


-
海南热带雨林国家公园黎母山位于海南岛的地理中心,是该岛热带雨林生态系统的核心保护区之一。这里不仅拥有丰富的人工林资源,更孕育着广袤的天然林。该区域内的平均气温维持在23.1 ℃,年平均降雨量高达
2343.1 mm,年平均相对湿度达到84%,水热条件极为优越[1 − 2]。保护区森林覆盖率达90%,植被类型为热带常绿季雨林[3 − 4],其中野生维管束植物共有190科759属1490 种,海南特有种为163种,占海南省特有树种的32.2%[4]。大蚕蛾科(Saturniidae)是鳞翅目(Lepidoptera)中因体型庞大而闻名的一大类群[5]。这类昆虫翅面色彩斑斓,常有艳丽夺目的图案,在收藏和工艺品领域备受青睐。更为重要的是,该科包含多种重要绢丝昆虫,其茧丝被广泛应用,具有重要的经济价值[6 − 7]。特别是樟蚕丝,因其独特性质,被视作高端钓鱼线以及外科手术缝合线的理想材料[8]。大蚕蛾种类繁多,全球分布极为广泛,尤其偏好热带和亚热带地区[9]。作为夜行性的食叶性昆虫[10],它们在生态系统中扮演着重要角色,是生物多样性不可或缺的一部分。
依据Kitching等人(2018)提出的分类系统,大蚕蛾科分为8个亚科,包括Agliinae、Arsenurinae、Ceratocampinae、Cercophaninae、Hemileucinae、Oxyteninae、Salassinae和Saturniinae,共计约180属
3454 种[11],展现了其丰富的物种多样性。1996年出版的《中国动物志》详细记载了我国15属58种大蚕蛾,其中海南省记录9属15种[8]。2002年《海南森林昆虫》记录海南省拥有9属19种[12]。海南岛的昆虫种类丰富,具有典型的热带特征,与南亚岛国的昆虫区系有许多相似之处[12]。昆虫资源潜力巨大,其中海南特有种,如海南小字大蚕蛾Cricula hainanensis[13]、海南树天蚕Lemaireia hainana[14]、海南鸮目大蚕蛾Salassa shuyiae[15]。这些特有种的存在,不仅丰富了海南岛的生物多样性,也为研究热带雨林生态系统的稳定性、物种适应性及生态演替提供了独特的视角。
黎母山景区系海南热带雨林国家公园的重要组成部分,属一般控制区及天窗区域。其独特的生态环境和丰富的生物多样性使其成为生物多样性保护的重要区域,也是开展热带雨林生态学研究、昆虫学研究以及生态旅游的理想场所,具有极高的科学研究和保护价值。
本研究旨在通过灯诱法和踏查,系统调查黎母山景区大蚕蛾科的物种多样性、飞行规律、趋光节律及区系特征,为海南热带雨林国家公园的生物多样性保护和资源利用提供科学依据。
-
研究结合了不定期调查与系统监测。2020–2023年期间进行不定期灯诱和踏查。自2023年10月起至2024年9月进行系统监测,每月于残月和蛾眉月期间,在黎母山(天窗区)3个地点——七家坊管护站(19.1025°N,109.4347°E)、学而山房(19.1020°N,109.4415°E)、锦绣湖(19.1019°N,109.4434°E),开展两次连续的灯诱活动(高压汞灯500 W,E40白光220 V,灯诱帐帐篷145 cm × 145 cm × 205 cm)。每晚19:00开灯,于21:00、00:00、03:00和06:00四个时间点逐次检查并记录所诱集的大蚕蛾种类、个体数量和性别(同一个体在其首次出现的时间点记录,后续时段不再重复记录),每种最多采集2对样本。
-
诱集到的雄成虫用98%乙醇处死后制作针插标本。获得的雌虫用于人工套卵,或从户外踏查中获得幼虫,进行室内饲养观察(温度26 ℃,湿度80%),并记录生命周期。
-
主要参考《中国昆虫志》第五卷[8]、《蝴蝶与蛾》[16]、《中华美蛾》[17]、《中国蛾类图鉴》[18]、《秦岭昆虫志》[19]、《庐山蝶蛾志》[20]、《蛾类图册》[21],同时借助昆虫爱好者社区(
https://www.inaturalist.org/home )和https://eol.org/pages/51538242/articles 进行物种辅助鉴定。对照《拉英汉昆虫学词典》[22]校正中文名信息。 -
计算公式:Rai = Ni/N × 100%,Rai表示物种i的相对多度,Ni代表物种i的个体数量,N为所有物种个体总数。个体数占群落总数量的10.00%以上者为优势种,1%–10%为常见种,小于1%的为稀有种[23]。
-
本研究使用非参数检验中的Kruskal-Wallis方差分析比较不同时间点所采集蛾类的翅展与体长的形态参数差异,显著度设定p < 0.05,数据分析和绘图在Excel 2022和Origin 2024中进行。
-
从2020年10月至2024年10月,经过70次踏查和灯诱,我们记录到黎母山景区大蚕蛾科共有2个亚科、9属和15种(表1),未发现其他6个亚科。
表 1 黎母山15种大蚕蛾物种名录及国内分布
Table 1. Checklist and domestic distribution of 15 Saturniidae species in Limushan
亚科/属
Subfamily/Gene种
Species国内分布
Distribution in China大蚕蛾亚科 Saturniinae 大蚕蛾属
Attacus乌桕大蚕蛾
Attacus atlas海南、贵州、云南、香港、江西、福建、广东、广西、湖南、台湾[8, 12, 17–18] 樗蚕蛾属
Samia王氏樗蚕
Samia wangi广东、湖南、江西、福建、香港、台湾、海南、海南、西藏、四川、陕西、浙江[27] 小字大蚕蛾属
Cricula海南小字大蚕蛾
Cricula hainanensis海南特有种[13] 树天蚕属
Lemaireia海南树天蚕
Lemaireia hainana海南特有种[14] 霖王蛾属
Rinaca藏霖王蛾
Rinaca thibeta海南、云南、福建[8, 17] 希霖王蛾
Rinaca simla福建、四川、广东、海南、云南[8, 12, 18] 目大蚕蛾属
Saturnia樟蚕
Saturnia pyretorum吉林、河南、陕西、甘肃、湖北、浙江、安徽、海南、贵州、内蒙古、黑龙江、辽宁、江苏、四川、广东、广西、河北、山东、湖南、福建、江西、云南、湖北、台湾[8, 12, 17 − 19] 长翅樟蚕
Saturnia roseata重庆、海南[28] 尾大蚕蛾属
Actias华尾大蚕蛾
Actias sinensis陕西、甘肃、广东、海南、江西、湖南、广西、台湾[8, 12, 17 − 19] 绿尾大蚕蛾
Actias selene陕西、甘肃、海南、江苏、江西、河南、河北、浙江、湖南、广东、福建、广西、台湾、云南、西藏、辽宁、湖北、四川、吉林[8, 12, 18 − 20] 豹大蚕蛾属
Loepa黄豹大蚕蛾
Loepa katinka河南、河北、宁夏、西藏、安徽、浙江、福建、江西、四川、广东、广西、云南、海南[8, 12, 20] 红豹大蚕蛾
Loepa anthera四川、海南、西藏、福建、广东、广西、云南[8, 12, 17–18] 柞蚕属
Antheraea钩翅柞蚕
Antheraea assamensis广东、云南、海南[8, 12, 17–18] 明目柞蚕
Antheraea frithi陕西、湖南、福建、西藏、云南、海南[8, 17, 19, 21] 猫目大蚕蛾亚科 Salassinae 猫目大蚕蛾属
Salassa海南鸮目大蚕蛾
Salassa shuyiae海南特有种[15] 其中,猫目大蚕蛾亚科Salassinae仅有1属1种——海南鸮目大蚕蛾Salassa shuyiae;大蚕蛾亚科Saturniinae为优势亚科,包括8属14种。
-
本次试验中,从2023年10月至2024年9月,各个月份均成功诱集到大蚕蛾(图1)。所获个体中以雄蛾为主,占总捕获量的88.74%,雌蛾仅占11.26%。6月物种数达到最大值,共7种;3月个体数最多,共38只。2023年10月和2024年1、2、5、7、9月各种群数量明显下降,可能与调查前期持续强降雨或高温影响。
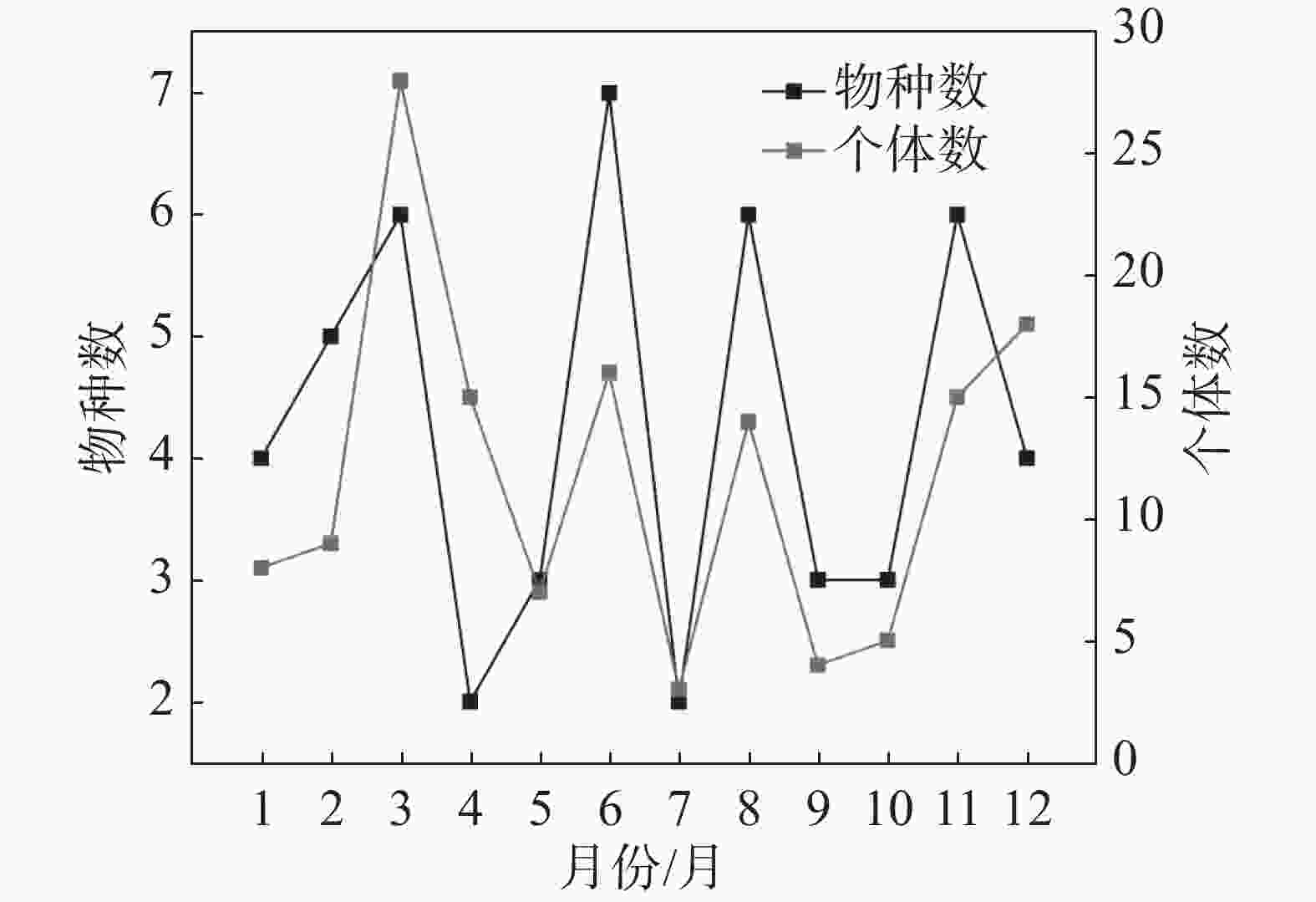
图 1 黎母山大蚕蛾科昆虫物种数和个体数量的月动态变化
Figure 1. Monthly dynamics of species richness and individual abundance of Saturniidae in Limushan
-
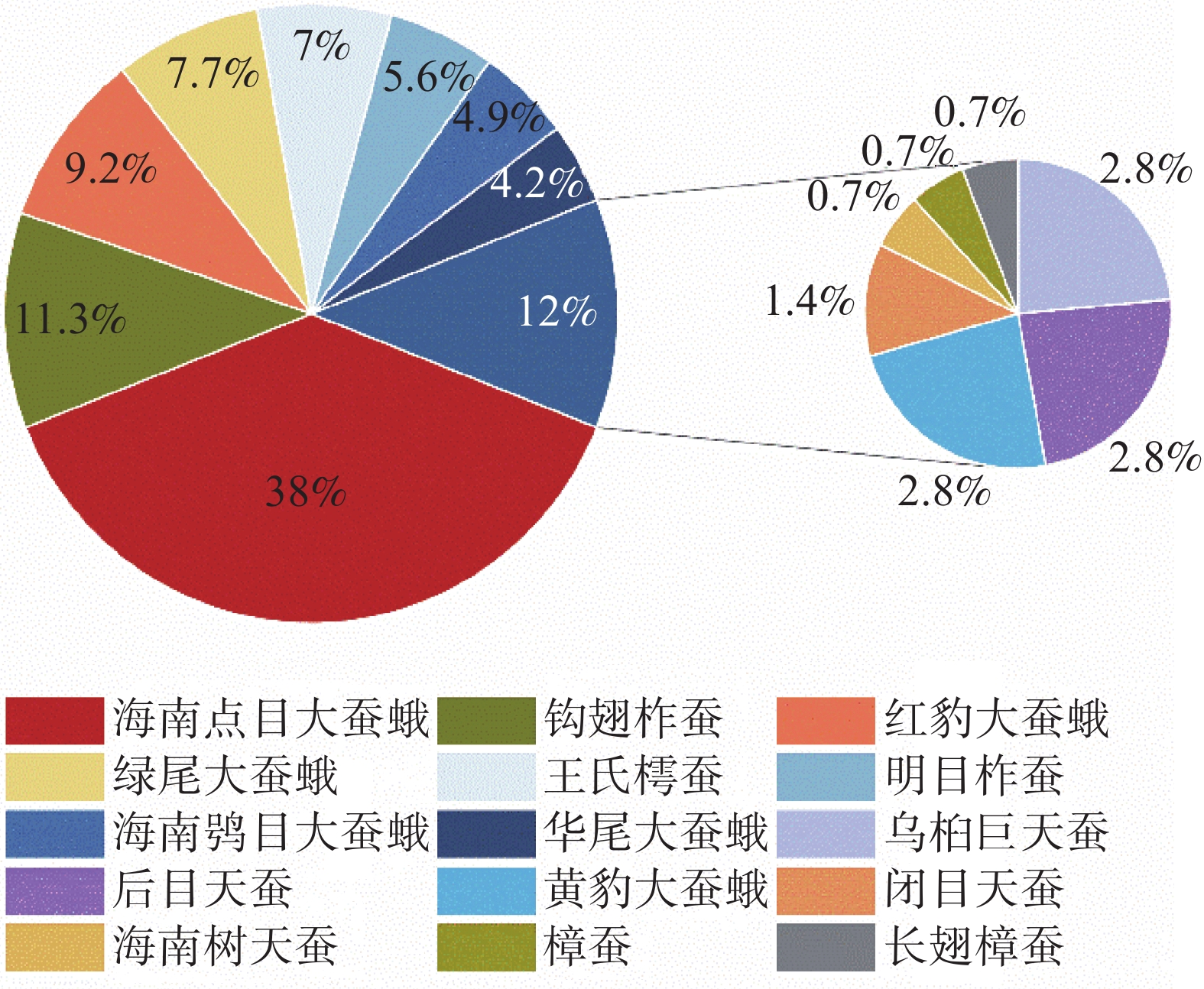
根据全年大蚕蛾数量统计结果表明(图2),海南小字大蚕蛾和钩翅柞蚕是黎母山的为优势种,分别占总个体数的38.03%和11.27%;藤豹大蚕蛾、绿尾大蚕蛾、王氏樗蚕、明目柞蚕、海南鸮目大蚕蛾、华尾大蚕蛾、乌桕大蚕蛾、希霖王蛾、黄豹大蚕蛾和藏霖王蛾为常见种,共占总数48.59%;海南树天蚕、樟蚕、长翅樟蚕为稀有种,仅占2.1%。
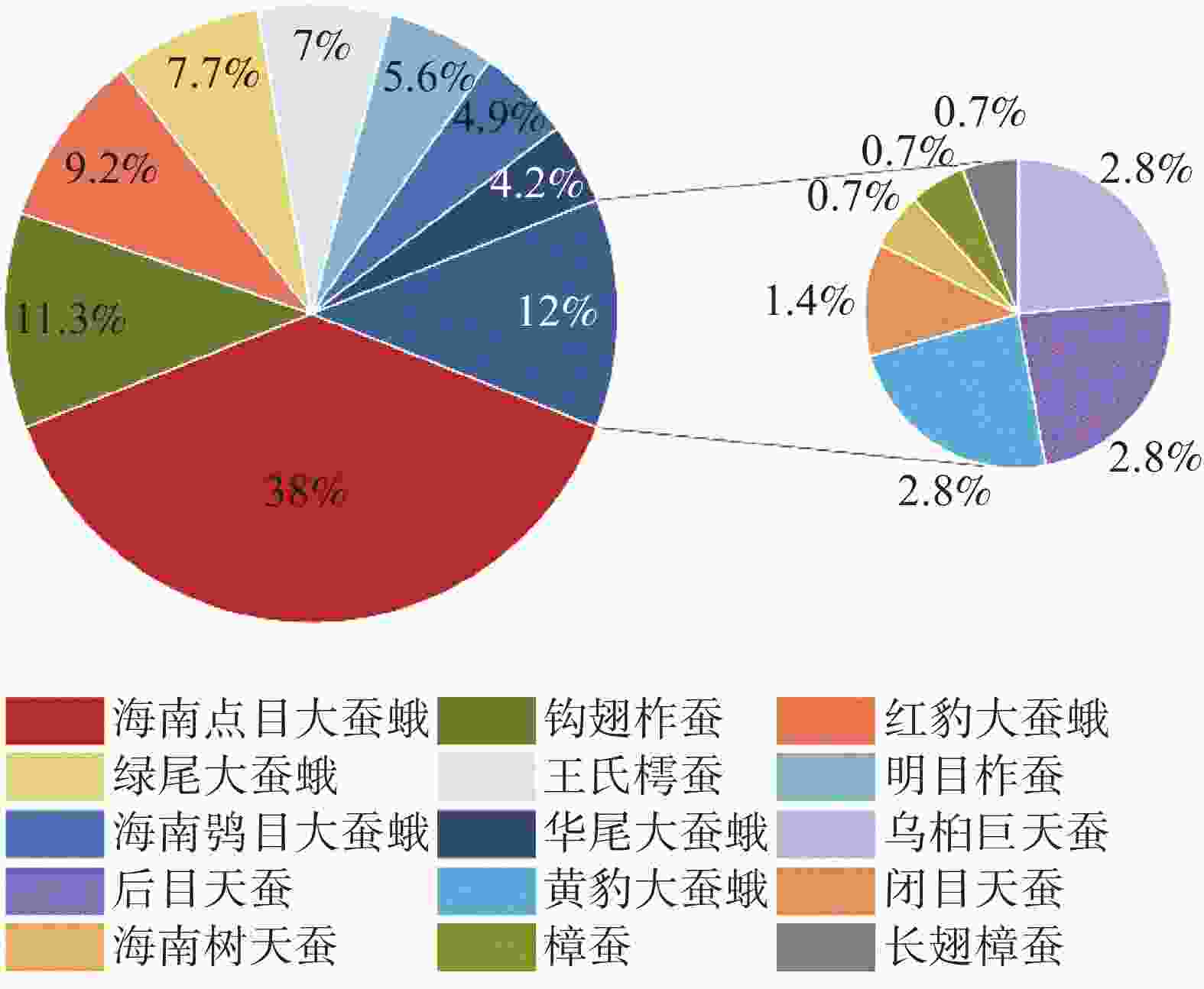
图 2 黎母山大蚕蛾科昆虫相对多度
Figure 2. Relative abundance of Saturniidae in Limushan
-
通过对15种大蚕蛾灯诱数据的飞行期分析(图3)发现,其羽化规律可分为三类:第一类包括海南树天蚕、长翅樟蚕、樟蚕、藏霖王蛾、希霖王蛾及海南鸮目大蚕蛾等6种,集中出现在10月至次年1月,仅具单一峰期,表现出明显的一化性特征;第二类为黄豹大蚕蛾和藤豹大蚕蛾,前者出现在11月至次年3月,后者见于3月至8月,均具有2个明显峰期,初步推断为二化性昆虫;第三类包括钩翅柞蚕、明目柞蚕、王氏樗蚕、乌桕大蚕蛾、海南小字大蚕蛾、华尾大蚕蛾和绿尾大蚕蛾,其中前4种集中出现于3月至9月,并呈现2–3月、5–6月及8–9月共3个发生期;后3种几乎全年可见,且存在世代重叠现象。室内饲养显示,海南小字大蚕蛾和钩翅柞蚕的世代周期为2–2.5个月,华尾大蚕蛾为1.5–2.5个月,乌桕大蚕蛾为2.5–3.5个月。综上,推测这7种大蚕蛾在当地每年可发生三代及以上,属于多化性昆虫。
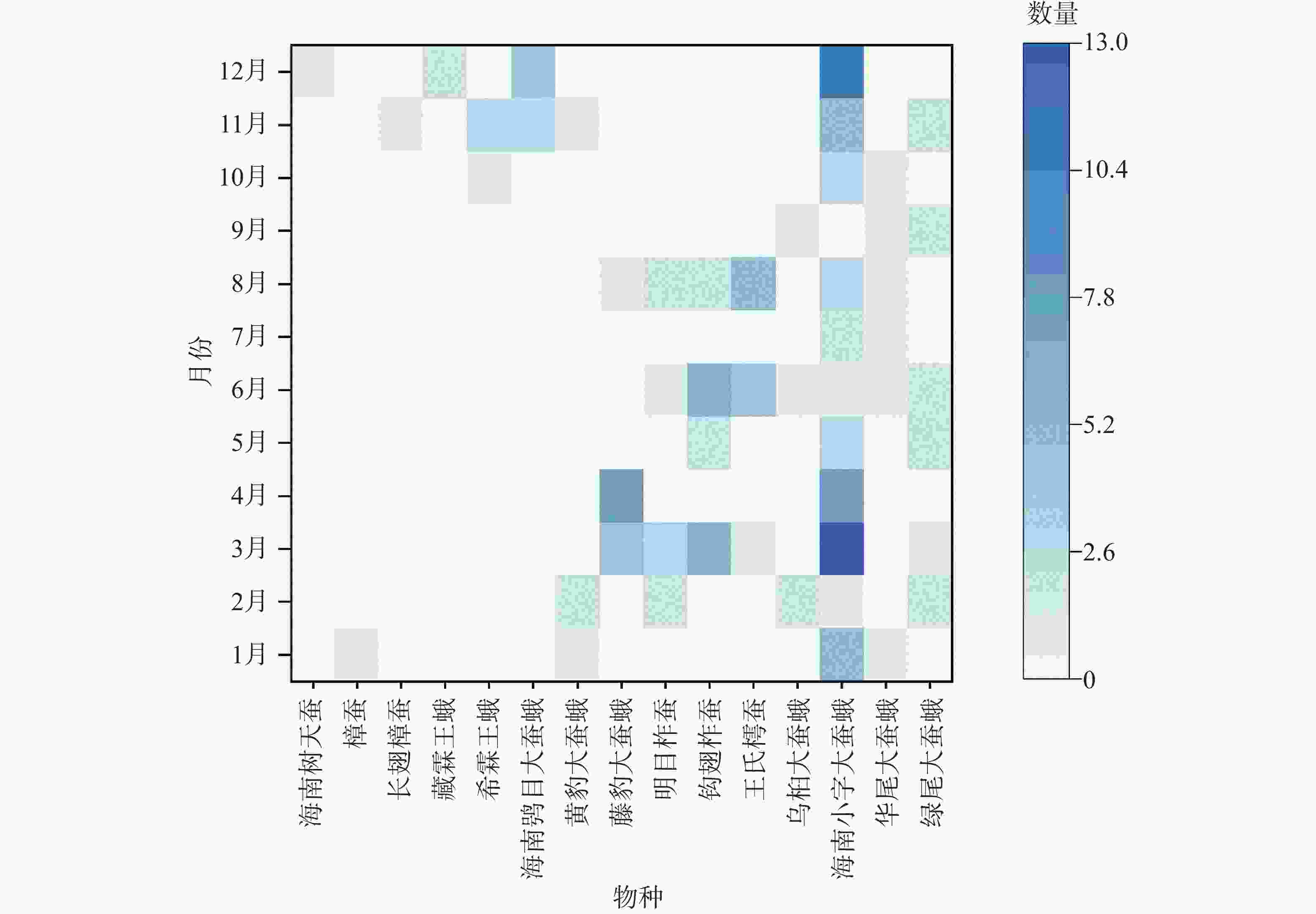
图 3 黎母山大蚕蛾科昆虫不同物种的飞行期分布与数量
Figure 3. Flight period distribution and abundance of Saturniidae in Limushan
-
从不同时间段大蚕蛾上灯数据分析(图4)表明,其趋光活动呈现出明显的时间节律。个体在19:00–21:00、21:00–00:00、00:00–03:00、03:00–06:00四个时间段上灯的比例分别为4.23%、9.86%、29.58%和56.34%,表明绝大多数个体集中于后半夜活动,尤其是03:00–06:00为活动高峰。
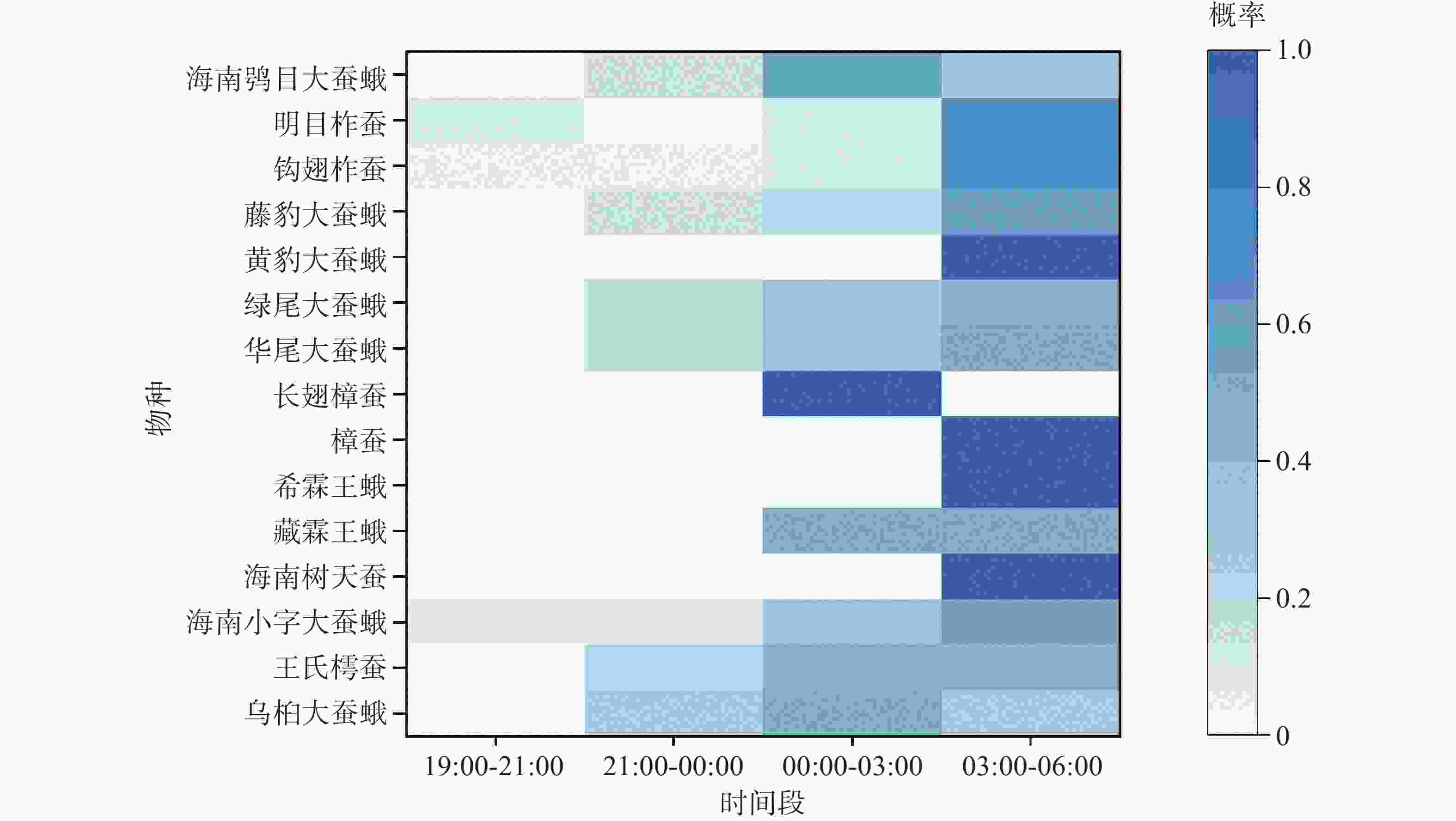
图 4 黎母山大蚕蛾科昆虫不同时间段的上灯频率
Figure 4. Light-attraction frequency of Saturniidae at different time periods in Limushan
从物种特异性来看,黄豹大蚕蛾、海南树天蚕、希霖王蛾、樟蚕、明目柞蚕、钩翅柞蚕及藤豹大蚕蛾等7种几乎集中在03:00–06:00出现;而海南鸮目大蚕蛾和乌桕大蚕蛾、长翅樟蚕则主要在00:00–03:00出现。
-
通过结合表1和昆虫区系资料(见表3),我们发现黎母山景区大蚕蛾科昆虫的分布特点:在世界动物区系中黎母山的大蚕蛾在东洋界均有分布,部分物种在古北界也有分布;在中国动物区系中主要分布在华南、华中、西南区。总体来说,黎母山大蚕蛾科的区系特征表现出显著的热带区系特征,与海南岛的自然环境相吻合。
表 3 黎母山大蚕蛾在世界动物区系和中国动物区系中的分布型及占比
Table 3. Distribution patterns and proportions of Saturniidae from Limushan in global and Chinese zoogeographical regions
序号
Number分布型
Distribution Patterns物种数
Species numbers占比
Total Species Percentage(%)世界动物区划 1 东洋界-古北界 8 53.33% 2 东洋界-古北界 7 46.67% 中国动物区划 1 华南区 3 20% 2 华南区-华中区 1 6.67% 3 华南区-华中区-西南区 5 33.33% 4 华南区-华中区-西南区-华北区 1 6.67% 5 华南区-华中区-西南区-华北区-青藏区 2 13.34% 6 西南区-华南区-华中区-华北区-蒙新区-青藏区 1 6.67% 7 西南区-华南区-华中区-东北区-华北区-蒙新区 1 6.67% 8 西南区-华南区-华中区-东北区-华北区-蒙新区-青藏区 1 6.67% 表 2 黎母山大蚕蛾科昆虫区系
Table 2. Fauna of Saturniidae in Limushan
序号
number种名
species name世界动物区划
Zoogeographic fauna of the world中国动物区划
Zoogeographic fauna of ChinaA B C D E F a b c d e f g 1 乌桕大蚕蛾 √ √ √ √ √ 2 王氏樗蚕 √ √ √ √ √ √ √ 3 海南小字大蚕蛾 √ √ 4 海南树天蚕 √ √ 5 藏霖王蛾 √ √ √ √ 6 希霖王蛾 √ √ √ √ √ 7 樟蚕 √ √ √ √ √ √ √ √ 8 长翅樟蚕 √ √ √ 9 华尾大蚕蛾 √ √ √ √ √ 10 绿尾大蚕蛾 √ √ √ √ √ √ √ √ √ 11 黄豹大蚕蛾 √ √ √ √ √ √ √ √ 12 藤豹大蚕蛾 √ √ √ √ 13 钩翅柞蚕 √ √ √ √ 14 明目柞蚕 √ √ √ √ √ √ √ 15 海南鸮目大蚕蛾 √ √ 注:“√”表示有分布,A为古北界,B为东洋界,C为新北界,D为非洲界,E为新热带区,F为澳洲界;a为西南区,b为华中区,c为华南区,d为东北区,e为华北区,f为蒙新区,g为青藏区 -
根据全国大蚕蛾昆虫的资料,我国大蚕蛾科昆虫共计有15属58种。而黎母山景区所记录到的大蚕蛾科昆虫分别占全国大蚕蛾科昆虫属数的60%和种数的25.86%。值得注意的是,海南黎母山景区的面积相较于全国总面积而言,不足0.36%,然而,就是在这样相对狭小的区域内,却蕴藏着如此丰富的大蚕蛾科昆虫资源。这一现象充分表明,黎母山地区在大蚕蛾科昆虫的物种多样性方面,具备极高的研究价值。
2002年出版的《海南森林昆虫》[12]该书中记录了9属19种大蚕蛾。黎母山四年实地调查中发现15种,与书中记录相符的有8种,其中包括王氏樗蚕、明目柞蚕、藏霖王蛾、长翅樟蚕、海南鸮目大蚕蛾以及海南树天蚕等6种大蚕蛾在《海南森林昆虫》中未被记录,后4种成虫羽化时间集中在11月到翌年1月,这可能与当时对海南冬季的调查有限有关。另外,《海南森林昆虫》中的点目天蚕蛾Cricula andrei于2004年被Brechlin重新定义为新种——海南小字大蚕蛾Cricula hainanensis[13]。
该书中记录的10种大蚕蛾(如黄尾大蚕蛾Actias heterogyna、角斑樗蚕蛾Samia watsoni、蓖麻蚕蛾Samia ricina、细带樗蚕蛾Samia insularis、樗蚕蛾Samia cynthia、豹大蚕蛾Loepa oberthuri、目豹大蚕蛾Loepa damartis、银杏大蚕蛾Dictyoploca japonica、黄目大蚕蛾Caligula anna、小字大蚕蛾Cricula trifenestrata)在本次4年调查中均未发现。
受人为干扰与气候变化双重因素的影响,全球生物多样性正呈现出衰退趋势,其中昆虫数量和多样性的急剧减少尤为显著。据多项研究指出,全球范围内已有超过20%的昆虫物种数量出现下降,部分地区的昆虫数量更是锐减近98%[29 – 30]。其下降速度已达到脊椎动物的两倍[31]。也有研究表明,当前超过40%的昆虫种类数量正在下降,地面上的昆虫数量几乎下降了98%[32];另有研究发现,森林中的部分昆虫,如甲虫正面临灭绝的威胁,而蝴蝶的数量也在急剧减少[33]。全球生物多样性下降的趋势,以及本次调查的时空局限性,可能是导致未能发现历史记录的10个物种的原因之一。考虑到我们的调查是在海拔600多米处通过灯诱方式进行的,那些未被记录到的10种大蚕蛾,很可能栖息在低海拔地区,或是周边其他我们尚未涉足的国家公园核心区内。
-
本研究结合文献,总结了黎母山15种大蚕蛾在不同地区的飞行期及化性(表4)。乌桕大蚕蛾、华尾大蚕蛾、钩翅柞蚕、明目柞蚕在黎母山地区的年世代数目,与《中华美蛾》[17]《中国蛾类图鉴Ⅳ》[18]有所不同。上述物种在海南表现为多化性昆虫。这一现象可能与海南独特的气候条件密切相关,特别是其全年充足的热量积累密切相关,为部分化性昆虫提供了更为适宜的生长环境,从而促使它们的世代数目在海南地区有所增加。在4年的调查中,黎母山的大蚕蛾成虫飞行期涵盖了全年多个月份,揭示了、其种群发生的持续性。同一种大蚕蛾在不同地区还可能表现出的化性差异,这进一步印证了昆虫化性的多样性和复杂性。
表 4 15种大蚕蛾昆虫不同地区飞行期及化性
Table 4. Flight periods and voltinism of 15 Saturniidae from Limushan compared to those in other regions
物种
Species飞行期及化性 Flight period and Voltinism 国内其他地区 海南(黎母山) 乌桕大蚕蛾 Attacus atlas 中国南部、香港、台湾,4–9月,一化或二化性[17];
江西、福建,4–5月及7–8月,二化性[18]3–9月,多化性 王氏樗蚕 Samia wangi 未见记录 3–9月,多化性 海南小字大蚕蛾 Cricula hainanensis 未见记录 几乎全年可见,多化性 海南树天蚕 Lemaireia hainana 未见记录 10月至次年1月,一化性 藏霖王蛾 Rinaca thibeta 西南、华南,10–11月,一化性[17] 10月至次年1月,一化性 希霖王蛾 Rinaca simla 未见记录 10月至次年1月,一化性 樟蚕 Saturnia pyretorum 台湾北部及海南,12月及次年1–2月,一化性[18];
河北、江西、福建、广西、广东,三月中旬[21];
2–4月,一化性[17]10月至次年1月,一化性 长翅樟蚕 Saturnia roseata 未见记录 10月至次年1月,一化性 华尾大蚕蛾 Actias sinensis 中国南部、西南、香港、台湾,4–9月[17];
江西、湖南,4月,二化性[18]几乎全年可见,多化性 绿尾大蚕蛾 Actias selene 中国大部分地区,4–10月,二化或三化[17];
河北、河南、浙江、湖南、江西、广东、福建、台湾,
5、7、9月,二化或三化[18]几乎全年可见,多化性 黄豹大蚕蛾 Loepa katinka 华南、西南,4月[17] 11月至次年3月,二化性 藤豹大蚕蛾 Loepa anthera 广东、广西、云南,4–9月,二化性[17];
福建,7月[18]3–8月,二化性 钩翅柞蚕(琥珀蚕)Antheraea assamensis 南部、西南部,4–8月,二化性[17];
云南,4月出现[18]3–9月,多化性 明目柞蚕 Antheraea frithi 中国南部、西南部,4–8月,二化性[17];
云南,5月[18, 21]3–9月,多化性 海南鸮目大蚕蛾 Salassa shuyiae 未见记录 10月至次年1月,一化性 -
昆虫的趋光性受多种因素影响,不同物种的上灯时间存在差异。如螟蛾科(Pyralidae)昆虫通常在上半夜更为活跃,而夜蛾科(Noctuidae)昆虫则倾向于在下半夜活动[34];斜纹夜蛾Spodoptera litura的活跃时段为23:00–03:00(次日),而甜菜夜蛾Spodoptera exigua的活跃时间则为01:00–04:00[35]。在水稻田中,稻纵卷叶螟Cnaphalocrocis medinalis的上灯高峰出现在20:00–21:00以及次日1:00–3:00,而黄翅大白蚁Macrotermes barneyi的上灯高峰则集中在19:00–21:00[36]。在黎母山有85.92%的大蚕蛾出现在后半夜00:00–06:00,最活跃的时间段为03:00–06:00。这一发现进一步证实了不同昆虫物种在趋光性上的时间差异。
周阳等于19:30–22:30在不同的林分中采用灯诱帐与聚光灯诱箱(二者灯泡规格一样,主要是光域不一样)相比,发现灯诱帐的形态参数均显著高于聚光灯诱箱,前者采集的蛾类物种体型更大、飞行能力更强[37]。但我们在固定地点、固定时间(19:00–06:00(次日)、采用统一的灯诱方法,并测量了所有样本的翅展(54–220 mm)与体长(15.32–43.34 mm)。统计分析表明,四个不同时间段诱集的大蚕蛾,与其翅展和体长均无显著性差异
性别等生理因素均会对昆虫的趋光行为造成影响。大量灯诱试验表明,在被光捕获的昆虫物种中,雌雄比例往往存在显著差异。在本次试验中,通过灯诱捕获的昆虫雌雄比例为1∶12。Nowinszky等人对匈牙利林业用诱捕灯在森林中诱集到的32种大型鳞翅目昆虫进行了分析,发现其中有29种昆虫的雄虫数量高于雌虫[38]。徐练发现了玉米螟Pyrausta nubilalis雄虫的趋光反应值整体上高于雌虫[39]。刘寿明等人也证实了当单独使用杀虫灯时,被诱捕的雌蛾数量(864头)显著少于雄蛾数量(
1954 头)[40]。性别差异可能源于多种因素。部分昆虫的雄成虫复眼较大,对光线更为敏感,因此雄性飞蛾可能比雌性飞蛾更容易受到光源的吸引[41]。值得注意的是,我们还发现被捕获的雌性成虫具有较高的携卵率。程文杰和张锦芳等人(2011,2022)认为,这可能是因为雄性飞蛾具有较强的活动能力,需要以灯光为导航来寻找交配的雌性,而已交配的雌性飞蛾则可能以灯光为导航来寻找合适的产卵地点[41–42]。
尽管许多研究已经从不同角度对昆虫的趋光行为进行了解释,但要想获得更加全面和完善的解释,还需要进一步的研究。
-
从生态区系的角度分析,黎母山景区的大蚕蛾科昆虫展现出鲜明的东洋区特征。同时该区域有46.67%的物种与古北界存在重叠。类似地,海南南丽湖国家湿地公园的蝴蝶分布也呈现出以东古北界与东洋界种分布类型为主[43];而在海南五指山保国村蝴蝶东洋界与古北界的共有种达到34种,占比达42.5%[44]。这些实例表明了海南昆虫东洋界与古北界之间的物种交流情况。就中国动物区系成分而言,海南在中国动物区系型中属于华南区,但黎母山景区的大蚕蛾科昆虫却与相邻的华中区、西南区拥有较多的共有成分。这种热带和亚热带特点的显著体现,很可能与这三个区域地理位置相邻、物种间相互渗透密切相关。这一现象进一步说明,黎母山地区的气候和生物学特性对昆虫物种的分布产生了深远影响。
作为我国首批5个海南热带雨林国家公园之一的黎母山,它是保护全球生物多样性的热点地区,具有重要的国家代表性和全球保护意义[45]。大蚕蛾是黎母山物种多样性的重要组成部分,黎母山景区大蚕蛾的寄主较为广泛,是重要的森林害虫之一,但实际上并未引发灾害性后果。相反,绝大多数大蚕蛾科昆虫在维护保护区生态系统的稳定方面发挥着重要作用,并具备潜在的经济价值。因此,对于黎母山景区的大蚕蛾科昆虫,我们不能仅仅将其视为害虫而加以控制,还应将其视为宝贵的资源昆虫,采取科学的管理和合理的开发利用策略。这要求我们不仅要保护该区域的大蚕蛾多样性,还要加强对大蚕蛾赖以生存和繁衍的自然资源和环境的保护力度。通过合理开发昆虫资源,我们可以促进森林生态系统的良性循环,实现生态、经济和社会的可持续发展。
致谢:感谢原浙江大学昆虫科学研究所张传溪教授对物种鉴定给予的指导。
Species Diversity and Annual Dynamics of Saturniidae in Limushan
-
摘要: 本研究针对海南热带雨林国家公园的黎母山景区进行大蚕蛾科的种类调查。通过2020–2024年的灯诱和踏查,共记录到黎母山大蚕蛾科15种,涵盖2个亚科、9属,其中有3个特有种。通过分析大蚕蛾的飞行期和上灯时间,发现大蚕蛾的活跃期主要集中在11月至8月之间,有56.34%的个体在03:00–06:00时间段被诱集。结合饲养记录,初步判断一化性昆虫有6种,二化性昆虫有2种,多化性昆虫有7种。此外,黎母山大蚕蛾科的区系特征与热带区系高度吻合,进一步证实了该地区独特的生态环境和丰富的生物多样性。本研究为进一步的物种保护和资源利用提供了科学依据。Abstract: This study investigated the diversity and phenology of Saturniidae moths in Limushan, Hainan Tropical Rainforest National Park. Through light trapping and transect surveys from 2020 to 2024, we recorded 15 Saturniidae species comprising 2 subfamilies and 9 genera, including 3 Hainan-endemic species. Analysis of flight periods revealed peak activity between November and the following August, with 56.34% of moths showing peak phototactic response at 03:00–06:00. Laboratory rearing indicated 6 univoltine species, 2 bivoltine species, and 7 multivoltine species. The faunal composition of Saturniidae in Limushan aligned with tropical zoogeographic regions, reflecting the area's distinctive ecological environment and biodiversity. These findings offer critical data for future species conservation and resource utilization .
-
图 3 黎母山大蚕蛾科昆虫不同物种的飞行期分布与数量
Fig. 3 Flight period distribution and abundance of Saturniidae in Limushan
图 4 黎母山大蚕蛾科昆虫不同时间段的上灯频率
Fig. 4 Light-attraction frequency of Saturniidae at different time periods in Limushan
表 1 黎母山15种大蚕蛾物种名录及国内分布
Table 1 Checklist and domestic distribution of 15 Saturniidae species in Limushan
亚科/属
Subfamily/Gene种
Species国内分布
Distribution in China大蚕蛾亚科 Saturniinae 大蚕蛾属
Attacus乌桕大蚕蛾
Attacus atlas海南、贵州、云南、香港、江西、福建、广东、广西、湖南、台湾[8, 12, 17–18] 樗蚕蛾属
Samia王氏樗蚕
Samia wangi广东、湖南、江西、福建、香港、台湾、海南、海南、西藏、四川、陕西、浙江[27] 小字大蚕蛾属
Cricula海南小字大蚕蛾
Cricula hainanensis海南特有种[13] 树天蚕属
Lemaireia海南树天蚕
Lemaireia hainana海南特有种[14] 霖王蛾属
Rinaca藏霖王蛾
Rinaca thibeta海南、云南、福建[8, 17] 希霖王蛾
Rinaca simla福建、四川、广东、海南、云南[8, 12, 18] 目大蚕蛾属
Saturnia樟蚕
Saturnia pyretorum吉林、河南、陕西、甘肃、湖北、浙江、安徽、海南、贵州、内蒙古、黑龙江、辽宁、江苏、四川、广东、广西、河北、山东、湖南、福建、江西、云南、湖北、台湾[8, 12, 17 − 19] 长翅樟蚕
Saturnia roseata重庆、海南[28] 尾大蚕蛾属
Actias华尾大蚕蛾
Actias sinensis陕西、甘肃、广东、海南、江西、湖南、广西、台湾[8, 12, 17 − 19] 绿尾大蚕蛾
Actias selene陕西、甘肃、海南、江苏、江西、河南、河北、浙江、湖南、广东、福建、广西、台湾、云南、西藏、辽宁、湖北、四川、吉林[8, 12, 18 − 20] 豹大蚕蛾属
Loepa黄豹大蚕蛾
Loepa katinka河南、河北、宁夏、西藏、安徽、浙江、福建、江西、四川、广东、广西、云南、海南[8, 12, 20] 红豹大蚕蛾
Loepa anthera四川、海南、西藏、福建、广东、广西、云南[8, 12, 17–18] 柞蚕属
Antheraea钩翅柞蚕
Antheraea assamensis广东、云南、海南[8, 12, 17–18] 明目柞蚕
Antheraea frithi陕西、湖南、福建、西藏、云南、海南[8, 17, 19, 21] 猫目大蚕蛾亚科 Salassinae 猫目大蚕蛾属
Salassa海南鸮目大蚕蛾
Salassa shuyiae海南特有种[15]  下载: 导出CSV
下载: 导出CSV
表 3 黎母山大蚕蛾在世界动物区系和中国动物区系中的分布型及占比
Table 3 Distribution patterns and proportions of Saturniidae from Limushan in global and Chinese zoogeographical regions
序号
Number分布型
Distribution Patterns物种数
Species numbers占比
Total Species Percentage(%)世界动物区划 1 东洋界-古北界 8 53.33% 2 东洋界-古北界 7 46.67% 中国动物区划 1 华南区 3 20% 2 华南区-华中区 1 6.67% 3 华南区-华中区-西南区 5 33.33% 4 华南区-华中区-西南区-华北区 1 6.67% 5 华南区-华中区-西南区-华北区-青藏区 2 13.34% 6 西南区-华南区-华中区-华北区-蒙新区-青藏区 1 6.67% 7 西南区-华南区-华中区-东北区-华北区-蒙新区 1 6.67% 8 西南区-华南区-华中区-东北区-华北区-蒙新区-青藏区 1 6.67%
下载: 导出CSV
表 2 黎母山大蚕蛾科昆虫区系
Table 2 Fauna of Saturniidae in Limushan
序号
number种名
species name世界动物区划
Zoogeographic fauna of the world中国动物区划
Zoogeographic fauna of ChinaA B C D E F a b c d e f g 1 乌桕大蚕蛾 √ √ √ √ √ 2 王氏樗蚕 √ √ √ √ √ √ √ 3 海南小字大蚕蛾 √ √ 4 海南树天蚕 √ √ 5 藏霖王蛾 √ √ √ √ 6 希霖王蛾 √ √ √ √ √ 7 樟蚕 √ √ √ √ √ √ √ √ 8 长翅樟蚕 √ √ √ 9 华尾大蚕蛾 √ √ √ √ √ 10 绿尾大蚕蛾 √ √ √ √ √ √ √ √ √ 11 黄豹大蚕蛾 √ √ √ √ √ √ √ √ 12 藤豹大蚕蛾 √ √ √ √ 13 钩翅柞蚕 √ √ √ √ 14 明目柞蚕 √ √ √ √ √ √ √ 15 海南鸮目大蚕蛾 √ √ 注:“√”表示有分布,A为古北界,B为东洋界,C为新北界,D为非洲界,E为新热带区,F为澳洲界;a为西南区,b为华中区,c为华南区,d为东北区,e为华北区,f为蒙新区,g为青藏区
下载: 导出CSV
表 4 15种大蚕蛾昆虫不同地区飞行期及化性
Table 4 Flight periods and voltinism of 15 Saturniidae from Limushan compared to those in other regions
物种
Species飞行期及化性 Flight period and Voltinism 国内其他地区 海南(黎母山) 乌桕大蚕蛾 Attacus atlas 中国南部、香港、台湾,4–9月,一化或二化性[17];
江西、福建,4–5月及7–8月,二化性[18]3–9月,多化性 王氏樗蚕 Samia wangi 未见记录 3–9月,多化性 海南小字大蚕蛾 Cricula hainanensis 未见记录 几乎全年可见,多化性 海南树天蚕 Lemaireia hainana 未见记录 10月至次年1月,一化性 藏霖王蛾 Rinaca thibeta 西南、华南,10–11月,一化性[17] 10月至次年1月,一化性 希霖王蛾 Rinaca simla 未见记录 10月至次年1月,一化性 樟蚕 Saturnia pyretorum 台湾北部及海南,12月及次年1–2月,一化性[18];
河北、江西、福建、广西、广东,三月中旬[21];
2–4月,一化性[17]10月至次年1月,一化性 长翅樟蚕 Saturnia roseata 未见记录 10月至次年1月,一化性 华尾大蚕蛾 Actias sinensis 中国南部、西南、香港、台湾,4–9月[17];
江西、湖南,4月,二化性[18]几乎全年可见,多化性 绿尾大蚕蛾 Actias selene 中国大部分地区,4–10月,二化或三化[17];
河北、河南、浙江、湖南、江西、广东、福建、台湾,
5、7、9月,二化或三化[18]几乎全年可见,多化性 黄豹大蚕蛾 Loepa katinka 华南、西南,4月[17] 11月至次年3月,二化性 藤豹大蚕蛾 Loepa anthera 广东、广西、云南,4–9月,二化性[17];
福建,7月[18]3–8月,二化性 钩翅柞蚕(琥珀蚕)Antheraea assamensis 南部、西南部,4–8月,二化性[17];
云南,4月出现[18]3–9月,多化性 明目柞蚕 Antheraea frithi 中国南部、西南部,4–8月,二化性[17];
云南,5月[18, 21]3–9月,多化性 海南鸮目大蚕蛾 Salassa shuyiae 未见记录 10月至次年1月,一化性
下载: 导出CSV
-
[1] 熊桃. 基于生态保护的海南黎母山国家森林公园总体规划设计研究[D]. 长沙: 中南林业科技大学, 2019. [2] 涂志华, 周凌峰, 黄艳萍, 等. 海南黎母山热带云雾林3种优势树种根际土壤微生物量季节动态及根际效应[J]. 生态科学, 2023, 42(2): 119 − 126. doi: 10.14108/j.cnki.1008-8873.2023.02.015 [3] 龚世平, 史海涛, 谢才坚, 等. 海南岛黎母山四眼斑水龟对春季生境的选择性[J]. 动物学研究, 2005, 26(2): 142 − 146. doi: 10.3321/j.issn:0254-5853.2005.02.006 [4] 陈加利, 徐诗涛, 廖建和, 等. 海南热带雨林沟谷植物群落区系地理研究[J]. 广东农业科学, 2015, 42(6): 130 − 135. doi: 10.16768/j.issn.1004-874x.2015.06.006 [5] FRIEDLANDER T P, PEIGLER R S, REGIER J C, et al. Monophyly, composition, and relationships within Saturniinae (Lepidoptera: Saturniidae): evidence from two nuclear genes[J]. Insect Systematics & Evolution, 2002, 33(1): 9 − 21. [6] 江秀均, 刘敏, 储一宁, 等. 红河州野生大蚕蛾科绢丝昆虫资源调查初报[J]. 中国蚕业, 2010, 31(4): 28 − 31. doi: 10.16839/j.cnki.zgcy.2010.04.010 [7] DEVI B, CHUTIA M, BHATTACHARYYA N. Food plant diversity, distribution, and nutritional aspects of the endemic golden silk producing silkworm, Antheraea assamensis - a review[J]. Entomologia Experimentalis et Applicata, 2021, 169(3): 237 − 248. doi: 10.1111/eea.13021 [8] 朱弘复, 王林瑶. 中国动物志 昆虫纲 第5卷 鳞翅目 蚕蛾科、大蚕蛾科、网蛾科[M]. 北京: 科学出版社, 1996. [9] ZHANG Z Q. Animal biodiversity: an outline of higher-level classification and survey of taxonomic richness[M]. Auckland: Magnolia Press, 2011. [10] SANTOS F L, CASAGRANDE M M, MIELKE O H H. Saturniidae and Sphingidae (Lepidoptera, Bombycoidea) assemblage in Vossoroca, Tijucas do Sul, Paraná, Brazil[J]. Anais da Academia Brasileira de Ciências, 2015, 87(2): 843 − 860. doi: 10.1590/0001-3765201520140368 [11] KITCHING I J, ROUGERIE R, ZWICK A, et al. A global checklist of the Bombycoidea (Insecta: Lepidoptera)[J]. Biodiversity Data Journal, 2018, 6: e22236. doi: 10.3897/BDJ.6.e22236 [12] 黄复生. 海南森林昆虫[M]. 北京: 科学出版社, 2002. [13] Brechlin R. Drei neue Saturniiden (Lepidoptera) aus China[J]. Cricula australosinica, 2004: 17 − 25 [14] NÄSSIG W A, WANG M. First record of the saturniid genus Lemaireia Nässig & Holloway, 1987 from Hainan island (PR China) with the description of a new species (Lepidoptera: Saturniidae)[J]. Nachrichten des Entomologischen Vereins Apollo, Frankfurt am Main, Neue Folge, 2006, 27(1/2): 23 − 25. [15] ZHANG W W, KOHLL S. Salassa shuyiae n. sp. , a new giant silkmoth from Hainan, China (Lepidoptera, Saturniidae, Salassinae)[J]. Nachrichten des entomologischen Vereins Apollo, Frankfurt am Main, Neue Folge, 2008, 29(1/2): 47 − 52. [16] 大卫·卡特. 蝴蝶与蛾: 世界各地500多种蝴蝶与蛾的彩色图鉴[M]. 北京: 中国友谊出版公司, 1997. [17] 吴云. 中华美蛾[M]. 郑州: 河南科学技术出版社, 2017. [18] 中国科学院动物研究所. 中国蛾类图鉴Ⅳ[M]. 北京: 科学出版社, 1981. [19] 杨星科, 薛大勇. 秦岭昆虫志, 8 鳞翅目. 大蛾类[M]. 西安: 世界图书出版社, 2017. [20] 方育卿. 庐山蝶蛾志[M]. 南昌: 江西高校出版社, 2003. [21] 朱弘复. 蛾类图册[M]. 北京: 科学出版社, 1975. [22] 彩万志. 拉英汉昆虫学词典[M]. 郑州: 河南科学技术出版社, 2022. [23] 李再霖, 彭锋, 王兴民, 等. 潮州单丛茶区不同海拔瓢虫科昆虫群落结构与多样性[J]. 茶叶科学, 2025, 45(1): 99 − 109. doi: 10.13305/j.cnki.jts.2025.01.012 [24] 中国科学院自然区划工作委员会. 中国动物地理区划与中国昆虫地理区划[M]. 北京: 科学出版社, 1959. [25] 章士美, 赵泳祥. 中国农林昆虫地理分布[M]. 北京: 中国农业出版社, 1996. (3-7, 297, 298 [26] 张荣祖. 中国动物地理[M]. 北京: 科学出版社, 1999. [27] NAUMANN S, PEIGLER R S. Four new species of the silkmoth genus Samia (Lepidoptera: Saturniidae)[J]. Nachrichten des Entomologischen Vereins Apollo, Frankfurt am Main, Neue Folge, 2001, 22(2): 75 − 83. [28] BRECHLIN R. Zwei neue Arten des genus (Subgenus) Saturnia Schrank, 1802 (Lepidoptera: Saturniidae)[J]. Entomo-Satsphingia, 2009, 2(2): 37 − 42. [29] DEUTSCH C A, TEWKSBURY J J, TIGCHELAAR M, et al. Increase in crop losses to insect pests in a warming climate[J]. Science, 2018, 361(6405): 916 − 919. doi: 10.1126/science.aat3466 [30] OUTHWAITE C L, MCCANN P, NEWBOLD T. Agriculture and climate change are reshaping insect biodiversity worldwide[J]. Nature, 2022, 605(7908): 97 − 102. doi: 10.1038/s41586-022-04644-x [31] WAGNER D L, GRAMES E M, FORISTER M L, et al. Insect decline in the Anthropocene: death by a thousand cuts[J]. Proceedings of the National Academy of Sciences of the United States of America, 2021, 118(2): e2023989118. doi: 10.1073/pnas.2023989118 [32] SÁNCHEZ-BAYO F, WYCKHUYS K A G. Worldwide decline of the entomofauna: a review of its drivers[J]. Biological Conservation, 2019, 232: 8 − 27. doi: 10.1016/j.biocon.2019.01.020 [33] WERMEILLE E, CHITTARO Y, GONSETH Y. Rote Liste der Tagfalter und Widderchen: Papilionoidea, Hesperioidea und Zygaenidae: Gefährdete Arten der Schweiz, stand 2012[M]. Bern: Bundesamt für Umwelt BAFU, 2014. [34] 刘立春. 昆虫趋光行为的初步观察[J]. 南京农业大学学报, 1982, 5(2): 52 − 59. [35] 桑文, 蔡夫业, 王小平, 等. 农用诱虫灯田间应用现状与展望[J]. 中国植保导刊, 2018, 38(10): 26 − 30. doi: 10.3969/j.issn.1672-6820.2018.10.004 [36] 杨菁菁, 梁朝巍, 沈斌斌, 等. 昆虫扑灯节律研究[J]. 安徽农业科学, 2012, 40(1): 210 − 212. doi: 10.13989/j.cnki.0517-6611.2012.01.179 [37] 周阳, 权子豪, 段曼微, 等. 基于林分昆虫多样性调查的传统灯诱法改进效果分析[J]. 生态学报, 2023, 43(16): 6851 − 6861. doi: 10.5846/stxb202205131348 [38] NOWINSZKY L, PUSKÁS J. Sex ratio analysis of some Macrolepidoptera species collected by Hungarian forestry light traps[J]. Acta Silvatica et Lignaria Hungarica, 2015, 11(2): 99 − 110. doi: 10.1515/aslh-2015-0008 [39] 徐练. 异色瓢虫、玉米螟和烟粉虱的趋光性研究[D]. 长沙: 湖南农业大学, 2016. [40] 刘寿明, 谭放军, 周程爱. 性信息素对灯光诱捕甜菜夜蛾效果的影响[J]. 湖南农业科学, 2022(3): 64 − 66. doi: 10.16498/j.cnki.hnnykx.2022.003.014 [41] 程文杰, 郑霞林, 王攀, 等. 昆虫趋光的性别差异及其影响因素[J]. 应用生态学报, 2011, 22(12): 3351 − 3357. doi: 10.13287/j.1001-9332.2011.0472 [42] 张锦芳, 张阳, 徐文平, 等. 昆虫的趋光性及其应用于害虫治理的研究进展[J]. 世界农药, 2020, 42(11): 26 − 35. doi: 10.16201/j.cnki.cn10-1660/tq.2020.11.04 [43] 肖贝文, 叶琳雄, 胡荣, 等. 海南南丽湖国家湿地公园蝴蝶种类及区系分析[J]. 热带生物学报, 2017, 8(3): 341 − 347. doi: 10.15886/j.cnki.rdswxb.2017.03.015 [44] 林树鹏, 翁小理, 李丽敏, 等. 海南五指山保国村蝴蝶调查及区系分析[J]. 热带农业科学, 2022, 42(2): 42 − 47. doi: 10.12008/j.issn.1009-2196.2022.02.007 [45] 贾东雨. 海南热带雨林国家公园生态旅游研究[J]. 旅游纵览, 2024(11): 103 − 105. doi: 10.3969/j.issn.1004-3292.2024.11.034 -

 点击查看大图
点击查看大图
计量
- 文章访问数: 739
- HTML全文浏览量: 465
- 被引次数: 0
