

-
海洋底栖生物(Marine benthos)是指生活在海洋基底表面或沉积物中的各种生物[1],是海洋生物中种类最多及生态关系最复杂的类群,浅至潮间带,深至海洋深渊,均有底栖生物的分布[2]。它们通过彼此间的营养关系,充分利用水层沉降的有机碎屑,促进营养物质的分解,在海洋生态系统的能量流动和物质循环中起到重要作用[3],很多也是可供人类直接利用的海洋资源。大型底栖生物(Macrobenthos)是指不能通过孔径为0.5 mm网筛的底栖生物,除海岸带有大型海藻以外,其余大型底栖生物均为动物[4]。大型底栖生物由于其固定于底质内或短距离活动的生活习性限制,受到环境变化的影响较为持久,对海洋环境污染反应敏感,具有污染效应指示性[5],因此,对海洋底栖生物的种群结构研究可以反映其生存环境水体的质量情况,有利于了解海洋生态系统的物质和能量动态,对保护海洋生态、维持渔业资源的可持续利用具有重要意义。国内更多的底栖生物群落研究位于近海的滩涂[6]、海湾[7-8]、岛屿潮间带[9-10],如胶州湾、长江口、福建和浙江沿岸等海域研究较多,而对于远海岛礁如西沙群岛及南沙群岛的底栖生物群落研究较少。西沙群岛的动物区系属于印度-西太平洋区的印尼-马来亚区,这是世界上海洋生物种类最丰富的区域[11],但因其地理位置远、夏季台风多等多种因素,在西沙群岛乃至整个南海海域展开科研工作难度较大,已有的西沙群岛底栖生物研究多为十几年前,乃至几十年前的资料,时间距离远、间隔大。如1957—1981年中科院海洋研究所、动物研究所、南海海洋研究所、上海水产学院(今上海海洋大学)、北京自然博物馆及海南岛和南海水产部门等联合或单独的多次调查,中科院海洋研究所于1957—1980年间先后5次派调查组前往西沙群岛对大型底栖海藻进行系统采集和研究[12],1980年发表了西沙群岛及附近海域的多毛类动物地理学研究[13],2002年对西沙和南沙群岛的底栖生物种群对比[14],2007年对南沙渚碧礁底栖生物调查[15];2009年进行了建国以来最大规模的西沙群岛珊瑚礁科考[16],但近年对于西沙海域的底栖生物群落研究数据则少报道。珊瑚礁生态系统是海洋生态系统的重要组成部分,具极高的初级生产力和生物多样性,也因其较高的生态服务价值而备受关注,但全球气候变化和人类活动已严重威胁到珊瑚礁生态系统的健康状况[17]。为更加深入了解西沙海域的珊瑚礁生态系统,于2016—2019年期间对西沙群岛附近珊瑚礁区的大型底栖生物进行了连续的采样调查,系统分析了该海域大型底栖生物的种类组成、丰度和生物量的分布、生物多样性等生物学特性,为西沙海域生物资源的保护及合理利用提供基础数据及理论依据。
-
西沙群岛(15°46′~17°08′N,111°11′~112°54′E)位于南海西北部,海南岛东南方,海岸线长达518 km,由宣德群岛和永乐群岛组合而成,以永兴岛为政治经济文化中心,扼守着南中国海的门户,具有极其重要的国际地位[18]。
本研究在宣德群岛的永兴岛、西沙洲、赵述岛和北岛,西部永乐群岛的晋卿岛、甘泉岛附近海域设置了调查站位。站位选择在岛屿周围珊瑚礁分布集中,并能反映出珊瑚礁状况的区域。由于天气等各种因素影响,各年的调查时间、调查岛屿不尽相同,但同一岛屿不同年份的调查位置尽量保持一致。2016年11月于永兴岛、西沙洲、赵述岛和北岛开展调查;2017年6月于西沙洲、赵述岛、北岛、晋卿岛和甘泉岛开展调查,9月调查永兴岛;2018年8月进行了全部6个岛屿的调查工作;2019年5月进行了北岛、晋卿岛、甘泉岛3个岛屿的调查。
-
由于珊瑚礁区底质为坚硬的珊瑚礁石,常规的抓斗式样方采集法无法正常采样,故采用潜水员水肺潜水采集。在各个岛屿礁盘站位选取珊瑚礁集中、并能反映出附近区域珊瑚礁状态的地点划定4个样方,样方面积为(0.25 m×0.25 m),样方水深受采样时潮汐情况影响在5~10 m之间略有变化,平均7 m。捕捉样方里可见的大型底栖生物(不包括珊瑚和大型海藻及海草),再用网袋将样方内珊瑚石采集、打捞出水,研究人员在船上将采回的珊瑚石敲碎并采集其中的大型底栖生物,进行分类鉴定、计数及生物量测量计算等研究工作,工作标准参照GB/T 12763.6—2007进行。
-
按照调查年份、调查站位所处岛屿海域,把来自同一年份、相同岛屿海域的样品归为1个群落,将样品划分为19个群落,计算Shannon-Wiener多样性指数(H')、Margalef丰富度指数(d)、Pielou均匀度指数(J')、优势度指数(Y)、优势度D2(该群落的第一和第二优势种的个体数占群落总个体数之比)、Jaccard群落系数(J)等数据,对大型底栖生物群落结构的生态特征进行分析。各指数的计算公式如下:
$$H^{\prime}=-\sum \mathrm{P}_i \cdot \log _2 \mathrm{P}_i $$ (1) $$ d=(\mathrm{S}-1) / \ln N$$ (2) $$J^{\prime}=H^{\prime} / \log _2 \mathrm{~S} $$ (3) $$ Y=n_i / N \cdot {{\rm{f}}}_i $$ (4) $$ \mathrm{D}_2=\left(N_1+N_2\right) / N $$ (5) 式中,Pi为第i种的生物个数与该群落总生物个数之比值;N为该群落生物个体数;S为该群落生物种数,0< J'≤1;ni为第i种的个体数;fi为该种在群落内各站位出现的频率;N为群落内每个种出现的总个体数;N1,N2分别代表该群落的第一和第二优势种的个体数。
对不同年份的19个岛屿大型底栖生物群落样本之间,利用各群落样本各自包含的物种数和共有的物种数分别两两计算Jaccard群落系数:J=c/(a+b−c)。其中,a,b分别为两群落的种类数,c为两群落的共有种数,以J为相似系数计列群落系数三角矩阵,根据三角矩阵采用近邻联结法将各样本岛屿大型底栖生物群落以群落系数从高到低进行关联,作聚类谱系图,研究群落分布特征。目前Warwick[19-20]提出的丰度生物量比较曲线法(Abundance and Biomass Curves,ABC)被认为是评价及监测海洋污染的行之有效的方法[21-22],它能够反映污染物对生物的实际效应及综合影响,以及海洋环境中不同污染物对生物的协同与拮抗作用,尤其是长期的轻度污染效应,因此很多研究都将ABC曲线法结果作为评价底栖生物群落受扰动情况的依据[23-24]。
数据计算,图表、ABC曲线绘制采用Microsoft Excel 2019制作。调查站位图使用Arc GIS软件绘制,聚类分析图使用PAST软件[25]绘制。
-
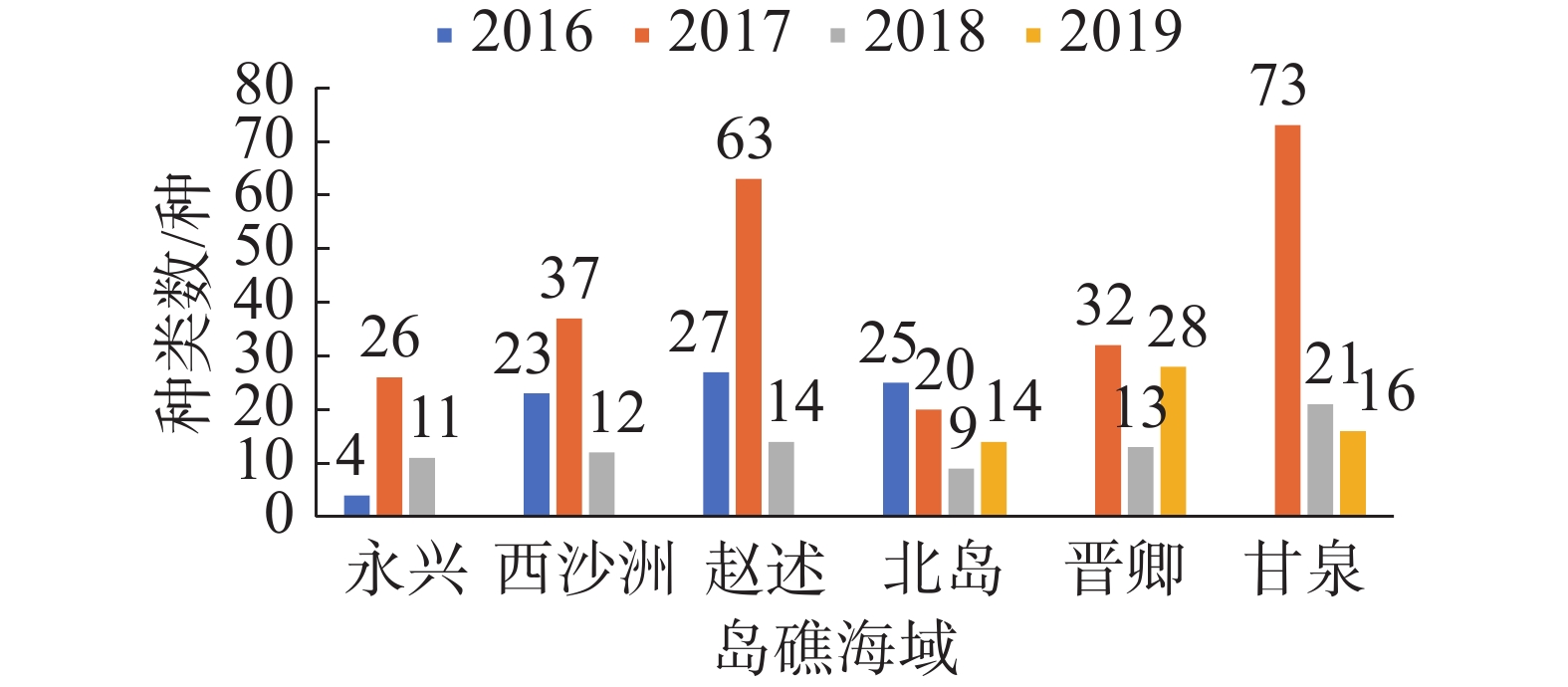
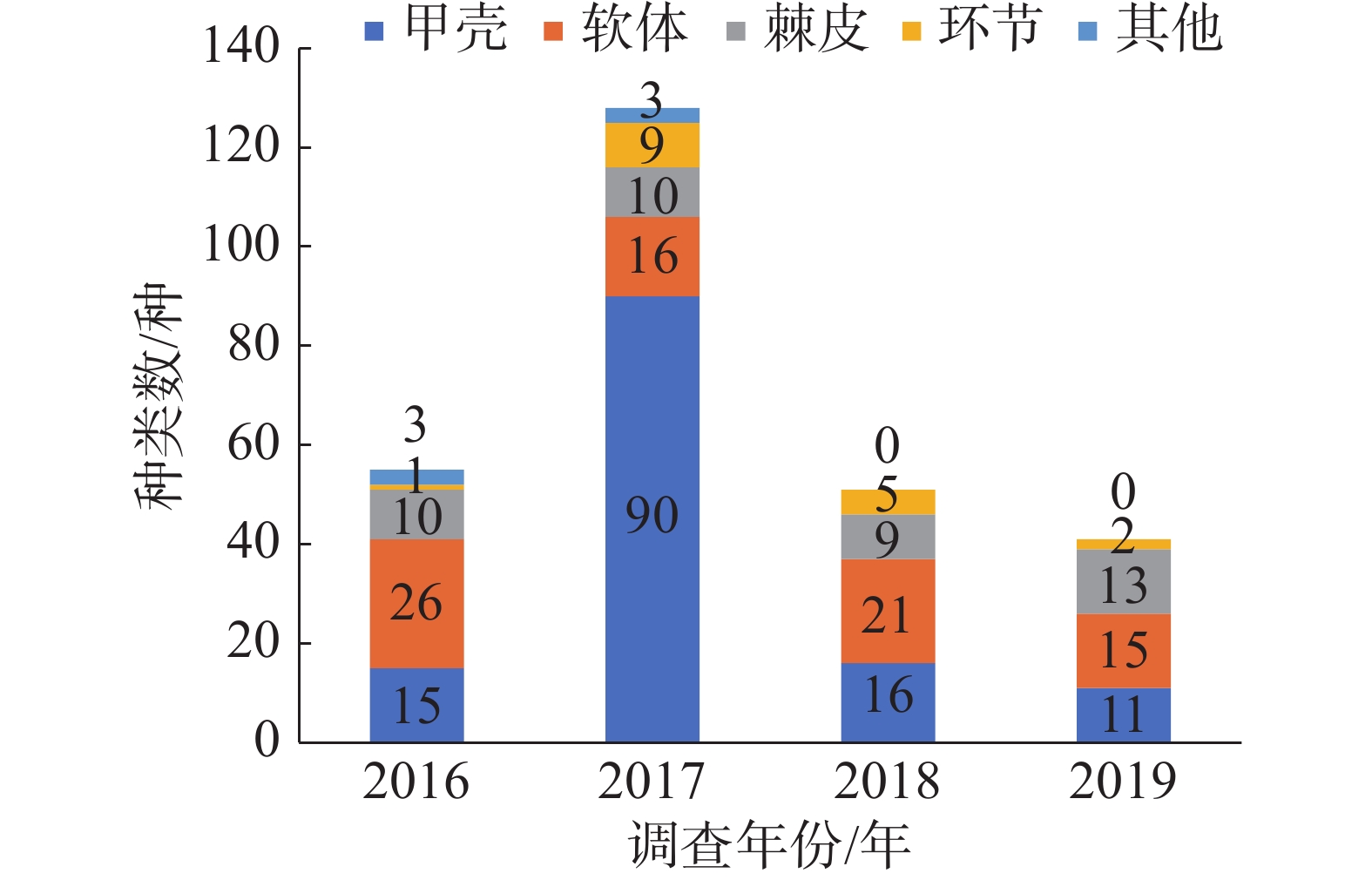
2016—2019年西沙附近海域珊瑚礁分布区共采获5个生物大类中的252种底栖生物。其中甲壳动物出现的种类最多,有120种;软体动物有72种;棘皮动物37种,环节动物17种,其他门类生物6种。各年度大型底栖生物各门类生物种数分布情况见图1,同一年度内各岛屿群落的大型底栖生物种类组成与该年度的总体生物组成基本近似。各门类生物种类占比一般是:软体动物>甲壳动物>棘皮动物>环节动物>其他门类,仅2017年除外。软体动物一般是西沙珊瑚礁区底栖生物群落中物种最丰富的门类,但2017年的各岛礁大型底栖生物群落的最优势生物类群为甲壳动物,其种类数及占比远远大于另外几大门类生物。
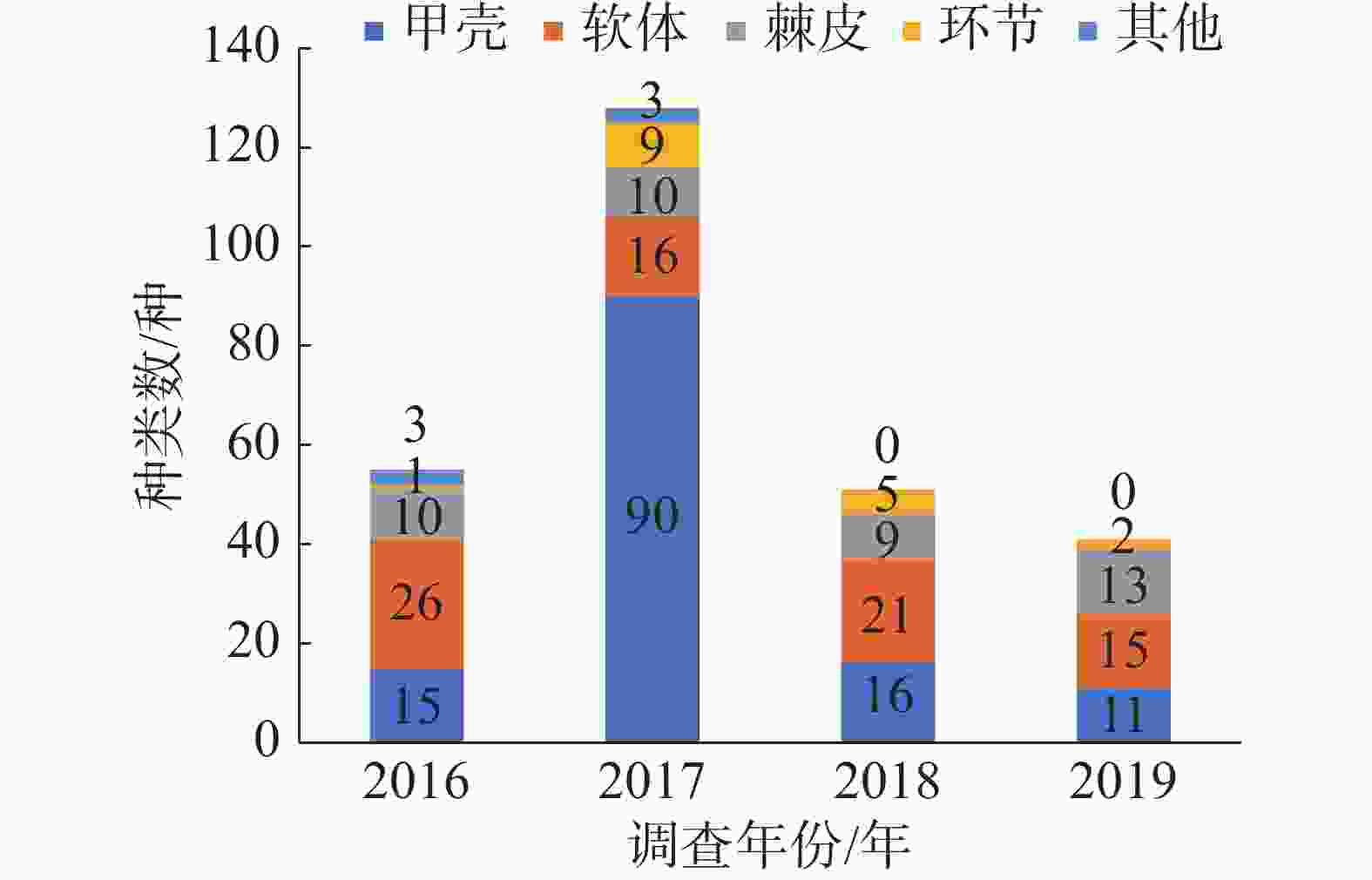
图 1 各年度大型底栖生物种类总体组成
各年度各岛礁海域的大型底栖生物群落种类数变化如图2,同一年度内,甘泉岛和赵述岛一般具有最大的生物种类数,西沙洲、晋卿岛次之,而北岛和永兴岛的生物种数一般较少。在不同年度之间,因为各年度之间采样的岛礁站位并不完全一致,因此不能简单对比各年度调查到的总生物种类数。分开观察各岛屿调查到的生物种数可发现,各海域的生物种类数一般在2016—2017年上升,2017年大型底栖生物的种类数达到1个高峰, 2018年和2019年出现明显回落,与调查到的总生物种数趋势一致。
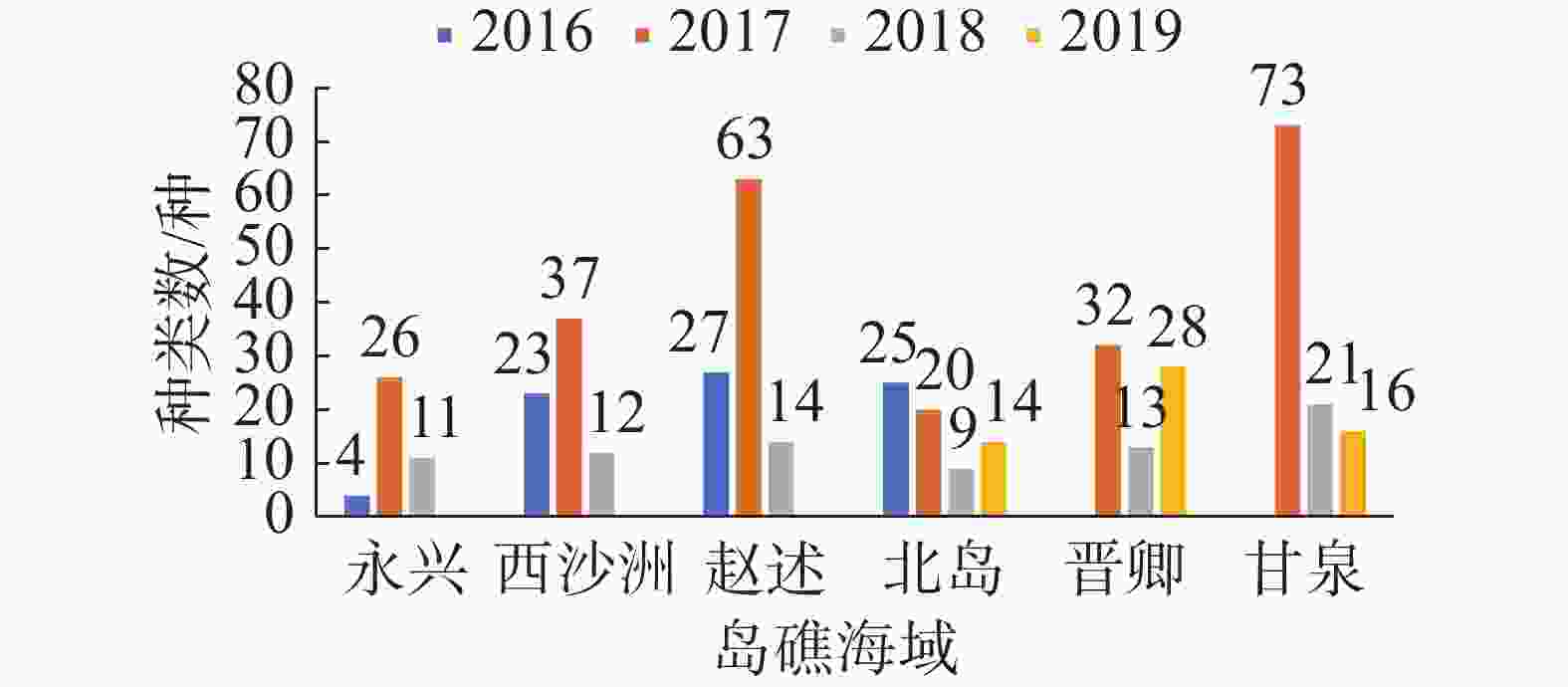
图 2 各岛礁海域大型底栖生物种类年度变化
-
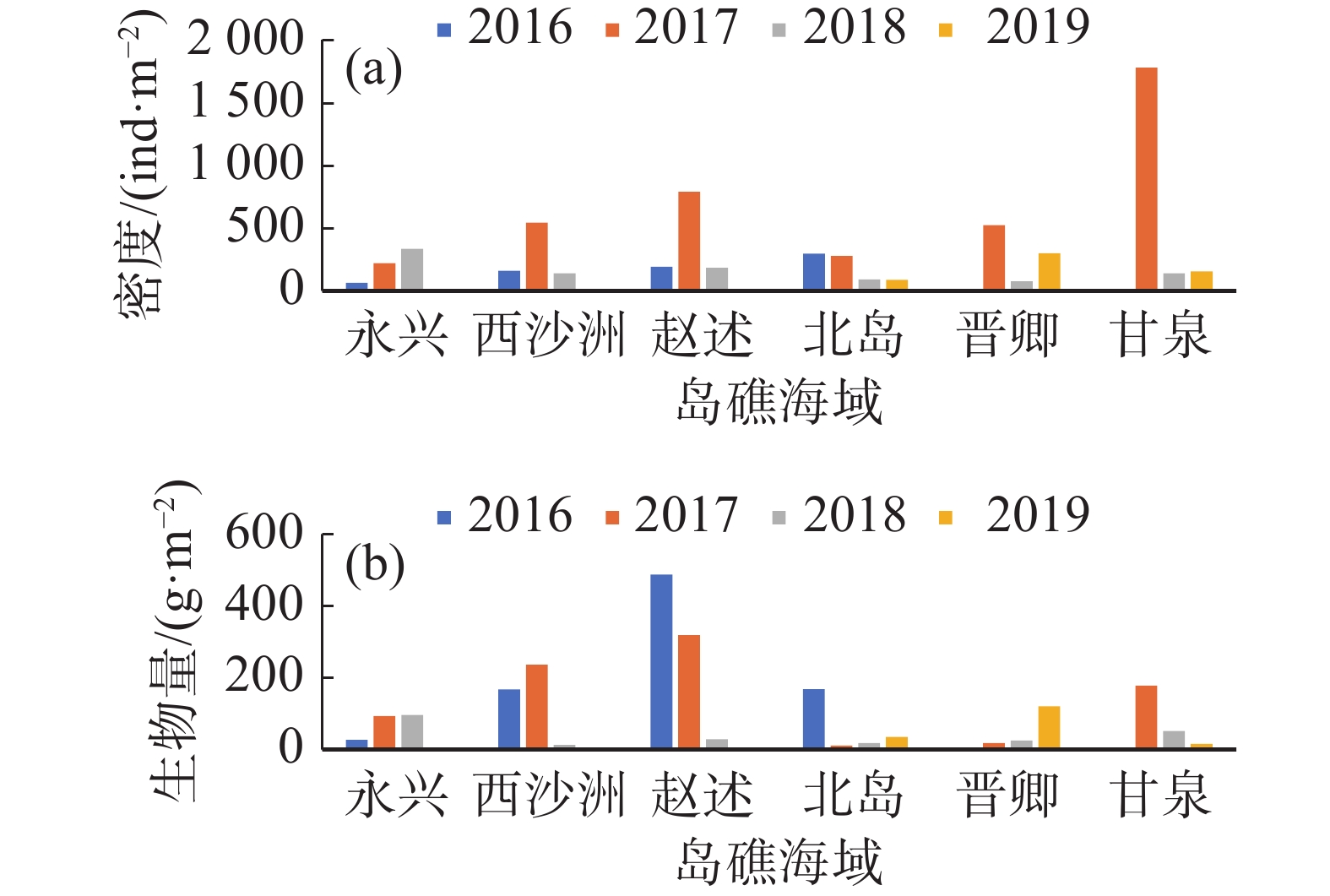
经计算,西沙群岛海域珊瑚礁区的平均大型底栖生物密度为(334.45±396.40) ind·m−2,生物量为(110.79±126.56) g·m−2,各年度各岛屿珊瑚礁区的大型底栖生物密度及生物量变化差距极大,表现了珊瑚礁大型底栖生物系统分布极不均匀的特征(图3)。
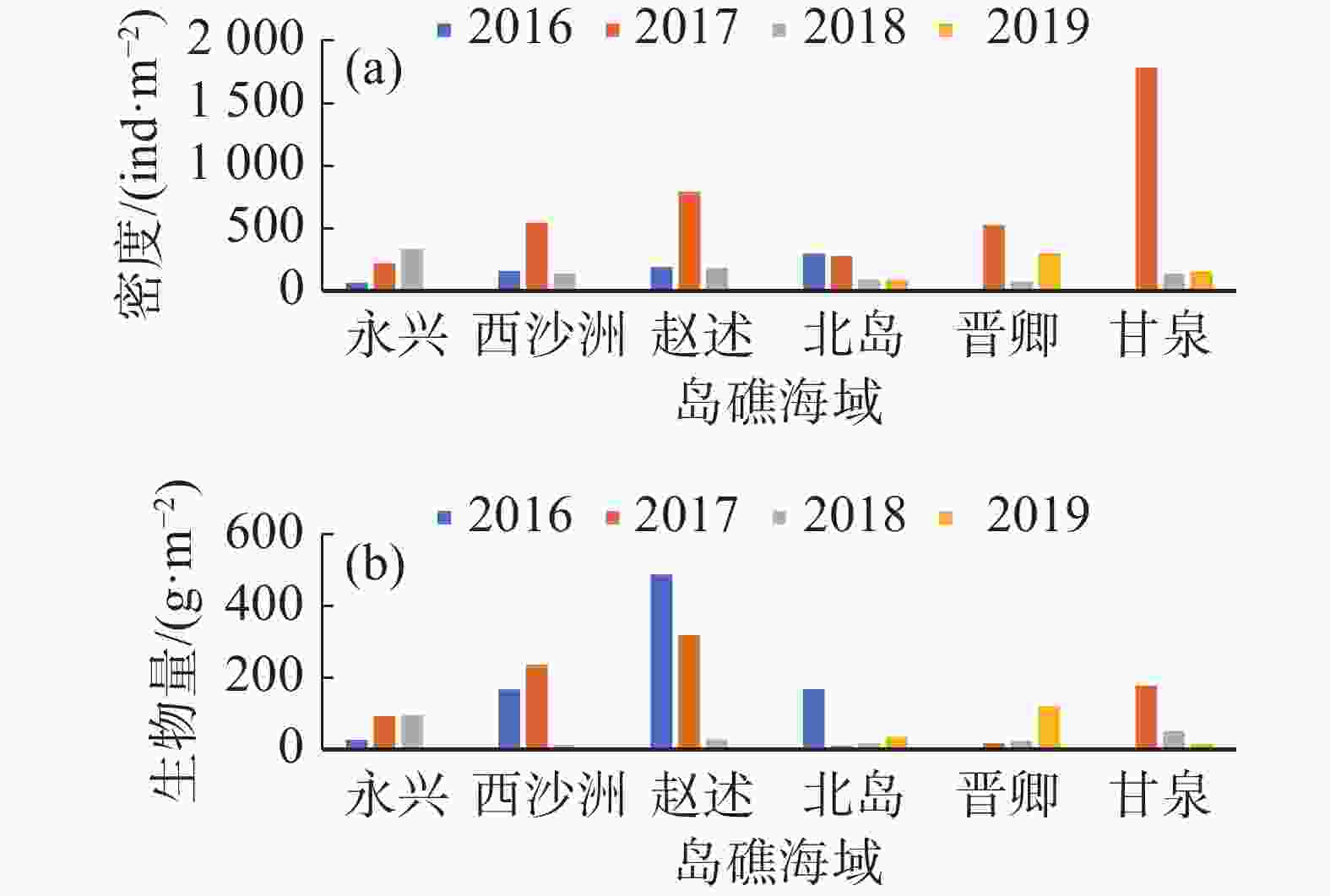
图 3 各年度西沙珊瑚礁区大型底栖生物密度(a)及生物量(b)
-
以单年度的岛屿海域作为生物组合单元,计算H',d,J',D2等各项数据,结果见表1。整个调查海域的平均指数为:H'=3.85±0.84,d=5.51±2.64,J'=0.89±0.05,D2=0.32±0.12。大部分的西沙岛礁珊瑚礁区大型底栖生物群落均具有以上特征,这些群落的大部分生物种类都是在样方里零星出现,种类数很多,但各种生物的数量都很少,多样性及均匀度高,而优势种的优势度低,除2016年的永兴岛群落因为种数实在太少仅有4种,导致虽然其均匀度为1(理论最大值),其多样性较低,而优势度较高。
表 1 西沙珊瑚礁区大型底栖生物多样性、均匀度和优势度
年份 指数 永兴 西沙洲 赵述 北岛 晋卿 甘泉 2016 H' 2.00 4.25 4.44 3.94 d 2.16 5.96 6.71 5.58 J' 1.00 0.94 0.93 0.85 D2 0.50 0.20 0.21 0.32 年份 指数 永兴 西沙洲 赵述 北岛 晋卿 甘泉 2017 H' 4.22 4.21 5.33 3.92 4.37 5.52 d 5.72 6.82 10.98 4.86 6.82 12.48 J' 0.90 0.81 0.89 0.91 0.87 0.89 D2 0.25 0.34 0.17 0.28 0.28 0.13 年份 指数 永兴 西沙洲 赵述 北岛 晋卿 甘泉 2018 H' 3.08 2.87 3.16 3.05 3.58 4.10 d 3.28 3.09 3.40 2.55 4.08 5.63 J' 0.89 0.80 0.83 0.96 0.97 0.93 D2 0.48 0.54 0.43 0.35 0.26 0.23 年份 指数 永兴 西沙洲 赵述 北岛 晋卿 甘泉 2019 H' 3.48 4.06 3.49 d 4.21 6.25 4.09 J' 0.91 0.84 0.87 D2 0.41 0.36 0.41 -
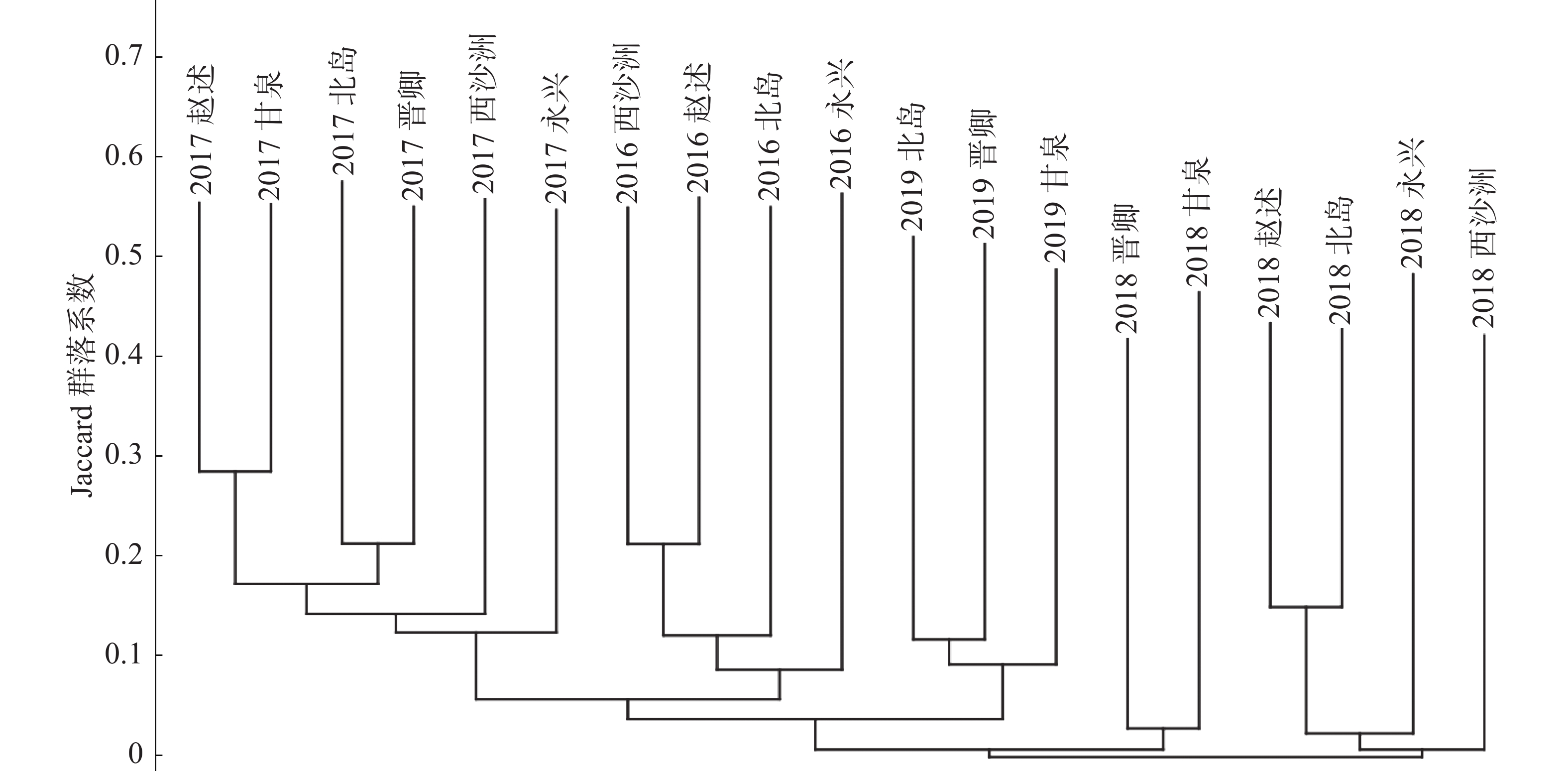
对不同年份的19个岛屿大型底栖生物群落样本之间,利用各群落样本各自包含的物种数和共有的物种数分别两两计算Jaccard群落系数,列出群落系数三角矩阵,根据三角矩阵采用近邻联结法将各样本岛屿大型底栖生物群落以群落系数从高到低进行关联,作出聚类谱系图(图4);由图5可以看出,各群落以调查年份为共同特征汇聚,2016、2017、2019年的调查结果都各自汇聚成枝,不同调查年份之间的群落系数则非常低。
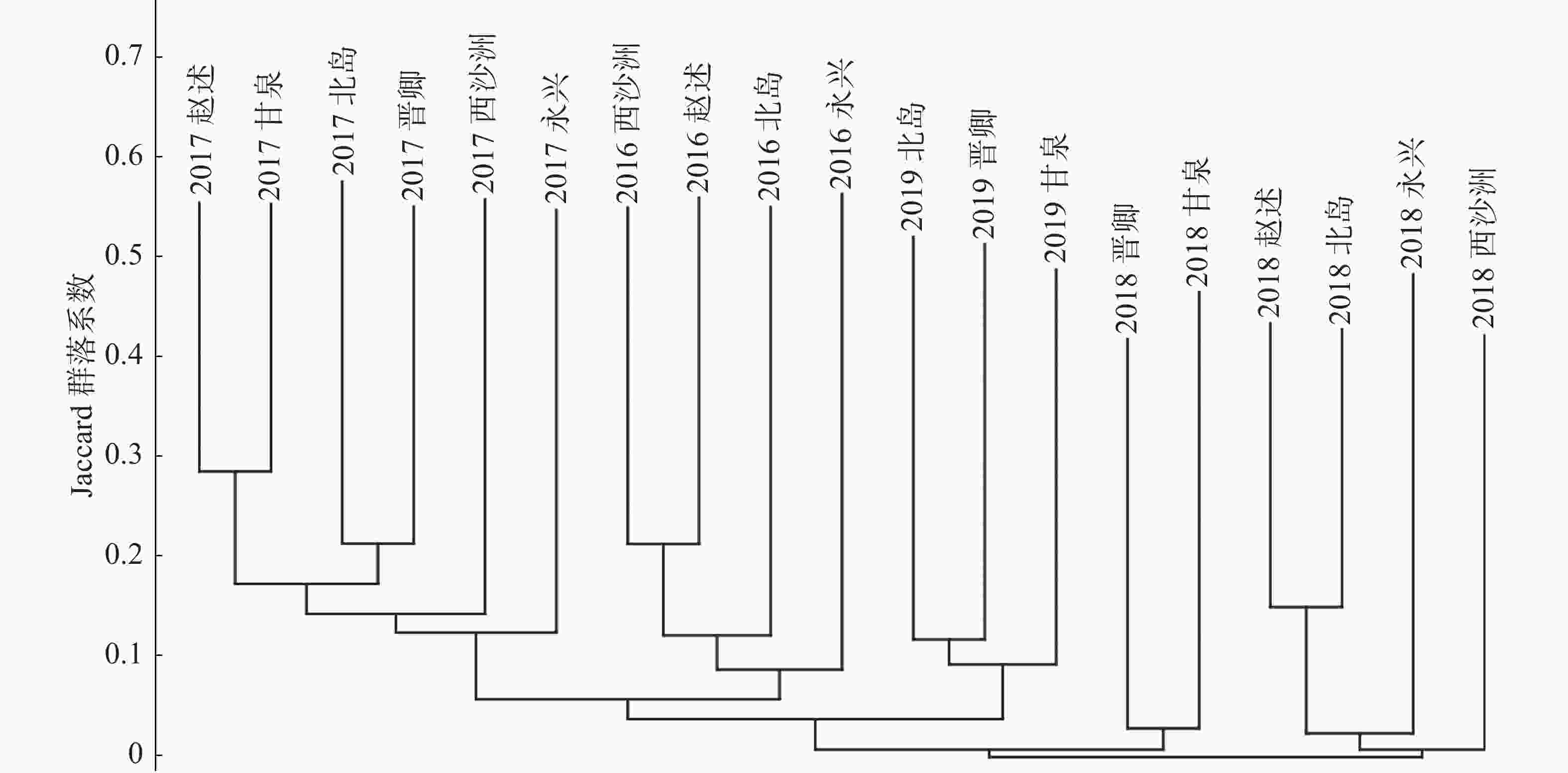
图 4 西沙珊瑚礁区大型底栖生物群落种类组成的聚类谱系图
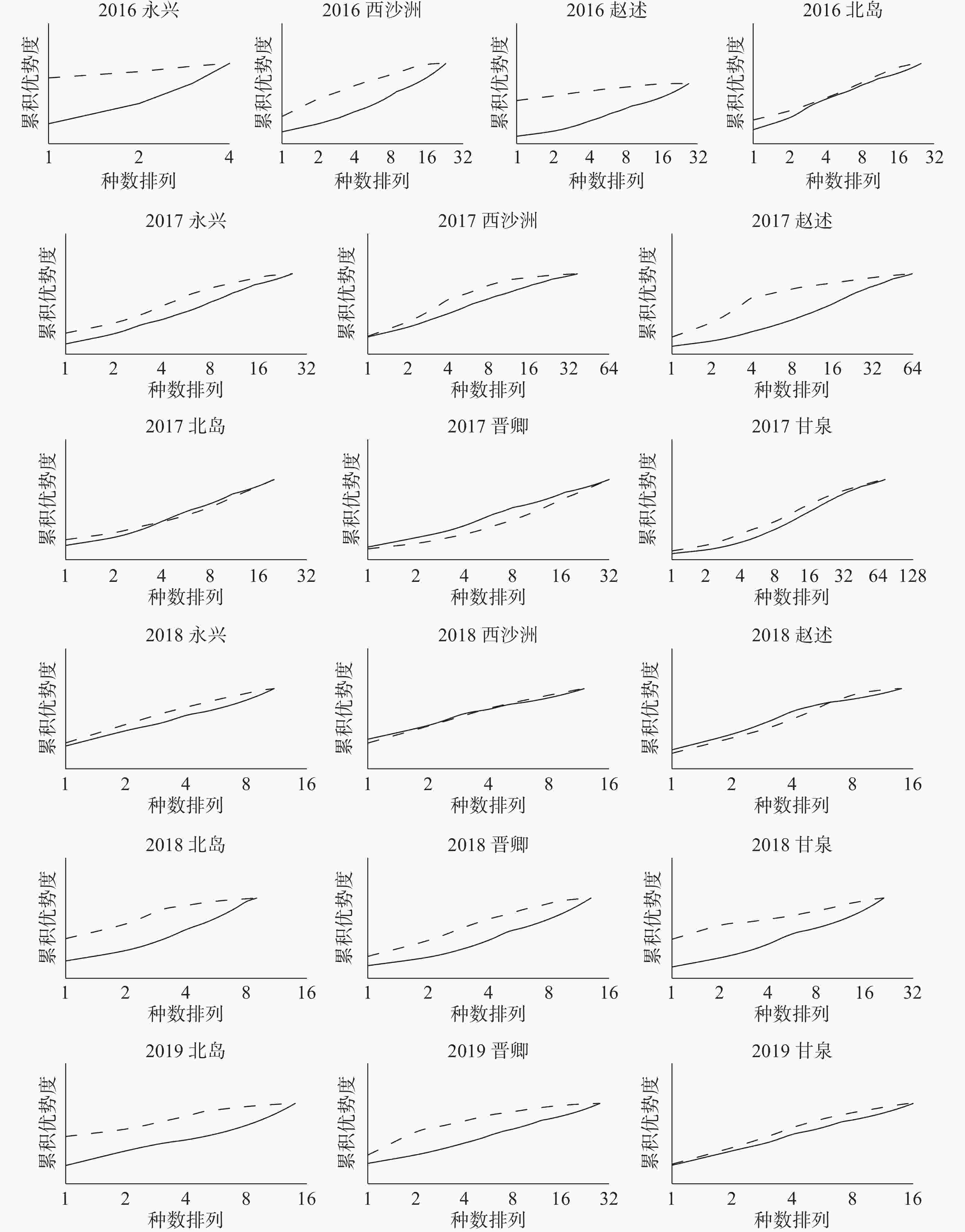
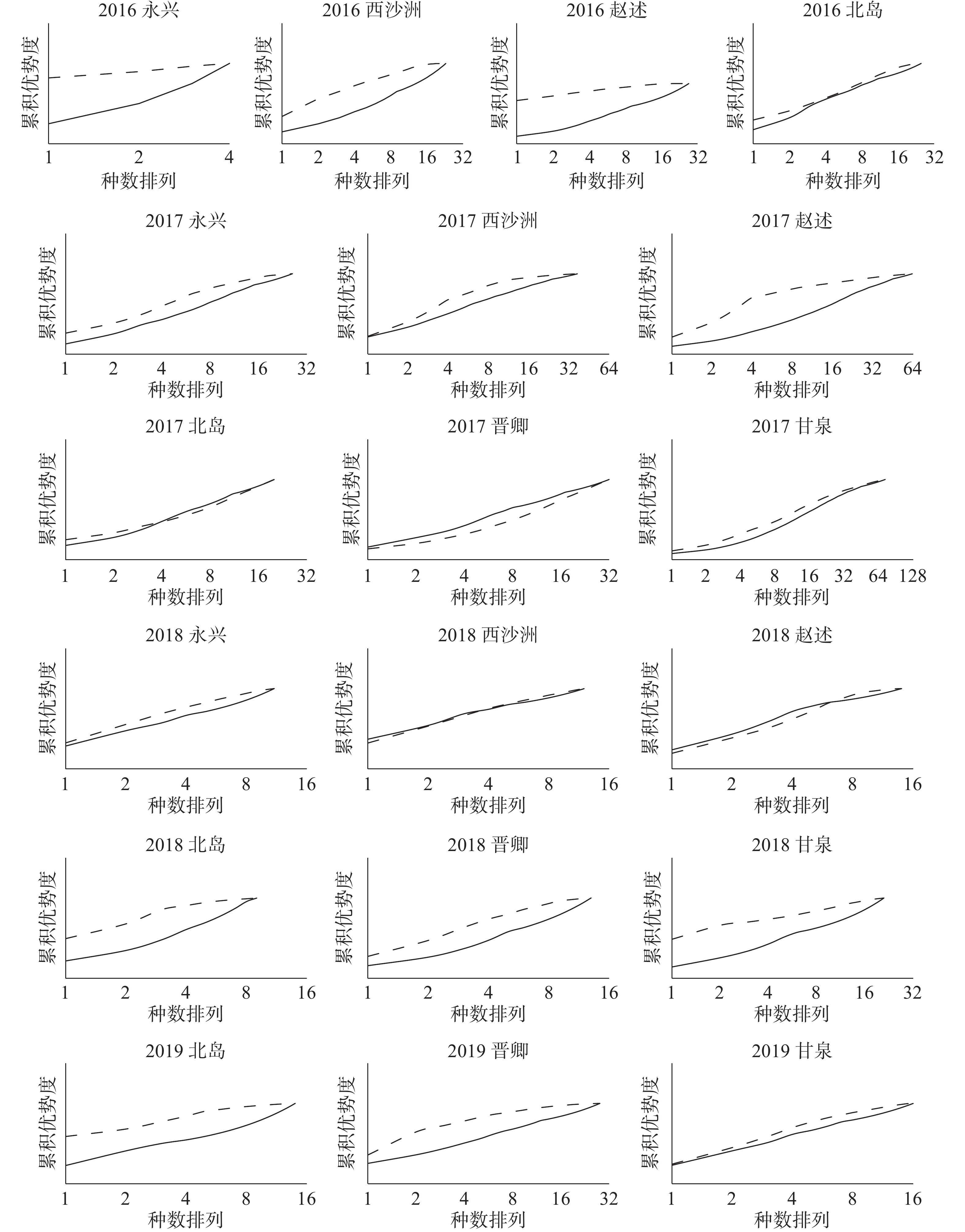
图 5 西沙珊瑚礁区大型底栖生物群落ABC曲线
-
将不同年份的19个岛屿大型底栖生物群落样本分别作出ABC曲线(图5)来判断群落是否受到扰动。由图可知,调查时间段内的大部分西沙海域岛屿的大型底栖生物群落,其生物量曲线(虚线)都是位于丰度曲线(实线)之上的,这说明群落中有一个或几个大型的种占优势,且每个种有几个个体,种内生物量的分布比丰度分布更具优势,表明这些群落的海洋环境相对稳定,底栖生物群落结构也相对平衡稳定,没有受到扰动或扰动程度较小;而2017年的北岛群落,2018年的西沙洲、赵述岛群落,K-优势度曲线图上的生物量曲线与丰度曲线相互交叉或重叠,表明这些群落生物量占优势的大个体已经消失。随机的个体较小的种类开始占据优势数量,在此情况下种内丰度与生物量的分布优势较难分辨,说明这些群落受到了中度污染或扰动;在整个研究范围内,仅有一个群落即2017年的晋卿岛群落,其丰度曲线整条均位于生物量曲线上方,表明该群落的底栖生物个体数由若干个体极小的种占据优势,种内丰度的分布比生物量分布更显优势,群落受到了严重扰动。
-
观察西沙群岛珊瑚礁区各年度大型底栖生物各门类生物种类数分布情况,同一年度内各岛屿群落的大型底栖生物种类组成与该年度的总体生物组成基本近似。各门类生物种类占比大小一般是:软体动物>甲壳动物>棘皮动物>环节动物>其他门类,仅2017年除外。软体动物一般是西沙珊瑚礁区底栖生物群落中物种最丰富的门类,这与大亚湾[26]、徐闻珊瑚礁自然保护区[27]及三亚蜈支洲岛[28]等地进行的珊瑚礁区大型底栖生物研究一致;但2017年的各岛礁大型底栖生物群落的最优势生物类群为甲壳动物,其种类数及占比远远大于另外几大门类生物,这种底栖生物群落的种类组成方式与2011年三亚珊瑚礁区的研究结果相近[29]。在其他非珊瑚礁底质的环境,优势类群为多毛类环节动物和棘皮动物,如2005—2009年于胶州湾进行的调查[3]、2015年于烟台进行的调查[30],结果都显示多毛类占据底栖生物类群的最主要部分;2018年于亚龙湾进行的研究则显示棘皮动物为大型底栖生物的优势类群[31],以上的生物种类分布情况则未在西沙珊瑚礁区的大型底栖生物群落中出现。
各海域的生物种类数一般在2016—2017年上升,2017年大型底栖生物的种类数达到1个高峰, 2018年和2019年出现明显回落,与调查到的总生物种数趋势一致。该现象的成因尚待更多研究揭示,长棘海星的爆发是一个非常值得重视的因素。虽然由于采样方法限制,长棘海星本身没有出现在本研究的样方中,但根据资料显示,西沙群岛的长棘海星爆发约以15年为周期,2006—2010年为其破坏期,使得西沙海域的珊瑚覆盖率从60%降至不到5%,2011—2017年西沙群岛的长棘海星密度较低(每公顷个体数少于1),珊瑚有所恢复;2018年长棘海星密度开始增加,分布密度为每公顷400只左右,2019年已达到每公顷上千只[32-33],这与本研究结果发现的大型底栖生物种群变动规律可以相互印证。
在底栖生物的生物量和栖息密度方面,本研究参考了国内一些大型底栖生物历史研究的数据并与本次研究进行比对(表2),在2011年我国的4个边缘海的大型底栖生物研究中列出了我国各海域的年平均生物量和栖息密度[3],与本研究的数据进行对比,可以看出,西沙群岛海域珊瑚礁区的大型底栖动物的栖息密度数据均大于南黄海、东海、南海海域的年总平均值,低于渤海和北黄海;而平均生物量则大于各海区的年平均生物量数据,表明西沙群岛珊瑚礁区海域在整个南海乃至我国边缘海海域中皆属丰富度相对较高的海域。将西沙群岛海域大型底栖生物群落的特征与国内其他珊瑚礁区底栖动物群落的特征数据进行对比,可发现西沙珊瑚礁区的大型底栖生物密度与同为远海岛礁的南沙渚碧礁[15]的数据较为接近,生物量略高于后者。而与近岸的几个珊瑚礁区如大亚湾[26] 、徐闻[27]、三亚[29]等相比,可发现近岸珊瑚礁区的生物密度明显高于远海珊瑚礁区,生物量一般也明显超出后者。分析其原因可能是:近海海区受到陆地径流和上升流影响,营养盐丰富,初级生产力较高;而西沙群岛作为远海岛礁,在初级生产力方面虽高于普通海盆区域,但与接近大陆和海南岛的近岸区域相比仍较低[34],导致虽然同是南海热带珊瑚礁区的大型底栖生物群落,远海岛礁群落的物种数量可能较多,但多数物种都是以单个小个体出现在样方中,在生物种类丰富度高于近海的同时,生物密度和生物量都不及近海珊瑚礁区。
表 2 我国各边缘海及珊瑚礁区底栖生物平均密度及生物量
在生物多样性与丰富度指数方面,H',d,J',D2从不同侧面测定生物群落中总生物数量/种类数以及各个种的生物数量之间的配置关系,通常用于群落样本生物种类多样性的综合分析。若前三者较大而最后者较小,说明多样性较高,生物群落环境的底质越好[35],反之则说明多样性较低。一些研究中将多样性指数(H')作为判断底栖生物群落是否受扰动的依据,如蔡立哲等[35]、李永祺等[36]采用的多样性指数与水质污染程度的分级标准,即H'小于1,严重污染;1<H'<2,中等污染;2<H'<3,轻度污染;H'大于3,清洁;根据这个标准,本研究中西沙群岛各岛屿的底栖生物群落均未受到中度以上的污染扰动,但是在2016年永兴岛、2018年西沙洲站位可能受到轻度的污染。然而,考虑到H'是生物多样性的反映,生物多样性受干扰的原因应是非常多元的,不可能只与底质或水体的污染有关,而西沙海域作为热带海域,珊瑚礁生境相对于其他生境又明显会有较高的丰富度,因此以上标准是否适用值得商榷,实际上该标准在国内也很少用于评价沉积环境质量。本研究使用ABC曲线分析方法也发现有一些H'较高的底栖生物群落实际上还是受到了一定程度的扰动,尤其在2017年和2018年较为严重。如2017年的北岛群落,2018年的西沙洲、赵述岛群落,K-优势度曲线图上的生物量曲线与丰度曲线相互交叉或重叠,表明这些群落受到了中度污染或扰动; 2017年的晋卿岛群落,其丰度曲线整条均位于生物量曲线上方,表明群落受到了严重扰动。
聚类分析结果显示,调查活动的时间对于群落组成相似度的影响要远大于地理位置的影响,导致这种结果最主要的原因可能是调查季节因素的影响。南海海域的调查工作往往由于各种因素无法保持在每年的相同时段开展,使得调查季节发生很大差异。如前所述,本研究的4个年度的调查季节各不相同,最早有5月初,最晚可达11月初。西沙群岛属于南海的中心地带,不受陆地条件影响,海流受季风左右,表层海水的流向、路径、强度均与季风有关,冬季(10月—翌年4月)海流向西岸集流,夏季(5—8月)则向东北,在季风转换的4月和10月海流不稳定[37]。海流的变化会影响整个海域的生物群落结构,导致其发生较大变化。
许多研究认为近年的珊瑚礁生态系统存在退化趋势[38-39],西沙海域的珊瑚覆盖率从2005年的56.20%下降至2012年的不足10%[40];李元超等[41]对西沙海域永兴岛和七连屿的研究结果表明,该区域内10年来的造礁石珊瑚种类下降明显,而造礁石珊瑚作为礁栖大型底栖生物的主要生存环境,其种群的变动与大型底栖生物的群落状况可以相互印证表明,西沙海域已经受到了一定程度的扰动,其原因可能是人类活动如船体抛锚、潜水活动损坏[42]、珊瑚病害、长棘海星爆发、全球气候变化等因素,但具体原因尚不能定论,需要结合环境因子和其他资料进行更多的研究分析。
-
通过分析2016—2019年西沙群岛珊瑚礁区大型底栖生物的群落种类组成、生物密度和生物量数据,计算群落多样性、丰富度、均匀度等指数,并与国内其他研究对比发现:西沙群岛珊瑚礁区大型底栖生物群落总体上以软体动物和甲壳动物为主,种类数、多样性相较于近岸珊瑚礁区底栖生物群落较高,而生物密度、生物量则较低;通过对各岛屿群落样本的聚类分析发现:调查时间的变化比地理位置的变化对底栖生物群落的结构有更大影响;通过多样性指数和ABC曲线分析,发现西沙海域的大型底栖生物群落受到了一定程度的扰动,其原因很有可能是长棘海星的爆发,具体扰动因素尚待更多资料进一步分析。
随着社会的不断进步与发展,我国对海洋资源的开发力度越来越大,近海海洋生态环境受到巨大的人类活动压力变得越来越脆弱,渔业资源日渐萎缩。在此情势下,我国的海洋资源开发活动自然而然会将眼光转向深远海,南海海域必将是我国未来海洋资源开发的重中之重,对南海海洋资源的开发利用和保护需要对该海域的生态环境投入持续关注并深入研究。大型底栖生物对海洋环境污染反应敏感,具有污染效应指示性。因此对南海海域大型底栖生物群落的长期的宏观变化规律作出详细的阐述,对其现状做出科学的评价,并对其未来发展趋势作出准确的预测,对我国海域海洋生物资源的开发和生物多样性保护、海洋环境变化的监测等均具有重要的现实意义。本研究表明,西沙海域的岛礁珊瑚礁大型底栖生物群落已经受到了一定程度的扰动,这种扰动和变化的成因为何,影响程度大小尚不明朗,需要更多的综合性调查研究来揭示。需要进行长期的调查观测、数据分析,结合水质、气象等多角度研究,了解其群落的变化规律和与人类活动、全球气候变化、环境因子的响应机制,才能从根本上阐明它们在海洋生态系统中的地位和作用,从而有效保护海洋生物资源和海洋生物多样性,进而对其充分开发、合理利用。
A survey of macrobenthos communities in coral reef areas of Xisha Islands
-
摘要: 为了解西沙群岛的珊瑚礁区大型底栖生物群落组成和分布特征,于2016—2019年期间对西沙群岛附近珊瑚礁区的大型底栖生物进行了连续的采样调查,系统分析了该海域大型底栖生物的种类组成、丰度和生物量的分布、生物多样性等生物学特性。结果表明,5个生物大类中的252种底栖生物中,甲壳动物出现的种类最多,有120种;其次为软体动物,有72种;棘皮动物37种,环节动物17种,其他门类生物6种。大型底栖生物密度为(334.45±396.40) ind·m−2,生物量为(110.79±126.56)g·m−2,Shannon-Wiener多样性指数(H’)为3.85±0.84、丰富度指数(d)为5.51±2.64、均匀度指数(J’)为0.89±0.05;多样性指数显示,2016年永兴岛及2018年西沙州海域受到轻度扰动;群落聚类分析显示,该区域大型底栖生物群落随时间变化较显著;丰度-生物量比较曲线法(ABC曲线)评价显示,2017年和2018年的部分岛屿大型底栖生物群落受到一定程度干扰。结果说明西沙群岛珊瑚礁区大型底栖生物群落总体上以软体动物和甲壳动物为主,调查时间的变化比地理位置的变化对底栖生物群落的结构有更大影响。Abstract: A survey was made of macrobenthos from 2016 to 2019 to understand the composition and distribution characteristics of macrobenthos community in the coral reef areas of Xisha islands. Four quadrats were arranged in each coral reef area of Xisha islands and the quadrat size was 0.25 m × 0.25 m. A systematic analysis was made of the species composition, richness, biomass distribution and biodiversity of the macrobenthos collected. The analysis showed that 252 species of macrobenthos were found in 5 biological groups, of which crustaceans were the most common species and had 120 species, followed by mollusc (72 species), echinoderms (37 species), annelids (17 species) and other phyla (6 species). Density of macrobenthos was (334.45±396.40) ind·m−2, biomass (110.79±126.56) g·m−2, Shannon Wiener diversity index (H') 3.85±0.84, Richness index (d) 5.51 ±2.64 and Evenness index (J') 0.89±0.05. According to the diversity index, Yongxing island in 2016 and Xishazhou in 2018 were slightly disturbed. The cluster analysis of the community showed that the macrobenthos communities in these areas changed significantly over time. An abundance/biomass comparison curve (ABC method) was used to assess the status of the macrobenthos communities, and the results showed that macrobenthos communities in some islands of Xisha Islands in 2017 and 2018 were disturbed to some extent. It is concluded that the macrobenthos communities in the coral reef areas of Xisha Islands are generally mainly composed of crustaceans and mollusks, and that the change in time of survey has a greater effect on the structure of macrobenthos community than the change in geological location.
-
Key words:
- Xisha Islands /
- coral reef /
- macrobenthos /
- community structure /
- ABC curve
-
表 1 西沙珊瑚礁区大型底栖生物多样性、均匀度和优势度
年份 指数 永兴 西沙洲 赵述 北岛 晋卿 甘泉 2016 H' 2.00 4.25 4.44 3.94 d 2.16 5.96 6.71 5.58 J' 1.00 0.94 0.93 0.85 D2 0.50 0.20 0.21 0.32 年份 指数 永兴 西沙洲 赵述 北岛 晋卿 甘泉 2017 H' 4.22 4.21 5.33 3.92 4.37 5.52 d 5.72 6.82 10.98 4.86 6.82 12.48 J' 0.90 0.81 0.89 0.91 0.87 0.89 D2 0.25 0.34 0.17 0.28 0.28 0.13 年份 指数 永兴 西沙洲 赵述 北岛 晋卿 甘泉 2018 H' 3.08 2.87 3.16 3.05 3.58 4.10 d 3.28 3.09 3.40 2.55 4.08 5.63 J' 0.89 0.80 0.83 0.96 0.97 0.93 D2 0.48 0.54 0.43 0.35 0.26 0.23 年份 指数 永兴 西沙洲 赵述 北岛 晋卿 甘泉 2019 H' 3.48 4.06 3.49 d 4.21 6.25 4.09 J' 0.91 0.84 0.87 D2 0.41 0.36 0.41  下载: 导出CSV
下载: 导出CSV
-
[1] 李太武. 海洋生物学[M]. 北京: 海洋出版社, 2013. [2] 李荣冠. 中国海陆架及邻近海域大型底栖生物[M]. 北京: 海洋出版社, 2003. [3] 李新正. 我国海洋大型底栖生物多样性研究及展望: 以黄海为例[J]. 生物多样性, 2011, 19(6): 676 − 684. [4] 沈国英. 海洋生态学. [M]. 3版. 北京: 科学出版社, 2010. [5] 田胜艳, 于子山, 刘晓收. 丰度/生物量比较曲线法监测大型底栖动物群落受污染扰动的研究[J]. 海洋通报, 2006, 25(1): 92 − 96. doi: 10.3969/j.issn.1001-6392.2006.01.013 [6] 夏江宝, 李传荣, 许景伟, 等. 黄河三角洲滩涂区大型底栖动物群落数量特征[J]. 生态环境学报, 2009, 18(4): 1368 − 1373. doi: 10.3969/j.issn.1674-5906.2009.04.029 [7] 张 萍, 缴建华, 孙万胜, 等. 渤海湾天津近岸海域大型底栖动物群落结构及次级生产力的初步研究[J]. 大连海洋大学学报, 2016, 31(3): 324 − 330. doi: 10.16535/j.cnki.dlhyxb.2016.03.017 [8] 袁涛萍, 李恒翔, 李 路. 夏季大亚湾大型底栖动物群落结构[J]. 热带海洋学报, 2017, 36(1): 41 − 47. [9] 朱四喜, 郭宝英, 杨红丽, 等. 浙江六横岛潮间带大型底栖动物的群落特征研究[J]. 水生态学杂志, 2010, 3(6): 19 − 22. doi: 10.15928/j.1674-3075.2010.06.004 [10] 周时强, 郭 丰, 吴荔生, 等. 福建海岛潮间带底栖生物群落生态的研究[J]. 海洋学报, 2001, 23(5): 104 − 109. [11] 齐钟彦. 西沙群岛的海洋生物考察[J]. 海洋科学, 1979(S1): 62 − 65. [12] 夏邦美, 王广策, 王永强. 三沙市南海诸岛底栖海藻区系调查及其与其它相关区系的比较分析[J]. 海洋与湖沼, 2013, 44(6): 1681 − 1704. doi: 10.11693/hyhz20131000145 [13] 吴宝铃, 孙瑞平, 陈 木. 西沙群岛及其附近海域多毛类动物地理学的研究[J]. 海洋学报, 1980(1): 111 − 130. [14] 李新正, 王永强. 南沙群岛与西沙群岛及其邻近海域海洋底栖生物种类对比[J]. 海洋科学集刊, 2002(1): 20 − 21. [15] 李新正, 李宝泉, 王洪法, 等. 南沙群岛渚碧礁大型底栖动物群落特征[J]. 动物学报, 2007, 53(1): 83 − 94. doi: 10.3969/j.issn.1674-5507.2007.01.010 [16] 许 红, 史国宁, 廖宝林, 等. 中国海洋的珊瑚-珊瑚礁: 南海中央区珊瑚-珊瑚礁生物多样性特征[J]. 古地理学报, 2021, 23(4): 771 − 788. doi: 10.7605/gdlxb.2021.04.043 [17] 吴钟解, 唐 佳, 蔡文启, 等. 海南岛东海岸珊瑚礁生态系统中抗生素抗性基因的赋存特征[J]. 热带生物学报, 2022, 13(5): 464 − 471. doi: 10.3969/j.issn.1674-7054.2022.5.hnrdnydxxb202205007 [18] 刘晓红. 浅谈西沙群岛海域生态环境现状[J]. 化工管理, 2020(20): 54 − 55. doi: 10.3969/j.issn.1008-4800.2020.20.027 [19] WARWICK R M, PEARSON T H, ROSWAHYUMI. Detection of pollution effects on marine macrobenthos further evaluation of the species abundance biomass method [J]. Marine Biology, 1987, 95: 193 − 200. doi: 10.1007/BF00409005 [20] WARWICK R M, CLARKEK R. Relearning the ABC: taxonomic changes and abundance biomass relationship in disturbed benthic communities [J]. Marine Biology, 1994, 118: 737 − 744. [21] 周细平, 徐帅良, 吴培芳, 等. 九龙江口海门岛红树林湿地大型底栖动物群落生态研究[J]. 应用海洋学学报, 2019, 38(1): 24 − 32. doi: 10.3969/J.ISSN.2095-4972.2019.01.003 [22] 胡桂坤, 秦璐璐, 李郁郁, 等. 基于ABC曲线的天津潮间带生物群落受扰动的分析[J]. 天津科技大学学报, 2019, 34(5): 57 − 62. [23] 唐以杰, 余世孝, 柯芝军, 等. 用ABC曲线法评价湛江红树林自然保护区的环境状况[J]. 广东教育学院学报, 2006, 26(3): 70 − 74. [24] 韩庆喜, 袁泽轶, 李宝泉, 等. 山东荣成临洛北湾夏季大型底栖动物群落生态学初步研究[J]. 海洋科学, 2012, 36(9): 17 − 23. [25] HAMMER ø, HARPER D A T, RYAN P D. PAST: Paleontological Statistics Software Package for Education and Data Analysis [J]. Palaeontologia Electronica, 2001, 4(1): 1 − 9. [26] 朱文涛, 秦传新, 段丁毓, 等. 大亚湾典型珊瑚礁区域冬春季节底栖生物分布研究[J]. 海洋渔业, 2020, 42(2): 170 − 182. doi: 10.3969/j.issn.1004-2490.2020.02.005 [27] 王丽荣, 陈锐球, 赵焕庭. 徐闻珊瑚礁自然保护区礁栖生物初步研究[J]. 海洋科学, 2008, 32(2): 56 − 62. [28] 马文刚, 夏景全, 魏一凡, 等. 三亚蜈支洲岛海洋牧场近岛区底表大型底栖动物群落结构及评价[J]. 热带海洋学报, 2022, 41(3): 135 − 146. doi: 10.11978/2021125 [29] 董 栋, 李新正, 王洪法, 等. 海南岛三亚珊瑚礁区大型底栖动物群落特征[J]. 海洋科学, 2015, 39(3): 83 − 91. [30] 杨 东, 周政权, 张建设, 等. 烟台牟平海洋牧场夏季大型底栖动物群落特征[J]. 海洋科学, 2017, 41(5): 134 − 143. [31] 黄丁勇, 王建佳, 陈甘霖, 等. 亚龙湾珊瑚礁大型礁栖生物的群落结构及生态警示[J]. 生态学杂志, 2021, 40(2): 412 − 426. doi: 10.13292/j.1000-4890.202102.033 [32] 李元超, 吴钟解, 梁计林, 等. 近15年西沙群岛长棘海星暴发周期及暴发原因分析[J]. 科学通报, 2019, 64(33): 3478 − 3484. [33] 李元超, 梁计林, 吴钟解, 等. 长棘海星的暴发及其防治[J]. 海洋开发与管理, 2019, 36(8): 9 − 12. doi: 10.3969/j.issn.1005-9857.2019.08.002 [34] 宋星宇, 刘华雪, 黄良民, 等. 南海北部夏季基础生物生产力分布特征及影响因素[J]. 生态学报, 2010, 30(23): 6049 − 6417. [35] 蔡立哲, 马 丽, 高 阳. 海洋底栖动物多样性指数污染程度评价标准的分析[J]. 厦门大学学报(自然科学版), 2002, 41(5): 641 − 646. [36] 李永祺, 丁美丽. 海洋污染生物学[M]. 北 京: 海洋出版社. 1991: 445-449. [37] 叶锦昭. 西沙群岛环境水文特征[J]. 中山大学学报(自然科学版), 1996, 35(S1): 15 − 21. [38] 李元超, 杨 毅, 郑新庆, 等. 海南三亚后海海域珊瑚礁生态系统的健康状况及其影响因素[J]. 生态学杂志, 2015, 34(4): 1105 − 1112. doi: 10.13292/j.1000-4890.20150304.018 [39] 吴钟解, 张光星, 陈石泉, 等. 海南西瑁洲岛周边海域造礁石珊瑚空间分布及其生态系统健康评价[J]. 应用海洋学学报, 2015, 34(1): 133 − 140. doi: 10.3969/J.ISSN.2095-4972.2015.01.017 [40] 吴钟解, 王道儒, 涂志刚, 等. 西沙生态监控区造礁石珊瑚退化原因分析[J]. 海洋学报, 2011, 33(4): 140 − 146. [41] 李元超, 陈石泉, 郑新庆, 等. 永兴岛及七连屿造礁石珊瑚近10年变化分析[J]. 海洋学报, 2018, 40(8): 99 − 111. [42] 张 瑜, 任瑜潇, 刘相波, 等. 潜水旅游对珊瑚礁生态系统影响研究的进展[J]. 热带生物学报, 2021, 12(2): 261 − 270. doi: 10.15886/j.cnki.rdswxb.2021.02.016 -

 点击查看大图
点击查看大图
图(5) / 表 (2)
计量
- 文章访问数: 482
- HTML全文浏览量: 238
- PDF下载量: 28
- 被引次数: 0





