

-
蚊子(Diptera: Culicidae)作为全球范围内传播人类和动物病原体的重要媒介,对公共卫生和动物健康构成了重大威胁[1]。蚊子能够携带并传播多种病原体,包括疟原虫(Plasmodium spp.)[2]、班氏丝虫(Wuchereria bancrofti)、马来丝虫(Brugia malayi)[3]、乙型脑炎病毒(Japanese encephalitis virus, JEV)[4]、登革热病毒(Dengue virus, DENV)、寨卡病毒(Zika virus, ZIKV)[5]以及基孔肯雅热病毒(Chikungunya virus, CHIKV)[6]等。全球已知蚊子种类约
3500 种,而在中国,已记录的种类超过400种[7]。在这些众多种类中,伊蚊属(Aedes)[8]、按蚊属(Anopheles)和库蚊属(Culex)因其在疾病传播中的显著作用而尤为重要[9]。海南岛位于北纬18°10′至20°10′,东经108°37′至111°03′[10],属于热带季风海洋性气候区,全年高温高湿的气候条件为蚊子的繁殖提供了极为有利的环境[11]。蚊子的活动不仅通过叮咬吸血和骚扰人类和动物,影响其正常生活,还通过传播疾病给生猪养殖产业带来重大经济损失[12]。蚊子体内寄居着众多共生微生物,包括细菌、真菌、古菌、病毒以及一些小型原生生物。这些微生物对蚊子的生理机能具有重要影响,它们协助蚊子进行食物消化,供应必需的营养,提升宿主的免疫功能和解毒能力,并对蚊子的生长、寿命以及繁殖行为产生影响[13]。微生物与蚊子之间的复杂交互是科研的重点领域之一。然而,关于海南岛猪场蚊子共生微生物的研究相对较少。
宏基因组学技术为研究昆虫共生微生物群落提供了一种无需培养的方法,进而快速鉴定包括古菌、细菌、真菌和病毒在内的多样化微生物。该技术在揭示宿主生理过程中微生物的作用方面具有显著优势,为生物防治策略的开发提供了新的研究途径[14]。对这些共生关系的深入研究有助于开发新型生物杀虫剂[15],并为未来的害虫生物防治提供新的思路。
-
Ezup柱式动物基因组DNA抽提试剂盒、琼脂糖和DL 2000 DNA Marker购自上海生物工程股份有限公司;DNA抽提试剂盒(E.Z.N.A.® Soil DNA Kit)购自美国Omega Bio-Tek公司;Goldview染料购自苏州金唯智生物科技有限公司;2X Taq Plus Master Mix II(Dye Plus)购自南京诺维赞生物科技股份有限公司;光触媒诱蚊灯(功夫小帅)购自河南智科弘润环保科技有限公司;连续变倍体视显微镜(XTL-BM-7B)购自上海彼爱姆光学仪器制造有限公司;PCR仪(Tprofessional)购自德国Biometra 公司;小型台式高速冷冻离心机(5415R)购自Eppendorf公司;琼脂糖水平电泳仪(Mini-PROTEAN Tetra)购自北京六一仪器厂产品;凝胶成像分析系统(GelDoc XR+)购自美国Bio-Rad公司。
-
2024年7月至2024年9月,在海南文昌市、定安县、白沙县、乐东县4个不同市县的猪场,于日落前1小时放置光触媒诱蚊灯,日出后1小时收集蚊虫。收集的蚊虫保存在95%酒精中,置于−20 ℃环境。
-
收集的蚊虫通过体视显微镜观察形态进行分组,参照《中国重要医学昆虫分类与鉴别》进行形态学鉴定。随后按照分组,每组取出2只蚊子,使用DNA提取试剂盒(生工公司)提取DNA,对提取的DNA的COI基因进行PCR扩增,使用1%的琼脂糖凝胶电泳,胶回收产物送至生工生物公司测序,测序结果与NCBI数据库比对,最终确定蚊子的种类及数量。
-
分别对4个不同猪场收集到的蚊子采用95%的酒精及ddH2O进行清洗,去除表面的微生物。清洗过后的蚊子置于超净台中风干水分,每个猪场各称量20 mg蚊子(大约10只)使用E.Z.N.A.® Soil DNA Kit(Omega Bio-Tek公司,美国)提取试剂盒提取总DNA,使用NanoDrop2000检测DNA纯度,使用TBS-380检测DNA浓度,使用1% 琼脂糖凝胶电泳检测DNA完整性。质检合格的DNA送至武汉百奥维凡生物科技有限公司构建PE文库和上机测序。
-
使用软件fastp(version 0.20.0)去除剪切后长度小于50 bp、平均碱基质量值低于20以及含N碱基的原始序列数据。同时使用软件BWA(version 0.7.9a)将原始序列数据比对蚊子宿主DNA序列,并去除比对相似性高宿主序列。利用拼接软件MEGAHIT(version 1.1.2)对优化序列进行拼接组装,使用 Prodigal软件(
http://metagene.cb.k.u-tokyo.ac.jp/ )对拼接结果中≥500 bp的连续序列进行预测,之后使用软件CD-HIT(version 4.6.1)对所有样品预测出来的基因序列进行聚类(参数设置为相似度≥95%,覆盖度≥90%),去除冗余,构建非冗余基因集。使用软件Diamond(version 0.8.35)将非冗余基因集的氨基酸序列与NR数据库进行比对(BLASTP比对参数设置期望值e-value为1e-5),获得物种注释。然后与KEGG数据库进行比对,获得基因对应的KEGG功能。 -
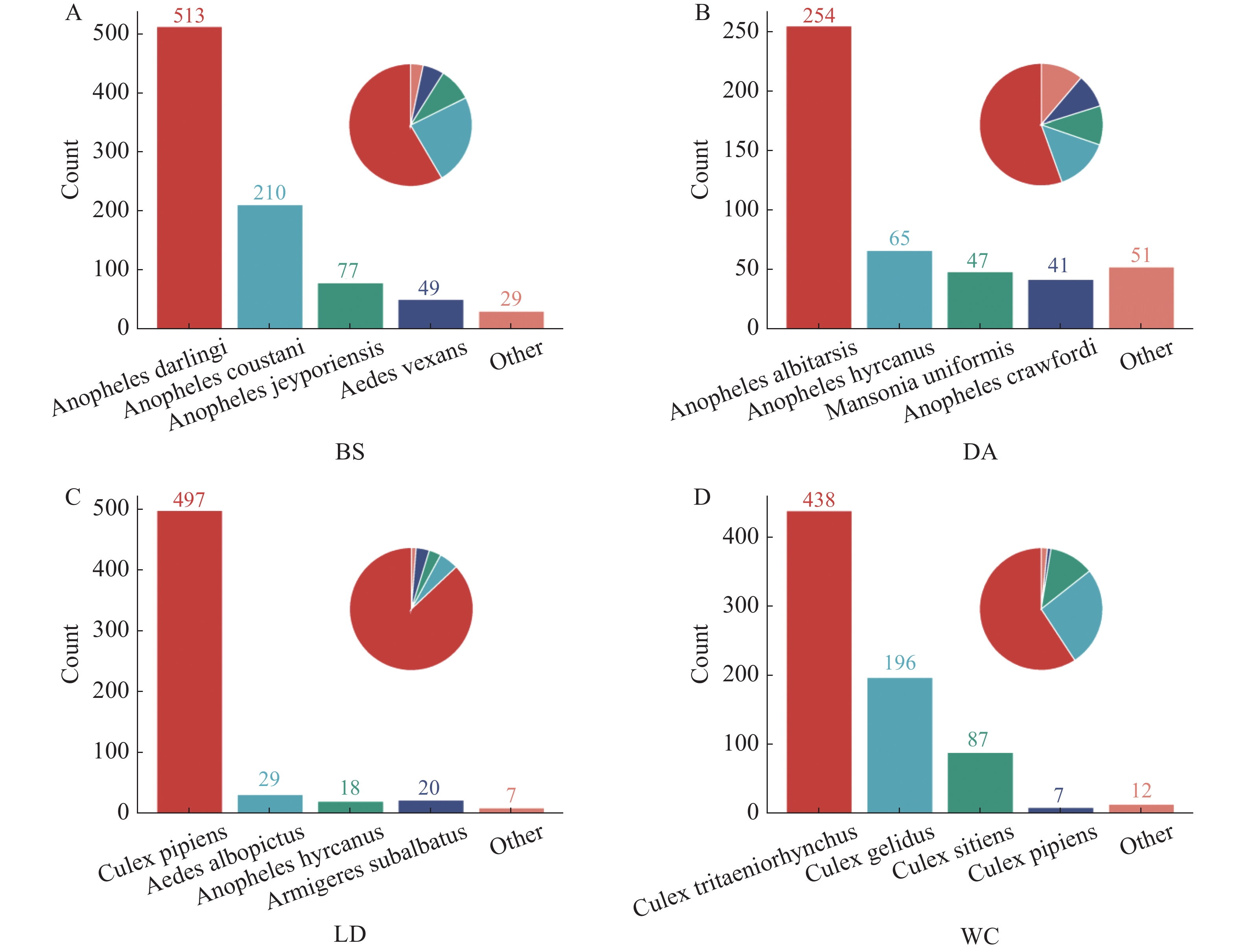
2024年7月至2024年9月,在白沙黎族自治县打安镇可程村、定安县黄竹镇同古村、乐东黎族自治县抱罗村和文昌市罗豆农场4个不同市县的猪场进行采样(猪场地理位置及气候条件见表1),提取总DNA,对COI基因进行扩增,凝胶电泳结果见图1。测序结果显示白沙采集的优势蚊种为达氏按蚊,定安采集的优势蚊种为中华按蚊,乐东采集的优势蚊种为尖音库蚊,文昌采集的优势蚊种为三带喙库蚊,占比分别为58.43%、55.46%、88.65%和57.84%。见图2。
图 1 海南不同地区猪场蚊媒COI基因的凝胶电泳图
Figure 1. Gel electrophoresis map of mosquito-borne COI gene in pig farms in different regions of Hainan
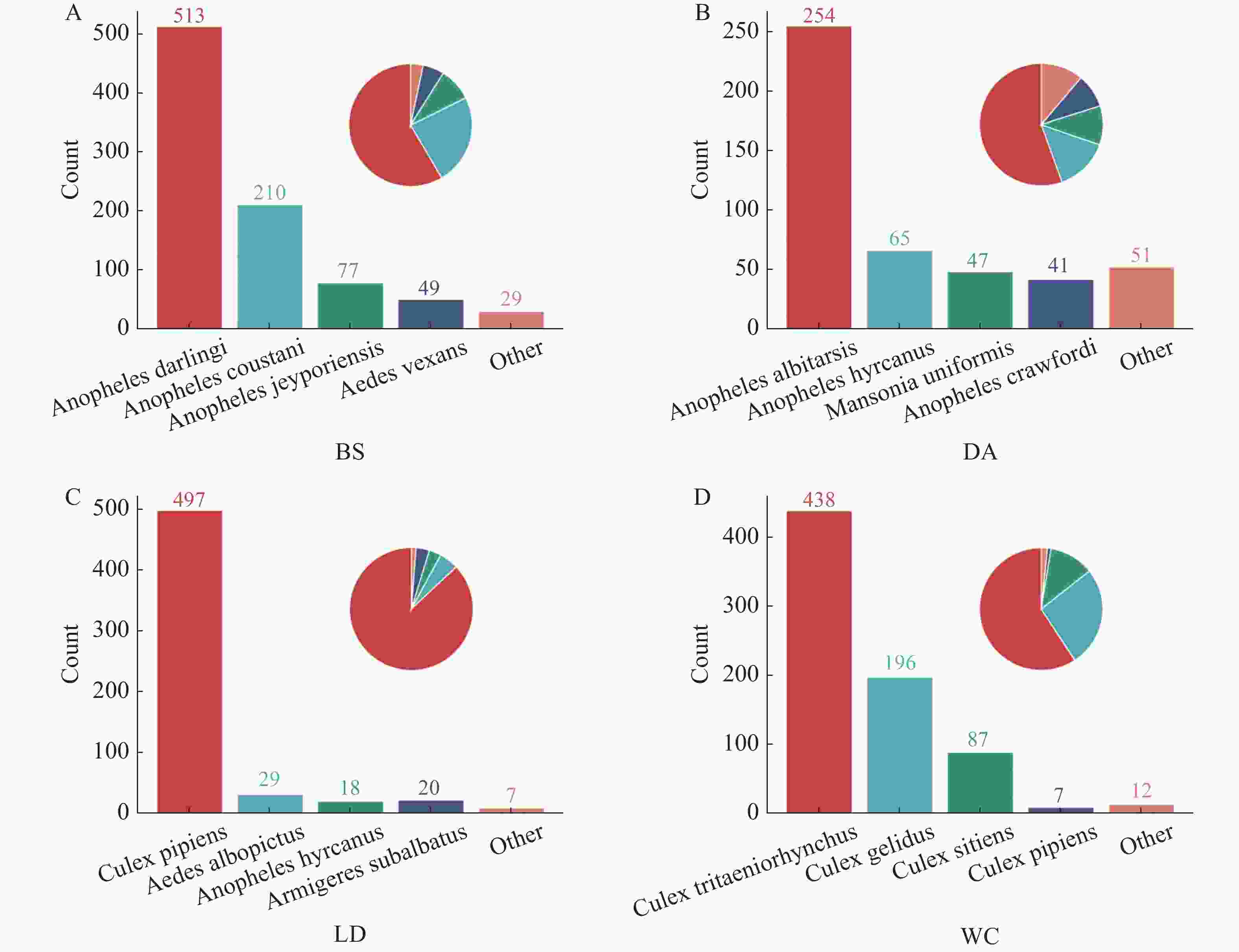
图 2 4个猪场蚊子种类分类
Figure 2. Classification of mosquito species in 4 pig farms
-
4个蚊子样本宏基因测序共得到原始数据48.3G,有效数据47.97G,数据有效率为99.32%,GC占比36.19%,Q30占比97.96%。见表2
表 2 2024年4个不同市县的4个不同猪场宏基因组测序数据
Table 2. Metagenomic sequencing data from four different pig farms in four different cities and counties in 2024
样本名称 原始数据(G) 有效数据(G) 有效数据率/% GC/% Q30/% BS 12.03 11.95 99.33 30.73 97.86 DA 12.06 11.98 99.34 37.51 98.09 LD 11.79 11.70 99.24 37.85 97.88 WC 12.42 12.34 99.36 38.65 97.99 注:BS为白沙采集的样本;DA为定安采集的样本;LD为乐东采集的样本;WC为文昌采集的样本。GC为有效数据中G和C两种碱基占总碱基的百分比;Q30是测序数据中每个碱基的Phred质量分数至少为30(识别错误率不超过0.1%)。 -
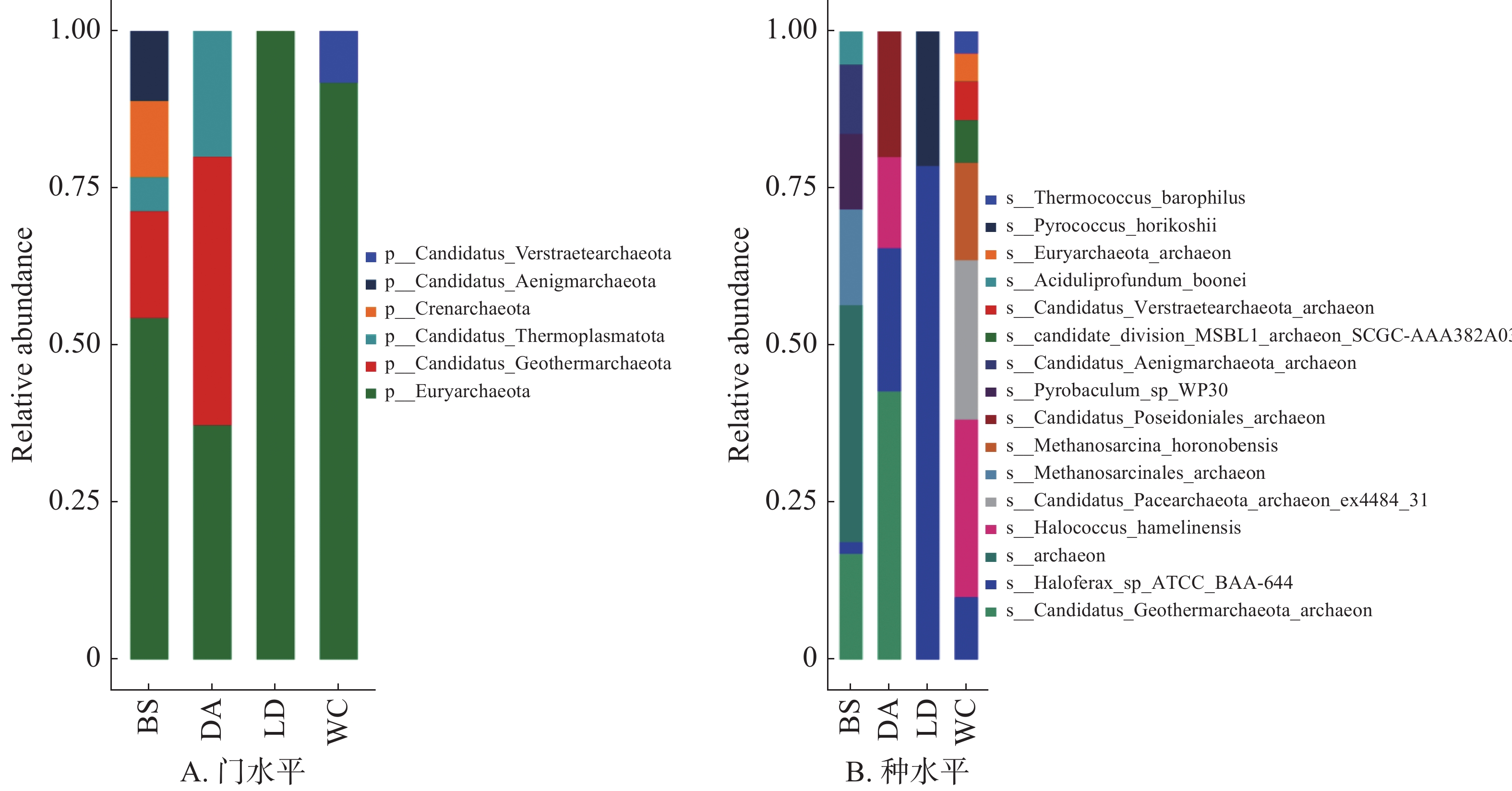
在门水平上(图3A),广古菌门(Euryarchaeota)为BS、LD、WC的优势门,在LD中占比为100%,在DA中为第二优势门,占比为37.25%,而热古菌门(Candidatus Geothermarchaeota)则为DA的优势门。

图 3 不同猪场的蚊子携带的古生菌不同水平上的相对丰度
Figure 3. Relative abundance of archaea carried by mosquitoes in different pig farms at different levels
在种水平上(图3B),4个样本共检测出16种古细菌。相对丰度较高的前5个物种分别为Candidatus Geothermarchaeota archaeon、Candidatus Pacearchaeota archaeon ex4484 31、Halococcus hamelinensis、Haloferax sp ATCC BAA-644和archaeon,检出的古细菌中尚未出现对人或猪产生疾病影响的物种的报道。
-
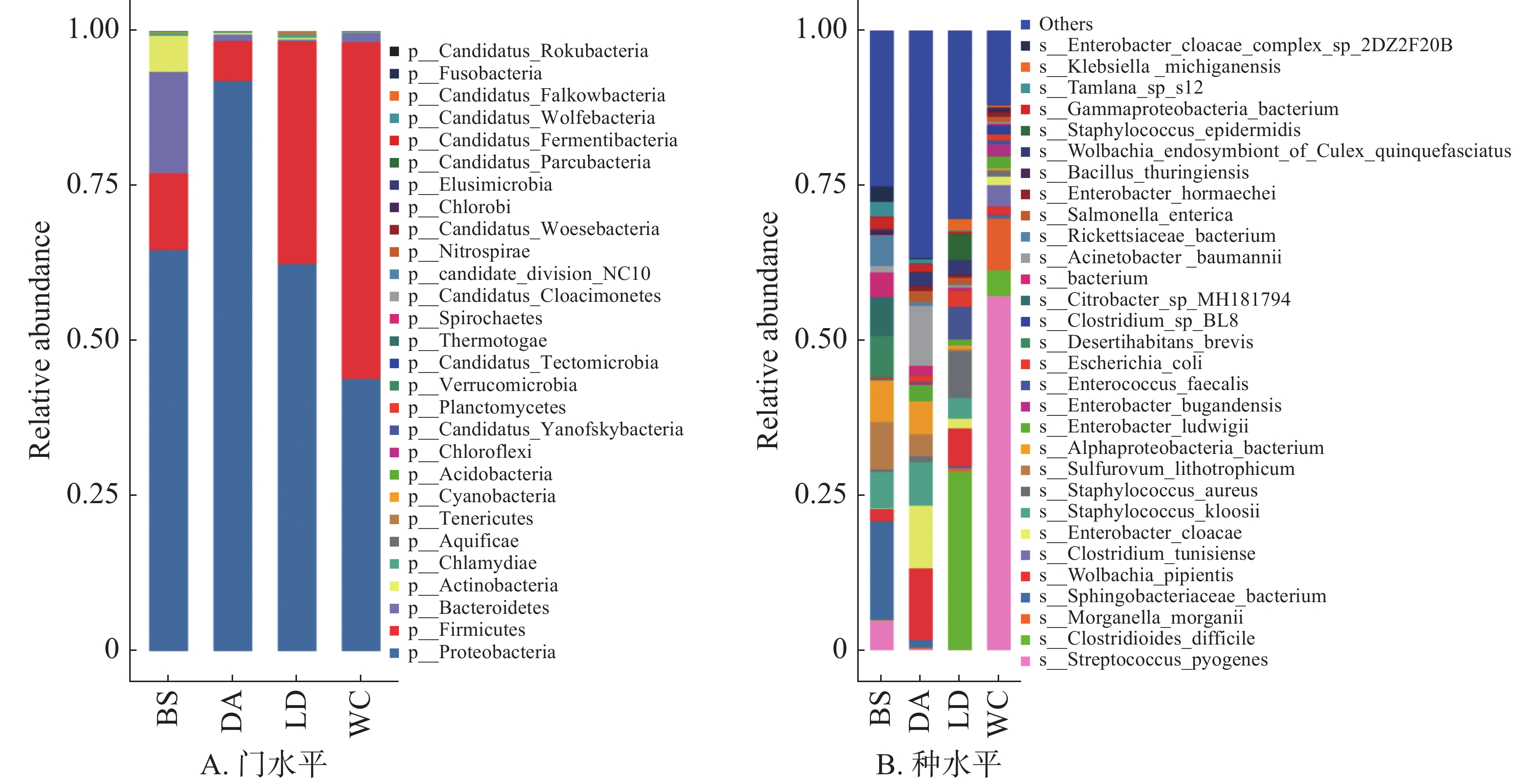
在门水平上(图4A),变形菌门(Proteobacteria)为4个样本的优势菌门,厚壁菌门(Firmicutes)为DA、LD和WC的第二大优势门。
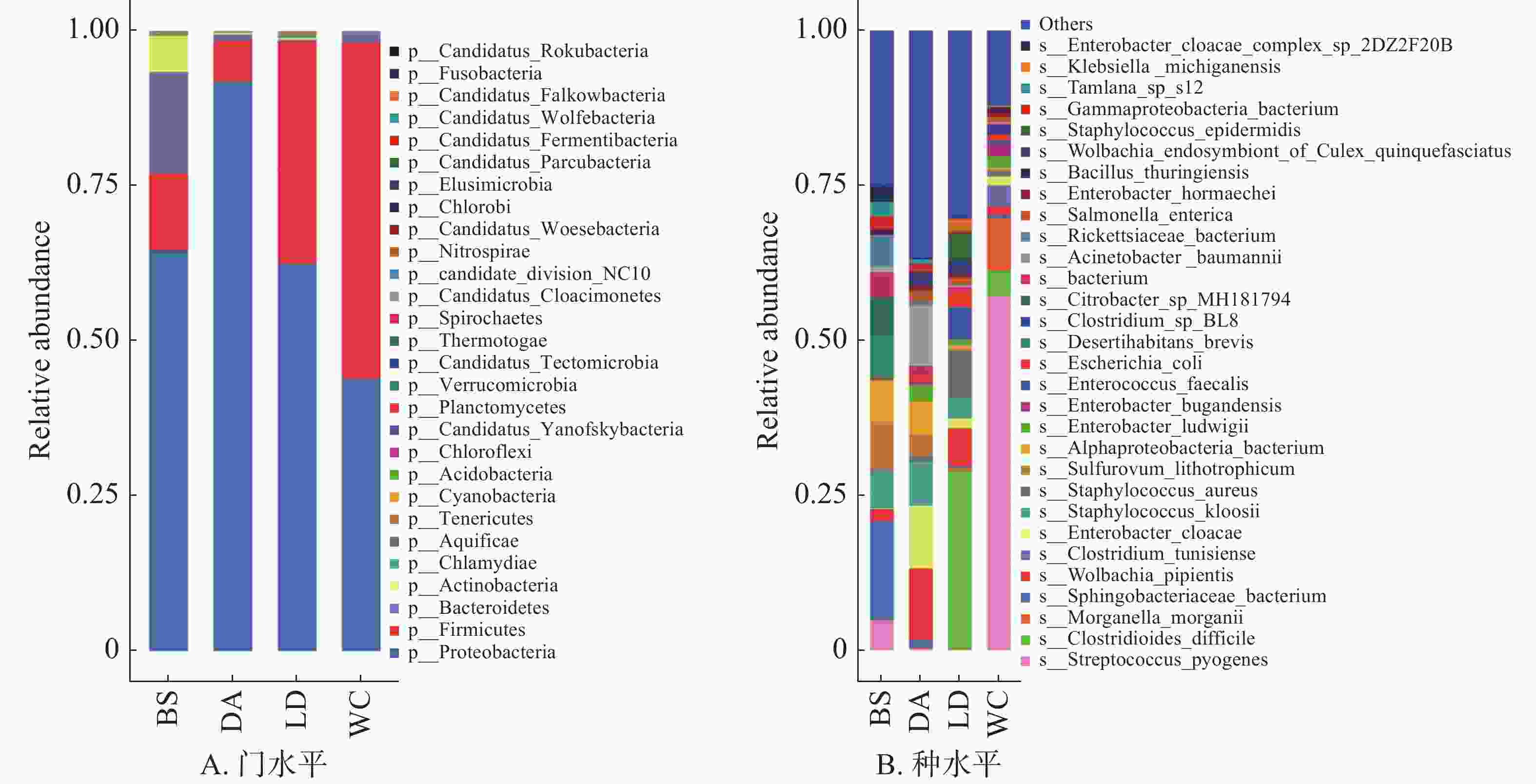
图 4 不同猪场的蚊子携带的细菌不同水平上的相对丰度
Figure 4. Relative abundance of bacteria carried by mosquitoes in different pig farms at different levels
共有
1357 种细菌注释到种水平上,相对丰度排名前30的物种箱式图见图4B。其中能导致人兽共患病的细菌有毛肠短状螺旋体(Brachyspira pilosicoli)、布鲁氏菌(Brucella melitensis)、流产衣原体(Chlamydia abortus)、钩端螺旋体(Leptospira interrogans)和软骨滑液菌(Waddlia chondrophila),BS含有布鲁氏菌,DA含有钩端螺旋体,LD含有毛肠短状螺旋体、流产衣原体和软骨滑液菌,WC含有流产衣原体和软骨滑液菌。毛肠短状螺旋体和钩端螺旋体能引发钩端螺旋体病,可以导致从轻微疾病到严重疾病,如器官损伤、脑膜炎和致命的出血性疾病[16]。布鲁氏菌[17]、流产衣原体[18]和软骨滑液菌[19]均会导致流产和不育症,对动物生产和繁殖以及人类健康造成极大的危害。其余少部分物种,如鲍曼不动杆菌(Acinetobacter baumannii)、蜡样芽孢杆菌(Arcobacter butzleri)、肉毒梭菌(Clostridium botulinum)和迟钝爱德华氏菌(Edwardsiella tarda)等大多为条件致病菌,主要与人类或动物的胃肠疾病、中毒以及感染有关。
-
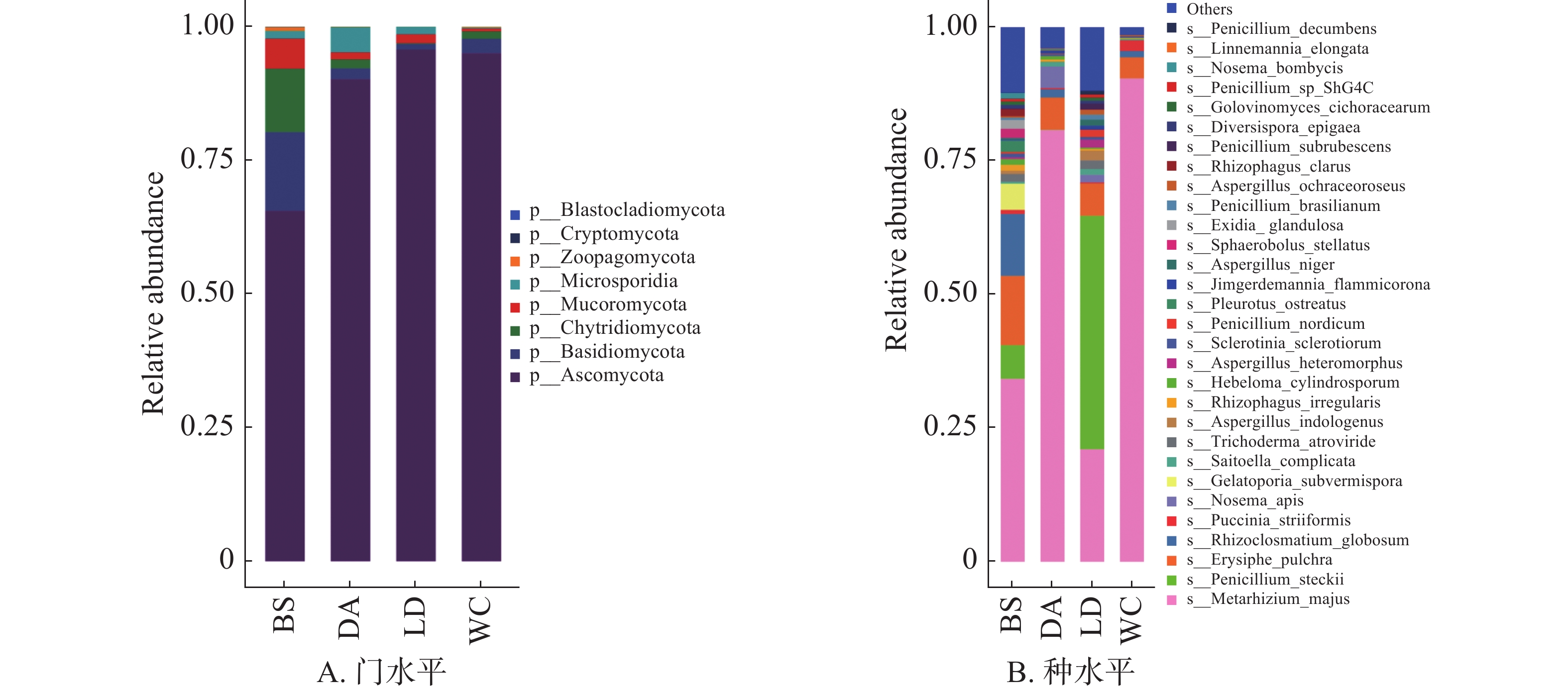
4个样本的真菌分布在门水平上具有较高的相似性,子囊菌门(Ascomycota)为4个样本的优势门(图5B),在BS中占比超过65%,其余3个样本中占比超过90%。
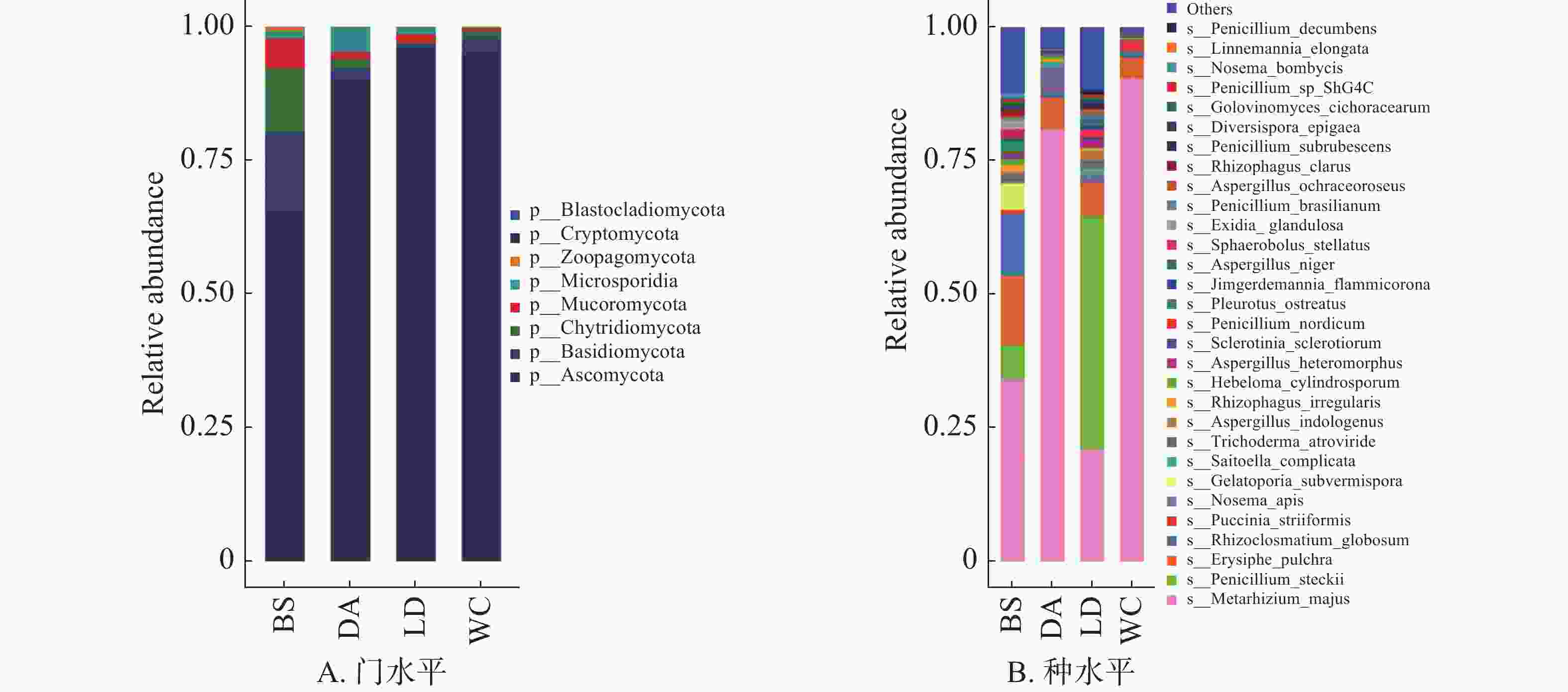
图 5 不同猪场的蚊子携带的真菌不同水平上的相对丰度
Figure 5. Relative abundance of fungi carried by mosquitoes in different pig farms at different levels
共有303种真菌注释到种水平上,图5B的箱式图仅展示排名前30的物种。其中有10余种会对人类和动物产生危害,如枝顶孢霉(Acremonium asperulatum)、土曲霉(Aspergillus terreus)和犬小孢子菌(Microsporum canis)等,主要表现为产生毒素或导致感染。
-
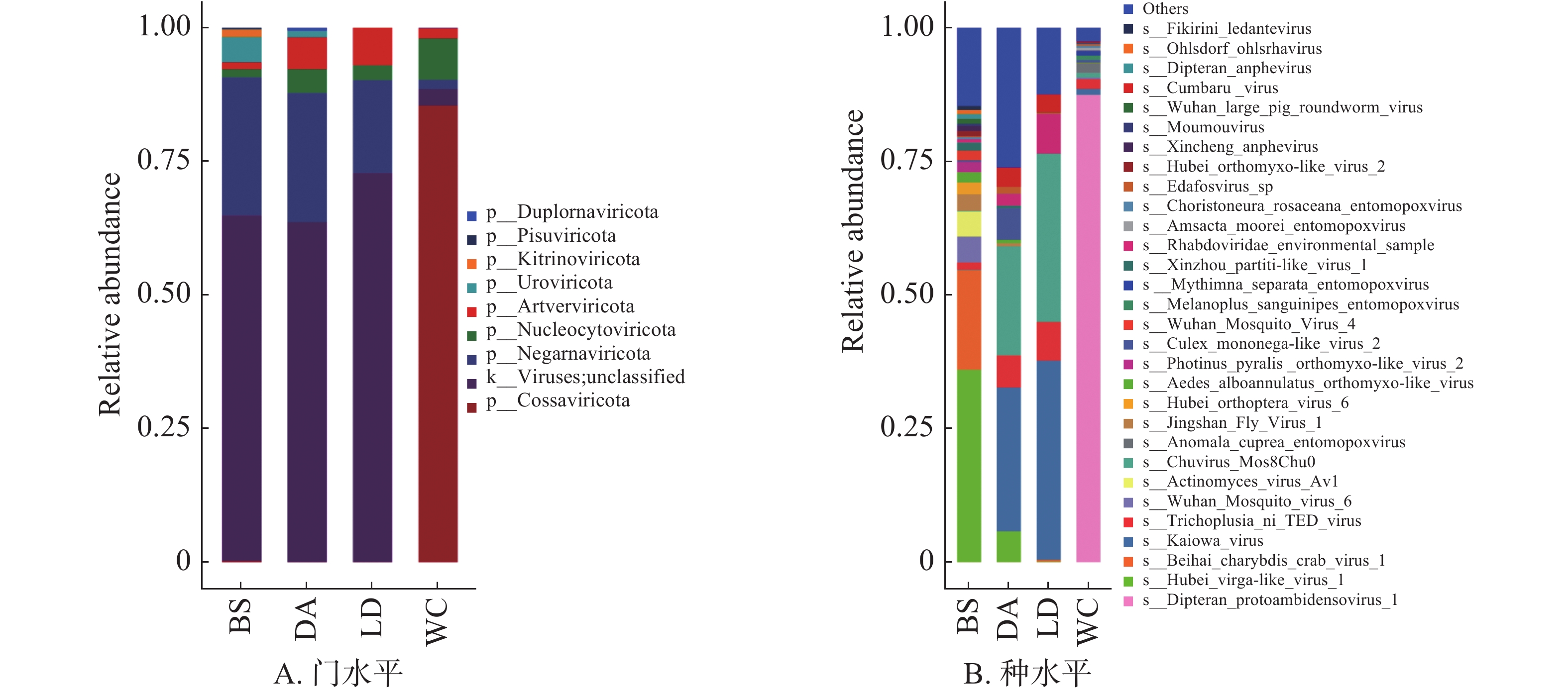
BS、DA、LD和WC分别有35.57%、36.39%、27.21%和96.93%可以注释到门水平(图6A),前3个样本的优势门均为负链RNA病毒门(Negarnaviricota),而WC的优势门为科萨特病毒门(Cossaviricota)。在种水平上(图6B),4个样本共注释到102种病毒,病毒数量排名为BS>WC>DA>LD,湖北virga样病毒1(Hubei virga-like virus 1)为BS的优势病毒,卡伊奥瓦病毒(Kaiowa virus)为DA和LD的优势病毒,双翅目原病毒1(Dipteran protoambidensovirus 1)为WC的优势病毒,占比超过87%。
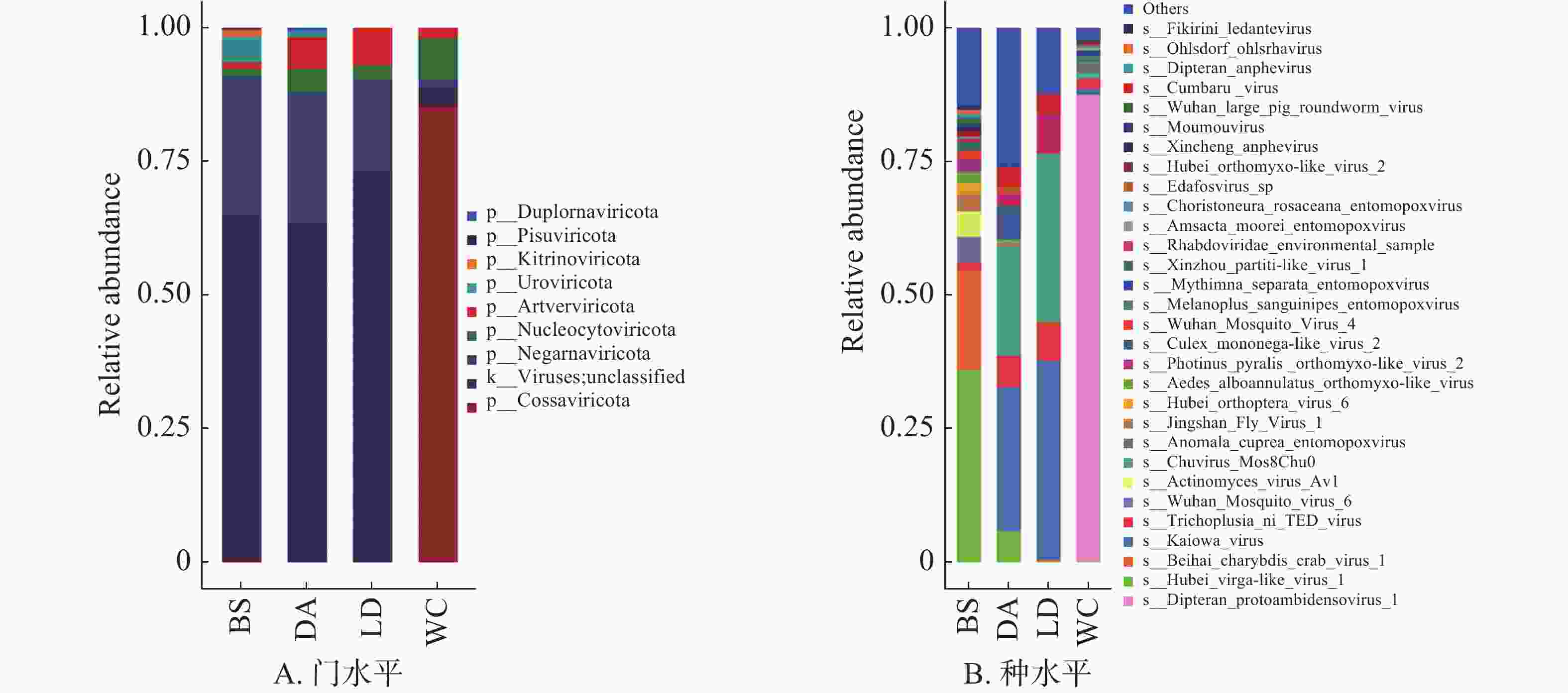
图 6 不同猪场的蚊子携带的病毒不同水平上的相对丰度
Figure 6. Relative abundance of mosquito-borne viruses at different levels in different pig farms
检测出的病毒以蚊作为传播媒介或宿主,感染昆虫、细菌和哺乳动物,如棉褐带卷蛾昆虫痘病毒'L'型(Adoxophyes honmai entomopoxvirus)和摩尔斯毛虫昆虫痘病毒(Amsacta moorei entomopoxvirus)可以感染特定的昆虫,而噬菌体(Actinomyces virus Av1)可以侵入细菌,埃及伊蚊病毒(Aedes aegypti anphevirus)和按蚊环状病毒(Anopheles annulipes orbivirus)可以感染哺乳动物。
-
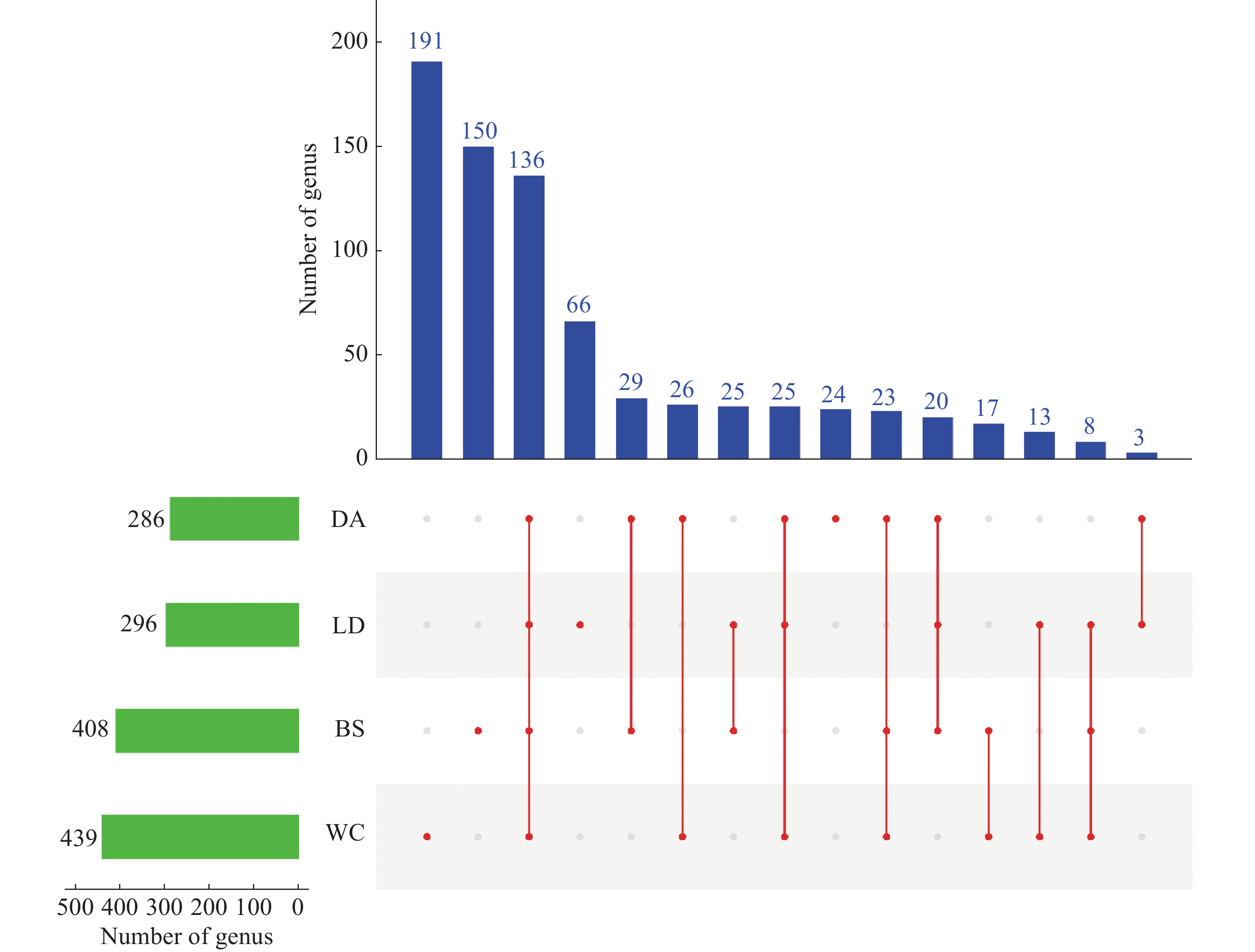
从基于属水平的upset plot图(图7)可以得到,4个样本共含有
1429 属,排名丰富度为WC>BS>LD>DA,4个样本共有136属,BS、DA、LD、WC独有的属数量分别为150、24、66和191。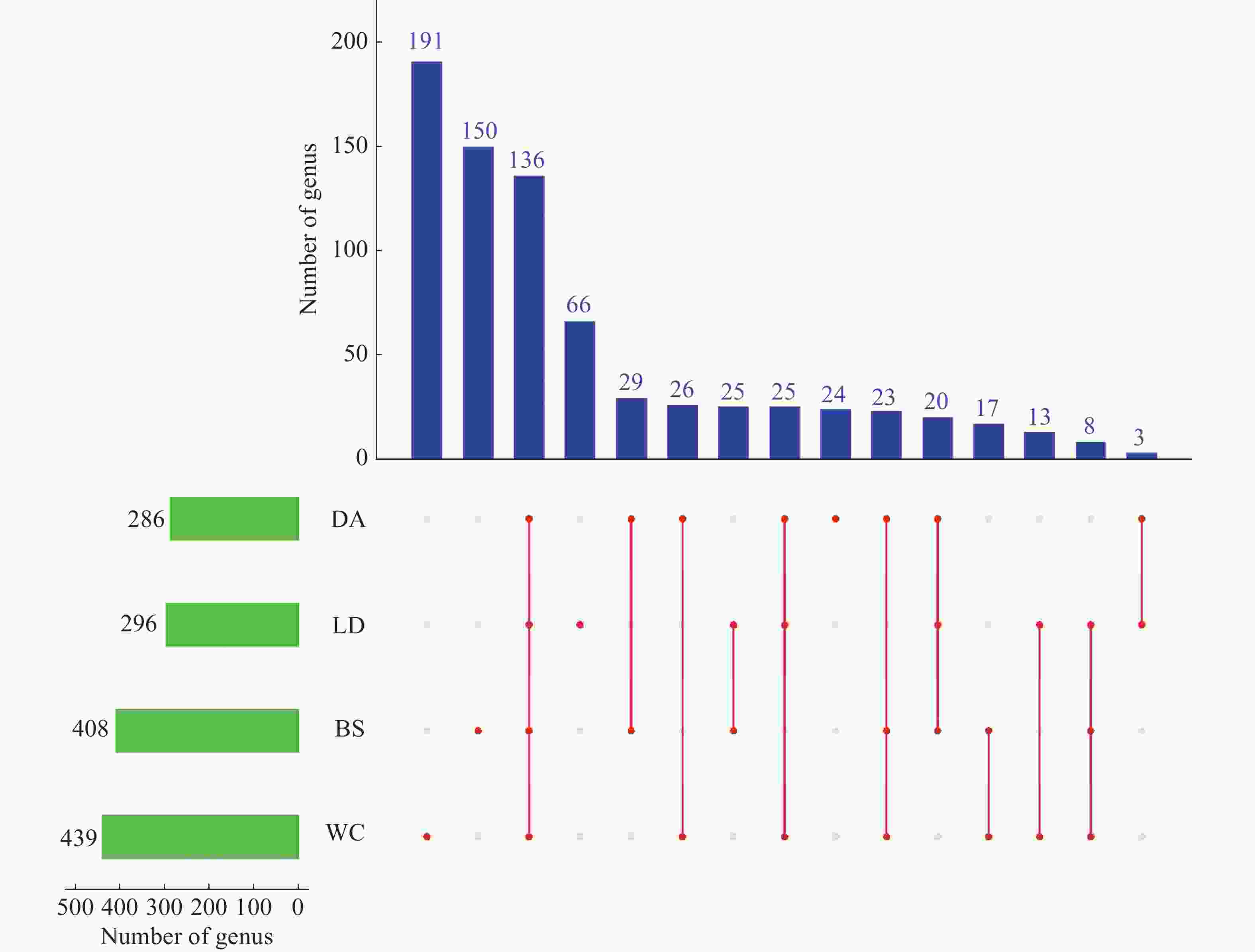
图 7 2024年4个不同市县4个不同猪场的蚊子的微生物多样性分析
Figure 7. Analysis of microbial diversity of mosquitoes in 4 different pig farms in 4 different cities and counties in 2024
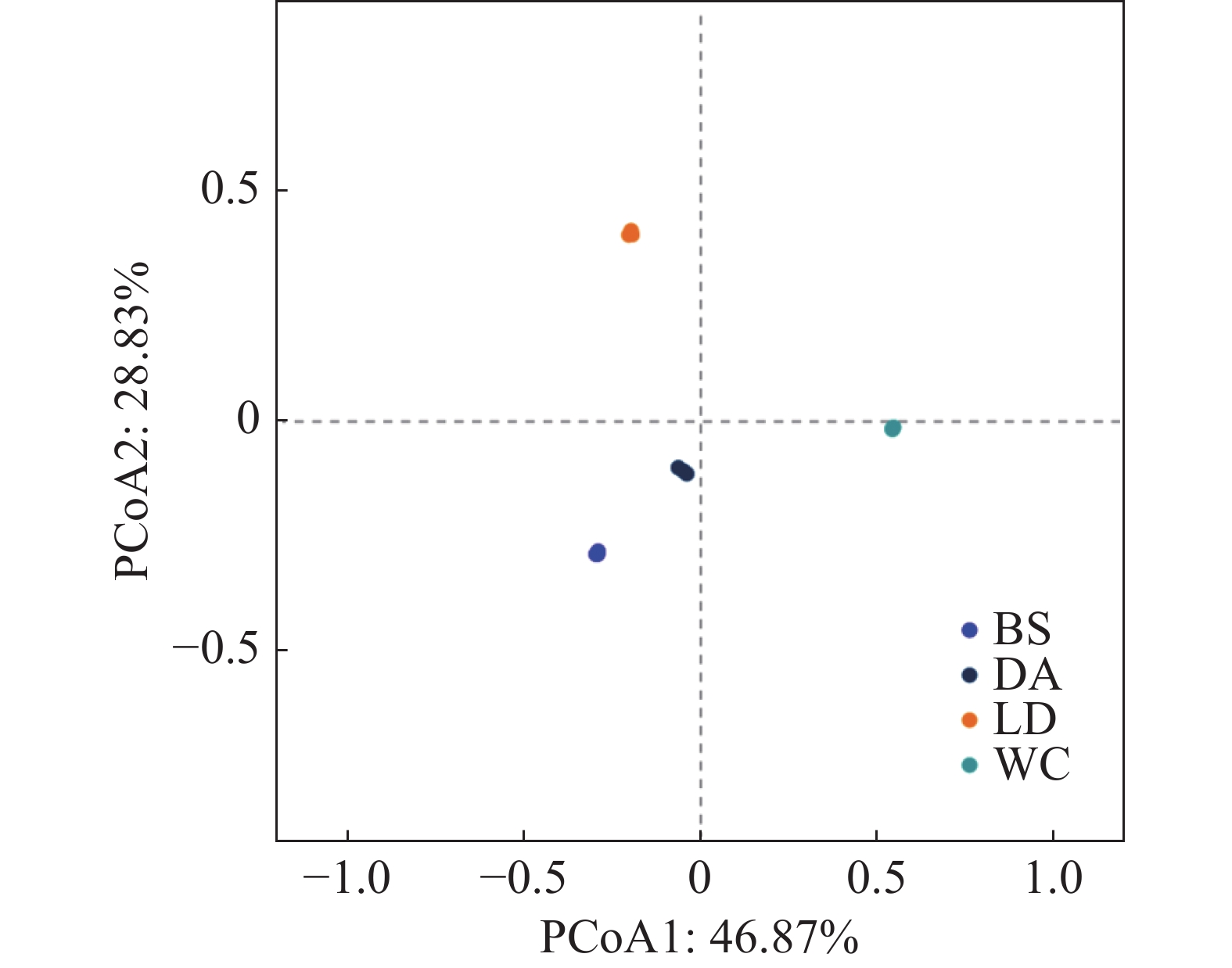
从基于4个样本属水平的主坐标分析可知(见图8),第一主坐标轴(PCoA1)解释了总变异的46.87%,第二主坐标轴(PCoA2)解释了28.83%的变异。分析结果显示,样本BS与样本DA在PCoA图中的空间距离最短,表明它们在微生物群落结构上具有较高的相似性。样本DA与样本WC的空间距离相对较近,暗示它们之间也存在一定程度的相似性。相比之下,样本LD在PCoA图中与其他样本的空间距离最远,表明其微生物群落结构与其他样本存在显著差异。
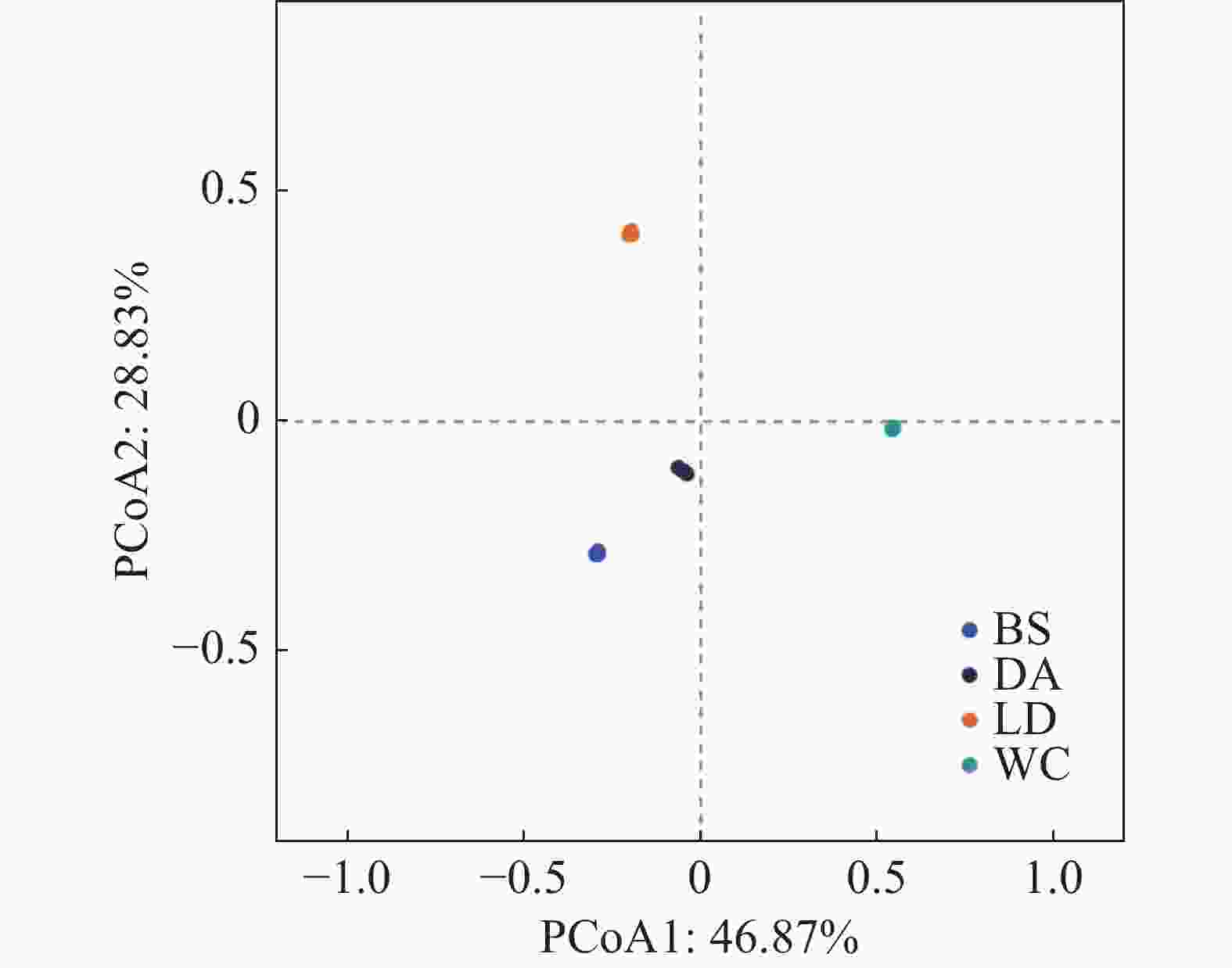
图 8 2024年4个不同市县4个不同猪场的蚊子属水平的主坐标分析
Figure 8. Principal coordinate analysis of mosquitoes at the genus level in the four pig farms in four different cities and counties in 2024
-
将
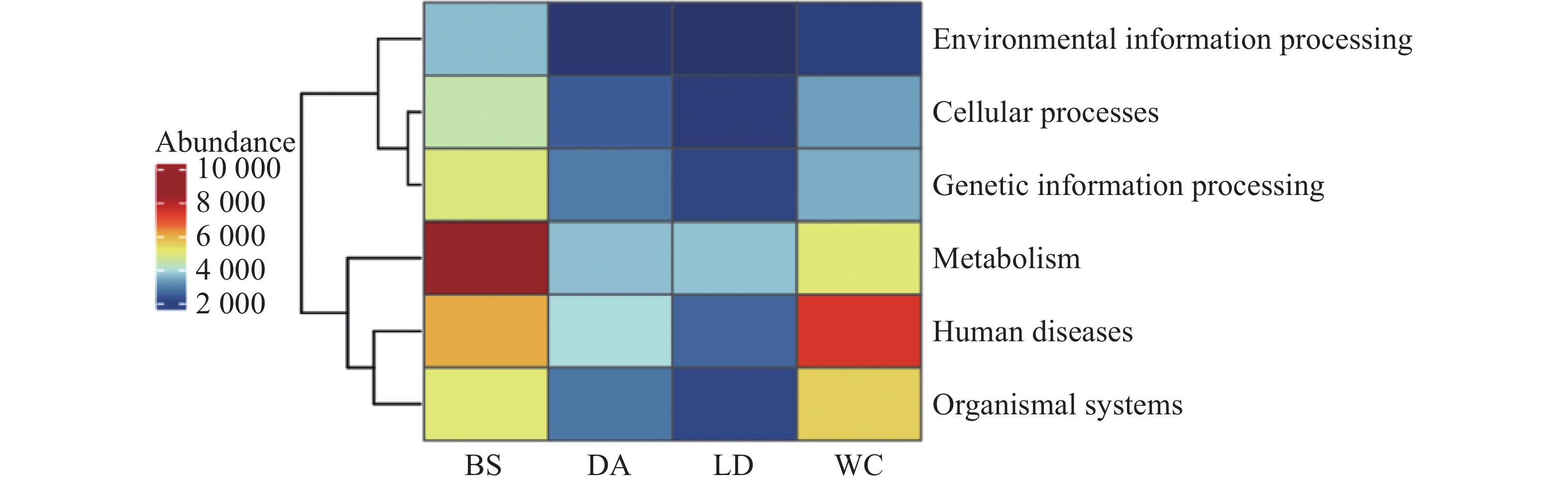
68645 个非冗余基因与KEGG数据库进行比对。KEGG数据库注释到6个一级代谢通路,分别为代谢(Metabolism)、遗传信息处理(Genetic Information Processing)、环境信息处理(Environmental Information Processing)、细胞过程(Cellular Processes)、有机系统(Organismal Systems)和人类疾病(Human Diseases)通路。其中基因功能注释热图(见图9)显示BS除人类疾病通路之外的其余5个一级通路相较于其他样本的相对丰度更高,其中代谢通路在BS中相对丰度最高。而WC的人类疾病通路的相对丰度更高,DA和LD各通路的相对丰度水平类似。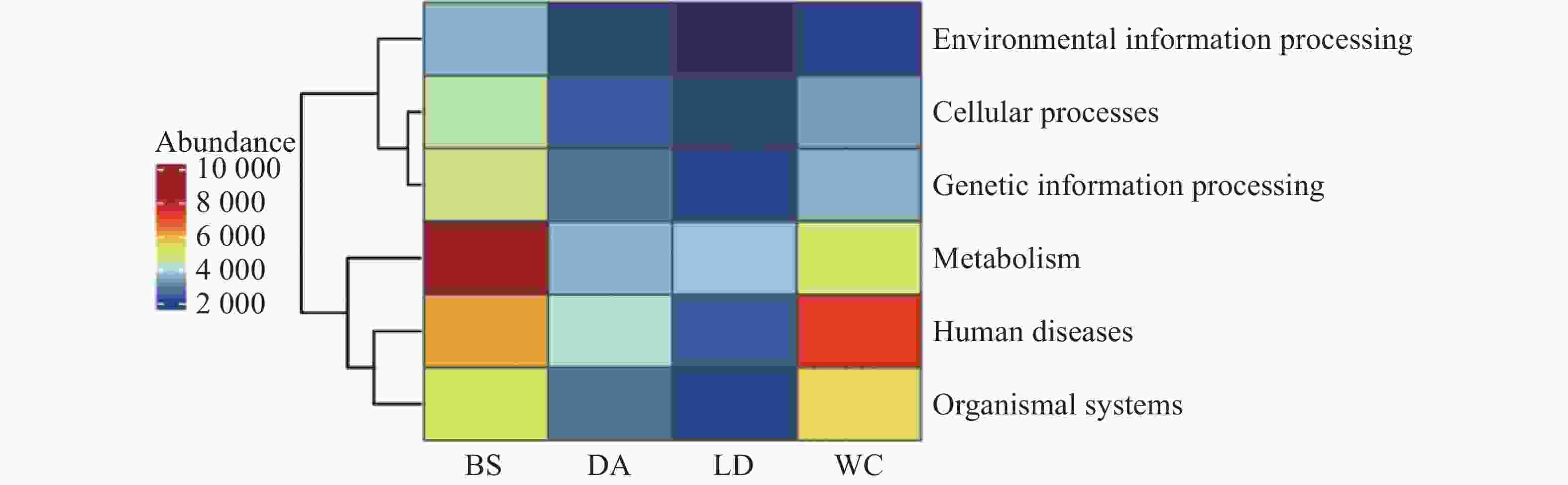
图 9 2024年4个不同市县4个不同猪场的蚊子基因功能注释热图
Figure 9. Heatmap of mosquito gene function annotation in 4 different pig farms in 4 different cities and counties in 2024
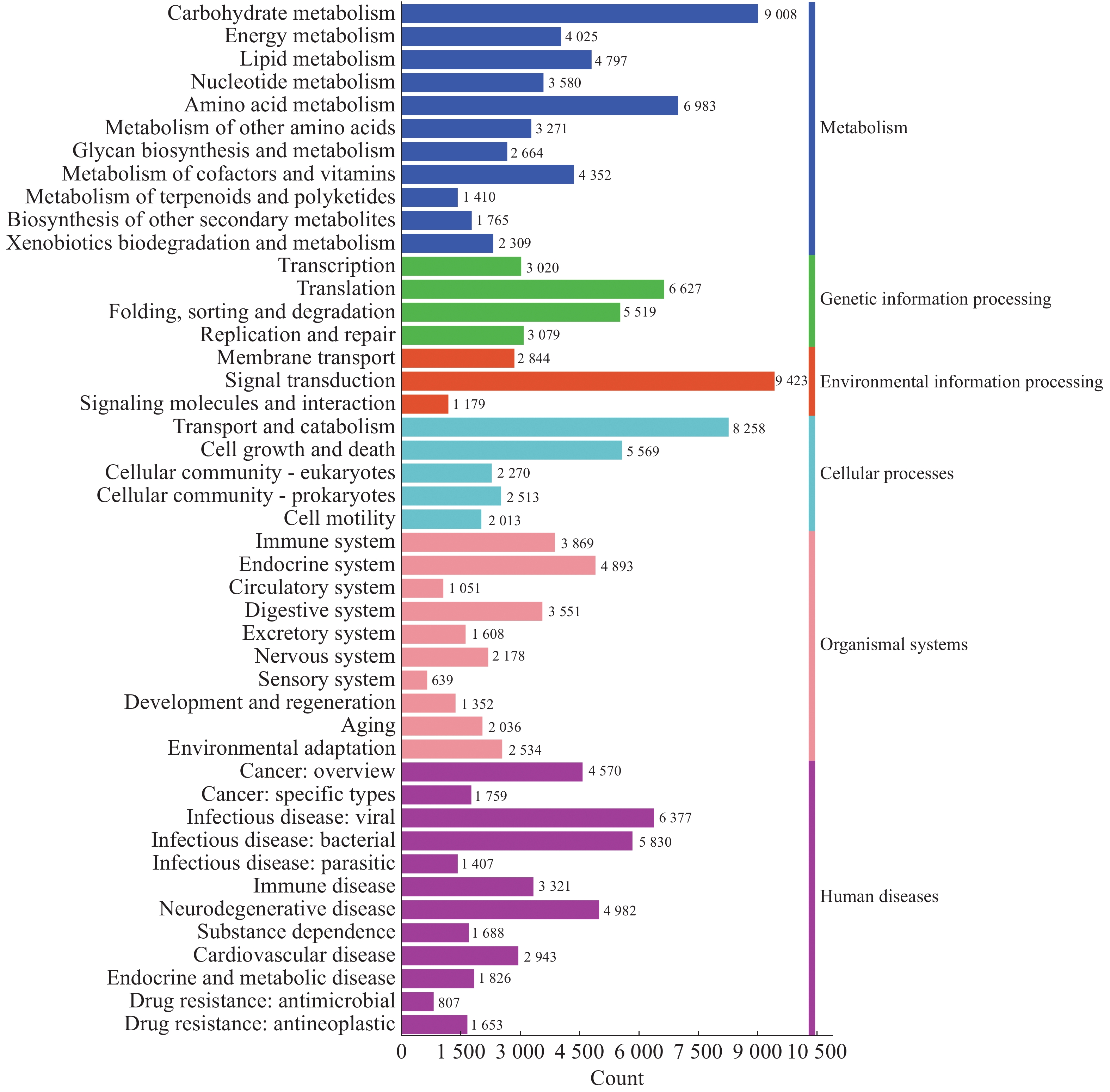
而从KEGG数据库可以注释到的二级通路相对应基因数量图来看(图10),一共可以注释到45个二级通路。在一级通路中,人类疾病通路含有的二级通路最多,为12个;代谢通路含有的基因数目最多,有
44164 个基因;环境信息处理通路含有的二级通路最少,相应的基因数量也最少,分别为3个二级通路和13446 个基因。信号转导(Signal transduction)通路含有的基因数目在二级通路中最多,有9423 个基因;其次为糖代谢通路(Carbohydrate metabolism),有9008 个基因;最少的为感受系统通路(Sensory system),有639个基因。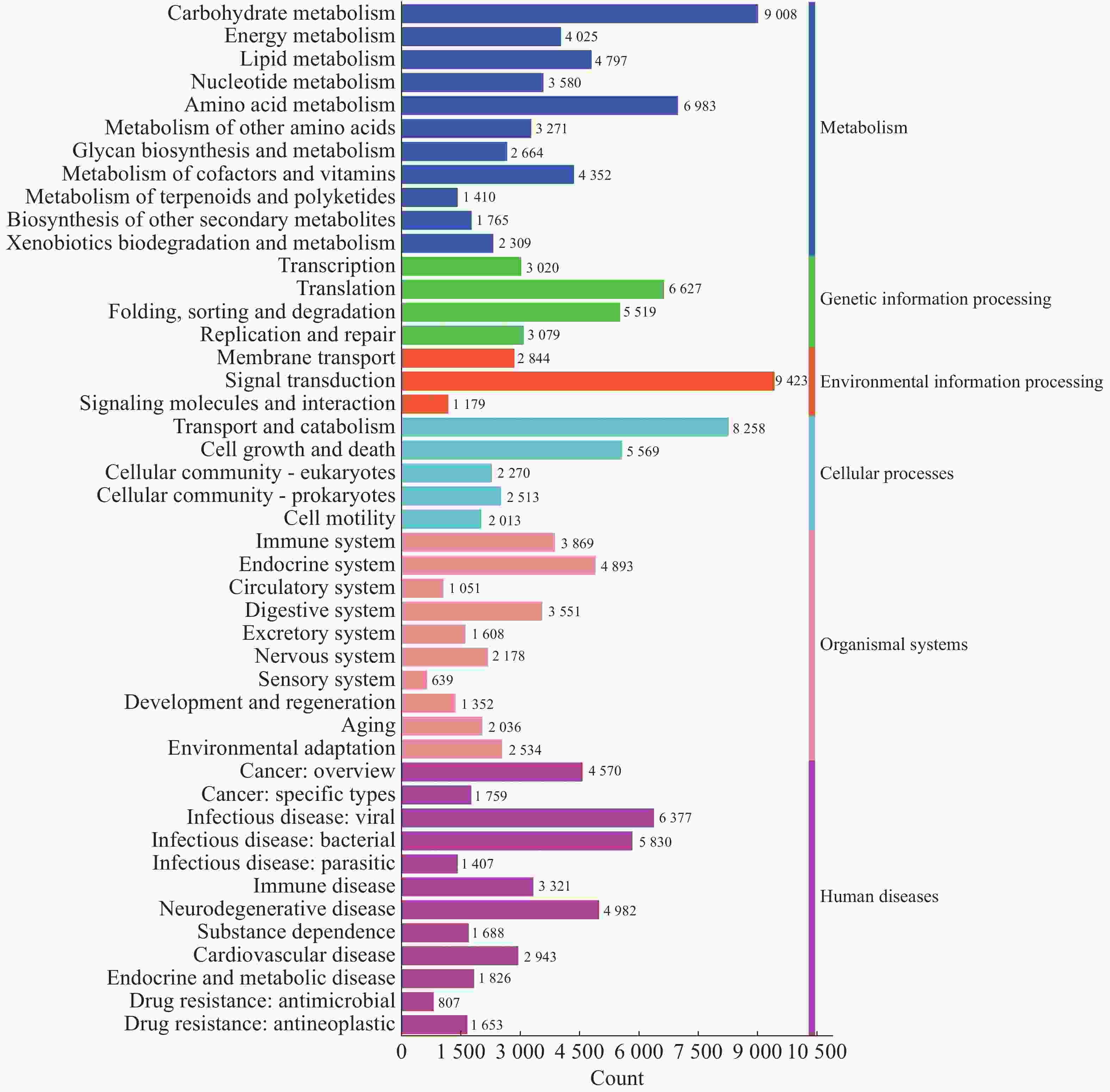
图 10 2024年4个不同市县4个不同猪场的蚊子样本的代谢通路注释基因数目统计
Figure 10. Statistics of the number of annotated genes in the metabolic pathways of mosquito samples from four different pig farms in four different cities and counties in 2024
-
昆虫及其微生物的研究对于理解生态系统、控制害虫、开发新药物等领域至关重要[20]。然而由于蚊子及其共生微生物的数量和种类繁多,且存在难以培养的种类,所以我们采用了宏基因组学的方法探究海南不同地区猪场的蚊子及其微生物。
此次采样的4个地区分别位于海南的东部、西部和中部,东部的文昌和西部的乐东地区处于沿海,中部的定安和白沙地区植被较为茂密,且白沙地区的自然环境更为原始,海拔相对于其他3处更高,所以各个地区的样本具有具有独特的生态特征和生物多样性。在4个不同的猪场优势蚊种各不相同,白沙、定安、乐东和文昌采样猪场的优势蚊种分别为达氏按蚊、中华按蚊、尖音库蚊和三带喙库蚊。
古细菌是一类单细胞微生物,属于古菌域,与细菌域和真核生物域共同构成生物分类学中的三大域[21]。古细菌与细菌在形态上有许多相似之处,如都是单细胞、没有细胞核等[22],但在分子水平上,它们与真核生物更为接近[23]。广古菌门包含了古菌中的大多数种类,它们可以在多种不同的极端环境中生存,包括产甲烷菌、嗜盐古菌、极端嗜热的好氧和厌氧古菌[24]。
变形菌门为样本中的第一大细菌门,其在微生物学中具有重要作用,在污水处理系统中,大多数脱氮细菌隶属于变形菌门,它们在反硝化中起着主导作用[25]。此外,变形菌门中的一些成员能够吞噬细菌和其他有机物[26],显示出其在自然界中的捕食者角色。然而,变形菌门也存在一些潜在的危害。它们中包括一些病原菌,如大肠杆菌、沙门氏菌、志贺氏菌[27]等,这些细菌可以引起人类和动物的疾病。继变形菌门之后的优势细菌门为厚壁菌门,在昆虫的肠道微生物群落中,厚壁菌门是主要的组成部分之一,可以协助宿主昆虫进行营养代谢,提供食物中缺乏的养分,促进昆虫的生长和繁殖[28]。同时厚壁菌门的细菌能通过分泌抗菌肽[29]、毒素等增强昆虫对寄生物的防御能力和抗病性。4个不同样本中含有的毛肠短状螺旋体、流产衣原体、软骨滑液菌、钩端螺旋体和毛肠短状螺旋体对人和动物的健康存在严重威胁,主要表现为流产、不孕[17 − 19]、器官损伤、脑膜炎和致命的出血性疾病等[16],需要重点关注防范,做好疾病预防治疗工作。
子囊菌门是4个样本共有的优势真菌门,其中包含的一些真菌物种是昆虫病原真菌[30],它们能够抑制昆虫种群,这些真菌可以作为生物控制手段,用于控制害虫数量。然而某些真菌在适宜条件下会产生次级代谢产物,即真菌毒素[31],如样本中含有的土曲霉可以产生土曲霉毒素。真菌毒素的产生受到产毒菌自身因素、宿主、环境的影响,产毒菌基因组中存在真菌毒素合成基因簇是真菌毒素产生的根本原因,而环境因素如光照、温度、水活度等可作为环境信号,通过细胞转导途径传递到产毒菌,激活或抑制真菌毒素合成基因簇的表达,从而影响真菌毒素的合成[32],进而对宿主健康产生影响。
负链RNA病毒门为三个样本的共有优势病毒门,科萨特病毒门为WC样本的优势门。对102种病毒分析得知,不同地区的蚊子携带的病毒具有一定的差异性,BS样本中含有病毒数量最多,对人和猪有威胁性的病毒占比也更高,可能的原因是白沙采集地点为高山丛林且养殖模式为放养型。病毒分析提示,白沙和文昌两处猪场蚊子携带病毒致病风险较高。
通过分析可以得知不同的猪场蚊子都有其独特的微生物群落,这可能与蚊子种类和生活环境相关。蚊子种类组成和生存环境越相似,共生微生物群落越相似。
从基因注释数据分析,白沙样本除人类疾病通路之外的其余5个一级通路相较于其他样本的相对丰度更高,其中代谢通路在白沙样本中相对丰度最高。而文昌样本的人类疾病通路的相对丰度更高,定安和乐东各通路的相对丰度水平类似。研究表明,不同的环境对于蚊子共生微生物有较大的影响,同时还会影响其基因表达。根据KEGG数据统计分析,一级通路中的代谢通路为第一大通路,包括碳水化合物代谢、氨基酸代谢、脂质代谢和辅因子和维生素代谢通路等二级通路,这与蚊子的营养获取、能量产生、代谢[33]和自噬-溶酶体信号通路密切相关。这些代谢通路在蚊子的生理过程中发挥着重要作用,影响着它们的生长、繁殖和对病原体的防御能力。一级通路中的第二大通路为人类疾病通路,包括病毒感染、细菌感染、癌症和神经退行性疾病通路等二级通路,而蚊子作为多种疾病的传播媒介,其肠道微生物群与这些疾病的传播密切相关[13]。蚊子通过吸食感染者的血液来获取病原体,而肠道微生物群的组成和稳定性直接影响蚊子对病原体的易感性和传播能力[34]。例如,研究发现肠道菌群通过参与色氨酸代谢,影响蚊子对疟原虫的抵抗力[35]。此外,某些肠道菌如粘质沙雷氏菌(Serratia marcescens)能通过分泌增效因子蛋白增强蚊子对病毒的易感性,从而调控蚊虫传播病毒的能力[36]。这些研究结果表明,蚊子的肠道微生物群不仅参与了其代谢过程,还可能影响人类疾病的传播,这对于蚊媒传染病的防控提供了新的干预靶点。
本研究采用宏基因组学方法探究海南不同地区猪场蚊子及其微生物,发现各地区蚊子样本微生物群落组成独特,与蚊子种类和生态环境密切相关。细菌、真菌和病毒分析表明,存在部分对哺乳动物健康产生危害的物种,需要重点防范。基因注释分析显示,代谢通路和人类疾病通路为蚊子共生微生物的主要功能,影响蚊子的生长、繁殖和疾病传播能力。研究结果为理解蚊子共生微生物群落及其功能提供新见解,对蚊媒传染病防控具有重要意义。
Survey and analysis of mosquito species and microbial metagenomics in the pig farms from different regions of Hainan
-
摘要: 为了解海南省不同地区猪场蚊子种类及其携带病原微生物的情况,为猪场蚊媒疫病的科学防控提供可靠数据支持,本研究使用光触媒诱蚊灯在海南省白沙黎族自治县(BS)、定安县(DA)、乐东黎族自治县(LD)和文昌市(WC)等4个不同市县的猪场采集蚊子,通过形态学和PCR方法鉴定蚊媒种类。再通过宏基因组学测序方法分析蚊媒微生物群落构成及基因功能。结果显示,海南不同地区的猪场优势蚊种各不同,BS、DA、LD和WC的优势蚊种分别为达氏按蚊、中华按蚊、尖音库蚊和三带喙库蚊,占比分别为58.43%、55.46%、88.65%和57.84%。经宏基因组学微生物测序分析,BS、LD、WC等地区的蚊媒微生物在门水平上,广古菌门为优势古细菌门,而DA猪场的蚊媒优势古细菌门则为热古菌门;变形菌门是4地区猪场蚊媒的优势细菌门;子囊菌门为4个地区蚊媒的优势真菌门;BS、DA和LD的优势病毒门均为负链RNA病毒门,而WC的优势病毒门为科萨特病毒门。种水平上,毛肠短状螺旋体(Brachyspira pilosicoli)、布鲁氏菌(Brucella melitensis)、流产衣原体(Chlamydia abortus)、钩端螺旋体(Leptospira interrogans)和软骨滑液菌(Waddlia chondrophila)等细菌、枝顶孢霉(Acremonium asperulatum)、土曲霉(Aspergillus terreus)和犬小孢子菌(Microsporum canis)等真菌及埃及伊蚊病毒(Aedes aegypti anphevirus)和按蚊环状病毒(Anopheles annulipes orbivirus)等病毒对人和猪存在致病威胁。此外,海南不同地区猪场蚊媒的微生物在门水平上保持较高的稳定性,在种水平上呈现多样性,存在较高的致病风险。该研究为海南猪场的蚊媒及其蚊媒疾病防控提供科学依据,同时为评估该地区蚊媒病在公共卫生中的威胁提供了基础数据。Abstract: An attempt was made to understand the mosquito species and their carrying pathogenic microorganisms in pig farms from different regions of Hainan Province, and to provide reliable data support for the scientific prevention and control of mosquito-borne diseases in pig farms. Light catalyst mosquito traps were used to collect mosquitoes from four different pig farms in Baisha Li Autonomous County (BS), Ding 'an County (DA), Ledong Li Autonomous County (LD) and Wenchang City (WC) in Hainan Province. The mosquito species collected were identified by using morphological characteristics and the PCR method, and the composition of mosquito-borne microbial community and gene function were analyzed by using metagenomic sequencing. The results showed that the dominant mosquito species in the pig farms of different regions of Hainan were different. The dominant mosquito species in BS, DA, LD and WC were Anopheles dabryanus, Anopheles sinensis, Culex pipiens pallens and Culex tritaeniorhynchus, accounting for 58.43 %, 55.46 %, 88.65 % and 57.84 %, respectively. The analysis of metagenomic microbial sequencing showed the mosquito-borne microorganisms in BS, LD, WC and other regions were at the phylum level. Euryarchaeota was the dominant archaea, while the dominant archaea in DA pig farm was Thermoarchaeota. Proteobacteria was the dominant phylum of mosquito vectors in the pig farms from the four regions. Ascomycota was the dominant phylum of mosquitoes in the 4 regions; negarnaviricota was the dominant virus phylum in BS, DA and LD, while Cossaviricota was the dominant virus phylum in WC. At the species level bacteria such as Brachyspira pilosicoli, Brucella melitensis, Chlamydia abortus, Leptospira interrogans and Waddlia chondrophila, fungi such as Acremonium asperulatum, Aspergillus terreus and Microsporum canis, and viruses such as Aedes aegypti anphevirus and Anopheles annulipes orbivirus pose a pathogenic threat to humans and pigs. Moreover, the mosquito-borne microorganisms from the pig farms in different regions of Hainan maintain high stability at the phylum level and diversity at the species level, and there is a high risk of pathogenicity. This study provides a scientific basis for the prevention and control of mosquitoes and mosquito-borne diseases in Hainan pig farms, and provides basic data for assessing the threat of mosquito-borne diseases in public health in this area.
-
Key words:
- mosquito /
- macrogene /
- microorganisms /
- pig farms
-
图 2 4个猪场蚊子种类分类
注:图A为白沙(BS)样本;图B为定安(DA)样本;图C为乐东(LD)样本;图D为文昌(WC)样本。
Fig. 2 Classification of mosquito species in 4 pig farms
图 3 不同猪场的蚊子携带的古生菌不同水平上的相对丰度
Fig. 3 Relative abundance of archaea carried by mosquitoes in different pig farms at different levels
图 4 不同猪场的蚊子携带的细菌不同水平上的相对丰度
Fig. 4 Relative abundance of bacteria carried by mosquitoes in different pig farms at different levels
图 5 不同猪场的蚊子携带的真菌不同水平上的相对丰度
Fig. 5 Relative abundance of fungi carried by mosquitoes in different pig farms at different levels
图 6 不同猪场的蚊子携带的病毒不同水平上的相对丰度
Fig. 6 Relative abundance of mosquito-borne viruses at different levels in different pig farms
图 7 2024年4个不同市县4个不同猪场的蚊子的微生物多样性分析
注:BS、DA、LD和WC分别表示白沙、定安、乐东和文昌的蚊子样本。左侧柱状图表示各样本含有属的数量。下方矩阵的单个点代表各样本特有的属,连线表示样本间共有属。右上方柱状图显示不同样本组合中共有属的数量。
Fig. 7 Analysis of microbial diversity of mosquitoes in 4 different pig farms in 4 different cities and counties in 2024
图 8 2024年4个不同市县4个不同猪场的蚊子属水平的主坐标分析
注:BS、DA、LD和WC分别表示白沙、定安、乐东和文昌的蚊子样本。样本点之间的距离越近,表明它们在属水平上的物种组成越相似。轴标签的百分比表示各坐标轴占菌群结构总变异的解释度,这两个轴的解释度越高,表明模型具有更好的效果,能够更好地展示样本间的菌群结构差异。
Fig. 8 Principal coordinate analysis of mosquitoes at the genus level in the four pig farms in four different cities and counties in 2024
图 9 2024年4个不同市县4个不同猪场的蚊子基因功能注释热图
注:BS、DA、LD和WC分别表示白沙、定安、乐东和文昌的蚊子样本。横坐标为各样本,纵坐标为6个一级通路的相对丰度,图中颜色深浅代表相对丰度的大小。
Fig. 9 Heatmap of mosquito gene function annotation in 4 different pig farms in 4 different cities and counties in 2024
图 10 2024年4个不同市县4个不同猪场的蚊子样本的代谢通路注释基因数目统计
注:横坐标代表基因数目,纵坐标代表二级通路名称。右侧图注代表二级通路对应的一级通路名称。
Fig. 10 Statistics of the number of annotated genes in the metabolic pathways of mosquito samples from four different pig farms in four different cities and counties in 2024
表 2 2024年4个不同市县的4个不同猪场宏基因组测序数据
Table 2 Metagenomic sequencing data from four different pig farms in four different cities and counties in 2024
样本名称 原始数据(G) 有效数据(G) 有效数据率/% GC/% Q30/% BS 12.03 11.95 99.33 30.73 97.86 DA 12.06 11.98 99.34 37.51 98.09 LD 11.79 11.70 99.24 37.85 97.88 WC 12.42 12.34 99.36 38.65 97.99 注:BS为白沙采集的样本;DA为定安采集的样本;LD为乐东采集的样本;WC为文昌采集的样本。GC为有效数据中G和C两种碱基占总碱基的百分比;Q30是测序数据中每个碱基的Phred质量分数至少为30(识别错误率不超过0.1%)。  下载: 导出CSV
下载: 导出CSV
-
[1] LIPPI C A, MUNDIS S J, SIPPY R, et al. Trends in mosquito species distribution modeling: insights for vector surveillance and disease control[J]. Parasites & Vectors, 2023, 16(1): 302. doi: 10.1186/s13071-023-05912-z [2] CARRILLO-BUSTAMANTE P, COSTA G, LAMPE L, et al. Evolutionary modelling indicates that mosquito metabolism shapes the life-history strategies of Plasmodium parasites[J]. Nature Communications, 2023, 14(1): 8139. doi: 10.1038/s41467-023-43810-1 [3] GLEAVE K, COOK D, TAYLOR M J, et al. Filarial infection influences mosquito behaviour and fecundity[J]. Scientific Reports, 2016, 6: 36319. doi: 10.1038/srep36319 [4] KRAMBRICH J, AKABERI D, LINDAHL J F, et al. Vector competence of Swedish Culex pipiens mosquitoes for Japanese encephalitis virus[J]. Parasites & Vectors, 2024, 17(1): 220. doi: 10.1186/s13071-024-06269-7 [5] OLMO R P, TODJRO Y M H, AGUIAR E R G R, et al. Mosquito vector competence for dengue is modulated by insect-specific viruses[J]. Nature Microbiology, 2023, 8(1): 135 − 149. doi: 10.1038/s41564-022-01289-4 [6] HIGGS S, VANLANDINGHAM D. Chikungunya virus and its mosquito vectors[J]. Vector-Borne and Zoonotic Diseases, 2015, 15(4): 231 − 240. doi: 10.1089/vbz.2014.1745 [7] ATONI E, ZHAO L, HU C, et al. A dataset of distribution and diversity of mosquito-associated viruses and their mosquito vectors in China[J]. Scientific Data, 2020, 7(1): 342. doi: 10.1038/s41597-020-00687-9 [8] LI Y P, AN Q, SUN Z, et al. Distribution areas and monthly dynamic distribution changes of three Aedes species in China: Aedes aegypti, Aedes albopictus and Aedes vexans[J]. Parasites & Vectors, 2023, 16(1): 297. doi: 10.1186/s13071-023-05924-9 [9] BHUVANESWARI A, SHRIRAM A N, RAJU K H K, et al. Mosquitoes, lymphatic filariasis, and public health: a systematic review of Anopheles and Aedes surveillance strategies[J]. Pathogens, 2023, 12(12): 1406. doi: 10.3390/PATHOGENS12121406 [10] ZHONG R, LIU S L, CHEN S Q, et al. Satellite observations reveal anthropogenic pressure significantly affects the suspended particulate matter concentrations in coastal waters of Hainan Island[J]. Journal of Environmental Management, 2024, 365: 121617. doi: 10.1016/j.jenvman.2024.121617 [11] LI Y J, ZHOU G F, ZHONG S F, et al. Spatial heterogeneity and temporal dynamics of mosquito population density and community structure in Hainan Island, China[J]. Parasites & Vectors, 2020, 13(1): 444. doi: 10.1186/s13071-020-04326-5 [12] STELDER J J, MIHALCA A D, OLESEN A S, et al. Potential mosquito vector attraction to- and feeding preferences for pigs in Romanian backyard farms[J]. Frontiers in Veterinary Science, 2022, 9: 1046263. doi: 10.3389/fvets.2022.1046263 [13] GAO H, CUI C L, WANG L L, et al. Mosquito microbiota and implications for disease control[J]. Trends in Parasitology, 2020, 36(2): 98 − 111. doi: 10.1016/j.pt.2019.12.001 [14] MILANESE A, MENDE D R, PAOLI L, et al. Microbial abundance, activity and population genomic profiling with mOTUs2[J]. Nature Communications, 2019, 10(1): 1014. doi: 10.1038/s41467-019-08844-4 [15] ŞENGÜL DEMIRAK M Ş, CANPOLAT E. Plant-based bioinsecticides for mosquito control: impact on insecticide resistance and disease transmission[J]. Insects, 2022, 13(2): 162. doi: 10.3390/insects13020162 [16] ADLER B, DE LA PEÑA MOCTEZUMA A. Leptospira and leptospirosis[J]. Veterinary Microbiology, 2010, 140(3/4): 287 − 296. doi: 10.1016/j.vetmic.2009.03.012 [17] GŁOWACKA P, ŻAKOWSKA D, NAYLOR K, et al. Brucella – virulence factors, pathogenesis and treatment[J]. Polish Journal of Microbiology, 2018, 67(2): 151 − 161. doi: 10.21307/pjm-2018-029 [18] WEN Y, DU Y J, SHI X Y, et al. Clinical diagnosis and treatment of seven patients diagnosed pneumonia caused by Chlamydia abortus: a case series report[J]. Frontiers in Medicine, 2024, 11: 1406737. doi: 10.3389/fmed.2024.1406737 [19] CORSARO D, VENDITTI D. Emerging chlamydial infections[J]. Critical Reviews in Microbiology, 2004, 30(2): 75 − 106. doi: 10.1080/10408410490435106 [20] ISLAM W, ADNAN M, SHABBIR A, et al. Insect-fungal-interactions: a detailed review on entomopathogenic fungi pathogenicity to combat insect pests[J]. Microbial Pathogenesis, 2021, 159: 105122. doi: 10.1016/j.micpath.2021.105122 [21] SPANG A, CACERES E F, ETTEMA T J G. Genomic exploration of the diversity, ecology, and evolution of the archaeal domain of life[J]. Science, 2017, 357(6351): eaaf3883. doi: 10.1126/science.aaf3883 [22] ZILLIG W. Comparative biochemistry of Archaea and Bacteria[J]. Current Opinion in Genetics & Development, 1991, 1(4): 544 − 551. doi: 10.1016/S0959-437X(05)80206-0 [23] FORTERRE P. Archaea: a goldmine for molecular biologists and evolutionists[M]//FERREIRA-CERCA S. Archaea: Methods and Protocols. New York: Humana, 2022: 1-21. doi: 10.1007/978-1-0716-2445-6_1 [24] ESSER D, HOFFMANN L, PHAM T K, et al. Protein phosphorylation and its role in archaeal signal transduction[J]. FEMS Microbiology Reviews, 2016, 40(5): 625 − 647. doi: 10.1093/femsre/fuw020 [25] BROMAN E, ZILIUS M, SAMUILOVIENE A, et al. Active DNRA and denitrification in oxic hypereutrophic waters[J]. Water Research, 2021, 194: 116954. doi: 10.1016/j.watres.2021.116954 [26] WU X Q, WU L Y, LIU Y N, et al. Microbial interactions with dissolved organic matter drive carbon dynamics and community succession[J]. Frontiers in Microbiology, 2018, 9: 1234. doi: 10.3389/fmicb.2018.01234 [27] BAUMGART S, PHAN T, MCKEW G. Epidemiology and antimicrobial resistance rates for Shigella species in a resource-rich setting[J]. Pathology, 2025, 57(1): 94 − 99. doi: 10.1016/J.PATHOL.2024.07.004 [28] LI H W, ZHAO C W, YANG Y, et al. The influence of gut microbiota on the fecundity of Henosepilachna vigintioctopunctata (Coleoptera: Coccinellidae)[J]. Journal of Insect Science, 2021, 21(4): 15. doi: 10.1093/jisesa/ieab061 [29] GANESAN N, MISHRA B, FELIX L, et al. Antimicrobial peptides and small molecules targeting the cell membrane of Staphylococcus aureus[J]. Microbiology and Molecular Biology Reviews, 2023, 87(2): e0003722. doi: 10.1128/mmbr.00037-22 [30] BRANINE M, BAZZICALUPO A, BRANCO S. Biology and applications of endophytic insect-pathogenic fungi[J]. PLoS Pathogens, 2019, 15(7): e1007831. doi: 10.1371/journal.ppat.1007831 [31] LIEW W P P, MOHD-REDZWAN S. Mycotoxin: its impact on gut health and microbiota[J]. Frontiers in Cellular and Infection Microbiology, 2018, 8: 60. doi: 10.3389/fcimb.2018.00060 [32] CACERES I, KHOURY A A, KHOURY R E, et al. Aflatoxin biosynthesis and genetic regulation: a review[J]. Toxins, 2020, 12(3): 150. doi: 10.3390/toxins12030150 [33] RIVERA-PÉREZ C, CLIFTON M E, NORIEGA F G. How micronutrients influence the physiology of mosquitoes[J]. Current Opinion in Insect Science, 2017, 23: 112 − 117. doi: 10.1016/j.cois.2017.07.002 [34] DENNISON N J, JUPATANAKUL N, DIMOPOULOS G. The mosquito microbiota influences vector competence for human pathogens[J]. Current Opinion in Insect Science, 2014, 3: 6 − 13. doi: 10.1016/j.cois.2014.07.004 [35] FENG Y B, PENG Y Q, SONG X M, et al. Anopheline mosquitoes are protected against parasite infection by tryptophan catabolism in gut microbiota[J]. Nature Microbiology, 2022, 7(5): 707 − 715. doi: 10.1038/s41564-022-01099-8 [36] WU P, SUN P, NIE K X, et al. A gut commensal bacterium promotes mosquito permissiveness to arboviruses[J]. Cell Host & Microbe, 2019, 25(1): 101-112. e5. doi: 10.1016/j.chom.2018.11.004 -

 点击查看大图
点击查看大图
计量
- 文章访问数: 784
- HTML全文浏览量: 639
- 被引次数: 0
