

-
种群的结构和动态是种群生态学研究的核心内容,种群结构反映了种群内不同类型个体的数量、大小特征,种群动态则反映了种群的数量动态和变化趋势[1 − 2],通常使用年龄结构、生存曲线、死亡曲线和生命表等形式开展种群的结构和动态研究,相关研究可以为种群保护提供理论依据。众多学者针对种群结构和数量动态开展了相关研究,如张金蜂等[3]通过年龄结构对长白山红松(Pinus koraiensis)种群进行研究,结果表明红松种群属于增长型种群,但幼苗死亡率高,应加强幼苗的抚育;张维伟等[4]根据黄土高原南部3种类型的麻栎林中的4种乔木种群的年龄结构预测其在20~50年后的发展趋势,提出麻栎林经营应以顺应自然演替规律的近自然经营方式为主的建议;夏丹等[5]通过编制静态生命表、存活曲线分析和生存分析等方式研究海南卷萼兜兰(Paphiopedilum appletonianum)的种群结构和数量动态变化,提出了以增加种群个体数量和扩大种群分布范围为目标的两种保护方式;林春惠等[6]通过编制静态生命表和绘制生存曲线、生存力函数曲线的方式,对广东省紫金县的两个大苞山茶(Camellia granthamiana)种群开展了种群结构和动态特征的相关研究,结果表明大苞山茶种群幼龄个体死亡率高,但仍有一定的更新潜力,建议加强幼龄个体的保护和减少人为的干扰。
海南是中国最南端的茶叶产区,出产的茶享有“华夏第一早春茶”的美誉,是世界优质大叶茶的重要产地[7]。野茶是栽培茶种质资源的重要来源,海南分布的野茶初步认定为普洱茶[8](Camellia sinensis var. assamica),广泛分布于五指山、琼中、昌江、白沙等中西部山区的热带雨林中,由于分布的广泛性和生境的复杂性,使得海南成为重要的茶树种质资源库。针对于海南野茶的研究,目前大多关注群落特征、分类地位和品质鉴定方面,如朱子丞等[9]针对鹦哥岭的野茶进行了群落物种组成和竞争关系的调查研究,苏凡等[10]基于形态学特征和psbA-trnH叶绿体编码基因序列明确了五指山野茶的分类地位,刘肖冰等[11]和苏凡等[12]针对五指山野茶的生理生化成分和营养保健功能进行了相关研究。而针对海南野茶种群的相关研究,最早可追溯到20世纪90年代郭远安[13]对海南的野茶进行的调查,当时发现野茶分布范围广且资源十分丰富。但受到人为生产活动的影响,海南野茶的生境呈现破碎化,分布范围和种群数量不断减少[14],目前已被列为国家Ⅱ级保护植物。李苑菱等[15]于2013—2015年对海南岛的野茶进行了分布特征和种群结构的调查研究,发现野茶为聚集分布,呈稳定型种群结构。海南热带雨林国家公园于2021年正式设立,在其核心区内分布着大量的野茶,野茶不仅是当地热带雨林食物链中重要的一环,对雨林生态系统完整性起着重要作用,同时还是当地村民栽培茶树种质资源的重要来源,野茶叶也是当地村民的收入来源之一。基于野茶资源的重要性,探明保护区不同区域野茶的种群现状和动态变化,对于针对性地开展野茶种群保护具有重要意义。
-
海南热带雨林国家公园位于海南岛中南部山区,是我国首批正式设立的5个国家公园之一,其东起吊罗山,西至尖峰岭,南自保亭毛感乡,北至黎母山,地理位置为18°33′16″—19°14′16″N,108°44′32″—110°04′43″E。海南热带雨林国家公园范围内共涵盖了五指山、琼中、白沙和昌江等9个市县,总面积达4 269 km2,其中核心保护区面积2 331 km2,占国家公园总面积的54.6%,一般控制区面积1 938 km2,占国家公园总面积的45.4%。海南热带雨林是中国最集中、类型最多样、保存最完好且连片面积最大的大陆性岛屿型热带雨林,包含了鹦哥岭、霸王岭、黎母山、吊罗山等著名山体,不仅是热带生物多样性保护的关键地区,也是全球热带雨林的重要组成部分和全球重要的种质资源基因库[16]。同时,它还是海南省水源保护的重点关注地[17]。海南热带雨林国家公园地处热带,热量条件优渥;多年平均降雨量为1 759 mm,雨量充沛。保护区内的植被类型包括了山地雨林、低地雨林和低地雨林次生林等,土壤有明显的垂直地带性,随海拔升高由砖红壤逐步过渡为赤红壤、黄壤和草甸土[18]。
-
基于海南野茶分布范围较广的特性,本研究采用样方法:于2023年4—12月在海南热带雨林国家公园内共设置了36个20 m×20 m的群落样方(其中鹦哥岭24个、霸王岭5个、黎母山4个、吊罗山3个),调查记录样方内的所有野茶个体。其中,胸径(DBH)≥1 cm的野茶个体记录树高、胸径,胸径小于1 cm的野茶个体记录数量。参考李苑菱等[15]对海南岛野茶的径级划分,将野茶的径级分为6级,Ⅰ级为幼苗(DBH<1 cm)、Ⅱ级为幼树(1≤DBH<5 cm);Ⅲ级(5 cm≤DBH<10 cm)和Ⅳ级(10 cm≤DBH<15 cm)为中树的两个阶段;Ⅴ级为大树(15 cm≤DBH<20 cm);Ⅵ级为老树(DBH≥20 cm)。
-
对海南热带雨林国家公园整体及各个区域的野茶种群进行径级结构的统计分析,绘制径级结构金字塔,计算存活率、死亡率、生命期望和消失率,绘制静态生命表、存活率与累计死亡率曲线、死亡率和消失率曲线。
存活率表示存活的程度,计算公式:
$$ {s} _{ {x} } = {l} _{ {x+1} } / {l} _{ {x} {。}} $$ (1) 死亡率表示死亡的程度,计算公式:
$$ {q} _{ {x} } = {d} _{ {x+1} } {/} {d} _{ {x} {。}} $$ (2) 生命期望表示x径级的个体在未来所能存活的平均年数期望,计算公式:
$$ {e} _{ {x} } = {T} _{ {x} } {/} {l} _{ {x} {。}} $$ (3) 消失率表示各龄级内的消失率,计算公式:
$$ {K} _{ {x} } =\ln {l} _{ {x} } -\ln{l} _{ {x+1} {。}} $$ (4) 其中,lx为野茶种群在x径级时的个体数;dx为野茶种群在x径级时的死亡个体数;Tx为从x径级开始,超过x径级的存活下来的个体数之和,Tx=∑(lx+lx)/2。
-
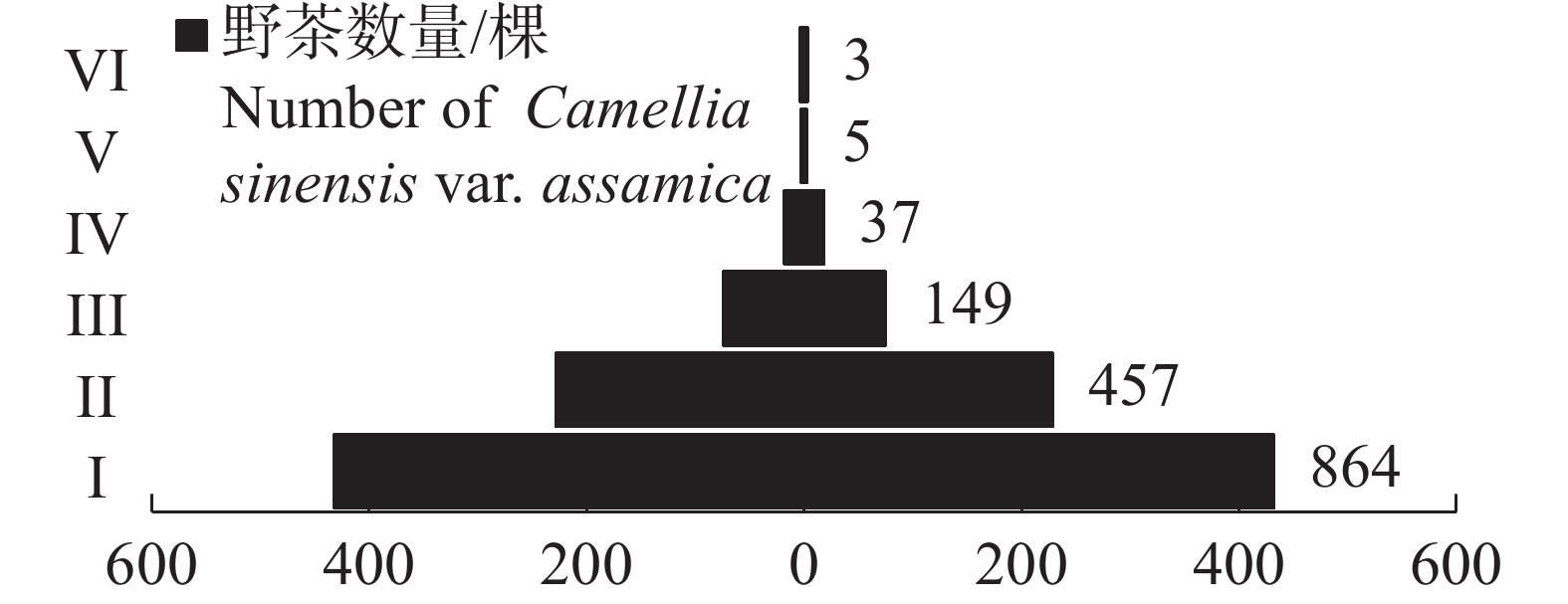
经调查,在海南热带雨林国家公园设置的36个样方中共记录到
1515 棵野茶(图1)。其中,幼苗达864棵,占比最大,为57.03%;其次为幼树,共记录到457棵,占比为30.17%;中树记录到149棵,占比9.83%;老树仅记录到3棵,占比最小,为0.02%。野茶种群的径级结构呈现典型的上尖下宽的金字塔结构,非繁殖期个体占比最大,繁殖期个体占比适中,繁殖后期个体占比较小,为典型的增长型种群。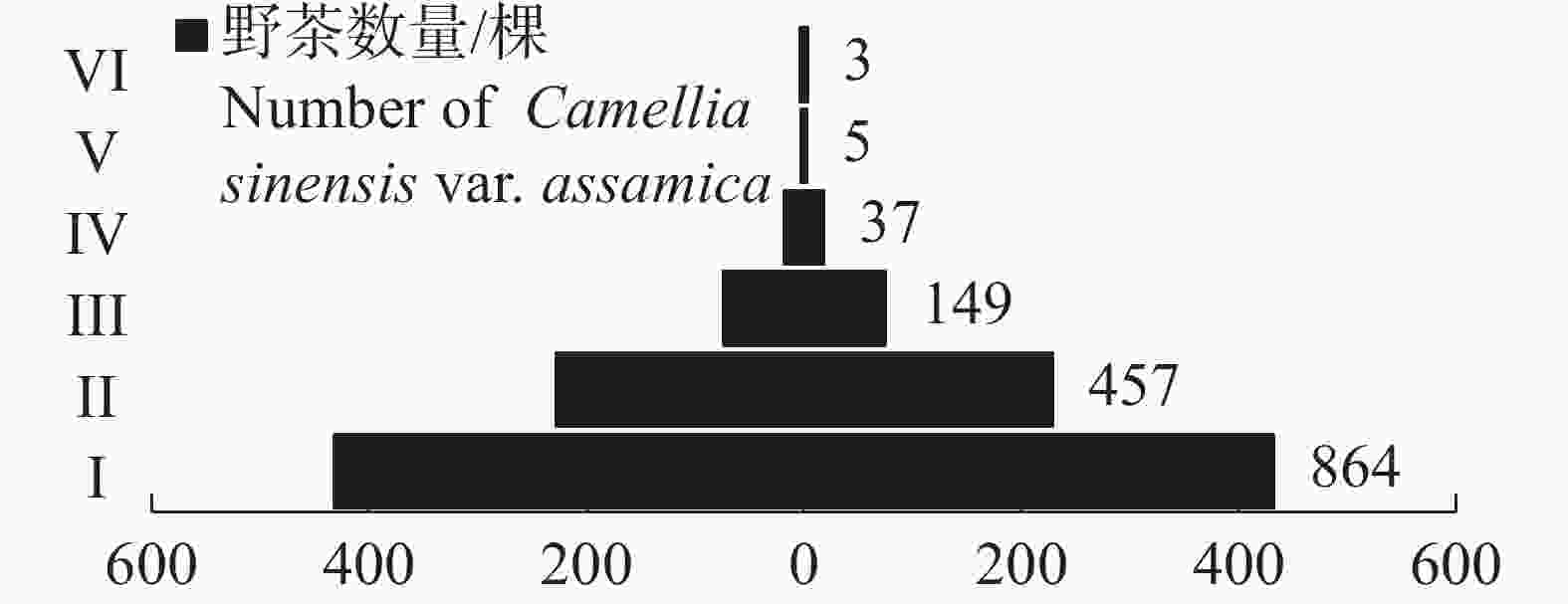
图 1 海南热带雨林国家公园野茶种群的径级结构
Figure 1. Diameter class structure of Camellia sinensis var. assamica populations in Hainan Tropical Rainforest National Park
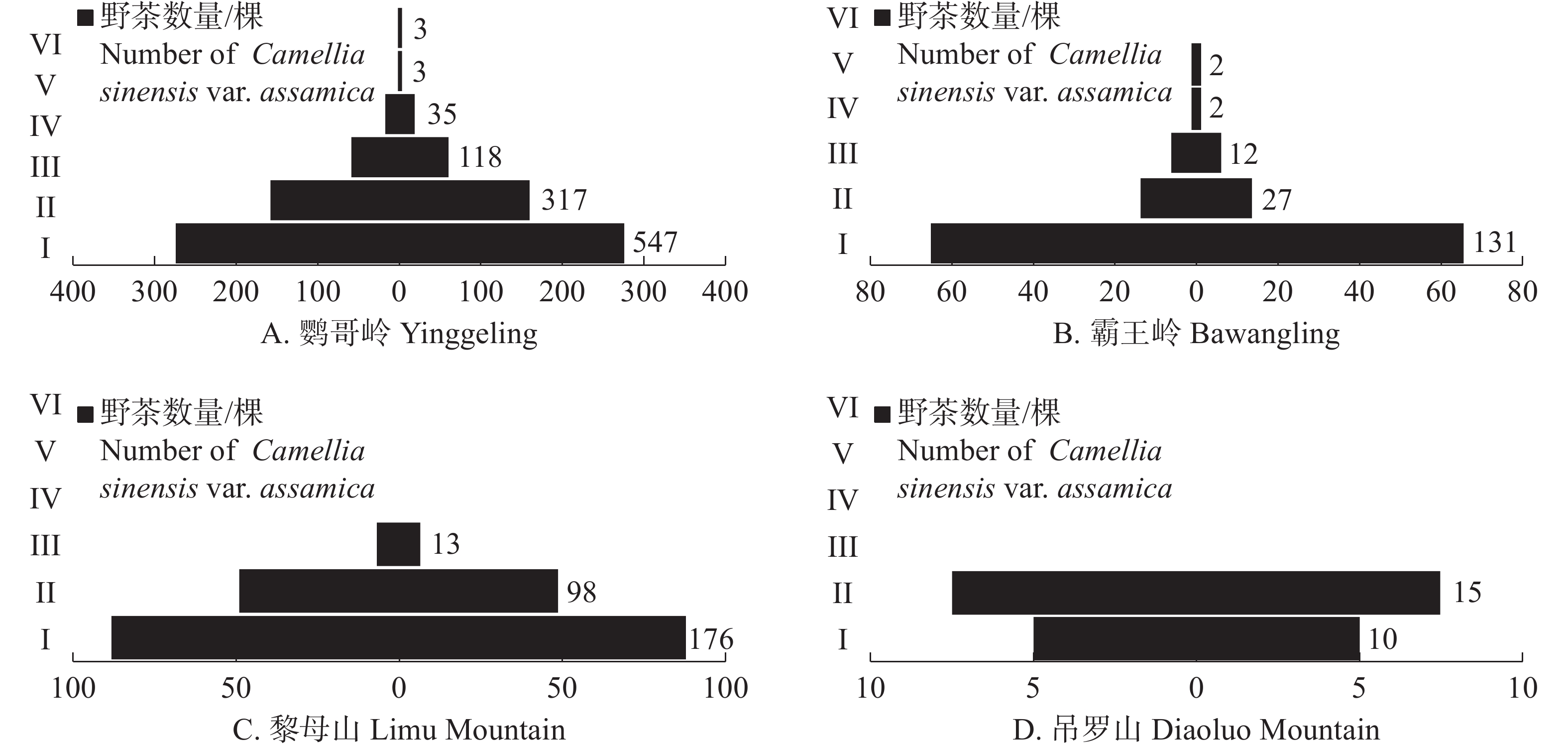
从海南热带雨林国家公园不同区域的野茶种群径级结构上看(图2),鹦哥岭为径级结构最完整的区域,6个径级皆有分布;吊罗山为径级结构最不完整的区域,仅记录到了Ⅰ、Ⅱ两个径级的野茶个体。鹦哥岭、霸王岭野茶种群径级结构为上尖下宽的增长型种群(图2−A,B),表现为野茶幼苗最多,主体为小胸径的野茶个体,中树、大树占据一定比例;黎母山野茶种群幼苗较多,但其集中在Ⅰ、Ⅱ、Ⅲ 3个径级,虽然为增长型种群,但缺乏大树和老树个体,径级结构不完整(图2−C);吊罗山野茶种群幼树最多,未记录到中树及以上径级,且幼苗相对较少(图2−D)。
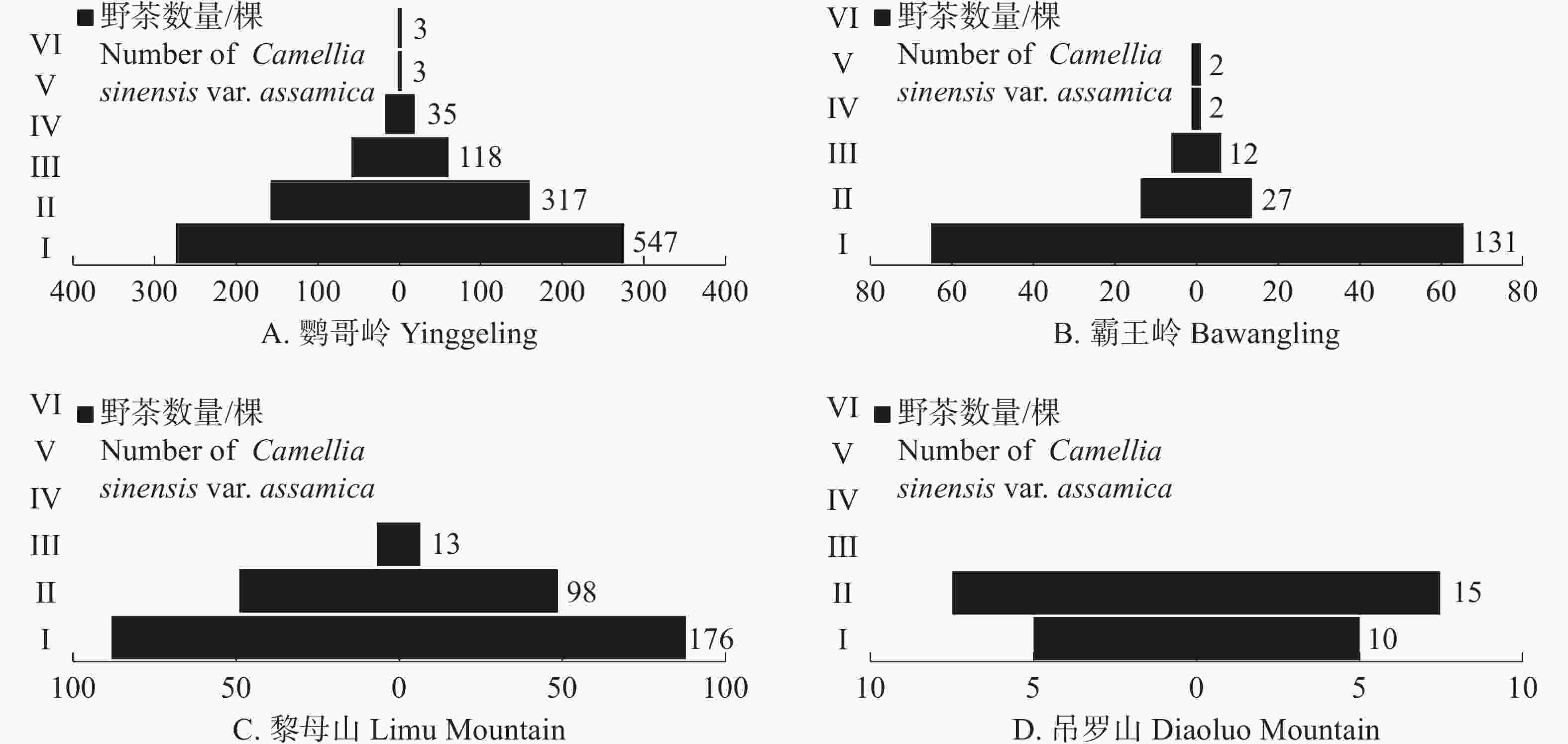
图 2 不用区域的野茶种群径级结构
Figure 2. Trail structure of Camellia sinensis var. assamica populations in different regions
从不同区域的径级分布上看,霸王岭和黎母山的茶种群虽然能萌发大量幼苗,但是在往后生长的过程中,幼苗大量死亡,应加强对于其野茶幼苗的抚育。而鹦哥岭的幼苗同样较多,但是各径级的数量相对稳定。吊罗山的径级结构中,大径级野茶个体和幼苗都较少,除了加强野茶幼苗的抚育外,还得加强幼树的保护,确保其能向大胸径野茶个体转化,促进其种群更新。野茶个体在大树、老树阶段的生存力较弱,容易遭受病虫害,应加强对大树、老树个体的保护。
-
从表1可以看出,海南热带雨林国家公园中的野茶种群各径级的个体数差别较大,Ⅰ径级野茶幼苗数量多,占比大,种群具有较大的增长潜力。随着径级的不断增大,存活率sx不断下降。野茶存活率在Ⅰ到Ⅱ径级的过程下降最快,变幅达到20.29%,说明在种群发展的过程中有许多幼苗被淘汰。生命期望值反映了各径级的平均期望的生存能力,在Ⅰ到Ⅳ径级生命期望值呈现下降趋势,而到了V阶段后有所上升,这说明野茶个体进入大树期后,生存力增强。
表 1 野茶种群的静态生命表
Table 1. Static life table of Camellia sinensis var. assamica population
x lx sx/% dx qx/% Tx ex Kx Ⅰ 864 52.89 407 47.11 757.5 0.88 0.64 Ⅱ 457 32.60 308 67.40 325.5 0.71 1.12 Ⅲ 149 24.83 115 77.18 97 0.65 1.39 Ⅳ 34 13.51 29 86.49 22.5 0.60 2.00 Ⅴ 5 60.00 2 40.00 4 0.80 0.51 Ⅵ 3 — — — — — — 注:x为野茶径级,共分6级;lx为野茶种群在x径级时的存活数;sx为野茶种群的存活率;dx为x径级到x+1径级过渡时死亡的野茶个体数;qx为野茶种群从x径级到x+1径级过渡时的死亡率;Tx为从x径级开始,超过x径级的存活下来的个体数之和;ex为生命期望,为x径级的个体在未来所能存活的平均年数期望;Kx为消失率;“—”表示无。 Note: x is the diameter class of Camellia sinensis var. assamica, divided into 6 levels; lx is the number of individuals in the Camellia sinensis var. assamica population at diameter class x; sx is the survival rate of the Camellia sinensis var. assamica population; dx is the number of Camellia sinensis var. assamica individuals that died at the transition from diameter class x to x +1; qx is the mortality rate of the Camellia sinensis var. assamica population at the transition from diameter class x to x +1; Tx is the sum of the number of individuals that survived beyond diameter class x from diameter class x onward; ex is the life expectancy, which is the i expectation of the average number of years that individuals of diameter class x will survive in the future; Kx is the disappearance rate;“—” means none. -
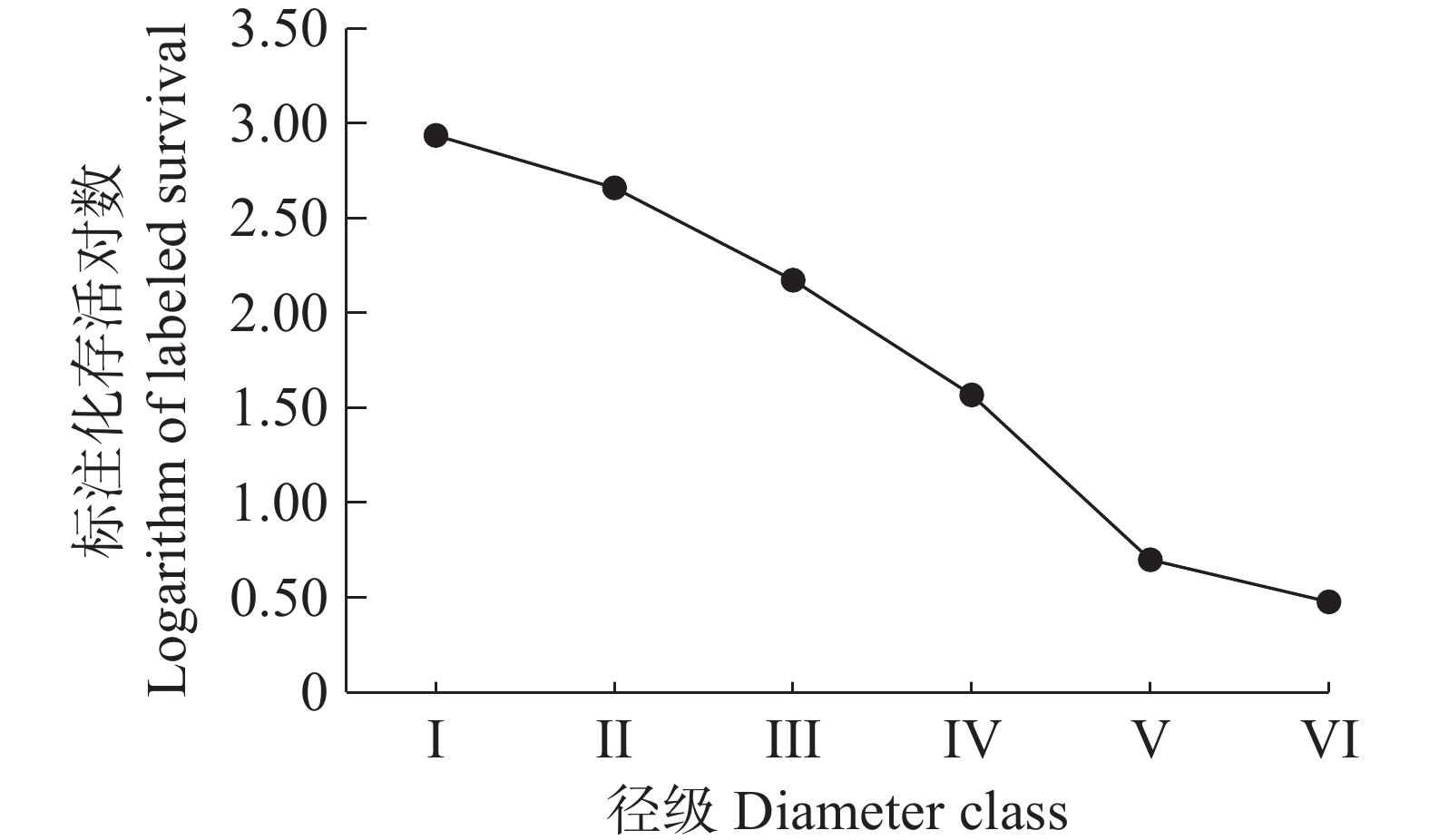
从野茶种群的存活曲线图(图3)可以看出,野茶种群的存活曲线接近于Ⅰ型,野茶幼苗的存活率较高,大树个体的死亡率较高,各径级的个体都能存活到生理年龄。
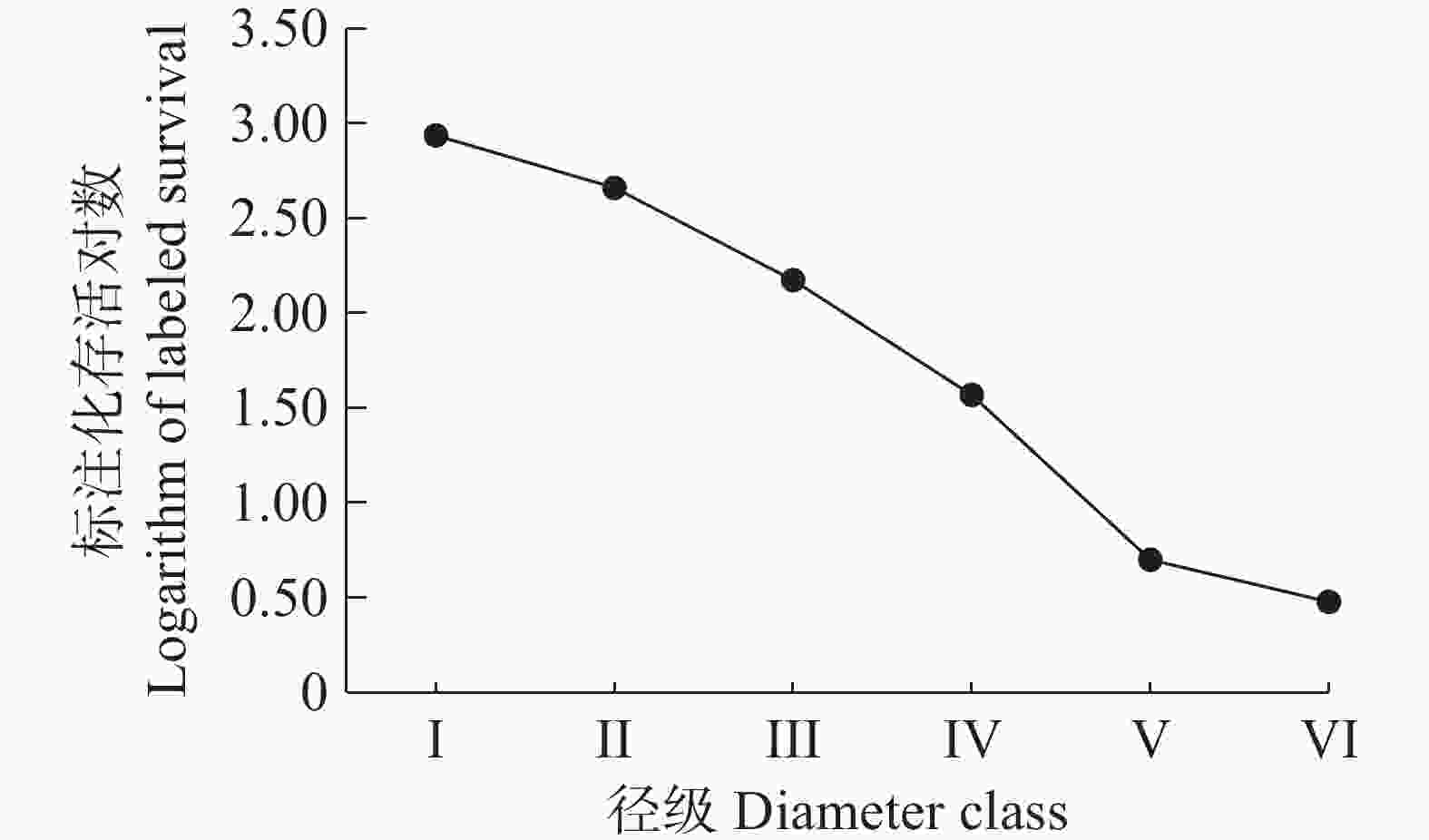
图 3 野茶种群的存活曲线图
Figure 3. Survival curves of Camellia sinensis var. assamica populations
-
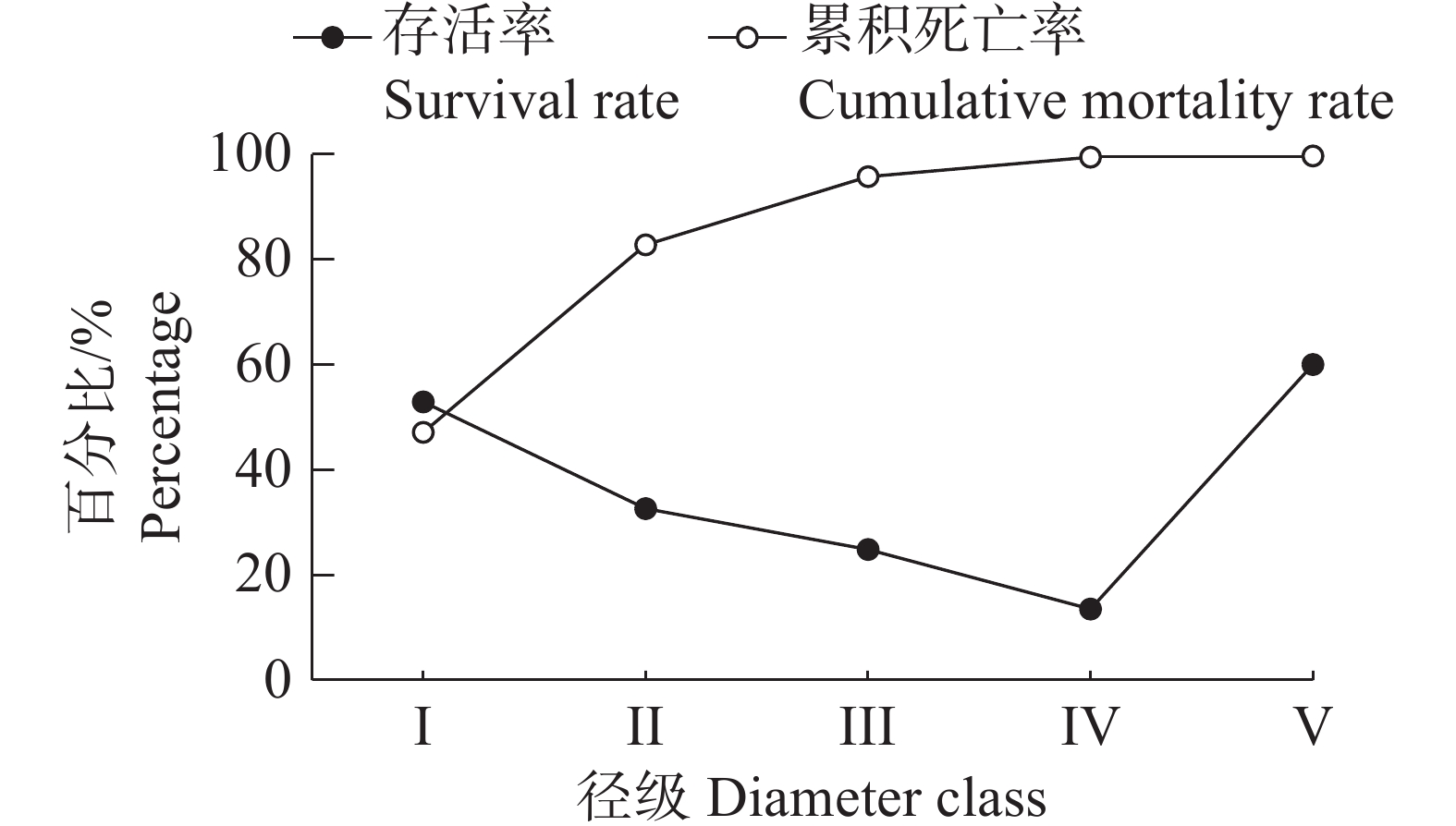
从图4可以看出,野茶种群的存活率随着径级的不断增加而逐渐下降,累积死亡率则呈现不断升高的趋势,两者呈现互补状态。在Ⅰ到Ⅱ径级之间,存活率和累积死亡率相近,表明野茶种群在这个阶段达到了平衡状态,之后累计死亡率大于存活率。存活率在Ⅰ~Ⅲ径级间下降较快,这是由于Ⅰ阶段的幼苗在向Ⅱ阶段的幼树成长的过程中,容易被其他树种竞争掉,所以这时候的存活率下降的最快,而与之对应的是累积死亡率在同这一阶段上升得最快。从Ⅲ阶段开始,曲线逐渐变得平缓,主要是因为野茶个体达到一定的大小和高度,生存力较强,这时候的累积死亡率变动较小。V阶段后存活率变得极低,是因为野茶的大树和老树个体开始进入生理衰老阶段。
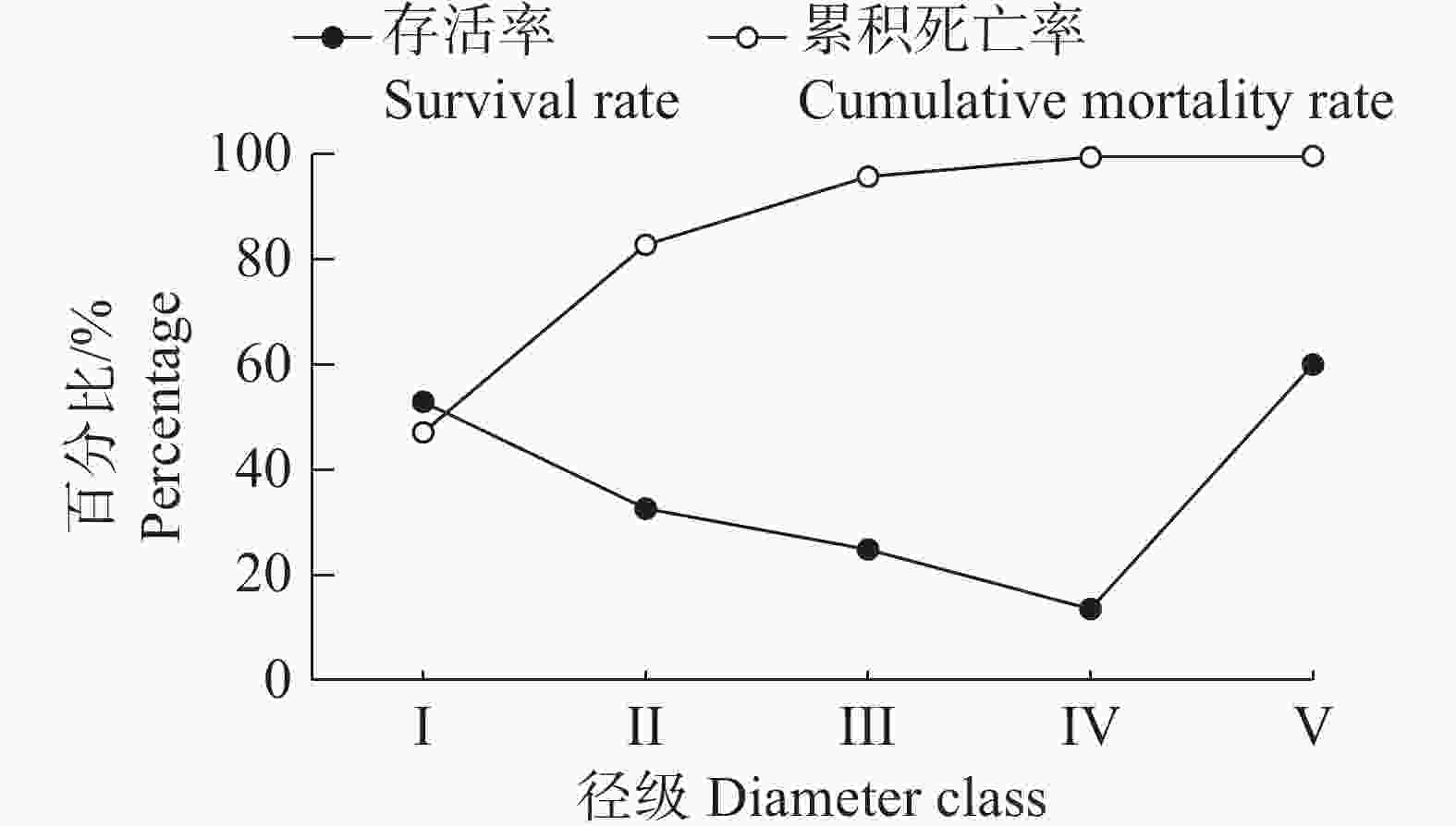
图 4 野茶种群存活率和累积死亡率曲线
Figure 4. Survival and cumulative mortality curves of Camellia sinensis var. assamica populations
-
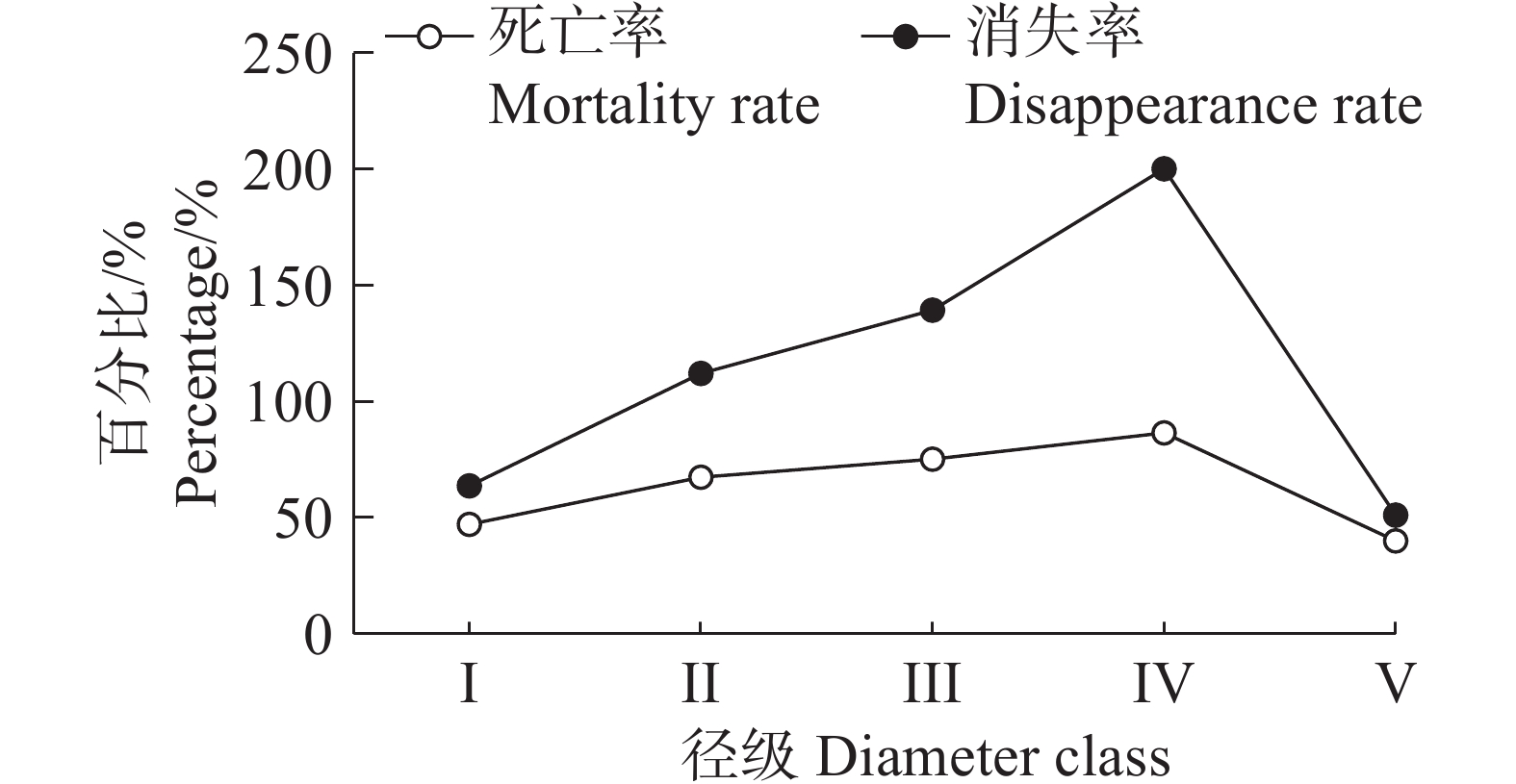
从图5可以看出,海南热带雨林国家公园中的野茶种群的死亡率和消失率的变化是一致的,都呈现随着径级的不断增大而上升,到达峰值后开始下降的趋势,两者在上升阶段都呈现“快–慢–快”的模式。死亡率的变化模式主要是因为野茶在幼苗时竞争力较弱,只有部分能存活下来并长大,随着个体的不断成长,占据一定的空间,活力增强,但依然会有大量个体被淘汰,而到了Ⅳ阶段后,野茶达到中树水平,此时的野茶占据一定优势,死亡率开始下降。在Ⅳ径级时,野茶种群的死亡率和消失率都达到峰值,说明野茶种群在这个径级时,种群数量急剧下降。
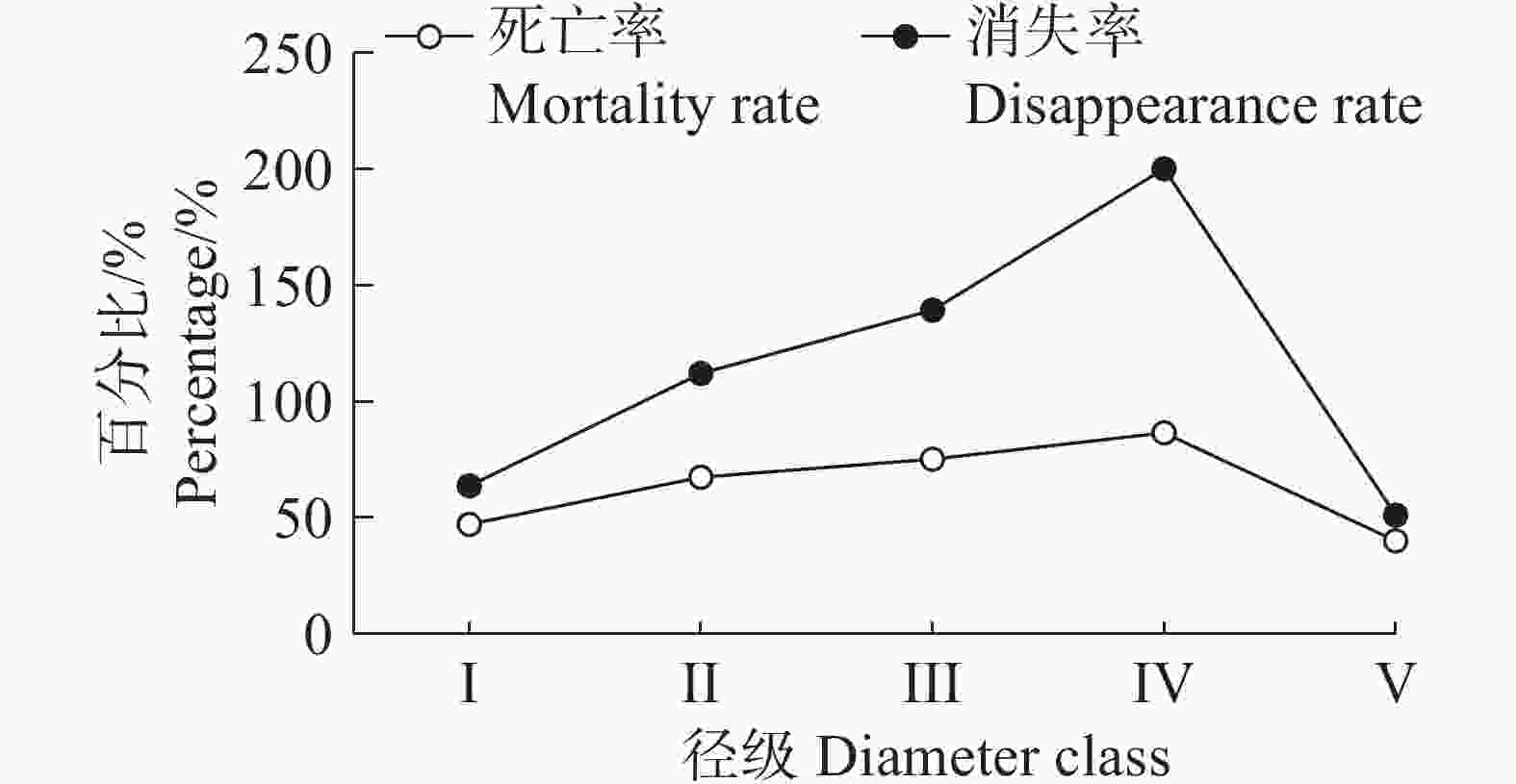
图 5 野茶种群死亡率和消失率曲线
Figure 5. Mortality and disappearance rate curves of Camellia sinensis var. assamica populations
-
海南热带雨林国家公园野茶种群的径级呈现典型“上尖下宽”的金字塔结构,为增长型种群,种群中幼苗占比最大,主体为小径级个体,大树、老树较少。存活曲线接近Deevey-Ⅰ型,幼苗存活率较高,从中树向大树过渡的过程中死亡率较高。老树较容易受到环境改变和病虫害的负面影响,应该采取相应措施开展保护,如立地环境研究、级别评定和挂牌等[19]。古茶树通常是指树龄达百年的野生茶树或栽培茶树,对茶资源的更新和可持续利用具有重要作用[20],而分布于自然保护区内的古茶树同时具备重要的历史价值和科学研究价值,应该重点开展保护[21]。
虽然海南热带雨林国家公园的野茶种群整体上呈增长趋势,但不同区域的野茶种群表现出不同的发展趋势。鹦哥岭、霸王岭的野茶径级结构较为完整,为增长型种群;黎母山野茶的径级结构不太完整,缺少大树和老树;吊罗山的野茶径级结构最不完整,只有小树和幼苗两个径级的个体,缺乏中树及以上个体。在保护时,应结合具体情况,采取不同的保护措施。鹦哥岭、霸王岭的野茶种群情况良好,应重点对古茶树个体进行保护;黎母山野茶种群大多为小径级个体,而野茶在小径级阶段受到的竞争强度较大[9],应保护小径级野茶个体,使其顺利过渡到大树阶段,实现种群的有效更新;吊罗山地区的野茶种群受到的破坏最为严重,在实际的调查中,调查区域内未记录到中树及以上个体,同时观察到较多的大树被砍伐,降为灌木状,而针对结实情况的调查结果显示,吊罗山野茶样方内的可繁殖个体都仅记录到零星的花和果。在之后的保护中,吊罗山地区应严格限制野茶果实的采摘,确保一定量的野茶幼苗的萌发,同时控制野茶叶片的采摘,确保小胸径茶树能够顺利过渡到成树阶段,实现种群的更新。从野茶种群的存活率、死亡率和消失率上看,野茶个体虽然存活率高,但在幼苗时期会产生自疏作用,许多幼苗会被淘汰掉。而在中树之前,野茶个体受到的竞争压力也会使得其死亡率较高,当达到中树阶段后,野茶个体占据一定的空间和资源,生存力增强,死亡率开始下降。
海南野茶的种群分布类型属于聚集分布[14],这与其繁殖方式有极大关系。由于其种子的传播、扩散能力较弱,所以较好的保护方式为就地保护,保护野茶的生境,同时也要减少人为活动带来的负面影响。由于茶组植物间会通过花粉传播进行基因交流,在人类的引种栽培活动下,野生种群的基因多样性可能会受到人工种群的影响[22],这需要后续开展进一步的研究。
Study of the Camellia sinensis var. assamica population in Hainan Tropical Rainforest National Park
-
摘要: 为探明海南热带雨林国家公园野茶的种群现状和动态变化,为后续种群的保护提供科学依据,采用样方法对国家公园内不同区域的野茶种群进行调查分析。结果表明:1)海南热带雨林国家公园野茶种群中幼苗数量较多,主体为小径级个体,大树、老树较少,为增长型种群;2)野茶种群的生存曲线接近Deevey-Ⅰ型,幼苗的存活率较高,中树第二阶段死亡率较高;3)针对不同区域,应采取不同的保护措施,如鹦哥岭、霸王岭区域野茶幼苗数量丰富,种群发展稳定,应重点保护大树和老树个体;黎母山区域缺乏大径级野茶个体,应重点保护中树个体,使其顺利过渡到大树阶段;吊罗山区域野茶种群更新受阻,应开展小径级野茶个体的抚育工作,使其顺利向成树过渡,实现种群更新。调查结果表明当地居民对野茶资源的保护意识较薄弱,应积极开展野茶资源保护的相关科普活动,增强当地居民的保护意识。Abstract: In order to explore the population status and dynamic changes of Camellia sinensis var. assamica in Hainan Tropical Rainforest National Park, and to provide a scientific theoretical basis for the protection of the subsequent populations, this study used quadrat survey methods to investigate the Camellia sinensis var. assamica populations in different regions of the national park. The results are as follows: (1) The Camellia sinensis var. assamica population in Hainan Tropical Rainforest National Park has a high number of seedlings, the main body of which are small-diameter individuals, and fewer individuals of large and old trees, which is a growing population. (2) The survival curve of the Camellia sinensis var. assamica population is close to the Deevey-I type, with a high survival rate of seedlings and a high mortality rate of medium-sized trees in the second stage. (3) Different protection measures should be taken in different regions: Yinggeling and Bawang Mountain regions have abundant Camellia sinensis var. assamica seedlings and stable population development, so the focus should be on the protection of large and old trees; Limu Mountain region lacks large-diameter Camellia sinensis var. assamica individuals, so the focus should be on the protection of medium-sized individuals, so as to enable them to smoothly transition to the large-diameter stage; Diaoluo Mountain region is hindered in the regeneration of the Camellia sinensis var. assamica populations, so the nurturing of small-sized Camellia sinensis var. assamica individuals should be carried out, so as to make the transition to adult trees and realize the population regeneration smoothly. The local residents' awareness of Camellia sinensis var. assamica resources protection is relatively weak, so we should actively carry out science popularization activities related to the protection of Camellia sinensis var. assamica resources to enhance the protection awareness of the local residents.
-
Key words:
- diameter class structure /
- static life table /
- population dynamics
-
图 2 不用区域的野茶种群径级结构
Fig. 2 Trail structure of Camellia sinensis var. assamica populations in different regions
图 4 野茶种群存活率和累积死亡率曲线
Fig. 4 Survival and cumulative mortality curves of Camellia sinensis var. assamica populations
图 5 野茶种群死亡率和消失率曲线
Fig. 5 Mortality and disappearance rate curves of Camellia sinensis var. assamica populations
表 1 野茶种群的静态生命表
Table 1 Static life table of Camellia sinensis var. assamica population
x lx sx/% dx qx/% Tx ex Kx Ⅰ 864 52.89 407 47.11 757.5 0.88 0.64 Ⅱ 457 32.60 308 67.40 325.5 0.71 1.12 Ⅲ 149 24.83 115 77.18 97 0.65 1.39 Ⅳ 34 13.51 29 86.49 22.5 0.60 2.00 Ⅴ 5 60.00 2 40.00 4 0.80 0.51 Ⅵ 3 — — — — — — 注:x为野茶径级,共分6级;lx为野茶种群在x径级时的存活数;sx为野茶种群的存活率;dx为x径级到x+1径级过渡时死亡的野茶个体数;qx为野茶种群从x径级到x+1径级过渡时的死亡率;Tx为从x径级开始,超过x径级的存活下来的个体数之和;ex为生命期望,为x径级的个体在未来所能存活的平均年数期望;Kx为消失率;“—”表示无。 Note: x is the diameter class of Camellia sinensis var. assamica, divided into 6 levels; lx is the number of individuals in the Camellia sinensis var. assamica population at diameter class x; sx is the survival rate of the Camellia sinensis var. assamica population; dx is the number of Camellia sinensis var. assamica individuals that died at the transition from diameter class x to x +1; qx is the mortality rate of the Camellia sinensis var. assamica population at the transition from diameter class x to x +1; Tx is the sum of the number of individuals that survived beyond diameter class x from diameter class x onward; ex is the life expectancy, which is the i expectation of the average number of years that individuals of diameter class x will survive in the future; Kx is the disappearance rate;“—” means none.  下载: 导出CSV
下载: 导出CSV
-
[1] 张金峰, 葛树森, 梁金花, 等. 长白山阔叶红松林紫椴种群结构与动态特征[J]. 生态学报, 2022, 42(13): 5381 − 5390. doi: 10.5846/stxb202106191629 [2] 余潇, 代嫚婷, 普甜, 等. 珍稀濒危植物蒜头果种群结构及动态特征研究[J]. 西部林业科学, 2023, 52(3): 8 − 16. doi: 10.16473/j.cnki.xblykx1972.2023.03.002 [3] 张金峰, 葛树森, 梁金花, 等. 长白山阔叶红松林红松种群年龄结构与数量动态特征[J]. 植物生态学报, 2022, 46(6): 667 − 677. doi: 10.17521/cjpe.2021.0498 [4] 张维伟, 尹代皓, 雷雨婷, 等. 黄土高原南部麻栎不同群落类型乔木种群结构及演替模拟[J]. 生态学报, 2024, 44(6): 2572 − 2581. doi: 10.20103/j.stxb.202110263011 [5] 夏丹, 王伟锋, 李东海, 等. 濒危野生卷萼兜兰种群分布格局和数量动态特征[J]. 热带亚热带植物学报, 2024, 32(5): 579 − 588. doi: 10.11926/jtsb.4817 [6] 林春惠, 顾惠怡, 叶钦良, 等. 珍稀濒危植物大苞山茶种群结构与动态特征[J]. 植物生态学报, 2023, 47(12): 1684 − 1692. doi: 10.17521/cjpe.2022.0488 [7] 张威, 古小玲. 海南自由贸易港建设背景下茶产业发展优劣势分析与对策建议[J]. 热带农业工程, 2023, 47(5): 68 − 71. [8] 杨小波. 海南植物图志[M]. 北京. 科学出版社, 2015. [9] 朱子丞, 戚春林, 杨小波, 等. 鹦哥岭野茶群落物种组成与竞争关系研究[J]. 林草资源研究, 2023(6): 129 − 136. doi: 10.13466/j.cnki.lczyyj.2023.06.016 [10] 苏凡, 杨小波, 李东海. 基于形态学特征和psbA-trnH叶绿体编码基因序列明确五指山野生茶的分类地位[J]. 热带作物学报, 2019, 40(8): 1565 − 1570. doi: 10.3969/j.issn.1000-2561.2019.08.016 [11] 刘肖冰, 陈冬梅, 林娟. 海南五指山野生茶制得红茶和绿茶的生化品质成分[J]. 东北林业大学学报, 2019, 47(4): 68 − 70. doi: 10.3969/j.issn.1000-5382.2019.04.014 [12] 苏凡, 杨小波, 李东海, 等. 五指山水满茶的茶多酚、总黄酮含量与抗氧化活性研究[J]. 热带作物学报, 2018, 39(4): 675 − 680. doi: 10.3969/j.issn.1000-2561.2018.04.010 [13] 郭远安. 海南岛野生茶树的调查[J]. 广东农业科学, 1991(2): 45. doi: 10.16768/j.issn.1004-874x.1991.02.024 [14] 李苑菱, 张丽, 杨小波, 等. 海南岛野生茶树空间分布及种群动态研究[J]. 林业资源管理, 2017(2): 81 − 87. doi: 10.13466/j.cnki.lyzygl.2017.02.014 [15] 李苑菱. 海南岛野生茶树种群生态学研究[D]. 海口: 海南大学, 2017. [16] 方发之, 吴二焕, 麦有专, 等. 海南热带雨林国家公园珍稀濒危观赏植物调查研究[J]. 南方林业科学, 2023, 51(2): 40 − 46. doi: 10.16259/j.cnki.36-1342/s.2023.02.009 [17] 刘阳. 海南热带雨林国家公园生态保护治理研究[J]. 资源与人居环境, 2023(7): 61 − 64. doi: 10.3969/j.issn.1672-822X.2023.07.020 [18] 李国华, 郭向阳, 李霖明, 等. 海南热带雨林国家公园不同植被类型的大型真菌多样性[J]. 生物多样性, 2022, 30(7): 176 − 188. doi: 10.17520/biods.2022110 [19] 曾煜, 陈悦. 云南古茶树资源保护的地方立法与制度的完善[J]. 西南林业大学学报(社会科学), 2020, 4(4): 18 − 23. [20] 蒋会兵, 唐一春, 陈林波, 等. 云南省古茶树资源调查与分析[J]. 植物遗传资源学报, 2020, 21(2): 296 − 307. doi: 10.13430/j.cnki.jpgr.20190422001 [21] 何祖普. 以云南省为谈例我国古茶树和古茶园的法律保护机制[J]. 福建茶叶, 2016, 38(4): 280 − 281. doi: 10.3969/j.issn.1005-2291.2016.04.179 [22] LI M M, MEEGAHAKUMBURA M K, WAMBULWA M C, et al. Genetic analyses of ancient tea trees provide insights into the breeding history and dissemination of Chinese Assam Tea (Camellia sinensis var. assamica)[J]. Plant Diversity, 2024, 46(2): 229 − 237. doi: 10.1016/j.pld.2023.06.002 -

 点击查看大图
点击查看大图
计量
- 文章访问数: 838
- HTML全文浏览量: 285
- PDF下载量: 10
- 被引次数: 0




