

-
抗生素在医疗以及农业生产(如水产养殖)中已被广泛应用 [1-3]。由于滥用抗生素及抗生素的处理不当和去除效率低等原因,造成抗生素及其代谢物随污水排放进入河流、湖泊及海洋等水环境中。目前,从水环境中检测到的抗生素浓度通常为每升水几纳克到几百微克不等[4],环境中残留的抗生素能够诱导抗生素耐药菌(Antibiotic Resistance Bacteria,ARB)和抗生素抗性基因(Antibiotic Resistance Genes,ARGs)的产生[5]。目前在医院废水、养殖废水、天然水体、空气和土壤等环境中均已检测到ARGs[6]。ARGs已成为一种新型环境污染物[7],具有传播性和持久性的特点[8],对生物健康和生态环境产生不利影响[9]。例如,ARGs能够通过水平基因转移(Horizontal Gene Transfer,HGT)促使未接触过抗生素的病原菌获得一种或者多种耐药性,这些具有耐药性的病原菌可能导致人类疾病治疗失效、治疗周期延长和死亡率增加[10-11]。因此,抗生素耐药性(主要是ARB和ARG)已被世界卫生组织纳入21世纪以来严重威胁人类健康的挑战之一[12],相关研究已发展成全球关注的热点问题。
珊瑚礁生态系统是海洋生态系统的重要组成部分,具有极高的初级生产力和生物多样性,也因其较高的生态服务价值而备受关注,但全球气候变化和人类活动已严重威胁到珊瑚礁生态系统的健康状况[13-14],其中,抗生素也已成为珊瑚礁生态系统的潜在威胁因素之一。文献[15-16]的研究结果表明,南海珊瑚礁生态系统中存在多种类型的抗生素(磺胺类、四环素类、喹诺酮类、大环内酯类、氯霉素类等)污染,总浓度最高可达441 ng·L−1。室内实验进一步揭示抗生素能扰乱珊瑚中微生态的平衡,对珊瑚的生长具有负面影响[17]。LIU等[16]也检测到南海北部礁区海水中的ARGs(sul1、sul2、cfr、cmla、floR、qnrA,qnrD和qnrS)污染,总丰度为2.11×104~8.00×106 copies·L−1。由此可见,珊瑚礁生态系统已面临ARGs污染的威胁。此外,人类食用的鱼类中约10%产自珊瑚礁区,珊瑚礁区的潜水观光是滨海旅游区最受欢迎的项目之一,这些均能够增加人类直接或间接感染ARGs的风险。综上,查明珊瑚礁生态系统中ARGs的赋存特征具有重大意义。
我国珊瑚礁生态系统属于印度-太平洋区系,占全球珊瑚礁总面积的2.57%,主要分布在南海诸岛和海南岛周边海域[18]。本研究选取海南岛东海岸8个珊瑚礁生态系统,测定了礁区海水中磺胺类、喹诺酮类和氯霉素类共3类7种ARGs(sul1、sul2、qnrB、qnrS、cmlA1-01,cmlA1-02和cmx(A))的丰度和12种水质参数(盐度、5种营养盐和7种金属离子浓度),旨在阐明海南岛东海岸珊瑚礁生态系统中ARGs的赋存特征,以期揭示人类活动对海南岛近岸生态系统中ARGs分布和传播的影响,并为当地政府制定减少ARGs污染和保护珊瑚礁的对策提供理论依据。
-
于2019年10月选择海南岛东海岸8个礁区(东郊、铜鼓岭、长圮港、龙湾、加井岛、分界洲、后海和鹿回头)作为采样站点(图1),其中,铜鼓岭、分界洲、加井岛、后海和鹿回头是著名的旅游景点;加井岛和分界洲远离海岸,受陆地污染较小;长圮港和东郊靠近河流汇集地,易受到生活污水与农业废水等陆源污染物影响,在长圮港附近还分布着大片养殖高位池[19-20]。
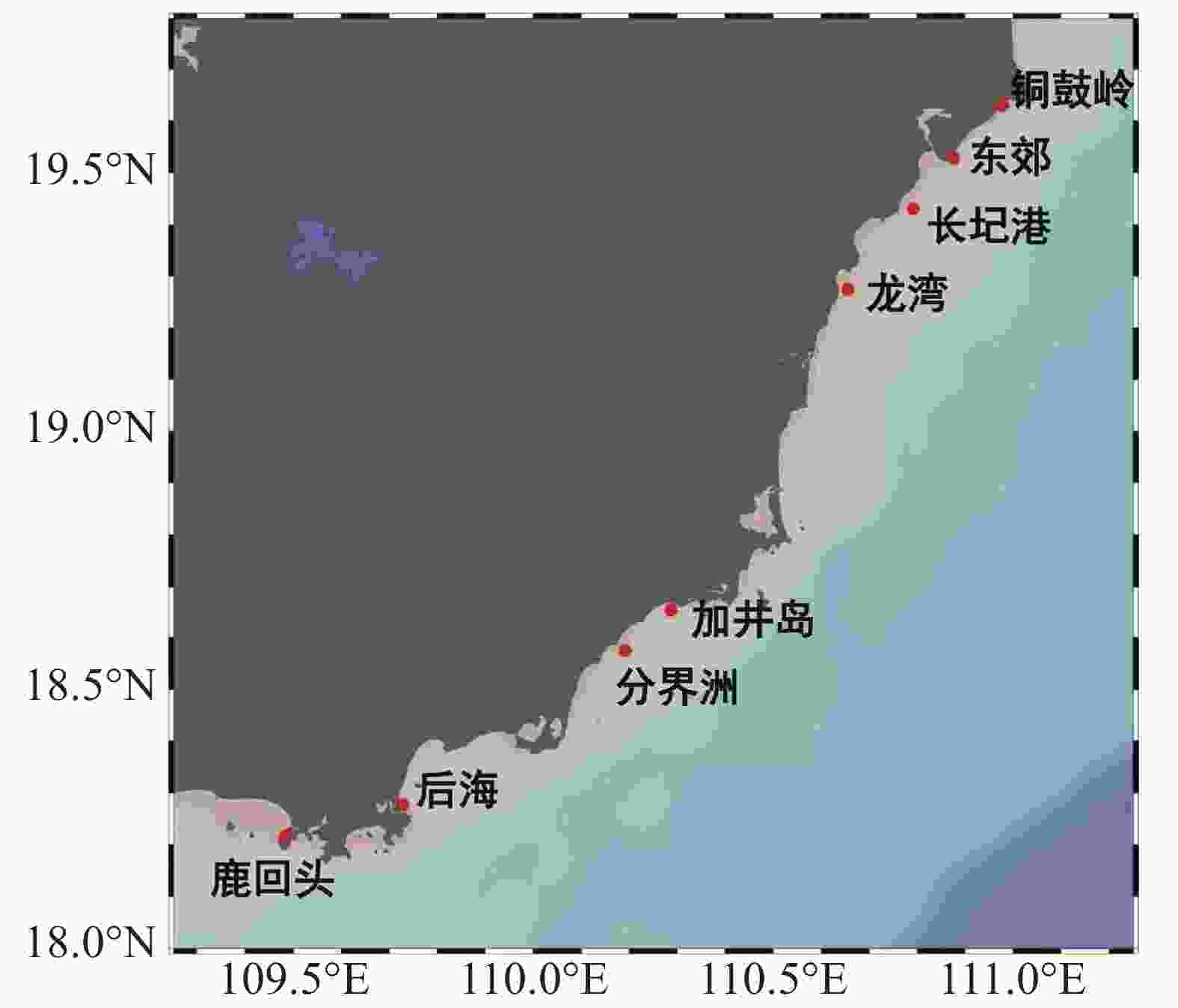
图 1 采样站点
在每个采样站点选择3个不同深度(分别为3、6和9 m),采用有机玻璃采水器(北京普力特仪器有限公司,中国)采集2 L底层海水。立即用0.2 μm聚碳酸酯膜(Merck Millipore公司,美国)过滤1 L海水,然后将滤膜置于冻存管中并置于干冰中保存,待用于后续的DNA提取和ARGs定量。剩余1 L海水的固定和运输方法具体参照《海洋监测规范》(GB 17378.4—2007)[21]。
-
本实验中检测的海水水质指标包括:盐度、NH3−-N、NO2−-N、NO3−-N、PO43−-P、Hg、As、Cu、Pb、Zn,Cd和Cr。上述指标的测定按照《海洋监测规范》(GB 17378.4—2007)和《海洋监测技术规程》(HY/T 147.1—2013)开展[21-22],各检测指标的分析方法详见表1。
表 1 海水样品检测项目分析方法
检测项目 分析方法 分析仪器 检测标准 盐度 盐度计法 盐度计 GB 17378.4—2007(29.1) PO43−-P 磷钼蓝分光光度法 分光光度计 GB 17378.4—2007(39.1) NO2−-N 萘乙二胺分光光度法 分光光度计 GB 17378.4—2007(37) NO3−-N 锌镉还原法 分光光度计 GB/T 12763.4—2007(11) NH3−-N 靛酚蓝分光光度法 分光光度计 GB 17378.4—2007(36.1) Cu 电感耦合等离子体质谱法 电感耦合等离子体质谱仪 HY/T 147.1-2013(5) Pb Zn Cr Cd Hg 原子荧光法 原子荧光光度计 GB 17378.4—2007(5.1) As 原子荧光法 原子荧光光度计 GB 17378.4—2007(11.1) -
在超净工作台中将滤膜剪碎后,采用DNA Clean & Concentrator试剂盒(Zymo Research,美国)提取滤膜上的海水总DNA[23],接着采用Nanodrop微量紫外分光光度计(Maestrogen,中国)和琼脂糖凝胶电泳法测定海水总DNA的浓度、纯度及完整性,将检测合格的DNA样品置于−80 ℃保存备用。
-
扩增ARGs片段,扩增体系:2.5 μL 10×PCR Buffer(Mg2+ plus),2 μL dNTP(2.5 mmol·L−1),1 μL正向引物(10 μmol·L−1),1 μL反向引物(10 μmol·L−1),1 μL DNA模板(所有海水总DNA的混合液),0.15 μL rTaq,17.35 μL DEPC水;扩增条件:94 ℃ 5 min,35个循环(94 ℃ 30 s、退火温度30 s、72 ℃ 1 min),72 ℃ 10 min,最后4 ℃储存;ARGs的引物序列及其退火温度参考LU等[24]的研究,具体参数如表2所示。PCR扩增产物经1%琼脂糖凝胶电泳检测后,采用SanPrep柱式DNA胶回收试剂盒(生工生物工程有限公司,中国)进行目的片段的回收纯化。将目的片段连接到pMD19-T载体[宝生物工程(大连)有限公司,中国]中并转化至Trans5α感受态细胞(全式金生物技术有限公司,中国)。每种转化中挑选至少3个单克隆菌落经PCR鉴定并测序验证。选择验证正确的阳性单克隆菌液过夜培养,按照SanPrep柱式DNA小量抽提试剂盒(生工生物工程有限公司,中国)说明书提取含有ARGs片段的质粒。采用Nanodrop微量紫外分光光度计测定提取质粒的浓度,并根据以下公式计算ARGs的拷贝数[25]。
表 2 引物序列及PCR 反应条件
抗性基因 引物序列 目的基因长度/bp 退火温度/℃ sul1 F:CGCACCGGAAACATCGCTGCAC
R:TGAAGTTCCGCCGCAAGGCTCG162 65 sul2 F:TCCGGTGGAGGCCGGTATCTGG
R:CGGGAATGCCATCTGCCTTGAG190 62 cmlA1-01 F: TAGGAAGCATCGGAACGTTGAT
R: CAGACCGAGCACGACTGTTG101 60 cmlA1-02 F: AGGAAGCATCGGAACGTTGA
R: ACAGACCGAGCACGACTGTTG101 62 qnrB F: CAGATTTYCGCGGCGCAAG
R: TTCCCACAGCTCRCAYTTTTC134 55 qnrS F: GTATAGAGTTCCGTGCGTGTGA
R: GGTTCGTTCCTATCCAGCGATT189 62 cmx(A) F: GCGATCGCCATCCTCTGT
R: TCGACACGGAGCCTTGGT107 60 $$ \begin{split} &\; 拷贝数= 6.02 \times 10^{23} \times (C \times 10^{-9}\div M)\text{,} \end{split}$$ 式中,C代表质粒浓度(ng·μL−1);M=(载体片段碱基对+目的基因碱基对)×660,本研究所用pMD19-T载体的长度为2 692 bp。
-
采用SYBR Green荧光定量PCR方法[16]检测海水DNA中3类7种ARGs的拷贝数,包括磺胺类(sul1,sul2)、喹诺酮类(qnrB,qnrS)和氯霉素类[cmlA1-01,cmlA1-02和cmx(A)]的ARGs [12, 24]。使用经准确定量浓度的质粒制备含梯度浓度ARGs(108~103 copies·μL−1)的标准液DNA,最后采用荧光定量PCR仪(朗基Q2000B型,中国)定量标准液和海水DNA中的ARGs拷贝数,同时为确保反应无污染设定空白对照(以DEPC水代替DNA模板,NTC)。反应体系和反应条件参考厂家说明书进行设置,反应体系:10 μL TB Green Premix Ex Taq Ⅱ(Takara,日本 or 宝生物工程(大连)有限公司,中国),0.8 μL正向引物(10 μmol·L−1),0.8 μL反向引物(10 μmol·L−1),2 μL DNA模板(总量 < 100 ng),6.4 μL DEPC水;反应条件:95 ℃ 30 s,40个循环(95 ℃ 30 s,退火温度30 s),溶解曲线的温度从60 ℃升高至95 ℃。待反应完成后,通过仪器自带的软件判定引物扩增特异性(溶解曲线呈现单峰)。根据Ct值(每个反应管内的荧光信号到达设定的域值时所经历的循环数,Ct值 < 33为阳性)与标准液DNA中ARGs拷贝数的对数值成反比线性关系,绘制标准曲线(R2 > 0.99),并计算待测海水DNA中的ARGs拷贝数。
-
所有数据以平均值±标准差表示,统计分析在IBM SPSS Statistics 20软件中完成。采用Spearman相关性分析探索海水中ARGs的拷贝数和水质参数间的相关性,以P < 0.05为差异。
-
海水盐度、营养盐和金属离子浓度见表3。各采样地点海水中的平均Pb离子含量在1.09~3.21 μg·L−1内,符合Ⅱ类水质设置的范围(1~5 μg·L−1)。此外,鹿回头礁区海水中的平均Zn离子浓度为20.59 μg·L−1,其数值在Ⅱ类水质规定的阈值范围内(20~50 μg·L−1)。在8个珊瑚礁礁区中,铜鼓岭、分界洲岛和加井岛礁区的水质优于其他站点,长圮港礁区中的营养盐含量较高,而东郊、龙湾和鹿回头礁区中部分金属离子浓度较高。
表 3 不同珊瑚礁生态系统中的水质参数
水质参数 取样点 东郊 铜鼓岭 长圮港 龙湾 盐度 33.404 - 33.379 33.467 33.467 33.445 32.63 32.783 32.724 32.562 32.17 32.355 NO2−-N 0.002 0.001 0.001 0.001 0.001 0.001 0.003 0.003 0.001 0.05 0.022 0.01 NO3−-N 0.038 0.052 0.046 0.03 0.027 0.014 0.077 0.078 0.056 0.221 0.104 0.125 NH3−-N 0.011 0.007 0.015 0.001 0.008 0.011 0.009 0.017 0.001 0.003 0.002 0.009 DIN 0.051 0.06 0.062 0.032 0.036 0.026 0.089 0.098 0.058 0.274 0.128 0.144 PO43−-P 0.002 0.004 0.005 0.005 0.005 0.001 0.008 0.008 0.002 0.004 0.002 NA Hg 0.036 0.045 0.05 0.04 0.044 0.04 0.034 0.018 0.039 0.023 0.022 0.043 As 1.2 1.227 2.465 3.075 0.797 0.758 1.898 1.085 0.94 0.852 0.946 2.532 Cu 0.71 0.27 0.39 0.53 0.42 0.65 0.66 0.55 0.47 1.2 2.63 0.79 Pb 4.03 1.63 1.76 0.59 1.12 1.56 1.88 1.01 0.72 1.06 2.01 1.99 Zn 7.1 NA NA 14.7 1.2 1.53 3.36 1.87 2.03 6.2 25.63 NA Cd 0.095 0.105 0.1 0.09 0.095 0.1 0.095 0.095 0.095 0.28 0.94 0.095 Cr 0.65 0.61 0.32 0.63 0.63 0.57 0.56 0.52 0.44 0.75 1.03 0.56 水质参数 取样点 鹿回头 后海 分界洲 加井岛 盐度 32.575 32.644 32.238 32.861 32.828 32.804 33.453 33.429 33.439 33.46 33.491 33.456 NO2−-N 0.002 0.005 0.002 0.001 0.001 0.001 0.001 0.001 0.001 0.001 NA 0.001 NO3−-N 0.056 0.079 0.062 0.048 0.031 0.064 0.057 0.042 0.031 0.037 0.034 0.04 NH3−-N 0.024 0.022 0.019 0.008 0.004 0.003 0.01 0.007 0.008 0.005 0.004 0.011 DIN 0.082 0.106 0.083 0.057 0.036 0.068 0.068 0.05 0.04 0.043 0.038 0.052 PO43−-P 0.004 0.004 0.001 NA NA NA NA 0.001 0.002 NA 0.005 NA Hg 0.026 0.02 0.069 0.014 0.024 0.004 0.033 0.016 0.004 0.023 0.008 0.004 As 0.739 0.727 1.065 0.73 0.723 0.698 1.033 1.949 0.91 1.35 1.061 0.943 Cu 0.35 0.59 0.46 1.02 0.98 2.45 0.5 0.25 0.66 0.56 0.67 0.3 Pb 2.77 1.37 1.83 0.73 2.63 2.71 2.98 3.7 2.95 2.18 2.06 3.68 Zn 31.11 15.21 15.46 16.27 16.74 24.86 1.7 16.61 2.17 2.63 7.94 0.57 Cd 0.095 0.08 0.07 0.08 0.08 0.925 0.115 0.085 0.09 0.105 0.115 0.085 Cr 9.89 0.65 0.44 0.45 0.49 1.24 0.51 0.66 0.48 0.52 0.57 0.46 注:“NA”代表未检出,“-”代表样品丢失。营养盐相关指标单位为mg·L−1,金属离子相关指标单位为μg·L−1。 -
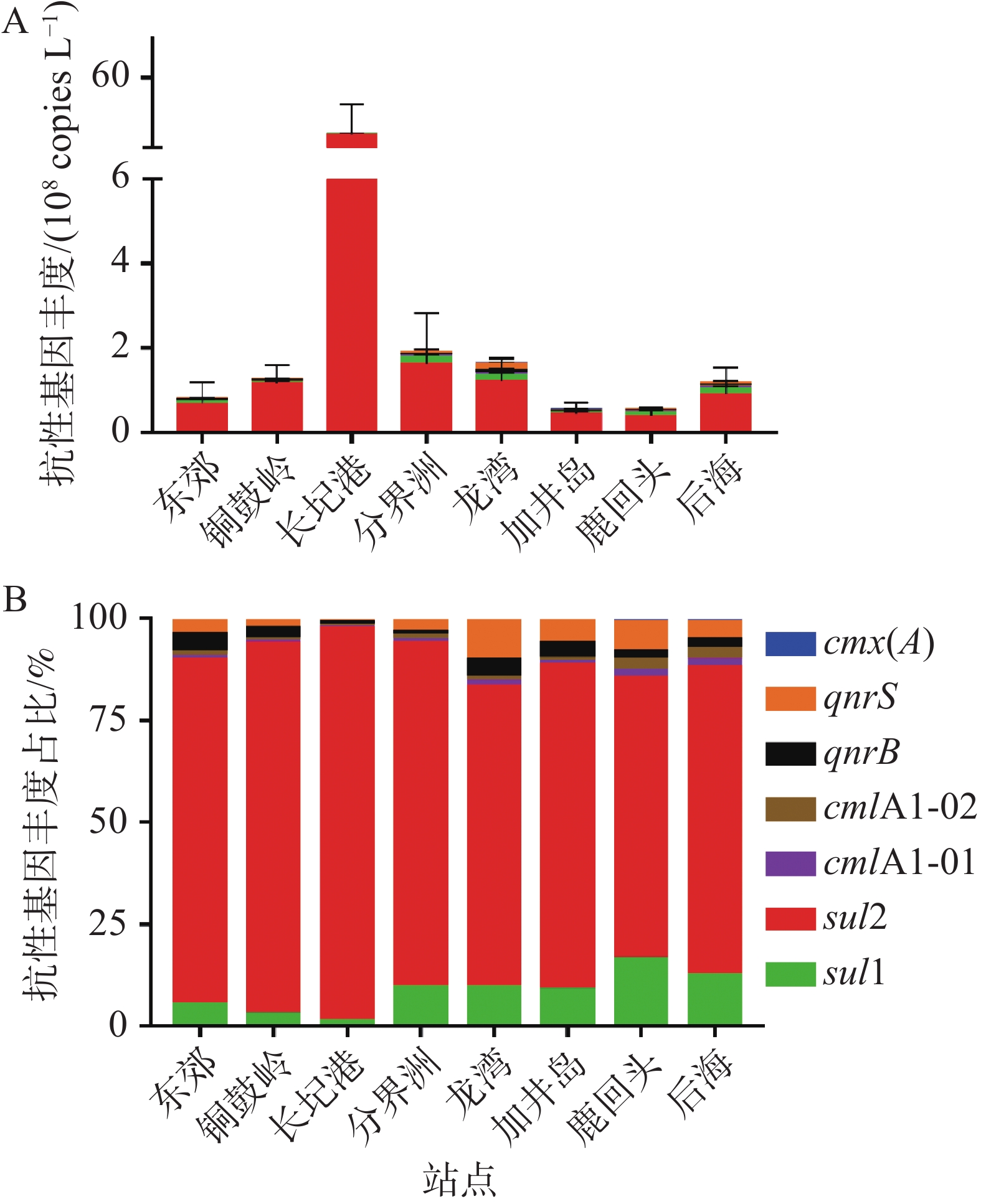
海南岛东海岸珊瑚礁区海水中ARGs的丰度如图2-A所示,在8个取样站点均检测到3类7种ARGs(sul1、sul2、qnrB、qnrS、cmlA1-01,cmlA1-02和cmx(A)),各站点ARGs的总拷贝数为5.51×107~1.71×109 copies·L−1。其中,长圮港具有最高的ARGs总拷贝数(1.71×109 copies·L−1),分界洲次之(1.92×108 copies·L−1),鹿回头(5.74×107 copies·L−1)和加井岛(5.51×107 copies·L−1)具有相对较低的ARGs拷贝数。进一步分析可知,sul2是所有取样站点中占比最高的ARG,拷贝数范围为3.96×107~1.65×109 copies·L−1,占各站点ARGs总拷贝数的68.99%~96.35%。cmx(A)的拷贝数在各站点中均为最低(2.08×105~3.48×106 copies·L−1),低于各站点ARGs总拷贝数的1.21%(图2)。
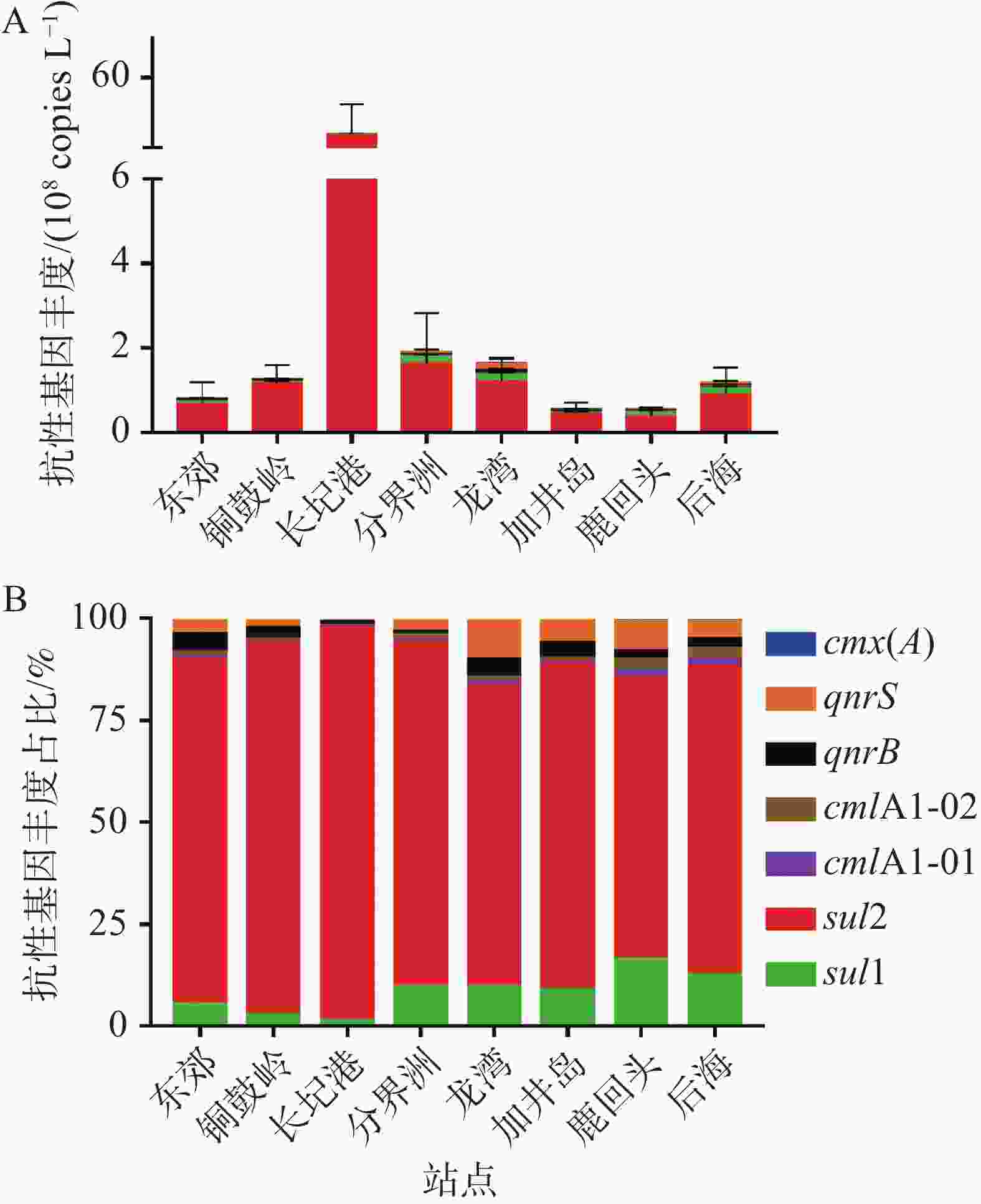
图 2 海南岛东海岸珊瑚礁生态系统中抗生素抗性基因的分布特征
-
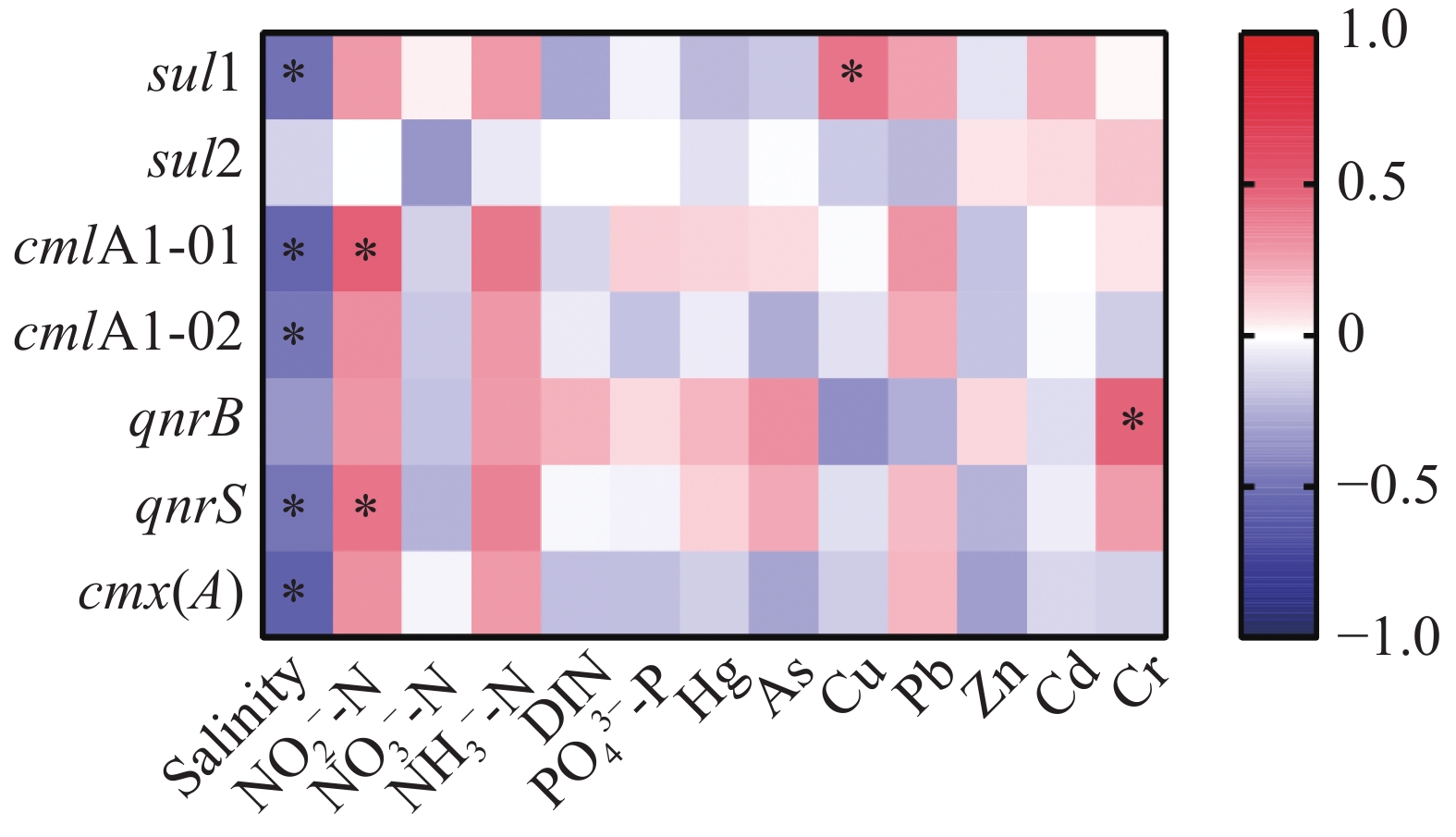
礁区海水中3类7种ARGs的丰度和水质指标的相关性分析如图3所示,sul1、cmlA1-01、cmlA1-02、qnrS和cmx(A)与盐度均呈显著(P < 0.05)负相关性,相关系数(ρ)分别为−0.50、−0.54、−0.47、−0.48和−0.59。cmlA1-01和qnrS与NO2−-N显著(P < 0.05)正相关,ρ分别为0.49和0.41。此外,磺胺类ARG sul1和铜离子浓度显著正相关(ρ=0.41,P < 0.05),喹诺酮类ARG qnrB和镉离子浓度显著正相关(ρ=0.48,P < 0.05)。
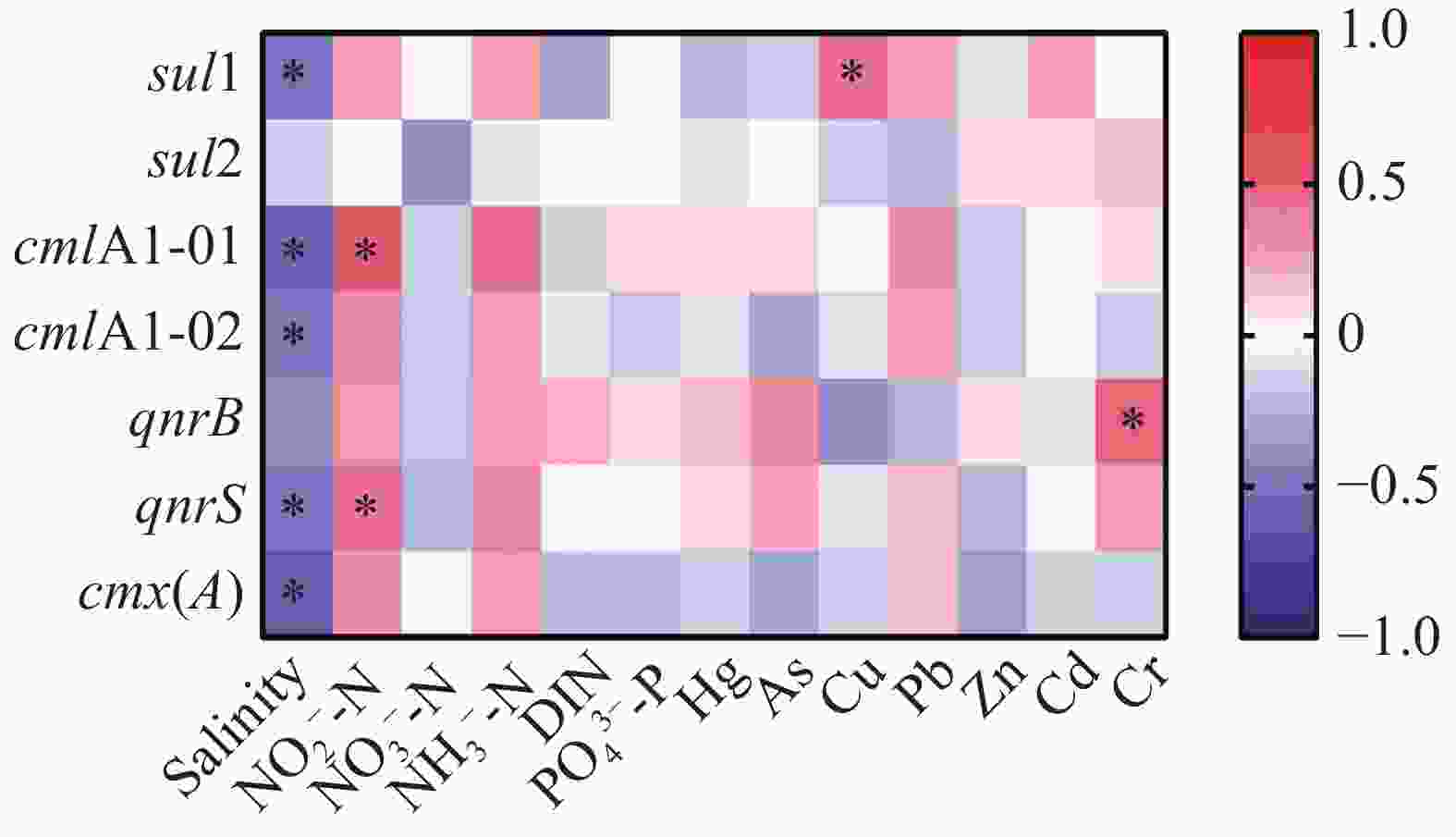
图 3 礁区海水中抗性基因的丰度和水质指标间的相关性分析
-
ARGs对海洋生物健康和生态环境造成的负面影响不容小觑。然而,珊瑚礁生态系统中ARGs污染的相关研究却鲜有报道[16, 26]。本研究调查了海南岛东海岸8个岸礁生态系统中3类7种ARGs的分布,以及ARGs丰度与海水水质指标的联系。
本研究发现磺胺类、喹诺酮类和氯霉素类的ARGs普遍存在于海南岛东海岸珊瑚礁生态系统的海水中。与之类似,在海南东寨港、外伶仃岛、庙湾岛和大鹏湾区域的海水也检测到上述类型的ARGs[16, 27-28]。各研究中检测的ARGs种类存在差异,因此很难比较ARGs的总拷贝数。在本研究中,长圮港中ARGs总拷贝数最高,分界洲次之,而鹿回头和加井岛较低。长圮港的高ARGs丰度可能与其所处区域现存的大量水产养殖业密切相关。集约化养殖导致水产养殖疾病频发,促使抗生素使用增加[29-30],从而造成严重的ARGs污染。加井岛和分界洲岛均离岸较远,加井岛属于无人岛,而分界洲岛是著名的旅游景点,分界洲岛比加井岛居住着更多的居民和游客,人类和动物医学中使用的抗生素通过废水排放到海水中造成ARGs污染[31]。导致两岛ARGs总拷贝数出现差异的主要原因可能是生活污水的排放量不同。鹿回头的珊瑚礁生态系统位于国家级珊瑚礁保护区内,附近酒店和游客数目相对较少,这也许是其低的ARGs总拷贝数的原因。在本研究中,海南岛东海岸珊瑚礁生态系统中存在多种类型的ARGs污染,ARGs的分布可能和水产养殖和生活污水排放等人类活动密切相关。
sul2是海南岛东海岸珊瑚礁海水中占比最高的ARGs,最高丰度为1.65×109 copies·L−1,其最高丰度值与海南岛东寨港海水中的sul2丰度(5.13×108 copies·L−1)相近[27],低于长江下游水体中的sul2丰度(3.30×108 copies·mL−1)[32]。磺胺类抗生素已广泛应用于人类和动物医学,其理化特征(低螯合能力、低结合常数、高水溶性和稳定性)导致在水环境中易产生相应的ARB,进而促使磺胺类ARGs成为水环境中的主导类型[33]。本研究中cmx(A)含量较低的原因可能是这些地区氯霉素抗生素的污染水平较低。总体而言,珊瑚礁海区存在较高丰度的磺胺类ARGs,笔者推测长期高浓度的sul2污染可能会对珊瑚礁生态系统健康产生不利影响。
珊瑚礁生态系统中的ARGs能够通过食物链以及潜水活动等方式进入人体,威胁人类健康[34]。因此,积极探索影响ARGs的丰度及其传播的环境因子显得尤为重要。在本研究中,Spearman相关性分析表明,大部分ARGs(包括sul1、cmlA1-01、cmlA1-02、qnrS和cmx(A))的丰度与海水盐度显著负相关。高盐能够抑制携带ARGs细菌的生长(通过影响细胞结构和生理功能),进而减少环境中ARGs的丰度[35]。同时,低盐度的礁区可能与生活污水或河流注入有关,其中含有人类或动物医学产生的抗生素可能诱导了高丰度的ARB和ARGs。BERGERON等[36]对沼泽样品的研究表明,ARGs的丰度在低盐条件下(6 PSU)多于在高盐(12 PSU)条件下的。LIU等[16]和阮孙兰[25]的研究也表明部分ARGs的丰度和盐度显著负相关。室内研究也表明高盐能够导致水体中ARGs丰度降低约50%[37]。此外,本研究揭示部分ARGs拷贝数与NO2−-N,Cu和Cr离子浓度显著正相关。ARGs丰度的变化可能和水体营养盐成分的变化相关,富营养化的水体有利于ARB的生长,进而促进ARGs的传播[4]。然而,阮孙兰[25]的研究表明亚硝酸盐和大多数目标抗性基因成负相关,造成该差异的具体机理还需要进一步研究。作为抗生素的替代品,金属化合物(例如CuSO4)被广泛应用于饲料添加剂中,用以预防疾病和促进生物生长[37]。目前,已有多项研究表明重金属对于抗生素抗性基因的分布和传播具有重要作用[38]。金属化合物能够在环境中普遍并持续存在,金属抗性基因通常与ARGs位于同一可移动遗传元件(Mobile Genetic Element,MGE)上,促使重金属在ARGs增殖和传播中起关键作用[39]。HU等[40]的研究也表明Cu离子浓度增加能够促使环境中形成高丰度的ARGs。WARDWELL等[41]研究表明,泥沼样品中Hg含量不仅能够增强细菌对Hg的抗性,也能够增加细菌的多重抗药性。综上所述,多种环境压力的共同选择,尤其是盐度、营养盐和金属离子浓度的选择及其诱导的细菌群落变化是ARGs在礁区海水中传播的主要驱动因素。污水处理厂可通过严格控制这些环境因子,来减少礁区中抗性基因的输入量。
本研究结果表明,海南岛东海岸珊瑚礁生态系统中存在ARGs污染,且水产养殖附近的礁区海水中ARGs污染程度最高。此外,ARGs的分布和海水盐度、营养盐和金属离子浓度含量显著相关。因此,通过调整环境中的盐度、营养盐和金属离子浓度可影响礁区ARGs的传播。
Occurrence and characteristics of antibiotic resistance genes in the coral reef ecosystems along the east coast of Hainan Island
-
摘要: 为了解海南岛近岸珊瑚礁生态系统中抗生素抗性基因(Antibiotic Resistance Genes,ARGs)的赋存特征,本研究借助绝对定量PCR技术分析了海南岛东海岸8个珊瑚礁礁区海水中的磺胺类(sul1、sul2)、喹诺酮类(qnrB、qnrS)和氯霉素类ARGs(cmlA1-01、cmlA1-02、cmx(A))的丰度及其与海水盐度、营养盐(NH3−-N、NO2−-N、NO3−-N、PO43−-P)和金属离子(Hg、As、Cu、Pb、Zn、Cd、Cr)浓度的相关性。结果发现,在8个调查站点海水中的磺胺类、喹诺酮类和氯霉素类共3类7种ARGs的检出率为100%,且sul2在所有调查站点中都是丰度最高的ARG(3.96×107~1.65×109 copies·L−1)。此外,5种ARGs[sul1、qnrS、cmlA1-01、cmlA1-02、cmx(A)]的丰度与盐度显著负相关,而qnrS和cmlA1-01的丰度均与NO2−-N浓度显著正相关,sul1和qnrB的丰度分别与Cu和Cr浓度显著正相关。结果表明,海南岛东海岸珊瑚礁生态系统中存在多种类型的ARGs污染,盐度、营养盐和金属离子可能是驱动礁区ARGs传播的主要环境因素。Abstract: To understand the occurrence and characteristics of antibiotic resistance genes (ARGs) in the coastal coral reef ecosystem of Hainan Island, a survey was made in the seawater of the coral reef ecosystems along the east coast of Hainan Island, and the sea water at different depths was sampled at 8 survey sites to analyze its abundances of ARGs of sulfonamides (sul1 and sul2), quinolones (qnrB and qnrS), and chloramphenicol (cmlA1-01, cmlA1-02, and cmx(A)), and its correlation between ARGs abundances and environmental conditions including salinity, nutrient (NH3−-N, NO2−-N, NO3−-N, and PO43−-P), and metal (Hg, As, Cu, Pb, Zn, Cd, and Cr) ion content by using real-time PCR-based absolute quantification. The results showed that the detection rates of 7 ARGs in the 3 types of sulfonamide, quinolone and chloramphenicol in the seawater at the 8 survey sites were 100%, and that sul2 (3.96×107−1.65×109 copies·L−1) was the most abundant in the seawater in all the coral reef ecosystems. Moreover, the absolute abundances of five ARGs (sul1, qnrS, cmlA1-01, cmlA1-02, and cmx(A)) were negatively correlated with salinity. Meanwhile, the absolute abundances of both cmlA1-01 and qnrS were positively correlated with the NO2−-N content, and sul1 and qnrB were positively correlated with Cu and Cr, respectively. These results suggest that there should be many types of ARG pollutions in the coral reef ecosystems along the east coast of Hainan Island. Salinity, nutrients and metal ions may be the main environmental factors driving the spread of ARGs in the coral reef area.
-
Key words:
- coral reef /
- environmental pollution /
- antibiotic resistance gene /
- nutrients /
- metal ion
-
表 1 海水样品检测项目分析方法
检测项目 分析方法 分析仪器 检测标准 盐度 盐度计法 盐度计 GB 17378.4—2007(29.1) PO43−-P 磷钼蓝分光光度法 分光光度计 GB 17378.4—2007(39.1) NO2−-N 萘乙二胺分光光度法 分光光度计 GB 17378.4—2007(37) NO3−-N 锌镉还原法 分光光度计 GB/T 12763.4—2007(11) NH3−-N 靛酚蓝分光光度法 分光光度计 GB 17378.4—2007(36.1) Cu 电感耦合等离子体质谱法 电感耦合等离子体质谱仪 HY/T 147.1-2013(5) Pb Zn Cr Cd Hg 原子荧光法 原子荧光光度计 GB 17378.4—2007(5.1) As 原子荧光法 原子荧光光度计 GB 17378.4—2007(11.1)  下载: 导出CSV
下载: 导出CSV
表 2 引物序列及PCR 反应条件
抗性基因 引物序列 目的基因长度/bp 退火温度/℃ sul1 F:CGCACCGGAAACATCGCTGCAC
R:TGAAGTTCCGCCGCAAGGCTCG162 65 sul2 F:TCCGGTGGAGGCCGGTATCTGG
R:CGGGAATGCCATCTGCCTTGAG190 62 cmlA1-01 F: TAGGAAGCATCGGAACGTTGAT
R: CAGACCGAGCACGACTGTTG101 60 cmlA1-02 F: AGGAAGCATCGGAACGTTGA
R: ACAGACCGAGCACGACTGTTG101 62 qnrB F: CAGATTTYCGCGGCGCAAG
R: TTCCCACAGCTCRCAYTTTTC134 55 qnrS F: GTATAGAGTTCCGTGCGTGTGA
R: GGTTCGTTCCTATCCAGCGATT189 62 cmx(A) F: GCGATCGCCATCCTCTGT
R: TCGACACGGAGCCTTGGT107 60
下载: 导出CSV
表 3 不同珊瑚礁生态系统中的水质参数
水质参数 取样点 东郊 铜鼓岭 长圮港 龙湾 盐度 33.404 - 33.379 33.467 33.467 33.445 32.63 32.783 32.724 32.562 32.17 32.355 NO2−-N 0.002 0.001 0.001 0.001 0.001 0.001 0.003 0.003 0.001 0.05 0.022 0.01 NO3−-N 0.038 0.052 0.046 0.03 0.027 0.014 0.077 0.078 0.056 0.221 0.104 0.125 NH3−-N 0.011 0.007 0.015 0.001 0.008 0.011 0.009 0.017 0.001 0.003 0.002 0.009 DIN 0.051 0.06 0.062 0.032 0.036 0.026 0.089 0.098 0.058 0.274 0.128 0.144 PO43−-P 0.002 0.004 0.005 0.005 0.005 0.001 0.008 0.008 0.002 0.004 0.002 NA Hg 0.036 0.045 0.05 0.04 0.044 0.04 0.034 0.018 0.039 0.023 0.022 0.043 As 1.2 1.227 2.465 3.075 0.797 0.758 1.898 1.085 0.94 0.852 0.946 2.532 Cu 0.71 0.27 0.39 0.53 0.42 0.65 0.66 0.55 0.47 1.2 2.63 0.79 Pb 4.03 1.63 1.76 0.59 1.12 1.56 1.88 1.01 0.72 1.06 2.01 1.99 Zn 7.1 NA NA 14.7 1.2 1.53 3.36 1.87 2.03 6.2 25.63 NA Cd 0.095 0.105 0.1 0.09 0.095 0.1 0.095 0.095 0.095 0.28 0.94 0.095 Cr 0.65 0.61 0.32 0.63 0.63 0.57 0.56 0.52 0.44 0.75 1.03 0.56 水质参数 取样点 鹿回头 后海 分界洲 加井岛 盐度 32.575 32.644 32.238 32.861 32.828 32.804 33.453 33.429 33.439 33.46 33.491 33.456 NO2−-N 0.002 0.005 0.002 0.001 0.001 0.001 0.001 0.001 0.001 0.001 NA 0.001 NO3−-N 0.056 0.079 0.062 0.048 0.031 0.064 0.057 0.042 0.031 0.037 0.034 0.04 NH3−-N 0.024 0.022 0.019 0.008 0.004 0.003 0.01 0.007 0.008 0.005 0.004 0.011 DIN 0.082 0.106 0.083 0.057 0.036 0.068 0.068 0.05 0.04 0.043 0.038 0.052 PO43−-P 0.004 0.004 0.001 NA NA NA NA 0.001 0.002 NA 0.005 NA Hg 0.026 0.02 0.069 0.014 0.024 0.004 0.033 0.016 0.004 0.023 0.008 0.004 As 0.739 0.727 1.065 0.73 0.723 0.698 1.033 1.949 0.91 1.35 1.061 0.943 Cu 0.35 0.59 0.46 1.02 0.98 2.45 0.5 0.25 0.66 0.56 0.67 0.3 Pb 2.77 1.37 1.83 0.73 2.63 2.71 2.98 3.7 2.95 2.18 2.06 3.68 Zn 31.11 15.21 15.46 16.27 16.74 24.86 1.7 16.61 2.17 2.63 7.94 0.57 Cd 0.095 0.08 0.07 0.08 0.08 0.925 0.115 0.085 0.09 0.105 0.115 0.085 Cr 9.89 0.65 0.44 0.45 0.49 1.24 0.51 0.66 0.48 0.52 0.57 0.46 注:“NA”代表未检出,“-”代表样品丢失。营养盐相关指标单位为mg·L−1,金属离子相关指标单位为μg·L−1。
下载: 导出CSV
-
[1] YANG Y, SONG W, LIN H, et al. Antibiotics and antibiotic resistance genes in global lakes: A review and meta-analysis [J]. Environment International, 2018, 116: 60 − 73. doi: 10.1016/j.envint.2018.04.011 [2] 叶必雄, 张岚. 环境水体及饮用水中抗生素污染现状及健康影响分析[J]. 环境与健康杂志, 2015, 32(2): 173 − 178. [3] 朱光平, 吴南翔, 范宏亮. 环境中抗生素抗性基因的去除方法[J]. 环境与职业医学, 2019, 36(12): 1168 − 1174. [4] WANG Z, HAN M, LI E, et al. Distribution of antibiotic resistance genes in an agriculturally disturbed lake in China: Their links with microbial communities, antibiotics, and water quality [J]. Journal of Hazardous Materials, 2020, 393: 122426. doi: 10.1016/j.jhazmat.2020.122426 [5] SHARMA V K, JOHNSON N, CIZMAS L, et al. A review of the influence of treatment strategies on antibiotic resistant bacteria and antibiotic resistance genes [J]. Chemosphere, 2016, 150: 702 − 714. doi: 10.1016/j.chemosphere.2015.12.084 [6] DONG H, CHEN Y, WANG J, et al. Interactions of microplastics and antibiotic resistance genes and their effects on the aquaculture environments [J]. Journal of Hazardous Materials, 2021, 403: 123961. doi: 10.1016/j.jhazmat.2020.123961 [7] PRUDEN A, PEI R, STORTEBOOM H, et al. Antibiotic resistance genes as emerging contaminants: studies in northern Colorado [J]. Environmental Science & Technology, 2006, 40(23): 7445 − 7450. [8] MAO D, YU S, RYSZ M, et al. Prevalence and proliferation of antibiotic resistance genes in two municipal wastewater treatment plants [J]. Water Research, 2015, 85: 458 − 466. doi: 10.1016/j.watres.2015.09.010 [9] LIU X, LU S, GUO W, et al. Antibiotics in the aquatic environments: a review of lakes, China [J]. Science of the Total Environment, 2018, 627: 1195 − 1208. doi: 10.1016/j.scitotenv.2018.01.271 [10] BUENO I, WSILLIANM-NGUYEN J, HWANG H, et al. Impact of point sources on antibiotic resistance genes in the natural environment: A systematic review of the evidence [J]. Animal Health Research Reviews, 2017, 18(2): 112 − 127. doi: 10.1017/S146625231700007X [11] WANG G, ZHOU S, HAN X, et al. Occurrence, distribution, and source track of antibiotics and antibiotic resistance genes in the main rivers of Chongqing city, southwest China [J]. Journal of Hazardous Materials, 2020, 389: 122110. doi: 10.1016/j.jhazmat.2020.122110 [12] ZHANG Y, LU J, WU J, et al. Potential risks of microplastics combined with superbugs: Enrichment of antibiotic resistant bacteria on the surface of microplastics in mariculture system [J]. Ecotoxicology and Environmental Safety, 2020, 187: 109852. doi: 10.1016/j.ecoenv.2019.109852 [13] 赵美霞, 余克服, 张乔民. 珊瑚礁区的生物多样性及其生态功能[J]. 生态学报, 2006, 26(1): 186 − 194. doi: 10.3321/j.issn:1000-0933.2006.01.025 [14] 黄荣永, 余克服, 王英辉, 等. 珊瑚礁遥感研究进展[J]. 遥感学报, 2019, 23(6): 1091 − 1112. [15] ZHANG R, ZHANG R, YU K, et al. Occurrence, sources and transport of antibiotics in the surface water of coral reef regions in the South China Sea: Potential risk to coral growth [J]. Environmental Pollution, 2018, 232: 450 − 457. doi: 10.1016/j.envpol.2017.09.064 [16] LIU S, SU H, PAN Y F, et al. Spatial and seasonal variations of antibiotics and antibiotic resistance genes and ecological risks in the coral reef regions adjacent to two typical islands in South China Sea [J]. Marine Pollution Bulletin, 2020, 158: 111424. doi: 10.1016/j.marpolbul.2020.111424 [17] SWEET M J, CROQUER A, BYTHELL J C. Dynamics of bacterial community development in the reef coral Acropora muricata following experimental antibiotic treatment [J]. Coral Reefs, 2011, 30(4): 1121. doi: 10.1007/s00338-011-0800-0 [18] 周国伟, 黄晖, 喻子牛, 等. 造礁石珊瑚与其共生藻(Symbiodinium)共生研究进展[J]. 生态学报, 2009, 29(8): 4397 − 4407. [19] 吴钟解, 吴瑞, 王道儒, 等. 海南岛东、南部珊瑚礁生态健康状况初步分析[J]. 热带作物学报, 2011, 32(1): 122 − 130. doi: 10.3969/j.issn.1000-2561.2011.01.025 [20] 朱志雄, 马坤, 方彰胜, 等. 海南省麒麟菜自然保护区海草资源分布及保护建议[J]. 广东农业科学, 2017, 44(4): 90 − 98. [21] 国家质量监督检验检疫总局. 国家标准化管理委员会. GB 17378.4—2007. 海洋监测规范第4部分: 海水分析 [s]. 北京: 中国标准出版社, 2007. [22] 国家海洋局. HY/T 147.1-2013. 海洋监测技术规程第1部分: 海水 [s]. 北京: 国家海洋局, 2013. [23] YUAN J, LI M, LIN S. An improved DNA extraction method for efficient and quantitative recovery of phytoplankton diversity in natural assemblages [J]. PLoS One, 2015, 10(7): e0133060. doi: 10.1371/journal.pone.0133060 [24] LU J, ZHANG Y, WU J, et al. Effects of microplastics on distribution of antibiotic resistance genes in recirculating aquaculture system [J]. Ecotoxicology and Environmental Safety, 2019, 184: 109631. doi: 10.1016/j.ecoenv.2019.109631 [25] 阮孙兰. 海南东寨港红树林典型区域抗生素抗性基因时空分布特征及影响因素研究 [D]. 海口: 海南大学, 2018. [26] SU H, HU X, XU W, et al. Diversity, abundances and distribution of antibiotic resistance genes and virulence factors in the South China Sea revealed by metagenomic sequencing [J]. Science of The Total Environment, 2022, 814: 152803. doi: 10.1016/j.scitotenv.2021.152803 [27] 姜春霞, 黎平, 李森楠, 等. 海南东寨港海水和沉积物中抗生素抗性基因污染特征研究[J]. 生态环境学报, 2019, 28(1): 128 − 135. [28] LI W, SU H, CAO Y, et al. Antibiotic resistance genes and bacterial community dynamics in the seawater environment of Dapeng Cove, South China [J]. Science of the Total Environment, 2020, 723: 138027. doi: 10.1016/j.scitotenv.2020.138027 [29] 王熙涛, 卢燕丹, 王丽丽, 等. 新型抗生素替代品防治水产动物细菌性疾病的研究进展[J]. 饲料与畜牧, 2015(4): 18 − 22. [30] 吕可, 赵前程, 刘婧懿, 等. 噬菌体在水生动物病害防治中的应用问题和解决策略[J]. 水产科学, 2020, 39(6): 964 − 971. [31] 苏建强, 黄福义, 朱永官. 环境抗生素抗性基因研究进展[J]. 生物多样性, 2013, 21(4): 481 − 487. [32] ZHANG G, LU S, WANG Y, et al. Occurrence of antibiotics and antibiotic resistance genes and their correlations in lower Yangtze River, China [J]. Environmental Pollution, 2020, 257: 113365. doi: 10.1016/j.envpol.2019.113365 [33] SUZUKI S, OGO M, MILLER T W, et al. Who possesses drug resistance genes in the aquatic environment?: Sulfamethoxazole (SMX) resistance genes among the bacterial community in water environment of Metro-Manila, Philippines [J]. Frontiers in Microbiology, 2013, 4: 102. [34] DANG C, XIA Y, ZHENG M, et al. Metagenomic insights into the profile of antibiotic resistomes in a large drinking water reservoir [J]. Environment International, 2020, 136: 105449. doi: 10.1016/j.envint.2019.105449 [35] LIU M, LI Q, SUN H, et al. Impact of salinity on antibiotic resistance genes in wastewater treatment bioreactors [J]. Chemical Engineering Journal, 2018, 338: 557 − 563. doi: 10.1016/j.cej.2018.01.066 [36] BERGERON S, BROWN R, HOMER J, et al. Presence of antibiotic resistance genes in different salinity gradients of freshwater to saltwater marshes in southeast Louisiana, USA [J]. International Biodeterioration & Biodegradation, 2016, 113: 80 − 87. [37] SEILER C, BERENDONK T U. Heavy metal driven co-selection of antibiotic resistance in soil and water bodies impacted by agriculture and aquaculture [J]. Frontiers in Microbiology, 2012, 3: 399. [38] 张俊亚, 魏源送, 陈梅雪, 等. 畜禽粪便生物处理与土地利用全过程中抗生素和重金属抗性基因的赋存与转归特征研究进展[J]. 环境科学学报, 2015, 35(4): 935 − 946. [39] HU H W, WANG J T, LI J, et al. Long-term nickel contamination increases the occurrence of antibiotic resistance genes in agricultural soils [J]. Environmental Science & Technology, 2017, 51(2): 790 − 800. [40] HU H W, WANG J T, LI J, et al. Field-based evidence for copper contamination induced changes of antibiotic resistance in agricultural soils [J]. Environmental Microbiology, 2016, 18(11): 3896 − 3909. doi: 10.1111/1462-2920.13370 [41] WARDWELL L H, JUDE B A, MOODY J P, et al. Co-selection of mercury and antibiotic resistance in sphagnum core samples dating back 2000 years [J]. Geomicrobiology Journal, 2009, 26(5): 351 − 360. doi: 10.1080/01490450902889072 -

 点击查看大图
点击查看大图
图(3) / 表 (3)
计量
- 文章访问数: 704
- HTML全文浏览量: 255
- PDF下载量: 35
- 被引次数: 0

