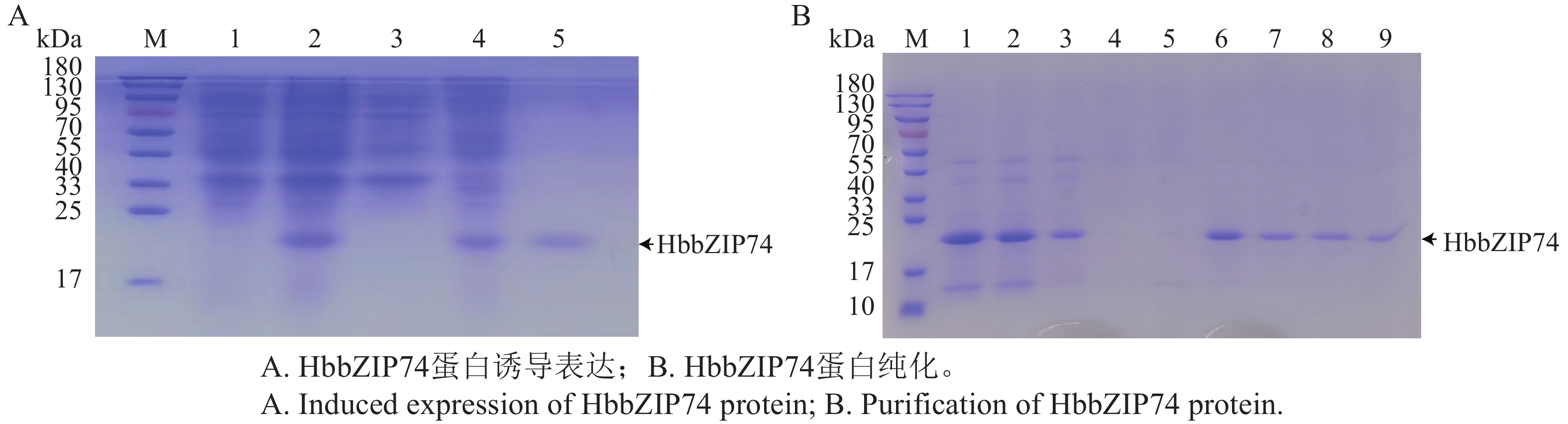
HbbZIP74-His重组蛋白的可溶性进行鉴定如图9-A所示,诱导样品上清液中(泳道3)无目的重组蛋白,而沉淀中(泳道4)包含目的蛋白,说明HbbZIP74-His重组蛋白主要以不可溶的包涵体形式存在。将沉淀中的包涵体蛋白进行纯化,SDS-PAGE电泳检测,在裂解液洗涤后杂蛋白基本被洗除,在洗脱液和10 mmol·L−1咪唑洗脱液中则无明显的条带,在50 mmol·L−1咪唑洗脱液泳道出现目的条带,后续纯化可用50 mmol·L−1咪唑洗脱液进行洗脱(图9-B)。洗脱后收集蛋白经透析后可获得纯化复性后的 HbbZIP74蛋白(图9-A,泳道5)。
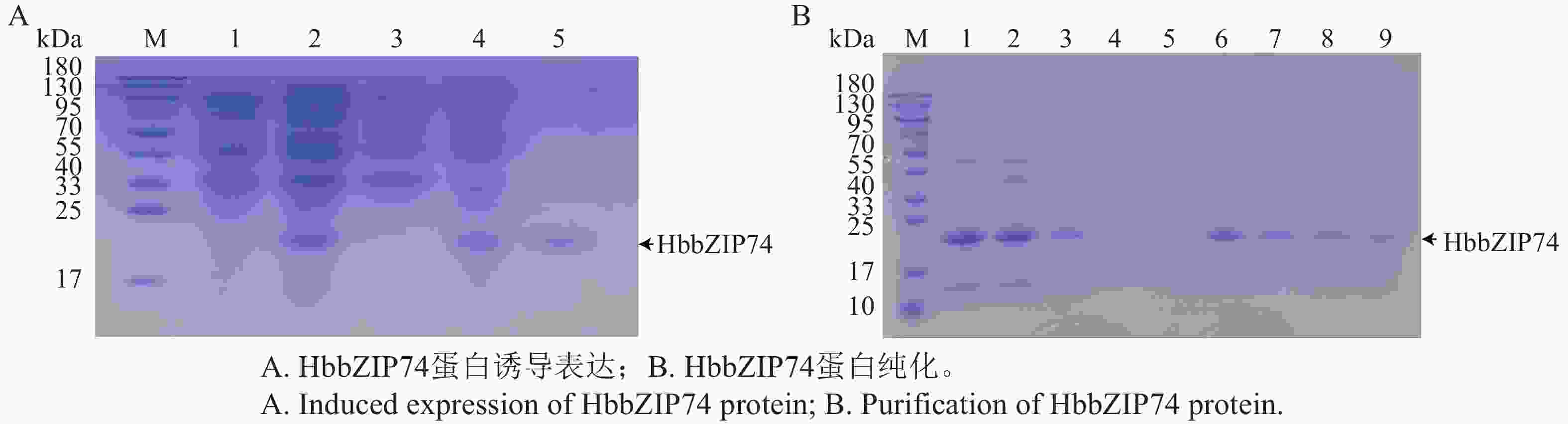
图 9 HbbZIP74蛋白诱导表达及纯化
Figure 9. Induced expression and purification of HbbZIP74 protein
-
bZIP转录因子作为植物中最大的转录因子家族之一,在植物生长发育、种子成熟和萌发、花发育、应对生物和非生物胁迫响应调控中的重要作用已有深入研究,在参与植物次生代谢,调控植物次生代谢产物的生物合成中也发挥了极其重要的作用。大量研究表明,bZIP转录因子可有效调节植物萜类化合物、黄酮类化合物和生物碱的生物合成。青蒿(Artemisia annua)中,除AabZIP外[25],AabZIP9也可以通过促进ADS和CYP71AV1的表达,正向调控青蒿素的积累[29]。GbbZIP08和GbbZIP15参与了银杏青花素的合成[30]。雷公藤(Tripterygium wilfordii)bZIP转录因子TwTGA1可以调节雷公藤内酯和倍半萜吡啶生物碱的生物合成[31]。天然橡胶是产胶植物体内合成的次生代谢产物,短角蒲公英(Taraxacum brevicorniculatum)和橡胶树中已有bZIP转录因子促进天然橡胶生物合成基因的表达,调节橡胶生物合成的报道[2,26]。本课题组前期通过酵母单杂交技术,在胶乳cDNA文库中筛选到与法尼基焦磷酸合酶、橡胶转移酶、小橡胶粒子蛋白和橡胶延长因子等橡胶生物合成相关酶和蛋白基因互作的bZIP转录因子HbbZIP74,表明其可能与天然橡胶生物合成调控相关。蛋白多序列比对显示,HbbZIP74具有bZIP转录因子的保守结构域,属于bZIP转录因子家族成员。亚细胞定位预测HbbZIP74定位于细胞核中,符合转录因子的一般特征。系统发育分析显示,HbbZIP74属于bZIP家族的S亚家族,该家族基因包含一类无内含子基因,编码蛋白分子质量一般约20 kDa,和C亚家族能形成同源和异源二聚体,在参与植物胁迫应答中发挥重要功能[15]。
橡胶树乳管细胞是特异化合成和储存天然橡胶的场所,上下贯通分布于橡胶树树皮的韧皮部。乳管的数量是决定天然橡胶产量的关键因素[32],天然橡胶生物合成相关基因在乳管细胞中高丰度表达[33]。HbbZIP74在胶乳中的表达量最高,树皮中次之,都显著高于根、叶和花等组织中的表达,暗示HbbZIP74可能与天然橡胶合成存在密切关系。目前,已经证实茉莉酸[14]、乙烯[34]、脱落酸和水杨酸[35]等激素在天然橡胶生物合成中起着重要的调控作用。本研究发现,茉莉酸、乙烯、脱落酸和水杨酸均能显著诱导胶乳中HbbZIP74的表达,乙烯对其影响最为显著,并可以持续到48 h,茉莉酸也使其表达量提高5倍多;脱落酸和水杨酸对HbbZIP74影响趋势比较一致,能使HbbZIP74的表达提高2~3倍,但在12 h时HbbZIP74表达量有明显的下降,显示激素对基因的调控存在复杂的机制。综上所述,HbbZIP74可能通过茉莉酸、乙烯、脱落酸和水杨酸信号途径参与对天然橡胶生物合成的调控。
转录因子编码蛋白的表达和纯化是开展其转录调控功能研究的基础,纯化蛋白可通过凝胶阻滞实验确定与靶基因启动子DNA的体外结合,以及结合部位,通过pull-down实验鉴定转录因子的互作蛋白。本研究成功构建了HbbZIP74的原核表达载体,并对HbbZIP74-His重组蛋白的最优表达条件进行摸索。除基因本身特性外,诱导剂浓度、诱导温度和时间都是影响蛋白表达的因素。本研究共使用5种浓度的IPTG诱导蛋白表达,结果表明,IPTG浓度的提高有助于蛋白表达量的增加,当浓度到1.0 mmol·L−1时,重组蛋白表达水平达到峰值;温度对重组蛋白的表达影响较大,16 ℃下细菌生长较慢,5 h几乎没有重组蛋白表达,20、28和37 ℃均有重组蛋白表达,且37 ℃重组蛋白表达量最高,这与37 ℃是大肠杆菌最适生长温度有关;HbbZIP74-His重组蛋白在37 ℃对诱导响应较快,在1 h就有较多表达,3 h后达到稳定表达状态。目前认为,包涵体的形成是由于蛋白在细胞内积累速率过快,没有正确折叠而聚集沉淀,HbbZIP74-His重组蛋白在37 ℃表达速率较快,可溶性检测显示主要以包涵体形式存在。本研究尝试降低诱导温度至16 ℃,降低诱导剂IPTG浓度为0.1 mmol·L−1,延长诱导时间至20 h,仍然无法获得可溶重组目的蛋白,推测形成包涵体可能与蛋白自身结构有较大关系。由于无法获得可溶重组蛋白,在37 ℃使用终浓度1.0 mmol·L−1 IPTG诱导3 h即为HbbZIP74-His蛋白快速高效表达的最优化条件。
通常包涵体蛋白缺乏生物活性,需要分离、纯化和复性用于后续研究。本研究中使用碱性缓冲液溶解包涵体,上清液经滤膜过滤后,使用Ni-NTA柱进行亲和纯化,咪唑洗脱后获得目标重组蛋白,洗脱液使用透析复性、富集,获得可用于下一步研究的HbbZIP74-His纯化蛋白。这一结果将为验证HbbZIP74与靶标基因启动子结合,HbbZIP74与其他蛋白相互作用提供了前提,也为深入研究转录因子HbbZIP74在天然橡胶生物合成中的功能奠定了基础。
本研究克隆了HbbZIP74基因,qRT-PCR分析结果显示,HbbZIP74在橡胶树胶乳和树皮中的表达水平较高,茉莉酸、脱落酸、乙烯和水杨酸处理,可以诱导胶乳中HbbZIP74的表达。通过构建pET28-HbbZIP74原核表达载体,在大肠杆菌(E. coli)BL21(DE3)菌株中进行异源表达和重组蛋白纯化,筛选出HbbZIP74-His重组蛋白的最佳表达条件为1 mmol·L−1 IPTG,37 ℃诱导3 h,重组蛋白主要存在于包涵体中。通过Ni-NTA亲和层析柱纯化获得大小约22 kDa的HbbZIP74-His重组蛋白,为进一步探究HbbZIP74在天然橡胶生物合成中的功能奠定基础。
橡胶树HbbZIP74基因的表达分析及蛋白原核表达纯化
DOI: 10.15886/j.cnki.rdswxb.20250015
 CSTR: 32425.14.j.cnki.rdswxb.20250015
CSTR: 32425.14.j.cnki.rdswxb.20250015
Expression analysis and prokaryotic expression purification of HbbZIP74 gene in rubber tree
-
摘要: 为研究巴西橡胶树(Hevea brasiliensis)bZIP转录因子在天然橡胶生物合成中的作用,本研究在实验室前期工作基础上,从橡胶树中克隆了1个bZIP类转录因子HbbZIP74基因,并对其进行了生物信息学分析和表达分析,成功构建pET28-HbbZIP74原核表达载体,在大肠杆菌(Escherichia coli) BL21(DE3)菌株中进行异源表达并对重组蛋白进行纯化。结果表明,HbbZIP74基因开放读码框(ORF)为486 bp,编码161个氨基酸,含有bZIP结构域,属于bZIP类转录因子S亚家族。HbbZIP74在胶乳和叶片中表达量较高,茉莉酸、脱落酸、乙烯和水杨酸可诱导胶乳中HbbZIP74表达。HbbZIP74-His重组蛋白异源表达最佳条件为1 mmol·L−1 IPTG, 37 ℃诱导3 h,但主要存在于包涵体中。通过Ni-NTA亲和层析柱纯化获得大小约22 kDa的重组蛋白,与预期结果一致。该研究为进一步探究HbbZIP74在天然橡胶生物合成中的功能奠定了基础。Abstract: In order to assess the role of the bZIP transcription factor in the biosynthesis of natural rubber in Hevea brasiliensis, HbbZIP74 gene was successfully cloned based on previous work in our laboratory. Bioinformatics analysis and expression pattern analysis were performed. The pET28-HbbZIP74 expression vector was constructed, and the recombinant protein was expressed in the strain Escherichia coli BL21 (DE3) and then purified. The results showed that the open reading frame (ORF) of the gene HbbZIP74 is 486 bp, encoding 161 amino acids, with a bZIP domain, classifying it as a bZIP transcription factor S subgroup. The expression level of HbbZIP74 is higher in latex and leaves, and its expression can be induced by jasmonic acid, abscisic acid, ethylene, and salicylic acid in latex. The optimal condition for the heterologous expression of the HbbZIP74-His recombinant protein was induced at 1 mmol·L−1 IPTG and 37 ℃ for 3 h. The recombinant protein was mainly accumulated in inclusion bodies. The recombinant protein, approximately 22 kDa in size, was purified using Ni-NTA affinity chromatography, which was consistent with expectations. This study lays the foundation for further exploration of the role of HbbZIP74 in the biosynthesis of natural rubber.
-
Key words:
- Hevea brasiliensis /
- bZIP transcription factor /
- gene cloning /
- gene expression /
- protein purification
-
图 2 HbbZIP74蛋白结构分析
A. aHbbZIP74蛋白保守结构域示意图;B. HbbZIP74蛋白二级结构示意图;C. HbbZIP74蛋白三级结构预测示意图;D. HbbZIP74亲水/疏水性分析图。
Fig. 2 Structural analysis of HbbZIP74 protein
A. Schematic diagram of conserved domains in HbbZIP74 protein; B. Schematic diagram of secondary structure of HbbZIP74 protein; C. Predicted tertiary structure of HbbZIP74 protein; D. Hydrophilicity/hydrophobicity analysis of HbbZIP74 protein.
图 3 HbbZIP74与其他物种氨基酸序列同源比对
注:Basic region,高度保守的N-x7-R/K基序;Leucine zipper,亮氨酸/疏水氨基酸拉链。
Fig. 3 Homologous alignment of HbbZIP74 amino acid sequences with other species
Note: Basic region, Highly conserved N-x7-R/K motif; Leucine zipper, Leu/hydrophobic amino acid zipper.
图 5 橡胶树不同组织HbbZIP74表达情况
注:不同小写字母表示差异显著(P<0.05),下同。
Fig. 5 Expression of HbbZIP74 in different tissues of the rubber tree
Note:Different lowercase letters indicated significant difference (P < 0.05), similarly hereinafter.
图 6 激素处理对胶乳中HbbZIP74表达的影响
Fig. 6 Expressions of HbbZIP74 in latex under different hormone treatments
图 7 重组载体pET28-HbbZIP74的构建
注:1~3为pET28-HbbZIP74同源重组转化后的阳性克隆验证。
Fig. 7 Construction of recombinant vector pET28-HbbZIP74
Note: 1−3 means positive clone verification after homologous recombination and transformation of pET28-HbbZIP74.
图 8 HbbZIP74蛋白诱导表达条件筛选
注:M,蛋白分子质量Marker;泳道1均未加IPTG;A泳道1~6,IPTG终浓度分别为0、0.1、0.3、0.5、1.0、2.0 mmol·L−1;B泳道1(16 ℃,0 mmol·L−1),泳道2~5,1 mm·L−1IPTG在16、20、28、37 ℃诱导。C泳道1~6,1 mm·L−1IPTG诱导0、1、2、3、4、5 h。
Fig. 8 Screening of conditions for expression of HbbZIP74 protein by inducing
Note: M, Protein molecular weight marker; Lane 1 all none IPTG; Lanes 1−6 in A, IPTG final concentrations of 0, 0.1, 0.3, 0.5, 1.0, and 2.0 mmol L−1, respectively; Lane 1 (16 °C, 0 mmol·L−1), lanes 2−5 in B, induced by 1 mmol·L−1 IPTG at 16, 20, 28, and 37 °C, respectively; Lanes 1−6 in C, induced by 1 mmol·L−1 for 0, 1, 2, 3, 4, and 5 h, respectively.
图 9 HbbZIP74蛋白诱导表达及纯化
注:M,蛋白分子质量Marker;A) 泳道1,未诱导样品;泳道2,诱导样品全菌液;泳道3,诱导样品上清;泳道4,诱导样品沉淀;泳道5,纯化后的蛋白。B) 1,蛋白裂解液;2,裂解液过滤后蛋白;3,上柱流穿液;4,洗脱液;5,10 mmol·L−1咪唑洗脱液;6~7,50 mmol·L−1咪唑洗脱液;8~9,100 mmol·L−1咪唑洗脱液。
Fig. 9 Induced expression and purification of HbbZIP74 protein
Note: M, Protein molecular weight marker; A) Lane 1, Uninduced sample; Lane 2, Induced sample whole cell lysate; Lane 3, Induced sample supernatant; Lane 4, Induced sample pellet; Lane 5, Purified protein; B) 1, Protein lysate; 2, Protein after lysate filtration; 3, Flow-through after column; 4, Elution buffer; 5, 10 mmol·L−1 imidazole elution buffer; Lanes 6−7, 50 mmol·L−1 imidazole elution buffer; Lanes 8−9, 100 mmol L−1 imidazole elution buffer.
-
[1] Cherian S, Ryu S B, Cornish K. Natural rubber biosynthesis in plants, the rubber transferase complex, and metabolic engineering progress and prospects [J]. Plant Biotechnology Journal, 2019, 17(11): 2041−2061. https://doi.org/10.1111/pbi.13181 doi: 10.1111/pbi.13181 [2] Bangrak P, Chotigeat W. Molecular cloning and biochemical characterization of a novel cystatin from Hevea rubber latex [J]. Plant Physiology and Biochemistry, 2011, 49(3): 244−250. https://doi.org/10.1016/j.plaphy.2010.12.007 doi: 10.1016/j.plaphy.2010.12.007 [3] Tang C, Yang M, Fang Y, et al. The rubber tree genome reveals new insights into rubber production and species adaptation [J]. Nature Plants, 2016, 2(6): 16073. https://doi.org/10.1038/nplants.2016.73 doi: 10.1038/nplants.2016.73 [4] De G N, Gholami A, Goormachtig S, et al. Transcriptional machineries in jasmonate-elicited plant secondary metabolism [J]. Trends in Plant Science, 2012, 17(6): 349−359. https://doi.org/10.1016/j.tplants.2012.03.001 doi: 10.1016/j.tplants.2012.03.001 [5] Wang Y, Guo D, Li H L, et al. Characterization of HbWRKY1, a WRKY transcription factor from Hevea brasiliensis that negatively regulates HbSRPP [J]. Plant Physiology and Biochemistry, 2013, 71: 283−289. https://doi.org/10.1016/j.plaphy.2013.07.020 doi: 10.1016/j.plaphy.2013.07.020 [6] Guo D, Yi H Y, Li H L, et al. Molecular characterization of HbCZF1, a Hevea brasiliensis CCCH-type zinc finger protein that regulates hmg1 [J]. Plant Cell Reports, 2015, 34(9): 1569−1578. https://doi.org/10.1007/s00299-015-1809-6 doi: 10.1007/s00299-015-1809-6 [7] Li H L, Wei L R, Guo D, et al. HbMADS4, a MADS-box transcription factor from Hevea brasiliensis, negatively regulates HbSRPP [J]. Frontiers in Plant Science, 2016, 7: 1709. [8] Li H L, Qu L, Guo D, et al. Histone deacetylase interacts with a WRKY transcription factor to regulate the expression of the small rubber particle protein gene from Hevea brasiliensis [J]. Industrial Crops and Products, 2020, 145: 111989. https://doi.org/10.1016/j.indcrop.2019.111989 doi: 10.1016/j.indcrop.2019.111989 [9] Guo D, Li H L, Wang Y, et al. A myelocytomatosis transcription factor from Hevea brasiliensis positively regulates the expression of the small rubber particle protein gene [J]. Industrial Crops and Products, 2019, 133: 90−97. https://doi.org/10.1016/j.indcrop.2019.01.052 doi: 10.1016/j.indcrop.2019.01.052 [10] Wang Y, Zhan D F, Li H L, et al. Transcriptome-wide identification and characterization of MYB transcription factor genes in the laticifer cells of Hevea brasiliensis [J]. Frontiers in Plant Science, 2017, 8: 1974. https://doi.org/10.3389/fpls.2017.01974 doi: 10.3389/fpls.2017.01974 [11] Wang Y, Zhan D F, Li H L, et al. Identification and characterization of the MADS-box genes highly expressed in the laticifer cells of Hevea brasiliensis [J]. Scientific Reports, 2019, 9(1): 12673. https://doi.org/10.1038/s41598-019-48958-9 doi: 10.1038/s41598-019-48958-9 [12] Qu L, Li H L, Guo D, et al. HbWRKY27, a group IIe WRKY transcription factor, positively regulates HbFPS1 expression in Hevea brasiliensis [J]. Scientific Reports, 2020, 10(1): 20639. https://doi.org/10.1038/s41598-020-77805-5 doi: 10.1038/s41598-020-77805-5 [13] Guo D, Yang Z P, Li H L, et al. The 14-3-3 protein HbGF14a interacts with a RING zinc finger protein to regulate expression of the rubber transferase gene in Hevea brasiliensis [J]. Journal of Experimental Botany, 2018, 69(8): 1903−1912. https://doi.org/10.1093/jxb/ery049 doi: 10.1093/jxb/ery049 [14] Deng X, Guo D, Yang S, et al. Jasmonate signalling in the regulation of rubber biosynthesis in laticifer cells of rubber tree, Hevea brasiliensis [J]. Journal of Experimental Botany, 2018, 69(15): 3559−3571. https://doi.org/10.1093/jxb/ery169 doi: 10.1093/jxb/ery169 [15] Dröge-laser W, Snoek B L, Snel B, et al. The Arabidopsis bZIP transcription factor family: an update [J]. Current Opinion in Plant Biology, 2018, 45: 36−49. https://doi.org/10.1016/j.pbi.2018.05.001 doi: 10.1016/j.pbi.2018.05.001 [16] 张瑜, 原淑佳, 李瑞锋, 等. 植物bZIP转录因子生物学功能研究进展[J]. 山西中医药大学学报, 2023, 24(2): 221−225. [17] Ellenberger T E, Brandl C J, Struhl K, et al. The GCN4 basic region leucine zipper binds DNA as a dimer of uninterrupted alpha helices: crystal structure of the protein-DNA complex [J]. Cell, 1992, 71(7): 1223−1237. https://doi.org/10.1016/S0092-8674(05)80070-4 doi: 10.1016/S0092-8674(05)80070-4 [18] Landschulz W H, Johnson P F, Mcknight S L. The leucine zipper: a hypothetical structure common to a new class of DNA binding proteins [J]. Science, 1988, 240(4860): 1759−1764. https://doi.org/10.1126/science.3289117 doi: 10.1126/science.3289117 [19] Guo Z, Dzinyela R, Yang L, et al. bZIP transcription factors: structure, modification, abiotic stress responses and application in plant improvement [J]. Plants, 2024, 13(15): 2058. https://doi.org/10.3390/plants13152058 doi: 10.3390/plants13152058 [20] Zhang M, Liu Y, Shi H, et al. Evolutionary and expression analyses of soybean basic Leucine zipper transcription factor family [J]. BMC Genomics, 2018, 19(1): 159. https://doi.org/10.1186/s12864-018-4511-6 doi: 10.1186/s12864-018-4511-6 [21] Li D, Fu F, Zhang H, et al. Genome-wide systematic characterization of the bZIP transcriptional factor family in tomato (Solanum lycopersicum L.) [J]. BMC Genomics, 2015, 16: 771. https://doi.org/10.1186/s12864-015-1990-6 doi: 10.1186/s12864-015-1990-6 [22] Duan L, Mo Z, Fan Y, et al. Genome-wide identification and expression analysis of the bZIP transcription factor family genes in response to abiotic stress in Nicotiana tabacum L [J]. BMC Genomics, 2022, 23(1): 318. https://doi.org/10.1186/s12864-022-08547-z doi: 10.1186/s12864-022-08547-z [23] Zhang B, Feng C, Chen L, et al. Identification and functional analysis of bZIP genes in cotton response to drought stress [J]. International Journal of Molecular Sciences, 2022, 23(23): 14894. https://doi.org/10.3390/ijms232314894 doi: 10.3390/ijms232314894 [24] Hu W, Yang H, Yan Y, et al. Genome-wide characterization and analysis of bZIP transcription factor gene family related to abiotic stress in cassava [J]. Scientific Reports, 2016, 6: 22783. https://doi.org/10.1038/srep22783 doi: 10.1038/srep22783 [25] Zhang F, Fu X, Lv Z, et al. A basic leucine zipper transcription factor, AabZIP1, connects abscisic acid signaling with artemisinin biosynthesis in Artemisia annua [J]. Molecular Plant, 2015, 8(1): 163−175. https://doi.org/10.1016/j.molp.2014.12.004 doi: 10.1016/j.molp.2014.12.004 [26] 赵同, 张盛敏, 曹杰, 等. 橡胶草bZIP基因家族鉴定及胶乳高表达基因功能分析[J]. 南方农业学报, 2025, 56(9): 2723−2735. https://doi.org/10.3969/j.issn.2095-1191.2025.09.003 doi: 10.3969/j.issn.2095-1191.2025.09.003 [27] Chang C, Liu Z, Wang Y, et al. A bZIP transcription factor, CaLMF, mediated light-regulated camptothecin biosynthesis in Camptotheca acuminata [J]. Tree Physiology, 2019, 39(3): 372−380. [28] Guo D, Li H L, Zhu J H, et al. HbTGA1, a TGA transcription factor from Hevea brasiliensis, regulates the expression of multiple natural rubber biosynthesis genes [J]. Frontiers in Plant Science, 2022, 13: 909098. https://doi.org/10.3389/fpls.2022.909098 doi: 10.3389/fpls.2022.909098 [29] Shen Q, Huang H, Zhao Y, et al. The transcription factor Aabzip9 positively regulates the biosynthesis of artemisinin in Artemisia annua [J]. Frontiers in Plant Science, 2019, 10: 1294. https://doi.org/10.3389/fpls.2019.01294 doi: 10.3389/fpls.2019.01294 [30] Han H, Xu F, Li Y, et al. Genome-wide characterization of bZIP gene family identifies potential members involved in flavonoids biosynthesis in Ginkgo biloba L. [J]. Scientific Reports, 2021, 11(1): 23420. https://doi.org/10.1038/s41598-021-02839-2 doi: 10.1038/s41598-021-02839-2 [31] Han J, Liu H T, Wang S C, et al. A class I TGA transcription factor from Tripterygium wilfordii Hook.f. modulates the biosynthesis of secondary metabolites in both native and heterologous hosts [J]. Plant Science, 2020, 290: 110293. https://doi.org/10.1016/j.plantsci.2019.110293 doi: 10.1016/j.plantsci.2019.110293 [32] 史敏晶, 张世鑫, 晁金泉, 等. 橡胶树1981'IRRDB野生种质预割诱导次生乳管分化的比较分析[J]. 热带作物学报, 2022, 43(2): 303−310. https://doi.org/10.3969/j.issn.1000-2561.2022.02.010 doi: 10.3969/j.issn.1000-2561.2022.02.010 [33] 张豪, 张华, 黎晓思, 等. 橡胶树无性系热研879胶乳HbPPO2基因克隆及表达分析[J]. 南方农业学报, 2025, 56(9): 2793−2804. https://doi.org/10.3969/j.issn.2095-1191.2025.09.009 doi: 10.3969/j.issn.2095-1191.2025.09.009 [34] Gao L, Sun Y, Wu M, et al. Physiological and proteomic analyses of molybdenum- and ethylene-responsive mechanisms in rubber latex [J]. Frontiers in Plant Science, 2018, 9: 621. https://doi.org/10.3389/fpls.2018.00621 doi: 10.3389/fpls.2018.00621 [35] Tungngoen K, Viboonjun U, Kongsawadworakul P, et al. Hormonal treatment of the bark of rubber trees (Hevea brasiliensis) increases latex yield through latex dilution in relation with the differential expression of two aquaporin genes [J]. Journal of Plant Physiology, 2011, 168(3): 253−262. https://doi.org/10.1016/j.jplph.2010.06.009 doi: 10.1016/j.jplph.2010.06.009 -

 点击查看大图
点击查看大图
计量
- 文章访问数: 1110
- HTML全文浏览量: 734
- PDF下载量: 46
- 被引次数: 0
