


-
植物群落是种类多样的植物在特定时间和空间下的有机组合,其内部存在着错综复杂的相互关系,可通过物种组成、植物区系成分、外貌特征和物种多样性等特征来反映[1-3]。植物群落结构作为生态系统功能和形成过程的基础[4],决定了物种在群落内的生态位和竞争格局,能够反映不同种群个体在群落中的组成情况及更新机制[5],是群落形成的主要机制之一,在森林生态系统中起着重要的作用。物种多样性一直是生态学领域研究的一个热点问题[6],是衡量群落结构和功能复杂性的尺度,不仅可以反映群落与环境的相互关系[7],也可以揭示群落的发展阶段、更新和生境状况等特征[8-11],对群落的稳定和发展具有重要作用。
南亚松(Pinus latteri )别名海南松,是一种典型的热带针叶林树种,主要分布于马来半岛、中南半岛、菲律宾等地区,在我国的海南、广西南部和广东南部也有分布。南亚松具有耐贫瘠、耐干旱、耐高温、耐火烧和耐海盐风等特点,是热带地区荒山荒地景观美化和森林产业发展等的优质树种,也是我国南部沿海生态造林的主要树种,具有很好的应用前景[12-14]。与其他松类树种相比,国内外学者对南亚松的研究虽然较早,研究报道尚为稀少,研究主要涉及以下几个方面:1)树脂道和松脂的特性研究[15-16];2)南亚松产脂力的研究[17];3)松香和松节油的特性研究[18-20];4)南亚松木材材性研究[21-22];5)南亚松林基地建设与管理研究[23-24];6)南亚松育苗和造林技术研究[25-26];7)南亚松种群结构与分布格局研究[27~28];8)南亚松遗传多样性研究[29]。对南亚松群落结构及物种多样性特征进行系统且全面的调查研究还鲜有报道。南亚松多为散生,而在我国海南热带雨林国家公园管理局霸王岭分局拥有面积较大、分布相对集中的南亚松群落[12,30-31],同时,该区域也是海南长臂猿(Nomascus hainanus)全球唯一的栖息地[32]。相关研究表明,海南长臂猿主要活动在霸王岭海拔 800~1 200 m的热带雨林内[33],与南亚松群落的分布范围高度重叠,表明两者存在潜在的紧密联系。因此,南亚松群落的稳定演替发展对海南长臂猿的长远发展至关重要。近年来,因长时间的人类活动及其他原因,南亚松林的分布面积在不断减少,群落的物种多样性也逐渐丧失,这可能会不利于海南长臂猿的生存和繁衍。开展南亚松群落结构及物种多样性调查,探明该区域南亚松群落的性质是必要的,这不仅能给南亚松群落的物种多样性保护与可持续利用提供理论依据,对海南长臂猿的长远发展也具有特殊意义。本研究以海南热带雨林国家公园管理局霸王岭分局的南亚松林为研究对象,对其物种组成、地理成分、群落结构、物种多样性进行调查,旨在为南亚松群落的物种多样性保护与持续利用提供参考。
-
海南热带雨林国家公园管理局霸王岭分局位于海南岛西南部,地理坐标为108°55′58″~109°17′39″E,18°48′36″~19°12′38″N,面积872.16 km2,约占海南热带雨林国家公园总面积的1/5,是位于海南热带雨林国家公园西部的第二个片区,区域范围主要包括昌江县的七叉镇、王下乡,东方市的东河镇、江边乡,以及白沙县的青松乡。
海南热带雨林国家公园管理局霸王岭分局地形复杂,以山地为主,可分为北部和南部两块山体,海拔范围为100~1 654 m;该地区属热带季风气候,四季不明显,受季风影响较大;年均温度21~24 ℃;平均年降雨量1 490~1 657 mm,分布不均匀,雨量主要集中在7−10月;随海拔升高雨量逐渐增加,相对湿度加大,山上多见雾露,年平均相对湿度84.2%;土壤主要为山地砖红壤和山地黄壤。
-
于2022年4月至8月,在海南热带雨林国家公园管理局霸王岭分局南亚松群落中选取61个10 m×10 m的样方开展调查,总面积为6 100 m2(表1)。对样方内的所有植物进行记录,其中,乔木和灌木记录其物种名称、胸径、高度和冠幅;灌木小苗、草本及藤本记录其物种名、数量;并记录样方的经纬度、海拔、坡度、坡向、土壤类型、郁闭度等信息。对现场无法鉴定的植物进行拍照并采集标本,带回到实验室进行室内鉴定。
表 1 南亚松林调查样方统计表
序号 海拔/m 样方大小/m2 样方数/个 面积/m2 1 600~700 100 19 1 900 2 700~800 100 22 2 200 3 800~900 100 20 2 000 -
根据调查样方实际情况,将群落内木本植物划分为8个径级(DBH),分别为:1 cm≤DBH≤3 cm、3 cm<DBH≤6 cm、6 cm<DBH≤10 cm、10 cm<DBH≤20 cm、20 cm<DBH≤30 cm、30 cm<DBH≤40 cm、40 cm<DBH≤50 cm、DBH>50 cm。树高结构(H)划分为5层,分别为1 m≤H≤5 m、5 m<H≤10 m、10 m<H≤15 m、15 m<H≤20 m、H>20 m。
群落中南亚松的径级划分为6个,分别为:1 cm≤DBH≤10 cm、10 cm<DBH≤20 cm、20 cm<DBH≤30 cm、30 cm<DBH≤40 cm、40 cm<DBH≤50 cm、DBH>50 cm。树高结构划分为5层,5 m<H≤10 m、10 m<H≤15 m、15 m<H≤20 m、20 m<H≤25 m、H>25 m。
-
根据调查数据计算物种丰富度指数(S)、重要值(IV)、Shannon-Wiener指数(H´)、Simpson指数(P)、Pielou指数(E)。
(1)物种丰富度指数(S):
$$ S =样方中出现物种数目。 $$ (2)重要值 (IV):
$$ IV=(相对密度+相对显著度+相对频度)/3\text{;} $$ $$ \begin{split} 相对密度=& 某个物种的个体数/\\& 全部物种的个体数 \times 100\text{%}\text{;} \end{split}$$ $$ \begin{split} 相对显著度=& 某个物种的胸高断面积/ \\&全部物种的总胸高断面积 \times 100\text{%} \text{;} \end{split}$$ $$ \begin{split} & 相对频度=某个物种在样方中出现的次数/ \\& \quad \quad \quad 全部物种在样方中出现的总次数 \times 100\text{%}。 \end{split}$$ (3)Shannon-wiener指数值计算公式为:
$$ {H^{'}} = - \sum\limits_{i = 1}^S {Pi\ln Pi} \text{,} $$ 式中,S 表示调查样方的物种数目;Pi表示第i个物种的相对密度(Pi = Ni/N;N为所有物种的个数之和;Ni代表第i种的个体数)。
(4)Simpson指数计算公式为:
$$ P = 1 - \sum\limits_{i = 1}^S {P{i^2}}, $$ 式中,S 表示调查样方的物种数目;Pi表示第i个物种的相对密度(Pi = Ni/N;N为所有物种的个数之和;Ni代表第i种的个体数)。
(5)Pielou指数计算公式为:
$$ E = {H^{'}}/\ln S \text{,} $$ 式中,H´表示Shannon-wiener指数,S 表示调查样方的物种数目。
-
原始数据的记录、统计和绘图等运用办公软件EXCEL2019、ArcMap10.2进行处理。
-
从表2可知,霸王岭南亚松群落物种丰富,在此次调查中共记录到230个物种,隶属66科146属。按照植物类群来分,蕨类植物有3科4属4种;种子植物有63科142属226种,其中,裸子植物有3科3属4种,被子植物60科139属222种(含单子叶植物7科11属11种和双子叶植物55科128属211种)。
表 2 霸王岭南亚松群落的物种组成
植物类群 科数 属数 种数 蕨类植物 3 4 4 裸子植物 3 3 4 被子植物 60 139 222 单子叶植物 7 11 11 双子叶植物 55 128 211 合计 66 146 230 群落的优势物种是群落组成特征的基础,对群落的构建起着决定性作用。霸王岭南亚松群落植物在科的分级上,以含少数种的科(1~10 种)占主导地位,其中,物种数≥10种的科有5科,分别为樟科(Lauraceae)(22种)、大戟科(Euphorbiaceae)(20种)、茜草科(Rubiaceae)(17种)、壳斗科(Fagaceae)(14种)、桃金娘科(Myrtaceae)(13种),5科共含38属86种,占其总属数的26.03 %,总种数的37.39%,是该南亚松林群落植物的主要组成科(表3)。
表 3 霸王岭南亚松群落的优势科属
序号 科名 属数 种数 属名 种数 1 樟科(Lauraceae) 8 22 蒲桃属(Syzygium) 10 2 大戟科(Euphorbiaceae) 13 20 山矾属(Symplocos) 7 3 茜草科(Rubiaceae) 11 17 柯属(Lithocarpus) 6 4 壳斗科(Fagaceae) 3 14 青冈属(Cyclobalanopsis) 5 5 桃金娘科(Myrtaceae) 3 13 琼楠属(Beilschmiedia) 5 6 — — 木姜子属(Litsea) 5 合计 38(26.03) 86(37.39) 38(16.52) 注:“— ”表示没有,括号内数据为占比(%)。 霸王岭南亚松群落中的优势属不明显,大部分属仅含1~4种,物种数≥5种的有6属,分别是蒲桃属(Syzygium)、山矾属(Symplocos)、柯属(Lithocarpus)、青冈属(Cyclobalanopsis)、琼楠属(Beilschmiedia)、木姜子属(Litsea),共含38个种 ,占总种数的16.52%(表3)。
优势种是依据物种重要值的大小来确定,如表4所示,在南亚松群落中,最主要的5种植物依次为南亚松(Pinus latteri )、九节(Psychotria asiatica)、银柴(Aporosa dioica)、托盘青冈(Quercus patelliformis)、黄牛木(Cratoxylum cochinchinense),其重要值分别为29.01%、8.82%、5.56%、5.46%、5.36%。在南亚松群落230种植物中重要值前10名的植物物种仅占总物种数4.35%的情况下,其重要值高达75.02%。
表 4 霸王岭南亚松群落植物重要值前10名物种
序号 物种名 相对密度/% 相对频度/% 相对显著度/% 重要值/% 1 南亚松(Pinus latteri) 18.90 8.01 60.14 29.02 2 九节(Psychotria asiatica) 19.22 6.96 0.30 8.83 3 银柴(Aporosa dioica) 11.65 4.07 0.97 5.56 4 托盘青冈(Quercus patelliformis) 2.81 3.02 10.56 5.46 5 黄牛木(Cratoxylum cochinchinense) 10.24 5.25 0.59 5.36 6 木荷(Schima superba) 6.53 4.46 3.04 4.68 7 黄杞(Engelhardia roxburghiana) 6.35 5.64 1.72 4.57 8 广东山胡椒(Lindera kwangtungensis) 9.34 3.02 1.25 4.54 9 野漆(Toxicodendron succedaneum) 5.58 4.72 0.55 3.62 10 细齿叶柃(Eurya nitida) 3.85 3.94 2.40 3.40 -
参照吴征镒等的划分标准[34-36],对霸王岭南亚松群落内植物科属的分布类型进行划分,蕨类植物除外。结果(表5)表明,在科级水平上,将研究区种子植物63科地理成分划分为9个类型,热带成分区分布科占主导地位,共计40科,占总科数(除世界广布科外)的80.00%,具有极强的热带性质。其中,以泛热带广布类型为主,共计29科,主要有大戟科、樟科、芸香科(Rutaceae)等;其次是世界广布科14个,占总科数的20.63%,包含茜草科、蔷薇科(Rosaceae)和禾本科(Rosaceae)等。温带成分区分布的科只有10个,占总科数(除世界广布科外)的20.00%,温带成分中,北温带分布8科,以壳斗科为主,东亚及北美间断分布2科,分别是木兰科(Magnoliaceae)、鼠刺科(Iteaceae)。
表 5 霸王岭南亚松群落种子植物的分布区类型
分布类型 科数 属数 1.世界广布 13 3 2.泛热带广布 29(58.00) 36(25.90) 3.热带亚洲和热带南美间断分布 5(10.00) 4(2.88) 4.旧世界热带 2(4.00) 15(10.79) 5.热带亚洲至热带大洋洲 1(2.00) 26(18.71) 6.热带亚洲至热带非洲 — 8(5.76) 7.热带亚洲 3(6.00) 35(25.18) 小计 热带区分布(2~7) 40(80.00) 124(89.21) 8.北温带 8(16.00) 4(2.88) 9.东亚及北美间断 2(4.00) 9(6.47) 14.东亚 — 1(0.72) 小计 温带区分布(8~12) 10(20.00) 14(10.07) 15.中国特有 — 1(0.72) 合计 63(100.00) 142(100.00) 注:“—” 表示没有,括号内数据为占比(%)。 在属级水平上,将研究区种子植物142属地理成分划分为11个类型,热带成分区分布属占绝对优势,共计124属,占总属数(除世界广布属外)的89.21%;以泛热带广布和热带亚洲类型为主,其中,泛热带广布类型有36属,占总属数(除世界广布属外)的25.90%,包括榕属(Ficus)、合欢属(Albizia)、九节属(Psychotria)等;热带亚洲类型有35属,占总属数(除世界广布属外)的25.18%,包括润楠属(Machilus)、山茶属(Camellia)、乌桕属(Triadica)等。温带成分区分布的属有14个,占总属数(除世界广布科外)的10.07%,中国特有分布区仅1属为杉木属 (Cunninghamia)。
-
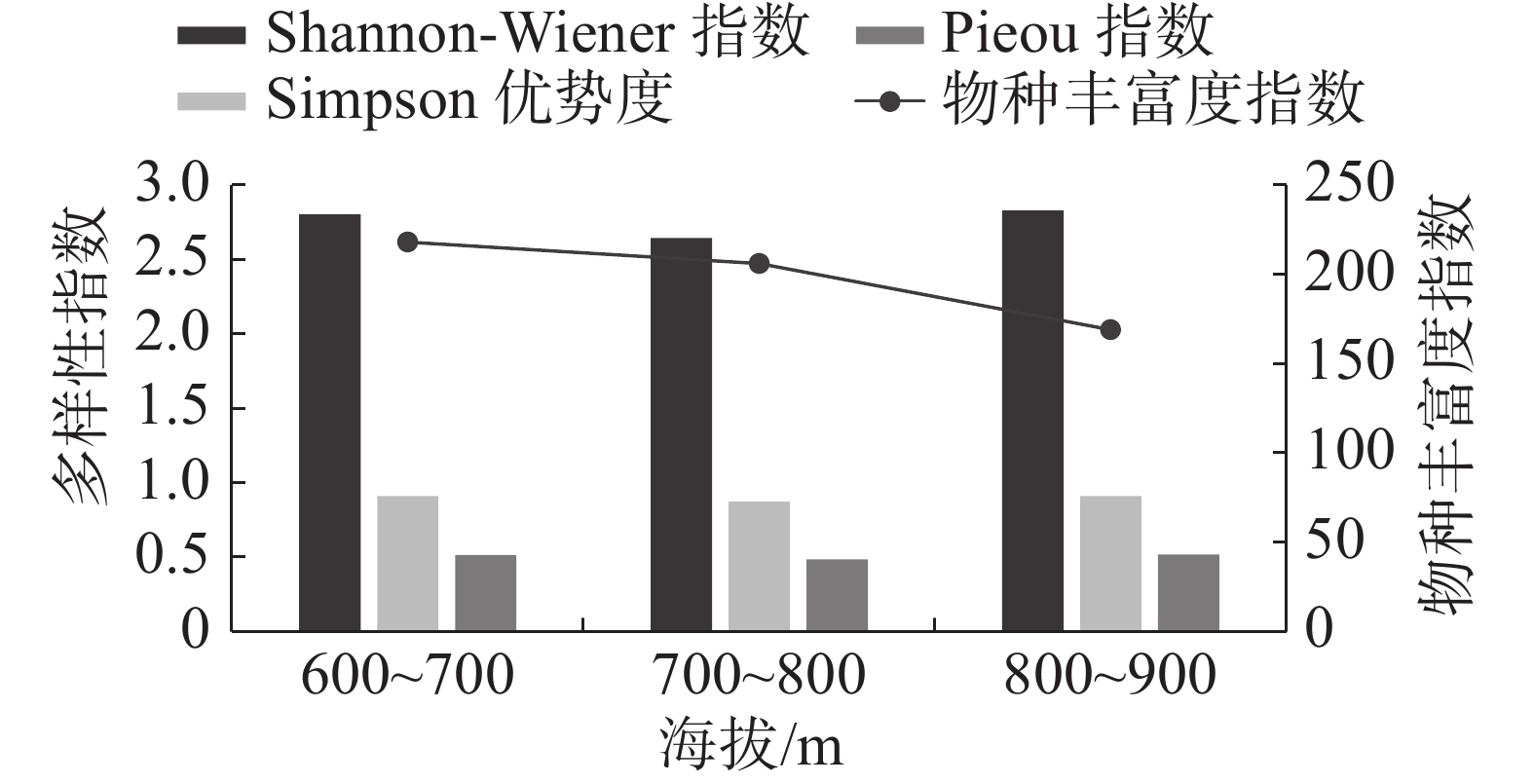
因不同海拔梯度的植物群落生境不同,所以发育着不同的植物群落。群落多样性指数也表现出随海拔梯度变化而分异的现象[37]。如图1所示,不同海拔梯度南亚松群落的物种多样性具有差异,其中,物种丰富度指数的变化趋势随着海拔的升高而降低;Shannon-Wiener指数、Simpson优势度指数和Pieou均匀度指数均随海拔的升高呈现出先降低再增加的趋势,变化趋势相同。

图 1 霸王岭不同海拔南亚松群落物种多样性
-
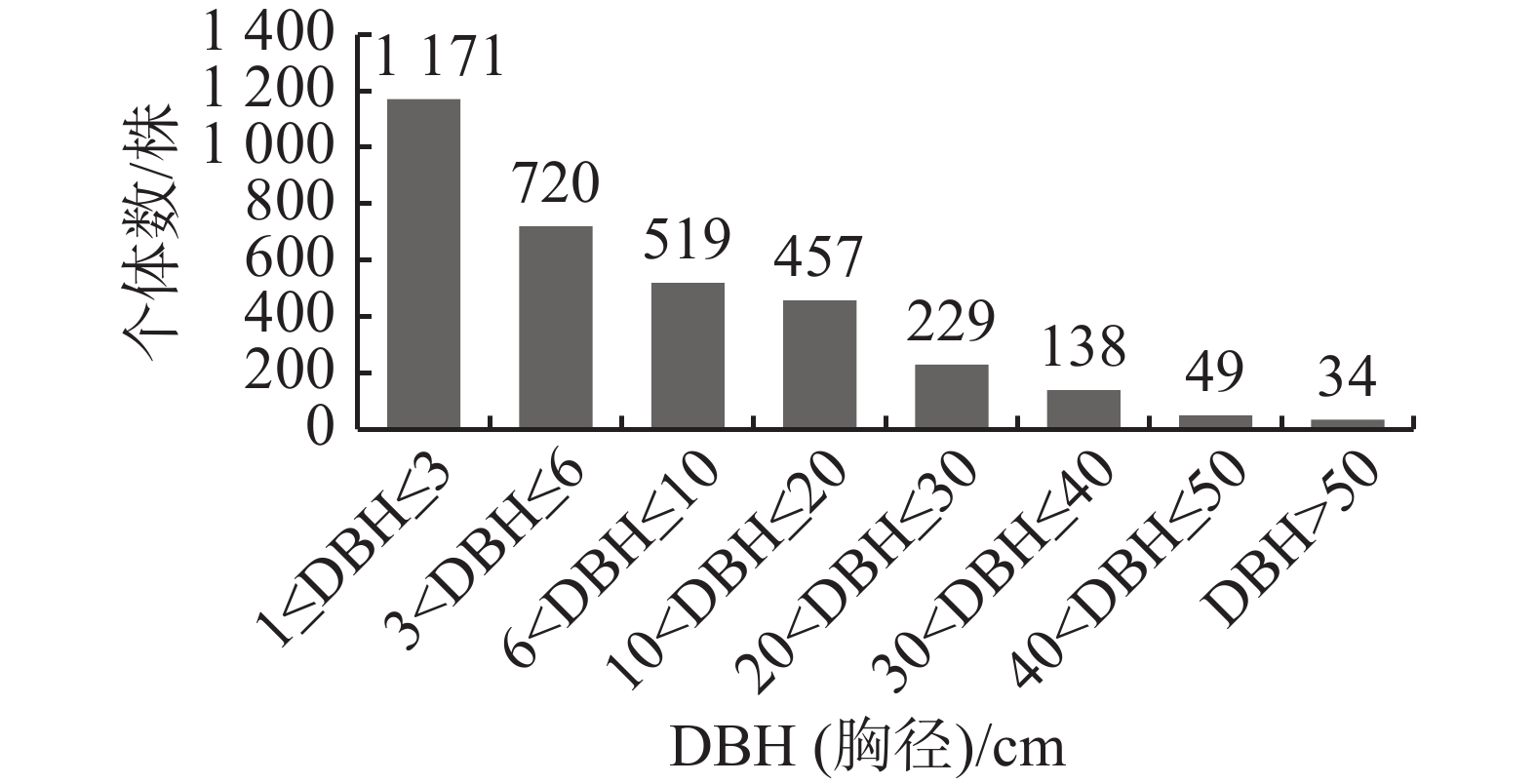
物种的径级分布不仅能反映群落的生长发育情况,还可以作为评价群落稳定性和预测演替趋势的重要指标[11]。如图2所示,霸王岭南亚松群落的径级结构呈典型的倒“J”型。样方内所有DBH(胸径)≥1 cm的物种共3 317株,其中,胸径在1 cm≤DBH≤3 cm的个体数最多,有1 171株,占总个体数的35.3%, 胸径在3 cm<DBH≤6 cm的有720株,占总个体数的21.71%;随着径级的增加,个体间竞争加剧,株数逐渐减少,但在每个径级都有一定数量的个体分布;大径级个体对维持群落乔木层的稳定性至关重要,倒“J”型的径级结构表明南亚松群落自我更新状况较好,群落能稳定演替发展。
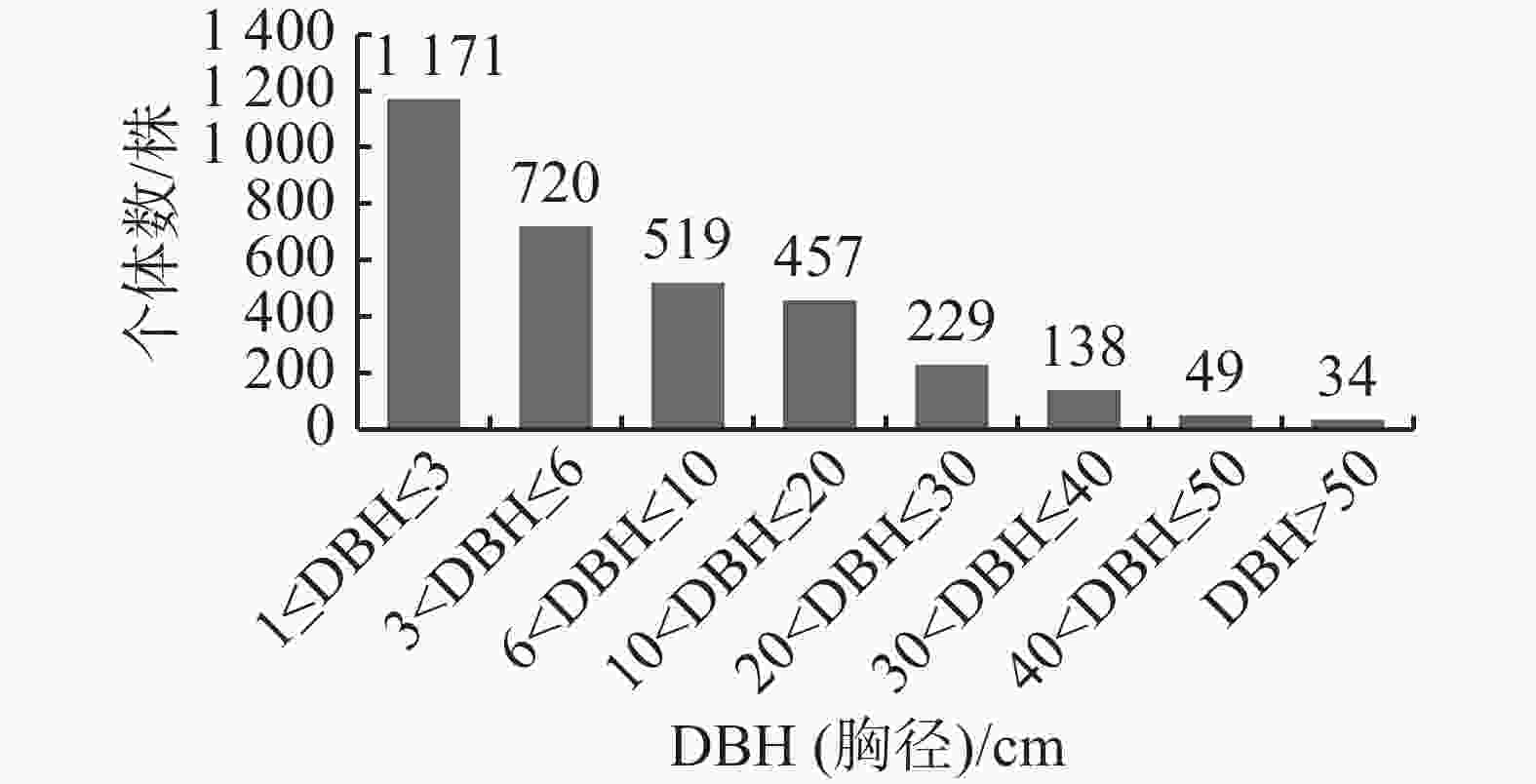
图 2 霸王岭南亚松群落径级结构
-
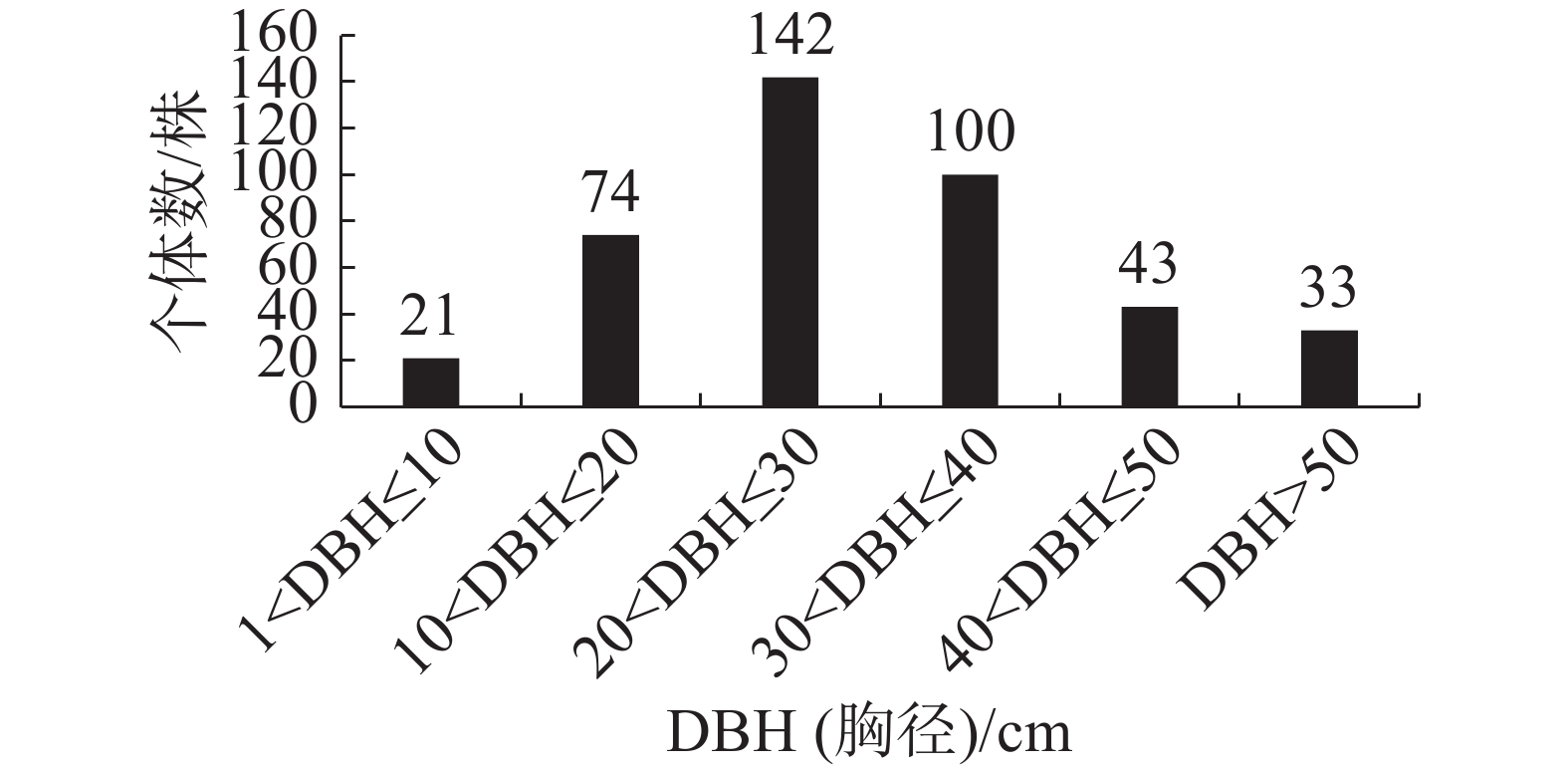
群落内南亚松的径级结构如图3所示,样方内共有413株南亚松,其中,胸径在20 cm<DBH≤30 cm的个体数最多,有142株,占南亚松总个体数的34.38%;其次是胸径在30 cm<DBH≤40 cm的个体,有100株,占南亚松总个体数的24.21%;胸径在1 cm<DBH≤10 cm的个体数最少,有21株,占南亚松总个体数的5.08%。
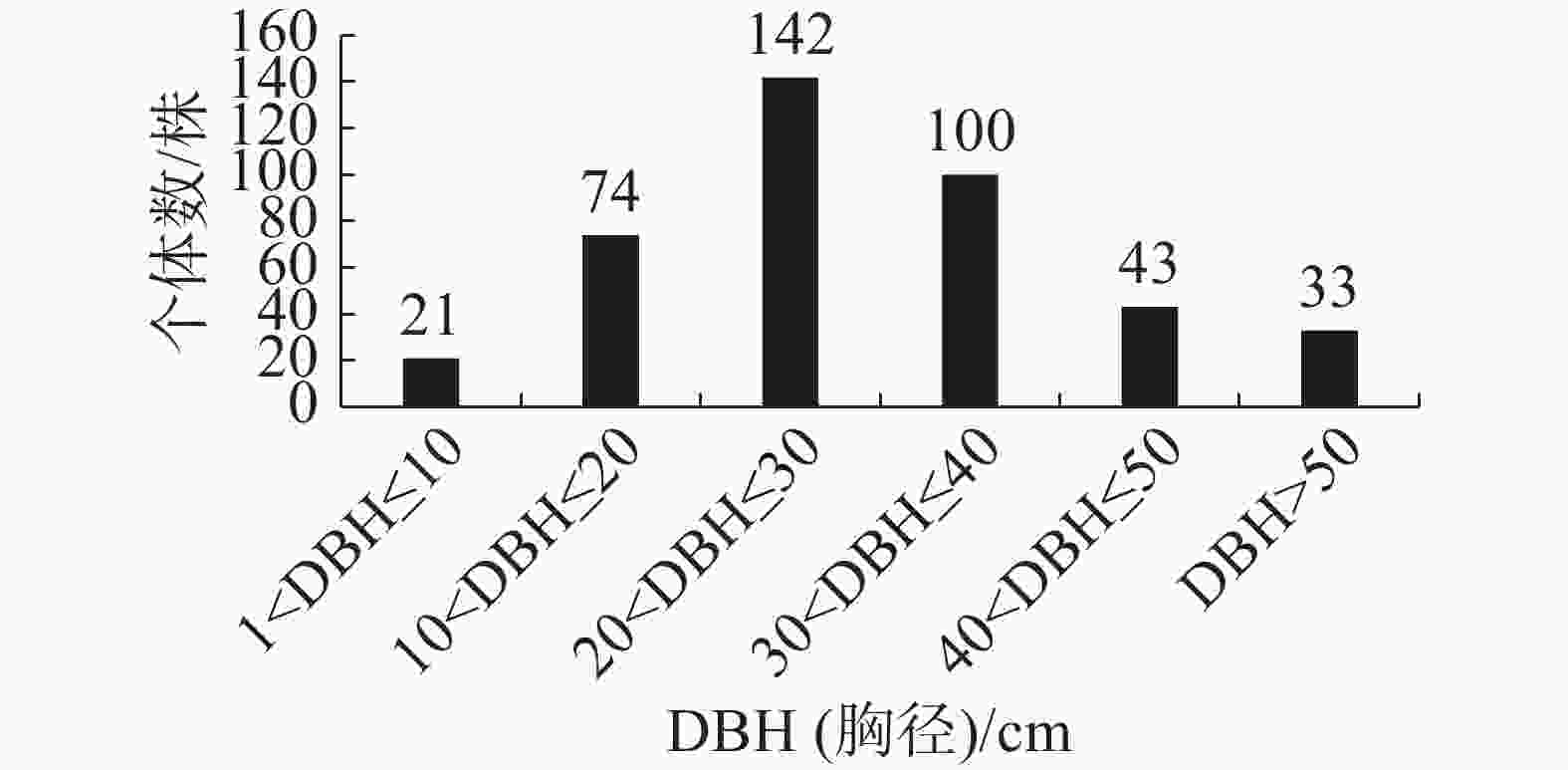
图 3 南亚松径级结构
-
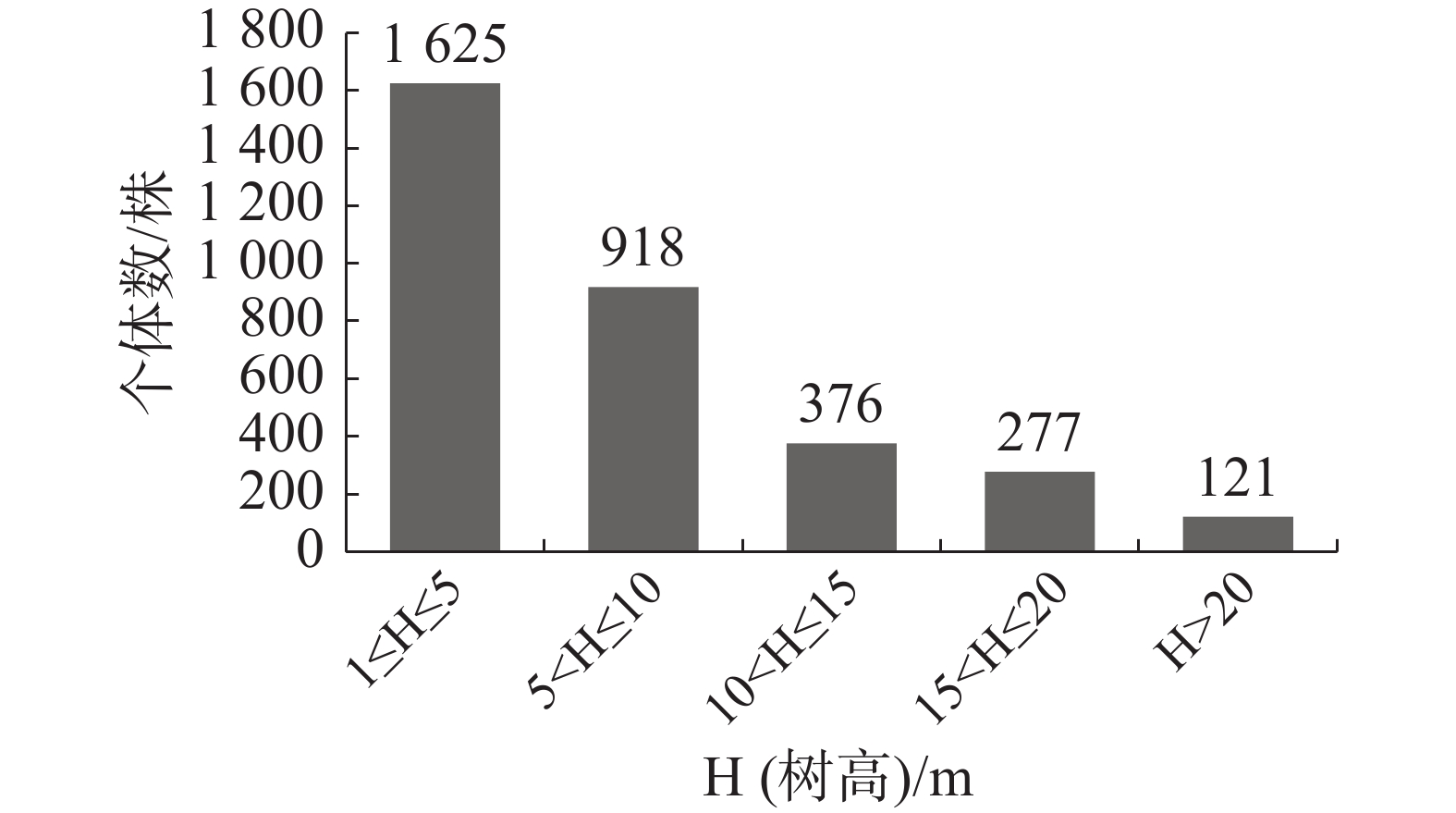
如图4所示,南亚松群落树高结构呈倒“J”型,其中,树高在1 m≤H(树高)≤5 m范围的个体数最多,有1 625株,占总个体数的49.0%,树高在5 m<H≤10 m的有918株,占总个体数的27.68%;随着高度的增加,株数逐渐减少,但每个树高级都有一定数量个体分布,说明立木株数在垂直分布上是连续的,没有明显的断层现象,群落较为成熟。
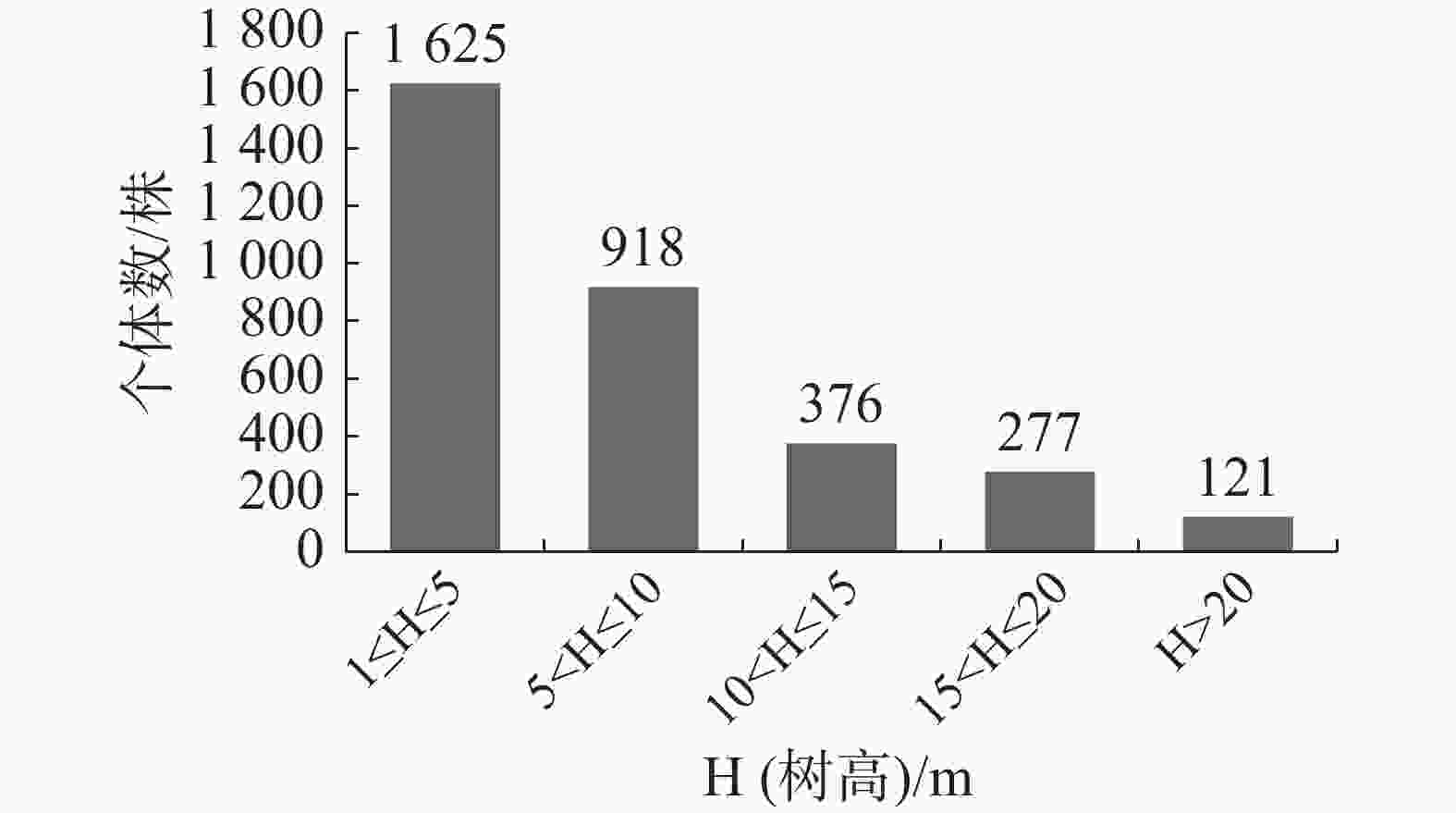
图 4 霸王岭南亚松群落树高结构
-
群落内南亚松的树高结构如图5所示,在15 m<H≤20 m范围的南亚松最多,有179株,占南亚松总个体数的43.34%;较矮与较高的个体数稀少,分别占南亚松总个体数的8.47%和7.99%。
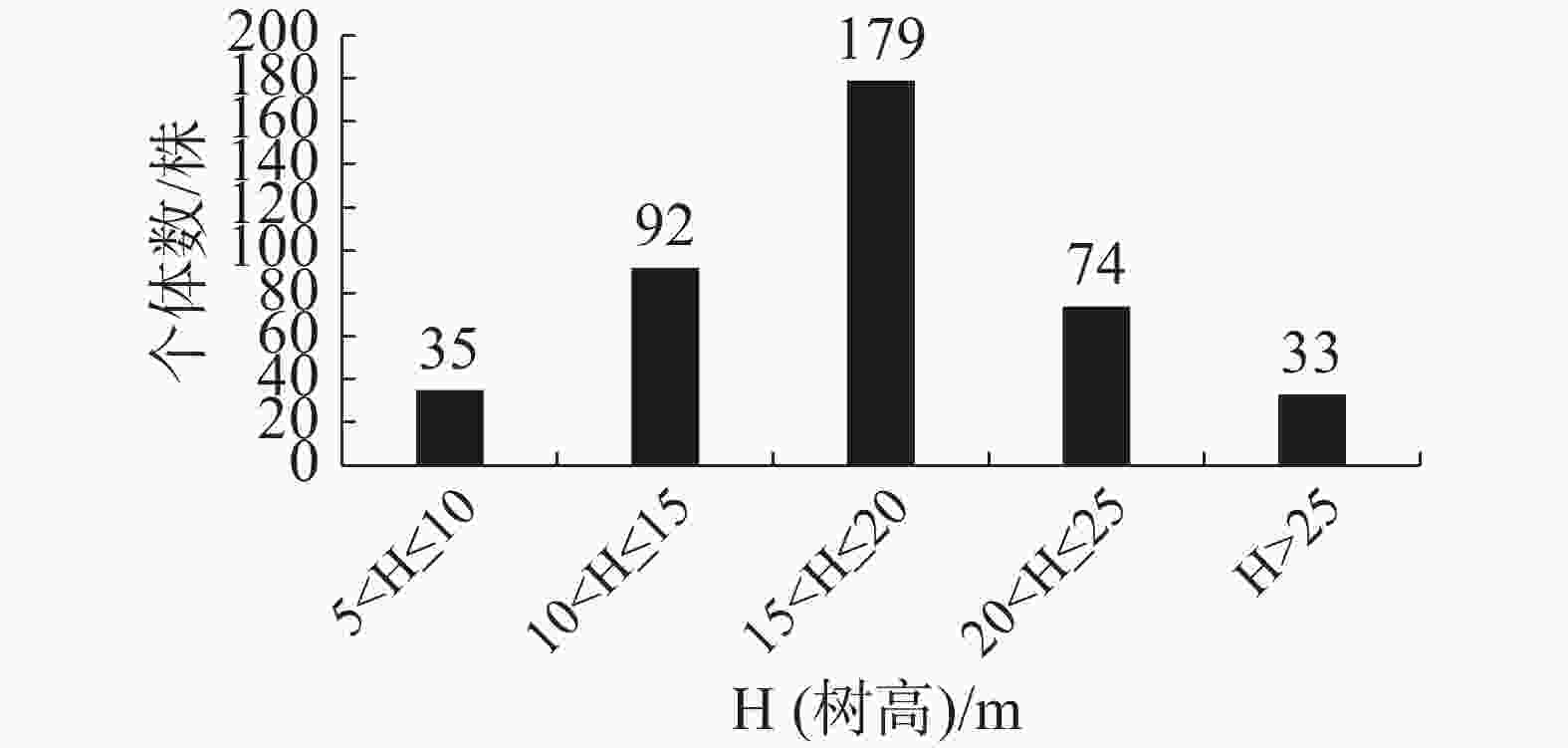
图 5 南亚松树高结构
-
霸王岭南亚松群落植物种类多样,在此次调查中,共记录到230个物种,隶属66科146属。其中,以樟科、大戟科、茜草科、壳斗科、桃金娘科为主要优势科,南亚松、九节、银柴、托盘青冈、黄牛木为主要优势种,优势属不明显。群落物种多样性与以往学者对海南霸王岭南亚松群落物种多样性的初步调查研究[28]对比发现,有百余种是未曾记录过的;而与海南长臂猿的食源和夜宿植物组成对比,发现南亚松群落内有58种海南长臂猿食源植物,占总物种数的25.22%,有53种海南长臂猿夜宿植物,占总物种数的23.04%[38-39]。可见,南亚松群落的稳定演替发展对海南长臂猿的长远发展至关重要。
霸王岭南亚松群落植物地理联系广泛,在科级水平上有9个分布类型,属级水平上有11个分布类型。分布以热带成分占据绝对优势,其中,以泛热带分布为主,说明区系组成含有强烈的泛热带性质;而温带区分布以北温带分布为主,与霸王岭是壳斗科为优势的山地雨林以北温带分布为主相符[40]。一般来说,随海拔升高,人为干扰减弱,中国特有分布将增加[41]、植物区系地理成分更加复杂[42]。而霸王岭南亚松群落中国特有属分布仅杉木属一个,表明霸王岭南亚松群落物种特有性弱,所占比例甚小的区系特征,这可能与地理分布范围狭窄,生态环境相似,人为干扰导致该地区在土地历史上没有长期孤立时段,不能形成自己独特的环境等因素有关。
霸王岭南亚松群落的物种丰富度指数的变化趋势随着海拔的升高而降低,其中,海拔梯度为600~700 m和700~800 m的物种丰富度指数明显高于800~900 m海拔梯度的物种丰富度指数,这可能与群落本身物种组成、结构和生境等特征有关,也表明南亚松群落的物种丰富度指数与海拔的变化密切相关。Shannon-Wiener指数、Simpson优势度指数和Pieou均匀度指数均随海拔的升高呈先降低再升高的趋势,变化趋势相同。关于物种多样性随海拔梯度变化的分布规律,虽然已有报道,但至今尚未达成共识。目前,分布规律主要有5种变化模式,分别是正相关、中度膨胀理论、中等海拔较低、负相关和无关[43]。Robert等[44]的研究结果表明,物种多样性随着海拔的升高呈先降低再升高的变化趋势,为中等海拔较低模式。本研究的结果与其相符,导致这种结果的原因可能有以下两点:首先,样方的分布范围不同,植物的生境也有所差别,因此,物种多样性随海拔变化的分布规律也有所差异;其次,由于南亚松的分布范围有限,海拔梯度的设置没有达到物种多样性的峰值,因此,物种多样性随海拔变化的变化模式也不同。
植物群落结构是个体对外界干扰强度、立地条件优劣、环境适应性和多样性的反映,被认为是影响群落物种组成和多样性的最重要因素之一[45]。径级和树高结构作为群落最基本的林分结构,是植物生长与环境关系的综合反映,也是评价植物群落稳定性、生长发育状况、预测群落结构发展的重要指标[46]。霸王岭南亚松群落结构复杂,其径级结构和树高结构都呈典型的倒“J”型,其中,胸径在1 cm≤DBH≤3 cm的个体数最多,有1 171株,占总个体数的35.3%,其次是3 cm≤DBH≤6 cm有720株,占总个体数的21.71%;树高在1 m≤H≤5 m范围的个体数最多,有1 625株,占总个体数的49.0%,其次5 m≤H≤10 m有918株,占总个体数的27.68%;随着径级和树高的增加,株数逐渐减少,但在每个径级和树高范围内都有一定数量个体分布,与臧润国等[47]、刘万德等[3]分别对霸王岭热带山地雨林和热带季雨林群落结构特征的研究结果相似。这些结果说明南亚松群落的更新层发育良好,不同世代间没有明显的断层现象,可以维持乔木层树种的更新,能够稳定演替发展。群落内南亚松的径级和树高结构都呈钟形,其中,胸径在20 cm≤DBH≤30 cm的个体数最多,共有179株,较小和较大范围的个体数稀少,通过与群落径级结构对比,可发现,在1 cm≤DBH≤10 cm范围,群落共有2 410株中只有21株是南亚松,10 cm≤DBH≤20 cm范围内457株中只有74株南亚松;而在20 cm≤DBH≤30 cm范围229株中有142株就是南亚松,30 cm≤DBH≤40 cm范围138株中有100株是南亚松。南亚松树高结构与群落树高结构的对比规律也是如此。南亚松作为该群落的先锋物种,已在该地生长多年,是该群落主要的大径级个体,所以其径级和树高结构都呈钟形,对维持群落稳定性起关键作用;而南亚松群落的径级和树高结构都呈倒“J”型是因为群落经过多年的演替,引入了许多其他物种,所以群落中小径级所含个体多,物种丰富,说明该群落更新良好。
综上可知,霸王岭南亚松群落物种组成丰富,含有一定比例的海南长臂猿食源和夜宿植物;其地理联系广泛,群落结构复杂,稳定性和物种多样性较高,在没有人为干扰下群落能够稳定演替发展。
海南霸王岭南亚松群落结构及物种多样性特征
DOI: 10.15886/j.cnki.rdswxb.20220089
 CSTR: 32425.14.j.cnki.rdswxb.20220089
CSTR: 32425.14.j.cnki.rdswxb.20220089
Community structure and species diversity of Pinus latteri forests in Bawangling Ridge, Hainan
-
摘要: 为了解海南霸王岭南亚松(Pinus latteri)的群落结构和物种多样性特征,采用样方法对南亚松群落的物种组成、地理成分、群落结构及物种多样性进行调查。结果表明:1)霸王岭南亚松群落植物种类丰富,共有66科146属230种,以樟科(Lauraceae)、大戟科(Euphorbiaceae)、茜草科(Rubiaceae)为主要优势科,南亚松(Pinus latteri )、九节(Psychotria asiatica)、银柴(Aporosa dioica)为主要优势种,优势属不明显;2)南亚松群落的植物地理联系广泛,以热带成分为绝对优势,含杉木属(Cunninghamia)一个特有属;3)不同海拔梯度南亚松群落的物种多样性有差异,其中,物种丰富度指数随海拔的升高而降低,而Shannon-Wiener指数、Simpson优势度指数和Pielou指数均随海拔的升高呈现先降低再增加的趋势;4)南亚松群落结构复杂,其径级和树高结构都是典型的倒“J”型,群落中南亚松的径级和树高结构呈钟形,说明南亚松群落稳定性高,更新状态良好,能够稳定演替发展。Abstract: A survey ofPinus latteri community in Bawangling Ridge, Hainan was made by using sample plotting, and the species composition, geographic composition, community structure, and species diversity ofthe P. latteri community were analyzed to have clear understanding of the community structure and species diversity of P. latteri therein. The survey indicated that the P. latteri community in Bawangling is rich in plant species and composed of 66 families, 146 genera, and 230 species, with Lauraceae, Euphoribiaceae, Rubiaceae as dominant families and P. latteri Mason, Psychotria asiatica, and Aporosa dioica as dominant species. Dominating genus is not immediately apparent. The species diversity of the P. latteri community is different and varies along the altitudinal gradient, where the species richness index decreased with the rise of the altitude, and the Shannon-Wiener index, and Pielou index decreased and then increased with the rise of the altitude. TheP. latteri community has extensive phytogeographic associations, with its tropical distribution being dominant and containing one endemic genus Cunninghamia. The P. latteri community has a complex structure with a typical inverted "J" shape in diameter and height distribution, and the diameter and tree height of P. latteri in the community are bell-shaped in structure, indicating that the P. latteri community is high in stability with good regeneration and can hence have a steady succession.
-
Key words:
- Pinus latteri /
- community structure /
- species diversity /
- Bawangling Ridge
-
表 1 南亚松林调查样方统计表
序号 海拔/m 样方大小/m2 样方数/个 面积/m2 1 600~700 100 19 1 900 2 700~800 100 22 2 200 3 800~900 100 20 2 000  下载: 导出CSV
下载: 导出CSV
表 2 霸王岭南亚松群落的物种组成
植物类群 科数 属数 种数 蕨类植物 3 4 4 裸子植物 3 3 4 被子植物 60 139 222 单子叶植物 7 11 11 双子叶植物 55 128 211 合计 66 146 230
下载: 导出CSV
表 3 霸王岭南亚松群落的优势科属
序号 科名 属数 种数 属名 种数 1 樟科(Lauraceae) 8 22 蒲桃属(Syzygium) 10 2 大戟科(Euphorbiaceae) 13 20 山矾属(Symplocos) 7 3 茜草科(Rubiaceae) 11 17 柯属(Lithocarpus) 6 4 壳斗科(Fagaceae) 3 14 青冈属(Cyclobalanopsis) 5 5 桃金娘科(Myrtaceae) 3 13 琼楠属(Beilschmiedia) 5 6 — — 木姜子属(Litsea) 5 合计 38(26.03) 86(37.39) 38(16.52) 注:“— ”表示没有,括号内数据为占比(%)。
下载: 导出CSV
表 4 霸王岭南亚松群落植物重要值前10名物种
序号 物种名 相对密度/% 相对频度/% 相对显著度/% 重要值/% 1 南亚松(Pinus latteri) 18.90 8.01 60.14 29.02 2 九节(Psychotria asiatica) 19.22 6.96 0.30 8.83 3 银柴(Aporosa dioica) 11.65 4.07 0.97 5.56 4 托盘青冈(Quercus patelliformis) 2.81 3.02 10.56 5.46 5 黄牛木(Cratoxylum cochinchinense) 10.24 5.25 0.59 5.36 6 木荷(Schima superba) 6.53 4.46 3.04 4.68 7 黄杞(Engelhardia roxburghiana) 6.35 5.64 1.72 4.57 8 广东山胡椒(Lindera kwangtungensis) 9.34 3.02 1.25 4.54 9 野漆(Toxicodendron succedaneum) 5.58 4.72 0.55 3.62 10 细齿叶柃(Eurya nitida) 3.85 3.94 2.40 3.40
下载: 导出CSV
表 5 霸王岭南亚松群落种子植物的分布区类型
分布类型 科数 属数 1.世界广布 13 3 2.泛热带广布 29(58.00) 36(25.90) 3.热带亚洲和热带南美间断分布 5(10.00) 4(2.88) 4.旧世界热带 2(4.00) 15(10.79) 5.热带亚洲至热带大洋洲 1(2.00) 26(18.71) 6.热带亚洲至热带非洲 — 8(5.76) 7.热带亚洲 3(6.00) 35(25.18) 小计 热带区分布(2~7) 40(80.00) 124(89.21) 8.北温带 8(16.00) 4(2.88) 9.东亚及北美间断 2(4.00) 9(6.47) 14.东亚 — 1(0.72) 小计 温带区分布(8~12) 10(20.00) 14(10.07) 15.中国特有 — 1(0.72) 合计 63(100.00) 142(100.00) 注:“—” 表示没有,括号内数据为占比(%)。
下载: 导出CSV
-
[1] 李俊清. 森林生态学[M].2版. 北京: 高等教育出版社, 2010: 211-272. [2] SCHWILKD W, ACKERLYD. Limiting similarity and functional diversity along environmental gradients [J]. Ecology Letters, 2005, 8(3): 272 − 281. doi: 10.1111/j.1461-0248.2004.00720.x [3] 刘万德, 臧润国, 丁易. 海南岛霸王岭两种典型热带季雨林群落特征[J]. 生态学报, 2009, 29(7): 3465 − 3476. doi: 10.3321/j.issn:1000-0933.2009.07.003 [4] 陈云, 王婷, 李培坤, 等. 河南木札岭温带落叶阔叶林群落特征及主要乔木空间分布格局[J]. 植物生态学报, 2016, 40(11): 1179 − 1188. doi: 10.17521/cjpe.2016.0192 [5] 冯立新, 买凯乐, 杨善云, 等. 桂西北红椿(Toona ciliata)群落结构与更新特征[J]. 东北林业大学学报, 2022, 50(7): 28 − 32. [6] 刘欢, 李文君, 陈杰, 等. 陕西米仓山自然保护区6种典型天然林群落的物种多样性[J]. 西北农林科技大学学报(自然科学版), 2017, 45(7): 29 − 39. [7] 茹文明, 张金屯, 张峰, 等. 历山森林群落物种多样性与群落结构研究[J]. 应用生态学报, 2006, 17(4): 561 − 566. [8] 郑凤英, 杜伟, 苟学文. 威海市区黑松林群落的物种多样性特征[J]. 生态环境, 2008, 17(5): 1965 − 1969. [9] LIM B K, ENGSTROM M. Species diversity of bats (Mammalia: Chiroptera) in Iwokrama Forest, Guyana, and the Guianan subregion: implications for conservation [J]. Biodiversity and Conservation, 2001, 10(4): 613 − 657. doi: 10.1023/A:1016660123189 [10] VELLAK K, PAAL J. Diversity of bryophyte vegetation in some forest types in Estonia: a comparison of old unmanaged and managed forest [J]. Biodiversity and Conservation, 1999, 8(12): 1595 − 1620. doi: 10.1023/A:1008927501623 [11] PANDEY S , SHUKLA R. Plant diversity in managed sal (Shorea robusta Gaertn. ) forests of Gorakhpur, India: species composition, regeneration and conservation [J]. Biodiversity and Conservation, 2003, 12(11): 2295 − 2319. doi: 10.1023/A:1024589230554 [12] 陈焕镛. 海南植物志: 第1卷[M]. 北京: 科学出版社, 1964: 211-212. [13] 中国植被编辑委员会. 中国植被[M]. 北京: 科学出版社, 1980: 247-249. [14] 广东省植物研究所. 广东植被[M]. 北京: 科学出版社, 1976: 98-105. [15] 张梦琴, 邓昭成. 南亚松木材树脂道的扫描电镜观察初报[J]. 南京林产工业学院学报, 1983, 26(3): 147 − 149. [16] 马文秀. 南亚松松脂二元树脂酸鉴定及含量测定[J]. 南京林产工业学院学报, 1988, 31(4): 89 − 95. [17] 张梦琴, 邓昭成. 南亚松产脂力的研究[J]. 林产化学与工业, 1990, 10(1): 29 − 37. [18] 宋湛谦. 我国松香、松节油的化学改性和利用[J]. 林产化学与工业, 1982, 2(1): 22 − 25. [19] 粟子安, 梁志勤, 秦文龙, 等. 中国松香、松节油主要化学组成的研究[J]. 林业科学, 1980, 16(3): 214 − 220. [20] 王石发. 柬埔寨南亚松松香和松节油的化学组成特征[J]. 林产化学与工业, 2007, 27(5): 31 − 36. [21] 徐慧兰, 陈虎, 颜培栋, 等. 海南南亚松木材材性的研究[J]. 中南林业科技大学学报, 2017, 37(5): 92 − 95. [22] 徐慧兰, 颜培栋, 杨章旗. 南亚松木材材性研究Ⅱ: 天然林群落间的比较[J]. 中南林业科技大学学报, 2020, 40(11): 167 − 172. [23] 邓昭成. 海南霸王岭林区南亚松林基地的经营管理[J]. 海南林业科技, 1988, 4(5): 29 − 33. [24] 邓昭成. 南亚松树基地建设及松脂生产经营管理[J]. 林产化学与工业, 1993, 13(增刊): 84 − 86. [25] 韩超, 徐建民, 童清, 等. 南亚松引种种源/家系苗期选择研究[J]. 中国农学通报, 2014, 30(19): 7 − 12. [26] 陶辉光. 南亚松造林技术研究[J]. 海南林业科技, 1991, 8(2): 13 − 16. [27] 黄清麟, 陈永富, 杨秀森. 海南霸王岭林区南亚松天然林乔木层结构特征研究[J]. 林业科学研究, 2002, 15(6): 741 − 745. [28] 黄运峰, 杨小波, 党金玲, 等. 海南霸王岭南亚松种群结构与分布格局[J]. 福建林业科技, 2009, 36(2): 1 − 5. doi: 10.3969/j.issn.1002-7351.2009.02.001 [29] 陈虎, 颜培栋, 杨章旗. 海南霸王岭南亚松天然林群落遗传多样性的SSR分析[J]. 植物遗传资源学报, 4, 15(4): 706-712. [30] 中国科学院中国植物志编辑委员会. 中国植物志: 第7卷[M]. 北京: 科学出版社, 1978: 260-263. [31] 广东省林业科学研究所. 海南主要经济树木[M]. 北京: 农业出版社, 1964: 46-551. [32] LI P, GARBER P A, BI Y, et al. Diverse grouping and mating strategies in the Critica11y Endangered Hainan gibbon nomascus hainanus [J]. Primates; Journal of Primatology, 2022, 63(3): 237 − 243. doi: 10.1007/s10329-022-00983-5 [33] LIU Z H, ZHANG Y Z, JIANG H S, et al. Population structure of Hylobates concolor in Bawanglin Nature Reserve, Hainan, China [J]. American Journal of Primatology, 1989, 19(4): 247 − 254. doi: 10.1002/ajp.1350190406 [34] 吴征镒. 《世界种子植物科的分布区类型系统》的修订[J]. 云南植物研究, 2003, 24(5): 535 − 538. [35] 吴征镒. 中国种子植物属的分布区类型[J]. 云南植物研究, 1991, 13(S4): 1 − 139. [36] 吴征镒, 周浙昆, 孙航. 种子植物分布区类型及其起源和分化[M].昆明: 云南科技出版社, 2006. [37] 马克平, 黄建辉, 于顺利, 等. 北京东灵山地区植物群落多样性的研究Ⅱ丰富度、均匀度和物种多样性指数[J]. 生态学报, 1995, 15(3): 268 − 277. doi: 10.3321/j.issn:1000-0933.1995.03.006 [38] 唐玮璐, 毕玉, 金崑. 海南热带雨林国家公园海南长臂猿食源植物组成[J]. 野生动物学报, 2021, 42(3): 675 − 685. [39] 唐玮璐, 金崑. 海南热带雨林国家公园海南长臂猿夜宿生境选择初步研究[J]. 北京林业大学学报, 2021, 43(2): 113 − 126. [40] 吕晓波. 海南霸王岭以壳斗科植物为优势的山地雨林群落结构及物种多样性研究[D]. 海口: 海南大学, 2012. [41] 苏文苹. 高黎贡山南段东西坡中山湿性常绿阔叶林群落学比较研究[D]. 昆明: 西南林学院, 2007. [42] 巫翠华, 张利利, 乔卫国, 等. 紫柏山自然保护区植物多样性垂直分布格局及其影响因素[J]. 福建农林大学学报(自然科学版), 2021, 50(5): 630 − 635. [43] 王小娜, 徐当会, 王谢军, 等. 祁连山灌丛群落结构特征随海拔梯度和经度的变化[J]. 生态环境学报, 2022, 31(2): 231 − 238. [44] PEET R K. Forest Vegetation of the colorado front range: patterns of species diversity [J]. Vegetatio, 1978, 37(2): 65 − 78. doi: 10.1007/BF00126830 [45] 郝建锋, 德艺, 李艳, 等. 海拔高度对江油地区杉木人工林群落结构和物种多样性的影响[J]. 西北植物学报, 2014, 34(12): 2544 − 2552. doi: 10.7606/j.issn.1000-4025.2014.12.2544 [46] 谢小魁, 苏东凯, 刘正纲, 等. 长白山原始阔叶红松林径级结构模拟[J]. 生态学杂志, 2010, 29(8): 1477 − 1481. [47] 臧润国, 杨彦承, 蒋有绪. 海南岛霸王岭热带山地雨林群落结构及树种多样性特征的研究[J]. 植物生态学报, 2001, 25(3): 270 − 275. -

 点击查看大图
点击查看大图
图(5) / 表 (5)
计量
- 文章访问数: 2036
- HTML全文浏览量: 946
- PDF下载量: 38
- 被引次数: 0








