



-
木麻黄(Casuarina equisetifolia)是属于双子叶植物纲(Dicotyledoneae)、木麻黄科(Casuarinaceae)、木麻黄属(Casuarina)的一类绿化常绿乔木,在中国广东、广西、海南等省区均有栽培。由于木麻黄具有速生、抗风、固沙、耐盐、耐干旱、耐瘠薄等特点,故成为我国南方滨海防风固沙不可缺少的一类优良树种[1]。海南省于1955年开始建设包括木麻黄林在内的沿海防护林。目前,海南全境木麻黄纯林与混交林总面积超过5万hm2,分布在海口、儋州、文昌、万宁等沿海12个市县的73个乡镇[2]。木麻黄海防林的建设在抵御台风,保持水土,改善自然生态,促进经济建设上均发挥了举足轻重的作用[3]。据报道,木麻黄病害目前总计有14种,分为非侵染性病害、细菌性病害、真菌性病害与其他病害。其中非侵染性病害中的衰退病与细菌性病害中的青枯病为主要病害,其余均为真菌性病害[4]。海南沿海种植多年的木麻黄海防林,由于其茎干易受到台风、害虫的危害,造成大量伤口,故易被病原木腐真菌侵染定殖而发生真菌性根腐和茎腐,甚至部分重病株整株死亡[5]。在木麻黄灵芝病害方面,陈礼浪与吴如慧[6-8]分别鉴定木麻黄红根病和茎腐病病原菌为热带灵芝(Ganoderma tropicum)与二孢假芝(Amauroderma subresinosum)并进行了生物学特性测定;单金雪[9]发现南方灵芝(Ganoderma australe)能引起木麻黄茎腐病并完成了其生物学特性研究。笔者在海南省昌江县棋子湾海滨木麻黄林中发现独体木麻黄活树上有一种灵芝菌引起的茎腐病,田间发病率2%,重病株整株枯死,此灵芝菌的担子果有明显的同心环纹与漆样光泽,与报道的热带灵芝和南方灵芝均不相同,而且经初步镜检发现其担孢子形态也不同于热带灵芝等已知种,推测可能为狭长孢灵芝(Ganoderma boninense Pat.)。狭长孢灵芝是一类在油棕上引起严重破坏性茎基腐病的重要灵芝病原菌[10],近年来在海南与台湾的槟榔上也有发现[11]。程乐乐等[7]和陈礼浪等[12]在调查海南棕榈科植物木腐菌时曾在槟榔活树、椰子和木麻黄死树上发现狭长孢灵芝。目前,还未见狭长孢灵芝引起的木麻黄活立木茎腐病的详细报道。本研究从木麻黄活树茎基部进行取样,对病害灵芝菌样本进行病原菌分离、鉴定和致病性测定,并对其生物学特性进行了研究,为进一步研究此类病菌及生产上茎腐病的防治提供参考。
-
样本于2018年1月采自海南省昌江县昌化镇棋子湾发病木麻黄活立木,担子果编号为HNCJ201801001。用保鲜袋包装样本并带回实验室备用。
-
马铃薯葡萄糖琼脂(PDA)、Richard、燕麦(OMA)、马铃薯蔗糖琼脂(PSA)、玉米(CMA)培养基的配置参考文献[13]的方法,木麻黄茎干煎汁培养基的配置参考文献[14]的方法,木屑培养基的配置参考文献[15]的方法。
-
OMEGA FungalDNA Kit购自美国OMEGA公司, DL2000 DNAMarker购自北京庄盟国际生物基因科技有限公司。2xSanTaq-Mix购自生工生物股份有限公司。摄影生物显微镜为日本Olympus BX-51。PCR 产物测序与相关引物的合成由铂尚生物技术有限公司完成。引物序列见表1。
-
采用组织分离法,在新鲜担子果表面用含70%酒精的医用无菌棉擦拭、消毒,再用无菌的手术刀切去表层,切取5 mm×5 mm的内层菌肉放到PDA 培养基上,于28 ℃恒温箱中培养一段时间后,对生长出来的形态符合的菌株进行纯化,将获得的编号为HNCJ201801001菌株转入PDA培养基于28 ℃黑暗条件下扩繁,保存和备用。
-
从长有HNCJ201801001菌株的PDA培养基上切取1 cm×1 cm的菌丝块,放置在高压蒸汽灭菌后的木屑培养基中,透明胶带封口后在黑暗常温状态下培养21 d,当整个培养基表面布满菌丝,可用于致病性测定。对照组采用接入空白PDA培养基块的木屑培养基,其他条件与处理方式均与处理组接种体相同。
-
对待接种木麻黄幼树的茎基部用70%酒精进行消毒,再用手术刀削开表皮将制备好的接种体贴在切口上,在下方用无菌水润湿过的棉花保湿,最后用保鲜袋与胶带进行固定。处理组设置5个重复,对照组使用空白木屑培养基。完成后定期浇水,定期观察苗木长势并拍照。
-
将木屑培养基装满300 mL组培瓶并灭菌,挑取分离到的菌株放入组培瓶中28 ℃黑暗培养,待组培瓶表面长满菌丝时开盖,放入盛有无菌水的玻璃瓶中进行诱导,每隔10 d观察灵芝长势并拍照。
-
(1)形态鉴定:参考《中国真菌志》[18]与《中国大型真菌》[19]等文献,对担子果进行宏观形态鉴定。切取担子果内部组织,制片,用光学显微镜进行显微观察,测量并拍照。(2)分子鉴定:参考邢佳慧[20]、袁滨[21]、OGHENEKARO[22]文献,选用并合成用于扩增ITS基因与LSU基因的引物。对纯化后的HNCJ201801001菌株在PDA培养基中进行预培养,5 d后收集菌丝体,使用OMEGA Fungal DNA Kit 提取其总DNA。用合成的2对引物扩增ITS与LSU基因。PCR产物经电泳检测和纯化回收后送往公司测序。将获得的测序结果在NCBI上进行比对并提交NCBI。
在GenBank上搜寻灵芝与其他属中同时含有ITS与LSU基因的菌株,下载其ITS与LSU序列并用Sequence Matrix软件进行拼接,用MEGA7.0对包含HNCJ201801001在内的所有序列通过NJ法构建进化树。
-
将HNCJ201801001菌株在PDA培养基上28 ℃培养5 d,然后在菌落边缘打取直径5 mm的菌饼,接种到各组的培养基上,每组设3个重复。每组除所研究变量外,其他培养条件组均统一设为28 ℃,24 h黑暗,PDA培养基。5 d后使用十字交叉法测量菌落直径。试验组条件设置:
(1)温度:设置15、20、25、28、30、32、35、37,40 ℃共9个处理组。
(2)光照:设置24 h黑暗、24 h光照、12 h光暗交替3个处理组。
(3)pH值:用NaOH与HCl将培养基pH依次调节到2~10,共9个处理组。
(4)培养基:设置PDA、PSA、Ricards、燕麦(OMA)、玉米(CMA)和木麻黄茎干煎汁培养基6个处理组。
研究取得的数据用Excel软件统计记录,分析并生成图表。使用SAS8.1软件处理数据。
-
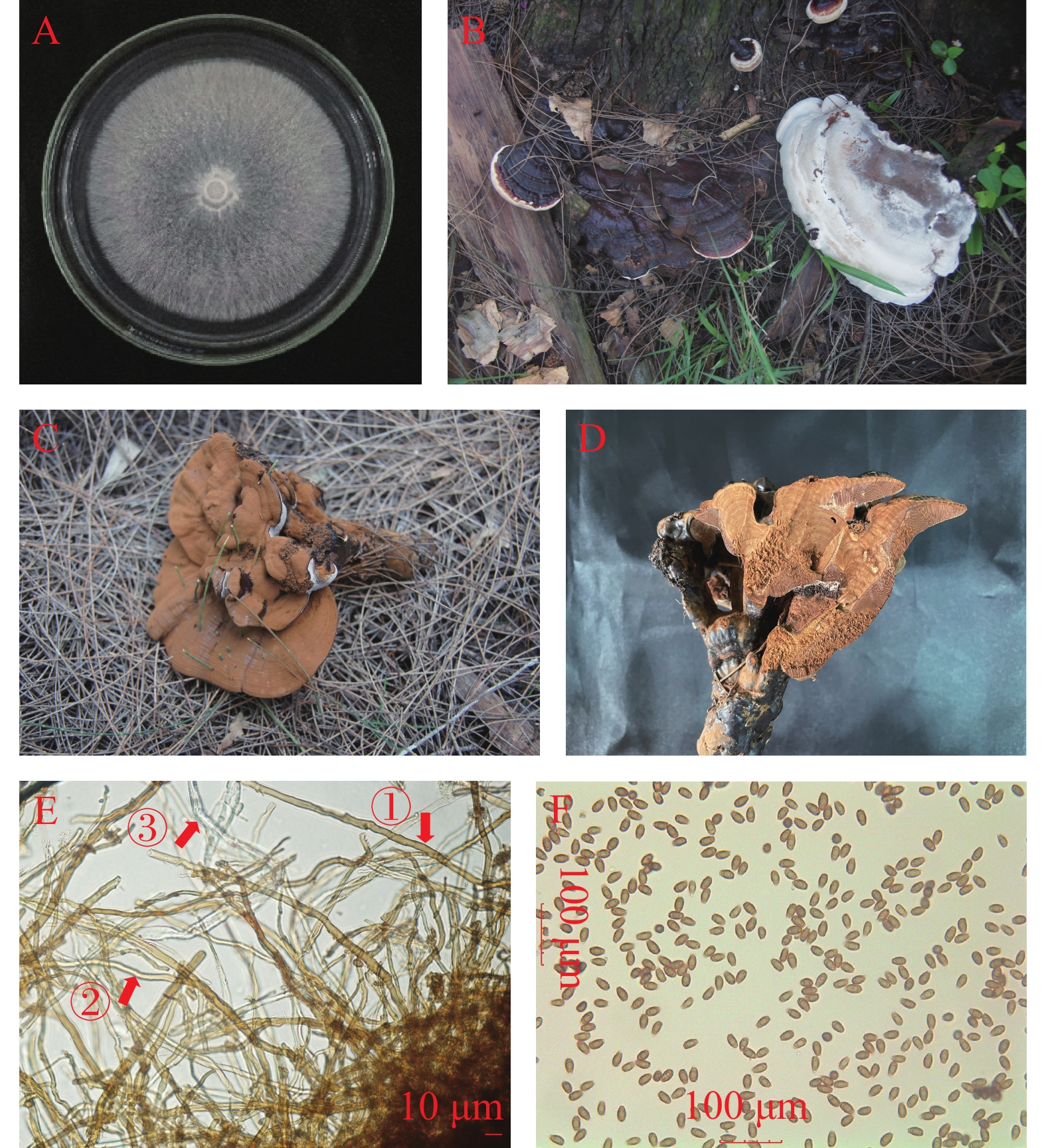
田间调查中发现,木麻黄活树受侵染后,叶片失绿无光泽,继而发黄脱落,部分小枝条枯死,树冠稀疏,生长不良,重病植株在3~5 a内整株枯死。可以观察到红褐色或漆黑色、有柄和无柄的担子果在病树的下部茎干上和地面附近的茎基部长出,产生担孢子的担子果表覆盖1层土褐色的担孢子粉,担子果上表面有明显的细密同心环纹与漆样光泽(图1-A,B,C,D,E)。病树茎基部木质部组织白腐,病株在田间发病率为2%左右。田间发病植株的地上部分症状与热带灵芝(G. tropicum)引起的红根病、南方灵芝(G. australe)和二孢假芝(Amauroderma subresinosum)引起的茎腐病症状相似,均会呈现树冠稀疏、生长不良等的相同症状,主要区别在于病树茎干基部生长出的病菌担子果形态不同[7-9]。

图 1 木麻黄茎腐病田间发病症状
-
接种60 d后,检查HNCJ201801001菌株对木麻黄树接种部位的侵染的情况,发现木麻黄接种处长出白色菌丝,病原菌已经成功侵染植株。120 d后,组织白色腐朽,木麻黄顶部枝叶开始发黄。5个月后,接种的木麻黄幼树树冠稀疏,生长发育不良,8个月后整株枯死,与田间症状基本一致,对照组无明显变化。在病株接种部位再分离得到相同的菌株,显示所分离的HNCJ201801001菌株为木麻黄茎腐病的致病菌。将HNCJ201801001菌株接入装满木屑培养基的350 mL组培瓶中,30 d后菌丝长满整个培养基,120 d后长出与病原样本的形态相一致的担子果(图2-A,B,C,D)。

图 2 木麻黄茎腐病病原菌的致病性测定
-
(1)菌落特征:菌株生长呈圆形,5 d内可以生长至直径8 cm。菌丝纯白色,菌丝浓密,平伏生长,无气味,老化后转为淡黄色,边缘整齐(图3-A)。(2)担子果:担子果一年生,有柄或无柄,新鲜担子果菌肉呈软木栓质,干燥后转为木质。菌盖扇形,表面基部中央凸起或叠生一至多个小扇形担子果,具有漆状光泽,自基部向外有明显的放射状皱纹和致密的环纹,幼小子实体浅黄色,成熟后表面从橙色到淡红褐色,老化后为非常暗褐色,大多数漆黑色;菌盖边缘薄或厚钝,边缘厚0.2~0.7 cm,幼时奶油色,随着担子果成熟与老化逐渐变为黄褐色与紫褐色;下表面的近基部呈灰褐色,靠菌盖边缘处渐呈灰白色。担子果大小为(14~18)cm×(9~12)cm,基部厚2~5 cm,柄粗1.8~2.7 cm,柄长5.5~6.8 cm;菌肉不分层,呈褐色,接近菌管处为深褐色,担子果中存在有黑色壳质层结构[18](图3-B,C,D)。

图 3 木麻黄灵芝茎腐病病原菌形态与显微结构
(3)显微特征:从自然寄主上生长的担子果上切取菌组织和挑取担孢子粉进行镜检。菌丝三体型,骨架菌丝厚壁实心,呈树枝状,直径3~5 µm。缠绕菌丝无色,厚壁,直径1~2 µm。生殖菌丝透明薄壁,直径4~7 µm。担孢子为狭长的卵圆形,部分顶端平截,双层壁,外壁平滑,无色透明,内壁有小刺,浅黄褐色,部分有油滴。大小为(8.9 ~ 11.7)µm×(4.2 ~ 5.5)µm,平均长为10.15 µm,平均宽为5.01 µm,长宽比Q=1.679 ~ 2.373(图3-E,F)。
-
测序结果显示菌株HNCJ201801001的rDNA-ITS序列为609 bp(序列号:OK643485.1),LSU序列为590 bp(序列号:OK643883.1)。将2种基因拼接后与1.8中获得的序列进行联合建树(表2)。结果表明,HNCJ201801001菌株与狭长孢灵芝遗传距离最小聚为一类(图4)。结合形态学分析结果,将引起海南昌江木麻黄茎腐病的病原灵芝菌鉴定为狭长孢灵芝(Ganoderma boninense Pat.)。
表 2 用于分子鉴定的菌株及其ITS与LSU基因登录号
种名 菌株号 ITS基因 LSU基因 Ganoderma australe ZRL20151500 LT716076.1 KY418900.1 Ganoderma gibbosum Pvc62 MK280717.1 MK215785.1 Ganoderma adspersum MFLU 19-2220 MN396655.1 MN428663.1 Ganoderma lingzhi HFJAU0581 MH294333.1 MH294366.1 Lentinus tigrinus CBS 249.39 MH856001.1 MH867501.1 Ganoderma lucidum GL-1 GU726919.1 MH873099.1 Ganoderma boninense WD 2028 KJ143905.1 KU220015.1 Ganoderma tsunodae WD2034 AB588989.1 AB368069.1 Ganoderma resinaceum CBS 220.36 MH855781.1 MH867290.1 Ganoderma nasalanense LPDR17060211 MK345441.1 MK346831.1 Ganoderma nasalanense LPDR17060212 MK345442.1 MK346832.1 Haddowia longipes LPDR17072708 MK345423.1 MK346828.1 Ganoderma orbiforme URM 86213 MK119829.1 MK119909.1 Ganoderma ecuadoriense URM 89449 MK119828.1 MK119908.1 
图 4 HNCJ201801001菌株与灵芝属相关种基于ITS与LSU基因构建的系统发育树
-
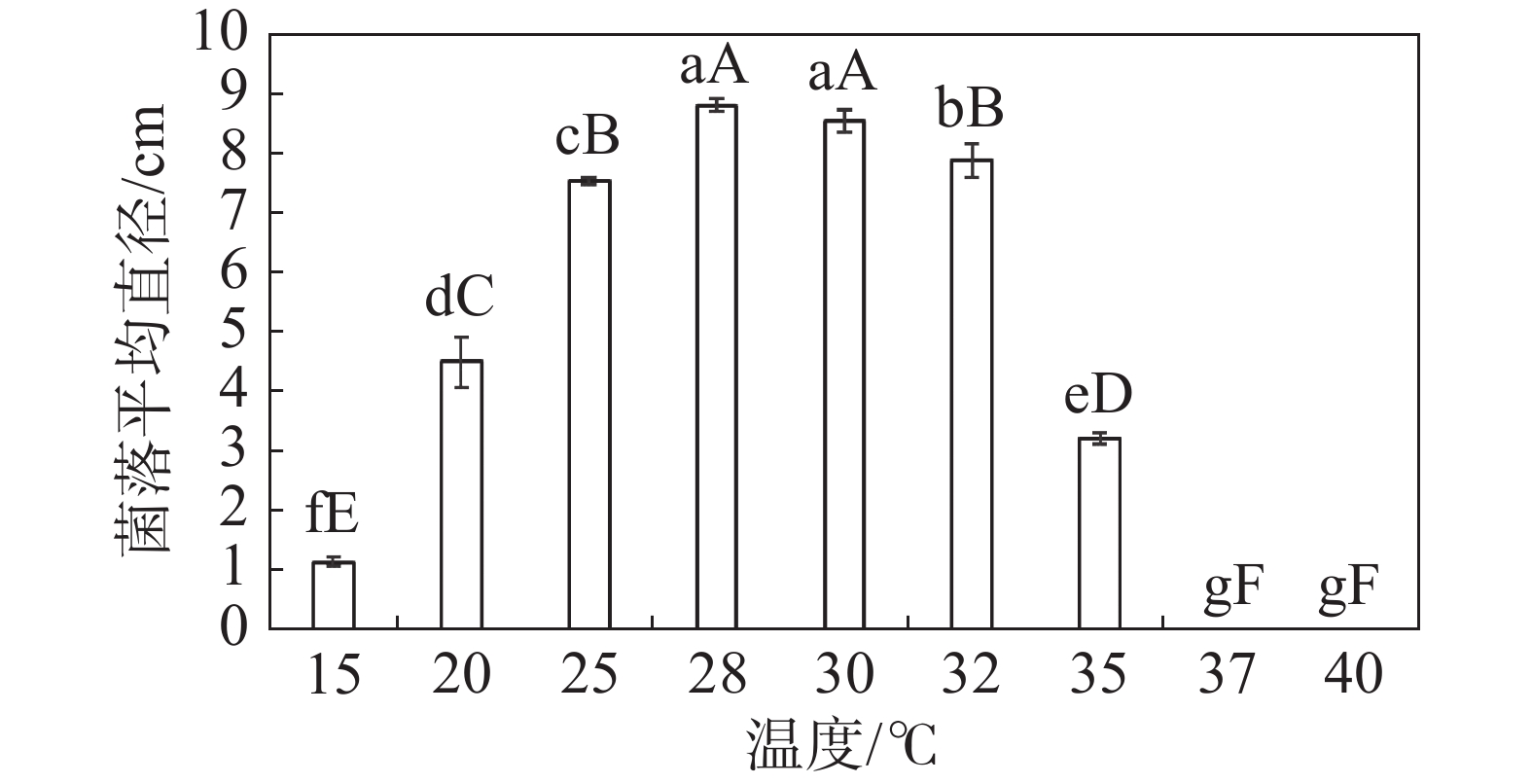
在不同温度下,HNCJ201801001 菌株生长情况差异显著。HNCJ201801001菌株在15~35 ℃的温度条件下均可生长,适宜生长温度为25~32 ℃,其中28 ℃时生长最好。37 ℃时病原菌完全停止生长,由此可得该病菌最适生长温度为28 ℃(图5)。
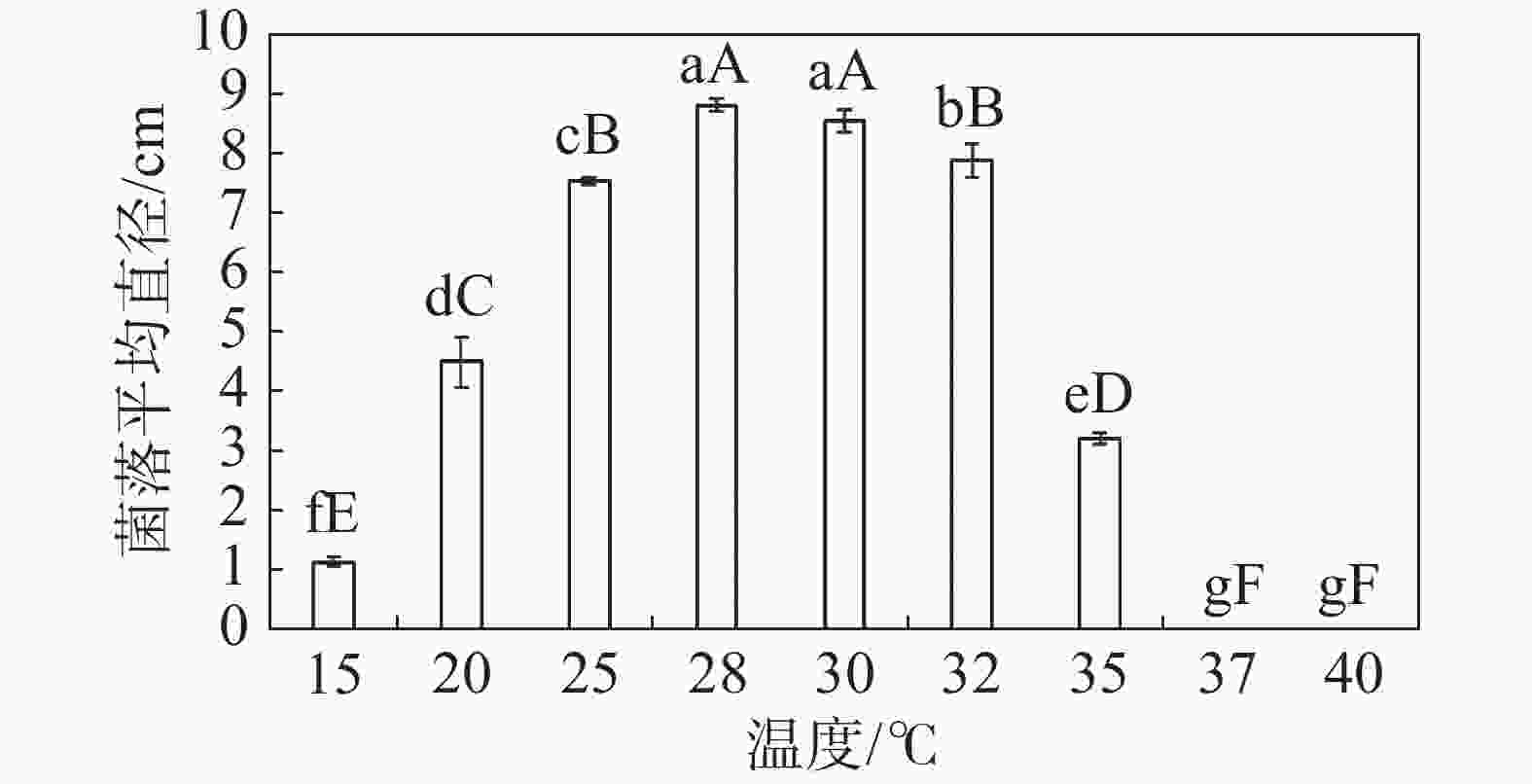
图 5 不同温度对木麻黄茎腐病病原菌HNCJ201801001菌丝生长的影响
-
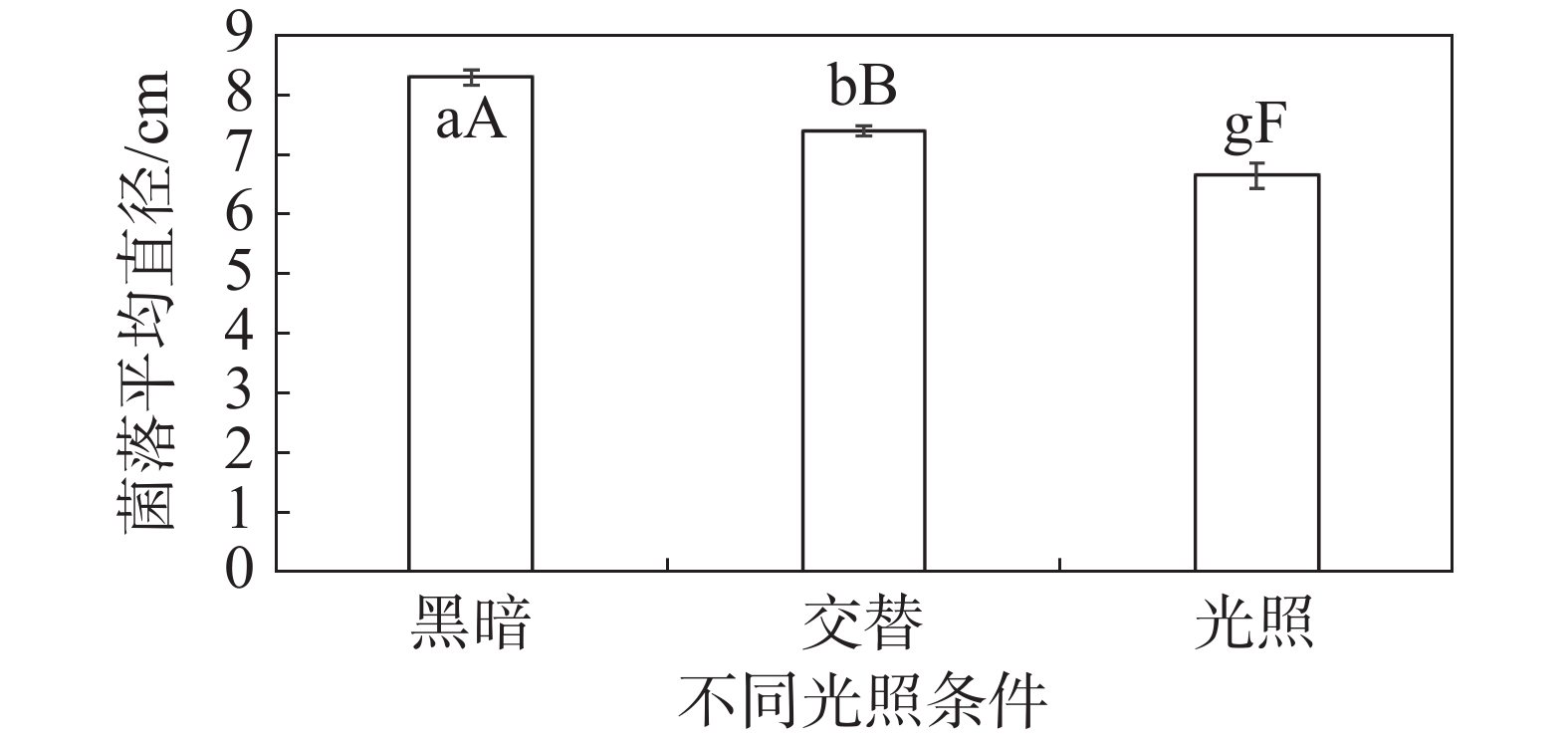
HNCJ201801001菌株在3种光照情况下的菌落直径有一定差异。在24 h光照条件下,HNCJ201801001菌株菌丝生长最慢,边缘透明稀薄,长势最弱。在12 h光暗交替条件下,HNCJ201801001菌株菌丝生长较慢,边缘厚度不均匀。在24 h黑暗状态下菌落生长最快,菌落生长均匀,边缘整齐,厚度最厚(图6)。
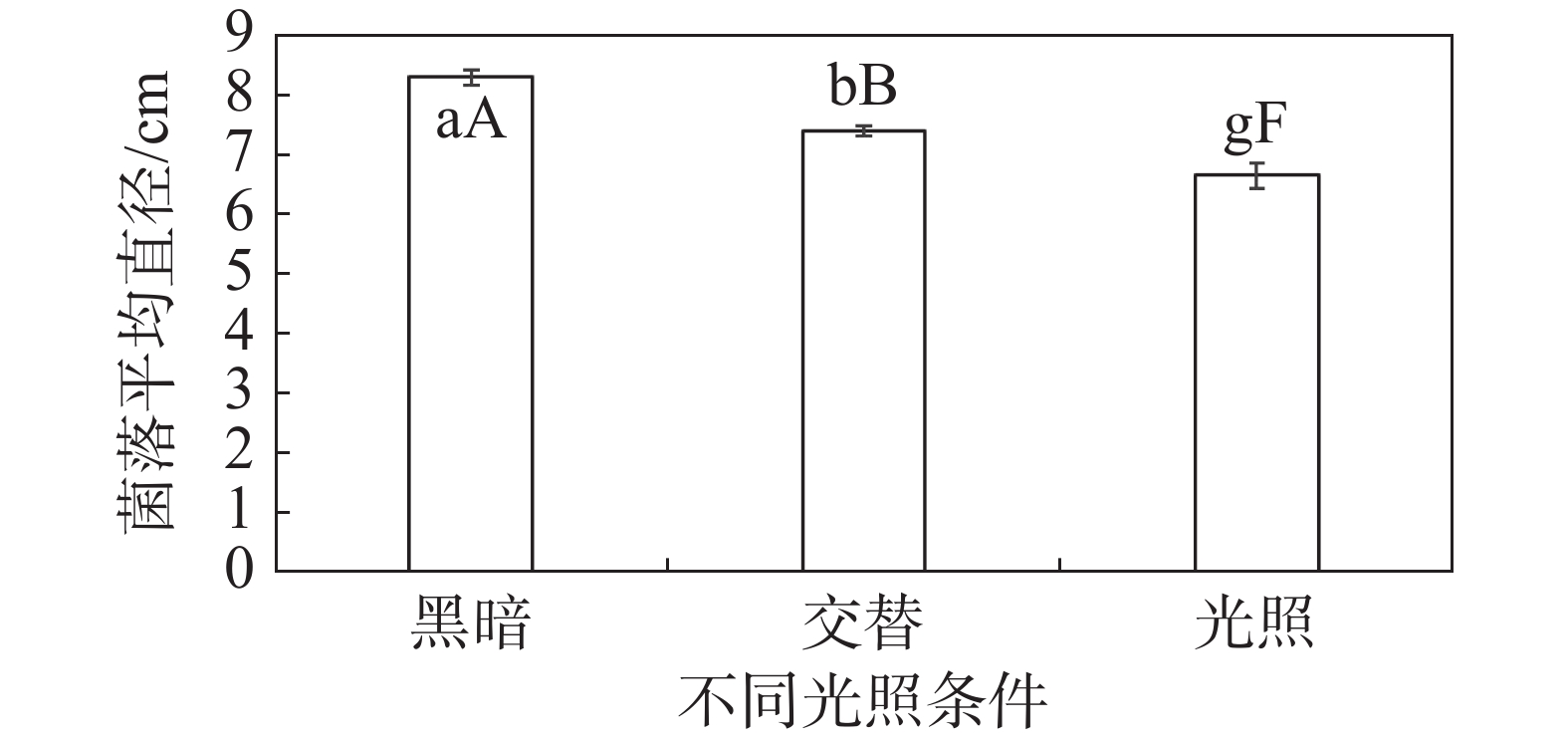
图 6 不同光照对木麻黄茎腐病病原菌HNCJ201801001菌丝生长的影响
-
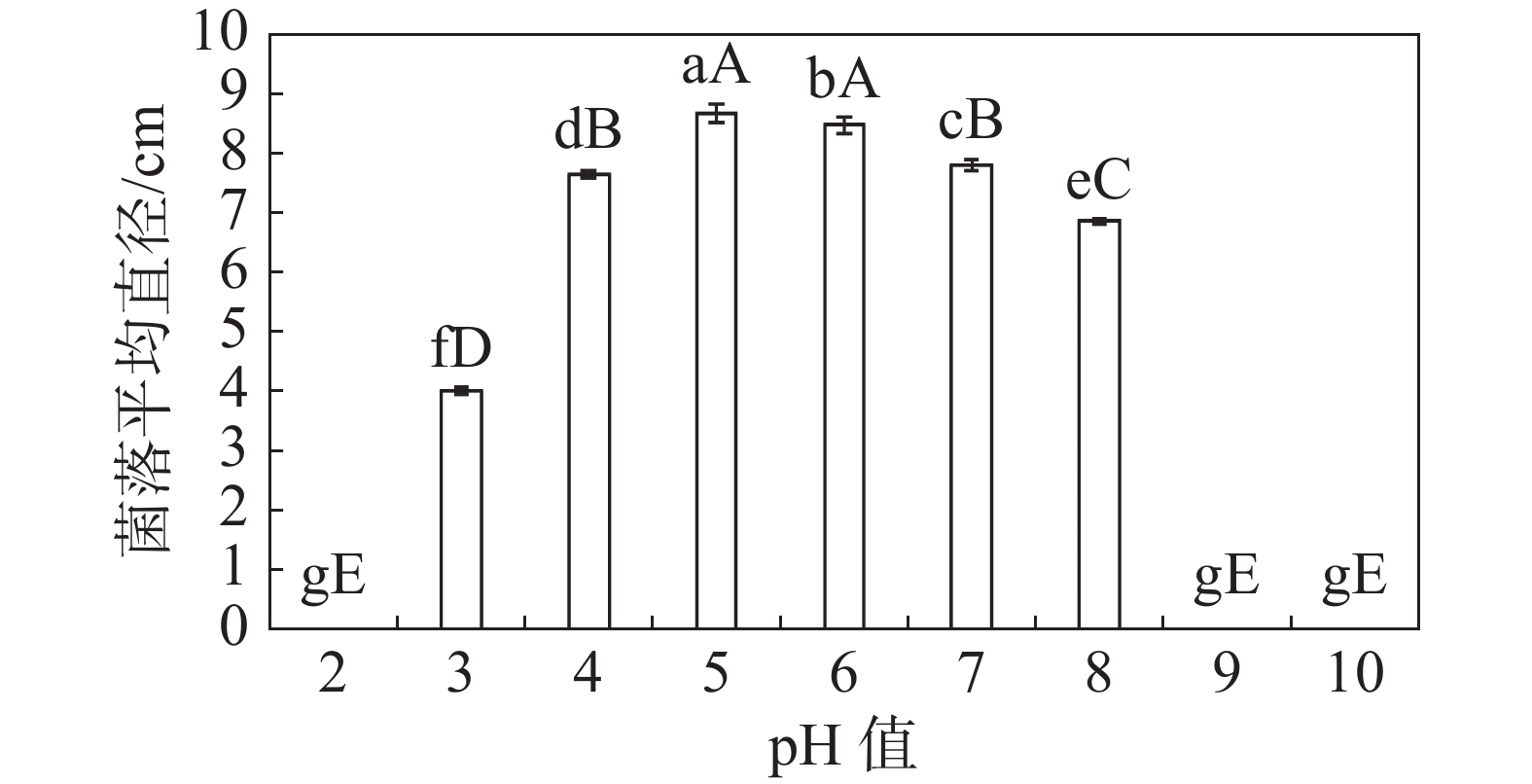
HNCJ201801001菌株在pH 3~8之间均可以正常生长,pH 4~7时生长得比较快。pH从2到5时,菌株生长速度随pH升高而加快。pH从5到9时,菌株生长速度随pH升高而减慢,最适生长pH为5。实验结果表明,弱酸性环境相比中性与碱性环境更适合该致病菌株的生长(图7)。

图 7 不同pH对木麻黄茎腐病病原菌HNCJ201801001菌丝生长的影响
-
HNCJ201801001菌株在6种培养基上均可以生长。不同处理组菌落的生长速率与状况有明显差异,其中,PDA与PSA培养基上的菌落菌丝层厚,生长最快;木麻黄茎干煎汁与燕麦培养基(CMA)上的菌落生长直径较大,但菌丝层薄于PDA与PSA培养基;在玉米(OMA)培养基上的菌落生长较慢, 菌丝层较稀薄;在Richards培养基上的菌落菌丝层接近透明,呈不规则多边形,生长最慢,长势极差(表3)。
表 3 不同培养基培对木麻黄茎腐病病原菌HNCJ201801001菌丝生长的影响
培养基 菌落直径/cm 菌丝生长状态 PDA 8.38 bB 菌丝层厚,厚薄均匀,
圆形,长势优良。Richard 4.23 fF 菌丝层稀疏近透明,
形状不规则,长势极差。燕麦 7.51 cC 菌丝层较厚,厚薄均匀,
圆形,长势良好。PSA 9.17 aA 菌丝层厚,厚薄均匀,
圆形,长势优良。玉米 6.25 eE 菌丝层薄,厚薄均匀,
近圆形,长势差。木麻黄茎干 7.13 dD 菌丝层较厚,厚薄均匀,
圆形,长势良好。 -
灵芝是灵芝属(Ganoderma)真菌的总称,目前世界范围内灵芝属有131个种,其中中国有23个种[23]。灵芝是一类重要的药用真菌,在中国作为滋补强壮的名贵药材,已有超过2000 a的药用历史。灵芝属真菌均为木腐菌,其中的部分种类会引起树木的根腐病或茎基腐病,导致树木枯死,如灵芝(Ganoderma lucidum)、橡胶树灵芝(G. pseudoferreum) 、有柄树舌(G. applanatum)和热带灵芝(G. tropicum)等。NAIDU等[24]于1966年报道灵芝菌寄主植物有19科36属48种,是一类重要的林木病原木腐菌。狭长孢灵芝(G. boninense)在马来西亚与印度尼西亚等国家是油棕(Elaeis guineensis)茎基腐病的主要病原菌,油棕被感染后引起茎基腐病并导致整株死亡,造成经济的损失超过5亿美元[25]。狭长孢灵芝在我国的分布以海南省为主[23]。目前,国内外对狭长孢灵芝的相关报道主要集中在化学成分、病害防治和基因组等几个方面[26-28],尚未出现关于狭长孢灵芝引起木麻黄活立木茎腐病的研究报道。
本研究结合形态学和分子生物学方法,确定了引起海南昌江木麻黄活立木茎腐病的病原菌为狭长孢灵芝。生物学特性测定结果显示,25~32 ℃均较适合该病原菌生长,其最适生长温度为28 ℃,最适pH为5;光照影响菌丝生长,病原菌在完全黑暗状态下生长速率最快;狭长孢灵芝适宜在PDA与PSA培养基上生长。程乐乐等[12]研究了引起海南陵水槟榔茎腐病的狭长孢灵芝,其适宜生长温度为26~30 ℃,最适生长温度为28 ℃,最适pH为5~6,光照抑制菌丝生长,最优碳源为蔗糖,与本研究结果基本一致,但陵水槟榔狭长孢灵芝在34 ℃以上不生长,而木麻黄狭长孢灵芝菌株在35 ℃依然可以生长,这可能是由于地区环境条件与寄主物种不同导致。笔者在田间调查时于海口桂林洋、澄迈桥头、昌江湿地公园、万宁乌场等地海滨多年生长的木麻黄活树和死树茎基部均发现生长有狭长孢灵芝的担子果,田间发病率约2%。
目前,国内有不少由灵芝属病原菌引起木麻黄茎腐或根腐病的报道。陈礼浪等[6]发现热带灵芝能导致木麻黄红根病;吴如慧等[8]与单金雪等[9]分别报道二孢假芝与南方灵芝是引起木麻黄茎腐病病原菌。
与本研究中的狭长孢灵芝相比,热带灵芝新鲜担子果同样具有漆状光泽,菌肉褐色,但其表面由中央到边缘为红褐色、紫红色逐渐变浅至黄白至白色。热带灵芝主要导致红根病,在田间症状上会表现出病根表面粘泥沙的特点,且水洗后可见枣红色革质菌膜。二孢假芝与南方灵芝均会导致木麻黄基部茎干组织白腐,但南方灵芝无光泽,担子果多为土褐色,菌肉肉桂色,有黑色壳质层,而二孢假芝表面有一定光泽,表面红褐色或紫褐色,多数有同心环带,但其菌肉一般为白色或浅黄色。本研究中的狭长孢灵芝,担子果上表面红褐色,有漆状光泽,与南方灵芝特征不同。菌盖边缘为橘黄色,靠近中部变为红或暗紫色,有明显的同心环纹和放射状突起纵脊,可与热带灵芝相区分。菌肉为褐色,长有黑色壳质层,区别于二孢假芝。狭长孢灵芝同样也能导致木麻黄活树茎干组织白腐。灵芝属中多种木腐真菌均可以导致木麻黄茎腐或根腐病的发生。
Identification and biological characteristics of the pathogen causing stem rot of Casuarina equisetifolia
-
摘要: 在海南省昌江县棋子湾生长的木麻黄(Casuarina equisetifolia)上发现一种新的灵芝茎腐病(Ganoderma sp.)。通过组织分离法从病原菌担子果中分离出代表菌株HNCJ201801001。对该菌株进行致病性测定、担子果诱导、形态学鉴定,构建rDNA-ITS、LSU系统发育树分析,完成生物学特性测定。依据形态学与分子检测结果,确定海南省昌江县棋子湾木麻黄茎腐病的病原菌为狭长孢灵芝(Ganoderma boninense)。对其进行生物学特性测定,结果表明,该菌的最适温度为28 ℃,最适pH为5.0,黑暗条件更有利于菌丝生长,在PDA和PSA培养基上生长较好。Abstract: A new stem rot caused by Ganoderma sp. was found on Casuarina equisetifolia growing in Qizi Bay, Changjiang County, Hainan Province. A representative strain HNCJ201801001 was isolated from basidiocarps of the pathogen by tissue isolation method. The pathogenicity determination, basidiocarp induction and morphological identification of the strain were carried out, a phylogenetic tree using rDNA-ITS and LSU genes was constructed for analysis of the strain, and the biological characteristics of the strain were determined. According to the results of morphological and molecular detection, the pathogen of C. equisetifolia stem rot in Qizi Bay, Changjiang County, Hainan Province was identified as Ganoderma boninense. The biological characteristics of G. boninense were determined and the results showed that the growth of G. boninense was optimum when cultured at temperature 28 ℃, pH5.0 in dark. Dark condition was more conducive to mycelial growth of G. boninense, and G. boninense grew better on PDA and PSA mediums than on the other mediums.
-
Key words:
- Casuarina equisetifolia /
- stem rot /
- Ganoderma boninense /
- biological characteristics
-
表 2 用于分子鉴定的菌株及其ITS与LSU基因登录号
种名 菌株号 ITS基因 LSU基因 Ganoderma australe ZRL20151500 LT716076.1 KY418900.1 Ganoderma gibbosum Pvc62 MK280717.1 MK215785.1 Ganoderma adspersum MFLU 19-2220 MN396655.1 MN428663.1 Ganoderma lingzhi HFJAU0581 MH294333.1 MH294366.1 Lentinus tigrinus CBS 249.39 MH856001.1 MH867501.1 Ganoderma lucidum GL-1 GU726919.1 MH873099.1 Ganoderma boninense WD 2028 KJ143905.1 KU220015.1 Ganoderma tsunodae WD2034 AB588989.1 AB368069.1 Ganoderma resinaceum CBS 220.36 MH855781.1 MH867290.1 Ganoderma nasalanense LPDR17060211 MK345441.1 MK346831.1 Ganoderma nasalanense LPDR17060212 MK345442.1 MK346832.1 Haddowia longipes LPDR17072708 MK345423.1 MK346828.1 Ganoderma orbiforme URM 86213 MK119829.1 MK119909.1 Ganoderma ecuadoriense URM 89449 MK119828.1 MK119908.1  下载: 导出CSV
下载: 导出CSV
表 3 不同培养基培对木麻黄茎腐病病原菌HNCJ201801001菌丝生长的影响
培养基 菌落直径/cm 菌丝生长状态 PDA 8.38 bB 菌丝层厚,厚薄均匀,
圆形,长势优良。Richard 4.23 fF 菌丝层稀疏近透明,
形状不规则,长势极差。燕麦 7.51 cC 菌丝层较厚,厚薄均匀,
圆形,长势良好。PSA 9.17 aA 菌丝层厚,厚薄均匀,
圆形,长势优良。玉米 6.25 eE 菌丝层薄,厚薄均匀,
近圆形,长势差。木麻黄茎干 7.13 dD 菌丝层较厚,厚薄均匀,
圆形,长势良好。
下载: 导出CSV
-
[1] 黄桂华, 仲崇禄, 张勇. 我国木麻黄科植物遗传改良研究进展[J]. 广东林业科技, 2005(4): 65 − 69. [2] 刘成路, 冉焰辉, 陶悠, 等. 海南岛海岸线木麻黄林现状调查[J]. 林业资源管理, 2013(2): 102 − 106. doi: 10.3969/j.issn.1002-6622.2013.02.019 [3] 胡小婵, 温春生. 海南省海防林研究[J]. 防护林科技, 2009(2): 63 − 64. doi: 10.3969/j.issn.1005-5215.2009.02.024 [4] 谢银燕, 王松, 吴春银, 等. 木麻黄病虫害及其防治的最新进展[J]. 江苏农业科学, 2019, 47(20): 36 − 41. [5] 杨彬. 海南岛木麻黄海防林天然更新特征、影响因素及评价[D]. 海口: 海南师范大学, 2019. [6] 陈礼浪. 海南木麻黄木腐菌的多样性及病原木腐菌致病性研究[D]. 海口: 海南大学, 2016. [7] 陈礼浪, 李增平. 木麻黄红根病病原菌鉴定及其生物学特性测定[J]. 热带作物学报, 2016, 37(6): 1188 − 1193. doi: 10.3969/j.issn.1000-2561.2016.06.023 [8] 吴如慧, 李增平, 陈礼浪. 木麻黄茎腐病病原菌的鉴定及其生物学特性测定[J]. 热带作物学报, 2019, 40(2): 334 − 340. [9] 单金雪, 李增平, 张宇, 等. 一种木麻黄灵芝茎腐病病原鉴定及生物学特性[J]. 热带生物学报, 2021, 12(1): 88 − 95. [10] 曹旭, 商亮, 李晶, 等. 利用微生物防治油棕榈茎基底感染狭长孢灵芝(Ganoderma boninense)研究[J]. 黑龙江科学, 2020, 11(14): 1 − 3. doi: 10.3969/j.issn.1674-8646.2020.14.001 [11] 蔡志浓, 安宝贞, 林筑苹, 等. 灵芝引起的槟榔基腐病[J]. 台湾农业研究, 2018, 67(3): 318 − 322. [12] 程乐乐, 李增平, 蒙汉华. 槟榔茎基腐病病原菌鉴定及其生物学特性测定[J]. 热带作物学报, 2017, 38(1): 119 − 125. doi: 10.3969/j.issn.1000-2561.2017.01.021 [13] 方中达. 植病研究方法[M]. 北京: 中国农业出版社, 1998: 46 − 50. [14] 李增平. 普通植物病理学实验、实习指导[M]. 北京: 中国农业出版社, 2017: 71 − 78. [15] 高秀兵, 李增平, 李晓娜, 等. 橡胶树几种根病的人工接种方法[J]. 热带作物学报, 2010, 31(4): 626 − 630. [16] 文庭池, 邓春英, 吴兴亮. 海南岛7种野生灵芝的形态与分子鉴定[J]. 贵州科学, 2015, 33(6): 1 − 8. doi: 10.3969/j.issn.1003-6563.2015.06.001 [17] 胡真臻, 李增平, 单金雪,等. 橡胶树灵芝茎腐病病原菌鉴定及其生物学特性测定[J]. 热带作物学报, 2021, 42(2): 488 − 494. [18] 赵继鼎, 张小青. 中国真菌志: 第18卷 灵芝科[M]. 北京: 科学出版社, 2000: 28 − 143. [19] 卯晓岚. 中国大型真菌[M]. 郑州: 河南科学技术出版社, 2000. [20] 中国菌物学会. 中国菌物学会2018年学术年会论文汇编[C]. 北京: 中国菌物学会: 2018. [21] 袁滨, 严俊杰, 柯丽娜, 等. 基于ITS系列分析鉴定野生灵芝属菌种[J]. 中国食用菌, 2018, 37(2): 62 − 66. [22] OGHENEKARO A O, MIETTINEN O, OMORUSI V I, et al. Molecular phylogeny of Rigidoporus microporus isolates associated with white rot disease of rubber trees (Hevea brasiliensis)[J]. Fungal Biology, 2014, 118(5 − 6): 495 − 506. [23] 邢佳慧. 灵芝属的物种多样性、分类与系统发育研究[D]. 北京: 北京林业大学, 2019. [24] NAIDU GVB, KUMAR SNS, SANNAMARAPPA M, et al. Ganoderma lucidum (Leys.) Karst. on arecanut palm: A review and further observations [J]. Mysore Hort. Soc, 1966, 1(3): 14 − 20. [25] HUSHIARIAN R, YUSOF N A, DUTSE S W. Detection and control of Ganoderma boninense: strategies and perspectives [J]. Springerplus, 2013, 2(1): 1 − 12. doi: 10.1186/2193-1801-2-1 [26] SUSANTO A, SUDHARTO P S, PURBA R Y. Enhancing biological control of basal stem rot disease (Ganoderma boninense) in oil palm plantations [J]. Mycopathologia, 2005, 159(1): 153 − 157. doi: 10.1007/s11046-004-4438-0 [27] 于晓丹, 王一菲, 姜霁航, 等. 狭长孢灵芝液体发酵上清液抗氧化活性研究[J]. 菌物学报, 2020, 39(1): 84 − 98. [28] PILOTTI C A, SANDERSON F R, AITKEN E. Genetic structure of a population of Ganoderma boninense on oil palm [J]. Plant Pathology, 2003, 52(4): 455 − 463. doi: 10.1046/j.1365-3059.2003.00870.x -

 点击查看大图
点击查看大图

图(7) / 表 (3)
计量
- 文章访问数: 693
- HTML全文浏览量: 235
- PDF下载量: 12
- 被引次数: 0
