


-
金枪鱼隶属鲈形目(Pereiformes)鲭科(Scombridae)金枪鱼属(Thunnus)。金枪鱼属有黄鳍金枪鱼Thunnus albacares、蓝鳍金枪鱼Thunnus maccoyii、大眼金枪鱼Thunnus obesus、大西洋蓝鳍金枪鱼Thunnus thynnus、太平洋蓝鳍金枪鱼Thunnus orientalis、长鳍金枪鱼Thunnus alalunga、黑鳍金枪鱼Thunnus atlanticus及青干金枪鱼Thunnus tonggol共8个物种。金枪鱼肉是典型的高蛋白,低热量的海洋食品,富含对人体有益的不饱和脂肪酸(如二十二碳六烯酸及十二碳五烯酸)[1]。食用金枪鱼对保持人体营养平衡、减肥及降低胆固醇含量等具有积极作用[2]。金枪鱼具有较高的经济价值,是远洋商业捕捞的主要对象之一。为保护金枪鱼种群资源及确保捕捞的可持续发展,相关国家先后联合成立了5个政府间区域性金枪鱼渔业管理组织(Tuna Regional Fisheries Management Organizations,tRFMOs)。
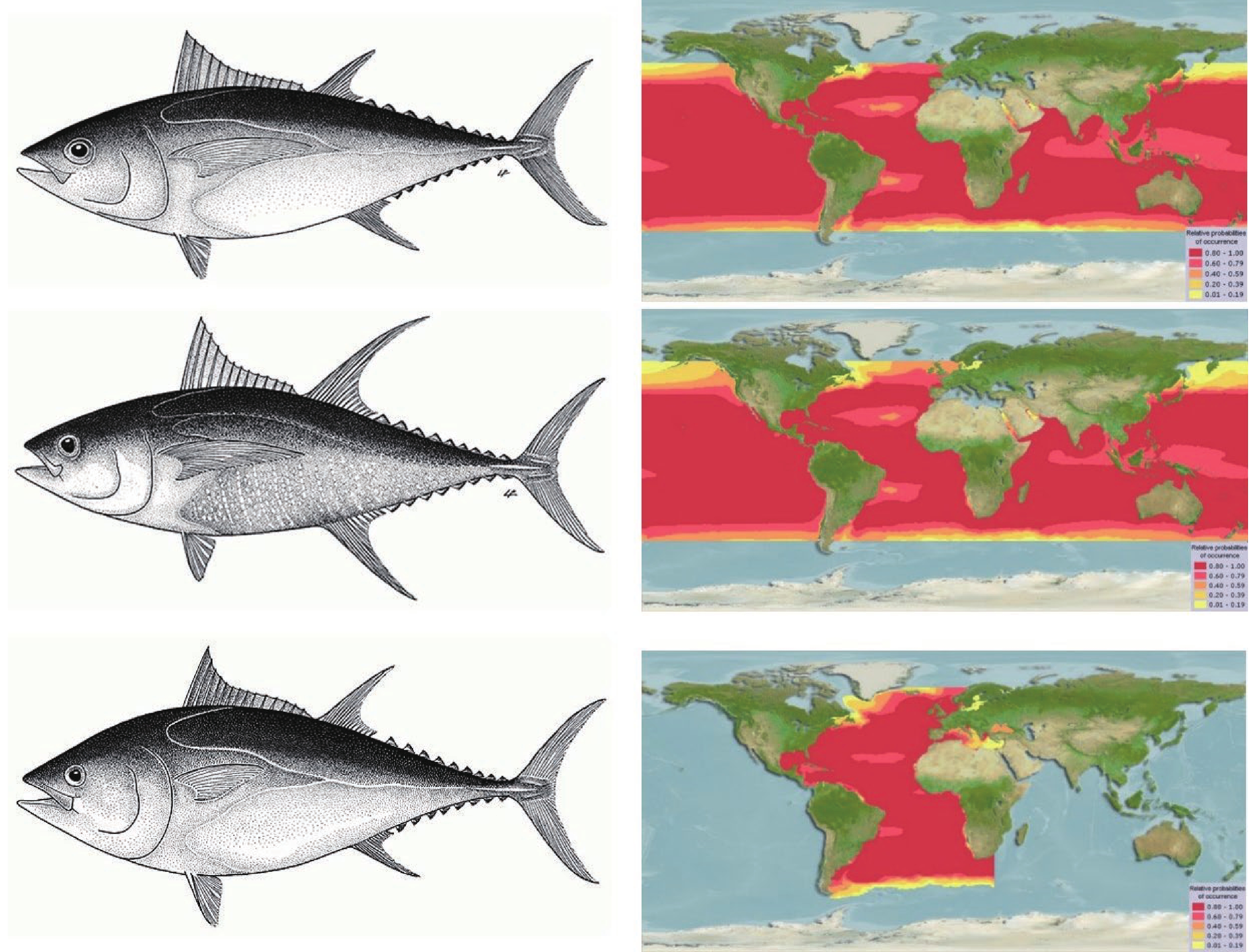
金枪鱼作为一种生活在海洋中上层的大洋洄游性高营养级捕食者,为提高种群对外界环境的适合度,在鱼龄与生长、摄食及繁殖3个生物学方面形成了特有的生物学特征以应对多种外界不利因素(如洄游导致的生存环境高不确定性、高商业价值导致的高被捕捞风险及金枪鱼种间生态位重叠等)。生长方面,金枪鱼幼鱼具有体长增速较快的特点,该特点降低了幼鱼被其他肉食鱼类捕获的概率,提高了金枪鱼幼鱼的存活率,而成年后体长增速下降、体质量增速增加,为成年金枪鱼的繁殖活动打下坚实基础[3]。在摄食方面,为适应大洋洄游的特点,金枪鱼进化为机会捕食者(opportunistic predators),可根据外界环境中饵料丰度的变化改变自身的摄食习性[4],大大提高了洄游时个体的存活率。在繁殖方面,金枪鱼会根据自身身体状况及外界环境,对繁殖的适合程度做出相对应的策略(reproductive strategies),以提高后代的存活率[5]。因此,本研究以经济价值较高、研究最为深入的黄鳍金枪鱼,大眼金枪鱼及大西洋蓝鳍金枪鱼(简称蓝鳍金枪鱼)3种金枪鱼为例(图1),总结金枪鱼在生物学鱼龄与生长、摄食及繁殖3个方面的研究进展。同时,针对金枪鱼生物学研究领域现存的不足,从研究方法、设备和技术手段等为金枪鱼的保护及相关生物学研究提供重要的理论依据。
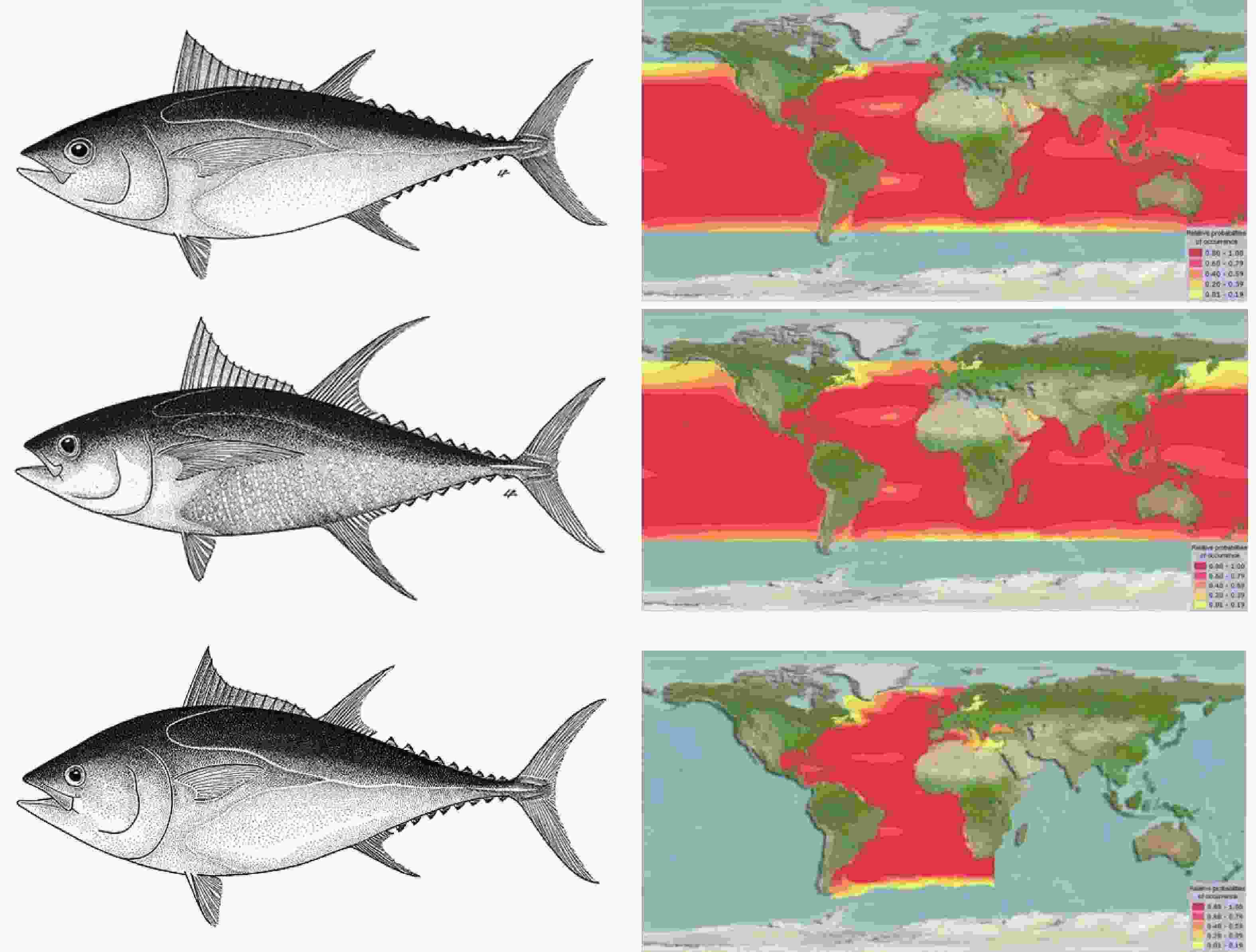
图 1 大眼金枪鱼(上)、黄鳍金枪鱼(中)、蓝鳍金枪鱼(下)及各自分布海域
-
鉴定材料选取的合适与否,对鉴定金枪鱼个体鱼龄及判断种群鱼龄结构的准确度具有重要影响。与高速游泳且大洋洄游的高营养级捕食者剑鱼Xiphias gladius相似,金枪鱼鱼龄鉴定材料主要有2种,分别是鱼体内硬骨组织中的耳石与脊椎[6-8],其中,脊椎被选作鉴定材料的频率最高。通常金枪鱼鱼龄是通过鉴定样本的单一硬骨组织材料所得,但吕凯凯[9]在对耳石、鳍棘及脊椎等多种鉴定材料进行比较后发现,用单一的鉴定材料判断鱼龄准确度不高,如,耳石截面存在明显日轮,但过于密集,读数困难;鳍棘截面上存在清晰的轮纹,但受脉管化现象影响,只能准确鉴定1~3龄的金枪鱼;脊椎存在副轮和边缘轮现象,鉴定准确度受此影响而较低。因此,吕凯凯认为多种硬骨组织材料同时鉴定所得结果更为准确。目前,相关研究领域普遍将“耳石-脊椎”组合作为鉴定金枪鱼鱼龄的最佳选择。金枪鱼寿命较长,平均寿命15 a[10]。与生态位相似的剑鱼及同属鲭科的鲣鱼Katsuwonus pelamis相比,金枪鱼寿命周期较雌性剑鱼短(雌性剑鱼寿命可达15~32 a),但长于鲣鱼(鲣鱼体型较金枪鱼小,但发育迅速,寿命短,仅8~12 a)[6]。目前记载鱼龄最大的蓝鳍金枪鱼叉长427 cm,体质量726 kg,20龄[11]。
-
与鲣鱼和剑鱼相似,金枪鱼幼鱼时期体长生长增速较快,且0~1龄体长增速最快,以大眼金枪鱼为例,年平均体长增长59 cm[10]。有研究表明,产卵前,金枪鱼、剑鱼及鲣鱼体长的增速较快,未产卵及未性成熟的幼鱼一般体型呈修长状,产卵后侧重于体质量的增加,所以金枪鱼成鱼体型多呈短粗状[3]。金枪鱼不同时段生长侧重不同的原因可能是受到igfl,igflr及mTOR等金枪鱼相关生长基因的调控。
与大多数“单拐点”生长增速(在性成熟前后出现增速拐点)的鱼类不同,金枪鱼体长生长增速具有多拐点的特征。该特征导致金枪鱼体长与时间之间的关系并非为“S型”,具体表现为:金枪鱼0~1龄时期增速最大(拐点1),而在2龄时出现生长增速最低值(拐点2),之后增速上升,在产卵后增速开始缓慢下降(拐点3)[10]。0~1龄时期体长增速最大(拐点1),可能是生长跨度较大的鱼类为提高自身存活率而具有的共同特性(与剑鱼相似);产卵后体长增速下降则是受到基因的调控,产卵后侧重体质量的增加;有学者认为,鱼类初次性成熟会消耗大量能量,导致该时段用于体型生长的能量大大减少,进而体长生长增速下降,而金枪鱼初次性成熟在2龄,因此最低生长增速在2龄时出现[12]。
-
根据已捕获样本的生长参数(叉长、体质量、鱼龄)建立模型,如代表叉长与体质量之间关系的幂函数W = aLb及描述金枪鱼生长特性的生长模型。在多种生长模型中选择与金枪鱼生长特性最适配的一种,对于能否准确描述金枪鱼的生长及其特性十分关键。因此,多位学者对现存的多种生长模型(如von Bertalanffy,Gompertz及Richards)进行赤池信息准则(Akaike information criterion,AIC)检测,结果表明,在现存模型中von Bertalanffy生长模型与金枪鱼适配性最高[13]。此后,von Bertalanffy生长模型被广泛用于描述金枪鱼的生长及生长特性(表1)。然而,在von Bertalanffy生长模型是否真正适用于描述金枪鱼生长特性的问题上,研究学者之间一直存在争议。WCPFC(Western and Central Pacific Fisheries Commission,中西太平洋渔业委员会)的研究员Lehodey[14]通过“电子标记+再捕获”法发现黄鳍金枪鱼生长具有“两段性”特征后(即多体长增速拐点),最先对von Bertalanffy生长模型是否最合适提出质疑。Lehodey[14]推测,由于金枪鱼生长跨度大,在不同的生长阶段生长发育、捕食活动及产热等生命活动耗能相差较大,导致不同生长阶段用于体型中体长增长的能量存在巨大差异,使得成年与未成年金枪鱼之间体长变化呈现明显的“两段性”,这种特征使得von Bertalanffy生长模型无法准确描述金枪鱼的生长特性。除上述原因外,还有学者对von Bertalanffy生长模型输入的相关生长参数要求进行分析,认为von Bertalanffy生长模型不适合用于描述金枪鱼的生长特性,因为von Bertalanffy生长模型的准确度与生长参数的覆盖范围广度密切相关,但金枪鱼取样困难,所得参数无法保证覆盖范围的全面性。例如,在野外取样中往往难以捕获体型大的个体,数据上缺少大体型金枪鱼的生长参数,最终会导致von Bertalanffy生长模型中参数渐进体长(L∞)较实际值偏小,对鱼群整体的生长描述产生偏差[15-16]。
表 1 不同海域von Bertalanffy生长模型拟合大眼金枪鱼、黄鳍金枪鱼及蓝鳍金枪鱼生长参数
种类 捕捞方式 海域 生长系数K 渐进体长/cm 生长起点鱼龄t0 肥满度a(×10−4) 幂指数b 参考文献 大眼金枪鱼 延绳钓 太平洋 0.18 178.41 −2.50 − − [17] 大眼金枪鱼 延绳钓 太平洋 0.20 208.70 −0.99 − − [18] 大眼金枪鱼 延绳钓 太平洋 − − − 5.15 2.76 [19] 大眼金枪鱼 延绳钓 大西洋 0.12 264.02 −0.68 − − [20] 大眼金枪鱼 延绳钓 印度洋 0.13 207.14 −1.89 − − [8] 黄鳍金枪鱼 延绳钓 太平洋 0.25 181.81 −0.48 5.55 2.72 [13] 黄鳍金枪鱼 延绳钓 太平洋 − − − 2.19 2.94 [21] 黄鳍金枪鱼 延绳钓 大西洋 0.27 230.70 −0.08 − − [22] 黄鳍金枪鱼 延绳钓 大西洋 − − − 0.85 3.14 [23] 黄鳍金枪鱼 延绳钓 印度洋 0.18 272.70 −0.27 1.59 3.30 [24] 蓝鳍金枪鱼 延绳钓 大西洋 314.90 0.09 −0.15 − − [25] 蓝鳍金枪鱼 延绳钓 大西洋 304.00 0.10 −0.65 − − [26] 蓝鳍金枪鱼 延绳钓 大西洋 373.08 0.07 −1.76 − − [27] -
在稳定同位素分析等方法兴起之前,“胃容物鉴定”是确定金枪鱼食物组成的唯一方法。但金枪鱼消化能力强,解剖之前胃容物消化殆尽的情况时有发生[28],且部分被捕捞的金枪鱼会出现反胃现象,将胃中的食物吐出[29-30]。因此,研究金枪鱼的食性仅用胃容物鉴定的方法不够准确。现阶段,金枪鱼食性分析将传统方法与现代技术相结合,其结果的准确性大大提高,常用的有“胃容物鉴定+稳定同位素分析”方法和“胃容物鉴定+脂肪酸分析鉴定”方法。与其他大洋性洄游鱼类(如剑鱼,旗鱼Istiophorus platypterus)相似,金枪鱼是机会捕食者,具有根据生境中饵料丰度变化调整自身捕食行为的能力。因此,不同海域的同种或不同种金枪鱼优势饵料相差较大(表2)。食性除了受海域影响外,不同金枪鱼种群之间,因生理生化特性不同,种间食性差异也较大[31]。例如,同一海域内,黄鳍金枪鱼与大眼金枪鱼优势饵料不同,二者基本不存在食性竞争,究其原因在于大眼金枪鱼比黄鳍金枪鱼具有更强的耐低温以及耐低氧能力,因此,大眼金枪鱼能在更深的海域进行捕食[31]。同样的现象也在蓝鳍金枪鱼与大眼金枪鱼之间被发现。
表 2 不同海域大眼金枪鱼,黄鳍金枪鱼及蓝鳍金枪鱼主要食物组成
有学者认为,金枪鱼可能并不是“机会捕食者”,最先提出该观点的是Chase[39],他发现洄游至新英格兰海域的蓝鳍金枪鱼数量与该海域中鲱鱼的种群资源量密切相关。Butler[40]在其他海域观察到类似现象,并推测蓝鳍金枪鱼洄游至新英格兰海域的目的是为了捕食该海域资源量丰富且高能量的鲱鱼,为自身的生长发育及繁殖提供能量,因此,他认为蓝鳍金枪鱼可能并不是完全的机会捕食者。但在对剑鱼食性深入研究后,Ménard[41]认为机会捕食者并不是指自身无“喜好”饵料的一类捕食者,而是指当“喜好”的饵料资源量匮乏时,具有转变主要饵料种类能力的一类生物,如剑鱼“喜好”捕食头足类,当头足类在当地海域资源量充足时,头足类为剑鱼的第一捕食目标,而当头足类资源量匮乏时,剑鱼则会转而寻找在该海域资源量丰富的其他鱼类或海洋生物为食。
-
金枪鱼与其他生长跨度大的鱼类(如剑鱼)相似,随着生长发育以及口器发育逐渐完善,会发生食性转变。有学者将胃含物分析(%IRI相对重要性指数百分率)和稳定同位素分析方法结合,发现台湾西南海域的黄鳍金枪鱼大约在叉长50 cm时发生食性转变[35]。叉长小于50 cm的黄鳍金枪鱼主要以虾和飞鱼幼鱼为食,叉长不小于50 cm的黄鳍金枪鱼以头足类和鲭鱼等硬骨鱼为主要饵料[35](图2)。
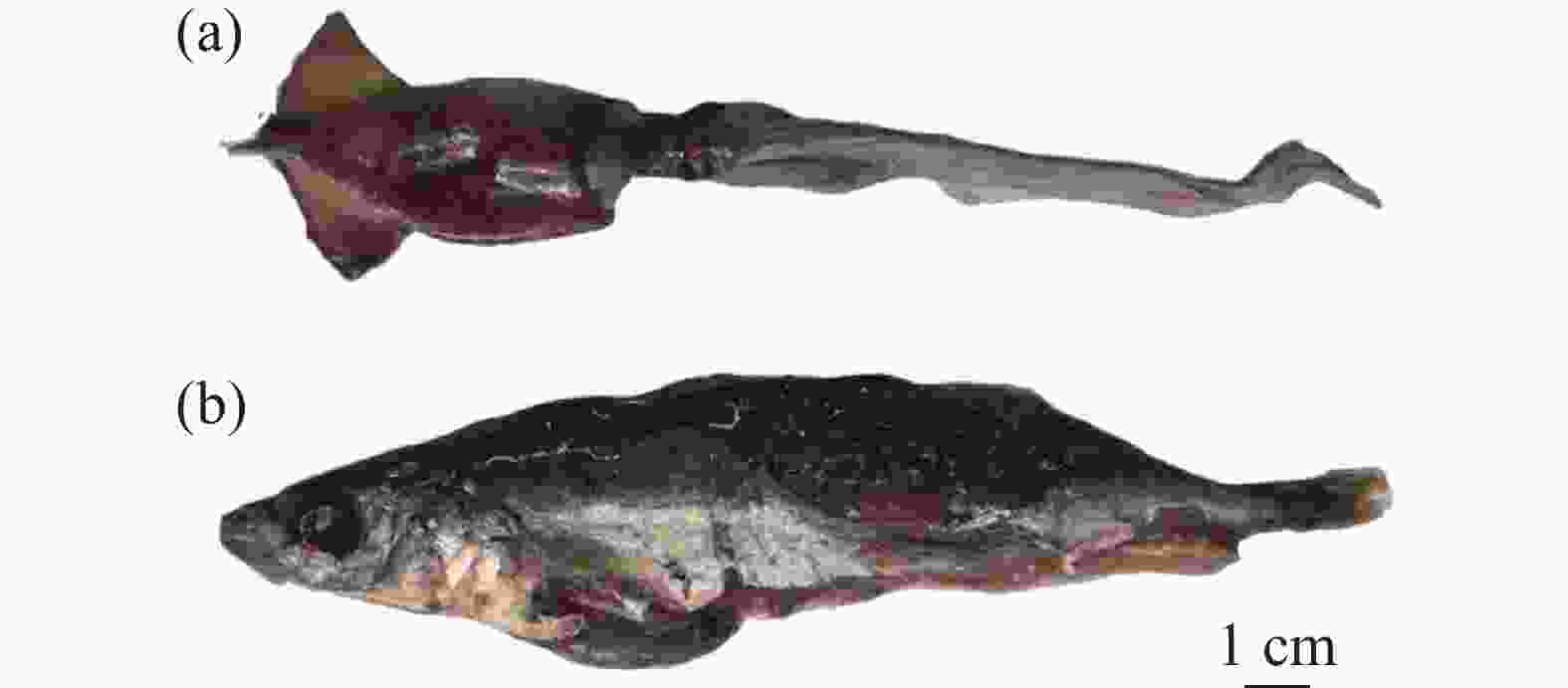
图 2 南海黄鳍金枪鱼胃容物实物图
与剑鱼相比,金枪鱼食性转变并不“彻底”。金枪鱼所捕获的猎物最大尺寸虽然随着金枪鱼自身体型的增大而增加,但成年金枪鱼的优势饵料仍与幼年金枪鱼的优势饵料存在重叠部分[41]。相比之下,体型小的剑鱼主要以海洋中上层鱼类为食,而成年后的剑鱼主要以海洋中下层的头足类为食[41],两者之间基本不存在优势饵料的重叠。导致以上区别的主要原因,可能是两者在捕食能力上存在差距。剑鱼作为游泳速度最快的鱼类(最高时速可达130 km·h−1),捕获猎物的能力远高于金枪鱼[42]。不仅如此,成年剑鱼的耐低温耐低氧能力强于成年金枪鱼,因此成年剑鱼可频繁且持久地在海洋中下层对头足类捕食,而成年金枪鱼与幼年金枪鱼均主要在海洋中上层捕食[43]。综上所述,局限于生理特性等其他因素的影响(主要是耐低温及耐低氧能力),金枪鱼成鱼与幼鱼捕食海域的空间维度存在重叠,导致金枪鱼在生长发育时虽食性发生改变,但食性转变并不“彻底”,成鱼与幼鱼之间的优势饵料仍存在重叠部分。
金枪鱼营养级随着体型增大而增加,但由于成年金枪鱼与剑鱼主要捕食空间维度不同,且海洋中下层头足类平均15N含量高于中上层硬骨鱼类,导致成年金枪鱼的平均营养级低于成年剑鱼,且金枪鱼平均营养级随体型变化的幅度小于剑鱼;体型对金枪鱼和剑鱼营养级变化的影响,剑鱼较金枪鱼更加明显[42]。
-
金枪鱼的种群性别比例与剑鱼相似,二者均不为1∶1[44-45]。金枪鱼种群内雄性多于雌性,且当叉长超过某个拐点时(如黄鳍金枪鱼的拐点为180 cm),叉长在该拐点以上的均为雄性[46]。剑鱼则与之相反,剑鱼种群内雌性多于雄性,叉长在拐点(200 cm)以上的均为雌性。金枪鱼种群中雄性多于雌性的原因可能为雌性个体性成熟后繁殖活动消耗较多能量,导致雌性性成熟个体的死亡率高于雄性[47]。剑鱼种群中雌性多于雄性的原因在于雌雄个体之间存在生长增速差异,即同龄个体中雌性剑鱼叉长大于雄性剑鱼,导致了高叉长组内无雄性剑鱼的情况发生[48]。
-
金枪鱼精巢为一对细长的性腺,呈长条状,黏附在腹腔背部。组织学观察发现,精巢是由精子发生同步进行的许多小裂片组成的外围区域和高度发达的输精管道系统组成。根据金枪鱼精巢发育的特点,将金枪鱼精巢发育分为5个阶段,分别是:未成熟期精巢呈细长带状,灰色,组织内多含精原细胞;发育期精巢增大,淡黄色,组织内仍以精原细胞为主,但出现少量精细胞;成熟期精巢肥大,纵向伸长,呈灰白色,轻压精巢可见精液流出(图3);排精期的精巢体型最大,约占腹腔的4/5,呈乳白色,轻压金枪鱼腹部可见乳白色的精液流出,组织内部充满精子以及排精后期精巢萎缩,灰白色,组织内存在大量的小裂片空腔,部分空腔还有少量残留精子[49]。
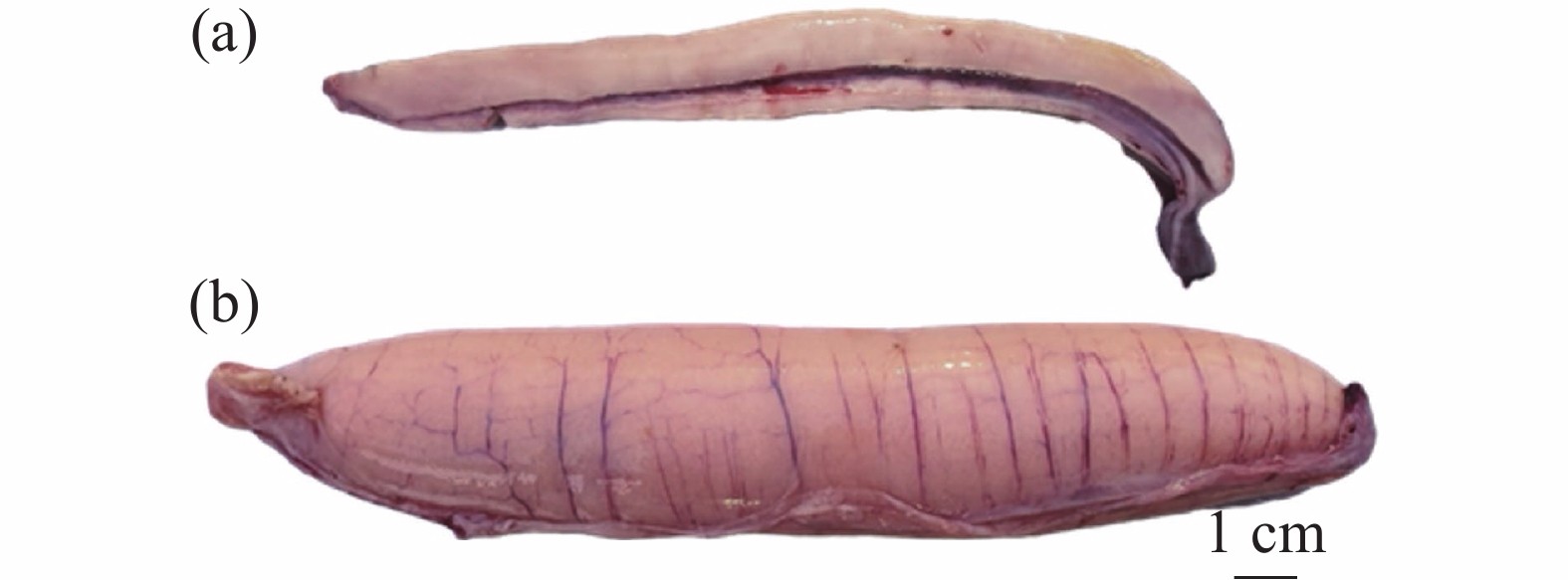
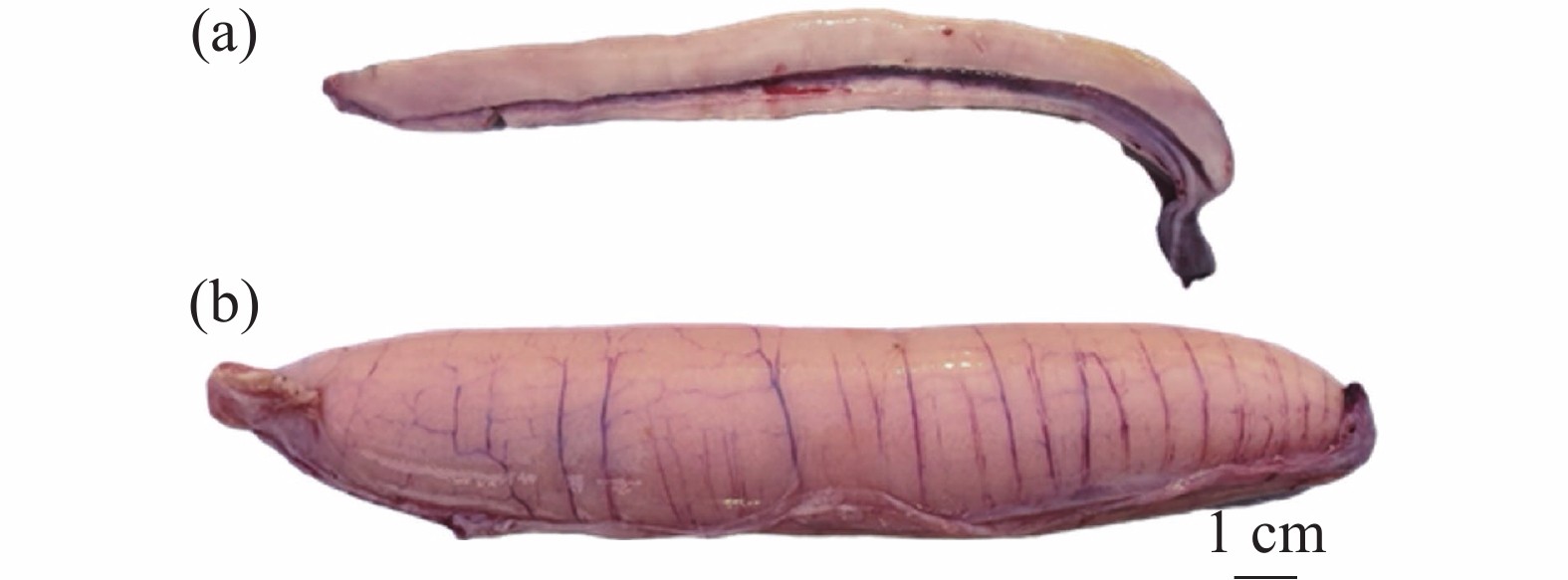
图 3 南海黄鳍金枪鱼性腺实物图
金枪鱼卵巢呈棒状(一对),由肠膜贴在腹腔背部上方左右两侧。根据卵巢的组织学特点,将金枪鱼卵巢发育分为5个时期:增值期、生长期、成熟期、排卵期及排卵后期(又称闭锁期)。各时期组织学特点分为:增殖期卵巢细长,肉眼无法看见颗粒状的卵细胞;生长期卵巢体型增大,血管明显,呈淡黄色,肉眼仍难以看见卵细胞;成熟期卵巢肥硕,组织上有明显的纵向血管,卵巢呈鲜黄色,肉眼可见明显的颗粒状卵细胞(图3);排卵期卵巢发育最大,有明显的纵向大血管,呈乳黄色,组织内充满了液化即将被排出的卵母细胞;排卵后期(闭锁期)卵巢变小,与生长期相似难以区分,组织内存在Ⅶ期卵母细胞且多有不规则空泡出现[50]。
-
组织学观察金枪鱼的卵巢发现,卵巢中卵母细胞为非同步成熟,即金枪鱼为多批次产卵鱼类,存在繁殖高峰期。一般金枪鱼的繁殖高峰为晚冬至初春(12月至翌年的1月),部分金枪鱼在晚夏(9月)也存在繁殖高峰[49]。通常金枪鱼是在凌晨产卵,2:00—3:00为产卵高峰,并且产卵经常持续到天亮时分[51]。温度是影响金枪鱼产卵的主要外界因素。一般金枪鱼产卵须表面水温在20 ℃以上,不同金枪鱼产卵温度不同,如大眼金枪鱼的产卵温度在24~28 ℃之间,而蓝鳍金枪鱼的产卵温度在21~26 ℃之间[52-53]。
几内亚湾为黄鳍金枪鱼与蓝鳍金枪鱼在大西洋西部的产卵地,地中海海域则是蓝鳍金枪鱼在大西洋东部的产卵地[1]。关于大眼金枪鱼与黄鳍金枪鱼在太平洋与印度洋的产卵地尚未有统一说法。此外,有学者认为蓝鳍金枪鱼在大西洋的产卵场不止上述两个,仍有小型或其他产卵场未被发现,值得继续勘探及研究[51]。
-
为了保证金枪鱼及其后代对生存环境有最大的适应性,金枪鱼种群在漫长的进化过程中形成了独特的繁殖策略。金枪鱼部分性成熟个体会在繁殖季节徘徊于产卵场外且不产卵或排精。在发现该特殊现象后,部分学者对此进行深入研究,发现该行为为金枪鱼的繁殖策略之一:跳过繁殖期[54]。发生“跳过繁殖期”的金枪鱼个体大多刚刚性成熟,且这一时段的金枪鱼自身机能没有达到繁殖的最佳状态[55]。通过执行跳过繁殖期的策略,刚刚性成熟的金枪鱼获得一段“繁殖空白期”,通过捕食饵料优化自身机体,提高自身的终身繁殖能力,以提高整个种群的后代存活率[55]。
除跳过繁殖期外,Reglero等[56]发现在产卵地还未达到最适产卵温度之前,部分金枪鱼提前产卵的现象。在深入研究后,他们进一步发现提前产卵时的温度虽不达最适温度的要求,但提前孵化的幼鱼所在环境中的饵料丰度远远高于之后在合适温度孵化的幼鱼所处环境中的饵料丰度,并且金枪鱼在幼鱼时期会在缺乏食物时将比自身体型较小的同类作为捕食目标。因此,“提前产卵”是金枪鱼提高后代存活率的繁殖策略之一[56]。
-
目前导致金枪鱼种群生物学相关资料相对缺乏的原因主要有2个方面:(1)金枪鱼是大洋洄游鱼类且人工养殖技术仅少数国家掌握,因此研究样本基本为野外捕获,获取研究样本困难。(2)相关研究普遍选用组织学等较为宏观的方法,无法从机理等方面深层次了解金枪鱼的特性。针对以上不足,已有学者将先进的设备及现代的研究方法运用到金枪鱼种群生物学研究中,如利用特殊装置模拟海洋环境,探究蓝鳍金枪鱼在大洋中的定位以及洄游的秘密;从分子方面研究金枪鱼的视觉系统,并与生物学分类接近的鱼类比较,发现相关关键基因并研究其分子机理,但运用上述技术以及设备研究金枪鱼种群生物学的实验室仅有少数。因此,为克服现阶段取样困难、研究方法整体水平偏低等问题,应着力更新金枪鱼研究工作者的实验设备,推动研究技术由传统组织学方法为主向现代组学等分子技术为主的转变,以此加深金枪鱼种群生物学的研究深度。
Research progress on age, growth, feeding and reproduction of bigeye tuna (Thunnus obesus), yellowfin tuna (Thunnus albacores) and bluefin tuna (Thunnus thynnus)
-
摘要: 以大眼金枪鱼Thunnus obesus,黄鳍金枪鱼Thunnus albacares及蓝鳍金枪鱼Thunnus maccoyii为例,围绕金枪鱼生物学中的鱼龄与生长、摄食及繁殖3个方面,综述了现阶段金枪鱼生物学研究的常用方法与主要的研究成果,即金枪鱼为提高种群的合适度,形成的多个独特的生物学特征,如多生长增速拐点、机会捕食者及多种繁殖策略等。同时,针对金枪鱼生物学研究领域现存的不足,从研究方法、设备和技术手段等方面提出未来研究建议。Abstract: Tunas are species of bony fish belonging to the family Scombridae, order Perciformes, and class Osteichthyes. They are high trophic predators with marine migratory behavior. Tunas have acquired various distinct biological traits in response to the pressures of human overfishing and the variable marine environments. Focused on Atlantic bluefin tuna (Thunnus thynnus), bigeye tuna (Thunnus obesus), and yellowfin tuna (Thunnus albacores), the most popular research methodologies and important research results in tuna biology, including age and growth, feeding, and reproduction are reviewed. The unique biological properties of tunas to boost the population's fitness (e.g., multiple growth inflection points, opportunistic predators, and multiple reproductive strategies, etc.) are illustrated. At the same time, the existing shortcomings in the field of tuna biology research are analyzed, based on which suggestions for future research, regarding research methods, equipment and technologies are hence proposed.
-
Key words:
- tuna /
- age and growth /
- feeding biology /
- reproductive biology
-
表 1 不同海域von Bertalanffy生长模型拟合大眼金枪鱼、黄鳍金枪鱼及蓝鳍金枪鱼生长参数
种类 捕捞方式 海域 生长系数K 渐进体长/cm 生长起点鱼龄t0 肥满度a(×10−4) 幂指数b 参考文献 大眼金枪鱼 延绳钓 太平洋 0.18 178.41 −2.50 − − [17] 大眼金枪鱼 延绳钓 太平洋 0.20 208.70 −0.99 − − [18] 大眼金枪鱼 延绳钓 太平洋 − − − 5.15 2.76 [19] 大眼金枪鱼 延绳钓 大西洋 0.12 264.02 −0.68 − − [20] 大眼金枪鱼 延绳钓 印度洋 0.13 207.14 −1.89 − − [8] 黄鳍金枪鱼 延绳钓 太平洋 0.25 181.81 −0.48 5.55 2.72 [13] 黄鳍金枪鱼 延绳钓 太平洋 − − − 2.19 2.94 [21] 黄鳍金枪鱼 延绳钓 大西洋 0.27 230.70 −0.08 − − [22] 黄鳍金枪鱼 延绳钓 大西洋 − − − 0.85 3.14 [23] 黄鳍金枪鱼 延绳钓 印度洋 0.18 272.70 −0.27 1.59 3.30 [24] 蓝鳍金枪鱼 延绳钓 大西洋 314.90 0.09 −0.15 − − [25] 蓝鳍金枪鱼 延绳钓 大西洋 304.00 0.10 −0.65 − − [26] 蓝鳍金枪鱼 延绳钓 大西洋 373.08 0.07 −1.76 − − [27]  下载: 导出CSV
下载: 导出CSV
-
[1] BLOCK B A, DEWAR H, BLACKWELL S B, et al. Migratory movements, depth preferences, and thermal biology of Atlantic bluefin tuna [J]. Science, 2001, 293(5533): 1310 − 1314. doi: 10.1126/science.1061197 [2] 李桂芬, 乐建盛. 金枪鱼的营养功效与开发加工[J]. 食品科技, 2003(9): 41 − 44. doi: 10.3969/j.issn.1005-9989.2003.09.016 [3] API M, BONFANTI E, LOMBARDO F, et al. Effects of age on growth in Atlantic bluefin tuna (Thunnus thynnus) [J]. General and Comparative Endocrinology, 2018, 265: 64 − 70. doi: 10.1016/j.ygcen.2018.01.010 [4] 郑晓春. 东太平洋大眼金枪鱼摄食生态及其栖息环境研究[D]. 上海: 上海海洋大学, 2014. [5] LUTCAVAGE M E, BRILL R W, SKOMAL G B, et al. Results of pop-up satellite tagging of spawning size class fish in the gulf of Maine: do north Atlantic bluefin tuna spawn in the mid-Atlantic? [J]. Canadian Journal of Fisheries and Aquatic Sciences, 1999, 56(2): 173 − 177. doi: 10.1139/f99-016 [6] TSERPES G, TSIMENIDES N. Determination of age and growth of swordfish, Xiphias gladius L., 1758, in the eastern Mediterranean using anal-fin spines [J]. Fishery Bulletin, 1995(93): 594 − 602. [7] 周成, 朱国平, 陈锦淘, 等. 印度洋南部大眼金枪鱼年龄鉴定及其与生长的关系[J]. 中国水产科学, 2012, 19(3): 536 − 544. [8] 李凤莹. 印度洋中南部大眼金枪鱼年龄与生长研究[D]. 上海: 上海海洋大学, 2014. [9] 吕凯凯. 马绍尔群岛海域大眼金枪鱼(Thunnus obesus)年龄与生长研究[D]. 上海: 上海海洋大学, 2011. [10] 李军, 李志凌, 叶振江. 大眼金枪鱼渔业现状和生物学研究进展[J]. 齐鲁渔业, 2005(12): 35 − 38. [11] 颜斐斐, 唐建业. 基于将大西洋蓝鳍金枪鱼列入 CITES 附录 I 提案的国际渔业管理分析[J]. 上海海洋大学学报, 2012, 21(5): 854 − 862. [12] LEHODEY P, HAMPTON J, LEROY B. Preliminary results on age and growth of bigeye tuna (Thunnus obesus) from the western and central Pacific Ocean as indicated by daily growth increments and tagging data, 1999[C], Standing Committee on Tuna and Billfish, 1999. [13] 赵海龙. 太平洋中部黄鳍金枪鱼耳石形态及微量元素[D]. 上海: 上海海洋大学, 2016. [14] LEHODEY P, LEROY B. Age and growth of yellowfin tuna (Thunnus albacares) from the western and central pacific ocean as indicated by daily growth increments and tagging data, 1999[C], Standing Committee on Tuna and Billfish, 1999. [15] DIAZ G A. A revision of western Atlantic bluefin tuna age of maturity derived from size samples collected by the Japanese longline fleet in the gulf of Mexico (1975-1980) [J]. ICCAT, 2010, 66(3): 1216 − 1226. [16] AILLOUD L E, LAURETTA M V, HANKE A R, et al. Improving growth estimates for Western Atlantic bluefin tuna using an integrated modeling approach [J]. Fisheries Research, 2017, 191: 17 − 24. doi: 10.1016/j.fishres.2017.02.013 [17] FARLEY J H, CLEAR N P, LEROY B, et al. Age, growth and preliminary estimates of maturity of bigeye tuna, Thunnus obesus, in the Australian region [J]. Marine and Freshwater Research, 2006, 57(7): 713. doi: 10.1071/MF05255 [18] SUN C, HUANG C, YEH S. Age and growth of the bigeye tuna, Thunnus obesus, in the western Pacific Ocean [J]. Fishery Bulletin (Washington, D. C. ), 2001, 99(3): 502. [19] 党莹超. 北太平洋大眼金枪鱼摄食生态及其栖息环境的研究[D]. 上海: 上海海洋大学, 2020. [20] ALVES A, DE BARROS P, PINHO M R. Age and growth studies of bigeye tuna Thunnus obesus from Madeira using vertebrae [J]. Fisheries Research, 2002, 54(3): 389 − 393. doi: 10.1016/S0165-7836(01)00268-5 [21] 冯波, 李忠炉, 侯刚. 南海大眼金枪鱼和黄鳍金枪鱼生物学特性及其分布[J]. 海洋与湖沼, 2014, 45(4): 886 − 894. doi: 10.11693/hyhz20130600081 [22] LESSA R, DUARTE-NETO P. Age and growth of yellowfin tuna (Thunnus albacares) in the western equatorial Atlantic, using dorsal fin spines [J]. Fisheries Research, 2004, 69(2): 157 − 170. doi: 10.1016/j.fishres.2004.05.007 [23] 宋利明, 陈新军, 许柳雄. 大西洋中部金枪鱼延绳钓渔场黄鳍金枪鱼(Thunnus albacares)生物学特性的初步研究[J]. 海洋与湖沼, 2004(6): 538 − 542. doi: 10.3321/j.issn:0029-814X.2004.06.009 [24] STéQUERT B, PANFILI J, DEAN J M. Age and growth of yellowfin tuna, Thunnus albacares, from the western Indian Ocean, based on otolith microstructure [J]. Oceanographic Literature Review, 1996, 12(43): 1275. [25] RESTREPO V R, DIAZ G A, WALTER J F, et al. Updated estimate of the growth curve of Western Atlantic bluefin tuna [J]. Aquatic Living Resources, 2010, 23(4): 335 − 342. doi: 10.1051/alr/2011004 [26] NEILSON J D, CAMPANA S E. A validated description of age and growth of western Atlantic bluefin tuna (Thunnus thynnus) [J]. Canadian Journal of Fisheries and Aquatic Sciences, 2008, 65(8): 1523 − 1527. doi: 10.1139/F08-127 [27] SANTAMARIA N, BELLO G, CORRIERO A, et al. Age and growth of Atlantic bluefin tuna, Thunnus thynnus (Osteichthyes: Thunnidae), in the Mediterranean Sea [J]. Journal of Applied Ichthyology, 2009, 25(1): 38 − 45. doi: 10.1111/j.1439-0426.2009.01191.x [28] PLEIZIER N K, CAMPANA S E, SCHALLERT R J, et al. Atlantic bluefin tuna (Thunnus thynnus) diet in the gulf of St. Lawrence and on the eastern Scotian Shelf [J]. Journal of Northwest Atlantic Fishery Science, 2012, 44: 67 − 76. doi: 10.2960/J.v44.m685 [29] SORELL J M, VARELA J L, GOñI N, et al. Diet and consumption rate of Atlantic bluefin tuna (Thunnus thynnus) in the strait of Gibraltar [J]. Fisheries Research, 2017, 188: 112 − 120. doi: 10.1016/j.fishres.2016.12.012 [30] LOGAN J M, RODRíGUEZ-MARíN E, GOñI N, et al. Diet of young Atlantic bluefin tuna (Thunnus thynnus) in eastern and western Atlantic foraging grounds [J]. Marine Biology, 2011, 158(1): 73 − 85. doi: 10.1007/s00227-010-1543-0 [31] ARTETXE-ARRATE I, FRAILE I, MARSAC F, et al. A review of the fisheries, life history and stock structure of tropical tuna (skipjack Katsuwonus pelamis, yellowfin Thunnus albacares and bigeye Thunnus obesus) in the Indian Ocean [J]. Advances in Marine Biology, 2021, 88: 39 − 89. [32] MOTEKI M, ARAI M, TSUCHIYA K, et al. Composition of piscine prey in the diet of large pelagic fish in the eastern tropical Pacific Ocean [J]. Fisheries Science, 2001, 67(6): 1063 − 1074. doi: 10.1046/j.1444-2906.2001.00362.x [33] 朱国平, 许柳雄, 周应祺, 等. 印度洋中西部和大西洋西部水域大眼金枪鱼的食性比较[J]. 生态学报, 2007(1): 135 − 141. doi: 10.3321/j.issn:1000-0933.2007.01.016 [34] VARELA J L, INTRIAGO K M, FLORES J C, et al. Feeding habits of juvenile yellowfin tuna (Thunnus albacares) in Ecuadorian waters assessed from stomach content and stable isotope analysis [J]. Fisheries Research, 2017, 194: 89 − 98. doi: 10.1016/j.fishres.2017.05.017 [35] WENG J S, LEE M A, LIU K M, et al. Feeding ecology of juvenile yellowfin tuna from waters southwest of taiwan inferred from stomach contents and stable isotope analysis [J]. Marine and Coastal Fisheries, 2015, 7(1): 537 − 548. doi: 10.1080/19425120.2015.1094157 [36] 朱国平, 许柳雄, 周应祺, 等. 印度洋中西部水域黄鳍金枪鱼的食性及其季节性变化[J]. 水产学报, 2008(5): 725 − 732. [37] ORTEGA-GARCíA S. Activity of the Mexican purse seine fleet and the feeding habits of yellowfin tuna [J]. Ciencias Marinas, 1992, 18(1): 139 − 149. doi: 10.7773/cm.v18i1.874 [38] BATTAGLIA P, ANDALORO F, CONSOLI P, et al. Feeding habits of the Atlantic bluefin tuna, Thunnus thynnus (L. 1758), in the central Mediterranean Sea (Strait of Messina) [J]. Helgoland Marine Research, 2013, 67(1): 97 − 107. doi: 10.1007/s10152-012-0307-2 [39] CHASE B C. Differences in diet of Atlantic bluefin tuna at five seasonal feeding grounds on the New England continental shelf [J]. Fishery Bulletin (Washington, D. C. ), 2002, 100(2): 168. [40] BUTLER C M, LOGAN J M, PROVAZNIK J M, et al. Atlantic bluefin tuna Thunnus thynnus feeding ecology in the northern gulf of Mexico: a preliminary description of diet from the western Atlantic spawning grounds [J]. Journal of Fish Biology, 2015, 86(1): 365 − 374. doi: 10.1111/jfb.12556 [41] MéNARD F, LORRAIN A, POTIER M, et al. Isotopic evidence of distinct foraging ecology and movement pattern in two migratory predators (yellowfin tuna and swordfish) of the western Indian Ocean [J]. Marine Biology, 2007, 153(2): 141 − 152. doi: 10.1007/s00227-007-0789-7 [42] YOUNG J, LANSDELL M, RIDDOCH S, et al. Feeding ecology of broadbill swordfish, Xiphias gladius, off eastern Australia in relation to physical and environmental variables [J]. Bulletin of Marine Science, 2006, 79(3): 793 − 809. [43] YOUNG J W, GUEST M A, LANSDELL M, et al. Discrimination of prey species of juvenile swordfish Xiphias gladius (Linnaeus, 1758) using signature fatty acid analyses [J]. Progress in Oceanography, 2010, 86(1-2): 139 − 151. doi: 10.1016/j.pocean.2010.04.028 [44] ZHU G, DAI X, XU L, et al. Reproductive biology of bigeye tuna, Thunnus obesus, (Scombridae) in the eastern and central tropical Pacific Ocean [J]. Environmental Biology of Fishes, 2010, 88(3): 253 − 260. doi: 10.1007/s10641-010-9636-7 [45] 朱国平, 许柳雄. 印度洋中西部大眼金枪鱼繁殖生物学的初步研究[J]. 水产科学, 2006(7): 364 − 366. doi: 10.3969/j.issn.1003-1111.2006.07.011 [46] WILD A. A review of the biology and fisheries for yellowfin tuna, Thunnus albacares, in the eastern Pacific Ocean [J]. Fao Fisheries Technical Paper, 1994, 37(4): 732 − 736. [47] 李攀, 陈锦淘, 朱国平, 等. 印度洋中南部大眼金枪鱼生物学特性研究[J]. 海洋渔业, 2010, 32(3): 283 − 288. doi: 10.3969/j.issn.1004-2490.2010.03.008 [48] ALıÇLı T Z. Age, sex ratio, length-weight relationships and reproductive biology of Mediterranean swordfish, Xiphias gladius L. , 1758, in the eastern Mediterranean [J]. African Journal of Biotechnology, 2012, 11(15). [49] 王修国. 东太平洋大眼金枪鱼、黄鳍金枪鱼生殖特征研究[D]. 上海: 上海海洋大学, 2010. [50] 翟天晨. 南太平洋雌性大眼金枪鱼繁殖生物学研究[D]. 上海: 上海海洋大学, 2015. [51] KARAKULAK S, ORAY I, CORRIERO A, et al. Evidence of a spawning area for the bluefin tuna (Thunnus thynnus L.) in the eastern Mediterranean [J]. Journal of Applied Ichthyology, 2004, 20(4): 318 − 320. doi: 10.1111/j.1439-0426.2004.00561.x [52] MEDINA A. Reproduction of Atlantic bluefin tuna [J]. Fish and Fisheries, 2020, 21(6): 1109 − 1119. doi: 10.1111/faf.12489 [53] ZOHAR Y, MYLONAS C C, ROSENFELD H, et al. Reproduction, broodstock management, and spawning in captive Atlantic bluefin tuna[M]//Benetti D D, Partridge G J, Buentello A. Advances in Tuna Aquaculture. San Diego: Academic Press, 2016: 159-188. [54] JORGENSEN C, ERNANDE B, FIKSEN O, et al. The logic of skipped spawning in fish [J]. Canadian Journal of Fisheries and Aquatic Sciences, 2006, 63(1): 200 − 211. doi: 10.1139/f05-210 [55] GALUARDI B, ROYER F, GOLET W, et al. Complex migration routes of Atlantic bluefin tuna (Thunnus thynnus) question current population structure paradigm [J]. Canadian Journal of Fisheries and Aquatic Sciences, 2010, 67(6): 966 − 976. doi: 10.1139/F10-033 [56] REGLERO P, URTIZBEREA A, TORRES A P, et al. Cannibalism among size classes of larvae may be a substantial mortality component in tuna [J]. Marine Ecology Progress Series, 2011, 433: 205 − 219. doi: 10.3354/meps09187 -

 点击查看大图
点击查看大图
图(3) / 表 (2)
计量
- 文章访问数: 834
- HTML全文浏览量: 378
- PDF下载量: 65
- 被引次数: 0
