

-
红外相机监测作为一种非损伤性的野生动物监测方法[1],具有昼夜连续工作、人力投入少、对研究对象干扰小等优势[2-3],目前已被广泛应用于动物生态学的研究[4]。通过分析红外相机所拍摄的动物图像数据,可以获得物种丰富度、空间分布、活动节律和种群密度等信息[5-6],能一定程度弥补传统调查方法的不足,尤其是针对一些夜间活动的、隐蔽的和珍稀物种的调查更为有效[7]。我国最早在云南省高黎贡山地区和台湾地区使用红外相机技术监测野生动物[8-9]。经过20多年的发展,红外相机技术已发展成为地栖大中型兽类和鸟类的重要常规监测技术[10-11]。但该技术应用于高度树栖的林冠层动物多样性的研究相对较少,且布设相机台数少及安装高度较低,仅5~15 m[12],在林冠层中使用红外相机调查树栖哺乳动物的潜在效果在很大程度上仍是未知的。
海南长臂猿(Nomascus hainanus)为长臂猿科(Hylobatidae)、黑冠长臂猿属(Nomascus),是我国一级重点保护野生动物。由于在历史上受栖息地破坏及狩猎等因素导致海南长臂猿种群数量极低[13],2007年10月保护国际(CI)宣布海南长臂猿名列全球25种濒危灵长类物种之首[14]。我国高度重视该物种的保护,基于自然恢复方法的科学保护使得海南长臂猿成为全球20种长臂猿中唯一处于数量增长的类群[15],种群数量从20世纪80年代2群7~9只增至2022年的5群36只[16],目前仅分布在海南热带雨林国家公园霸王岭片区斧头岭及南崩岭区域[17],属于高度树栖的昼行性小型猿类[18-19],以成熟果实、嫩叶、嫩芽为食[20-21]。
海南长臂猿作为海南热带雨林国家公园野生动植物保护的标志性物种和生态指示种[22],对于该物种的保护和研究有助于提升热带雨林生态系统原真性和完整性。目前对海南长臂猿的研究集中在种群现状、食性和栖息地等方面[23-24],缺少海南长臂猿同域分布区伴生鸟兽多样性组成及季节性变化相关研究。本研究拟使用红外相机技术分析海南长臂猿林冠层同域分布物种的多样性、年活动格局以及海拔分布格局,探讨海南长臂猿与其同域分布鸟兽物种的共存机制,为未来基于生物多样性的海南长臂猿栖息地修复和生态廊道建设提供参考。
-
海南热带雨林国家公园霸王岭片区(以下简称为霸王岭片区)地处海南昌江县和白沙县境内(19º02′~19º08′N, 109º02′~109º13′E),总面积约850 km2,海拔75~1 655 m,年平均温度21.9 ℃,年平均相对湿度84.2%,年均降水量1 657 mm,属热带季风气候,旱季雨季分明,5—10月为雨季,11月至次年4月为旱季,4—8月常有台风;其核心保护区内覆盖有热带低地雨林、热带山地雨林、热带山地常绿阔叶林和热带山顶矮林[13]。国家一级重点保护动物有海南长臂猿、海南山鹧鸪(Arborophila ardens)、大灵猫(Viverra zibetha)、海南孔雀雉(Polyplectron katsumatae)等8种,国家二级重点保护动物有水鹿(Rusa unicolor)、白鹇(Lophura nycthemera)、蟒蛇(Python bivittatus)、猕猴(Macaca mulatta tcheliensis)和巨松鼠 (Ratufa bicolor)等50种[25]。
-
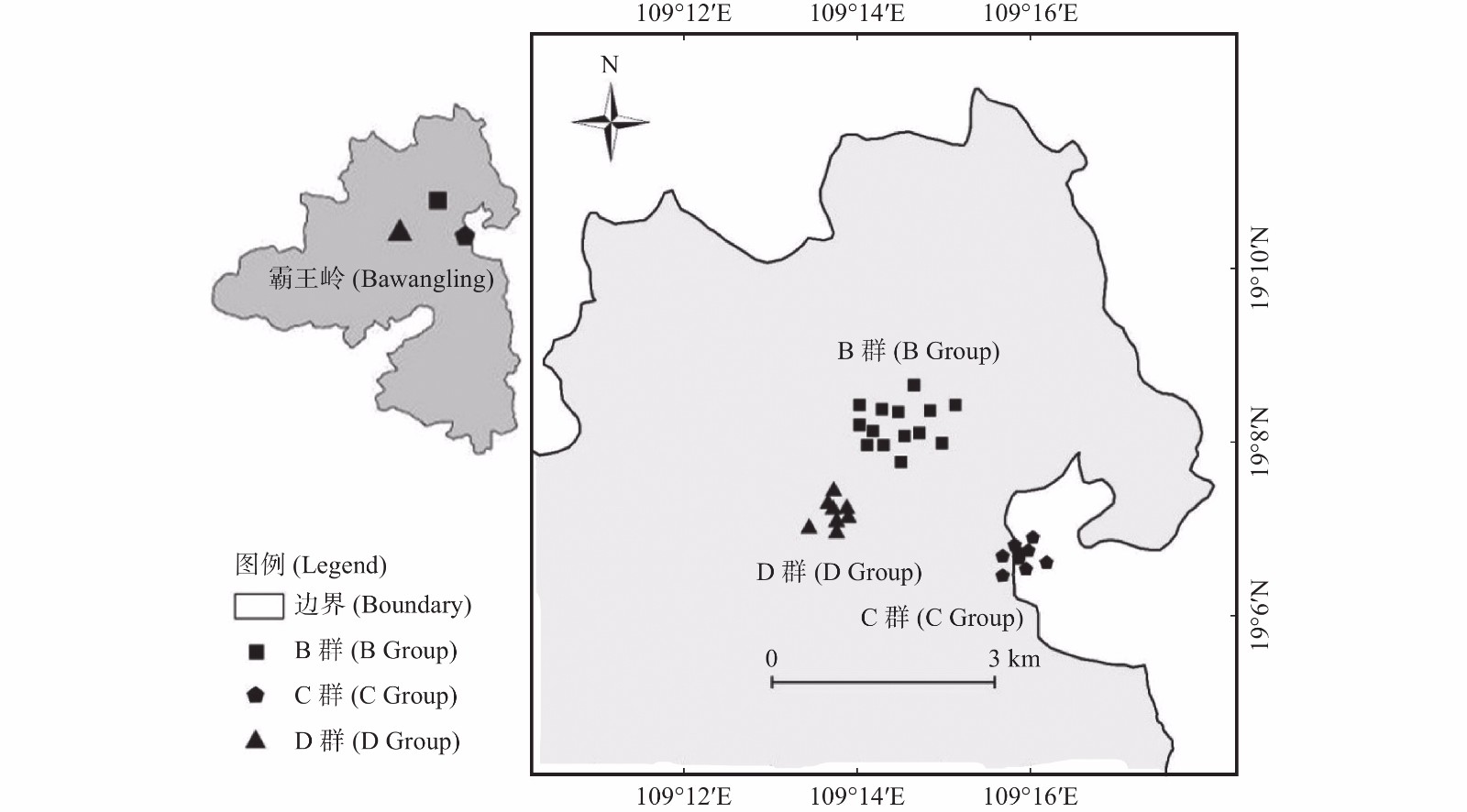
根据海南长臂猿各个家庭群分布现状、海拔分布和猿食植物分布,选择5个家庭群中的B、C、D群分布区作为研究区域,面积约15 km2。B、C、D群分布区主要为热带山地雨林,其他林型分布较少。2019年1月开始,布设33个红外相机监测位点66台红外相机(图1),红外相机安放主要考虑长臂猿实际利用区域(护林员记录的长臂猿活动强度及猿食植物分布),兼顾较高和较低海拔(600~800 m热带低地雨林布设16台,800~1 100 m热带山地雨林布设40台,1 100 m热带苔藓林布设10台)。将相机安装在离地13~25 m的树冠层,选取海南长臂猿可能经过或者附近有猿食植物且视野开阔的树干进行安放,避免树枝遮挡镜头;镜头与地面呈5°~15°俯视角,避免阳光直射。每个监测位点安装的2台相机机身相靠、镜头相反,各自反方向拍照,相邻位点之间的直线距离在100~500 m [26-27]。拍摄模式设置为连拍3张照片同时录制视频10 s,拍摄间隔为0 s,灵敏度为中。安装前进行性能测试,安装时记录每个位点相机的编号、GPS位点、海拔和生境因子等信息。
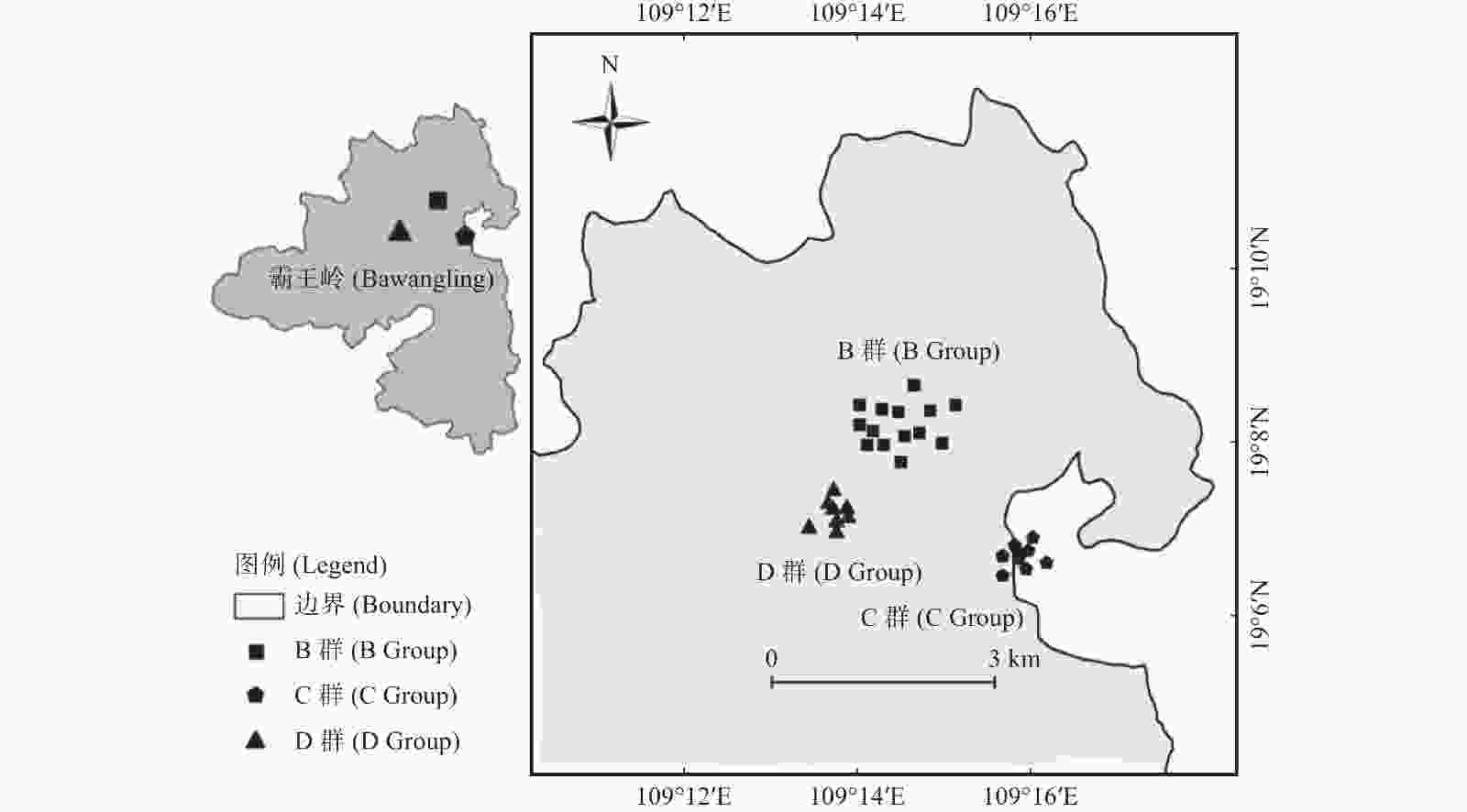
图 1 海南长臂猿分布区林冠层红外相机监测点分布图
2019年8月,完成第一次红外相机的数据回收(替换SD卡)和维护(更换电池),同时发现有11台红外相机长时间受到雨水冲刷从而彻底损坏,最终实际正常工作的红外相机为55台(海拔600~900 m 14台,海拔900~1 100 m 31台,海拔1 100 m以上10台)。2019年11月和2020年6月完成第二次和第三次数据回收,未有相机损坏。熟悉海南本土物种的专家人工对照片和视频进行物种识别,剔除模糊不清或没有明显物种特征的照片,所拍摄到照片和视频中的物种鉴定主要参考《中国鸟类分类与分布名录》和《中国哺乳动物多样性》[28-29]。
-
利用相对丰富度指数衡量海南长臂猿分布区林冠层兽类和鸟类各物种相对种群数量。独立有效照片是指同一相机位点含同种个体的相邻有效照片( 或视频)间隔时间至少为 30 min[30],1个相机工作日以单台红外相机在野外持续工作24 h为标准,每台红外相机有效工作日以该位点拍摄到的第一张野外工作照片(显示有正确设置信息)和最后一张野外工作照片的日期间隔为有效工作日。将拍摄到各种鸟类和兽类物种的独立有效照片与所拍摄物种独立有效照片总数的百分比作为每个物种的相对丰富度(relative abundance index, RAI) [31]。
-
依据统计所得数据,计算月相对丰富度指数(monthly relative abundance index, MRAI),分析独立有效照片较多的动物年活动规律。
-
根据海南长臂猿栖息地类型和相机实际安装情况将研究区域分为3个海拔梯度(600~800、 800~1 100、 1 100 m以上),并统计不同海拔梯度的独立有效照片,采用Shannon-Wiener指数、Pielou均匀度指数和Simpson指数判断不同海拔梯度的α多样性水平[32]。
Shannon-Wiener指数(H')是将物种丰富度与种的多度结合起来的指数,通常来说H'值越大,物种多样性越丰富。由于每个海拔梯度布设的红外相机台数的不同,在比较不同海拔梯度的物种数和多样性指数时,以每一台相机作为独立样本,计算每一台相机的物种数、多样性指数和均匀度指数,通过比较其平均值表示不同海拔梯度的物种多样性情况[33]。采用Kruskall-Wallis test来比较不同海拔梯度物种多样性的差异,差异显著水平设定为0.05。所有数据都在SPSS 24.0和Excel 2021上进行处理。
Pielou均匀度指数(J)反映物种分布的均匀程度,能够表征分类单元中每个物种个体间的差异。均匀度(J)数值越大,群落中物种个体间的差异越小。
物种优势度(C)采用Simpson指数计算,C值越大说明群落内物种数量分布越不均匀,优势种地位越突出[34]。
-
2019-01-20—2020-06-30期间,55台红外相机累计监测到16 266个有效相机工作日,共拍摄到兽类、鸟类照片和视频14 136份,鉴定出独立有效照片1 747张, 其中兽类1 569张(89.8%)、鸟类178张(10.2%)。记录到3目4科10种兽类,6目11科20种鸟类。拍摄到海南长臂猿(21个)、海南鼯鼠(Petaurista hainana)(16个)、红颊长吻松鼠(Dremomys rufigenis)(16个)和果子狸(Paguma larvata)(15个)的位点数居于前4位,其他兽类和鸟类较少(表1)。
表 1 海南长臂猿同域分布区林冠层兽类与鸟类动物名录
物种 保护等级 IUCN
红色名录独立有效
照片数相对丰富度
指数拍摄到该物种的
相机位点数兽类 灵长目 Carnivora
长臂猿科 Viverridae
海南长臂猿 Nomascus hainanus
猴科 Cercopithecidae
猕猴 Macaca mulatta tcheliensis
一级
二级
CR
CR
917
24
58.44
1.53
21(20)
4(0)食肉目 Carnivora
灵猫科 Viverridae
果子狸 Paguma larvata
椰子狸 Paradoxurus hermaphroditus
小灵猫 Viverricula indica
二级
一级
LC
LC
LC
33
45
1
2.10
2.87
0.06
15(10)
6(3)
1(1)啮齿目 Rodentia
松鼠科 Sciuridae
赤腹松鼠 Callosciurus erythraeus
巨松鼠 Ratufa bicolor
海南鼯鼠 Petaurista hainana
倭松鼠 Tamiops maritimus
红颊长吻松鼠 Dremomys rufigenis
二级
LC
NT
LC
15
24
279
6
225
0.96
1.53
17.78
0.38
14.34
4(2)
5(4)
16(15)
3(3)
16(19)鸟类 Birds 雀形目 Passeriformes
鸦科 Corvidae
白翅蓝鹊 Urocissa xanthomelana
黄胸绿鹊 Cissa hypoleuca
灰树鹊 Dendrocitta formosae
鹎科 Pycnonotidae
栗背短脚鹎 Hemixos castanonotus
鹃鵙科 Passeriformes
赤红山椒鸟 Pericrocotus flammeus
雀鹛科 Alcippeidae
灰眶雀鹛 Alcippe morrisonia
二级
NT
LC
LC
LC
LC
LC
4
8
2
2
1
7
2.25
4.50
1.12
1.12
0.56
3.96
3
5
2
2
1
1啄木鸟目 Capitonidae
啄木鸟科 Picidae
大黄冠啄木鸟 Chrysophlegma flavinucha
拟啄木鸟科 Capitonidae
黑眉拟啄木鸟 Psilopogon fabe二级
NT
LC
3
4
1.69
2.25
3
4鸮形目 Strigiformes
鸱鸮科 Strigidae
斑头鸺鹠 Glaucidium cuculoides
草鸮 Tyto longimembris
黄嘴角鸮 Otus spilocephalus
二级
LC
LC
LC
12
3
1
6.74
1.70
0.56
1
2
1鹰形目 Accipitriformes
鹰科 Accipitridae
鹰雕 Nisaetus nipalensis
二级
NT
11
6.18
4咬鹃目Trogoniformes
咬鹃科 Trogonidae
红头咬鹃 Harpactes erythrocephalus
二级
LC
1
0.56
1鸽形目 Columbiformes
鸠鸽科 Columbidae
厚嘴绿鸠 Treron curvirostra
山皇鸠 Ducula badia
绿翅金鸠 Chalcophaps indica
二级
二级
LC
LC
LC
36
80
2
20.23
44.92
1.12
5
5
2佛法僧目 Coraciiformes
蜂虎科 Meropidae
蓝须夜蜂虎 Nyctyornis athertoni
二级
LC
1
0.56
1注:CR:极危;NT:近危;LC:无危。( )表示该物种与海南长臂猿出现在同一相机的台数 。 -
记录到的10种兽类中,国家一级重点保护动物2种,即海南长臂猿和小灵猫(Viverricula indica);国家二级重点保护野生动物3种,分别是猕猴、巨松鼠和椰子狸(Paradoxurus hermaphroditus),国家一、二级重点保护动物合计占所有兽类物种数的50%。海南长臂猿被IUCN红色名录列为极危物种(CR),巨松鼠被列为近危物种(NT)。此外,兽类中相对丰富度指数居于前三的是海南长臂猿(RAI =58.44)、海南鼯鼠(RAI =17.78)和红颊长吻松鼠(RAI =14.34),其他兽类的相对丰富度指数均低于3,小灵猫仅有1张独立有效照片(表1)。
记录到的17种鸟类中,国家二级重点保护鸟类8种,分别是黄胸绿鹊(Cissa hypoleuca)、大黄冠啄木鸟(Chrysophlegma flavinucha)、斑头鸺鹠(Glaucidium cuculoides)、鹰雕(Nisaetus nipalensis)、红头咬鹃(Harpactes erythrocephalus)、厚嘴绿鸠(Treron curvirostra)、山皇鸠(Ducula badia)、和蓝须蜂虎(Nyctyornis athertoni),占所有鸟类物种数的53%。记录到的17种鸟类中,3种为近危物种(NT),分别是白翅蓝鹊(Urocissa xanthomelana)、大黄冠啄木鸟和鹰雕。鸟类中相对丰富度指数居于前三的是山皇鸠(RAI=44.92)、厚嘴绿鸠(RAI=20.23)和斑头鸺鹠(RAI=6.74);赤红山椒鸟(Pericrocotus flammeus)、红头咬鹃、黄嘴角鸮(Otus spilocephalus)和蓝须夜蜂虎的相对丰富度指数较低(表1)。
-
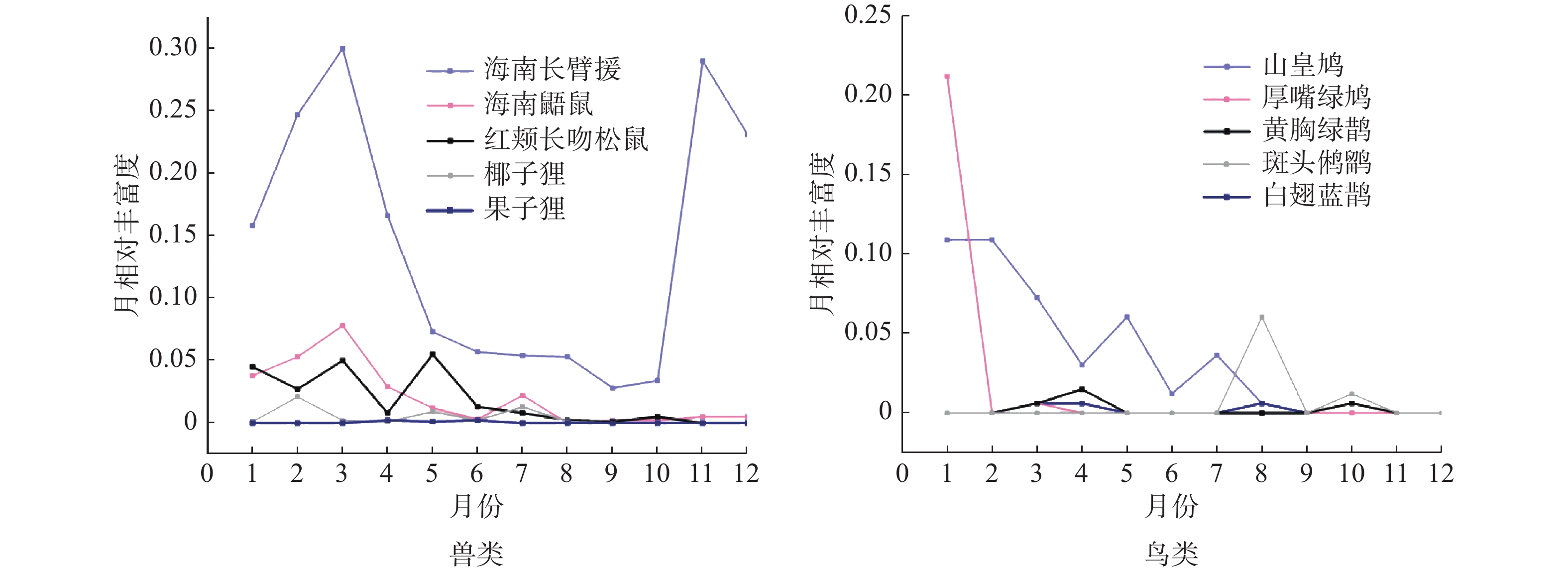
兽类各月份活动强度较高,鸟类则较低,在11、12月份未拍摄到鸟类照片和视频。海南长臂猿和海南鼯鼠年活动格局相似,且与季节(旱季和雨季)显著相关(海南长臂猿: χ2 = 8.308, df = 1, P = 0.002; 海南鼯鼠: χ2 = 3.124, df = 1, P = 0.047)。海南长臂猿在9月份活动强度最低,10月份快速上升,3月份达到全年活动峰值,旱季则较低。海南鼯鼠在3月份出现活动峰值,在5—10月份趋于平缓。其他兽类如红颊长吻松鼠、椰子狸、倭松鼠(Tamiops maritimus)的年活动格局均与季节无显著相关,在各个月份的活动强度较为平缓。5种鸟类的年活动格局均与季节无显著相关,其中山皇鸠和厚嘴绿鸠在1月份出现活动峰值,斑头鸺鹠和白翅蓝鹊在8月份出现峰值,黄胸绿鹊在4月份出现峰值,其他月份鸟类月相对丰富度较低,主要集中在1—4月份和7—8月份活动(图2)。
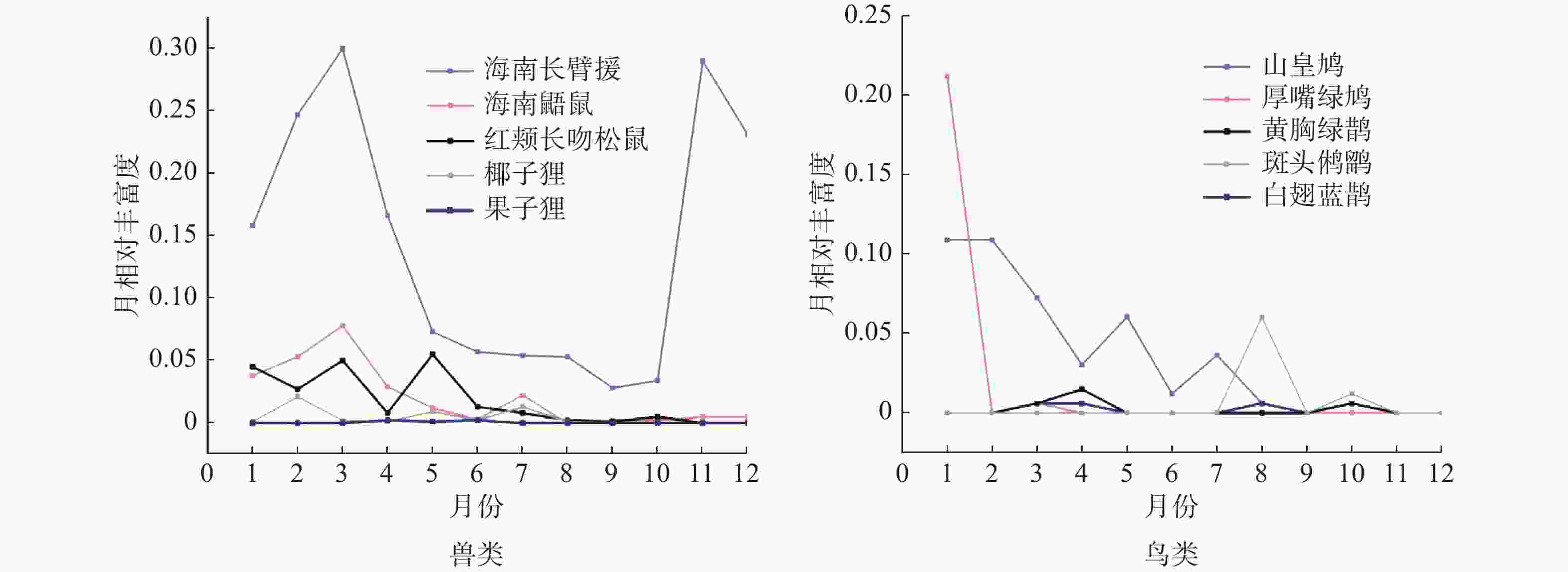
图 2 兽类和鸟类的年活动格局
-
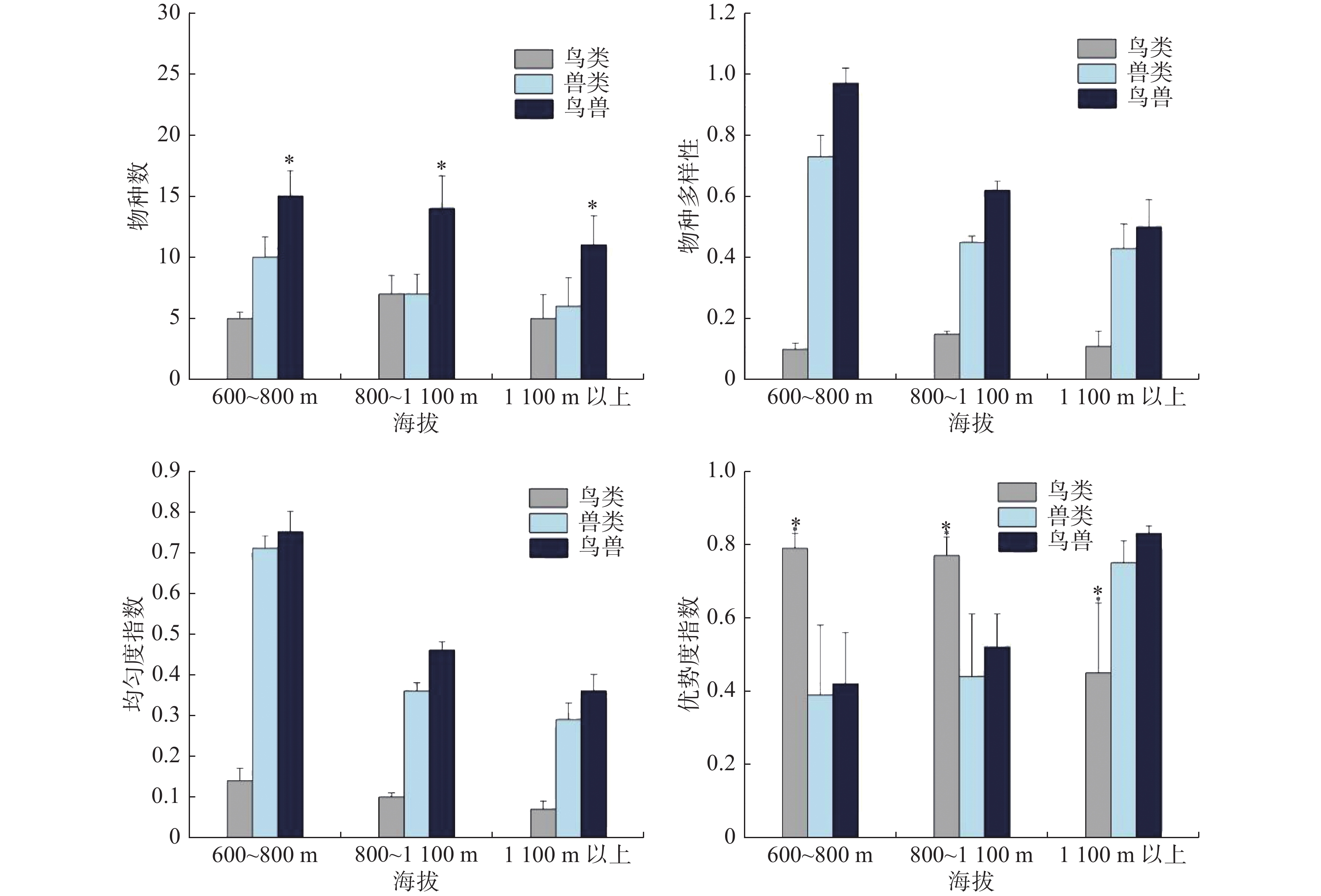
不同海拔梯度下鸟类和兽类物种数均不存在显著差异(鸟类: χ2 = 2.085, df = 2, P =0.339;兽类: χ2 = 5.53, df = 2, P = 0.061);鸟兽物种总数则存在显著差异(鸟兽: χ2 = 6.399, df = 2, P = 0.038),主要表现为,海拔1 100 m以上的鸟兽类物种总数低于其他海拔梯度(图3)。比较不同海拔梯度的物种多样性指数、均匀度指数以及优势度指数,只发现鸟类的优势度指数存在显著性差异性(鸟类: χ2 = 18.585, df = 2, P = 0.0001),主要表现为,鸟类优势度指数在海拔1 100 m以上明显低于其他海拔梯度;兽类和鸟兽的多样性指数和均匀度指数随着海拔的升高呈单调递减;兽类中海南鼯鼠和红颊长吻松鼠是优势种,优势度指数分别是0.13和0.10;鸟类中山皇鸠和厚嘴绿鸠是优势物种,优势度指数分别是0.40和0.20(图3)。
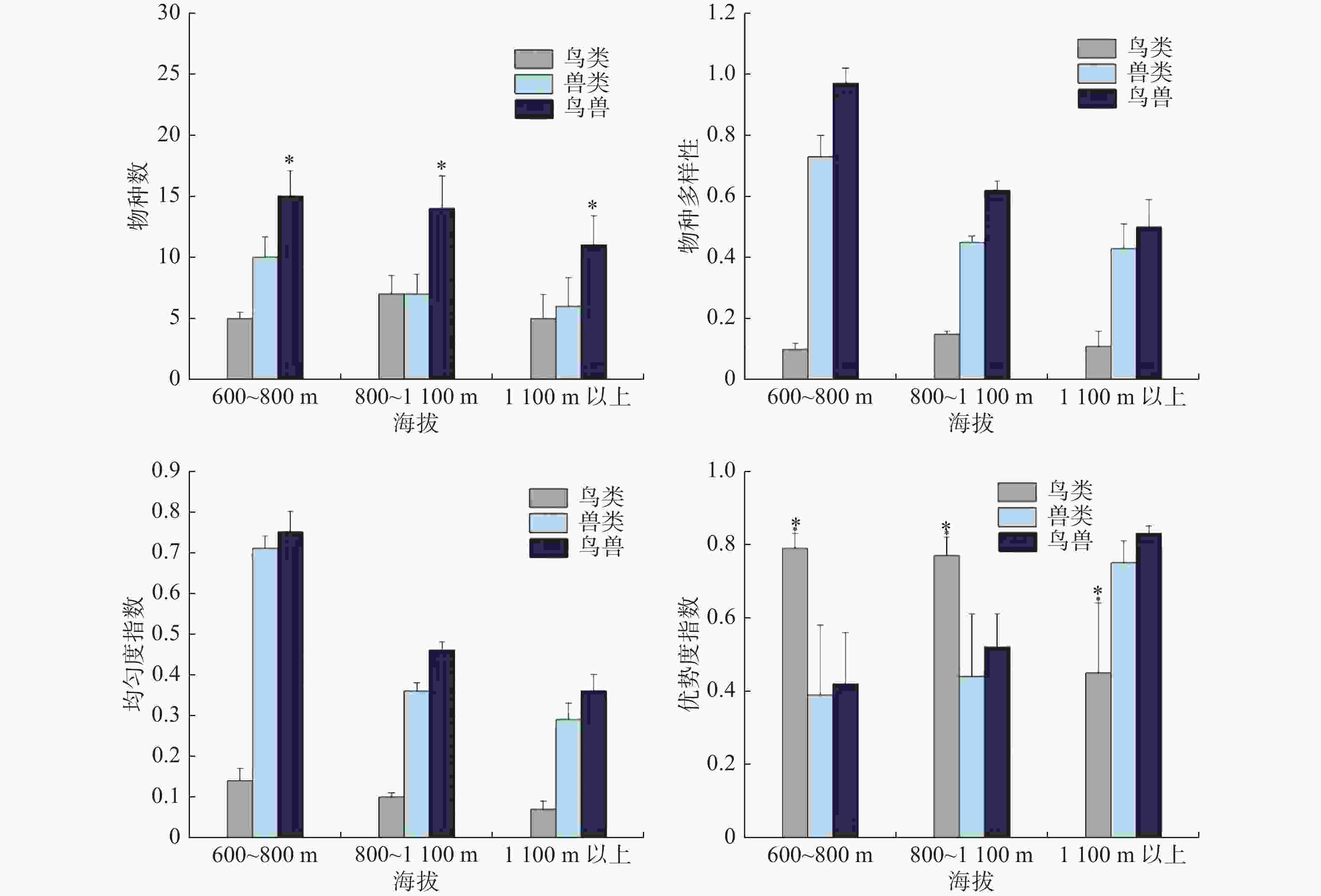
图 3 不同海拔梯度下鸟类和兽类的物种数和多样性分布差异
-
密切相关的同域物种会采用导致生态位分离或资源划分的过程,从而使物种共存成为可能[35-36]。本研究监测到与海南长臂猿同域分布的兽类包括巨松鼠、赤腹松鼠、海南鼯鼠,果子狸和椰子狸等,尽管这些兽类与海南长臂猿的食物生态位存在重叠,但在活动模式和空间利用的差异使它们可以共存[37-38]。在活动模式方面,海南长臂猿是昼型性灵长类,椰子狸、果子狸和海南鼯鼠则是典型的夜行性动物[39],这些物种时间生态位的高度分离有利于物种共存[35]。在空间利用方面,海南长臂猿是高度树栖动物,几乎不会下到地面寻找食物,而其他物种均有地面活动现象,如赤腹松鼠会在果实资源丰富的季节下到地面层搜寻食物[40]。此外,研究区域还发现另外一种灵长类猕猴,虽然该物种与海南长臂猿的食物、时间生态位高度重叠,但海南长臂猿出现位点并未发现猕猴,海南长臂猿在与猕猴的生态位竞争中可能处于优势。海南长臂猿缺少自然天敌,但少数大型猛禽,如蛇雕(Spilornis cheela)等可能会攻击长臂猿幼崽,本研究共记录到2种猛禽,即鹰雕和草鸮(Tyto longimembris),红外相机监测和人为跟踪均未发现对海南长臂猿幼崽的伤害行为。
-
物种丰富度的海拔分布格局可分为4种常见的格局:递减型、低高原型、低高原中峰型和中峰型[41],本研究兽类物种数虽然在海拔梯度间不存在显著性差异,但整体上表现为随着海拔的升高呈单调递减趋势,随着海拔的升高,温度、植被类型、食物资源等会呈现出一定的垂直分布和变化,进而对动物分布产生重要影响[42]。该研究区域海拔600~800 m、800~1 100 m、1 100 m 以上分别对应的是热带沟谷雨林、热带山地雨林和山地苔藓林。在山地苔藓林区域树木稀疏且缺少高大乔木,会引发食物资源短缺,物种间竞争变大,亦或不能为某些树栖动物提供隐蔽的栖息场所,如海南长臂猿会选择胸径更大,高度更高、冠幅更大、枝下更高的乔木夜宿[43]。鸟类的物种数指数先升高后减少,是典型的中峰模式,随海拔上升气温下降,导致鸟类在海拔1 100 m 以上的分布种数、分布数量逐渐下降,这可能是高海拔区域鸟类物种多样性指数较低的原因[33];分布区内低海拔区域较强的人为干扰,可能影响了鸟类的分布[34]。
海南长臂猿以及其他兽类年活动格局表明,海南长臂猿和海南鼯鼠的月活动强度有季节性变化,旱季月活动强度较高,雨季较低,这与海南长臂猿分布区的植被类型和气候有关。在雨季,霸王岭片区雨水充沛,大部分植物生长茂盛且处于结果期,故食物资源丰富且分布均匀;旱季降雨量偏低,植物果实减少引起海南长臂猿和海南鼯鼠的食物短缺。海南长臂猿等因移动需要付出一定的能量,可能也需要增加对叶芽类取食的策略或增加食物的摄入来弥补所消耗的能量[24],活动强度往往会因食物资源的时空变化而存在季节性差异[44]。雨季结束,旱季到来食物资源减少时,布设在果树附近的红外相机拍摄到的视频显示海南长臂猿和海南鼯鼠取果、采食画面的滞留时间变长,也表明旱季的月活动强度比雨季高。其他兽类在雨季和旱季活动强度相对稳定,与食性有关,如饮食灵活的猕猴可以通过改变其食料偏好来避免与其他类群的种间竞争[45]。鸟类的生殖期具有明显的季节性,生活在我国的鸟类一般在3~8月份生殖[46],研究区域的鸟类活动规律与此相符,活动峰值主要集中在1— 4月份和7—8月份。在繁殖期间,雌雄个体相遇的概率大幅提升,从而提高了月丰富度指数。然而,在1—2月份山皇鸠和厚嘴绿鸠的月丰富度指数高,这可能与该地区的温度较高有关。
-
传统的调查方法倾向于关注昼行性的野生动物,红外相机技术可以监测到动物夜间活动,从而实现对目标物种的24 h适时监测[47]。在监测活跃的、体型较大的树栖物种,如海南长臂猿等也特别高效,尽管该物种实际数量很少,但拍摄到最多独立有效照片,而且发现动物不会回避红外相机,在自然条件下各种行为可以详细地、近距离地记录下来[2]。此外,虽然树栖红外相机高度暴露在雨水和阳光下,但此次林冠层红外相机的故障率17%低于其他地方报道的热带雨林红外相机的故障率70%[48]。本研究结果表明,在树冠层中使用红外相机调查树栖哺乳动物效果与陆地相似,是调查林冠内隐蔽、夜间活动和活跃动物群落的有效方法。国内应用树栖红外相机调查林冠层野生动物的研究极少,可以在未来的调查工作中尝试定义和标准化安装规范,如树栖红外相机安装的高度和树种的选择以及考虑到热带雨林较高的生物多样性,在1个布设位点安装多个红外相机等。
致谢:海南热带雨林国家公园霸王岭片区对研究予以大力支持,护林员张志诚、李文永、谢赠南等对野外工作的顺利开展提供了不可或缺的帮助;陈少莲和杨川在鸟类鉴别方面给予许多帮助,谨此致谢。
Diversity of companion mammals and birds based on infrared camera trapping in the forest canopy of Hainan gibbon range
-
摘要: 为了探究海南长臂猿( Nomascus hainanus )同域分布的兽类和鸟类的多样性、年活动格局以及海拔分布格局, 2019年1月至2020年6月,在海南长臂猿分布区林冠层布设红外相机共记录到3目4科10种兽类,其中,国家一级重点保护动物2种,相对丰富度居于兽类前3位分别为海南长臂猿(58.44)、海南鼯鼠(Petaurista hainana)(17.78)和红颊长吻松鼠(Dremomys rufigenis)(14.34)。红外相机共记录到6目11科20种鸟类,其中,属于国家级保护动物10种,相对丰富度较高的鸟类有山皇鸠(Ducula badia)(44.92)、厚嘴绿鸠(Treron curvirostra)(20.23)和班头鸺鹠(Glaucidium cuculoides)(6.74)。海南长臂猿和海南鼯鼠的月相对丰富度表现出相似性,旱季活动较为频繁,雨季减弱;其他兽类和鸟类的月相对丰富度则与季节无显著相关。兽类多样性随着海拔升高呈单调递减趋势,在海拔600 ~ 800 m物种多样性水平最高;鸟类多样性指数随海拔增加而先增加后减小。本研究揭示了海南长臂猿同域分布区林冠层伴生鸟兽组成,为海南长臂猿栖息地修复及生态廊道建设提供了基于动物多样性的重要资料。Abstract: Infrared camera technology is increasingly used in wildlife diversity surveys and long-term monitoring, mainly focusing on the research of ground-dwelling birds and mammals, but less on highly arboreal endangered species, such as Hainan gibbons and associated species diversity. From January 2019 to June 2020, 66 infrared cameras were deployed in the forest canopy of the Hainan gibbon sympatric distribution area to explore the diversity of mammals and bird, the seasonality of activity intensity and changes with altitude gradients. A total of 10 species of mammals that belonged to 3 orders and 4 families were recorded, of which 2 species were under national Class II protection. The three mammals with the highest relative abundance were Hainan gibbon (Nomascus hainanus) (RAI=58.44), Petaurista hainana (RAI=17.78) and Dremomys rufigenis (RAI=14.34). There were 20 species of birds belonging to 6 orders and 11 families recorded, of which 10 species were listed under national Class II protection and three of them, Ducula badia, Treron curvirostra and Glaucidium cuculoides, were higher in relative abundance index (44.92, 20.23 and 6.74, respectively). Hainan gibbons and Petaurista hainana showed similar monthly relative abundance, with their activities being high in the dry season and low in the wet season; while other mammals and birds were not significantly correlated with seasons. The diversity of mammals showed a monotonical decrease with the increase of altitude, and had the highest level of species diversity in the range of 600 ~ 800 m asl. The diversity index of birds first increased and then decreased with the increase of altitude. This study revealed for the first time the composition of companion birds and mammals in the same area where Hainan gibbons were distributed, providing a reference for habitat restoration and ecological corridor construction of Hainan gibbons based on animal diversity in the Bawangling area of Hainan Tropical Rainforest National Park.
-
Key words:
- infrared camera /
- forest canopy /
- species diversity /
- Nomascus hainanus /
- sympatry
-
表 1 海南长臂猿同域分布区林冠层兽类与鸟类动物名录
物种 保护等级 IUCN
红色名录独立有效
照片数相对丰富度
指数拍摄到该物种的
相机位点数兽类 灵长目 Carnivora
长臂猿科 Viverridae
海南长臂猿 Nomascus hainanus
猴科 Cercopithecidae
猕猴 Macaca mulatta tcheliensis
一级
二级
CR
CR
917
24
58.44
1.53
21(20)
4(0)食肉目 Carnivora
灵猫科 Viverridae
果子狸 Paguma larvata
椰子狸 Paradoxurus hermaphroditus
小灵猫 Viverricula indica
二级
一级
LC
LC
LC
33
45
1
2.10
2.87
0.06
15(10)
6(3)
1(1)啮齿目 Rodentia
松鼠科 Sciuridae
赤腹松鼠 Callosciurus erythraeus
巨松鼠 Ratufa bicolor
海南鼯鼠 Petaurista hainana
倭松鼠 Tamiops maritimus
红颊长吻松鼠 Dremomys rufigenis
二级
LC
NT
LC
15
24
279
6
225
0.96
1.53
17.78
0.38
14.34
4(2)
5(4)
16(15)
3(3)
16(19)鸟类 Birds 雀形目 Passeriformes
鸦科 Corvidae
白翅蓝鹊 Urocissa xanthomelana
黄胸绿鹊 Cissa hypoleuca
灰树鹊 Dendrocitta formosae
鹎科 Pycnonotidae
栗背短脚鹎 Hemixos castanonotus
鹃鵙科 Passeriformes
赤红山椒鸟 Pericrocotus flammeus
雀鹛科 Alcippeidae
灰眶雀鹛 Alcippe morrisonia
二级
NT
LC
LC
LC
LC
LC
4
8
2
2
1
7
2.25
4.50
1.12
1.12
0.56
3.96
3
5
2
2
1
1啄木鸟目 Capitonidae
啄木鸟科 Picidae
大黄冠啄木鸟 Chrysophlegma flavinucha
拟啄木鸟科 Capitonidae
黑眉拟啄木鸟 Psilopogon fabe二级
NT
LC
3
4
1.69
2.25
3
4鸮形目 Strigiformes
鸱鸮科 Strigidae
斑头鸺鹠 Glaucidium cuculoides
草鸮 Tyto longimembris
黄嘴角鸮 Otus spilocephalus
二级
LC
LC
LC
12
3
1
6.74
1.70
0.56
1
2
1鹰形目 Accipitriformes
鹰科 Accipitridae
鹰雕 Nisaetus nipalensis
二级
NT
11
6.18
4咬鹃目Trogoniformes
咬鹃科 Trogonidae
红头咬鹃 Harpactes erythrocephalus
二级
LC
1
0.56
1鸽形目 Columbiformes
鸠鸽科 Columbidae
厚嘴绿鸠 Treron curvirostra
山皇鸠 Ducula badia
绿翅金鸠 Chalcophaps indica
二级
二级
LC
LC
LC
36
80
2
20.23
44.92
1.12
5
5
2佛法僧目 Coraciiformes
蜂虎科 Meropidae
蓝须夜蜂虎 Nyctyornis athertoni
二级
LC
1
0.56
1注:CR:极危;NT:近危;LC:无危。( )表示该物种与海南长臂猿出现在同一相机的台数 。  下载: 导出CSV
下载: 导出CSV
-
[1] KAYS R W, SLAUSON K M. Remote cameras [J]. Noninvasive Survey Methods for Carnivores, 2008, 32(3): 110 − 140. [2] NICHOLS J D, KARANTH K U. Camera Traps in Animal Ecology: Methods and Analyses [M]. New York: Springer, 2011. [3] 李晟, 王大军, 肖治术, 等. 红外相机技术在我国野生动物研究与保护中的应用与前景[J]. 生物多样性, 2014, 22(6): 685 − 690. [4] TROLLE M, KÉRY M. Camera-trap study of ocelot and other secretive mammals in the northern Pantanal [J]. Mammalia, 2005, 69(3/4): 409 − 416. doi: 10.1515/mamm.2005.032 [5] KARANTH K U, NICHOLS J D. Estimation of tiger densities in India using photographic captures and recaptures [J]. Ecology, 1998, 79(8): 2852 − 2862. doi: 10.1890/0012-9658(1998)079[2852:EOTDII]2.0.CO;2 [6] AZLAN J M, SHARMA D S K. The diversity and activity patterns of wild felids in a secondary forest in Peninsular Malaysia [J]. Oryx, 2006, 40(1): 36 − 41. doi: 10.1017/S0030605306000147 [7] MÉNDEZ-CARVAJAL P G. The Orion Camera System, a new method for deploying camera traps in tree canopy to study arboreal primates and other mammals: a case study in Panama [J]. Mesoamericana, 2014, 18(1): 9 − 23. [8] 马世来, HARRIS RB. 自动感应照像系统在野生动物调查中的应用[J]. 动物学研究, 1996, 17(4): 360. [9] PEI J C. An evaluation of using auto-trigger cameras to record activity patterns of wild animals [J]. Taiwan Journal of Forest Science, 1998, 13(4): 317 − 324. [10] ROWCLIFFE J M, CARBONE C. Surveys using camera traps: are we looking to a brighter future? [J]. Animal Conservation, 2008, 11(3): 185 − 186. doi: 10.1111/j.1469-1795.2008.00180.x [11] 朱淑怡, 段菲, 李晟. 基于红外相机网络促进我国鸟类多样性监测: 现状、问题与前景[J]. 生物多样性, 2017, 25(10): 1114 − 1122. [12] 房以好, 任国鹏, 高颖, 等. 红外相机安放于地面和林冠层对野生动物监测结果的影响[J]. 生物多样性, 2018, 26(7): 717 − 726. [13] 邓怀庆, 周江. 海南长臂猿研究现状[J]. 四川动物, 2015, 34(4): 635 − 640. doi: 10.11984/j.issn.1000-7083.2015.04.025 [14] 周江, 陈辈乐, 魏辅文. 海南长臂猿的家族群相遇行为观察[J]. 动物学研究, 2008, 9(6): 667 − 673. [15] 王祝华. 极濒危物种海南长臂猿添丁 这是个关于生态保护的中国好故事 [N]. 科技日报, 2021-09-28(6). [16] 李梦瑶, 于伟慧. “猿”来有喜 [N]. 海南日报, 2022-04-13(A06). [17] CHAN B P L, LO Y F P, MO Y. New hope for the Hainan gibbon: formation of a new group outside its known range [J]. Oryx, 2020, 54(3): 296 − 296. [18] ZHOU J, WEI F, LI M, et al. Reproductive characters and mating behavior of wild Nomascus hainanus [J]. International Journal of Primatology, 2008, 29(4): 1037 − 1046. doi: 10.1007/s10764-008-9272-7 [19] 李萍. 翻山越岭来看你——追踪海南长臂猿[J]. 大自然, 2021, 42(6): 13 − 17. doi: 10.3969/j.issn.0255-7800.2021.06.004 [20] 刘振河, 覃朝锋. 海南长臂猿栖息地结构分析[J]. 兽类学报, 1990, 10(3): 163 − 169. [21] 林家怡, 莫罗坚, 庄雪影, 等. 海南黑冠长臂猿主要摄食植物的区系分布多样性研究[J]. 热带林业, 2006, 34(3): 21 − 24. doi: 10.3969/j.issn.1672-0938.2006.03.006 [22] LIU H, MA H, CHEYNE S M, et al. Recovery hopes for the world's rarest primate [J]. Science, 2020, 368(6495): 1074 − 1074. [23] 陈升华, 杨世彬, 许涵, 等. 海南长臂猿栖息地森林群落组成结构与多样性分析[J]. 广西林业科学, 2009, 38(4): 207 − 212. doi: 10.3969/j.issn.1006-1126.2009.04.003 [24] 唐玮璐, 毕玉, 金崑. 海南热带雨林国家公园海南长臂猿食源植物组成[J]. 野生动物学报, 2021, 42(3): 675 − 685. doi: 10.3969/j.issn.1000-0127.2021.03.004 [25] 邓立斌, 陈端吕, 邓丽群. 基于层次分析法的自然保护区模糊综合评价研究——以海南霸王岭国家级自然保护区为例[J]. 中国农学通报, 2013, 29(29): 118 − 125. doi: 10.11924/j.issn.1000-6850.2012-3096 [26] 胡磊. 基于红外自动相机技术的马鹿和狍活动节律与马鹿集群行为 [D]. 北京: 北京林业大学, 2013. [27] 肖治术, 李欣海, 姜广顺. 红外相机技术在我国野生动物监测研究中的应用[J]. 生物多样性, 2014, 22(6): 683 − 684. [28] 郑光美. 中国鸟类分类与分布名录 [M] .3版. 北京: 科学出版社, 2017. [29] 蒋志刚, 刘少英, 吴毅, 等. 中国哺乳动物多样性[J]. 生物多样性, 2017, 25(8): 886. doi: 10.17520/biods.2017098 [30] TIMOTHY G. O'BRIEN, MARGARET F, et al Crouching tigers, hidden prey: Sumatran tiger and prey populations in a tropical forest landscape [J]. Animal Conservation, 2003, 6(2): 131 − 139. doi: 10.1017/S1367943003003172 [31] 武鹏峰, 刘雪华, 蔡琼, 等. 红外相机技术在陕西观音山自然保护区兽类监测研究中的应用[J]. 兽类学报, 2012, 32(1): 67 − 71. [32] 孙儒泳. 动物生态学原理 [M]. 北京: 北京师范大学出版社, 2001. [33] 汪国海, 李生强, 施泽攀, 等. 广西猫儿山自然保护区的兽类和鸟类多样性初步调查——基于红外相机监测数据[J]. 兽类学报, 2016, 36(3): 338 − 347. [34] 刘周, 周虎, 吴国生, 等. 三江源查旦乡夏季大中型兽类与鸟类物种多样性和垂直分布格局[J]. 兽类学报, 2020, 40(5): 520 − 531. [35] BROWN J H, LIEBERMAN G A. Resource utilization and coexistence of seed‐eating desert rodents in sand dune habitats [J]. Ecology, 1973, 54(4): 788 − 797. doi: 10.2307/1935673 [36] SCHOENER T W. Resource partitioning in ecological communities: Research on how similar species divide resources helps reveal the natural regulation of species diversity [J]. Science, 1974, 185(4145): 27 − 39. doi: 10.1126/science.185.4145.27 [37] TOKESHI M. Species Coexistence: Ecological and Evolutionary Perspectives [M]. New York:John Wiley & Sons, 2009. [38] SCHREIER B M, HARCOURT A H, COPPETO S A, et al. Interspecific competition and niche separation in primates: a global analysis [J]. Biotropica, 2009, 41(3): 283 − 291. doi: 10.1111/j.1744-7429.2008.00486.x [39] 徐龙辉. 海南岛生态系统与野生动物资源的发展问题[J]. 热带地理, 1982, 2(3): 27 − 30. doi: 10.13284/j.cnki.rddl.001732 [40] MCCAIN C M. Global analysis of bird elevational diversity [J]. Global Ecology and Biogeography, 2009, 18(3): 346 − 360. doi: 10.1111/j.1466-8238.2008.00443.x [41] GRACANIN A, MIKAC K M. Camera traps reveal overlap and seasonal variation in the diel activity of arboreal and semi-arboreal mammals [J]. Mammalian Biology, 2022, 102(2): 341 − 355. doi: 10.1007/s42991-021-00218-y [42] 龚正达, 吴厚永, 段兴德, 等. 云南横断山区蚤类物种多样性的地理分布趋势与重要环境因素的关系[J]. 生物多样性, 2001, 9(4): 319 − 328. doi: 10.3321/j.issn:1005-0094.2001.04.001 [43] 唐玮璐, 金崑. 海南热带雨林国家公园海南长臂猿夜宿生境选择初步研究[J]. 北京林业大学学报, 2021, 43(2): 113 − 126. [44] 林家怡. 海南长臂猿取食植物及其分布特点与保护对策[C]//广东省植物学会. 广东省植物学会第十六期学术研讨会论文集. 2007: 42-55. [45] DAMMHAHN M, KAPPELER P M. Comparative feeding ecology of sympatric Microcebus berthae and M. murinus [J]. International Journal of Primatology, 2008, 29(6): 1567 − 1589. doi: 10.1007/s10764-008-9312-3 [46] 冯昌章. 两种共域热带鸟类的繁殖策略比较 [D]. 海口: 海南师范大学, 2019. [47] THORN M, SCOTT D M, GREEN M, et al. Estimating brown hyaena occupancy using baited camera traps [J]. South African Journal of Wildlife Research-24-month delayed open access, 2009, 39(1): 1 − 10. doi: 10.3957/056.039.0101 [48] KAYS R, TILAK S, KRANSTAUBER B, et al. Monitoring wild animal communities with arrays of motion sensitive camera traps [J]. International Journal of Research and Reviews in Wireless Sensor Networks, 2011, 1(3): 19 − 29. -

 点击查看大图
点击查看大图
图(3) / 表 (1)
计量
- 文章访问数: 510
- HTML全文浏览量: 325
- PDF下载量: 49
- 被引次数: 0
