

-
豆大蓟马(Megalurothrips usitatus Bagnall)又称普通大蓟马或豆花蓟马,是豆科作物上的重要害虫,尤其对海南、广西、广东、贵州等地的豇豆造成了严重的危害和经济损失,成为这些地区豇豆生产的重要限制因素[1]。近年来,海南豇豆种植面积稳定在2万hm2以上,产量近60万t,仅次于辣椒,是海南冬季瓜菜的主要栽培品种之一[2]。蓟马早期危害可造成豇豆叶片皱卷、生长点坏死,开花结荚期受害可造成大量落花减产和豆荚“黑化”,严重影响产量和品质[3]。为了有效防控蓟马,已开展了大量有益的研究探索,如化学防治[4-7]、理化诱控防治[8-13]、生物防治[14-16]以及农业防治[17]等,但是目前生产上主要还是依赖于化学防治。由于豆大蓟马自身特性如虫体微小、隐匿危害、繁殖力强、发育历期短、世代重叠严重以及土壤中化蛹等,常造成对其的化学防治效果不理想,而高频次高剂量地施用化学农药可导致豆大蓟马对多种常用杀虫剂产生了不同程度的抗性[18-19]。提高防效和延缓抗药性产生的重要途径是科学地进行化学防治。科学化学防治一是要找准防治时期,二是要合理地选用杀虫剂。不同杀虫剂品种对豆大蓟马不同虫态有着较大的毒力差异[20],因此,需要根据豆大蓟马田间发生的主要虫态来选择性地使用杀虫剂,这就要求掌握不同杀虫剂对蓟马各个虫态的毒性。多种杀虫剂对豆大蓟马成虫和若虫的毒力[19-22] 已见报道,但未见对其卵和蛹的毒力报道。基于此,笔者选择生产上多种常用杀虫剂对豆大蓟马的卵和蛹进行室内毒力测定。
种群动态和体型特征是昆虫生态学研究的重要内容,对制定害虫防治策略具有重要意义。已有研究报道豆大蓟马在豇豆上的空间分布[23]、色板诱集[3,11-12]、踏查[24]和基于性比的成虫时空动态[25]研究。关于昆虫在体型上的差异及其影响方面的研究表明,体型大小与昆虫的生殖能力和抗逆性具有相关性,从而对害虫的种群动态变化产生影响[26-28]。目前关于豆大蓟马在海南的全年发生动态以及地理种群体型大小还缺乏系统的调查研究。因此,笔者以海南豇豆主产区澄迈县(海南北部)和三亚市(海南南部)为代表调查了豆大蓟马在海南豇豆的全年发生规律及不同地理位置种群的体型大小,以期为豆大蓟马的综合防治提供理论依据。
-
用于体型大小测定的豆大蓟马成虫分别在2022年4月采集于澄迈县大丰镇信宜村(19°51′N;110°2′E)、在2021年1月采集于三亚市崖州区南滨农场(18°22′N;109°11′E)、在2021年4月采集于儋州市宝岛新村豇豆田(19°30′N;109°29′E),室内饲养一代后的3日龄成虫用于体型大小测量。用于毒力测定的豆大蓟马虫源采自海南省澄迈县永发镇豇豆田,在室内用豇豆豆荚饲养多代后用于实验;饲养方法参照唐良德等[20]的方法。
-
97%高效氯氟氰菊酯(郑州农田化工有限公司),76%甲氨基阿维菌素苯甲酸盐(简称甲维盐,河北天顺生物工程有限公司),92%阿维菌素(南通农药剂型开发中心),96%吡虫啉(四川国光农化股份有限公司),96%啶虫脒(石家庄伊宏化工有限公司),97%溴虫腈、95%溴氰虫酰胺(湖北康宝泰精细化工有限公司),96%螺虫乙酯(广西南宁绿丰化工有限公司),上述药剂均为原药;99%绿颖矿物油(韩国SK杰克株式会社,市售),乙基多杀菌素(艾绿士® 60 g·L−1悬浮剂,美国科迪华公司,市售)。
-
根据海南南部冬季种植和北部冬春种植为主全年均可种植豇豆的特点,南部选择在三亚市崖州区南滨农场不定期对豇豆花期豆大蓟马进行随机采样调查(踏查),北部选择在澄迈县大丰镇全年种植豇豆调查豆大蓟马的发生动态,并对全年气温进行监测。采用小米智能温湿计(型号:LYWSD03MMC)每小时记录1次温度,以每日24时温度的平均值为当天平均气温。采用随机采样调查法,每次调查30朵花(芽)或叶片,将采集的花朵一一装入封口袋内带回实验室进行鉴定和计数,现场计数叶片和花芽上的豆大蓟马,以平均每花或每叶的蓟马数量绘制种群发生动态图。
-
体长和体宽是昆虫个体大小的度量[29-30]。采用基恩士(KEYENCE)超景深显微镜(型号:VHX-7000)对三亚、澄迈和儋州豆大蓟马雌雄个体分别进行体长和体宽的测量。测量虫态为各地理位置种群饲养一代后的3日龄成虫,测量前将待测蓟马样品置于−20℃冰箱中冷冻处理使其失去活动能力。
-
卵的毒力测定方法采用浸渍法。在玻璃瓶(200 mL)底部垫一层粗糙的手纸,放置一节长约5 cm的豇豆豆荚(切口两端平整不留豆粒孔),然后每瓶接入20头5日龄的蓟马雌成虫,用74 μm防虫网和橡皮筋扎紧封口。产卵24 h后,将豆荚取出放入预先配制好的药液中,浸渍10 s后取出转移到垫有滤纸的培养皿(Ф=9 cm)中,置于(26 ± 1)℃、光暗比14 h∶10 h、相对湿度65%的培养箱中,培养96 h后调查孵化的蓟马若虫数量,以清水浸渍为对照处理。因蓟马卵产于植物组织内无法观察,以孵化的若虫数表示卵的存活数,以清水对照处理卵孵化若虫数表示蓟马的产卵量[31-33]。每处理一节豆荚即一个重复,每个药剂设置5个浓度梯度,每个浓度4次重复。
蛹的毒力测定方法采用POTTER喷雾法,喷雾条件设定参照Tang等[34]报道的方法。在培养皿(Ф=6 cm)中分别接入20头蓟马蛹,置于POTTER 喷雾塔下喷雾(喷雾压强为100 kPa,沉降时间为10 s),每个处理喷雾量为1.5 mL,喷雾后将蓟马转移至预先铺垫一层湿润的粗糙手纸作为化蛹基质的培养皿(Ф=6 cm)中,用74 μm防虫网和橡皮筋扎紧封口。置于(26 ± 1)℃、光暗比14 h∶10 h、相对湿度65%的培养箱中,每48 h后观察记录蛹的存活情况,直至活虫全部羽化,以羽化数表示存活数。根据预试验,每个药剂配制5个浓度,每个浓度设4次重复,以喷清水处理为对照。
-
豆大蓟马的种群发生动态(数量)以及不同地理位置种群间体型大小的显著性差异在SPSS 17.0分析软件中采用Tukey法在0.05水平上进行比较。不同药剂对豆大蓟马卵和蛹的毒力测定实验中,因豆大蓟马卵产于植物组织内无法观察,以孵化的若虫数表示卵的存活数,卵的死亡数(响应数)以对照组卵孵化若虫数减去药剂处理组卵孵化若虫数计算。不同药剂对豆大蓟马的致死中浓度(LC50)采用SPSS分析软件中的Probit回归分析法计算。
-
由图1-A可知,豆大蓟马在豇豆的全生育期均可发生,以花期发生数量最大,苗期发生数量最低。豇豆属于循环花序植物,通常可采收2个花期,第2花期蓟马发生数量显著降低。从图中还可看出,第2茬和第3茬豇豆整个生育期气温明显高于第1茬,且豆大蓟马发生的种群数量也显著高于第1茬同期豆大蓟马发生的数量,表明气温显著影响豆大蓟马的发生。
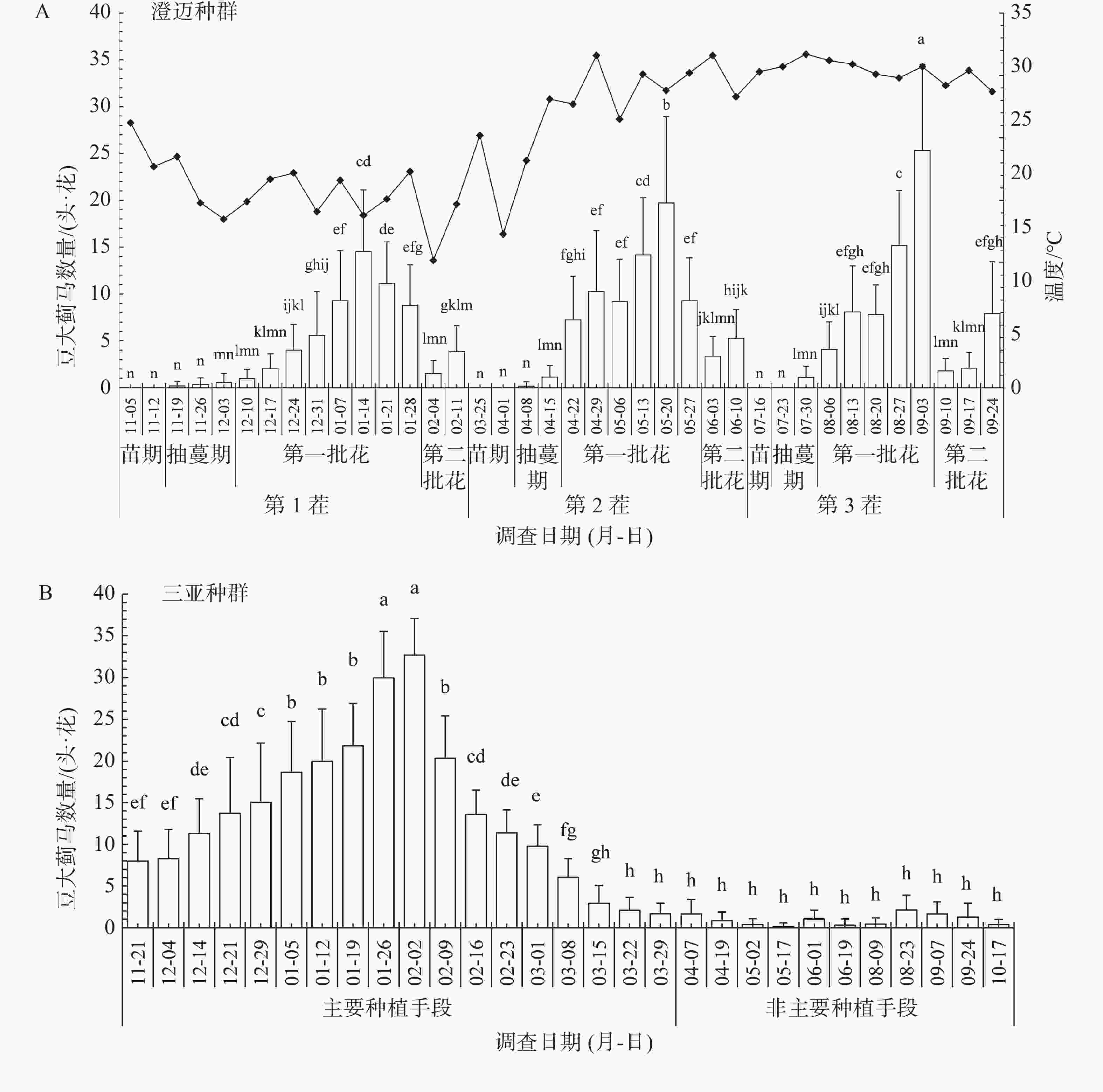
图 1 豆大蓟马在海南的种群发生动态
由图1-B可知,在主要种植时段(冬春季)豆大蓟马在豇豆上的发生数量呈明显单峰型分布,从12月中下旬到翌年的2月中下旬,豆大蓟马的种群一直维持在较高的数量水平(>10头·花),在1月下旬至2月初达到最高峰(约 30头·花)。在非主要种植时段(4月后)豆大蓟马数量迅速下降,并长时间维持在较低数量水平(<5头·花)。
-
通过对不同地理位置种群豆大蓟马体型的比较(表1),儋州种群的雌雄个体体长和体宽均显著大于三亚种群和澄迈种群,且三亚种群也显著大于澄迈种群。
表 1 豆大蓟马不同地理位置种群体型大小
样品 雌/雄虫 测量样
本数/只体长/μm 体宽/μm 三亚种群 雌虫 117 1493.59±12.18b 309.99±2.27b 澄迈种群 雌虫 99 1342.30±8.85c 203.19±1.53c 儋州种群 雌虫 109 1634.15±13.48a 318.35±2.22a 三亚种群 雄虫 114 1227.89±8.46b 221.29±1.66b 澄迈种群 雄虫 97 1072.41±8.73c 207.18±1.58c 儋州种群 雄虫 101 1315.08±10.53a 226.40±1.83a 注:同列不同字母数值间差异显著(P<0.05)。 -
不同药剂对豆大蓟马卵的毒力表现显著不同(表2)。其中,螺虫乙酯对豆大蓟马卵的活性最高(LC50=18.51 mg·L−1),乙基多杀菌素、溴氰虫酰胺和啶虫脒对豆大蓟马卵也表现较高的毒力,LC50分别为46.17、52.54和58.53 mg·L−1,其他测试药剂对豆大蓟马卵的活性较低(LC50 > 110 mg·L−1)。
表 2 不同药剂对豆大蓟马卵的毒力
药剂 毒力回归方程 LC50(mg·L−1) 95%置信区间值 拟合度 Chisquare df P 乙基多杀菌素 Y=−2.583+1.552x 46.17 37.34~55.05 0.079 3 0.994 高效氯氟氰菊酯 Y=−4.370+1.726x 340.18 294.27~391.49 2.812 3 0.422 螺虫乙酯 Y=−1.898+1.498x 18.51 14.16~22.77 4.661 3 0.198 甲维盐 Y=−3.290+1.532x 140.48 119.18~165.14 0.723 3 0.868 溴虫腈 Y=−3.570+1.733x 114.79 97.29~134.14 3.649 3 0.302 啶虫脒 Y=−3.093+1.750x 58.53 49.05~68.44 4.190 3 0.242 吡虫啉 Y=−4.188+1.959x 137.48 119.79~157.35 1.836 3 0.607 溴氰虫酰胺 Y=−2.800+1.628x 52.54 43.86~61.41 4.004 3 0.261 矿物油 Y=−5.761+1.669x 2835.21 2451.03~3294.32 4.194 3 0.241 不同药剂对豆大蓟马蛹的毒力也表现显著不同(表3)。乙基多杀菌素和溴虫腈对豆大蓟马蛹的活性最高,LC50分别为26.18 和27.71 mg·L−1,其次为高效氯氟氰菊酯(LC50=35.84 mg·L−1)和甲维盐(LC50=37.32 mg·L−1),再次是阿维菌素(LC50=56.31 mg·L−1)和溴氰虫酰胺(LC50=62.98 mg·L−1),新烟碱类杀虫剂吡虫啉和啶虫脒对豆大蓟马蛹的活性较低(90 mg·L−1 < LC50 < 115 mg·L−1)。
表 3 不同药剂对豆大蓟马蛹的毒力
药剂 毒力回归方程 LC50(mg·L−1) 95%置信区间 拟合度 Chisquare df P 乙基多杀菌素 Y=−2.436+1.718x 26.18 20.80~31.72 0.861 3 0.835 高效氯氟氰菊酯 Y=−2.829+1.820x 35.84 29.64~42.65 0.569 3 0.904 甲维盐 Y=−2.328+1.481x 37.32 29.70~45.84 0.639 3 0.887 阿维菌素 Y=−2.704+1.545x 56.31 44.00~69.17 2.618 3 0.454 溴虫腈 Y=−2.208+1.530x 27.71 21.57~34.12 3.508 3 0.320 啶虫脒 Y=−2.853+1.455x 91.36 72.37~112.61 0.839 3 0.840 吡虫啉 Y=−3.052+1.486x 113.41 91.63~139.44 0.734 3 0.865 溴氰虫酰胺 Y=−2.311+1.284x 62.98 47.40~79.77 2.859 3 0.414 -
明确害虫田间发生种群动态是开展害虫综合治理(IPM)的重要前提和基础。蓟马是典型的r-对策害虫,具有发育历期短、世代重叠严重、繁殖力强等特点,且既可营两性生殖又可营孤雌生殖,种群增长迅速。通过田间种群动态监测发现,豆大蓟马在海南全年均可发生,但受豇豆生育期显著影响,表现为花期发生数量显著大于其他生育期,与花期发生尤为严重的报道一致[3,35]。另外,在花期对蓟马监测发现在海南南部(三亚)豇豆主要种植时段发生数量大,非主要种植时段发生数量骤减,表明寄主及其生育期是影响豆大蓟马种群发生动态的重要因素;另一方面,昆虫作为一种变温动物,其发生又受到环境温度的显著影响[36-37],豆大蓟马在田间的种群发生动态也印证了这一现象。豆大蓟马在日平均气温30℃左右的环境下(第二茬和第三茬)种群发生数量显著高于同期温度下(< 25℃,第一茬)的种群数量。有研究表明豆大蓟马产卵量在30℃时最高,平均产卵量达到232.78粒·雌[38],这与本研究结果是十分吻合的。但温度过高,也不利于豆大蓟马的田间发生,正如本研究的调查结果发现在海南南部(三亚)豇豆的非主要种植时段,正值高温阶段,既不利于豇豆生长也不利于豆大蓟马的发生(图1-B)。
体型是昆虫最基本也是最重要特征之一,包括卵的大小、卵质量,蛹质量,成虫体长、体宽、体质量等,昆虫雌雄个体之间体型存在差异的现象被称为性体型二型性现象,同时这种现象存在着地理差异[27,39]。本研究发现豆大蓟马成虫不仅存在性体型二型性现象,而且发现各地理种群存在显著体型差异,体型从大到小依次为儋州种群 > 三亚种群 > 澄迈种群。有研究结果表明,环境因素在昆虫体型的地理差异中起到重要作用[40],包括温度、湿度、光照、光周期、寄主植物以及种群密度等[27-28]。本研究结果发现,豆大蓟马不同地理位置种群体型大小存在显著差异,但受何种因素调控还不得而知。此外,体型大小对昆虫的繁殖能力、竞争能力和抗逆性等生物学特性会产生重要影响[28,41]。总体来说,体型大的昆虫的繁殖能力更强,更具有交配选择优势,可提高后代的适合度[42];体型大的昆虫在食物的竞争、领地的侵占或保卫、天敌防御等方面均表现出一定的优势[43];体型大的昆虫的体内储存着相对较多的能量物质,对逆境的耐受能力也较强[44]。然而,本研究发现体型大的儋州种群田间发生并不严重,反而体型小的三亚种群和澄迈种群在田间发生更为严重(图1)。这可能是受当地的种植模式(儋州零星种植,三亚、澄迈规模种植)、寄主品种、环境因子以及人为干预(化学防治)等综合因素调控所致。豆大蓟马体型差异的生物学意义及其调控机制还有待于进一步的研究。
蓟马是缨翅目昆虫的总称,分为锥尾(锯尾)亚目和管尾亚目,属过渐变态昆虫,其生活史经历卵、若虫、蛹和成虫4个阶段。锥尾亚目具锯状产卵器将卵产于植物组织内,蛹通常在叶背面叶脉的交叉处、叶柄基部、萼片间、叶鞘间、果实凹陷处及枯枝落叶层或土壤等场所化蛹。豆大蓟马的卵在豇豆上主要产于花器、生长点和嫩荚中,老熟若虫转移到土壤中化蛹,这一习性给生产上防治豆大蓟马带来巨大挑战。在害虫整个生态控制体系中,需要各个环节均采取相应的措施,防治才能更有效和可持续的控制害虫的暴发危害[45]。生产上对豆大蓟马进行防治时,通常仅考虑肉眼可见的成虫和若虫,往往忽视了看不见的卵和蛹,缺乏防治的科学性和针对性,造成防效不理想。因此针对此类害虫防控时需要对其生活史的各个虫态采取有效措施,在对黄瓜作物上的西花蓟马和烟蓟马进行生物防治的例子很好地体现了这一防治理念[46-47]。本研究发现,乙基多杀菌素、螺虫乙酯有较好的杀卵效果,乙基多杀菌素、溴虫腈和高效氯氟氰菊酯有较好的灭蛹效果,这些杀虫剂对不同虫态的毒力差异与虫态习性、杀虫剂的作用方式紧密相关,蓟马的卵产在植物组织内,而化蛹则在土壤或落叶层中,因而杀卵就对药剂的内吸性或渗透性提出要求,而杀蛹则要求有较高的触杀活性[48-49]。另外,毒力差异也可能与毒力测定方法有关[50-51]。
种群动态监测对于豆大蓟马的田间用药防治具有很好的指导作用。通过调查发现,每一茬豇豆都存在明显的豆大蓟马发生高峰(图1-A),且高峰期均为豇豆花期,因而豇豆花期是化学防治豆大蓟马的主要时期。考虑到豆大蓟马世代发育历期短(30℃下为10.57 d)[36],因而花期豆大蓟马成虫数量大时,除了选用对成虫有较高活性的杀虫剂外,还需要选择具有高杀卵活性的药剂,做到既杀灭成虫又杀灭卵;而在若虫大量发生时,除了选用对若虫有较高活性的杀虫剂外,也需要选择对蛹有较高活性的药剂,做到若虫和蛹兼杀的效果。根据田间虫态发生规律选择防治药剂,有利于提高豆大蓟马防治的针对性,从而达到提高防效的目的,同时田间作业时要尽量对植株/地面垄沟喷雾均匀,以提高对豆大蓟马各虫态的防效。
-
豆大蓟马存在性体型二型性和多态性现象,在海南全年发生且是豇豆上的主要害虫,并受物候期和气温的双重影响。卵产于寄主组织中,在土层或落叶层化蛹,卵和蛹对化学杀虫剂表现出敏感性差异。其中,螺虫乙酯具有较高的杀卵活性,而乙基多杀菌素和溴虫腈具有较高的杀蛹活性,可推荐螺虫乙酯、乙基多杀菌素和溴虫腈作为豆大蓟马卵和蛹的田间防治药剂。
Occurrence of Megalurothrips usitatus in Hainan and preliminary screening of insecticides for control of its eggs and pupae
-
摘要: 为明确豆大蓟马(Megalurothrips usitatus)在海南的时空动态、种群体型差异及筛选高效卵和蛹杀灭剂,笔者系统调查了豆大蓟马在海南豇豆主产区的全年发生动态,测量了不同地理位置种群成虫的体型大小,并采用浸渍法和POTTER喷雾法分别测定了多种杀虫剂对豆大蓟马卵和蛹的室内毒力。结果表明,豆大蓟马在海南可全年发生,种群动态受寄主生育期和温度双重调控;儋州种群的雌雄个体的体长和体宽显著大于三亚种群和澄迈种群;螺虫乙酯对豆大蓟马卵具有较高的毒力(LC50=18.51 mg·L−1),而乙基多杀菌素和溴虫腈则对蛹具有较高的活性,LC50分别为26.18和27.71 mg·L−1,可推荐作为豆大蓟马卵和蛹的田间防治药剂。Abstract: To clarify the spatiotemporal dynamics of the thrips Megalurothrips usitatus Bagnall populations and the differences of the thrips populations in adult body size in Hainan, China and screen high effective insecticides for control of thrips eggs and pupae,a systematical investigation was made into the annual occurrence of M. usitatus on cowpea in Hainan, the adult body size of the M. usitatus populations at different geographical locations were measured, and the toxicity of various insecticides against the eggs and pupae of M. usitatus was tested by using the dip method and POTTER bioassay, respectively. The investigation showed that M. usitatus occured throughout the year in Hainan. Population dynamics were significantly regulated by host phenology and temperature. The adult body size measurement showed that the adult body length and body width of M. usitatus populations were significantly higher in Danzhou than in Sanya and Chengmai. The toxicity test showed that spirotetramat had a higher ovicidal activity (LC50=18.51 mg·L−1) against M. usitatus eggs, while spinosad and chlorfenapyr had a higher activity against M. usitatus pupae, with their LC50 values being 26.18 and 27.71 mg·L−1, respectively. These insecticides could hence be recommended as candidates for controlling M. usitatus eggs and pupae in the field.
-
Key words:
- Megalurothrips usitatus Bagnall /
- body size /
- insecticide /
- LC50 /
- Hainan
-
表 1 豆大蓟马不同地理位置种群体型大小
样品 雌/雄虫 测量样
本数/只体长/μm 体宽/μm 三亚种群 雌虫 117 1493.59±12.18b 309.99±2.27b 澄迈种群 雌虫 99 1342.30±8.85c 203.19±1.53c 儋州种群 雌虫 109 1634.15±13.48a 318.35±2.22a 三亚种群 雄虫 114 1227.89±8.46b 221.29±1.66b 澄迈种群 雄虫 97 1072.41±8.73c 207.18±1.58c 儋州种群 雄虫 101 1315.08±10.53a 226.40±1.83a 注:同列不同字母数值间差异显著(P<0.05)。  下载: 导出CSV
下载: 导出CSV
表 2 不同药剂对豆大蓟马卵的毒力
药剂 毒力回归方程 LC50(mg·L−1) 95%置信区间值 拟合度 Chisquare df P 乙基多杀菌素 Y=−2.583+1.552x 46.17 37.34~55.05 0.079 3 0.994 高效氯氟氰菊酯 Y=−4.370+1.726x 340.18 294.27~391.49 2.812 3 0.422 螺虫乙酯 Y=−1.898+1.498x 18.51 14.16~22.77 4.661 3 0.198 甲维盐 Y=−3.290+1.532x 140.48 119.18~165.14 0.723 3 0.868 溴虫腈 Y=−3.570+1.733x 114.79 97.29~134.14 3.649 3 0.302 啶虫脒 Y=−3.093+1.750x 58.53 49.05~68.44 4.190 3 0.242 吡虫啉 Y=−4.188+1.959x 137.48 119.79~157.35 1.836 3 0.607 溴氰虫酰胺 Y=−2.800+1.628x 52.54 43.86~61.41 4.004 3 0.261 矿物油 Y=−5.761+1.669x 2835.21 2451.03~3294.32 4.194 3 0.241
下载: 导出CSV
表 3 不同药剂对豆大蓟马蛹的毒力
药剂 毒力回归方程 LC50(mg·L−1) 95%置信区间 拟合度 Chisquare df P 乙基多杀菌素 Y=−2.436+1.718x 26.18 20.80~31.72 0.861 3 0.835 高效氯氟氰菊酯 Y=−2.829+1.820x 35.84 29.64~42.65 0.569 3 0.904 甲维盐 Y=−2.328+1.481x 37.32 29.70~45.84 0.639 3 0.887 阿维菌素 Y=−2.704+1.545x 56.31 44.00~69.17 2.618 3 0.454 溴虫腈 Y=−2.208+1.530x 27.71 21.57~34.12 3.508 3 0.320 啶虫脒 Y=−2.853+1.455x 91.36 72.37~112.61 0.839 3 0.840 吡虫啉 Y=−3.052+1.486x 113.41 91.63~139.44 0.734 3 0.865 溴氰虫酰胺 Y=−2.311+1.284x 62.98 47.40~79.77 2.859 3 0.414
下载: 导出CSV
-
[1] 卢维海, 韦莹军, 谭道朝, 等. 海南毒豇豆事件对广西植保的启示[J]. 广西农学报, 2010, 25(2): 86 − 87. [2] 海南省统计局, 国家统计局海南调查总队.海南统计年鉴[M]. 北京: 中国统计出版社, 2021: 280. [3] 唐良德, 梁延坡, 韩云, 等. 海南豇豆蓟马发生为害调查及蓝板监测技术研究[J]. 中国植保导刊, 2015, 35(3): 53 − 57. [4] 李现玲, 宋娜. 60 g•L-1乙基多杀菌素悬浮剂防治豇豆蓟马田间药效试验[J]. 广西植保, 2012, 25(3): 18 − 19. [5] 陈剑山, 李鹏, 刘奎, 等. 3 种药剂对豇豆蓟马的田间防效评价[J]. 中国植保导刊, 2015, 35(5): 66 − 57. [6] 唐良德, 林军, 付步礼, 等. 豇豆闭花前后施药对防治蓟马效果的影响[J]. 中国植保导刊, 2016, 36(3): 66 − 67. [7] 沈登荣, 何超, 曾通明, 等. 乙基多杀菌素与4种杀虫剂复配对普通大蓟马的毒力测定[J]. 西南师范大学学报(自然科学版), 2017, 42(12): 53 − 57. [8] CHANG N T. Color preference of thrips (Thysanoptera: Thripidae) in the adzuki bean field [J]. Plant Protection Bulletin, 1990, 32(4): 307 − 316. [9] 罗丰, 袁廷庆, 柯用春, 等. 不同颜色防虫网对豇豆生长特性、产量及蓟马发生量的影响[J]. 南方农业学报, 2014, 45(9): 1584 − 1588. [10] 张瑞敏, 付步礼, 邱海燕, 等. 防虫网对豇豆主要害虫的阻隔作用研究[J]. 中国植保导刊, 2015, 35(11): 36 − 38. doi: 10.3969/j.issn.1672-6820.2015.11.009 [11] TANG L D, ZHAO H Y, FU B L, et al. Colored sticky traps to selectively survey thrips in cowpea ecosystem [J]. Neotropical Entomology, 2016, 45(1): 96 − 101. doi: 10.1007/s13744-015-0334-1 [12] 闫凯莉, 唐良德, 吴建辉. 普通大蓟马对不同颜色的趋性及日节律调查[J]. 应用昆虫学报, 2017, 54(4): 639 − 645. [13] 李晓维, 罗雪君, 王丽坤, 等. 普通大蓟马聚集信息素的分离和鉴定[J]. 昆虫学报, 2019, 62(9): 1017 − 1027. [14] 唐良德, 王晓双, 赵海燕, 等. 大草蛉幼虫捕食豆大蓟马和豆蚜的功能反应及生长发育[J]. 中国生物防治学报, 2017, 33(1): 49 − 55. doi: 10.16409/j.cnki.2095-039x.2017.01.008 [15] LIU P P, JIA W T, ZHENG X, et al. Predation functional response and life table parameters of Orius sauteri (Hemiptera: Anthocoridae) feeding on Megalurothrips usitatus (Thysanoptera: Thripidae) [J]. Florida Entomologist, 2018, 101(2): 254 − 259. doi: 10.1653/024.101.0216 [16] DU C L, YANG B, WU J H, et al. Identification and virulence characterization of two Akanthomyces attenuatus isolates against Megalurothrips usitatus (Thysanoptera: Thripidae) [J]. Insects, 2019, 10(6): 168. doi: 10.3390/insects10060168 [17] 黄伟康, 孔祥义, 柯用春, 等. 普通大蓟马的研究进展[J]. 中国蔬菜, 2018(2): 21 − 27. [18] 唐良德, 赵海燕, 付步礼, 等. 海南地区豆大蓟马田间种群的抗药性监测[J]. 环境昆虫学报, 2016, 38(5): 1032 − 1037. [19] 唐良德, 赵海燕, 付步礼, 等. 海南普通大蓟马抗药性监测及对6 种杀虫剂的敏感性[J]. 环境昆虫学报, 2018, 40(5): 1175 − 1181. [20] 唐良德, 付步礼, 邱海燕, 等. 豆大蓟马对12种杀虫剂的敏感性测定[J]. 热带作物学报, 2015, 36(3): 570 − 574. [21] 孔祥义, 肖春雷, 刘勇, 等. 5种药剂对蓟马的室内毒力测定及防治效果研究[J]. 广东农业科学, 2012, 39(20): 70 − 72. doi: 10.3969/j.issn.1004-874X.2012.20.025 [22] 肖春雷, 刘勇, 吴青君, 等. 不同药剂对三亚地区豇豆上普通大蓟马的毒力[J]. 植物保护, 2014, 40(6): 164 − 166. doi: 10.3969/j.issn.0529-1542.2014.06.031 [23] 范咏梅, 童晓立, 高良举, 等. 普通大蓟马在海南豇豆上的空间分布型[J]. 环境昆虫学报, 2013, 35(6): 737 − 743. [24] 袁成明, 郅军锐, 李景柱, 等. 贵州省蔬菜蓟马的种类、分布及综合防治[J]. 湖北农业科学, 2008, 47(12): 1442 − 1444. doi: 10.3969/j.issn.0439-8114.2008.12.021 [25] 罗亚丽. 普通大蓟马性比的时空动态及温度和杀虫剂的干扰效应[D]. 海口: 海南大学, 2020. [26] 张礼生, 陈红印, 李保平. 天敌昆虫扩繁与应用[M]. 北京: 中国农业科学技术出版社, 2014: 69 − 70. [27] 匡先钜, 戈峰, 薛芳森. 昆虫体型及性体型二型性的地理变异[J]. 昆虫学报, 2015, 58(3): 351 − 360. [28] 黄韵姗, 张静宇, 蒋明星. 昆虫个体大小对其种群生物学的影响[J]. 生态学报, 2017, 37(7): 2158 − 2168. [29] ROGERS L E, HINDS W T, BUSCHBOM R L. A general weight vs. length relationship for insects [J]. Annals of the Entomological Society of America, 1976, 69: 387 − 389. doi: 10.1093/aesa/69.2.387 [30] GRUNER D S. Regressions of length and width to predict arthropod biomass in the Hawaiian Islands [J]. Pacific Science, 2003, 57: 325 − 336. doi: 10.1353/psc.2003.0021 [31] VAN RIJN P C J, MOLLEMA C, STEENHUIS-BROERS G M. Comparative life history studies of Frankliniella occidentalis and Thrips tabaci (Thysanoptera: Thripidae) on cucumber [J]. Bulletin of Entomological Research, 1995, 85: 285 − 297. doi: 10.1017/S0007485300034386 [32] ZHANG Z J, WU Q J, LI X F, et al. Life history of western flower thrips, Frankliniella occidentalis (Thysan., Thripae), on five different vegetable leaves [J]. Journal of Applied Entomology, 2007, 131: 347 − 354. doi: 10.1111/j.1439-0418.2007.01186.x [33] PARK C G, KIM H Y, LEE J H. Parameter estimation for a temperature-dependent development model of Thrips palmi Karny (Thysanoptera: Thripidae) [J]. Journal of Asia-Pacific Entomology, 2010, 13: 145 − 149. doi: 10.1016/j.aspen.2010.01.005 [34] TANG L D, QIU B L, CUTHBERTSON A G S, et al. Status of insecticide resistance and selection for imidacloprid resistance in the ladybird beetle Propylaea japonica (Thunberg) [J]. Pesticide Biochemistry and Physiology, 2015, 123: 87 − 92. doi: 10.1016/j.pestbp.2015.03.008 [35] 潘雪莲, 杨磊, 金海峰, 等. 豆大蓟马在海南发生及防治的研究进展[J]. 热带生物学报, 2021, 12(4): 508 − 513. doi: 10.3969/j.issn.1674-7054.2021.4.hnrdnydxxb202104015 [36] COLINET H, SINCLAIR B J, VERNON P, et al. Insects in fluctuating thermal environments [J]. Annual Review of Entomology, 2015, 60: 123 − 140. doi: 10.1146/annurev-ento-010814-021017 [37] 马罡, 马春森. 气候变化下极端高温对昆虫种群影响的研究进展[J]. 中国科学:生命科学, 2016, 46(5): 556 − 564. [38] 邱海燕, 刘奎, 李鹏, 等. 豆大蓟马的生物学特性研究[J]. 热带作物学报, 2014, 35(12): 2437 − 2441. doi: 10.3969/j.issn.1000-2561.2014.12.021 [39] HONĚK A. Intraspecific variation in body size and fecundity in insects: a general relationship [J]. Oikos, 1993, 66(3): 483 − 492. [40] CHOWN S L, GASTON K J. Body size variation in insects: a macroecological perspective [J]. Biological Reviews, 2010, 85(1): 139 − 169. doi: 10.1111/j.1469-185X.2009.00097.x [41] SIEMANN E, TILMAN D, HAARSTAD J. Insect species diversity, abundance and body size relationships [J]. Nature, 1996, 380(6576): 704 − 706. doi: 10.1038/380704a0 [42] FOX C W, MCLENNAN L A, MOUSSEAU T A. Male body size affects female lifetime reproductive success in a seed beetle [J]. Animal Behaviour, 1995, 50(1): 281 − 284. doi: 10.1006/anbe.1995.0242 [43] AMERI M, RASEKH A, MICHAUD J P. Body size affects host defensive behavior and progeny fitness in a parasitoid wasp, Lysiphlebus fabarum [J]. Entomologia Experimentalis et Applicata, 2014, 150(3): 259 − 268. doi: 10.1111/eea.12158 [44] GERGS A, JAGER T. Body size-mediated starvation resistance in an insect predator [J]. Journal of Animal Ecology, 2014, 83(4): 758 − 768. doi: 10.1111/1365-2656.12195 [45] 唐良德, 赵海燕, 秦双, 等. 蝇蛹俑小蜂生物学特性及土壤因子对其寄生的影响[J]. 中国生物防治学报, 2015, 31(4): 467 − 472. doi: 10.16409/j.cnki.2095-039x.2015.04.004 [46] WU S Y, GAO Y L, XU X N, et al. Evaluation of Stratiolaelaos scimitus and Neoseiulus barkeri for biological control of thrips on greenhouse cucumbers [J]. Biocontrol Science and Technology, 2014, 24(10): 1110 − 1121. doi: 10.1080/09583157.2014.924478 [47] WU S Y, ZHANG Z K, GAO Y L, et al. Interactions between foliage- and soil-dwelling predatory mites and consequences for biological control of Frankliniella occidentalis [J]. Biocontrol, 2016, 61: 717 − 727. doi: 10.1007/s10526-016-9762-z [48] 邹一桥, 郑炳宗. 常用农药对丽蚜小蜂和中华草岭的毒力测定[J]. 生物防治通报, 1988(3): 114 − 117. [49] 王晓, 陈鹏, 张硕, 等. 12种杀虫剂对日本通草蛉不同虫态的毒力及安全性评价[J]. 植物保护, 2019, 45(2): 211 − 217. doi: 10.16688/j.zwbh.2018179 [50] 汤方, 李生臣, 孔祥波, 等. 吡虫啉等杀虫剂对温室白粉虱及其两种天敌的选择毒力[J]. 农药学学报, 2007, 9(1): 88 − 91. doi: 10.3321/j.issn:1008-7303.2007.01.018 [51] 王伟, 荣晓东, 胡学难, 等. 几种杀虫剂对榕管蓟马的室内毒力测定[J]. 广东农业科学, 2009, 36(5): 112 − 113. doi: 10.16768/j.issn.1004-874x.2009.05.015 -

 点击查看大图
点击查看大图

图(1) / 表 (3)
计量
- 文章访问数: 271
- HTML全文浏览量: 180
- PDF下载量: 16
- 被引次数: 0
