


-
放线菌具有重要的生态学意义,也是很多生物活性物质的来源之一[1]。放线菌种类繁多,在陆地和水域生态系统中都有广泛分布。国内外的研究者在陆源放线菌及其活性物质方面已开展了较充分的研究,从相关菌株中也发现了许多结构新颖的活性化合物。目前对海洋环境中放线菌的研究还需进一步加强,特别是对深海区域放线菌的探索和开发尚处于起步阶段。地球表面约71%的面积是海洋,其中95%的海域深度超过1000 m[2],深海环境具有高压、低温、黑暗、寡营养等特征,生活在其中的生物也具备相应独特的适应能力,这些适应能力通常伴随着对基因、初级及次级代谢途径的调控[3]。研究发现在长期的进化过程中,深海放线菌等生物体内常产生有独特结构的次生代谢物,且具有一定的生物活性[4-5],而且这些活性物质具有是陆源微生物中未被发现过的新颖化合物结构[6],具有重要的研究价值。可见,广阔而独特的深海环境是研究和开发放线菌资源的良好场所。
南海是西太平洋最大的边缘海域,海域面积达350万km2,平均深度为1350 m,最深深度超过5.5 km[7-8]。冷泉是一种特殊的海底环境,在冷泉深部富含硫化氢、甲烷等碳氢化合物的流体会向海底表面渗漏或喷发,因此冷泉环境中含有大量的甲烷[9],因此冷泉环境中的微生物群落优势种属多为硫酸盐还原菌、好氧甲烷菌、氧甲烷菌、硫氧化菌等[10],该类群的冷泉微生物丰富度高于热液和普通海底沉积物[11]。不同冷泉的微生物群落存在差异[12]。南海冷泉区微生物资源多样性丰富[13],从南海冷泉区丰富的微生物资源中寻找新型次级代谢产物有利于加强对海马冷泉未知生物资源的开发与利用。已有许多新型活性物质来源于南海冷泉微生物[14-19],对微生物资源的利用推动了深海微生物资源的应用潜力评价。
海马冷泉位于南海西北陆坡,是南海2个活跃的冷泉之一[10],目前对海马冷泉微生物群落研究较少[20-23]。本研究主要在南海海马冷泉区,对沉积物中的放线菌进行分离纯化、培养鉴定、多样性分析及次级代谢产物抗菌活性的研究,旨在挖掘海马冷泉区可培养的放线菌资源,及其潜在的生物活性价值,为后续研究和开发冷泉放线菌资源提供参考资料。
-
沉积物样品由中国科学院深海科学与工程研究所“探索一号”科考船搭载的“深海勇士”号载人潜水器于2021年2月采集于南海海马冷泉区。样品采集后,放置4 ℃冰箱冷藏。巡航结束后,所有样品放在冰上运送到实验室,放置4 ℃保存备用。样品分为3组HM2、HM3、HM6(表1)。
表 1 深海沉积物样品信息表
样品 潜次 深度/m 坐标(经度E,纬度N) HM2-1a SY336 1365.0 110°28′20.67″,16°43′44.75″ HM2-1c SY336 1365.0 110°28′20.67″,16°43′44.75″ HM3-1a SY337 1383.0 110°27′31.32″,16°43′52.21″ HM3-1b SY337 1383.0 110°27′31.32″,16°43′52.21″ HM3-1c SY337 1383.0 110°27′31.32″,16°43′52.21″ HM3-1d SY337 1383.0 110°27′31.32″,16°43′52.21″ HM3-1f SY337 1383.0 110°27′31.32″,16°43′52.21″ HM6-1a SY340 1368.2 110°42′29.19″,16°25′40.40″ HM6-1b SY340 1368.2 110°42′29.19″,16°25′40.40″ HM6-1c SY340 1368.2 110°42′29.19″,16°25′40.40″ HM6-1d SY340 1368.2 110°42′29.19" ,16°25′40.40″ HM6-1e SY340 1368.2 110°42′29.19″,16°25′40.40″ HM6-1f SY340 1368.2 110°42′29.19″,16°25′40.40″ 注:样品名称最后的字母表示海水-沉积界面以下沉积深度,a:0~4 cm;b: 4~8 cm;c:8~12 cm;d:12~16 cm;e:16~20 cm;f:20~24 cm。 抗菌活性检测采用7种供试菌,即H119(粪肠球菌Enterococcus faecalis)、H57(甲氧西林耐药金黄色葡萄球菌methicillin-resistant Staphylococcus aureus, MRSA)、G280(甲氧西林敏感金黄色葡萄球菌methicillin-susceptible Staphylococcus aureus, MSSA)、H442(亚胺培南敏感鲍曼不动杆菌imipenem-susceptible Acinetobacter baumannii)、E292(亚胺培南耐药鲍曼不动杆菌imipenem-resistant Acinetobacter baumannii)、G1(高地芽孢杆菌Bacillus altitudinis)、F1(副溶血性弧菌Vibrio parahaemolyticus)。菌株分离、发酵、抑菌活性测试使用相应的培养基(表2)。
表 2 深海沉积物放线菌分离、发酵、活性测试所用培养基配方
用途 培养基名称 培养基组分 分离培养 ISP3琼脂培养基 燕麦 20 g,琼脂 18 g,七水合硫酸亚铁 0.001 g,四水合氯化亚锰 0.001 g,七水合硫酸锌 0.001 g,海水 1 L,pH=7.3 R2A琼脂培养基 R2A 18.1 g,海水 1 L,pH=7.0 分离/发酵培养 2216E培养基 Difco marine broth 2216E 37.4 g,琼脂 20 g,海水 1 L,pH=7.4 活性测试 LB培养基 胰蛋白胨 10 g,酵母提取物 5 g,氯化钠 10 g,ddH2O 1 L,琼脂 20 g,ddH2O 1 L,pH=7.4 -
在无菌条件下,称取0.1 g深海沉积物样品,加入1 mL无菌海水,振荡10 min,随后将沉积物悬浊液梯度稀释10−1到10−6,稀释倍数为10−6悬浊液100 μL于培养基中涂布,28 ℃倒置培养,每隔24 h观察菌落生长状况,待形成菌落后,挑取部分菌,采用三区划线法接种到分离培养基,重复纯化培养多次直至得到单一纯化菌株。随后用含25%甘油液体培养基冻存(−80 ℃)保种。
-
利用试剂盒提取DNA(TIANamp Bacteria DNA Kit),以16S rDNA序列通用引物27F/1492R[24]、Taq酶体系对其进行PCR扩增。对PCR产物验证通过后测序(擎科生物公司)。测序结果拼接完成后,上传EZbiocloud在线数据库(
https://www.ezbiocloud.net )进行对比分析,获得菌株的分类信息。以近缘菌株的16S rRNA基因序列作为参比对象,采用MEGA软件neighbor-joining构建系统发育树。 -
在2216E琼脂培养基上采用平板划线法培养菌株形成菌落,挑取单菌落,接种至5 mL发酵培养基中培养(28 ℃,180 r·min−1)3 d,获得种子菌液。随后,按照发酵培养基∶种子液(v∶v)= 100∶5的比例进行发酵培养(28 ℃,180 r·min−1)7 d,发酵液总体积为105 mL。发酵完成后,取乙酸乙酯与发酵液等体积充分混合,萃取3次,有机相合并后经旋转蒸发仪浓缩获得粗提物,−20℃保存备用。
-
采用K-B纸片扩散法药敏试验[25]检测放线菌发酵粗提物对7种供试菌的抑菌活性。用甲醇(分析纯)配制待测粗提物溶液(10 g·L−1),分2次滴加到灭菌滤纸片(直径6 mm)上,每次滴加5 μL,将风干的滤纸片贴在涂布了供试菌的固体培养基上(供试菌稀释至10−3涂布),采用万古霉素以及氯霉素(1 g·L−1)为阳性对照,甲醇溶液为空白对照,37 ℃培养24 h。通过与阳性对照的抑菌圈直径对比,判断粗提物的抑菌活性。
-
实验从HM2、HM3、HM6三组沉积样中分离出50株放线菌,从HM3样品组中分离的放线菌最多,共26株,其次是HM6样品组,19株,HM2样品组的最少,只有5株(图1)。这50株放线菌都属稀有类型,形态类似于细菌,无气生菌丝,隶属于7个属(图2),其中类诺卡式菌属(Nocardioides)22株、迪茨氏菌属(Dietzia)13株、乔治菌属(Georgenia)6株、考克氏菌属(Kocuria)4株、节杆菌属(Arthrobacter)2株、小单孢菌属(Micromonospora)2株、印度洋金色丸菌属(Chryseoglobus)1株。各放线菌属间的相对丰富度呈现一定差异,其中类诺卡氏菌属在3组样品中属于优势菌。采用2216E培养基、R2A琼脂培养基和ISP3琼脂培养基分别筛选到18、29和3株放线菌,R2A培养基更适合用于筛选深海放线菌。基于对潜在新菌种的界定[26-28],在这50株放线菌中,有6株与近缘菌株16S rRNA基因的相似度低于98.7%,初步判断为潜在新种(表3)。
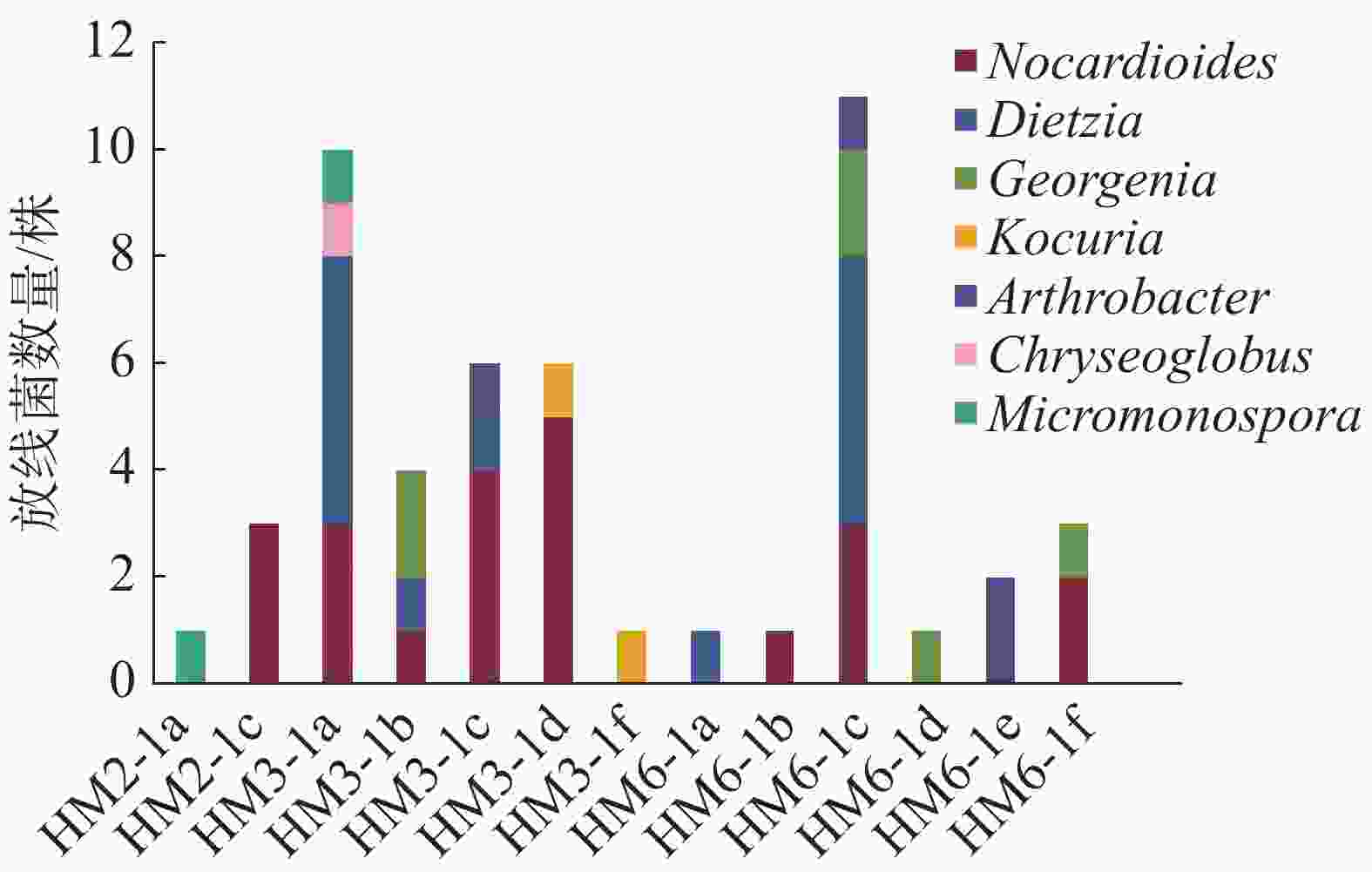
图 1 从各沉积样品中分离到的放线菌种类
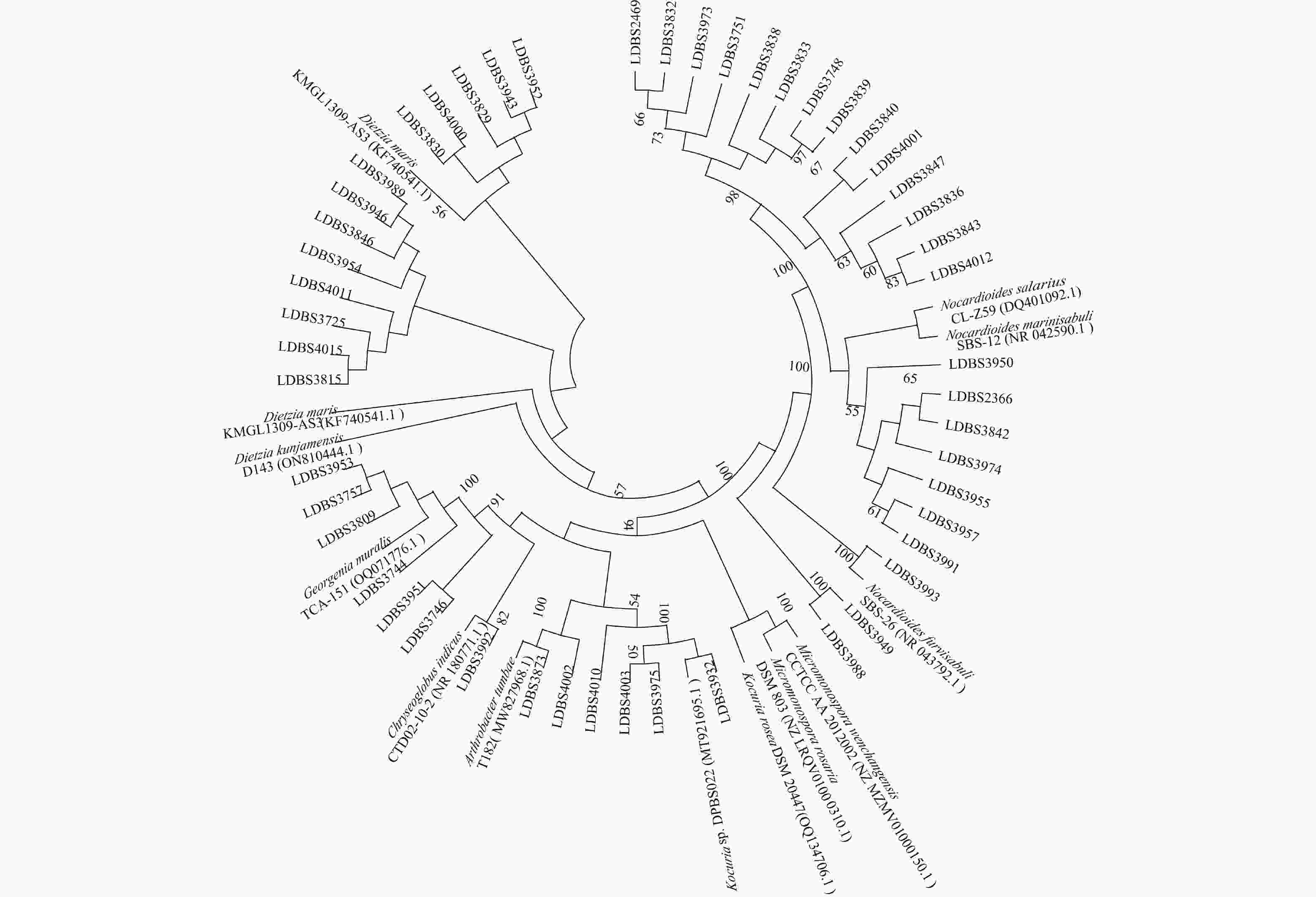
图 2 分离出的50株放线菌与近缘菌株的系统发育树
表 3 50株放线菌中潜在的新菌株
菌株编号 近缘菌株 相似度/% 样品来源 LDBS3725 Dietzia maris DSM 43672 98.66 HM3-1a LDBS3838 Nocardioides marinisabuli SBS-12 98.65 HM3-1c LDBS3839 Nocardioides marinisabuli SBS-12 98.58 HM3-1c LDBS3840 Nocardioides salarius CL-Z59 98.66 HM3-1c LDBS3974 Nocardioides salaries CL-Z59 98.52 HM3-1a LDBS3988 Micromonospora rosaria DSM 803 98.52 HM3-1a -
通过对50株放线菌的发酵液粗提物进行抑菌活性分析,发现7株放线菌的粗提物对3种供试菌显示出不同程度的抑菌作用,抑制活性比例为14%。其中,5株菌对F1有抑菌活性,4株对G280有活性,2株对H57有活性。编号为 LDBS2366和LDBS2469的菌株粗提物仅对 G280 有抑菌活性。 LDBS3832、 LDBS3950和 LDBS3974对F1有轻微抑菌活性,此外LDBS3974 对H57也有轻微抑菌活性。LDBS3949的菌株粗提物对H57、G280、F1均有抑菌活性。LDBS3988的菌株粗提物对G280和F1有抑菌活性。50株放线菌粗提物对H119、H422、E292、G1均没有活性。小单孢菌属的抑菌活性及抑菌范围都优于类诺卡式菌属(表4)。其中,小单孢菌属的菌株LDBS3949和LDBS3988呈现出良好的抑菌效果。
表 4 放线菌代表性菌株的抗菌活性
菌株编号及阳性对照 菌株名称 抑菌圈直径/mm H119 H57 G280 H422 E292 G1 F1 LDBS2366 Nocardioides sp. - - 10 - - - - LDBS2469 Nocardioides sp. - - 12 - - - - LDBS3832 Nocardioides sp. - - - - - - 7 LDBS3949 Micromonospora sp. - 15 12 - - - 22 LDBS3950 Nocardioides sp. - - - - - - 10 LDBS3974 Nocardioides sp. - 7 - - - - 7 LDBS3988 Micromonospora sp. - - 30 - - - 20 万古霉素 19 20 20 - - 18 20 氯霉素 - - - 20 20 - - 注:H119:粪肠球菌,H57:甲氧西林耐药金黄色葡萄球菌,G280:甲氧西林敏感金黄色葡萄球菌,H442:亚胺培南敏感鲍曼不动杆菌,E292:亚胺培南耐药鲍曼不动杆菌,G1:高地芽孢杆菌,F1:副溶血性弧菌;- :无活性,除H422和E292以氯霉素为阳性对照,其余病原菌以万古霉素为阳性对照。 -
从南海海马冷泉区的三组沉积物样品中分离到50株放线菌,属于5个科,7个属,其中类诺卡式菌属、迪茨氏菌属是优势菌,后者为海洋特有。这与从南大西洋深海沉积物分离出的36株放线菌不同,属于6个科,8个属,其中微球菌属、迪茨氏菌属和微杆菌属是优势菌[29],也有实验分离出132株放线菌,属于13个科,19个属,其中考克氏菌属、短杆菌属和链霉菌属是优势菌[30]。在这些沉积样品中,共有种属的放线菌有迪茨氏菌属、考克氏菌属、微球菌属、微杆菌属。不同沉积样品的放线菌种属存在着差异,这可能一方面是源自地域环境差异,另一方面是因不同的培养条件和分离方法造成的。基于对潜在新种的界定,本研究分离的放线菌中有6株的相似性低于98.7%初步认为是潜在新种,它们属于迪茨氏菌属、类诺卡式菌属、小单孢菌属。有研究发现Nocardioides sp. strain CF18利用单加氧酶(pBMO)在丁烷上生长,而pBMO与甲烷氧化细菌的甲烷单加氧酶(pMMO)基因序列相似[31],推测这些酶助力于类诺卡式菌属菌株在含大量甲烷环境的冷泉中能生存并成为优势属。
本研究中发现诺卡式菌属和小单孢菌属的发酵液粗提物都呈现出较好的抗菌活性,其中,小单孢菌属的LDBS3988和LDBS3949菌株对G+ 菌MRSA、MSSA,以及G− 菌V. parahaemolyticus都表现出抑制作用。Zhang等[32]曾从南海沉积物来源的小单孢菌属的次级代谢产物中分离到3个新化合物fluostatins I−K,小单孢菌属的次级代谢产物对金黄色葡萄球菌具有较好的抑制作用。除了能产多种化合物的小单孢菌属外,本研究分离到的类诺卡式菌属对MRSA、MSSA、V. parahaemolyticus也呈现出一定的抗菌活性,这是首次报道深海来源类诺卡式菌属有抑菌活性。尤其是小单孢菌属的2株菌LDBS3988和LDBS3949的发酵粗提物呈现较强的抑菌活性,具有较好的深入研究价值。
目前对南海冷泉沉积样品中硫化细菌的研究较多[33],但对放线菌的研究还较少。本研究揭示了冷泉区丰富的放线菌资源,这些具有活性的菌株可作为挖掘深海放线菌抗菌天然产物的重要资源。后期研究将通过优化培养条件、发酵条件、激活沉默次级代谢合成途径等手段进一步挖掘新颖活性化合物,为从深海极端环境中寻找新型抗生素提供参考资料。
南海冷泉区深海沉积物来源放线菌的分离培养及其抑菌活性
DOI: 10.15886/j.cnki.rdswxb.20230022
 CSTR: 32425.14.j.cnki.rdswxb.20230022
CSTR: 32425.14.j.cnki.rdswxb.20230022
Isolation, cultivation and antibacterial activity of actinomycetes from the sediments in the deep-sea cold seeps of South China Sea
-
摘要: 为挖掘南海冷泉区可培养放线菌资源及其潜在的生物活性,采用稀释涂布法分离培养了来源于南海冷泉区深海沉积物中的放线菌,利用16S rRNA基因测序鉴定菌株,构建系统发育树,并分析了放线菌的多样性;通过K-B法药敏试验对放线菌发酵粗提物进行抑菌活性测试。结果显示:分离获得的50株放线菌隶属于5个科,7个属,其中5株放线菌被初步判定为潜在新种。7株放线菌的发酵粗提物对副溶血性弧菌、甲氧西林敏感金黄色葡萄球菌和甲氧西林耐药金黄色葡萄球菌呈现不同程度的抑菌活性,其中活性最好的为小单孢菌属(Micromonospora)的2株菌,Micromonospora sp. LDBS3988抑菌活性最强,Micromonospora sp. LDBS3949抑菌活性较为广谱。Abstract: In order to explore the resources and potential bioactivity value of actinomycetes, actinomycetes were isolated from the deep-sea sediments in the cold seeps of South China Sea by serial dilution and spread plate technique and cultured. The strains of isolated actinomycetes were identified by using 16S rRNA gene sequencing, based on which a phylogenetic tree was constructed, and the diversity of actinomycetes were analyzed. The antibacterial activities of the crude extracts of the isolated actinomycetes were tested by using the K-B test for antibiotic susceptibility. The results showed that 50 isolated actinomyces belonged to 5 families and 7 genera, and that 5 strains of actinomycetes were preliminarily identified as potential new species. And the crude extracts of 7 strains showed different antibacterial activities against methicillin-resistant Staphylococcus aureus, methicillin-sensitive Staphylococcus aureus and Vibrio parahaemolyticus. Among the 7 strains, two strains of Micromonospora displayed the highest antibacterial activity. Micromonospora sp.LDBS3988 has the highest antibacterial activity, and Micromonospora sp.LDBS3949 has a broad-spectrum antibacterial activity.
-
Key words:
- deep-sea sediment /
- actinomycetes /
- antibacterial activity /
- cold seep
-
图 2 分离出的50株放线菌与近缘菌株的系统发育树
系统发育树为1000次Bootstrap分析所得结果;图中的数值是Bootstrap大于50%的结果,进化距离为0.005。
表 1 深海沉积物样品信息表
样品 潜次 深度/m 坐标(经度E,纬度N) HM2-1a SY336 1365.0 110°28′20.67″,16°43′44.75″ HM2-1c SY336 1365.0 110°28′20.67″,16°43′44.75″ HM3-1a SY337 1383.0 110°27′31.32″,16°43′52.21″ HM3-1b SY337 1383.0 110°27′31.32″,16°43′52.21″ HM3-1c SY337 1383.0 110°27′31.32″,16°43′52.21″ HM3-1d SY337 1383.0 110°27′31.32″,16°43′52.21″ HM3-1f SY337 1383.0 110°27′31.32″,16°43′52.21″ HM6-1a SY340 1368.2 110°42′29.19″,16°25′40.40″ HM6-1b SY340 1368.2 110°42′29.19″,16°25′40.40″ HM6-1c SY340 1368.2 110°42′29.19″,16°25′40.40″ HM6-1d SY340 1368.2 110°42′29.19" ,16°25′40.40″ HM6-1e SY340 1368.2 110°42′29.19″,16°25′40.40″ HM6-1f SY340 1368.2 110°42′29.19″,16°25′40.40″ 注:样品名称最后的字母表示海水-沉积界面以下沉积深度,a:0~4 cm;b: 4~8 cm;c:8~12 cm;d:12~16 cm;e:16~20 cm;f:20~24 cm。  下载: 导出CSV
下载: 导出CSV
表 2 深海沉积物放线菌分离、发酵、活性测试所用培养基配方
用途 培养基名称 培养基组分 分离培养 ISP3琼脂培养基 燕麦 20 g,琼脂 18 g,七水合硫酸亚铁 0.001 g,四水合氯化亚锰 0.001 g,七水合硫酸锌 0.001 g,海水 1 L,pH=7.3 R2A琼脂培养基 R2A 18.1 g,海水 1 L,pH=7.0 分离/发酵培养 2216E培养基 Difco marine broth 2216E 37.4 g,琼脂 20 g,海水 1 L,pH=7.4 活性测试 LB培养基 胰蛋白胨 10 g,酵母提取物 5 g,氯化钠 10 g,ddH2O 1 L,琼脂 20 g,ddH2O 1 L,pH=7.4
下载: 导出CSV
表 3 50株放线菌中潜在的新菌株
菌株编号 近缘菌株 相似度/% 样品来源 LDBS3725 Dietzia maris DSM 43672 98.66 HM3-1a LDBS3838 Nocardioides marinisabuli SBS-12 98.65 HM3-1c LDBS3839 Nocardioides marinisabuli SBS-12 98.58 HM3-1c LDBS3840 Nocardioides salarius CL-Z59 98.66 HM3-1c LDBS3974 Nocardioides salaries CL-Z59 98.52 HM3-1a LDBS3988 Micromonospora rosaria DSM 803 98.52 HM3-1a
下载: 导出CSV
表 4 放线菌代表性菌株的抗菌活性
菌株编号及阳性对照 菌株名称 抑菌圈直径/mm H119 H57 G280 H422 E292 G1 F1 LDBS2366 Nocardioides sp. - - 10 - - - - LDBS2469 Nocardioides sp. - - 12 - - - - LDBS3832 Nocardioides sp. - - - - - - 7 LDBS3949 Micromonospora sp. - 15 12 - - - 22 LDBS3950 Nocardioides sp. - - - - - - 10 LDBS3974 Nocardioides sp. - 7 - - - - 7 LDBS3988 Micromonospora sp. - - 30 - - - 20 万古霉素 19 20 20 - - 18 20 氯霉素 - - - 20 20 - - 注:H119:粪肠球菌,H57:甲氧西林耐药金黄色葡萄球菌,G280:甲氧西林敏感金黄色葡萄球菌,H442:亚胺培南敏感鲍曼不动杆菌,E292:亚胺培南耐药鲍曼不动杆菌,G1:高地芽孢杆菌,F1:副溶血性弧菌;- :无活性,除H422和E292以氯霉素为阳性对照,其余病原菌以万古霉素为阳性对照。
下载: 导出CSV
-
[1] SUBRAMANI R, AALBERSBERG W. Marine actinomycetes: an ongoing source of novel bioactive metabolites [J]. Microbiological Research, 2012, 167(10): 571 − 580. doi: 10.1016/j.micres.2012.06.005 [2] KAMJAM M, SIVALINGAM P, DENG Z, et al. Deep Sea actinomycetes and their secondary metabolites [J]. Frontiers in Microbiology, 2017, 8: 760. doi: 10.3389/fmicb.2017.00760 [3] SKROPETA D, WEI L. Recent advances in deep-sea natural products [J]. Natural Product Reports, 2014, 31(8): 999 − 1025. doi: 10.1039/C3NP70118B [4] WRIGHT P C, WESTACOTT R E, BURJA A M. Piezotolerance as a metabolic engineering tool for the biosynthesis of natural products [J]. Biomolecular Engineering, 2003, 20(4/5/6): 325 − 331. doi: 10.1016/s1389-0344(03)00042-x [5] MA L, BARTHOLOME A, TONG M H, et al. Identification of a fluorometabolite from Streptomyces sp. MA37: (2R3S4S)-5-fluoro-2, 3, 4-trihydroxypentanoic acid [J]. Chemical Science, 2015, 6(2): 1414 − 1419. doi: 10.1039/c4sc03540b [6] RAMALINGAM V, RAJARAM R. Antioxidant activity of 1-hydroxy-1-norresistomycin derived from Streptomyces variabilis KP149559 and evaluation of its toxicity against zebra fish Danio rerio [J]. RSC Advances, 2016, 6(20): 16615 − 16623. doi: 10.1039/C5RA22558B [7] SU J. Overview of the South China Sea circulation and its influence on the coastal physical oceanography outside the Pearl River Estuary [J]. Continental Shelf Research, 2004, 24(16): 1745 − 1760. doi: 10.1016/j.csr.2004.06.005 [8] 蔡观强, 彭学超, 张玉兰. 南海沉积物物质来源研究的意义及其进展[J]. 海洋科学进展, 2011, 29(1): 113 − 121. doi: 10.3969/j.issn.1671-6647.2011.01.014 [9] BOETIUS A, WENZHöFER F. Seafloor oxygen consumption fuelled by methane from cold seeps [J]. Nature Geoscience, 2013, 6(9): 725 − 734. doi: 10.1038/ngeo1926 [10] FENG D, QIU J W, HU Y, et al. Cold seep systems in the South China Sea: an overview [J]. Journal of Asian Earth Sciences, 2018, 168: 3 − 16. doi: 10.1016/j.jseaes.2018.09.021 [11] KNITTEL K, BOETIUS A. Anaerobic oxidation of methane: progress with an unknown process [J]. Annual Review of Microbiology, 2009, 63: 311 − 334. doi: 10.1146/annurev.micro.61.080706.093130 [12] VIGNERON A, CRUAUD P, PIGNET P, et al. Archaeal and anaerobic methane oxidizer communities in the Sonora Margin cold seeps, Guaymas Basin (Gulf of California) [J]. The ISME Journal, 2013, 7(8): 1595 − 1608. doi: 10.1038/ismej.2013.18 [13] 孙瑜, 牛明杨, 刘俏, 等. 南海Formosa冷泉区沉积物微生物多样性与分布规律研究[J]. 微生物学报, 2022, 62(6): 2001 − 2020. [14] HU X Y, LI X M, YANG S Q, et al. New cytochalasin derivatives from deep-sea cold seep-derived endozoic fungus Curvularia verruculosa CS-129 [J]. Chemistry & Biodiversity, 2022, 19(8): e202200550. doi: 10.1002/cbdv.202200550 [15] HU X Y, WANG C Y, LI X M, et al. Cytochalasin derivatives from the endozoic Curvularia verruculosa CS-129, a fungus isolated from the deep-sea squat lobster Shinkaia crosnieri living in the cold seep environment [J]. Journal of Natural Products, 2021, 84(12): 3122 − 3130. doi: 10.1021/acs.jnatprod.1c00907 [16] SONG Q, YANG S Q, LI X M, et al. Aromatic polyketides from the deep-sea cold-seep mussel associated endozoic fungus Talaromyces minioluteus CS-138 [J]. Marine Drugs, 2022, 20(8): 529. doi: 10.3390/md20080529 [17] HU X, LI X, YANG S, et al. Vercytochalasins A and B: Two unprecedented biosynthetically related cytochalasins from the deep-sea-sourced endozoic fungus Curvularia verruculosa [J]. Chinese Chemical Letters, 2023, 34(2): 107516. doi: 10.1016/j.cclet.2022.05.030 [18] YAN L H, LI P H, LI X M, et al. Chevalinulins A and B, proangiogenic alkaloids with a spiro[bicyclo[2.2. 2]octane-diketopiperazine]skeleton from deep-sea cold-seep-derived fungus Aspergillus chevalieri CS-122 [J]. Organic Letters, 2022, 24(14): 2684 − 2688. doi: 10.1021/acs.orglett.2c00781 [19] JIN E, LI H, LIU Z, et al. Antibiotic dixiamycins from a cold-seep-derived Streptomyces olivaceus [J]. Journal of Natural Products, 2021, 84(9): 2606 − 2611. doi: 10.1021/acs.jnatprod.1c00411 [20] NIU M, FAN X, ZHUANG G, et al. Methane-metabolizing microbial communities in sediments of the Haima cold seep area, northwest slope of the South China Sea [J]. FEMS Microbiology Ecology, 2017, 93(9): fix101. doi: 10.1093/femsec/fix101 [21] ZHUANG G C, XU L, LIANG Q, et al. Biogeochemistry, microbial activity, and diversity in surface and subsurface deep-sea sediments of South China Sea [J]. Limnology and Oceanography, 2019, 64(5): 2252 − 2270. doi: 10.1002/lno.11182 [22] JING H, WANG R, JIANG Q, et al. Anaerobic methane oxidation coupled to denitrification is an important potential methane sink in deep-sea cold seeps [J]. Science of the Total Environment, 2020, 748: 142459. doi: 10.1016/j.scitotenv.2020.142459 [23] LING J, GUAN H, LIU L, et al. The diversity, composition, and putative functions of gill-associated bacteria of bathymodiolin mussel and vesicomyid clam from Haima cold seep, South China Sea [J]. Microorganisms, 2020, 8(11): 1699. doi: 10.3390/microorganisms8111699 [24] WEISBURG W G, BARNS S M, PELLETIER D A, et al. 16S ribosomal DNA amplification for phylogenetic study [J]. Journal of Bacteriology, 1991, 173(2): 697 − 703. doi: 10.1128/jb.173.2.697-703.1991 [25] FOR THE BSAC WORKING PARTY ON SUSCEPTIBILITY TESTING, ANDREWS J M. BSAC standardized disc susceptibility testing method [J]. Journal of Antimicrobial Chemotherapy, 2001, 48(suppl_1): 43 − 57. doi: 10.1093/jac/48.suppl_1.43 [26] 肖珂, 周双清, 许云, 等. 海绵共附生放线菌的分离鉴定与抑菌活性分析[J]. 热带生物学报, 2020, 11(2): 156 − 162. [27] CHENG C, OTHMAN E M, REIMER A, et al. Ageloline A, new antioxidant and antichlamydial quinolone from the marine sponge-derived bacterium Streptomyces sp. SBT345 [J]. Tetrahedron Letters, 2016, 57(25): 2786 − 2789. doi: 10.1016/j.tetlet.2016.05.042 [28] KIM M, OH H S, PARK S C, et al. Towards a taxonomic coherence between average nucleotide identity and 16S rRNA gene sequence similarity for species demarcation of prokaryotes [J]. International Journal of Systematic and Evolutionary Microbiology, 2014, 64(Pt 2): 346−351. doi: 10.1099/ijs.0.059774-0 [29] 贾文文. 南大西洋深海放线菌的分离与多样性分析 [D]. 哈尔滨: 哈尔滨工业大学, 2013. [30] 张玉便, 张改云. 南大西洋深海沉积物中可培养放线菌的多样性[J]. 应用海洋学学报, 2014, 33(4): 508 − 515. doi: 10.3969/J.ISSN.2095-4972.2014.04.008 [31] SAYAVEDRA-SOTO L A, HAMAMURA N, LIU C W, et al. The membrane-associated monooxygenase in the butane-oxidizing Gram-positive bacterium Nocardioides sp. strain CF8 is a novel member of the AMO/PMO family [J]. Environmental Microbiology Reports, 2011, 3(3): 390 − 396. doi: 10.1111/j.1758-2229.2010.00239.x [32] ZHANG W, LIU Z, LI S, et al. Fluostatins i–k from the South China Sea-derived Micromonospora rosaria scsio N160 [J]. Journal of Natural Products, 2012, 75(11): 1937 − 1943. doi: 10.1021/np300505y [33] RUFF S E, BIDDLE J F, TESKE A P, et al. Global dispersion and local diversification of the methane seep microbiome [J]. Proceedings of the National Academy of Sciences of the United States of America, 2015, 112(13): 4015 − 4020. doi: 10.1073/pnas.1421865112 -

 点击查看大图
点击查看大图

图(2) / 表 (4)
计量
- 文章访问数: 1551
- HTML全文浏览量: 719
- PDF下载量: 38
- 被引次数: 0
